Interazioni tra luce e materia vivente -...
6
Interazioni tra luce e materia vivente CH3 CH3 CH3 IHIHIH LH H H H H2C i i C H3 H2 CH3 Alla luce sono dovuti tre processi fondamentali della vita: la fotosintesi, la visione e la fotoperiodicità (cioè la risposta delle piante e degli animali al ripetersi ciclico della notte e del giorno) S i ritiene che la vita si sia originata da una materia primordiale, per azione della luce del Sole su mo- lecole semplici, quando la crosta terre- stre iniziò a raffreddarsi. Probabilmen- te essa non subí mutamenti per perio- di di tempo indefiniti, ma, a un certo punto, con l'avvento della fotosintesi, alcuni organismi riuscirono a utilizza- re la luce del Sole più direttamente. Cosí succede anche attualmente nelle piante, le quali, mediante fotosintesi sfruttano la luce del Sole per le neces- sità energetiche di tutte le forme di vita. Quando si svilupparono le varie spe- cie di animali, vennero favorite, per quanto riguarda la sopravvivenza, solo quelle che erano maggiormente capaci di percepire quanto accadeva attorno a loro. Per utilizzare gli stimoli lumi- nosi, gli animali avevano bisogno di una specie di rivelatore : un tessuto, una macchia oculare, o un occhio. Il rivelatore doveva essere accoppiato a un sistema di risposta : un ganglio o un cervello. I segnali provenienti da questo sistema guidavano gli sposta- menti verso il cibo o lontano dal pe- ricolo. La fotosintesi e la visione non sono i soli tipi di utilizzazione delle fonti lu- minose. Sia gli animali che le piante, infatti, hanno sviluppato anche dei meccanismi di risposta ai cambiamenti ciclici quotidiani della luce e del buio. Tra l'altro, questa fotoperiodicità re- gola il ripetersi stagionale della fiori- tura delle piante, della metamorfosi de- gli insetti e delle nidiate degli uccelli. Per comprendere questi fenomeni ci si può domandare come la luce agisca sulla vita. Una parte della risposta è molto semplice : agisce eccitando par- ticolari molecole in grado di assorbirla. Più difficili da descrtvere sono le modi- ficazioni subite da queste molecole nel corso dell'assorbimento, ma si cono- di Sterling B. Hendricks scono attualmente abbastanza bene molti dettagli dei processi che interven- gono. Le nostre idee circa il modo in cui le reazioni molecolari sono colle- gate alle risposte degli organismi sono però ancora allo stadio di ipotesi. D iscutendo sulle attuali conoscenze delle interazioni tra la luce e la vita, tratteremo per prima cosa della visione, poiché questo fenomeno ha molti aspetti in comune sia con la fo- tosintesi che con la fotoperiodicità. In tutti e tre i processi la luce inizia la sua azione con l'assorbimento da parte di una piccola molecola colorata – il cromoforo – associata a una grande molecola di proteina. Nel caso della visione le molecole sensibili alla luce impartiscono una colorazione rosa vio- laceo alla retina. Nella retina dell'oc- chio umano vi sono circa 100 milioni di cellule a forma di bastoncelli e cin- que milioni a forma di cono. Ognuna di esse è collegata, attraverso una si- napsi a un neurite sensitivo che con- duce lo stimolo al cervello. Fotografie al microscopio elettronico dimostrano che le estremità dei bastoncelli e dei coni sono unite a sottili sacchi membra- nosi, ai quali si associano i cromofori che assorbono la luce. (La vista e la fotosintesi dipendono da questa asso- ciazione di un cromoforo con una membrana.) La luce, che eccita il cro- moforo, provoca nella membrana dei cambiamenti, che vengono percepiti sotto forma di stimoli dalla fibra ner- vosa. Per quanto riguarda la visione si co- noscono bene sia la struttura del cro- moforo ricevente sia il suo sistema di eccitazione luminosa. Il cromoforo ri- cettore è un'a'deide della vitamina A (in termini strutturistici 1-cis-retine- ne), come stabili George Wald della Harvard University. Si è trovato che questo cromoforo è associato a una pro- teina, l'opsina. Le opsine sono proteine grasse; hanno perciò affinità con la membrana del sacco, costituita princi- palmente da lipidi, cioè da grassi. Vi sono quattro tipi di opsine, uno nei bastoncelli e tre nei coni. Combi- nate con 1'1 1-cis-retinene, esse forma- no rispettivamente la rodopsina e tre tipi di iodopsine. Per l'eccitazione lu- minosa, tutte e quattro le opsine si mo- dificano allo stesso modo. Nei fenomeni della visione le mole- cole del cromoforo, della fotosintesi e della fotoperiodicità, sono notevolmen- te simili per la presenza di semplici e doppi legami alternati. Molecole di questo genere, conosciute dai chimici come « sistemi coniugati », sono strut- turalmente assai stabili, poiché non si può verificare la rotazione di gruppi atomici uniti tramite doppi legami. Ogni sistema coniugato, se è esteso in modo opportuno, ha un livello ener- getico tanto basso da poter essere ecci- tato dalla luce visibile. Quando il si- stema viene eccitato, si verifica un rilas- samento dei doppi legami, per cui la sua configurazione da cis diventa trans (si veda la figura in alto nella pagina a fronte). Questa capacità di cambiare forma è un elemento chiave sia nel fe- nomeno della visione sia in quello del- la fotoperiodicità. Nella fotosintesi, tut tavia, non si verificano cambiamenti di forma a causa della struttura fissa ad anello dei cromofori della clorofilla. Gli effetti della luce suPa visione e sulla fotoperiodicità sono determinabili con misure delle trasformazioni mole- colari provocate dalla eccitazione. Al- cune di queste trasformazioni sono molto rapide : possono aver luogo in meno di un milionesimo di secondo. Trasformazioni cosí ve l oci si possono seguire solo se l'eccitazione ha la du- rata di un brevissimo, ma intenso, lam- po di luce. Anche la misura del'e tra- sformazioni deve avvenire entro un tempo brevissimo. Il metodo impiega- to è l'eccitazione con un lampo di luce a temperatura ambiente, o anche mi- nore, seguita da fotoanalisi, che è la tecnica per cui George Porter e R.G.W. Norrish (insieme a Manfred Eigen), ottennero il premio Nobel per la chimica nel 1967 (si veda l'articolo Gli effetti chimici della luce di Gerald Oster, in e Le Scienze » n. 12, agosto 1969). Le basse temperature rallentano le trasformazioni molecolari e le ren- dono più facili da misurare. Quando il cromoforo viene eccitato dalla luce, si trasforma da cis a trans. Ne risulta che la rodopsina si converte in prelumirodopsina, con un cromoforo di configurazione trans. Il ruolo che la luce gioca nel fenomeno visivo si li. mita a consentire la realizzazione di questa trasformazione, che è seguita da molti rapidi cambiamenti nella strut- tura dell'opsina e nelle relazioni tra il cromoforo e l'opsina. Per avere una idea del tempo che lo stimolo impiega per arrivare dalla cellula della retina all'estremità del neurite, si pensi che l'eccitamento avviene a causa di feno- meni che hanno luogo nel primo mille- simo di secondo. L a sequenza delle trasformazioni mo- lecolari può essere seguita studian- do la rodopsina in soluzione. La prelu- mirodopsina, identificabile dal suo mas- simo di assorbimento della luce a 543 nanometri, può essere ottenuta a tem- perature inferiori a –140°C e rever- sibilmente trasformata in rodopsina. Quando la prelumirodopsina viene por- tata alla temperatura di –40 °C, si tra- sforma in lumirodopsina. La stessa tra- sformazione probabilmente avviene an- che alla temperatura del corpo umano, ma molto più rapidamente. Questi cambiamenti e quelli successivi, che comprendono la formazione di metaro- dopsine, danno luogo a trasformazioni della configurazione molecolare della opsina. Nei vertebrati si giunge infine alla dissociazione del cromoforo dalla proteina. La forma rilassata del retine- ne, completamente trans, deve essere ridotta a forma alcoolica e riossidata ad aldeide per rigenerare 1-cis-retinene. Non appena rigenerato, il cis-retinene si combina spontaneamente con l'op- sina per formare di nuovo la rodop- sina. Analisi delle variazioni del potenzia- le elettrico che si verificano simultanea- mente alle trasformazioni molecolari dimostrano che tali fenomeni hanno luogo entro 25 milionesimi di secondo dopo il lampo di luce. In un circuito che comprende i tes- suti circostanti e la retina, il potenzia- le della cornea è positivo, ma è segui- CH3 CH3 i H i H C1 C 1C H -? H H H2C L'Ha H3C- CH C i H2 CH3 C(1) H CIS TRANS La visione dipende da una molecola, detta cromoforo, sensibile alla luce, l'll-cis-retinene (a sinistra), che ha legami singoli e doppi alternati (in colore). Quando la luce eccita la molecola, questa passa dalla forma cis a quella trans (a destra), provocando una serie di complesse trasformazioni chimiche nella struttura delle proteine associate al cromoforo. COOH COOH ROSSO CH3 CH CH 3 CH2 CH2 CH3 CH3 CH CH3 CH2 CH2 CH3 I I i i I I HC — C C C C—C HC C I i ii ii i i i I O N C N ''C'.'" N ."- C' N -'0 HHH H H H ROSSO LONTANO La fotoperiodicità nelle piante dipende dal fitocromo, altra molecola sensibile alla luce. Come il retinene ha doppi legami coniugati (in colore). Quando si verifica uno stimolo luminoso, esso passa da una configurazione sensibile al rosso (in alto), a una sensibile al rosso lontano, a causa dello spostamento di due atomi d'idrogeno (in basso). H2C % CH CH3 I H i C C C / \ /CH3 H 3 C—C i i /C —CH2 C—N ,N=C / \ HC Mg /C H C—N N—C CH "C—CH3 / \ /C\ CH C C i CH2 CH3 CH3 CH3 CH3 CH2 HC—C i ,... 0 I H2 I H2 I H2 i H2 i C-0 \ CH3 H 3 C H C C H's- C C H C". C C ' '''0 O O H2 H2 H2 H2 H2 H2 H FITOLO CLOROFILLINA La fotosintesi dipende da molecole di differenti specie di clorofilla, che hanno diversi gruppi laterali. Questa molecola è quella della clorofilla a. Come i cromofori della visione e della fotoperiodicità, le molecole di clorofilla hanno doppi legami coniugati (in colore) i quali, però, formano un cappio chiuso entro la parte clorofillinica della molecola. Quando si ha l'eccitazione luminosa, la clorofilla trasferisce l'energia ricevuta a centri dove avvengono le trasformazioni chimiche (si veda la figura a pagina 70). COOH COOH i I CH3 CH2 CH2 i CH3 CH2' CH3 CH2 CH2 CCCC C I I II II I CH3 i CH3' CH3 CH2 C C C H H H H H H 60 61
Transcript of Interazioni tra luce e materia vivente -...

Interazioni tra luce emateria vivente
CH3 CH3 CH3IHIHIH
LH H H HH2C
ii C H3
H2 CH3
Alla luce sono dovuti tre processi fondamentali della vita: lafotosintesi, la visione e la fotoperiodicità (cioè la risposta dellepiante e degli animali al ripetersi ciclico della notte e del giorno)
S
i ritiene che la vita si sia originatada una materia primordiale, perazione della luce del Sole su mo-
lecole semplici, quando la crosta terre-stre iniziò a raffreddarsi. Probabilmen-te essa non subí mutamenti per perio-di di tempo indefiniti, ma, a un certopunto, con l'avvento della fotosintesi,alcuni organismi riuscirono a utilizza-re la luce del Sole più direttamente.Cosí succede anche attualmente nellepiante, le quali, mediante fotosintesisfruttano la luce del Sole per le neces-sità energetiche di tutte le forme divita.
Quando si svilupparono le varie spe-cie di animali, vennero favorite, perquanto riguarda la sopravvivenza, soloquelle che erano maggiormente capacidi percepire quanto accadeva attornoa loro. Per utilizzare gli stimoli lumi-nosi, gli animali avevano bisogno diuna specie di rivelatore : un tessuto,una macchia oculare, o un occhio. Ilrivelatore doveva essere accoppiato aun sistema di risposta : un ganglio oun cervello. I segnali provenienti daquesto sistema guidavano gli sposta-menti verso il cibo o lontano dal pe-ricolo.
La fotosintesi e la visione non sonoi soli tipi di utilizzazione delle fonti lu-minose. Sia gli animali che le piante,infatti, hanno sviluppato anche deimeccanismi di risposta ai cambiamenticiclici quotidiani della luce e del buio.Tra l'altro, questa fotoperiodicità re-gola il ripetersi stagionale della fiori-tura delle piante, della metamorfosi de-gli insetti e delle nidiate degli uccelli.
Per comprendere questi fenomeni cisi può domandare come la luce agiscasulla vita. Una parte della risposta èmolto semplice : agisce eccitando par-ticolari molecole in grado di assorbirla.Più difficili da descrtvere sono le modi-ficazioni subite da queste molecole nelcorso dell'assorbimento, ma si cono-
di Sterling B. Hendricks
scono attualmente abbastanza benemolti dettagli dei processi che interven-gono. Le nostre idee circa il modo incui le reazioni molecolari sono colle-gate alle risposte degli organismi sonoperò ancora allo stadio di ipotesi.
D iscutendo sulle attuali conoscenzedelle interazioni tra la luce e la
vita, tratteremo per prima cosa dellavisione, poiché questo fenomeno hamolti aspetti in comune sia con la fo-tosintesi che con la fotoperiodicità. Intutti e tre i processi la luce inizia lasua azione con l'assorbimento da partedi una piccola molecola colorata – ilcromoforo – associata a una grandemolecola di proteina. Nel caso dellavisione le molecole sensibili alla luceimpartiscono una colorazione rosa vio-laceo alla retina. Nella retina dell'oc-chio umano vi sono circa 100 milionidi cellule a forma di bastoncelli e cin-que milioni a forma di cono. Ognunadi esse è collegata, attraverso una si-napsi a un neurite sensitivo che con-duce lo stimolo al cervello. Fotografieal microscopio elettronico dimostranoche le estremità dei bastoncelli e deiconi sono unite a sottili sacchi membra-nosi, ai quali si associano i cromoforiche assorbono la luce. (La vista e lafotosintesi dipendono da questa asso-ciazione di un cromoforo con unamembrana.) La luce, che eccita il cro-moforo, provoca nella membrana deicambiamenti, che vengono percepitisotto forma di stimoli dalla fibra ner-vosa.
Per quanto riguarda la visione si co-noscono bene sia la struttura del cro-moforo ricevente sia il suo sistema dieccitazione luminosa. Il cromoforo ri-cettore è un'a'deide della vitamina A(in termini strutturistici 1-cis-retine-ne), come stabili George Wald dellaHarvard University. Si è trovato chequesto cromoforo è associato a una pro-
teina, l'opsina. Le opsine sono proteinegrasse; hanno perciò affinità con lamembrana del sacco, costituita princi-palmente da lipidi, cioè da grassi.
Vi sono quattro tipi di opsine, unonei bastoncelli e tre nei coni. Combi-nate con 1'1 1-cis-retinene, esse forma-no rispettivamente la rodopsina e tretipi di iodopsine. Per l'eccitazione lu-minosa, tutte e quattro le opsine si mo-dificano allo stesso modo.
Nei fenomeni della visione le mole-cole del cromoforo, della fotosintesi edella fotoperiodicità, sono notevolmen-te simili per la presenza di semplici edoppi legami alternati. Molecole diquesto genere, conosciute dai chimicicome « sistemi coniugati », sono strut-turalmente assai stabili, poiché non sipuò verificare la rotazione di gruppiatomici uniti tramite doppi legami.
Ogni sistema coniugato, se è estesoin modo opportuno, ha un livello ener-getico tanto basso da poter essere ecci-tato dalla luce visibile. Quando il si-stema viene eccitato, si verifica un rilas-samento dei doppi legami, per cui lasua configurazione da cis diventa trans(si veda la figura in alto nella pagina afronte). Questa capacità di cambiareforma è un elemento chiave sia nel fe-nomeno della visione sia in quello del-la fotoperiodicità. Nella fotosintesi, tuttavia, non si verificano cambiamentidi forma a causa della struttura fissaad anello dei cromofori della clorofilla.
Gli effetti della luce suPa visione esulla fotoperiodicità sono determinabilicon misure delle trasformazioni mole-colari provocate dalla eccitazione. Al-cune di queste trasformazioni sonomolto rapide : possono aver luogo inmeno di un milionesimo di secondo.Trasformazioni cosí ve l oci si possonoseguire solo se l'eccitazione ha la du-rata di un brevissimo, ma intenso, lam-po di luce. Anche la misura del'e tra-sformazioni deve avvenire entro un
tempo brevissimo. Il metodo impiega-to è l'eccitazione con un lampo di lucea temperatura ambiente, o anche mi-nore, seguita da fotoanalisi, che è latecnica per cui George Porter eR.G.W. Norrish (insieme a ManfredEigen), ottennero il premio Nobel perla chimica nel 1967 (si veda l'articoloGli effetti chimici della luce di GeraldOster, in e Le Scienze » n. 12, agosto1969). Le basse temperature rallentanole trasformazioni molecolari e le ren-dono più facili da misurare.
Quando il cromoforo viene eccitatodalla luce, si trasforma da cis a trans.Ne risulta che la rodopsina si convertein prelumirodopsina, con un cromoforodi configurazione trans. Il ruolo che laluce gioca nel fenomeno visivo si li.mita a consentire la realizzazione diquesta trasformazione, che è seguita damolti rapidi cambiamenti nella strut-tura dell'opsina e nelle relazioni tra ilcromoforo e l'opsina. Per avere unaidea del tempo che lo stimolo impiegaper arrivare dalla cellula della retinaall'estremità del neurite, si pensi chel'eccitamento avviene a causa di feno-meni che hanno luogo nel primo mille-simo di secondo.
La sequenza delle trasformazioni mo-lecolari può essere seguita studian-
do la rodopsina in soluzione. La prelu-mirodopsina, identificabile dal suo mas-simo di assorbimento della luce a 543nanometri, può essere ottenuta a tem-perature inferiori a –140°C e rever-sibilmente trasformata in rodopsina.Quando la prelumirodopsina viene por-tata alla temperatura di –40 °C, si tra-sforma in lumirodopsina. La stessa tra-sformazione probabilmente avviene an-che alla temperatura del corpo umano,ma molto più rapidamente. Questicambiamenti e quelli successivi, checomprendono la formazione di metaro-dopsine, danno luogo a trasformazionidella configurazione molecolare dellaopsina. Nei vertebrati si giunge infinealla dissociazione del cromoforo dallaproteina. La forma rilassata del retine-ne, completamente trans, deve essereridotta a forma alcoolica e riossidata adaldeide per rigenerare 1-cis-retinene.Non appena rigenerato, il cis-retinenesi combina spontaneamente con l'op-sina per formare di nuovo la rodop-sina.
Analisi delle variazioni del potenzia-le elettrico che si verificano simultanea-mente alle trasformazioni molecolaridimostrano che tali fenomeni hannoluogo entro 25 milionesimi di secondodopo il lampo di luce.
In un circuito che comprende i tes-suti circostanti e la retina, il potenzia-le della cornea è positivo, ma è segui-
CH3 CH3i H i H
C1C1C H-?H HH2C
L'Ha
H3C- CHC i H2 CH3
C(1)H
CIS
TRANS
La visione dipende da una molecola, detta cromoforo, sensibile alla luce, l'll-cis-retinene(a sinistra), che ha legami singoli e doppi alternati (in colore). Quando la luce eccita lamolecola, questa passa dalla forma cis a quella trans (a destra), provocando una seriedi complesse trasformazioni chimiche nella struttura delle proteine associate al cromoforo.
COOH COOH
ROSSO
CH3 CH CH 3 CH2 CH2 CH3 CH3 CH
CH3 CH2 CH2 CH3I I i i
I I
HC—C C C C—C HC CI i ii ii i i i I
O N C N ''C'.'" N ."- C' N -'0HHH H H H
ROSSO LONTANO
La fotoperiodicità nelle piante dipende dal fitocromo, altra molecola sensibile allaluce. Come il retinene ha doppi legami coniugati (in colore). Quando si verifica unostimolo luminoso, esso passa da una configurazione sensibile al rosso (in alto), a unasensibile al rosso lontano, a causa dello spostamento di due atomi d'idrogeno (in basso).
H2C %
CH CH3I H iC C C
/ \ /CH3H 3C—C i i /C —CH2
C—N ,N=C/ \
HC Mg /C HC—N N—C
CH "C—CH3/\ /C\
CH C Ci
CH2
CH3 CH3 CH3 CH3 CH2 HC—Ci ,...
0I H2 I H2 I H2 i H2 i C-0\
CH3H 3C H C C H's- C C H C". C C ''''0 O O
H2 H2 H2 H2 H2 H2 H
FITOLO CLOROFILLINA
La fotosintesi dipende da molecole di differenti specie di clorofilla, che hanno diversigruppi laterali. Questa molecola è quella della clorofilla a. Come i cromofori dellavisione e della fotoperiodicità, le molecole di clorofilla hanno doppi legami coniugati(in colore) i quali, però, formano un cappio chiuso entro la parte clorofillinica dellamolecola. Quando si ha l'eccitazione luminosa, la clorofilla trasferisce l'energia ricevutaa centri dove avvengono le trasformazioni chimiche (si veda la figura a pagina 70).
COOH COOHi I
CH3 CH2 CH2i
CH3 CH2' CH3 CH2 CH2
CCCC CI I II II I
CH3i
CH3' CH3 CH2
C C C
H H H H H H
60 61

LUCE
C/S-RETINENE
C773
to, in un millesimo di secondo, da unsegnale in aumento di segno opposto.Questi fenomeni si verificano duranteil periodo in cui sono presenti sia laprelumirodopsina sia la lumirodopsina.Il primo potenziale probabilmente ac-compagna la trasformazione della ro-dopsina in prelumirodopsina. Il suc-cessivo dipende nettamente dalla tem-peratura che si ha a una certa distanzadal luogo in cui la luce agisce, proba-bilmente nella membrana esterna deibastoncelli o dei coni.
Sarebbe interessante poter identifi-care queste variazioni di potenziale co-me primi stadi dell'eventuale eccitazio-
TRANS-RETINENE
PRELUM I-RODOPSINA
498 NANOMETRI 543 NANOM ETR I> — 140 °C
CROMOFOROLUCE
ROSSA
PROTEINA
FITOCROMOROSSO A
660 NANOMETRI 690 NANOMETRI
> — 110 °C
ne della fibra nervosa. Un'altra ipotesiè che l'eccitazione del nervo sia asso-ciata con la transizione che si ha trala metarodopsina I e la metarodopsi-na II (si veda la figura in alto in que-sta pagina).
La percezione dei colori dipende dal-le tre opsine, che si trovano ciascunain un differente cono. I loro spettri diassorbimento sono stati rilevati, e lecurve hanno dei picchi alle lunghezzed'onda di 450, 525 e 555 nanometri(rispettivamente nelle regioni azzurra,verde e gialla dello spettro visibile). Laattivazione con la luce porta alla stessasequenza di trasformazioni molecolari
descritta per la rodopsina. Le associa-zioni dei nervi con coni e bastoncellipermettono di fissare i dettagli sullaretina, o di registrarli, per la trasmis-sione dei segnali visivi; i tre tipi dicono assorbono in maniera diversa icolori emessi dall'immagine, permet-tendo cosí la visione cromatica.
La risposta delle piante alle variazio-ni di lunghezza del giorno e della
notte consiste in variazioni molecolari,provocate dalla luce, molto simili aquelle che si verificano per la visione.I fenomeni legati alla fotoperiodicitànon sono però conosciuti cosí bene co-
me quelli della visione, perciò li presen-teremo tramite alcuni esempi illustra-tivi. Il crisantemo e molte altre pianterispondono con la fioritura alla cre-scente lunghezza delle notti quando siapprossima la stagione umida. Le pian-te non fioriscono se vengono artificial-mente interrotti i lunghi periodi not-turni mediante breve esposizione allaluce verso mezzanotte. La luce rossa,con un massimo d'assorbimento a unalunghezza d'onda di 660 nanometri, èmolto efficace per prevenire la fioritu-ra. Il pigmento recettore della luce nel-le piante è quindi azzurro, colore com-plementare di quello rosso assorbito.Le piante fioriscono se dopo l'esposi-zione alla luce rossa vengono sottopo-ste a radiazioni di lunghezza d'onda allimite della visibilità, nel rosso lontano(730 nanometri).
Il comportamento della pianta orna-mentale chiamata kalanchoe illustrachiaramente la reversibilità della rea-zione. La luce rossa evidentemente tra-sforma il pigmento fotoreversibile nel-la forma che assorbe il rosso lontano.La pianta diviene incapace di fiorire,perché la luce del rosso lontano fa tor-nare il pigmento alla forma capace diassorbire il rosso, la quale impedisceappunto la prosecuzione della fioritura.Il controllo della fioritura tramite va-riazioni artificialmente indotte dellalunghezza della notte è un fattore mol-to importante per determinare quali va-rietà di soia, frumento o altre colturecommerciali, sia meglio utilizzare peravere buoni raccolti alle varie latitudi-ni con periodi diversi di luce e di buio.
Molti tipi di semi germinano solo seè stato attivato il pigmento fotorever-sibi!e. I semi di alcuni pini e lattughe,per esempio, non germinano in labora-torio se non sono stati esposti breve-mente alla luce rossa (o meglio a ra-diazioni luminose contenenti luce ros-sa). Se si fa seguire, alla attivazionecon luce rossa dei semi, una breve espo-sizione alle radiazioni del rosso lonta-no prima di riportarli al buio, essi ri-mangono allo stato di quiescenza. Il ci-clo di attivazione e disattivazione puòripetersi più volte; la germinazione olo stato di quiescenza dipendono daltipo dell'ultima esposizione della se-quenza descritta.
Il fabbisogno di luce per la germina-zione dei semi è responsabile della per-sistenza di erbacce nei campi coltivatia foraggio. Un seme che è nello statodi quiescenza, quando cade sul terreno,viene di solito coperto da uno strato diterra nel corso dell'inverno. Mentre ilseme è sepolto, il pigmento che con-trolla la germinazione si trasforma neltipo capace di assorbire la luce rossa;ora, il seme non germina fino a quando
La retina di un gatto, in sezione, ingrandita 670 volte. Lo strato di fibre nervose lasinistra) è quella parte della retina in contatto con il corpo vitreo dell'occhio. Primadi raggiungere i coni e i bastoncelli (a destra), la luce deve attraversare quest'ultimoe altri cinque strati di tessuto retinico. La microfotografia è dovuta a A. J. Ladman.
Questa fotografia al microscopio elettronico mostra i bastoncelli e i coni dell'occhioumano ingranditi 7200 volte. I pigmenti della visione sono concentrati in strati mem-branosi chiamati lamelle. La microfotografia è stata eseguita da Toichiro Kuwabara.
RODOPSINA
Le trasformazioni della rodopsina, il pigmento della vista deivertebrati, possono essere seguite in laboratorio a basse tem.perature. Compaiono quattro forme di pigmento (rettangoli),
11711
695 NANOMETR I
> — 70 °C
710 NANOMETRI
> — 35 .0
Anche le trasformazioni del fitocromo, il pigmento fotorever-sibile delle piante che interviene nella fotoperiodicità, possonoessere seguite a basse temperature. Come per la rodopsina,appaiono forme intermedie, con picchi di assorbimento della
\4'\i i 1i~
OPSINA OPSINA
LUMI-RODOPSINA
META-RODOPSINA I
META-RODOPSINA
TRANS-RETINENE+ OPSINA<
497 NANOMETRI 478 NANOMETRI 380 NANOMETRI 387 NANOMETRI> — 4000 > —15 OC > 000
C/S-RETINENE
Einik
OPSINA
RIDUZIONE - OSSIDAZIONEC/S-RETINENE+ OPSINA
quando la molecola di retinene (in nero) passa alla configura.zione trans e quindi si dissocia dall'opsina. Quando ritornaalla forma di cis-retinene, si ricombina e completa il ciclo.
710-740 NANOMETRI
> 000
C!7]FITOCROMO
(ROSSO LONTANO)
730 NANOMETRI
ROSSOLONTANO
luce caratteristici, prima che la forma iniziale del pigmento,che assorbe il rosso, risulti trasformata in quella che assor-be nel rosso lontano. Diversamente dal retinene il cromoforo(in nero) rimane associato alla proteina durante l'intero ciclo.
62 63
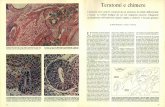
400 450 500 550
600
650
LUNGHEZZA D'ONDA (NANOMETRI)
La percezione del colore nell'uomo deriva dalla combinazione del retinene con trediverse opsine nei coni. I tre diversi pigmenti di iodopsina che si formano assorbonola maggior parte della luce visibile a tre lunghezze d'onda distinte. Le differenze trai segnali provenienti da ciascun gruppo di coni riflettono i colori dell'immagine.
AZZURRI° VERDEí \
%
n'I
/
/
/i
n•I
I%\
I1i,
I%\
\
n %%
%%\
\
\
s
,n
n
100
o
75
50
25
non viene esposto ancora alla luce so-lare tramite le pratiche di coltivazione.Quando avviene l'esposizione, la lucedel Sole ritrasforma alcuni dei pigmen-ti che assorbono il rosso in altri cheassorbono il rosso lontano, e inizia lagerminazione. È noto che i semi diuna comune erbaccia, il farinaccio sel-vatico, rimasti sepolti per 1700 anni,hanno poi dato luogo a germinazionequando furono esposti alla luce.
L'attivazione del pigmento fotorever-sibile controlla anche la crescita deglia'beri e di molte delle comuni pianteda fiori. Affinché queste piante cresca-no continuamente, devono essere sot-toposte per lunghi periodi alla lucediurna. Quando i giorni s'accorciano,l'accrescimento si ferma e i germogliritornano a quello stato di quiescenzache li protegge dalle basse temperatureinvernali.
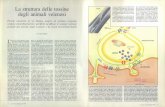
Il pigmento fotoreversibile delle pian-te è stato chiamato fitocromo. Pur es-sendo invisibile nei tessuti delle piantea causa della sua bassa concentrazione,è stato isolato con metodi largamenteusati nella preparazione di enzimi e al-tre proteine. Il pigmento è azzurro(si veda la figura nella pagina a fronte)e la sua fotoreversibilità è esattamentequella che ci si può attendere sulla ba-se della reazione della pianta allo sti-molo della luce.
64
La struttura chimica della molecoladi fitocromo dimostra che essa ha
una certa relazione con i pigmenti gial-lo-verdi della bile umana e con i pig-menti azzurri delle alghe azzurre. Lamolecola è infatti formata da un grup-po aperto di atomi, strettamente simileagli anelli delle molecole di clorofilla.Essa ha due gruppi laterali che possonopassare dalla forma cis a quella transper l'eccitazione luminosa. La trasfor-mazione più probabile dovuta a questaeccitazione, tuttavia, è un cambiamentodi posizione degli atomi di idrogenodella molecola.
Le variazioni che si verificano nellamolecola di fitocromo tramite eccita-zione ottenuta con un lampo di lucesono simili a quelle che si hanno nellarodopsina. La prima risposta all'eccita-zione ha luogo in pochi milionesimi disecondo e si ottiene una forma mole-
colare analoga a quella della prelumi-rodopsina. La trasformazione si fermaa questo punto se la temperatura è mi-nore di —110 °C. A questa temperaturale molecole possono essere ritrasfor-mate nella forma iniziale rossorecet-trice per azione della luce. A tempera-ture maggiori di —110°C si formanomolti più fitocromi di forma interme-dia prima che compaia la forma finaleatta ad assorbire radiazioni nel rossolontano. Questi stadi intermedi com-prendono anche alterazioni della for-ma molecolare della proteina associataal fitocromo, proprio come succede perl'opsina, che è la proteina della rodop-sina. Nella sua forma definitiva il fito-cromo è diverso dalla rodopsina inquanto la sua molecola, anziché disso-ciarsi, rimane legata alla proteina. Leradiazioni del rosso lontano possonoritrasformare la forma del fitocromoin quella che assorbe nel rosso, anchese si formano serie differenti di formemolecolari durante la riconversione.
Dopo l'eccitazione del fitocromo av-vengono lentamente sia la fioritura siala germinazione dei semi e molte altremanifestazioni della pianta. Diversa-mente da quanto si verifica nel feno-meno della visione, in cui la rispostasegue subito la rapida comparsa di mo-lecole intermedie, la risposta alla foto-periodicità dipende dalla presenza del-la forma finale del fitocromo, quellache assorbe le radiazioni del rosso lon-tano. Si conosce poco circa il modo incui le molecole dello stadio finale ese-guono questo tipo di lavoro. Una delleipotesi è quella che esse regolino laproduzione di enzimi controllando ilmateriale genetico nel nucleo della cel-lula. Un'altra ipotesi è quella per cuila maggiore solubilità nei lipidi rendapossibile un'unione delle molecole allemembrane della cellula, cioè alle pa-reti e alla membrana del nucleo. Unatrasformazione della molecola di fito-cromo influenzerebbe poi la permeabi-lità delle membrane e perciò anche ilfunzionamento della cellula.
L'esposizione delle piante a radiazio-ni corrispondenti alla lunghezza d'ondadell'azzurro e del lontano rosso nellospettro visibile contrasta l'azione dellaforma molecolare di fitocromo che as-
Il pigmento sensibile alla luce che regola le reazioni fotoperiodiche delle piante è visi.bile nella fotografia della pagina a fronte nei suoi due stati fondamentali. Questo pig-mento, chiamato fitocromo, viene studiato durante le ricorrenze stagionali, come la fio.ritura delle piante e la germinazione dei semi. Nel primo stato (a sinistra), il fitocromoè eccitabile tramite radiazioni del rosso lontano, nell'altro (a destra) tramite radiazionidi luce rossa. Alternando l'esposizione a questi due tipi di radiazioni il pigmento passareversibilmente dall'uno all'altro stato. Il fitocromo qui riportato è stato estratto dapianticelle di avena nel laboratorio di F. E. Mumford e E. L. Jenner del Centro di ri.cerca della Du Pont de Nemours. Il pigmento è contenuto in una cella quadrata diquarzo, adatta agli studi di assorbimento di luce, in soluzione allo 0,2 %. I numeriin trasparenza in alto nella cella indicano la lunghezza del percorso della luce: I cm.

PERIODI DIURNI
I I UCCELLI NORMALI
6 ORE 16 ORE500
UCCELLI CIECHI426
404
400
Nei passeri, la fotoperiodicità influenza anche altri fotorecettori oltre a quelli del-l'occhio. Il peso dei testicoli, infatti, sia nei passeri normali che in quelli ciechi (a si-nistra) rimane basso quando le gabbie vengono illuminate artificialmente per due mesial fine di simulare periodi diurni molto corti e lunghi intervalli notturni. Quando siinverte il trattamento, sottoponendo i passeri per due mesi a lunghi periodi diurni.il peso dei testicoli cresce allo stesso modo nei due casi (a destra). Questo esperimentoè stato realizzato da Michael Menaker e Henry Keatts della Università del Texas.
sorbe nel rosso lontano. Può darsi chel'eccitazione provocata dalla luce cor-rispondente alla lunghezza d'onda delrosso lontano provochi un continuo di-stacco, dalle membrane della cellula,delle molecole capaci di assorbire que-ste radiazioni. Un'eccitazione continuadi questo genere è quella che si verifi-ca, per esempio, durante i lunghi pe-riodi di esposizione alla luce che hannoun'influenza tanto spiccata sull'accresci-mento degli abeti Douglas. Gli alberidi questa specie rimangono nello statodi quiescenza, se vengono esposti allaluce diurna per 12 ore e per un ugualtempo alla luce notturna; mentre si ac-crescono regolarmente se aumenta ilperiodo di esposizione diurna.
fenomeno della fotoperiodicitànon è tipico delle piante : anche glianimali reagiscono alle variazioni dilunghezza del giorno e della notte. Lamigrazione e la riproduzione di moltespecie di uccelli, i cicli di attività dinumerosi mammiferi e le diapause (pe-riodi di inanimazione temporanea o diletargo) degli insetti sono controllate inquesto modo. Questi esempi di fotope-riodicità (e alcune reazioni meno netteche hanno luogo anche nell'uomo) di-pendono dall'azione di diversi ormoni
che lavorano in sequenza. Tali sequen-ze di azioni ormoniche possono avereun ritmo regolare. Esse costituisconouna base del cosiddetto ritmo circadia-no (cioè giornaliero) degli « orologi bio-logici ». Il ciclo di 24 ore di tali oro-logi è stabilito dalla luce.
Le diapause degli insetti sono fruttodell'azione reciproca di un ormone edella luce. Alcuni bachi da seta e lalarva del bruco della mela, per esem-pio, quando il periodo diurno è breve,passano nello stato di quiescenza. Inquesto stato, che aiuta gli insetti a so-pravvivere durante l'inverno, viene so-spesa, da parte di un gruppo di celluledella regione cerebrale centrale, la se-crezione di un ormone. L'ormone bloc-cato è il primo di una serie che portaall'ormone finale, l'ecdisone, che con-trolla la metamorfosi della pupa in unbruco adulto. Quando le cellule cere-brali della pupa quiescente vengonoesposte alla luce solare per molte ore,l'ormone del cervello viene liberato ela metamorfosi ha inizio. L'ecdisoneiniettato in una pupa in quiescenzaporta alla metamorfosi anche se i pe-riodi diurni sono brevi.
In questo caso l'unica azione dellaluce è quella diretta a favorire la se-
crezione di un ormone del cervello. Ilpigmento delle cellule cerebrali che as-sorbe la luce non è stato ancora iden-tificato, ma vi sono ricerche in atto daparte del Dipartimento dell'agricolturadegli Stati Uniti, appunto sulle reazionidei bruchi della mela alla luce, che so-no molto promettenti, per cui si speradi dare presto una risposta a questoquesito. Le regioni azzurro-verdi dellospettro (tra 500 e 560 nanometri), eprobabilmente ancor meglio lunghezzed'onda minori, sembrano essere moltoefficaci per interrompere la diapausa.Con una certa probabilità il pigmentoè del tipo della porfirina, con unastruttura centrale simile a quella adanello della clorofilla.
La dipendenza dell'uomo dall'orolo-gio biologico si manifesta con il disa-gio che egli sente quando la relazionetra il suo ritmo circadiano e il realeciclo diurno e notturno viene rapida-mente variata, come quando viaggia inaereo per distanze molto elevate, peresempio di molti gradi di longitudine.In questo caso i suoi controlli ormonicivengono disturbati o sfasati. I topi daipiedi bianchi e altri piccoli mammiferihanno periodi ciclici di attività che sem-brano essere regolati dalla luce.
In una prima fase i cicli di attivitàvengono influenzati da ormoni secretidalla regione ipotalamica del cervello.Non si sa se questa regione cerebralecontenga il pigmento recettore dellepiccole quantità di luce che possono pe-netrare nel cervello, oppure se essa ven-ga stimolata da un segnale provenien-te dall'occhio, o comunque dalla re-gione oculare. L'ipotalamo controlla laipofisi, che secerne ormoni che a lorovolta controllano le gonadi, la cortec-cia delle ghiandole surrenali e altri or-gani di questo tipo. Al momento attua-le, comunque, rimane ancora da stabi-lire sia l'esistenza di un pigmento re-sponsabile delle manifestazioni fotope-riodiche per i vertebrati, sia la sua po-sizione anatomica, sia il tipo di azio-ne che esplica a livello molecolare.
Eccitando un cromoforo, la luce agi-sce come stimolo sia nel caso della
fotorecezione che in quella della foto-periodicità, dando inizio a processi chedipendono, dal punto di vista energe-tico, dal metabolismo proprio dell'or-ganismo. Nel terzo importante campod'interazione tra luce e materia vivente— la fotosintesi — si verifica esattamen-te l'opposto: l'energia luminosa vieneutilizzata per produrre gli alimenti ne-cessari alla vita. Affinché questo acca-da vi devono essere : 1) un sistema re-cettore della luce, 2) un sistema per tra-sferire 1' energia tra le molecole e 3)
Porzione di bastoncello dell'occhio umano ingrandita 44 000 riempita dalle lamelle; quella pii interna (a destra) ha strut-volte al microscopio elettronico. La zona esterna (a sinistra) è tura complessa. Anche questa microfotografia è di T. Kuwabara.
Porzione di un cono dell'occhio umano, ingrandita 44 000 volte. Le lamelle sono differenti da quelle dei bastoncelli, in quantoSi notano le differenze tra la struttura interna e quella esterna. sono « impacchettate », alcune singolarmente, alcune a gruppi.
66 67

L'organo fotorecettore di un invertebrato (in questo caso di un granchio marino quiingrandito 8000 volte al microscopio elettronico) viene chiamato ommatidio e è unadelle 1000 unità di questo tipo che si trovano nell'occhio dell'animale. I fotorecet.tori si dipartono dal centro con struttura raggiata. Anche l'anello centrale è foto.sensibile. La foto è di William H. Miller della Yale University School of Medicine.
Lunghi periodi diurni influenzano l'accrescimento dell'abeteDouglas. Esposto a corti periodi diurni, o a tempi uguali diluce e buio, l'albero rimane nello stato di quiescenza (a sini-stra). L'eccitazione prodotta da ulteriori radiazioni provoca un
accrescimento. Un albero (ai centro) ha ricevuto un'ora di illu.minazione a bassa intensità nel periodo notturno di 12 ore; perun altro albero (a destra) il periodo diurno di 12 ore è sta•to allungato con otto ore di illuminazione a bassa intensità.
meccanismi per accoppiare l'energialuminosa alle trasformazioni chimiche.Le molecole di clorofilla (o meglio /emolecole di diversi tipi di clorofilla, chedifferiscono tra loro per i gruppi late-rali) costituiscono il principale sistemafotorecettore. Un elettrone che si trovanella molecola della clorofilla, tramitel'azione della luce visibile, viene ecci-tato a livelli energetici più alti. Questoelettrone ritorna al livello iniziale inmeno di un centomilionesimo di secon-do. Tale ritorno potrebbe produrre unariemissione della luce visibile assorbitama, ovviamente, in tal caso non si ot-terrebbe nessun vantaggio dal processofotosintetico. Il ritorno allo stato ini-ziale procede, invece, a gradini, duran-te ognuno dei quali l'energia necessariaper la fotosintesi viene trasferita lungouna catena di molecole. L'energia rice-vuta viene cosí resa solo in parte comeluce di lunghezza d'onda più alta, equindi di energia minore di quella del-
la luce assorbita. (Questa luce emessaè il rosso scuro, emissione caratteristi-ca della clorofilla quando viene eccita-ta alla fluorescenza.) L'energia vienetrasferita, tramite molecole di clorofil-la, a un recettore finale, costituito dauna molecola che utilizza questa ener-gia per le sintesi chimiche. Le molecoleche servono da veicolo per il trasferi-mento d'energia, in primo luogo devonotrovarsi fisicamente molto vicine l'unaall'altra, e poi devono costituire uncollegamento adeguato tra l'energia di-sponibile e quella che può essere as-sorbita.
I plastidi delle cellule delle piante,organelli contenenti i pigmenti di clo-rofilla, sono formati da strutture stra-tificate, chiamate lamelle, costituite pre-valentemente da grassi e proteine (siveda la figura in alto a pagina 70). Lamolecola di clorofilla ha una porzione(chiamata fitolo) che è solubile nei gras-si, e un corpo principale (clorofillina)
che ha invece affinità per le proteine.Queste proprietà conducono a un siste-ma strutturale in cui le molecole diclorofilla risultano strettamente impac-chettate.
Le lamelle contengono anche altremolecole provviste di legami coniugati.Tra queste vi sono dei carotenoidi chehanno una struttura simile al retinene,e la fitocianina, che è un cromoforoassai simile a quello del fitocromo. Que-ste molecole di contorno assorbonoanch'esse l'energia per trasferirla allemolecole di clorofilla.
L'energia accumulata viene infine por-tata dalla clorofilla a molecole re-
lativamente piccole che funzionano da« deposito ». In una lamella vi è unadi queste molecole ogni 500 molecoledi clorofilla. Questo basso numero di« depositi », mentre realizza una ap-prezzabile economia nel sistema a« gradini chimici » per la fotosintesi,costituisce ovviamente una strettoia,una specie di collo di bottiglia, perquanto riguarda i trasferimenti d'ener-gia. Quando la luce raggiunge un li-vello d'intensità di circa un quinto diquello della luce solare, l'energia giun-ge alle molecole più velocemente diquanto non possa essere utilizzata. Lasaturazione a questo livello d'intensitànon è nemmeno un buon compromes-so poiché, mediamente, la foglia di unapianta è un poco in ombra e raramen-te riceve una quantità d'energia moltoal di sopra del livello di un quinto.
L'energia accumulata nelle molecoleviene infine utilizzata per convertire lemolecole d'acqua in idrogeno e ossi-geno, e per produrre un composto ric-co di elettroni, che chiameremo e X »,il quale funziona da recettore finaledegli elettroni. Si forma come sottopro-dotto non utilizzabile un materiale os-sidato (cioè che ha ceduto elettroni),che nel caso delle piante verdi è ossi-geno. L'atmosfera terrestre, infatti,contiene l'ossigeno necessario alla vitadegli animali appunto come sottopro-dotto della fotosintesi.
La resa del processo chimico foto-sintetico diminuisce al crescere dellalunghezza d'onda delle radiazioni lu-minose assorbite. L'assorbimento nelrosso lontano dello spettro visibile, tut-tavia, può divenire efficace se sonopresenti radiazioni supplementari dilunghezza d'onda minore. Questo com-portamento può suggerire l'ipotesi del-la presenza di due stadi, anziché uno,nel fenomeno di trasferimento deglielettroni; forse ci sono due depositi dienergia che lavorano sinergicamente.Sono oggetto di ricerche attuali sia iprocessi associati all'accumulo di ener-gia, sia il modo in cui può essere coor-
dinato il flusso elettronico tra i due sta-di suddetti.
Possiamo fare una rappresentazionedel sistema di flusso elettronico, secon-do il modo attuale di vedere le cose(si vedano le figure a pagina 70).
I centri-trappole sono indicati conle lettere E e P. Con un primo assor-bimento di luce si pensa che un elet-trone sia trasferito da P a X (b): conla perdita di un elettrone P rimane os-sidato mentre X, che lo ha ricevuto, di-viene un composto ridotto, cioè ricco
di elettroni. Il secondo assorbimento diluce (a), realizza il trasferimento di unelettrone dall'acqua al punto E, ren-dendo E ricco di elettroni e ottenendo,dalle piante verdi, ossigeno libero comesostanza ossidata. Lo schema vienecompletato da un trasferimento elettro-nico dalla forma ridotta E a quella os-sidata P. Affinché si realizzino le fasidi trasporto elettronico dall'acqua aX, è necessaria anche una stretta par-tecipazione di porzioni delle lamelledei plastidi, ricche in lipidi.
Questo schema semplice può dareun'idea degli elementi interessati allafotosintesi, per quanto riguarda la par-te « foto », ma rimane molto da diresulla parte « sintesi ». La « sintesi »comprende un'estrazione che si puòutilizzare nei processi vitali. L'ossigeno,benché sia un prodotto ossidato, puòinfine ritornare al composto ridotto Xtramite la respirazione. X è un pro-dotto immediatamente utilizzabile, perreazioni esterne alle lamelle. Esso ser-ve come agente principale del trasferi-
68 69
telecomunicazionielettronica
commutazione e trasmissione telefonica e telegraficatrasmissione su filo
e via radio, anche tramite satelliti artificialitrasmissione dati, immagini e per teleoperazioni
elettroacusticaimpianti trasmittenti radiotelevisivitelefoni e citofoni
segnalamento ferroviario - segnalazione e allarme
SOCIETA' ITALIANA TELECOMUNICAZIONISIEMENS s.p.a. GRUPPO ST
Sede, direzione gen'
Sede di rappresentanze e stSicilia 111- 50135 Firenzevia Medina 40 - 10134 Torin
LUCE
X RIDOTTO
CLOROFILLAX OSSIDATO
T T -TL
T T 7' T CAROTENOIDE
ARIDOTTO
X + ++ OSSIDATO
PRIDOTTO
OSSIGENOLIBERO
X + + RIDOTTO
per il trasferimento d'energia, che comprende due « depositi »energetici, E e P (in colore, al centro), legati da un sistemache provvede al trasferimento elettronico, costituito dalle uni-tà A. Il sistema di trasferimento tramite il quale questo rag-gruppamento molecolare trasforma l'energia luminosa n frecce in-terne) in energia chimica (frecce esterne) è mostrato qui sotto.
ATP
ERIDOTTO
POSSIDATO
altre molecole di clorofilla eccitate dalle luce ( b). Il trasferi-mento termina quando P ritorna al livello energetico più basso,cedendo sia l'elettrone sia l'energia a X, un composto riccodi elettroni. Ciò porta X a un livello energetico tale da consen-tirgli di attivare il processo di riduzione dell'anidride carbo-nica (freccia in basso a destra); questa è la principale azionefotosintetica. Altri due eventi si hanno come conseguenza delpassaggio di elettroni. Ioni idrogeno (freccia in alto) forni-scono il gradiente d'energia necessario per trasformare l'adeno-sindifosfato (ADP) in adenosintrifosfato (ATP). In modo si»mile lo ione neutralizzato dalla perdita elettronica che iniziail processo si congiunge con altri componenti per formareacqua e liberando perciò l'ossigeno (freccia in alto a destra).
LUCE ADP —A-\> ATP
T
PROTEINA
CLOROFILLA
ACQUA
T TCAROTENOIDE
I fotorecettori delle piante sono formati da molecole schieratein ordine particolare. Dentro il cloroplasto le molecole di clo-rofilla sono unite sia per attrazione, sia per l'affinità della
coda » di fitolo per i lipidi e del corpo per le proteine. Inquesta struttura troviamo anche altri pigmenti, come i carote-noidi. Ogni 500 molecole circa di clorofilla si ha un centro
ADP
a21-9 -n 4e- + 4H + + 02
LUCEe
ENERGIA
EOSSIDATO
AOSSIDATO
I)
eLUCE
ENERGIA
••••
Rappresentazione schematica del trasferimento elettronico < agradini » responsabile della fotosintesi, processo che consistenell'utilizzazione dell'energia luminosa per ridurre l'anidridecarbonica a zucchero. Entro la zona che contiene i due depo-siti energetici E e P, l'acqua e i suoi componenti (rettangoloin alto) sono in uno stato di equilibrio finché parecchie mole-cole di clorofilla non trasferiscono l'energia ricevuta dalla luce(a) al primo deposito d'energia E. Questo, portato dall'energiaricevuta a un più alto stato eccitato, prende un elettrone (frec-cia colorata) dall'acqua, poi ritorna al livello energetico piùbasso; l'elettrone catturato viene liberato e passa attraverso ilsistema di trasferimento A. L'elettrone giunge al secondo depo-sito energetico, P, poco dopo che esso ha ricevuto energia da
mento d'energia nella riduzione dellaanidride carbonica a zucchero. Un'ul-teriore reazione trasforma X, median-te passaggi intermedi, e unitamente adanidride carbonica e acqua, in un Xpovero di elettroni, od ossidato, e inzuccheri contenenti fosforo. La reazionenecessita di energia maggiore di quellache il solo X può fornire. Questa ener-gia proviene dall'adenosintrifosfato(ATP). L'ATP si forma quando l'ade-nosindifosfato (ADP) perde acqua ereagisce con una molecola di fosfato.L'energia per la trasformazione di X sirende disponibile quando il fosfato chesi era addizionato all'ATP viene tra-sferito a qualche altra molecola o sisepara dall'ATP per idrolisi.
Ritornando alla nostra rappresenta-zione schematica, sembra che la quan-tità di energia ceduta nel trasferimen-to di elettroni da E a P sia sufficienteper ottenere ATP. Fino a tempi recentisi è pensato che questo fosse il metodopiù probabilmente utilizzato da almenoun gruppo di piante per ottenere ATP.Un'ipotesi che viene attualmente moltodiscussa è quella della comparsa di ioniidrogeno dentro le lamelle, durante iltrasferimento di elettroni che si ha do-po l'assorbimento della luce. Un gra-diente energetico adatto alla formazio-ne di ATP sarebbe, in questo caso, rea-lizzato dalla spiccata acidità di origineionica all'interno della membrana, ri-spetto all'ambiente esterno alla mem-brana stessa. Riguardo alla prima azio-ne della luce (b), gli elettroni eccitatida questi fenomeni possono anche ri-tornare attraverso X al punto di par-potenza P, accoppiandosi ad ATP lungoil cammino percorso, in un processoconosciuto come fosforilazione ciclica.
uesti aspetti del trasferimento dienergia nella fotosintesi sono stati
sviluppati principalmente negli ultimidieci anni. Vi è ancora molto da im-parare circa i dettagli a livello moleco-lare che riguardano la liberazione diossigeno, la formazione di ATP e ilcoordinamento del flusso elettroniconelle varie parti del processo. Nuovescoperte possono cambiare gli odierniconcetti sulla fotosintesi anche a livel-lo dei principi fondamentali. La situa-zione rispetto alle conoscenze attuali èpraticamente la stessa anche per quan-to riguarda la visione e la fotoperiodi-cità. L'esame delle trasformazioni chesi hanno immediatamente dopo l'assor-bimento della luce ha provato che que-sto è un campo di studio più fecondodi quel:o relativo alle ricerche sugli sta-di successivi, che riguardano la visione,l'accrescimento e il ritmo biologico.
OSSIGENO
A
T TLIPIDI
CLOROFILLA
CLOROFILLA
70
71