La membrana purpurea dei batteri alofili -...
5
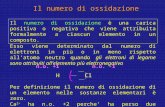
Particelle di batteriorodopsina, la proteina della membrana purpurea di IL halobium, sono ingrandite di 87 000 volte in questa microfoto- grafia elettronica di una cellula, ottenuta mediante criodecapaggio. La membrana, prima congelata e poi rotta, si è divisa lungo il piano centrale del doppio strato lipidico. Poiché la cellula è in pratica un cilindro con estremità emisferiche, le facce di frattura convesse appar- tengono alla porzione interna della membrana interna, o citopla- smatica e le facce concave alla porzione della membrana esterna. Le proteine immerse nella membrana appaiono sulle facce di frattura co- me particelle. Le regioni con struttura grossolana appartengono alla porzione principale della membrana cellulare, la cosiddetta membrana rossa. Le regioni a grana fine, invece, con un disegno percettibilmente esagonale, sono ammassi di membrana color porpora. Esse si trovano sulle facce convesse: le regioni corrispondenti sulle facce concave risul- tano essere lisce, completamente prive di particelle. La batteriorodop- sina rimane immersa, dopo la frattura, nella porzione citoplasmatica. In questa microfotografia elettronica, una sospensione di membrana color porpora appare ingrandita 67 000 volte. La sospensione è stata spruzzata su una superficie e mentre la pellicola si prosciugava si sono formate delle crepe, che hanno evidenziato i piani del reticolo esagonale. I batteri alofili, qui ingranditi 1850 volte, sono cellule mobili a forma di bastoncello, i cui flagel- li non sono visibili. Nella microfotografia è ripreso il mutante Halobacterium halobium R,. L a vita sulla Terra è basata sulla foto- sintesi, il processo attraverso il quale l'energia irradiata dal Sole viene trasformata in modo da mantenere l'attività metabolica degli organismi vi- venti. Solo le piante, alcuni batteri e le alghe azzurre possono attuare la fotosin- tesi in quanto sono gli unici organismi a possedere la sostanza critica che capta l'energia luminosa: la clorofilla. L'assor- bimento di questa energia da parte della clorofilla avvia il trasferimento di elet- troni lungo una catena di altre molecole di pigmento, sempre legate a membrane, e l'energia che si libera nel corso del tra- sporto degli elettroni viene trasformata nei legami ad alto contenuto energetico dell'adenosintrifostato (ATP), il traspor- tatore primario di energia delle cellule viventi. In questo modo l'energia diviene disponibile per la sintesi di quella fonda- mentale riserva di energia chimica sulla Terra che sono le molecole di amido, di cellulosa e di ossigeno libero. Abbiamo scoperto di recente un altro meccanismo fotosintetico basato su un diverso pigmento, molto affine alla ro- dopsina, che era stata considerata fino ad allora solo come un pigmento visivo dell'occhio degli animali. Il suo sistema si trova negli alobatteri o batteri alofili, microrganismi che necessitano per la lo- ro crescita di elevate concentrazioni di cloruro di sodio e quindi proliferano nei laghi salati naturali e nelle saline, dove l'acqua di mare viene fatta evaporare per produrre il sale. Il pigmento e il sistema che da esso dipende trasformano l'ener- gia luminosa solare in energia chimica, di cui i batteri necessitano per il loro metabolismo e per mantenere la loro in- tegrità cellulare in un ambiente caratte- rizzato da basse concentrazioni di ossige- no, da un'elevata intensità luminosa e da una forte pressione osmotica. Nel 1964 mi imbattei in un lavoro in cui si diceva che i batteri alofili erano privi della resistente parete cellula- re esterna, che protegge la membrana della maggior parte dei batteri e, inoltre, che questa membrana si disintegrava, a basse concentrazioni di sale, in subunità identiche di lipoproteina. Da molti anni stavo studiando la struttura e la funzione delle membrane cellulari e questa, degli alofili, mi parve una strana membrana; inoltre, la sua composizione pareva con- traddire alcune delle mie idee fondamen- tali su come sono organizzate le mem- brane cellulari. Così, quando alla fine di quell'anno, Robert Rowen venne a la- vorare nel mio laboratorio della Rocke- feller University, demmo uno sguardo ai batteri alofili. Ne emerse che quel lavoro era in errore: i batteri non mancano af- fatto di una parete cellulare e in realtà non è la membrana lipoproteica, ma una parete cellulare glicoproteica, a fornire le subunità identiche. Andando alla ricerca dei dati originali, eseguimmo alcune nuo- ve e interessanti osservazioni e conti- nuammo così lo studio dei batteri alofili. Halobacterium halobium, la specie sulla quale soprattutto lavoriamo, è una cellula a forma di bastoncino, del diame- tro di mezzo micrometro e della lunghez- za di forse 5 micrometri. Cresce in ma- niera ottimale in una soluzione 4,3 mola- re (M) di cloruro di sodio, cioè in 4,3 moli, equivalenti a circa 250 grammi di sale per litro di soluzione (l'acqua di ma- re è normalmente una soluzione circa 0,6 M). Il batterio non vive a concentrazioni inferiori a 3,0 M. Oltre a elementi nutri- tivi come gli amminoacidi o i peptidi (costituenti delle proteine), il suo terreno colturale ha bisogno di piccole concen- trazioni di ioni di potassio, di magnesio e di calcio e di tracce di ferro e di man- ganese. Le sue cellule non metabolizzano gli zuccheri anaerobicamente e abbiamo ammesso che, in mancanza di una simile fermentazione, l'unica fonte di energia doveva essere la fosforilazione ossidati- va, processo grazie al quale gli animali e la massima parte dei batteri trasformano l'energia che si libera durante la respira- zione, per ossidazione di amminoacidi, acidi grassi e carboidrati nei legami ad alto contenuto energetico dell'ATP. Quando abbiamo sospeso delle cellule di H. halobium in una miscela salina di partenza (cioè il terreno colturale senza le sostanze nutritive) e abbiamo diluito questo mezzo per esporre i batteri a de- crescenti concentrazioni di sale, abbiamo La membrana purpurea dei batteri alofili Un pigmento molto simile alla rodopsina, la «porpora visiva» dell'occhio animale, è contenuto nella membrana dei batteri alofili e permette loro di trasformare l'energia solare in energia chimica di Walther Stoeckenius 26 27
-
Upload
trinhthuan -
Category
Documents
-
view
219 -
download
0
Transcript of La membrana purpurea dei batteri alofili -...
Particelle di batteriorodopsina, la proteina della membrana purpureadi IL halobium, sono ingrandite di 87 000 volte in questa microfoto-grafia elettronica di una cellula, ottenuta mediante criodecapaggio.La membrana, prima congelata e poi rotta, si è divisa lungo il pianocentrale del doppio strato lipidico. Poiché la cellula è in pratica uncilindro con estremità emisferiche, le facce di frattura convesse appar-tengono alla porzione interna della membrana interna, o citopla-smatica e le facce concave alla porzione della membrana esterna. Le
proteine immerse nella membrana appaiono sulle facce di frattura co-me particelle. Le regioni con struttura grossolana appartengono allaporzione principale della membrana cellulare, la cosiddetta membranarossa. Le regioni a grana fine, invece, con un disegno percettibilmenteesagonale, sono ammassi di membrana color porpora. Esse si trovanosulle facce convesse: le regioni corrispondenti sulle facce concave risul-tano essere lisce, completamente prive di particelle. La batteriorodop-sina rimane immersa, dopo la frattura, nella porzione citoplasmatica.
In questa microfotografia elettronica, una sospensione di membranacolor porpora appare ingrandita 67 000 volte. La sospensione è stata
spruzzata su una superficie e mentre la pellicola si prosciugava si sonoformate delle crepe, che hanno evidenziato i piani del reticolo esagonale.
I batteri alofili, qui ingranditi 1850 volte, sono cellule mobili a forma di bastoncello, i cui flagel-li non sono visibili. Nella microfotografia è ripreso il mutante Halobacterium halobium R,.
L
a vita sulla Terra è basata sulla foto-sintesi, il processo attraverso ilquale l'energia irradiata dal Sole
viene trasformata in modo da mantenerel'attività metabolica degli organismi vi-venti. Solo le piante, alcuni batteri e lealghe azzurre possono attuare la fotosin-tesi in quanto sono gli unici organismi apossedere la sostanza critica che captal'energia luminosa: la clorofilla. L'assor-bimento di questa energia da parte dellaclorofilla avvia il trasferimento di elet-troni lungo una catena di altre molecoledi pigmento, sempre legate a membrane,e l'energia che si libera nel corso del tra-sporto degli elettroni viene trasformatanei legami ad alto contenuto energeticodell'adenosintrifostato (ATP), il traspor-tatore primario di energia delle celluleviventi. In questo modo l'energia divienedisponibile per la sintesi di quella fonda-mentale riserva di energia chimica sullaTerra che sono le molecole di amido, dicellulosa e di ossigeno libero.
Abbiamo scoperto di recente un altromeccanismo fotosintetico basato su undiverso pigmento, molto affine alla ro-dopsina, che era stata considerata finoad allora solo come un pigmento visivo
dell'occhio degli animali. Il suo sistemasi trova negli alobatteri o batteri alofili,microrganismi che necessitano per la lo-ro crescita di elevate concentrazioni dicloruro di sodio e quindi proliferano neilaghi salati naturali e nelle saline, dovel'acqua di mare viene fatta evaporare perprodurre il sale. Il pigmento e il sistemache da esso dipende trasformano l'ener-gia luminosa solare in energia chimica,di cui i batteri necessitano per il lorometabolismo e per mantenere la loro in-tegrità cellulare in un ambiente caratte-rizzato da basse concentrazioni di ossige-no, da un'elevata intensità luminosa e dauna forte pressione osmotica.
Nel 1964 mi imbattei in un lavoro incui si diceva che i batteri alofili
erano privi della resistente parete cellula-re esterna, che protegge la membranadella maggior parte dei batteri e, inoltre,che questa membrana si disintegrava, abasse concentrazioni di sale, in subunitàidentiche di lipoproteina. Da molti annistavo studiando la struttura e la funzionedelle membrane cellulari e questa, deglialofili, mi parve una strana membrana;inoltre, la sua composizione pareva con-
traddire alcune delle mie idee fondamen-tali su come sono organizzate le mem-brane cellulari. Così, quando alla fine diquell'anno, Robert Rowen venne a la-vorare nel mio laboratorio della Rocke-feller University, demmo uno sguardo aibatteri alofili. Ne emerse che quel lavoroera in errore: i batteri non mancano af-fatto di una parete cellulare e in realtànon è la membrana lipoproteica, ma unaparete cellulare glicoproteica, a fornire lesubunità identiche. Andando alla ricercadei dati originali, eseguimmo alcune nuo-ve e interessanti osservazioni e conti-nuammo così lo studio dei batteri alofili.
Halobacterium halobium, la speciesulla quale soprattutto lavoriamo, è unacellula a forma di bastoncino, del diame-tro di mezzo micrometro e della lunghez-za di forse 5 micrometri. Cresce in ma-niera ottimale in una soluzione 4,3 mola-re (M) di cloruro di sodio, cioè in 4,3moli, equivalenti a circa 250 grammi disale per litro di soluzione (l'acqua di ma-re è normalmente una soluzione circa 0,6M). Il batterio non vive a concentrazioniinferiori a 3,0 M. Oltre a elementi nutri-tivi come gli amminoacidi o i peptidi(costituenti delle proteine), il suo terrenocolturale ha bisogno di piccole concen-trazioni di ioni di potassio, di magnesioe di calcio e di tracce di ferro e di man-ganese. Le sue cellule non metabolizzanogli zuccheri anaerobicamente e abbiamoammesso che, in mancanza di una similefermentazione, l'unica fonte di energiadoveva essere la fosforilazione ossidati-va, processo grazie al quale gli animali ela massima parte dei batteri trasformanol'energia che si libera durante la respira-zione, per ossidazione di amminoacidi,acidi grassi e carboidrati nei legami adalto contenuto energetico dell'ATP.
Quando abbiamo sospeso delle celluledi H. halobium in una miscela salina dipartenza (cioè il terreno colturale senzale sostanze nutritive) e abbiamo diluitoquesto mezzo per esporre i batteri a de-crescenti concentrazioni di sale, abbiamo
La membrana purpurea deibatteri alofili
Un pigmento molto simile alla rodopsina, la «porpora visiva»dell'occhio animale, è contenuto nella membrana dei batteri alofilie permette loro di trasformare l'energia solare in energia chimica
di Walther Stoeckenius
26
27
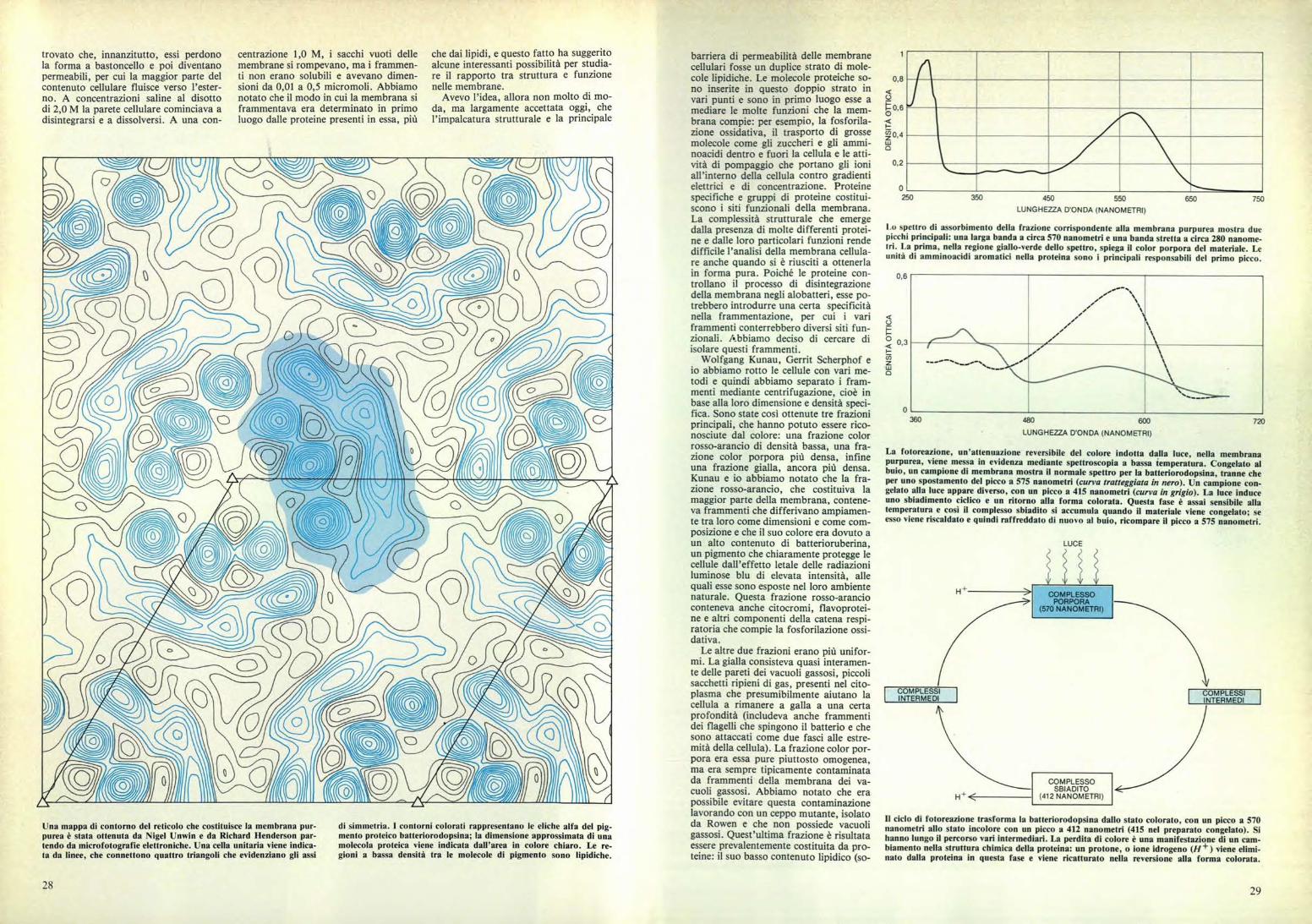
Una mappa di contorno del reticolo che costituisce la membrana pur-purea è stata ottenuta da Nigel Unwin e da Richard Henderson par-tendo da microfotografie elettroniche. Una cella unitaria viene indica-ta da linee, che connettono quattro triangoli che evidenziano gli assi
di simmetria. I contorni colorati rappresentano le eliche alfa del pig-mento proteico batteriorodopsina; la dimensione approssimata di unamolecola proteica viene indicata dall'area in colore chiaro. Le re-gioni a bassa densità tra le molecole di pigmento sono lipidiche.
1.—0,4z
o
0,8
o
0,6o
o250 350 450 550
650 750
LUNGHEZZA D'ONDA (NANOMETRI)
Lo spettro di assorbimento della frazione corrispondente alla membrana purpurea mostra duepicchi principali: una larga banda a circa 570 nanometri e una banda stretta a circa 280 nanome-tri. La prima, nella regione giallo-verde dello spettro, spiega il color porpora del materiale. Leunità di amminoacidi aromatici nella proteina sono i principali responsabili del primo picco.
0,2
0,6
„----,\\i
360
480 600
720
LUNGHEZZA D'ONDA (NANOMETRI)
La fotoreazione, un'attenuazione reversibile del colore indotta dalla luce, nella membranapurpurea, v iene messa in evidenza mediante spettroscopia a bassa temperatura. Congelato albuio, un campione di membrana mostra il normale spettro per la batteriorodopsina, tranne cheper uno spostamento del picco a 575 nanometri (curva tratteggiata in nero). Un campione con-gelato alla luce appare diverso, con un picco a 415 nanometri (curva in grigio). La luce induceuno sbiadimento ciclico e un ritorno alla forma colorata. Questa fase è assai sensibile allatemperatura e così il complesso sbiadito si accumula quando il materiale viene congelato; seesso viene riscaldato e quindi raffreddato di nuovo al buio, ricompare il picco a 575 nanometri.
LUCE
COMPLESSOPORPORA
(570 NANOMETRI)
COMPLESSOSBIADITO
(412 NANOMETRI)
COMPLESSIINTERMEDI
Il ciclo di fotoreazione trasforma la batteriorodopsina da lo stato colorato, con un picco a 570nanometri allo stato incolore con un picco a 412 nanometri (415 nel preparato congelato). Sihanno lungo il percorso vari intermediari. La perdita di colore è una manifestazione di un cam-biamento nella struttura chimica della proteina: un protone, o ione idrogeno ( H + ) viene elimi-nato dalla proteina in questa fase e viene ricatturato nella reversione alla forma colorata.
H+
COMPLESSIINTERMEDI
Fi+<
trovato che, innanzitutto, essi perdonola forma a bastoncello e poi diventanopermeabili, per cui la maggior parte delcontenuto cellulare fluisce verso l'ester-no. A concentrazioni saline al disottodi 2,0 M la parete cellulare cominciava adisintegrarsi e a dissolversi. A una con-
centrazione 1,0 M, i sacchi vuoti dellemembrane si rompevano, ma i frammen-ti non erano solubili e avevano dimen-sioni da 0,01 a 0,5 micromoli. Abbiamonotato che il modo in cui la membrana siframmentava era determinato in primoluogo dalle proteine presenti in essa, più
che dai lipidi, e questo fatto ha suggeritoalcune interessanti possibilità per studia-re il rapporto tra struttura e funzionenelle membrane.
Avevo l'idea, allora non molto di mo-da, ma largamente accettata oggi, chel'impalcatura strutturale e la principale
barriera di permeabilità delle membranecellulari fosse un duplice strato di mole-cole lipidiche. Le molecole proteiche so-no inserite in questo doppio strato invari punti e sono in primo luogo esse amediare le molte funzioni che la mem-brana compie: per esempio, la fosforila-zione ossidativa, il trasporto di grossemolecole come gli zuccheri e gli ammi-noacidi dentro e fuori la cellula e le atti-vità di pompaggio che portano gli ioniall'interno della cellula contro gradientielettrici e di concentrazione. Proteinespecifiche e gruppi di proteine costitui-scono i siti funzionali della membrana.La complessità strutturale che emergedalla presenza di molte differenti protei-ne e dalle loro particolari funzioni rendedifficile l'analisi della membrana cellula-re anche quando si è riusciti a ottenerlain forma pura. Poiché le proteine con-trollano il processo di disintegrazionedella membrana negli alobatteri, esse po-trebbero introdurre una certa specificitànella frammentazione, per cui i variframmenti conterrebbero diversi siti fun-zionali. Abbiamo deciso di cercare diisolare questi frammenti.
Wolfgang Kunau, Gerrit Scherphof eio abbiamo rotto le cellule con vari me-todi e quindi abbiamo separato i fram-menti mediante centrifugazione, cioè inbase alla loro dimensione e densità speci-fica. Sono state così ottenute tre frazioniprincipali, che hanno potuto essere rico-nosciute dal colore: una frazione colorrosso-arancio di densità bassa, una fra-zione color porpora più densa, infineuna frazione gialla, ancora più densa.Kunau e io abbiamo notato che la fra-zione rosso-arancio, che costituiva lamaggior parte della membrana, contene-va frammenti che differivano ampiamen-te tra loro come dimensioni e come com-posizione e che il suo colore era dovuto aun alto contenuto di batterioruberina,un pigmento che chiaramente protegge lecellule dall'effetto letale delle radiazioniluminose blu di elevata intensità, allequali esse sono esposte nel loro ambientenaturale. Questa frazione rosso-arancioconteneva anche citocromi, flavoprotei-ne e altri componenti della catena respi-ratoria che compie la fosforilazione ossi-dativa.
Le altre due frazioni erano più unifor-mi. La gialla consisteva quasi interamen-te delle pareti dei vacuoli gassosi, piccolisacchetti ripieni di gas, presenti nel cito-plasma che presumibilmente aiutano lacellula a rimanere a galla a una certaprofondità (includeva anche frammentidei flagelli che spingono il batterio e chesono attaccati come due fasci alle estre-mità della cellula). La frazione color por-pora era essa pure piuttosto omogenea,ma era sempre tipicamente contaminatada frammenti della membrana dei va-cuoli gassosi. Abbiamo notato che erapossibile evitare questa contaminazionelavorando con un ceppo mutante, isolatoda Rowen e che non possiede vacuoligassosi. Quest'ultima frazione è risultataessere prevalentemente costituita da pro-teine: il suo basso contenuto lipidico (so-
28 29

o 200
300TEMPO (SECONDI)
La luce influisce sul consumo di ossigeno dei batteri (in alto) e sulla acidità del mezzo (in basso).La pendenza della curva superiore è una misura della velocità del consumo di ossigeno. In unacamera chiusa, le cellule consumano rapidamente l'ossigeno al buio (aree ombreggiate); quandosi accende la luce (area bianca), c'è una diminuzione del 76 per cento in tale velocità (linea con-tinua); la velocità originaria si raggiunge di nuovo quando la luce viene spenta. La luce provocaanche un aumento dell'acidità del mezzo poiché i protoni sono riversati in esso dalle cellule.
100
6,4
6,3
6,2
6,1
60
75
50
25
• CARBONIO• AZOTO
O OSSIGENO
• IDROGENO
Il retinene, il cromoforo della batteriorodopsina, responsabile del suo assorbimento luminoso,è legato alla catena polipeptidica della proteina, come risulta da questo schema. Il retinene el'amminoacido (lisina) a cui esso si lega appaiono a sinistra. Quando si combinano tra loro, siha eliminazione di una molecola d'acqua, formatasi dall'atomo di ossigeno del gruppo termina-le aldeidico del retinene e da due atomi di idrogeno del gruppo amminico della lisina mentre tragli atomi di carbonio e di azoto si forma una base di Schiff. Essa perde e riacquista un protonedurante la fotoreazione, alternandosi tra due forme (a destra): protonata (1) e deprotonata (2).
lo il 25 per cento) spiega la sua elevatadensità. Studi di diffrazione dei raggi Xhanno permesso di ottenere diffratto-grammi ben definiti, con una risoluzioneelevata, il che implica che il materialedella membrana purpurea deve avere unastruttura che si avvicina, come regolari-tà, a quella dei cristalli proteici. Anche lemicrofotografie elettroniche hanno rive-lato una struttura regolare. Correlandole microfotografie del materiale che co-stituisce la frazione color porpora conmicrofotografie di membrane cellulari in-tere, abbiamo potuto stabilire, infine,che tale frazione non è affatto diffusa intutta la membrana, ma nella cellula inte-gra è presente come ammassi nell'internoe in collegamento con la membrana. Aquell'epoca, non conoscevamo ancora,quale potesse essere la funzione di talemembrana.
Nel 1967 mi trasferii dalla RockefellerUniversity alla Università di Cali-
fornia a San Francisco e soltanto nel
1969 con Allen E. Blaurock e DieterOesterhelt ripresi il lavoro sulla membra-na purpurea dei batteri. Blaurock analiz-zava la struttura della frazione nei parti-colari mediante diffrazione dei raggi X,mentre io raccoglievo dati complementa-ri di microscopia elettronica. Siamo riu-sciti così a discernere un reticolo esago-nale nel piano della membrana: la suacella unitaria, o più piccola unità ripeti-tiva, consisteva di 3 molecole di proteinae di 40 molecole di lipidi. Ciò concorda-va con il risultato ottenuto da Oesterhelt,secondo cui la membrana purpurea con-teneva soltanto una specie di proteinadal peso molecolare 26 000. Questa pro-teina sembrava possedere una elevatapercentuale di amminoacidi idrofobi, da-to che concordava con quelli ottenutimediante diffrazione dei raggi X i qualiindicavano che una buona parte di ognimolecola proteica deve essere sprofonda-ta nella regione più interna, idrofoba,della membrana.
Per molto tempo siamo rimasti per-
plessi di fronte alla peculiare pigmenta-zione della membrana purpurea. Gene-ralmente si determina la lunghezza d'on-da alla quale una sostanza biologica co-lorata assorbe la luce e si tenta quindi diestrarre e purificare il fattore responsa-bile del suo spettro di assorbimento. Unasospensione del materiale costitutivo del-la membrana ci aveva mostrato un'am-pia banda di assorbimento con un apicetra 560 e 570 nanometri (cioè nella zonagiallo-verde dello spettro visibile e unaforte punta di assorbimento, caratteristi-ca delle proteine, a 280 nanometri nellaregione ultravioletta. Ogniqualvolta, pe-rò, aggiungevamo un solvente o un de-tergente, nel tentativo di estrarre il cro-moforo (cioè il gruppo chimico respon-sabile dell'assorbimento luminoso e quin-di del colore), il colore sbiadiva.
È difficile, in retrospettiva, tracciarel'origine dell'idea che, alla fine, ha risol-to l'enigma. Può essere stato il fattodella decolorazione a opera dei solventio dei detergenti, o il fatto che la rodopsi-na sia stata nominata per caso in labora-torio, in quanto Blaurock, nel passato,aveva studiato la struttura delle mem-brane fotorecettrici dell'occhio, oppureil fatto che io stesso avevo notato una ri-sposta fototattica in H. halobium: quan-do l'intensità d'illuminazione diminuivanella zona rossa dello spettro, si verifica-va un'inversione nella direzione di nuotodi queste cellule batteriche. Qualunquesia stata la ragione, Oesterhelt decise divedere se il colore della membrana pur-purea non fosse dovuto per caso a unacromoproteina tipo rodopsina. Egli haseguito il procedimento stabilito per ana-lizzare la rodopsina e ha trovato che laproteina della membrana purpurea con-tiene un retinene per molecola. Il retine-ne, unito in complesso con varie protei-ne, od opsine, è il cromoforo di tutti ipigmenti visivi degli animali e si uniscein complesso con una pàrticolare opsinaper formare la rodopsina, il pigmentodei bastoncelli dell'occhio umano. Ab-biamo chiamato batteriorodopsina laproteina della membrana purpurea.
Il retinene è l'aldeide della vitaminaA. Nei pigmenti visivi è coniugato conuna proteina attraverso quella che è chia-mata una base di Schiff: un legame tra ilgruppo aldeidico (CHO) del retinene eun gruppo amminico (NI-1 2 ) di certe uni-tà amminoacidiche della proteina. Nellarodopsina, il pigmento visivo più noto, ilretinene, si lega in maniera specifica conil gruppo amminico di un'unità di lisina.Lo stesso avviene per la proteina dellamembrana purpurea (si veda la figura inquesta pagina). Il retinene non assorbenello spettro visibile; ha invece un'am-pia banda di assorbimento a circa 380nanometri e, nella forma legata con laproteina, dovrebbe assorbire a circa 370nanometri. D'altra parte, la principalebanda di assorbimento del materiale del-la membrana purpurea e della massimaparte dei pigmenti visivi è localizzata incorrispondenza di lunghezze d'onda mol-to più lunghe. Questo spostamento versoil rosso si spiega con le particolari condi-
zioni in cui il retinene forma un comples-so con la sua proteina specifica: gli am-minoacidi ai quali si lega formano appa-rentemente una specie di sacca idrofobache protegge la base di Schiff dall'am-biente acquoso (sia esso l'occhio o la so-spensione batterica), che altrimenti in-frangerebbe per idrolisi i legami dellabase di Schiff. Abbiamo notato che,quando questa speciale conformazionedella proteina viene distrutta da un sol-vente o da un detergente, il massimo diassorbimento si sposta da circa 570 a 370nanometri e, a questo punto, il retinenesi libera dalla proteina. Ciò spiega il no-stro insuccesso nell'estrarre un cromofo-ro con assorbimento a 570 nanometri e ilfatto che il colore sbiadisse.
T a similitudine della batteriorodopsinacon i pigmenti visivi ci ha subito
suggerito che la sua funzione nella cellu-la sia in relazione con le proprietà di as-sorbimento luminoso. Tale funzione po-teva essere quella di scatenare la rispostafototattica che avevamo osservato, maciò è parso improbabile per due ragioni.La prima era la grande quantità di mate-riale color porpora, sufficiente a costi-tuire fino al 50 per cento della membra-na superficiale della cellula; le rispostefototattiche si realizzano in altre cellulecon molto meno pigmento. La secondaera che la batteriorodopsina non sembra-va subire alcuna fotoreazione che potes-se determinare una risposta fototattica.Quando la rodopsina e altri pigmentivisivi vengono esposti alla luce, sbiadi-scono: perdono colore e il loro spettro diassorbimento si modifica. Il primo feno-meno è la prova della fotoreazione chescatena l'eccitamento visivo. La batterio-rodopsina, d'altra parte, non mostra al-cun cambiamento significativo nellospettro di assorbimento, quando vieneilluminata.
La forte concentrazione della membra-na purpurea nelle cellule mi ha fatto rite-nere che, se riuscivamo a trovare una fo-toreazione, era possibile che essa alimen-tasse qualche funzione metabolica piut-tosto che innescare una risposta senso-riale del tipo della fototassi. Un ruolosvolto nel metabolismo energetico do-vrebbe essere facilmente riconoscibilesotto forma di effetto sul contenuto diATP delle cellule; la luce potrebbe anchefornire un netto «risparmio» sull'altroprocesso di produzione di energia dellecellule, potrebbe cioè abbassare il livellodella respirazione. Lavorando prima conEva S.H. Kirsten e poi con Arlette Da-non, ho misurato la concentrazione diATP in cellule mantenute in varie condi-zioni di luminosità, ma gli esperimentihanno dato all'inizio risultati ambigui.D'altra parte, ho potuto presto convin-cermi che, esponendo le cellule alla luce,la respirazione veniva ridotta.
Alla fine, un'osservazione, che risalivaai nostri primi tentativi di estrazione, ci èservita come chiave per capire la foto-reazione della membrana purpurea e,quindi, la sua funzione. Oesterhelt avevanotato che, se si trattava una sospensio-
ne del materiale costituente la membranacon etere etilico, si otteneva uno sbiadi-mento reversibile man mano che l'etereevaporava. Anche se la luce non sembra-va esservi implicata, la reazione apparivainteressante. Così, un giorno, chiesi aMartin Brown, allora tecnico diplomatonel mio laboratorio, di ripetere l'esperi-mento con l'etere, ma sospendendo lamembrana purpurea in una soluzione sa-lina ad alta concentrazione, più vicinapossibile all'ambiente naturale, inveceche in acqua pura. Quasi subito egli notòche la membrana sbiadiva con l'eteresoltanto quando la sospensione venivailluminata con radiazioni dello spettrovisibile e che la reazione poteva essereinvertita mediante radiazioni provenientidalla regione dell'ultravioletto vicino.(Abbiamo trovato in seguito che l'ultra-
violetto non era indispensabile; la mem-brana sbiadita dalla luce ritornava al suocolore originario e alla sua condizione diassorbimento normale anche dopo esserestata posta per pochi secondi al buio).Avevamo così la prima dimostrazionedi una fotoreazione, anche se si trattavadi una fotoreazione che comportava lapartecipazione di un reagente chimicu.
Ho descritto la scoperta in una letteraa Oesterhelt, tornato all'Università diMonaco. Egli era in visita presso l'Isti-tuto Max Planck per la nutrizione diDortmund e, in quella sede, con BennoHess, il cui laboratorio era ben attrezza-to per simili studi, aveva cominciato aindagare sulla cinetica del cambiamentospettrale. L'apice di assorbimento a 412nanometri della membrana color porpo-ra, sbiadita dall'etere, venne attribuito a
30
31

FLAGELLI
LUCE
MEMBRANA PURPUREA
+ •4- a
+
+
MEMBRANA ROSSA
+
+
+
PARETE CELLULARE
▪
• s+•
• iINISI
ADP + Pi
ATP
ATPADP + Pi '
111+a+
▪ CATENARESPIRATORIA
111 02
FLAGELLI
Il trasporto di protoni, in una cellula di batterio alofilo, è correlato al metabolismo energetico.La membrana cellulare contiene zone di membrana purpurea e pigmenti della catena respirato-ria. Sia la luce, sia l'ossigeno possono promuovere l'eliminazione di protoni. Il processo alimen-tato dalla luce è mediato dalla membrana purpurea, mentre il processo alimentato dall'ossigenoavviene attraverso le catene respiratorie in presenza di un substrato ossidabile. Il risultantepotenziale di membrana (positivo all'esterno) e la concentrazione di protoni (maggiore all'ester-no) alimentano un flusso all'indietro di protoni tramite l'enzima ATPasi, il quale sintetizzal'adenosintrifosfato (A TP) a partire dall'adenosindifosfato (A D P) e da fosfato inorganico (Pi).
700650450 500 550 600LUNGHEZZA D'ONDA (NANOMETRI)
120
100
40
80
60
20
o
La sintesi di ATP può venire alimentata o dalla luce o dall'ossigeno. Quando alcune cellule diH. halobium vengono private di ossigeno (in un contenitore chiuso in cui viene fatto gorgogliaredell'azoto) e di luce (regione ombreggiata del grafico) la concentrazione di ATP diminuisce.Quando la luce viene accesa, il livello di ATP aumenta immediatamente. Esso diminuisce di nuo-vo al buio, ma viene reintegrato quando, nella sospensione, viene fatto gorgogliare dell'ossigeno.
0 6
0,7
5o
uJ
Oo
5i LIJ
CL
oo2
0,5
E
O
I
oo
00
La dipendenza della sintesi dell'ATP dalla membrana purpurea può essere dimostrata confron-tando la produzione di ATP a varie lunghezze d'onda con gli spettri della membrana e di unasospensione di cellule integre. Gli istogrammi mostrano le quantità di ATP sintetizzate quandole cellule vengono illuminate attraverso filtri che lasciano passare solo strette bande di lunghezzed'onda. Essi si adeguano allo spettro della membrana purpurea (curva continua in colore) piut-tosto che allo spettro ottenuto dalla sospensione di cellule integre (curva tratteggiata in colore).
o 15 30 45TEMPO (MINUTI)
60 75
AZOTO >.E—OSSIGENO
•
o
0,4
o
0,3
0,2
0.1
0
•
-----
una batteriorodopsina modificata, for-matasi e accumulatasi rapidamente allaluce e quindi degradatasi un poco piùlentamente al buio per riformare il com-plesso con assorbimento a 570 nanome-tri. Oesterhelt aveva inoltre scoperto chesia lo sbiadimento, sia la reazione inver-
sa si associavano alla liberazione e allaassunzione di ioni idrogeno, o protoni. Ilcomplesso sbiadito veniva «deprotonato»e la sua inversione verso il complessooriginario con assorbimento a 570 nano-metri era accompagnata da una «ripro-tonazione». Il movimento di protoni si
rifletteva in un cambiamento reversibiledel pH del mezzo, che diventava più aci-do quando i protoni erano liberati dallamembrana e meno acido quando la mem-brana si riprotonava. Oesterhelt avevaosservato analoghi cambiamenti di pH,indotti dalla luce, in sospensioni di cellu-le intatte.
Nel frattempo, studiavo con altri stru-menti le modificazioni nel potere di
assorbimento della membrana purpurea:in particolare mediante spettroscopia abassa temperatura, con Richard Loziernel laboratorio di Warren L. Butler pres-so l'Università della California a SanDiego, e mediante spettroscopia lampocon Richard A. Cone prèsso la JohnsHopkins University. Questi sistemi per-mettono di individuare cambiamentitransitori nel potere di assorbimento ehanno rivelato che i cambiamenti indottidalla luce, che avevamo osservato finoad allora solo in presenza di etere, avve-nivano, anche in assenza di etere, sia nelmateriale che costituisce la membranache nelle cellule intatte, se erano sospesinel mezzo salino e nel materiale dellamembrana sospeso in acqua pura. Ab-biamo inoltre trovato che avevamo a chefare con una fotoreazione ciclica, checomporta parecchi composti intermedi,tra i quali il complesso con massimo diassorbimento a 412 nanometri non erache uno dei tanti. L'assorbimento dellaluce nella banda di 570 nanometri fapassare più volte il pigmento attraversole varie fasi del ciclo, fino a quando laluce viene spenta; ogni ciclo si completain pochi millisecondi e si associa alla li-berazione e alla assunzione di protoni (siveda la figura in basso a pagina 29).L'etere modifica la cinetica del ciclo percui si accumula e viene facilmente messoin rilievo un complesso sbiadito, depro-tonato, con massimo di assorbimento a412 nanometri. Senza etere, non si accu-mulano significative quantità del com-plesso sbiadito, anche in condizioni dicontinua illuminazione (a meno che nonsi abbiano delle intensità elevatissime).Per questo motivo non eravamo riuscitia individuarlo.
Oesterhelt e io pubblicammo questi ri-sultati. Le nostre osservazioni chiave siriassumevano in un'acidificazione rever-sibile del mezzo, indotta dalla luce, nellesospensioni cellulari, nella liberazione eassunzione di protoni da parte dellamembrana purpurea durante la fotorea-zione e nel fatto che l'esposizione dellecellule intatte alla luce faceva abbassareil loro livello di respirazione. La libera-zione e l'assunzione di protoni da partedelle cellule intatte erano chiaramentediverse dal cambiamento nella protona-zione osservato durante la fotoreazionedella membrana purpurea isolata; nellesospensioni di cellule, il cambiamentoavveniva più lentamente, ma raggiunge-va un valore più alto. Nondimeno siamostati indotti a pensare che i due fenomenifossero strettamente correlati e abbiamopostulato che nella cellula integra lamembrana agisca da pompa di protoni,
suggerendo che la liberazione e l'assun-zione di questi ultimi hanno luogo suilati opposti della membrana. C'è una viaattraverso la membrana che permette aiprotoni di spostarsi dal sito'cli assunzio-ne a quello di liberazione, ma non in di-rezione opposta. In condizioni di illumi-nazione costante, il pigmento si riciclacontinuamente con un ritmo di parec-chie centinaia di volte al secondo, assu-mendo protoni sul lato citoplasmico, in-terno, della membrana e riversandoli nelmezzo, sul lato esterno di essa. Pertanto,tra l'interno della cellula e l'ambienteesterno si stabiliscono un gradiente diconcentrazione di protoni e un potenzia-le elettrico. L'energia luminosa si tra-sforma in un gradiente elettrochimico.
Questa interpretazione dei nostri risul-tati è stata suggerita dalla teoria «che-mioosmotica» della trasduzione di ener-gia, proposta alcuni anni fa da PeterMitchell dei Glynn Research Laborato-ries in Inghilterra. Questo autore soste-neva che l'energia derivata dal flusso dielettroni lungo la catena di trasporto de-gli elettroni o nella fosforilazione ossida-tiva (respirazione) o nella fotofosforila-zione (fotosintesi) viene innanzitutto im-magazzinato sotto forma di un gradienteelettrochimico di protoni attraverso lamembrana, contenente le catene di tra-sporto degli elettroni. Questa riserva dienergia serve per sintetizzare l'ATP dal-l'ADP (adenosindifosfato) e dal fosfatoinorganico, quando i protoni rifluisconoattraverso la stessa membrana attraversoun particolare enzima, l'ATPasi di mem-brana. Il meccanismo di pompaggio deiprotoni nella fosforilazione ossidativa enella fotosintesi è sicuramente molto di-verso da quello che avevamo postulatoper la membrana purpurea, ma il risulta-to in tutti e tre i casi è lo stesso: l'energiachimica libera proveniente da qualchesubstrato ossidabile o dalla luce si tra-sforma nell'energia libera di un gradien-te elettrochimico di protoni. La funzioneprimaria della membrana purpurea con-siste, dunque, nel fare in modo che la lu-ce diventi un'alternativa alla respirazionecome fonte di energia per i batteri alofi-li. Un'osservazione che concorda conquest'interpretazione è quella che le cel-lule sintetizzano una membrana purpu-rea soltanto quando crescono in presen-za di basse concentrazioni di ossigeno ealla luce.
Avevamo ancora da dimostrare che lecellule possono, di fatto, sintetizza-
re l'ATP mediante il gradiente generatodalla luce. I nostri primi esperimenti era-no stati inconcludenti, come abbiamopoi scoperto, perchè non ci eravamopreoccupati di stabilire per le cellule del-le condizioni sufficientemente anaerobi-che. Arlette Danon e io abbiamo conti-nuato a lavorare assieme su questo pro-blema e la Danon, tornata al WeizmannInstitute in Israele, è riuscita a realizzarel'esperimento. Ha sospeso i batteri inuna soluzione salina senza sostanze nu-tritizie e, al buio, vi ha fatto gorgogliaredell'azoto. In queste condizioni, la con-
32
33

Questa microfotografia elettronica ottenuta mediante criodecapaggio mostra vescicole lipidicheartificiali che incorporano frammenti di membrana purpurea ingrandite 77 000 volte. Le parti-celle che si notano qui sono prevalentemente sulle facce di frattura concave, non sulle facceconvesse come nelle cellule (si veda la figura in alto a pagina 27). Esse, cioè, sono incluse nellametà esterna della membrana che si è scissa; l'orientazione della batteriorodopsina nelle vescico-le e Opposta a quella dei batteri. I,a proteina, mentre pompa protoni entro le eseicole, fa‘oriseela loro uscita dalle cellule; alla luce l'ambiente delle vescicole tende a diminuire di acidità.
•n•
PAUL MUSSEN JOHN I CONGER JERONIE
LO SVILUPPODEL BAMBINOE LA PERSONALITA
WORM
1n1.1.1"•nn MIMI1 ~MI.1n1. .1n1111MIM.1 ••••nnn1n11n•
nn• 1n1, InLn, =.1n1,MéMIW.111n1•1/
Una strategia organica di « campi lunghi » e di . « primi piani » sulbambino e l'adolescente. L'incidenza dell'ambiente sociale e cultu-rale; le nuove conoscenze in settori quali la percezione, la memoriae il linguaggio; le teorie del comportamento, psicoanalitiche, piage-tiane. Il più noto manuale di psicologia dell'età evolutiva in America.Volume unico pp. 580, L. 9.800Edizione in cinque volumi:
periodo prenatale, pp. 108, L. 2.200; I primi due anni, pp. 80, L. 1,900;Linguaggio e sviluppo cognitivo, pp. 112, L. 2.200; Gli anni prescolari -La terza infanzia, pp. 148, L. 2.400; L'adolescenza, pp. 128, L. 2.200.
centrazione intracellulare di ATP si è ab-bassata rapidamente a circa il 30 per cen-to del livello originario ed è rimasta co-stante. Chiaramente le cellule sono ingrado di sospendere la massima parte deiprocessi che richiedono energia, quandonon sono disponibili fonti esterne di que-sta. Quando la sospensione cellulare vie-ne illuminata, il contenuto di ATP dellecellule sale rapidamente fino al livellooriginario o anche al di sopra; al buioquesto livello scende di nuovo. L'intro-duzione di aria (cioè di ossigeno) nellasospensione ha lo stesso effetto della lu-ce: il contenuto di ATP sale. Quandoabbiamo misurato l'accumulo di ATP incondizioni di illuminazione con diverselunghezze d'onda, abbiamo trovato chesoltanto le radiazioni aventi lunghezze dionda assorbite dalla membrana purpureasono efficaci nella sintesi di ATP.
Abbiamo continuato a raccogliere os-servazioni che erano se non altro concor-di con affermazioni più specifiche: chesia la respirazione, sia la luce generanoun gradiente elettrochimico di protoniattraverso la membrana cellulare e che ilgradiente alimenta la sintesi di ATP aopera dell'ATPasi della membrana. Ab-biamo potuto compiere questi lavori u-sando sostanze che interferiscono in ma-niera specifica con la sintesi di ATP.Innanzitutto abbiamo usato due sostanzechimiche che inibiscono direttamente laattività della ATPasi di membrana e am-bedue prevengono l'accumulo dell'ATP,determinato o dalla luce o dalla respira-zione. In secondo luogo abbiamo prova-to dei disaccoppianti, agenti che permet-tono al trasporto di elettroni di realizzar-si, ma che, di fatto, lo disaccoppianodalla fosforilazione e, pertanto, dalla sin-tesi di ATP che generalmente essa com-pie. La teoria di Mitchell afferma che idisaccoppianti rendono la membranamolto permeabile ai protoni, impedendocosì che si generi un gradiente elettrochi-
mico: difatti membrane modello, trattatecon disaccoppianti, diventano permeabiliai protoni. Nelle sospensioni di batterialofili, i disaccoppianti non solo hannoinibito l'accumulo di ATP determinatodalla luce o dalla respirazione, ma hannoanche impedito l'acidificazione del mez-zo, che riflette una deprotonazione. Ab-biamo provato, infine, parecchie sostan-ze che interferiscono in maniera specificacon la catena di trasporto degli elettroni,bloccando tutto l'accumulo di ATP de-terminato dalla respirazione, senza avereinvece alcun effetto sulla reazione indot-ta dalla luce. Esse hanno bloccato anchel'acidificazione del mezzo indotta dallarespirazione, ma non quella indotta dallaluce.
Tutte queste osservazioni erano inbuon accordo con il nostro postulatocentrale che la membrana purpurea èuna pompa di protoni azionata dalla lu-ce. Tuttavia, anche le cellule batterichesono sistemi enormemente complessi e illoro metabolismo energetico non è statoancora del tutto compreso, per cui pos-siamo solo formulare delle spiegazioniplausibili in alternativa alle nostre osser-vazioni.
T e cellule di Halobacterium halobiumpossono essere rotte sottoponendole
a un congelamento e a un disgelo alter-nati, a torsioni o a una rapida riduzionedella concentrazione salina del mezzoambiente. In tali condizioni, la cellula sisvuota della massima parte del suo con-tenuto e le membrane si saldano nuova-mente tra loro, formando delle vescicolecave, o sacchi, che si chiamano involucricellulari. Assicuratici soltanto che in unodi questi involucri venisse mantenuto laopportuna orientazione della membranapurpurea, abbiamo notato che questi sac-chi vuoti reagivano alla luce, generandoun gradiente elettrochimico di protoni.Ciò escludeva la possibilità che i sistemi
enzimatici nel citoplasma fossero i re-sponsabili dei vari eventi che avevamoosservato. Tuttavia rimaneva la possibi-lità che sistemi produttori di energia,non identificati, legati alla membranadella cellula, potessero svolgere un certoruolo. Per verificare questa ipotesi, do-vevamo rimuovere la porzione principale(rossa) della membrana cellulare e sosti-tuirla con una barriera alla permeabilità,che avesse proprietà analoghe, ma fossemetabolicamente inerte. Tutto questo eragià stato fatto in precedenza. Doppi stra-ti lipidici si formano spontaneamentequando i lipidi della membrana o copiesintetiche di essi vengono dispersi nel-l'acqua e i doppi strati tendono a forma-re minuscole vescicole; inoltre, vari com-ponenti si possono combinare con i lipidinel tentativo di ricostruire membranemodello funzionali.
Efraim Racker della Cornell Universi-ty, che con questi metodi aveva ricostrui-to con successo alcuni elementi dellacatena di trasporto degli elettroni dei mi-tocondri, ha accettato di collaborare conme, per cui mi sono recato a Ithaca conun preparato di membrana purpurea iso-lata. Il nostro primo esperimento ha a-vuto successo: abbiamo combinato fram-menti di membrana purpurea con vesci-cole fosfolipidiche artificiali incubandola miscela con un poco di detergente osottoponendola ad agitazione medianteonde sonore. Quando il preparato cosìottenuto è stato esposto alla luce, il mez-zo ha subito cambiamenti reversibili dipH, molto più imponenti di quanto po-tesse essere giustificato da una trasfor-mazione di tutta la proteina della mem-brana purpurea nella forma deprotona-ta. Come dato di fatto, le intensità lumi-nose erano tali che solo poca batterioro-dopsina avrebbe potuto essere deproto-nata; il preparato doveva pompare atti-vamente protoni. Abbiamo trovato chel'azione di pompaggio era sensibile al-l'effetto dei disaccoppianti, il che indica-va chiaramente che la luce stava stabi-lendo un gradiente di protoni attraversola membrana della vescicola lipidica. Da-to che la batteriorodopsina era la solaproteina presente nei nostri preparati eche essa funzionava nelle vescicole for-mate da vari lipidi, risultò evidente cheera questo pigmento a trasformare l'e-nergia luminosa per generare un gradien-te di protoni.
C'era una singolare differenza tra l'ef-fetto della luce sulle cellule intatte o sugliinvolucri cellulari e l'effetto su questevescicole lipidiche. In quest'ultimo casola luce produceva non una diminuzionedel pH del mezzo (acidificazione), ma unaumento (alcalinizzazione) e i protonivenivano pompati dal mezzo circostantenelle vescicole. Ciò ha fatto pensare chela batteriorodopsina fosse orientata inmaniera opposta nelle cellule o nelle ve-scicole, vale a dire che nelle vescicole illato «citoplasmatico» della membranafosse rivolto verso l'ambiente esterno.Siamo riusciti a verificare questa ipotesicon microfotografie elettroniche di pre-parati ottenuti mediante criodecapag-
gio. Se una membrana viene congelata equindi rotta, essa tende a scindersi lungoil piano che divide i due strati del doppiostrato lipidico. Avevamo notato che lemembrane di cellule intatte, sottoposte acriodecapaggio, presentavano due faccedi frattura chiaramente distinguibili. Lefacce convesse, che dovevano trovarsisulla metà interna, o citoplasmatica, del-la membrana spaccata, erano costellatedi proteine strettamente ammassate; lefacce concave, che erano le facce di frat-tura della metà membrana esterna erano,invece, lisce (si veda la figura in alto apagina 27). I preparati ottenuti per crio-decapaggio delle vescicole artificialimostrano un quadro opposto, confer-mando così che, in questo caso, la pro-teina è orientata in senso inverso (si vedala figura nella pagina a fronte).
In seguito le osservazioni sulle vescico-le lipidiche sono state ampliate da San--Bao Hwang e da altri nel mio laborato-rio, da V.P. Skulachev e collaboratoriall'Università statale di Mosca e da ricer-catori del laboratorio di Racker. Un ri-sultato merita una menzione speciale,perché costituisce un importante argo-mento in favore della teoria chemio-osmotica di Mitchell. Racker aveva giàrealizzato una sintesi di ATP incorpo-rando parti della catena di trasporto de-gli elettroni e ATPasi mitocondriale invescicole lipidiche e aggiungendo un sub-strato ossidabile. Se, come sostiene lateoria di Mitchell, la catena di trasportodegli elettroni funziona generando ungradiente di protoni elettrochimico, chea sua volta alimenta la sintesi di ATP ca-talizzata dall'ATPasi, allora si dovrebbepoter sostituire la batteriorodopsina e laluce al posto degli elementi della catena edel substrato ossidabile. Racker vi è riu-scito: ha incorporato l'ATPasi dei mito-condri di cuore di bue nelle vescicole co-stituite da lipidi e batteriorodopsina;quindi ha illuminato il preparato, ha sin-tetizzato ATP dall'ADP e da fosfatoinorganico.
Queste indagini chiudono il primo ca-pitolo dell'esplorazione di un nuovo si-stema fotosintetico. Abbiamo dimostra-to che un organismo privo di clorofillapuò captare e trasformare l'energia lumi-nosa e utilizzarla per sintetizzare ATP oper alimentare altri processi metabolici.Fa questo mediante una cromoproteinache assomiglia molto al pigmento visivodegli animali. Il pigmento è incorporatonella membrana cellulare, dove trasfor-ma l'energia luminosa in protoni di tra-sporto che l'attraversano, contro un po-tenziale elettrico e un gradiente di con-centrazione.
Un problema che va affrontato ora èquello della natura del meccanismo
intramolecolare grazie al quale l'assorbi-mento della luce fa muovere i protoniattraverso la membrana, meccanismo og-gi intensamente studiato in molti labora-tori, con sempre nuovi risultati. Al Me-dical Research Council Laboratory ofMolecular Biology di Cambridge, in In-ghilterra, Richard Henderson e Nigel
Unwin hanno sfruttato la disposizionecristallina della batteriorodopsina nellamembrana per ottenere una mappa tridi-mensionale della struttura proteica conuna nuova tecnica di ricostruzione dellamembrana, partendo da un numero ele-vato di microfotografie elettroniche otte-nute con esposizioni estremamente basseai fasci di elettroni. È questa la primastruttura di una proteina di membranache è stata determinata con una risolu-zione quasi atomica. Assieme alle analisidelle sequenze amminoacidiche, che ven-gono realizzate in parecchi altri labora-tori, una simile ricostruzione dovrebbedarci ulteriori informazioni sul meccani-smo di pompaggio. La nostra indaginesulla cinetica del ciclo di fotoreazione,assieme agli studi spettrografici compiutida Aaron Lewis alla Cornell University,hanno rivelato che è la base di Schiff delretinene che, in maniera specifica, perdeun protone per guadagnarne successiva-mente un altro, durante il ciclo, e per-tanto deve essere probabilmente interes-sata in maniera diretta nel movimentodei protoni.
Sulla base dei numerosi dati sinoraraccolti, si stanno preparando modellimolecolari del meccanismo di pompag-gio, che potranno essere controllati me-diante nuovi esperimenti. Ciò che ap-prenderemo potrà migliorare notevol-mente le nostre conoscenze a proposito
di altre pompe ioniche che agiscono inmolti altri tipi di membrane.
Un altro problema è come, in seguito,la cellula trasformi l'energia dell'ATP eregoli il flusso dell'energia attraverso suc-cessivi processi metabolici. In questo ca-so i preparati relativamente semplici de-gli involucri sono più facilmente analiz-zabili delle cellule intatte. Ricercatori del-l'Arnes Research Center della NationalAeronautics and Space Administration edella Cornell University hanno trovatoche la luce può alimentare l'assunzione,da parte degli involucri, di ioni sodio epotassio oltre che di amminoacidi e chequest'assunzione è indipendente dallasintesi di ATP.
La scoperta della membrana purpureaha cominciato a influenzare fortementeun certo numero di campi della biologiache hanno un elevato interesse, in parti-colare in relazione alla struttura e allafunzione delle membrane in generale e aisistemi di trasduzione dell'energia in par-ticolare. L'esame della relazione esisten-te tra batteriorodopsina e pigmenti visividell'occhio dovrebbe aprire una nuovaprospettiva sui meccanismi della visionee sulla sua evoluzione.
Non è inoltre escluso che potremo ap-prendere da questi batteri che sono ingrado di ricavare energia dalla luce comeanche noi potremmo fare un uso miglio-re del Sole come fonte di energia.
34
35