Flusso ematico e attività cerebrale -...
5
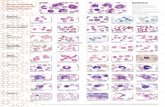
Flusso ematico e attività cerebrale Mediante l'impiego di un isotopo radioattivo è possibile osservare su uno schermo televisivo come il flusso ematico vari a seconda del livello di attività delle aree cerebrali di Niels A. Lassen, David H. Ingvar e Erik Skinhój / frenologi dell'inizio del XIX secolo credevano che le qualità morali di un individuo, come la creatività e l'ag- gressività, potessero essere dedotte dalla disposizione delle protuberanze sulla superficie del cranio, che riflettevano pre- sumibilmente una ipertrofia di zone spe- cifiche del cervello in esso contenuto. Sebbene questa ipotesi, fantasiosa ma tenace, sia ora del tutto priva di credito, per ironia della sorte essa contiene un germe di verità. Gli studi degli effetti del danno cerebrale sul comportamento e della stimolazione elettrica del cervello nel corso di interventi chirurgici, hanno rivelato che funzioni sensorie e motorie specifiche (piuttosto che tratti della per- sonalità) sono localizzate in determinate regioni della corteccia cerebrale, lo strato di cellule nervose, avente uno spessore di soli cinque millimetri, che costituisce la superficie del cervello. Nei nostri labora- tori dell'ospedale Bispebjerg di Copena- ghen e della Università di Lund, in Svezia, una nuova tecnica basata sull'uso di iso- topi radioattivi ci ha permesso di osserva- re direttamente la localizzazione delle funzioni nel cervello umano. I primi importanti tentativi di localizza- re le funzioni corticali furono compiuti nel 1861 dal medico francese Paul Broca, che, dopo la morte, sottopose ad autopsia due paralitici, entrambi paralizzati solo dal lato destro e sofferenti di una grave afasia che li aveva resi muti. Nel corso delle autopsie Broca scoprì un netto dete- rioramento del tessuto cerebrale in una zona dell'emisfero sinistro e concluse che la funzione del linguaggio è localizzata in quella regione del cervello. I ricercatori successivi adottarono il metodo indiretto di Broca, per localizzare molte altre fun- zioni corticali sulla base di insufficienze neurologiche e mentali derivate da danno cerebrale. Negli anni quaranta e cinquanta, nu- merosi neurochirurghi, e in particolare Wilder Penfield e i suoi colleghi del Mon- treal Neurological Institute, costruirono una mappa di varie aree della corteccia stimolandola elettricamente durante in- terventi chirurgici in cui la superficie del cervello doveva essere esposta a causa della natura della malattia. Le caratteri- stiche della risposta a ciascuna stimola- zione erano determinate sulla base delle osservazioni verbali del paziente che, nel corso della procedura, era sveglio. Per esempio, se venivano stimolati punti della area somatosensoria della corteccia, il paziente riferiva sensazioni di torpore o di prurito in zone specifiche della superficie del corpo. Tali studi di localizzazione, confortati da numerose osservazioni negli animali, hanno permesso la determinazione di una serie fondamentale di relazioni tra aree della corteccia e funzioni: il lobo occipita- le nella parte posteriore della corteccia media le funzioni visive, la corteccia po- stero-centrale media la sensibilità della pelle e dei muscoli, zone dei lobi tempora- li mediano la funzione uditiva, la cortec- cia antero-centrale media il controllo motorio, mentre l'area di Wernicke e quella di Broca (che, praticamente in ogni individuo, sono localizzate nell'emisfero cerebrale sinistro) mediano rispettiva- mente la comprensione e la sintesi del linguaggio. Zone relativamente estese della corteccia non sono attivate da stimo- li sensoriali né sono coinvolte nella me- diazione di attività motorie; queste regio- ni, tra cui quelle chiamate aree associati- ve, sono coinvolte nell'elaborazione delle afferenze sensorie, nella programmazio- ne di attività motorie e forse in funzioni puramente mentali. Sebbene gli studi sulle lesioni cerebrali e sulla elettrostimolazione abbiano forni- to una gran quantità di informazioni sulla localizzazione corticale delle funzioni, tali studi dimostrano limiti intrinseci. Come fu puntualizzato già nel 1874 da John Hughlings Jackson del National Hospital for Nervous Deseases di Londra «localiz- zare il danno che impedisce il linguaggio e localizzare il centro del linguaggio sono due cose diverse». Per esempio, come si vedrà in questo articolo, l'emisfero destro è molto più attivo mentre il soggetto sta parlando di quanto potrebbe sembrare in base agli studi basati esclusivamente sul danno tissutale. Un metodo più diretto per chiarire il problema della localizza- zione consiste nello studiare la corteccia cerebrale integra in diverse situazioni funzionali, mentre il cervello è «al lavo- ro». Una tecnica consiste nel registrare mediante minuscoli elettrodi la modifica- zione della velocità di scarica di ciascuna cellula nervosa della corteccia. Tuttavia per decifrare le interazioni enormemente complesse tra 10 miliardi di cellule nervo- se cerebrali non è sufficiente neppure una nutrita batteria di microelettrodi. Noi abbiamo adottato un metodo del tutto diverso e di più semplice e ampia applica- zione: la misura dell'incremento del flus- so ematico nelle aree della corteccia che vengono stimolate per l'esecuzione di specifiche attività sensoriali, motorie o mentali. I I nostro metodo si basa sul fatto che la circolazione del sangue attraverso i tessuti dell'organismo varia in rapporto al livello del metabolismo e all'attività fun- zionale di quei tessuti. Le relazioni reci- proche tra funzione, metabolismo ener- getico e flusso ematico furono postulate per la prima volta nel 1914 da Joseph Barcroft della Università di Cambridge, il quale affermò che un aumento del livello funzionale di un tessuto può essere man- tenuto solo aumentando il ritmo con cui viene consumato l'ossigeno. La maggior parte dell'energia utilizzabile nell'organi- smo deriva dalla scissione della molecola adenosintrifosfato (ATP) ricca di ener- gia, nei prodotti adenosindisfosfato (ADP) meno ricco di energia e fosforo inorganico (P i ). L'ATP viene poi ricostituito attraverso una reazione tra i suoi prodotti di scissio- ne, che richiede ossigeno e glucosio (fo- sforilazione ossidativa). Il tracciato allo stato di riposo dell'attività della cellula nen osa degli emisferi destro e sinistro della corteccia cerebrale normale, è stato riv elato misurando il flusso ematico regionale, che è strettamente asso- ciato al tasso metabolico e quindi all'atti% ità funzionale. Le figure sono state realizzate da un calcolatore sulla base di dati ottenuti individuan- do il passaggio dell'isotopo radioattivo xeno 133 attraverso la corteccia. La percezione sensoriale modifica il tracciato del flusso ematico della corteccia, permettendo di individuare la localizzazione di aree che mediano l'elaborazione delle informazioni sensoriali. In queste imma- gini compaiono solo gli scostamenti del tasso di flusso rispetto allo stato di riposo. A sinistra il soggetto seguiva con gli occhi un oggetto in Il movimento olontario atti, a parti differenti della corteccia centrale in rapporto alla zona del corpo interessata. Nell'indagine di sinistra il soggetto muoveva le dita della mano dalla parte del corpo opposta a quella dell'emisfero esaminato; l'area corrispondente alla mano e alle Ciascun pixel, o elemento di immagine, rappresenta un centimetro quadrato di corteccia. In base alla scala cromatica, il tasso medio di flusso è verde, i tassi fino al 20 per cento inferiori alla media sono az- zurro sfumato e i tassi fino al 20 per cento superiori sono rosso sfuma- to. Le immagini mostrano con chiarezza che anche nello stato di ripo- so le aree frontali sono molto più attive di altre zone della corteccia. movimento; la corteccia di associazione v isiv a è attiva, così come i campi visivi frontali e l'area motoria supplementare nella parte supe- riore del lobo frontale. A destra il medesimo soggetto ascoltava alcune parole; la corteccia uditiva del lobo temporale è attiva, come l'adia- cente area di Wernicke, che media la comprensione del linguaggio. dita della corteccia centrale e l'area motoria supplementare sono attive. Nell'indagine illustrata a destra venne chiesto al soggetto di muovere la bocca contando ripetutamente fino a venti: l'area motoria della bocca, l'area motoria supplementare e la corteccia uditiva appaiono attive. 32 D 33
Transcript of Flusso ematico e attività cerebrale -...
Flusso ematicoe attività cerebrale
Mediante l'impiego di un isotopo radioattivo è possibileosservare su uno schermo televisivo come il flusso ematicovari a seconda del livello di attività delle aree cerebrali
di Niels A. Lassen, David H. Ingvar e Erik Skinhój
/
frenologi dell'inizio del XIX secolocredevano che le qualità morali di unindividuo, come la creatività e l'ag-
gressività, potessero essere dedotte dalladisposizione delle protuberanze sullasuperficie del cranio, che riflettevano pre-sumibilmente una ipertrofia di zone spe-cifiche del cervello in esso contenuto.Sebbene questa ipotesi, fantasiosa matenace, sia ora del tutto priva di credito,per ironia della sorte essa contiene ungerme di verità. Gli studi degli effetti deldanno cerebrale sul comportamento edella stimolazione elettrica del cervellonel corso di interventi chirurgici, hannorivelato che funzioni sensorie e motoriespecifiche (piuttosto che tratti della per-sonalità) sono localizzate in determinateregioni della corteccia cerebrale, lo stratodi cellule nervose, avente uno spessore disoli cinque millimetri, che costituisce lasuperficie del cervello. Nei nostri labora-tori dell'ospedale Bispebjerg di Copena-ghen e della Università di Lund, in Svezia,una nuova tecnica basata sull'uso di iso-topi radioattivi ci ha permesso di osserva-re direttamente la localizzazione dellefunzioni nel cervello umano.
I primi importanti tentativi di localizza-re le funzioni corticali furono compiutinel 1861 dal medico francese Paul Broca,che, dopo la morte, sottopose ad autopsiadue paralitici, entrambi paralizzati solodal lato destro e sofferenti di una graveafasia che li aveva resi muti. Nel corsodelle autopsie Broca scoprì un netto dete-rioramento del tessuto cerebrale in unazona dell'emisfero sinistro e concluse chela funzione del linguaggio è localizzata inquella regione del cervello. I ricercatorisuccessivi adottarono il metodo indirettodi Broca, per localizzare molte altre fun-zioni corticali sulla base di insufficienzeneurologiche e mentali derivate da dannocerebrale.
Negli anni quaranta e cinquanta, nu-merosi neurochirurghi, e in particolareWilder Penfield e i suoi colleghi del Mon-treal Neurological Institute, costruirono
una mappa di varie aree della cortecciastimolandola elettricamente durante in-terventi chirurgici in cui la superficie delcervello doveva essere esposta a causadella natura della malattia. Le caratteri-stiche della risposta a ciascuna stimola-zione erano determinate sulla base delleosservazioni verbali del paziente che, nelcorso della procedura, era sveglio. Peresempio, se venivano stimolati punti dellaarea somatosensoria della corteccia, ilpaziente riferiva sensazioni di torpore o diprurito in zone specifiche della superficiedel corpo.
Tali studi di localizzazione, confortatida numerose osservazioni negli animali,hanno permesso la determinazione di unaserie fondamentale di relazioni tra areedella corteccia e funzioni: il lobo occipita-le nella parte posteriore della cortecciamedia le funzioni visive, la corteccia po-stero-centrale media la sensibilità dellapelle e dei muscoli, zone dei lobi tempora-li mediano la funzione uditiva, la cortec-cia antero-centrale media il controllomotorio, mentre l'area di Wernicke equella di Broca (che, praticamente in ogniindividuo, sono localizzate nell'emisferocerebrale sinistro) mediano rispettiva-mente la comprensione e la sintesi dellinguaggio. Zone relativamente estesedella corteccia non sono attivate da stimo-li sensoriali né sono coinvolte nella me-diazione di attività motorie; queste regio-ni, tra cui quelle chiamate aree associati-ve, sono coinvolte nell'elaborazione delleafferenze sensorie, nella programmazio-ne di attività motorie e forse in funzionipuramente mentali.
Sebbene gli studi sulle lesioni cerebralie sulla elettrostimolazione abbiano forni-to una gran quantità di informazioni sullalocalizzazione corticale delle funzioni, talistudi dimostrano limiti intrinseci. Comefu puntualizzato già nel 1874 da JohnHughlings Jackson del National Hospitalfor Nervous Deseases di Londra «localiz-zare il danno che impedisce il linguaggio elocalizzare il centro del linguaggio sono
due cose diverse». Per esempio, come sivedrà in questo articolo, l'emisfero destroè molto più attivo mentre il soggetto staparlando di quanto potrebbe sembrare inbase agli studi basati esclusivamente suldanno tissutale. Un metodo più direttoper chiarire il problema della localizza-zione consiste nello studiare la cortecciacerebrale integra in diverse situazionifunzionali, mentre il cervello è «al lavo-ro». Una tecnica consiste nel registraremediante minuscoli elettrodi la modifica-zione della velocità di scarica di ciascunacellula nervosa della corteccia. Tuttaviaper decifrare le interazioni enormementecomplesse tra 10 miliardi di cellule nervo-se cerebrali non è sufficiente neppure unanutrita batteria di microelettrodi. Noiabbiamo adottato un metodo del tuttodiverso e di più semplice e ampia applica-zione: la misura dell'incremento del flus-so ematico nelle aree della corteccia chevengono stimolate per l'esecuzione dispecifiche attività sensoriali, motorie omentali.
II nostro metodo si basa sul fatto che la circolazione del sangue attraverso i
tessuti dell'organismo varia in rapporto allivello del metabolismo e all'attività fun-zionale di quei tessuti. Le relazioni reci-proche tra funzione, metabolismo ener-getico e flusso ematico furono postulateper la prima volta nel 1914 da JosephBarcroft della Università di Cambridge, ilquale affermò che un aumento del livellofunzionale di un tessuto può essere man-tenuto solo aumentando il ritmo con cuiviene consumato l'ossigeno. La maggiorparte dell'energia utilizzabile nell'organi-smo deriva dalla scissione della molecolaadenosintrifosfato (ATP) ricca di ener-gia, nei prodotti adenosindisfosfato(ADP) meno ricco di energia e fosforoinorganico (P i ).
L'ATP viene poi ricostituito attraversouna reazione tra i suoi prodotti di scissio-ne, che richiede ossigeno e glucosio (fo-sforilazione ossidativa).
Il tracciato allo stato di riposo dell'attività della cellula nen osa degliemisferi destro e sinistro della corteccia cerebrale normale, è statoriv elato misurando il flusso ematico regionale, che è strettamente asso-ciato al tasso metabolico e quindi all'atti% ità funzionale. Le figure sonostate realizzate da un calcolatore sulla base di dati ottenuti individuan-do il passaggio dell'isotopo radioattivo xeno 133 attraverso la corteccia.
La percezione sensoriale modifica il tracciato del flusso ematico dellacorteccia, permettendo di individuare la localizzazione di aree chemediano l'elaborazione delle informazioni sensoriali. In queste imma-gini compaiono solo gli scostamenti del tasso di flusso rispetto allo statodi riposo. A sinistra il soggetto seguiva con gli occhi un oggetto in
Il movimento olontario atti, a parti differenti della corteccia centralein rapporto alla zona del corpo interessata. Nell'indagine di sinistra ilsoggetto muoveva le dita della mano dalla parte del corpo opposta aquella dell'emisfero esaminato; l'area corrispondente alla mano e alle
Ciascun pixel, o elemento di immagine, rappresenta un centimetroquadrato di corteccia. In base alla scala cromatica, il tasso medio diflusso è verde, i tassi fino al 20 per cento inferiori alla media sono az-zurro sfumato e i tassi fino al 20 per cento superiori sono rosso sfuma-to. Le immagini mostrano con chiarezza che anche nello stato di ripo-so le aree frontali sono molto più attive di altre zone della corteccia.
movimento; la corteccia di associazione v isiv a è attiva, così come icampi visivi frontali e l'area motoria supplementare nella parte supe-riore del lobo frontale. A destra il medesimo soggetto ascoltava alcuneparole; la corteccia uditiva del lobo temporale è attiva, come l'adia-cente area di Wernicke, che media la comprensione del linguaggio.
dita della corteccia centrale e l'area motoria supplementare sono attive.Nell'indagine illustrata a destra venne chiesto al soggetto di muovere labocca contando ripetutamente fino a venti: l'area motoria della bocca,l'area motoria supplementare e la corteccia uditiva appaiono attive.
32 D
33
'AREA MOTORIASUPPLEMENTARE
CORTECCIA MOTORIACORTECCIA SOMATOSENSORIA(PRIMARIA E D'ASSOCIAZIONE)
CORTECCIA PREMOTORIA
LOBO FRONTALE
CAMPO OCULAREFRONTALE
LOBO PARIETALE
LOBOOCCIPITALE
AREA PREFRONTALE
AREAVISIVAPRIMARIA
AREA DI BROCA(LATO SINISTRO)
AREA OLFATTORIA
CORTECCIA MOTORIA
CORTECCIA PREMOTORIA
LOBO TEMPORALE
CORTECCIA SOMATOSENSORIA(PRIMARIA E D'ASSOCIAZIONE)
AREA UDITIVAPRIMARIA
AREA D'ASSOCIAZIONEVISIVA
AREA D'ASSOCIAZIONEUDITIVA (COMPRENDENTEL'AREA DI WERNICKESUL LATO SINISTRO)
LOBO PARIETALEAREA PREFRONTALE
LOBO OCCIPITALE
AREA MOTORIASUPPLEMENTARE
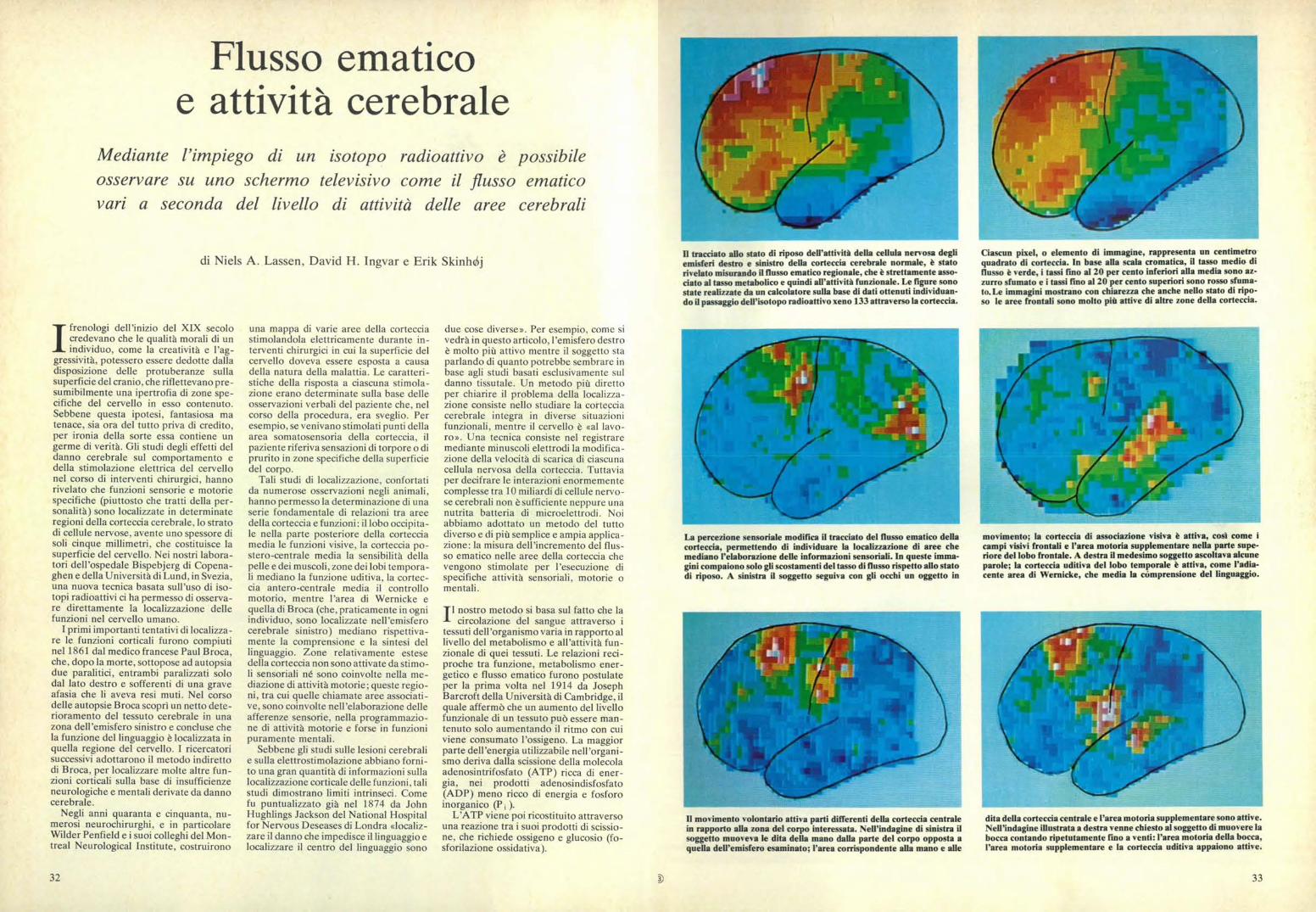
La mappa funzionale della corteccia cerebrale umana è stata ottenutacon metodiche diverse: studi sugli effetti di danni cerebrali, stimolazio-ne della corteccia esposta nel corso di interventi chirurgici, registrazionimediante elettrodi dell'attività corticale evocata da stimoli comporta-mentali o sensoriali e iniezione di isotopi radioattivi, secondo la tecnicadescritta in questo articolo. Ciascuna percezione sensoriale ha un'area
corticale primaria che riceve i prolungamenti provenienti da una super-ficie recettiv a periferica e un'area di associazione che elabora l'afferen-za sensoriale e la memorizza. L'area premotoria è coinvolta in attivitàcomplesse come lo scrivere a macchina. L'area motoria supplemen-tare programma i movimenti volontari sequenziali. L'area di Brocacontrolla la coordinazione dei muscoli coinvolti nel linguaggio.
ARTERIACEREBRALE
POSTERIOREARTERIA CEREBRALE MEDIA
ARTERIA CEREBRALEANTERIORE
LOBOOCCIPITALE
ARTERIA VERTEBRALE
Nel disegno sono rappresentate le arterie principali che riforniscono la corteccia cerebrale. Leregioni del linguaggio e quelle uditive sono irrorate dalla arteria cerebrale media. Le regionifrontale e della scissura interemisferica sono irrorate dall'arteria cerebrale anteriore. Entrambequeste arterie sono a loro volta ramificazioni dell'arteria carotide del collo. L'area visiv a primariasituata posteriormente nella corteccia, tuttavia, è irrorata dall'arteria cerebrale posteriore, che èuna ramificazione della arteria vertebrale del collo. Poiché lo xeno radioattivo v iene iniettatonella carotide, la corteccia visiNa primaria non compare nelle immagini del flusso ematico.
Poiché esiste un rapporto costante tra ilnumero di molecole di ATP rigenerate e ilnumero di molecole di ossigeno consuma-te nel processo, il livello funzionale di untessuto è strettamente associato alla suaassunzione di ossigeno. L'ossigeno vienefornito ai tessuti mediante il flusso emati-co e un aumento nella richiesta di ossige-no viene soddisfatto da un aumento delflusso di sangue ossigenato. Nel corso diuna attività muscolare intensa il flusso delsangue attraverso il tessuto muscolarepuò aumentare fino a venti volte i livellinormali, e si è anche trovato che il flussoematico negli altri organi si adatta ade-guatamente alle variazioni della richiestadi ossigeno. Poiché questa regolazionedel flusso ematico continua anche quandogli organi sono isolati e del tutto denerva-ti, essa sembra essere controllata da fatto-ri chimici rilasciati nel corso di processimetabolici locali, anche se l'esatto mec-canismo non è ancora chiarito.
Per quanto riguarda il cervello, le inter-relazioni tra attività funzionale, metaboli-smo e flusso ematico furono suggerite perla prima volta nel 1890 da Charles S. Roye da Charles S. Sherrington, che lavora-vano al Cambridge Pathological Labora-tory. Essi osservarono che entro alcunisecondi dall'insorgere di un attacco epi-lettico (una iperattività sincronizzata del-le cellule nervose che si esprime medianteconvulsioni muscolari) si manifestava unrigonfiamento del cervello, il che suggeri-va che si era verificato un massiccio au-mento del rifornimento ematico.
Indagini successive hanno dimostratoche il metabolismo cerebrale e il flussoematico subiscono un incremento anchedurante la normale attivazione di regionidella corteccia cerebrale coinvolte nellaesecuzione di compiti specifici. Nel 1937Cari F. Schmidt e James P. Hendrix dellaUniversity of Pennsylvania School ofMedicine, registrarono mediante termo-coppie un aumento strettamente circo-scritto del flusso ematico attraverso lacorteccia visiva di un gatto, quando veni-va illuminato un piccolo punto della reti-na dell'animale. Da allora sono stati rife-riti molti altri esempi di aumento localiz-zato del flusso ematico cerebrale, sia neglianimali che nell'uomo, ed è oggi certo chenel corso di una attività che coinvolge untipo specifico di funzione cerebrale si assi-ste a una modificazione locale dell'attivitàdella cellula nervosa e quindi del tassometabolico, che determina un aumentodel flusso ematico nella regione attivata.Questa scoperta ha reso possibile la loca-lizzazione di funzioni cerebrali studiandole variazioni regionali del flusso ematico.Si potrebbero dedurre le stesse informa-zioni, ugualmente bene, rintracciandomodificazioni del metabolismo locale(mediante la misurazione dell'assunzionesia di ossigeno che di glucosio), ma finorale funzioni della corteccia cerebrale del-l'uomo sono state localizzate solo me-diante misure di flusso ematico.
TI flusso ematico medio nel cervello1- umano fu determinato per la primavolta nel 1944, da Seymour S. Kety che
allora lavorava con Schmidt. Il metodo diKety consisteva nel misurare la velocitàcon cui il cervello si saturava o si desatu-rava di un gas inerte. Il soggetto inalavaper 10 minuti ossido d'azoto al 15 percento e, durante questo tempo, la pro-gressiva concentrazione del gas era se-guita dal prelievo di campioni di sanguearterioso e venoso proveniente dal cervel-lo. L'area compresa tra le curve di satura-zione arteriosa e venosa consentiva lamisura del flusso ematico medio, che nel-la norma è di circa 50 millilitri di sangueper 100 grammi di tessuto cerebrale perminuto. Seymour S. Kety misurò anche ladifferenza del contenuto in ossigeno trasangue venoso e sangue arterioso, che ènormalmente di circa 3,5 millilitri di ossi-geno per 100 grammi di tessuto cerebraleper minuto.
In seguito divenne possibile misurare ilflusso ematico in regioni circoscritte delcervello umano integro mediante l'uso diisotopi radioattivi. Nel 1961 i tre autorisvilupparono questa metodica partendoda studi sui gatti e arrivando ad applicarlanel campo della diagnostica umana. Ilmetodo implica l'uso di xeno 133, un iso-topo radioattivo del gas inerte xeno. Il gasradioattivo viene sciolto in soluzione fi-siologica e un piccolo volume (da due atre millimetri, che contengono da 3 a 5millicurie di radioattività) viene iniettatoin forma di bolo in una delle arterie prin-cipali del cervello. L'arrivo e la successivaeliminazione della radioattività da diver-se regioni cerebrali viene seguito minutoper minuto da un rivelatore a raggi gam-ma, consistente in una batteria di 254 ri-velatori a scintillazione collocati all'e-sterno, ciascuno dei quali viene collimatoin modo da permettere una scansione di
circa un centimetro quadrato della super-ficie cerebrale. L'informazione ottenutadai rivelatori viene elaborata da un picco-lo calcolatore digitale e rappresentata informa grafica su uno schermo televisivo acolori, in cui a ogni livello di flusso vieneassociato un colore o una tinta differenti.A causa della attenuazione della radia-zione proveniente dalle strutture cerebra-li più profonde, la radiazione gamma regi-strata proviene dalla corteccia cerebralesuperficiale. Così la tecnica con xeno ra-dioattivo fornisce un quadro abbastanzaspecifico della attività della corteccia ce-rebrale immediatamente al di sotto dellebatterie di rivelatori.
Abbiamo usato questa tecnica comeroutine in pazienti che dovevano esseresottoposti ad arteriografia. L'arteriogra-fia cerebrale comprende l'esecuzione diuna lastra radiografica dei vasi sanguignicerebrali, dopo iniezione nelle arterie diun mezzo di contrasto. Questa iniezioneviene fatta mediante catetere (un sottiletubo di plastica) inserito direttamentenella carotide interna per mezzo di unapuntura nel collo o fatto passare attraver-so l'arteria femorale per mezzo di unapuntura nell'inguine. Poiché per l'inie-zione di xeno radioattivo viene usato lostesso catetere, la misurazione del flussoematico cerebrale non comporta rischiulteriori per il paziente. (Il basso livellodella radiazione gamma emessa dalloxeno 133 non è considerato dannoso).
Nei nostri laboratori di Copenaghen edi Lund circa 500 pazienti sono stati sot-toposti a esame cerebrale con la tecnicabasata sull'uso di xeno radioattivo a scopodiagnostico per lo studio di colpi apoplet-tici, tumori o epilessia. Retrospettiva-mente possiamo dire circa 1'80 per cento
3435
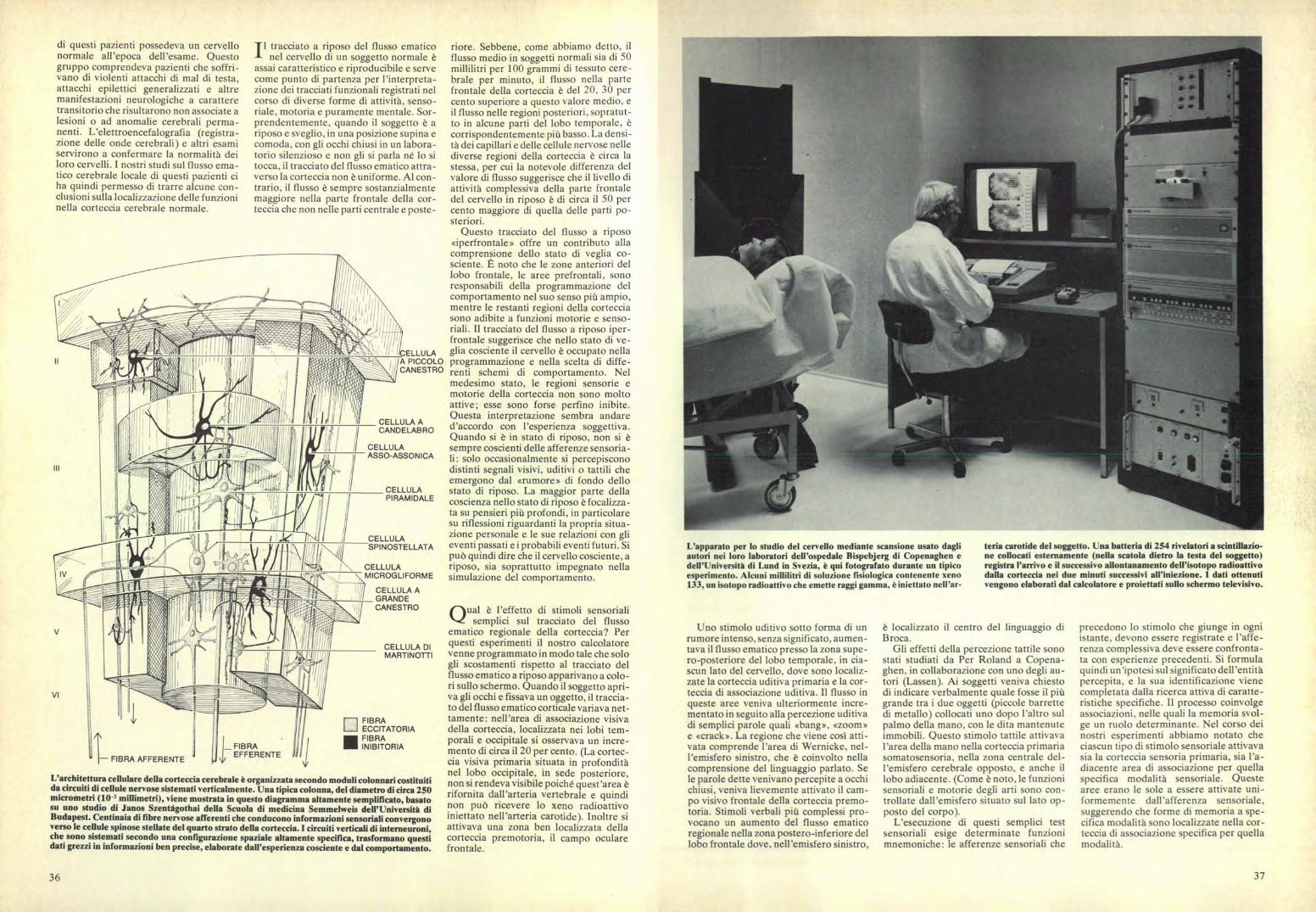
L'apparato per lo studio del cervello mediante scansione usato dagliautori nei loro laboratori dell'ospedale Bispebjerg di Copenaghen edell'Università di Lund in Sv ezia, è qui fotografato durante un tipicoesperimento. Alcuni millilitri di soluzione fisiologica contenente xeno133, un isotopo radioattivo che emette raggi gamma, è iniettato nell'ar-
teria carotide del soggetto. Una batteria di 254 rivelatori a scintillazio-ne collocati esternamente (nella scatola dietro la testa del soggetto)registra l'arrivo e il successivo allontanamento dell'isotopo radioattivodalla corteccia nei due minuti successivi all'iniezione. I dati ottenutivengono elaborati dal calcolatore e proiettati sullo schermo televisivo.
ELLULAA PICCOLOCANESTRO
ICELLULAASSO-ASSONICA
CELLULAPIRAMIDALE
CELLULA SPINOSTELLATA
CELLULAMICROGLIFORME
iÌi»i GRANDE
CELLULA A
CANESTRO
CELLULA ACANDELABRO
V
CELLULA DIMARTINOTTI
VI
T— FIBRA AFFERENTE
Il FIBRAEFFERENTE
FIBRAI ECCITATORIA
FIBRAM INIBITORIA
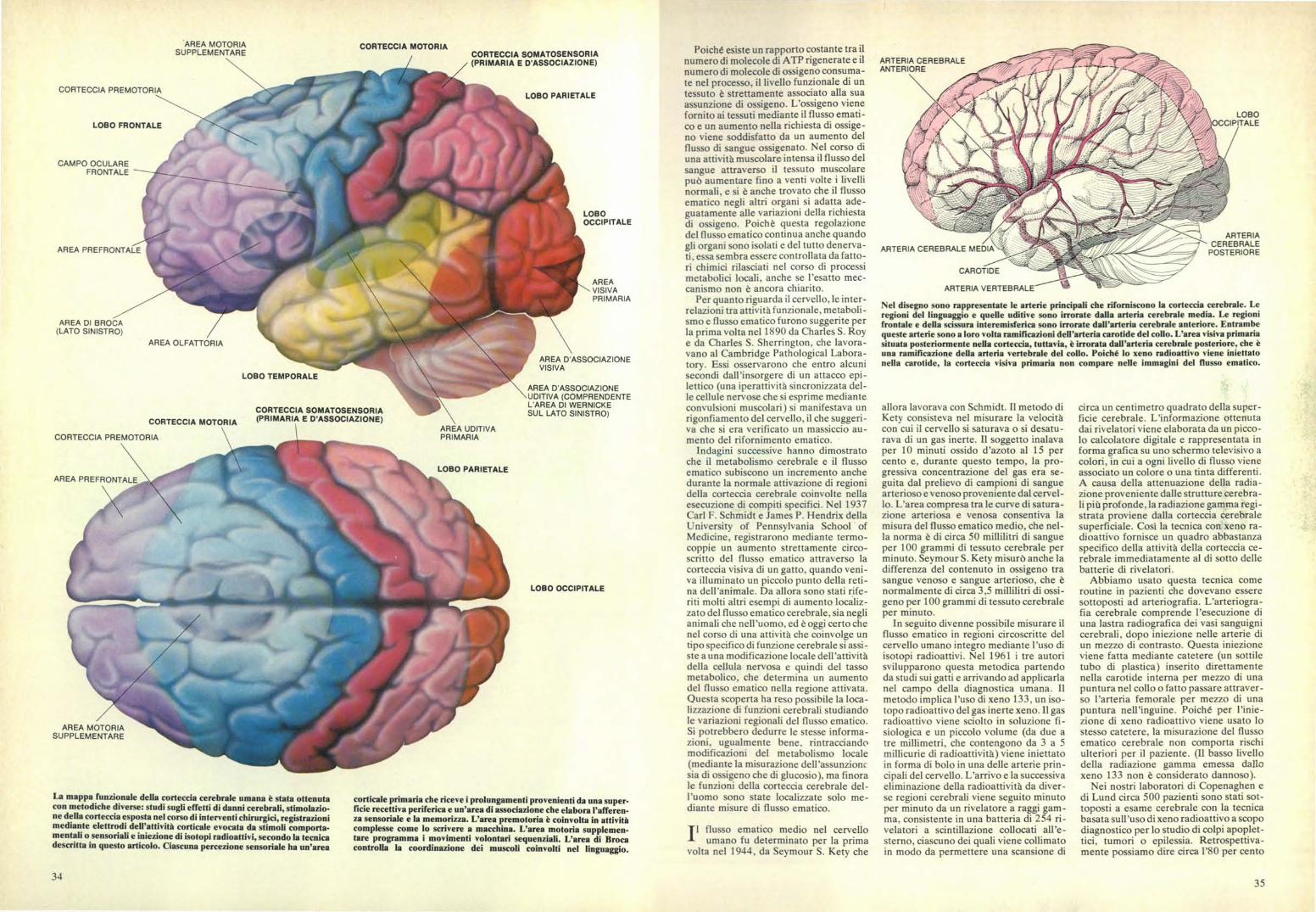
L'architettura cellulare della corteccia cerebrale è organizzata secondo moduli colonnari costituitida circuiti di cellule nervose sistemati verticalmente. Una tipica colonna, del diametro di circa 250micrometri (10 3 millimetri), viene mostrata in questo diagramma altamente semplificato, basatosu uno studio di Janos Szentàgothai della Scuola di medicina Semmelweis dell'Università diBudapest. Centinaia di fibre nervose afferenti che conducono informazioni sensoriali convergonoverso le cellule spinose stellate del quarto strato della corteccia. I circuiti verticali di interneuroni,che sono sistemati secondo una configurazione spaziale altamente specifica, trasformano questidati grezzi in informazioni ben precise, elaborate dall'esperienza cosciente e dal comportamento.
di questi pazienti possedeva un cervellonormale all'epoca dell'esame. Questogruppo comprendeva pazienti che soffri-vano di violenti attacchi di mal di testa,attacchi epilettici generalizzati e altremanifestazioni neurologiche a caratteretransitorio che risultarono non associate alesioni o ad anomalie cerebrali perma-nenti. L'elettroencefalografia (registra-zione delle onde cerebrali) e altri esamiservirono a confermare la normalità deiloro cervelli. I nostri studi sul flusso ema-tico cerebrale locale di questi pazienti ciha quindi permesso di trarre alcune con-clusioni sulla localizzazione delle funzioninella corteccia cerebrale normale.
Il tracciato a riposo del flusso ematico nel cervello di un soggetto normale è
assai caratteristico e riproducibile e servecome punto di partenza per l'interpreta-zione dei tracciati funzionali registrati nelcorso di diverse forme di attività, senso-riale, motoria e puramente mentale. Sor-prendentemente, quando il soggetto è ariposo e sveglio, in una posizione supina ecomoda, con gli occhi chiusi in un labora-torio silenzioso e non gli si parla né lo sitocca, il tracciato del flusso ematico attra-verso la corteccia non è uniforme. Al con-trario, il flusso è sempre sostanzialmentemaggiore nella parte frontale della cor-teccia che non nelle parti centrale e poste-
riore. Sebbene, come abbiamo detto, ilflusso medio in soggetti normali sia di 50millilitri per 100 grammi di tessuto cere-brale per minuto, il flusso nella partefrontale della corteccia è del 20, 30 percento superiore a questo valore medio, eil flusso nelle regioni posteriori, sopratut-to in alcune parti del lobo temporale, ècorrispondentemente più basso. La densi-tà dei capillari e delle cellule nervose nellediverse regioni della corteccia è circa lastessa, per cui la notevole differenza delvalore di flusso suggerisce che il livello diattività complessiva della parte frontaledel cervello in riposo è di circa il 50 percento maggiore di quella delle parti po-steriori.
Questo tracciato del flusso a riposo«iperfrontale» offre un contributo allacomprensione dello stato di veglia co-sciente. t noto che le zone anteriori dellobo frontale, le aree prefrontali, sonoresponsabili della programmazione delcomportamento nel suo senso più ampio,mentre le restanti regioni della cortecciasono adibite a funzioni motorie e senso-riali. Il tracciato del flusso a riposo iper-frontale suggerisce che nello stato di ve-glia cosciente il cervello è occupato nellaprogrammazione e nella scelta di diffe-renti schemi di comportamento. Nelmedesimo stato, le regioni sensorie emotorie della corteccia non sono moltoattive; esse sono forse perfino inibite.Questa interpretazione sembra andared'accordo con l'esperienza soggettiva.Quando si è in stato di riposo, non si èsempre coscienti delle afferenze sensoria-li : solo occasionalmente si percepisconodistinti segnali visivi, uditivi o tattili cheemergono dal «rumore» di fondo dellostato di riposo. La maggior parte dellacoscienza nello stato di riposo è focalizza-ta su pensieri più profondi, in particolaresu riflessioni riguardanti la propria situa-zione personale e le sue relazioni con glieventi passati e i probabili eventi futuri. Sipuò quindi dire che il cervello cosciente, ariposo, sia soprattutto impegnato nellasimulazione del comportamento.
Qual è l'effetto di stimoli sensoriali
semplici sul tracciato del flussoematico regionale della corteccia? Perquesti esperimenti il nostro calcolatorevenne programmato in modo tale che sologli scostamenti rispetto al tracciato delflusso ematico a riposo apparivano a colo-ri sullo schermo. Quando il soggetto apri-va gli occhi e fissava un oggetto, il traccia-to del flusso ematico corticale variava net-tamente: nell'area di associazione visivadella corteccia, localizzata nei lobi tem-porali e occipitale si osservava un incre-mento di circa il 20 per cento. (La cortec-cia visiva primaria situata in profonditànel lobo occipitale, in sede posteriore,non si rendeva visibile poiché quest'area èrifornita dall'arteria vertebrale e quindinon può ricevere lo xeno radioattivoiniettato nell'arteria carotide). Inoltre siattivava una zona ben localizzata dellacorteccia premotoria, il campo ocularefrontale.
Uno stimolo uditivo sotto forma di unrumore intenso, senza significato, aumen-tava il flusso ematico presso la zona supe-ro-posteriore del lobo temporale, in cia-scun lato del cervello, dove sono localiz-zate la corteccia uditiva primaria e la cor-teccia di associazione uditiva. Il flusso inqueste aree veniva ulteriormente incre-mentato in seguito alla percezione uditivadi semplici parole quali «bang», «zoom»e «crack». La regione che viene così atti-vata comprende l'area di Wernicke, nel-l'emisfero sinistro, che è coinvolto nellacomprensione del linguaggio parlato. Sele parole dette venivano percepite a occhichiusi, veniva lievemente attivato il cam-po visivo frontale della corteccia premo-toria. Stimoli verbali più complessi pro-vocano un aumento del flusso ematicoregionale nella zona postero-inferiore dellobo frontale dove, nell'emisfero sinistro,
è localizzato il centro del linguaggio diBroca.
Gli effetti della percezione tattile sonostati studiati da Per Roland a Copena-ghen, in collaborazione con uno degli au-tori (Lassen). Ai soggetti veniva chiestodi indicare verbalmente quale fosse il piùgrande tra i due oggetti (piccole barrettedi metallo) collocati uno dopo l'altro sulpalmo della mano, con le dita mantenuteimmobili. Questo stimolo tattile attivaval'area della mano nella corteccia primariasomatosensoria, nella zona centrale del-l'emisfero cerebrale opposto, e anche illobo adiacente. (Come è noto, le funzionisensoriali e motorie degli arti sono con-trollate dall'emisfero situato sul lato op-posto del corpo).
L'esecuzione di questi semplici testsensoriali esige determinate funzionimnemoniche: le afferenze sensoriali che
precedono lo stimolo che giunge in ogniistante, devono essere registrate e l'affe-renza complessiva deve essere confronta-ta con esperienze precedenti. Si formulaquindi un'ipotesi sul significato dell'entitàpercepita, e la sua identificazione vienecompletata dalla ricerca attiva di caratte-ristiche specifiche. Il processo coinvolgeassociazioni, nelle quali la memoria svol-ge un ruolo determinante. Nel corso deinostri esperimenti abbiamo notato checiascun tipo di stimolo sensoriale attivavasia la corteccia sensoria primaria, sia l'a-diacente area di associazione per quellaspecifica modalità sensoriale. Questearee erano le sole a essere attivate uni-formemente dall'afferenza sensoriale,suggerendo che forme di memoria a spe-cifica modalità sono localizzate nella cor-teccia di associazione specifica per quellamodalità.
36
37
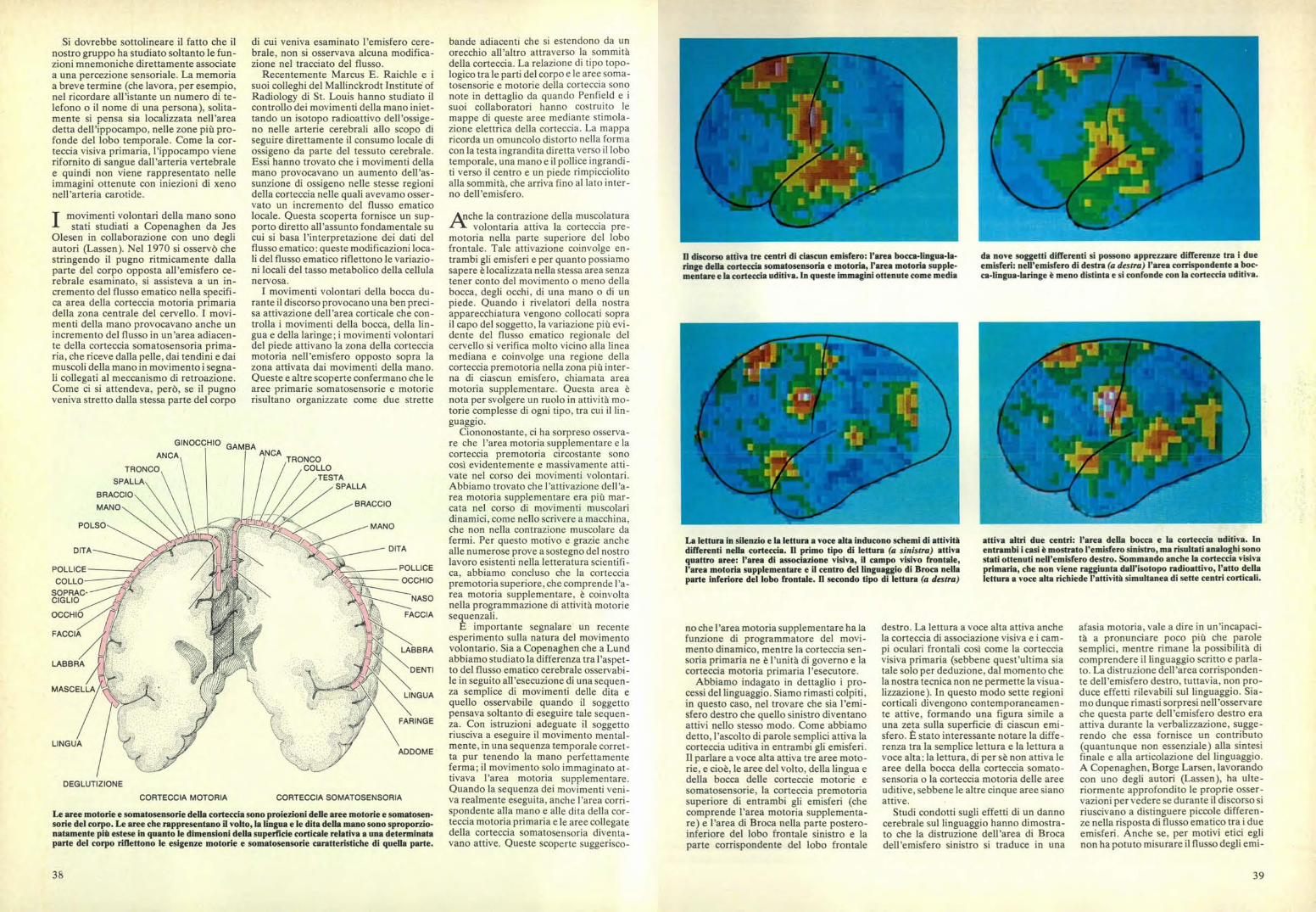
Il discorso attiva tre centri di ciascun emisfero: l'area bocca-lingua-la-ringe della corteccia somatosensoria e motoria, l'area motoria supple-mentare e la corteccia uditiva. In queste immagini ottenute come media
da nove soggetti differenti si possono apprezzare differenze tra i dueemisferi: nell'emisfero di destra (a destra) l'area corrispondente a boc-ca-lingua-laringe è meno distinta e si confonde con la corteccia uditiva.
La lettura in silenzio e la lettura a v oce alta inducono schemi di attivitàdifferenti nella corteccia. Il primo tipo di lettura (a sinistra) attivaquattro aree: l'area di associazione visiva, il campo visivo frontale,l'area motoria supplementare e il centro del linguaggio di Broca nellaparte inferiore del lobo frontale. Il secondo tipo di lettura (a destra)
attiva altri due centri: l'area della bocca e la corteccia uditiv a. Inentrambi i casi è mostrato l'emisfero sinistro, ma risultati analoghi sonostati ottenuti nell'emisfero destro. Sommando anche la corteccia v isivaprimaria, che non viene raggiunta dall'isotopo radioattivo, l'atto dellalettura a voce alta richiede l'attività simultanea di sette centri corticali.
DEGLUTIZIONE
CORTECCIA MOTORIA
CORTECCIA SOMATOSENSORIA
Le aree motorie e somatosensorie della corteccia sono proiezioni delle aree motorie e somatosen-sorie del corpo. Le aree che rappresentano il volto, la lingua e le dita della mano sono sproporzio-natamente più estese in quanto le dimensioni della superficie corticale relativa a una determinataparte del corpo riflettono le esigenze motorie e somatosensorie caratteristiche di quella parte.
GINOCCHIO GAMBA
ANCA
TRONCO
SPALLA
BRACCIO
MANO
POLLICE
COLLOSOPRAC-CIGLIO k,OCCHIO
FACCIA
LABBRA
MASCELLA
LINGUA
ANCATRONCO
COLLOTESTA
SPALLA
BRACCIO
MANO
DITA
POLLICE
OCCHIO'S\
LABBRA
DENTI
LINGUA
FARINGE
ADDOME
Si dovrebbe sottolineare il fatto che ilnostro gruppo ha studiato soltanto le fun-zioni mnemoniche direttamente associatea una percezione sensoriale. La memoriaa breve termine (che lavora, per esempio,nel ricordare all'istante un numero di te-lefono o il nome di una persona), solita-mente si pensa sia localizzata nell'areadetta dell'ippocampo, nelle zone più pro-fonde del lobo temporale. Come la cor-teccia visiva primaria, l'ippocampo vienerifornito di sangue dall'arteria vertebralee quindi non viene rappresentato nelleimmagini ottenute con iniezioni di xenonell'arteria carotide.
IT movimenti volontari della mano sono
stati studiati a Copenaghen da JesOlesen in collaborazione con uno degliautori (Lassen). Nel 1970 si osservò chestringendo il pugno ritmicamente dallaparte del corpo opposta all'emisfero ce-rebrale esaminato, si assisteva a un in-cremento del flusso ematico nella specifi-ca area della corteccia motoria primariadella zona centrale del cervello. I movi-menti della mano provocavano anche unincremento del flusso in un'area adiacen-te della corteccia somatosensoria prima-ria, che riceve dalla pelle, dai tendini e daimuscoli della mano in movimento i segna-li collegati al meccanismo di retroazione.Come ci si attendeva, però, se il pugnoveniva stretto dalla stessa parte del corpo
di cui veniva esaminato l'emisfero cere-brale, non si osservava alcuna modifica-zione nel tracciato del flusso.
Recentemente Marcus E. Raichle e isuoi colleghi del Mallinckrodt Institutè ofRadiology di St. Louis hanno studiato ilcontrollo dei movimenti della mano iniet-tando un isotopo radioattivo dell'ossige-no nelle arterie cerebrali allo scopo diseguire direttamente il consumo locale diossigeno da parte del tessuto cerebrale.Essi hanno trovato che i movimenti dellamano provocavano un aumento dell'as-sunzione di ossigeno nelle stesse regionidella corteccia nelle quali avevamo osser-vato un incremento del flusso ematicolocale. Questa scoperta fornisce un sup-porto diretto all'assunto fondamentale sucui si basa l'interpretazione dei dati delflusso ematico: queste modificazioni loca-li del flusso ematico riflettono le variazio-ni locali del tasso metabolico della cellulanervosa.
I movimenti volontari della bocca du-rante il discorso provocano una ben preci-sa attivazione dell'area corticale che con-trolla i movimenti della bocca, della lin-gua e della laringe ; i movimenti volontaridel piede attivano la zona della cortecciamotoria nell'emisfero opposto sopra lazona attivata dai movimenti della mano.Queste e altre scoperte confermano che learee primarie somatosensorie e motorierisultano organizzate come due strette
bande adiacenti che si estendono da unorecchio all'altro attraverso la sommitàdella corteccia. La relazione di tipo topo-logico tra le parti del corpo e le aree soma-tosensorie e motorie della corteccia sononote in dettaglio da quando Penfield e isuoi collaboratori hanno costruito lemappe di queste aree mediante stimola-zione elettrica della corteccia. La mapparicorda un omuncolo distorto nella formacon la testa ingrandita diretta verso il lobotemporale, una mano e il pollice ingrandi-ti verso il centro e un piede rimpicciolitoalla sommità, che arriva fino al lato inter-no dell'emisfero.
Ache la contrazione della muscolaturavolontaria attiva la corteccia pre-
motoria nella parte superiore del lobofrontale. Tale attivazione coinvolge en-trambi gli emisferi e per quanto possiamosapere è localizzata nella stessa area senzatener conto del movimento o meno dellabocca, degli occhi, di una mano o di unpiede. Quando i rivelatori della nostraapparecchiatura vengono collocati soprail capo del soggetto, la variazione più evi-dente del flusso ematico regionale delcervello si verifica molto vicino alla lineamediana e coinvolge una regione dellacorteccia premotoria nella zona più inter-na di ciascun emisfero, chiamata areamotoria supplementare. Questa area ènota per svolgere un ruolo in attività mo-torie complesse di ogni tipo, tra cui il lin-guaggio.
Ciononostante, ci ha sorpreso osserva-re che l'area motoria supplementare e lacorteccia premotoria circostante sonocosì evidentemente e massivamente atti-vate nel corso dei movimenti volontari.Abbiamo trovato che l'attivazione dell'a-rea motoria supplementare era più mar-cata nel corso di movimenti muscolaridinamici, come nello scrivere a macchina,che non nella contrazione muscolare dafermi. Per questo motivo e grazie anchealle numerose prove a sostegno del nostrolavoro esistenti nella letteratura scientifi-ca, abbiamo concluso che la cortecciapremotoria superiore, che comprende l'a-rea motoria supplementare, è coinvoltanella programmazione di attività motoriesequenzali.
E importante segnalare un recenteesperimento sulla natura del movimentovolontario. Sia a Copenaghen che a Lundabbiamo studiato la differenza tra l'aspet-to del flusso ematico cerebrale osservabi-le in seguito all'esecuzione di una sequen-za semplice di movimenti delle dita equello osservabile quando il soggettopensava soltanto di eseguire tale sequen-za. Con istruzioni adeguate il soggettoriusciva a eseguire il movimento mental-mente, in una sequenza temporale corret-ta pur tenendo la mano perfettamenteferma; il movimento solo immaginato at-tivava l'area motoria supplementare.Quando la sequenza dei movimenti veni-va realmente eseguita, anche l'area corri-spondente alla mano e alle dita della cor-teccia motoria primaria e le aree collegatedella corteccia somatosensoria diventa-vano attive. Queste scoperte suggerisco-
no che l'area motoria supplementare ha lafunzione di programmatore del movi-mento dinamico, mentre la corteccia sen-soria primaria ne è l'unità di governo e lacorteccia motoria primaria l'esecutore.
Abbiamo indagato in dettaglio i pro-cessi del linguaggio. Siamo rimasti colpiti,in questo caso, nel trovare che sia l'emi-sfero destro che quello sinistro diventanoattivi nello stesso modo. Come abbiamodetto, l'ascolto di parole semplici attiva lacorteccia uditiva in entrambi gli emisferi.Il parlare a voce alta attiva tre aree moto -rie, e cioè, le aree del volto, della lingua edella bocca delle corteccie motorie esomatosensorie, la corteccia premotoriasuperiore di entrambi gli emisferi (checomprende l'area motoria supplementa-re) e l'area di Broca nella parte postero-inferiore del lobo frontale sinistro e laparte corrispondente del lobo frontale
destro. La lettura a voce alta attiva anchela corteccia di associazione visiva e i cam-pi oculari frontali così come la cortecciavisiva primaria (sebbene quest'ultima siatale solo per deduzione, dal momento chela nostra tecnica non ne permette la visua-lizzazione). In questo modo sette regionicorticali divengono contemporaneamen-te attive, formando una figura simile auna zeta sulla superficie di ciascun emi-sfero. E stato interessante notare la diffe-renza tra la semplice lettura e la lettura avoce alta: la lettura, di per sè non attiva learee della bocca della corteccia somato-sensoria o la corteccia motoria delle areeuditive, sebbene le altre cinque aree sianoattive.
Studi condotti sugli effetti di un dannocerebrale sul linguaggio hanno dimostra-to che la distruzione dell'area di Brocadell'emisfero sinistro si traduce in una
afasia motoria, vale a dire in un'incapaci-tà a pronunciare poco più che parolesemplici, mentre rimane la possibilità dicomprendere il linguaggio scritto e parla-to. La distruzione dell'area corrisponden-te dell'emisfero destro, tuttavia, non pro-duce effetti rilevabili sul linguaggio. Sia-mo dunque rimasti sorpresi nell'osservareche questa parte dell'emisfero destro eraattiva durante la verbalizzazione, sugge-rendo che essa fornisce un contributo(quantunque non essenziale) alla sintesifinale e alla articolazione del linguaggio.A Copenaghen, Borge Larsen, lavorandocon uno degli autori (Lassen), ha ulte-riormente approfondito le proprie osser-vazioni per vedere se durante il discorso siriuscivano a distinguere piccole differen-ze nella risposta di flusso ematico tra i dueemisferi. Anche se, per motivi etici eglinon ha potuto misurare il flusso degli emi-
NASO
FACCIA
38
39
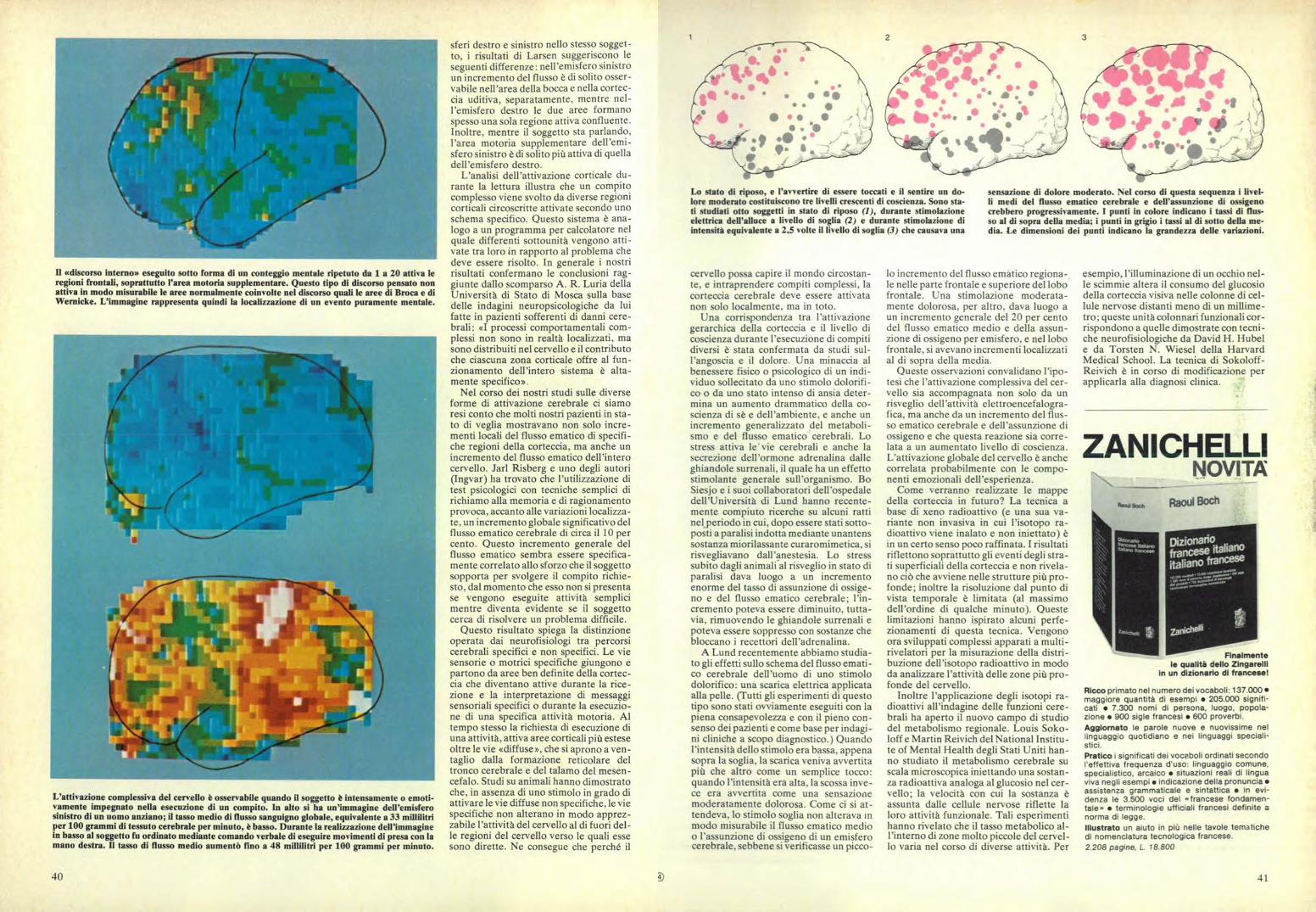
3
Lo stato di riposo, e l'av ertire di essere toccati e il sentire un do-lore moderato costituiscono tre livelli crescenti di coscienza. Sono sta-ti studiati otto soggetti in stato di riposo (1), durante stimolazioneelettrica dell'alluce a li% ello di soglia (2) e durante stimolazione diintensità equiv alente a 2,5 volte il livello di soglia (3) che causava una
sensazione di dolore moderato. Nel corso di questa sequenza i livel-li medi del flusso ematico cerebrale e dell'assunzione di ossigenocrebbero progressivamente, I punti in colore indicano i tassi di flus-so al di sopra della media; i punti in grigio i tassi al di sotto della me-dia. Le dimensioni dei punti indicano la grandezza delle variazioni.
risrmi Bort/
Finalmentele qualità dello Zingarelli
in un dizionario di francese!
Il «discorso interno» eseguito sotto forma di un conteggio mentale ripetuto da 1 a 20 attiva leregioni frontali, soprattutto l'area motoria supplementare. Questo tipo di discorso pensato nonattiva in modo misurabile le aree normalmente coinvolte nel discorso quali le aree di Broca e diWernicke. L'immagine rappresenta quindi la localizzazione di un evento puramente mentale.
L'attivazione complessiva del cervello è osservabile quando il soggetto è intensamente o emoti-vamente impegnato nella esecuzione di un compito. In alto si ha un'immagine dell'emisferosinistro di un uomo anziano; il tasso medio di flusso sanguigno globale, equivalente a 33 millilitriper 100 grammi di tessuto cerebrale per minuto, è basso. Durante la realizzazione dell'immaginein basso al soggetto fu ordinato mediante comando verbale di eseguire movimenti di presa con lamano destra. Il tasso di flusso medio aumentò fino a 48 millilitri per 100 grammi per minuto.
sferi destro e sinistro nello stesso sogget-to, i risultati di Larsen suggeriscono leseguenti differenze: nell'emisfero sinistroun incremento del flusso è di solito osser-vabile nell'area della bocca e nella cortec-cia uditiva, separatamente, mentre nel-l'emisfero destro le due aree formanospesso una sola regione attiva confluente.Inoltre, mentre il soggetto sta parlando,l'area motoria supplementare dell'emi-sfero sinistro è di solito più attiva di quelladell'emisfero destro.
L'analisi dell'attivazione corticale du-rante la lettura illustra che un compitocomplesso viene svolto da diverse regionicorticali circoscritte attivate secondo unoschema specifico. Questo sistema è ana-logo a un programma per calcolatore nelquale differenti sottounità vengono atti-vate tra loro in rapporto al problema chedeve essere risolto. In generale i nostririsultati confermano le conclusioni rag-giunte dallo scomparso A. R. Luna dellaUniversità di Stato di Mosca sulla basedelle indagini neuropsicologiche da luifatte in pazienti sofferenti di danni cere-brali: «I processi comportamentali com-plessi non sono in realtà localizzati, masono distribuiti nel cervello e il contributoche ciascuna zona corticale offre al fun-zionamento dell'intero sistema è alta-mente specifico».
Nel corso dei nostri studi sulle diverseforme di attivazione cerebrale ci siamoresi conto che molti nostri pazienti in sta-to di veglia mostravano non solo incre-menti locali del flusso ematico di specifi-che regioni della corteccia, ma anche unincremento del flusso ematico dell'interocervello. Jarl Risberg e uno degli autori(Ingvar) ha trovato che l'utilizzazione ditest psicologici con tecniche semplici dirichiamo alla memoria e di ragionamentoprovoca, accanto alle variazioni localizza-te, un incremento globale significativo delflusso ematico cerebrale di circa il 10 percento. Questo incremento generale delflusso ematico sembra essere specifica-mente correlato allo sforzo che il soggettosopporta per svolgere il compito richie-sto, dal momento che esso non si presentase vengono eseguite attività semplicimentre diventa evidente se il soggettocerca di risolvere un problema difficile.
Questo risultato spiega la distinzioneoperata dai neurofisiologi tra percorsicerebrali specifici e non specifici. Le viesensorie o motrici specifiche giungono epartono da aree ben definite della cortec-cia che diventano attive durante la rice-zione e la interpretazione di messaggisensoriali specifici o durante la esecuzio-ne di una specifica attività motoria. Altempo stesso la richiesta di esecuzione diuna attività, attiva aree corticali più esteseoltre le vie «diffuse», che si aprono a ven-taglio dalla formazione reticolare deltronco cerebrale e del talamo del mesen-cefalo. Studi su animali hanno dimostratoche, in assenza di uno stimolo in grado diattivare le vie diffuse non specifiche, le viespecifiche non alterano in modo apprez-zabile l'attività del cervello al di fuori del-le regioni del cervello verso le quali essesono dirette. Ne consegue che perché il
cervello possa capire il mondo circostan-te, e intraprendere compiti complessi, lacorteccia cerebrale deve essere attivatanon solo localmente, ma in toto.
Una corrispondenza tra l'attivazionegerarchica della corteccia e il livello dicoscienza durante l'esecuzione di compitidiversi è stata confermata da studi sul-l'angoscia e il dolore. Una minaccia albenessere fisico o psicologico di un indi-viduo sollecitato da uno stimolo dolorifi-co o da uno stato intenso di ansia deter-mina un aumento drammatico della co-scienza di sè e dell'ambiente, e anche unincremento generalizzato del metaboli-smo e del flusso ematico cerebrali. Lostress attiva le vie cerebrali e anche lasecrezione dell'ormone adrenalina dalleghiandole surrenali, il quale ha un effettostimolante generale sull'organismo. BoSiesjo e i suoi collaboratori dell'ospedaledell'Università di Lund hanno recente-mente compiuto ricerche su alcuni rattinel periodo in cui, dopo essere stati sotto-posti a paralisi indotta mediante unantenssostanza miorilassante curaromimetica, sirisvegliavano dall'anestesia. Lo stresssubito dagli animali al risveglio in stato diparalisi dava luogo a un incrementoenorme del tasso di assunzione di ossige-no e del flusso ematico cerebrale; l'in-cremento poteva essere diminuito, tutta-via, rimuovendo le ghiandole surrenali epoteva essere soppresso con sostanze chebloccano i recettori dell'adrenalina.
A Lund recentemente abbiamo studia-to gli effetti sullo schema del flusso emati-co cerebrale dell'uomo di uno stimolodolorifico: una scarica elettrica applicataalla pelle. (Tutti gli esperimenti di questotipo sono stati ovviamente eseguiti con lapiena consapevolezza e con il pieno con-senso dei pazienti e come base per indagi-ni cliniche a scopo diagnostico.) Quandol'intensità dello stimolo era bassa, appenasopra la soglia, la scarica veniva avvertitapiù che altro come un semplice tocco:quando l'intensità era alta, la scossa inve-ce era avvertita come una sensazionemoderatamente dolorosa. Come ci si at-tendeva, lo stimolo soglia non alterava inmodo misurabile il flusso ematico medioo l'assunzione di ossigeno di un emisferocerebrale, sebbene si verificasse un picco-
lo incremento del flusso ematico regiona-le nelle parte frontale e superiore del lobofrontale. Una stimolazione moderata-mente dolorosa, per altro, dava luogo aun incremento generale del 20 per centodel flusso ematico medio e della assun-zione di ossigeno per emisfero, e nel lobofrontale, si avevano incrementi localizzatial di sopra della media.
Queste osservazioni convalidano l'ipo-tesi che l'attivazione complessiva del cer-vello sia accompagnata non solo da unrisveglio dell'attività elettroencefalogra-fica, ma anche da un incremento del flus-so ematico cerebrale e dell'assunzione diossigeno e che questa reazione sia corre-lata a un aumentato livello di coscienza.L'attivazione globale del cervello è anchecorrelata probabilmente con le compo-nenti emozionali dell'esperienza.
Come verranno realizzate le mappedella corteccia in futuro? La tecnica abase di xeno radioattivo (e una sua va-riante non invasiva in cui l'isotopo ra-dioattivo viene inalato e non iniettato) èin un certo senso poco raffinata. I risultatiriflettono soprattutto gli eventi degli stra-ti superficiali della corteccia e non rivela-no ciò che avviene nelle strutture più pro-fonde; inoltre la risoluzione dal punto divista temporale è limitata (al massimodell'ordine di qualche minuto). Questelimitazioni hanno ispirato alcuni perfe-zionamenti di questa tecnica. Vengonoora sviluppati complessi apparati a multi-rivelatori per la misurazione della distri-buzione dell'isotopo radioattivo in mododa analizzare l'attività delle zone più pro-fonde del cervello.
Inoltre l'applicazione degli isotopi ra-dioattivi all'indagine delle funzioni cere-brali ha aperto il nuovo campo di studiodel metabolismo regionale. Louis Soko-loff e Martin Reivich del National Institu-te of Mental Health degli Stati Uniti han-no studiato il metabolismo cerebrale suscala microscopica iniettando una sostan-za radioattiva analoga al glucosio nel cer-vello; la velocità con cui la sostanza èassunta dalle cellule nervose riflette laloro attività funzionale. Tali esperimentihanno rivelato che il tasso metabolico al-l'interno di zone molto piccole del cervel-lo varia nel corso di diverse attività. Per
esempio, l'illuminazione di un occhio nel-le scimmie altera il consumo del glucosiodella corteccia visiva nelle colonne di cel-lule nervose distanti meno di un millime-tro; queste unità colonnari funzionali cor-rispondono a quelle dimostrate con tecni-che neurofisiologiche da David H. Hubele da Torsten N. Wiesel della HarvardMedical School. La tecnica di Sokoloff-Reivich è in corso di modificazione perapplicarla alla diagnosi clinica.
7ANICHELLINOVITA
Ricco primato nel numero dei vocaboli: 137.000maggiore quantità di esempi • 205.000 signifi-cati • 7.300 nomi di persona, luogo, popola-zione • 900 sigle francesi • 600 proverbi.Aggiornato le parole nuove e nuovissime nellinguaggio quotidiano e nei linguaggi speciali-
Pratico i significati dei voceboli ordinati secondol'effettiva frequenza d'uso: linguaggio comune,specialistico, arcaico • situazioni reali di linguaviva negli esempi • indicazione della pronuncia •assistenza grammaticale e sintattica • in evi-denza le 3.500 voci del francese fondamen-tale » • terminologie ufficiali francesi definite anorma di legge.Illustrato un aiuto in più nelle tavole tematichedi nomenclatura tecnologica francese.2.208 pagine, L. 18.800
40
D
41





![Infezioni Del Circolo Ematico[CCLA]](https://static.fdocumenti.com/doc/165x107/55cf9d58550346d033ad3904/infezioni-del-circolo-ematicoccla.jpg)