Ramapithecus - Kataweb
4
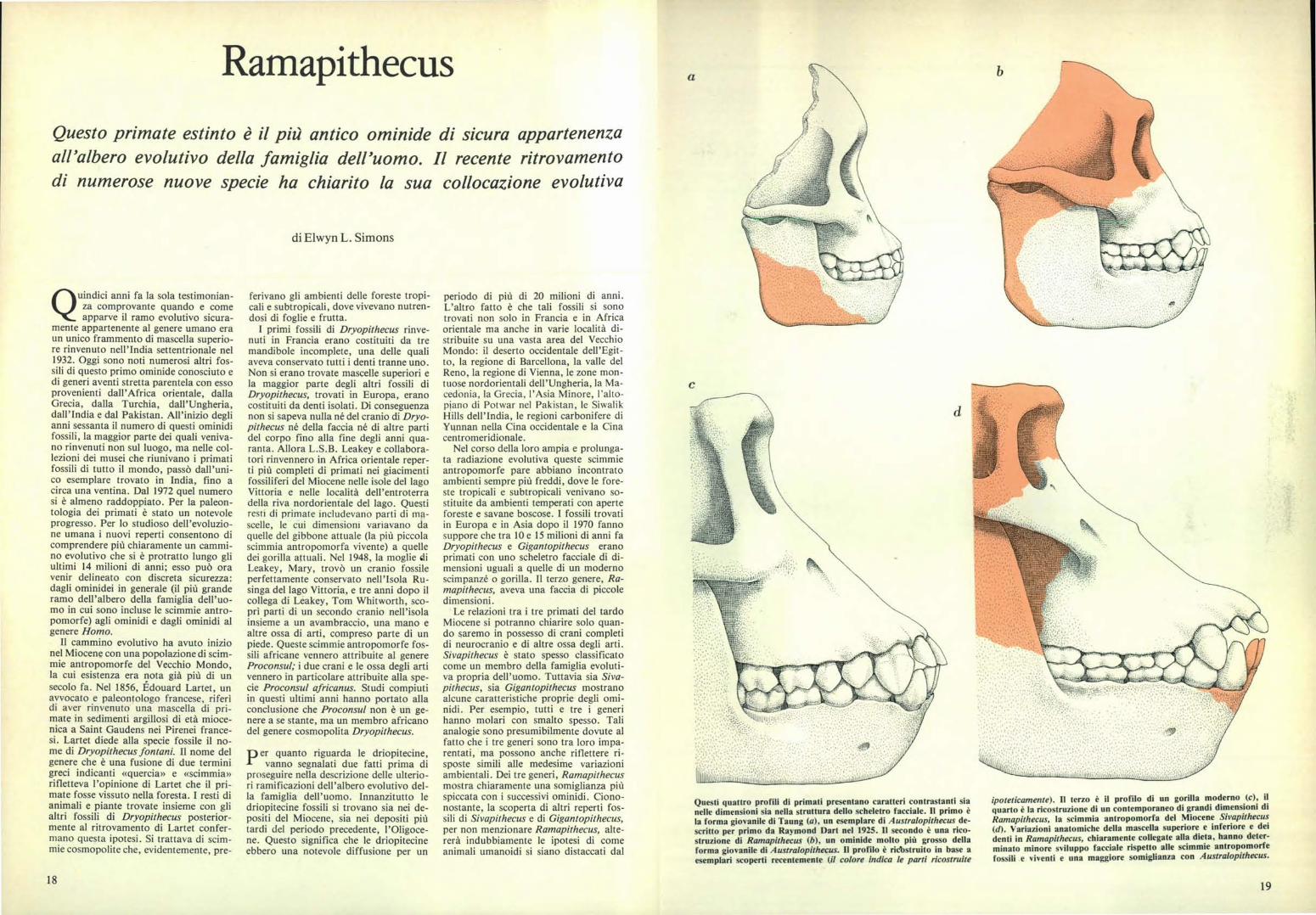
ipoteticamente). Il terzo è il profilo di un gorilla moderno (c), il quarto è la ricostruzione di un contemporaneo di grandi dimensioni di Ramapithecus, la scimmia antropomorfa del Miocene Sivapithecus (d). Variazioni anatomiche della mascella superiore e inferiore e dei denti in Ramapithecus, chiaramente collegate alla dieta, hanno deter- minato minore sviluppo facciale rispetto alle scimmie antropomorfe fossili e viventi e una maggiore somiglianza con Australopithecus. Q uindici anni fa la sola testimonian- za comprovante quando e come apparve il ramo evolutivo sicura- mente appartenente al genere umano era un unico frammento di mascella superio- re rinvenuto nell'India settentrionale nel 1932. Oggi sono noti numerosi altri fos- sili di questo primo ominide conosciuto e di generi aventi stretta parentela con esso provenienti dall'Africa orientale, dalla Grecia, dalla Turchia, dall'Ungheria, dall'India e dal Pakistan. All'inizio degli anni sessanta il numero di questi ominidi fossili, la maggior parte dei quali veniva- no rinvenuti non sul luogo, ma nelle col- lezioni dei musei che riunivano i primati fossili di tutto il mondo, passò dall'uni- co esemplare trovato in India, fino a circa una ventina. Dal 1972 quel numero si è almeno raddoppiato. Per la paleon- tologia dei primati è stato un notevole progresso. Per lo studioso dell'evoluzio- ne umana i nuovi reperti consentono di comprendere più chiaramente un cammi- no evolutivo che si è protratto lungo gli ultimi 14 milioni di anni; esso può ora venir delineato con discreta sicurezza: dagli ominidei in generale (il più grande ramo dell'albero della famiglia dell'uo- mo in cui sono incluse le scimmie antro- pomorfe) agli ominidi e dagli ominidi al genere Homo. Il cammino evolutivo ha avuto inizio nel Miocene con una popolazione di scim- mie antropomorfe del Vecchio Mondo, la cui esistenza era nota già più di un secolo fa. Nel 1856, tdouard Lartet, un avvocato e paleontologo francese, riferì di aver rinvenuto una mascella di pri- mate in sedimenti argillosi di età mioce- nica a Saint Gaudens nei Pirenei france- si. Lartet diede alla specie fossile il no- me di Dryopithecus fontani. Il nome del genere che è una fusione di due termini greci indicanti «quercia» e «scimmia» rifletteva l'opinione di Lartet che il pri- mate fosse vissuto nella foresta. I resti di animali e piante trovate insieme con gli altri fossili di Dryopithecus posterior- mente al ritrovamento di Lartet confer- mano questa ipotesi. Si trattava di scim- mie cosmopolite che, evidentemente, pre- ferivano gli ambienti delle foreste tropi- cali e subtropicali, dove vivevano nutren- dosi di foglie e frutta. I primi fossili di Dryopithecus rinve- nuti in Francia erano costituiti da tre mandibole incomplete, una delle quali aveva conservato tutti i denti tranne uno. Non si erano trovate mascelle superiori e la maggior parte degli altri fossili di Dryopithecus, trovati in Europa, erano costituiti da denti isolati. Di conseguenza non si sapeva nulla né del cranio di Dryo- pithecus né della faccia né di altre parti del corpo fino alla fine degli anni qua- ranta. Allora L.S.B. Leakey e collabora- tori rinvennero in Africa orientale reper- ti più completi di primati nei giacimenti fossiliferi del Miocene nelle isole del lago Vittoria e nelle località dell'entroterra della riva nordorientale del lago. Questi resti di primate includevano parti di ma- scelle, le cui dimensioni variavano da quelle del gibbone attuale (la più piccola scimmia antropomorfa vivente) a quelle dei gorilla attuali. Nel 1948, la moglie di Leakey, Mary, trovò un cranio fossile perfettamente conservato nell'Isola Ru- singa del lago Vittoria, e tre anni dopo il collega di Leakey, Tom Whitworth, sco- prì parti di un secondo cranio nell'isola insieme a un avambraccio, una mano e altre ossa di arti, compreso parte di un piede. Queste scimmie antropomorfe fos- sili africane vennero attribuite al genere Proconsul; i due crani e le ossa degli arti vennero in particolare attribuite alla spe- cie Proconsul africanus. Studi compiuti in questi ultimi anni hanno portato alla conclusione che Proconsul non è un ge- nere a se stante, ma un membro africano del genere cosmopolita Dryopithecus. P er quanto riguarda le driopitecine, vanno segnalati due fatti prima di proseguire nella descrizione delle ulterio- ri ramificazioni dell'albero evolutivo del- la famiglia dell'uomo. Innanzitutto le driopitecine fossili si trovano sia nei de- positi del Miocene, sia nei depositi più tardi del periodo precedente, l'Oligoce- ne. Questo significa che le driopitecine ebbero una notevole diffusione per un periodo di più di 20 milioni di anni. L'altro fatto è che tali fossili si sono trovati non solo in Francia e in Africa orientale ma anche in varie località di- stribuite su una vasta area del Vecchio Mondo: il deserto occidentale dell'Egit- to, la regione di Barcellona, la valle del Reno, la regione di Vienna, le zone mon- tuose nordorientali dell'Ungheria, la Ma- cedonia, la Grecia, l'Asia Minore, l'alto- piano di Potwar nel Pakistan, le Siwalik Hills dell'India, le regioni carbonifere di Yunnan nella Cina occidentale e la Cina centromeridionale. Nel corso della loro ampia e prolunga- ta radiazione evolutiva queste scimmie antropomorfe pare abbiano incontrato ambienti sempre più freddi, dove le fore- ste tropicali e subtropicali venivano so- stituite da ambienti temperati con aperte foreste e savane boscose. I fossili trovati in Europa e in Asia dopo il 1970 fanno suppore che tra 10 e 15 milioni di anni fa Dryopithecus e Gigantopithecus erano primati con uno scheletro facciale di di- mensioni uguali a quelle di un moderno scimpanzé o gorilla. Il terzo genere, Ra- mapithecus, aveva una faccia di piccole dimensioni. Le relazioni tra i tre primati del tardo Miocene si potranno chiarire solo quan- do saremo in possesso di crani completi di neurocranio e di altre ossa degli arti. Sivapithecus è stato spesso classificato come un membro della famiglia evoluti- va propria dell'uomo. Tuttavia sia Siva- pithecus, sia Gigantopithecus mostrano alcune caratteristiche proprie degli orni- nidi. Per esempio, tutti e tre i generi hanno molari con smalto spesso. Tali analogie sono presumibilmente dovute al fatto che i tre generi sono tra loro impa- rentati, ma possono anche riflettere ri- sposte simili alle medesime variazioni ambientali. Dei tre generi, Ramapithecus mostra chiaramente una somiglianza più spiccata con i successivi ominidi. Ciono- nostante, la scoperta di altri reperti fos- sili di Sivapithecus e di Gigantopithecus, per non menzionare Ramapithecus, alte- rerà indubbiamente le ipotesi di come animali umanoidi si siano distaccati dal Questi quattro profili di primati presentano caratteri contrastanti sia nelle dimensioni sia nella struttura dello scheletro facciale. Il primo è la forma gio. anile di Taung (a), un esemplare di Australopithecus de- scritto per primo da Raymond Dart nel 1925. Il secondo è una rico- struzione di Ramapithecus (b), un ominide molto più grosso della forma giovanile di Australopithecus. Il profilo è ricbstruito in base a esemplari scoperti recentemente (il colore indica le parti ricostruite Ramapithecus a Questo primate estinto è il più antico ominide di sicura appartenenza all'albero evolutivo della famiglia dell'uomo. Il recente ritrovamento di numerose nuove specie ha chiarito la sua collocazione evolutiva di Elwyn L. Simons 18 19
Transcript of Ramapithecus - Kataweb
ipoteticamente). Il terzo è il profilo di un gorilla moderno (c), il
quarto è la ricostruzione di un contemporaneo di grandi dimensioni diRamapithecus, la scimmia antropomorfa del Miocene Sivapithecus(d). Variazioni anatomiche della mascella superiore e inferiore e deidenti in Ramapithecus, chiaramente collegate alla dieta, hanno deter-minato minore sviluppo facciale rispetto alle scimmie antropomorfefossili e viventi e una maggiore somiglianza con Australopithecus.
Q
uindici anni fa la sola testimonian-za comprovante quando e comeapparve il ramo evolutivo sicura-
mente appartenente al genere umano eraun unico frammento di mascella superio-re rinvenuto nell'India settentrionale nel1932. Oggi sono noti numerosi altri fos-sili di questo primo ominide conosciuto edi generi aventi stretta parentela con essoprovenienti dall'Africa orientale, dallaGrecia, dalla Turchia, dall'Ungheria,dall'India e dal Pakistan. All'inizio deglianni sessanta il numero di questi ominidifossili, la maggior parte dei quali veniva-no rinvenuti non sul luogo, ma nelle col-lezioni dei musei che riunivano i primatifossili di tutto il mondo, passò dall'uni-co esemplare trovato in India, fino acirca una ventina. Dal 1972 quel numerosi è almeno raddoppiato. Per la paleon-tologia dei primati è stato un notevoleprogresso. Per lo studioso dell'evoluzio-ne umana i nuovi reperti consentono dicomprendere più chiaramente un cammi-no evolutivo che si è protratto lungo gliultimi 14 milioni di anni; esso può oravenir delineato con discreta sicurezza:dagli ominidei in generale (il più granderamo dell'albero della famiglia dell'uo-mo in cui sono incluse le scimmie antro-pomorfe) agli ominidi e dagli ominidi algenere Homo.
Il cammino evolutivo ha avuto inizionel Miocene con una popolazione di scim-mie antropomorfe del Vecchio Mondo,la cui esistenza era nota già più di unsecolo fa. Nel 1856, tdouard Lartet, unavvocato e paleontologo francese, riferìdi aver rinvenuto una mascella di pri-mate in sedimenti argillosi di età mioce-nica a Saint Gaudens nei Pirenei france-si. Lartet diede alla specie fossile il no-me di Dryopithecus fontani. Il nome delgenere che è una fusione di due terminigreci indicanti «quercia» e «scimmia»rifletteva l'opinione di Lartet che il pri-mate fosse vissuto nella foresta. I resti dianimali e piante trovate insieme con glialtri fossili di Dryopithecus posterior-mente al ritrovamento di Lartet confer-mano questa ipotesi. Si trattava di scim-mie cosmopolite che, evidentemente, pre-
ferivano gli ambienti delle foreste tropi-cali e subtropicali, dove vivevano nutren-dosi di foglie e frutta.
I primi fossili di Dryopithecus rinve-nuti in Francia erano costituiti da tremandibole incomplete, una delle qualiaveva conservato tutti i denti tranne uno.Non si erano trovate mascelle superiori ela maggior parte degli altri fossili diDryopithecus, trovati in Europa, eranocostituiti da denti isolati. Di conseguenzanon si sapeva nulla né del cranio di Dryo-pithecus né della faccia né di altre partidel corpo fino alla fine degli anni qua-ranta. Allora L.S.B. Leakey e collabora-tori rinvennero in Africa orientale reper-ti più completi di primati nei giacimentifossiliferi del Miocene nelle isole del lagoVittoria e nelle località dell'entroterradella riva nordorientale del lago. Questiresti di primate includevano parti di ma-scelle, le cui dimensioni variavano daquelle del gibbone attuale (la più piccolascimmia antropomorfa vivente) a quelledei gorilla attuali. Nel 1948, la moglie diLeakey, Mary, trovò un cranio fossileperfettamente conservato nell'Isola Ru-singa del lago Vittoria, e tre anni dopo ilcollega di Leakey, Tom Whitworth, sco-prì parti di un secondo cranio nell'isolainsieme a un avambraccio, una mano ealtre ossa di arti, compreso parte di unpiede. Queste scimmie antropomorfe fos-sili africane vennero attribuite al genereProconsul; i due crani e le ossa degli artivennero in particolare attribuite alla spe-cie Proconsul africanus. Studi compiutiin questi ultimi anni hanno portato allaconclusione che Proconsul non è un ge-nere a se stante, ma un membro africanodel genere cosmopolita Dryopithecus.
Per quanto riguarda le driopitecine,
vanno segnalati due fatti prima diproseguire nella descrizione delle ulterio-ri ramificazioni dell'albero evolutivo del-la famiglia dell'uomo. Innanzitutto ledriopitecine fossili si trovano sia nei de-positi del Miocene, sia nei depositi piùtardi del periodo precedente, l'Oligoce-ne. Questo significa che le driopitecineebbero una notevole diffusione per un
periodo di più di 20 milioni di anni.L'altro fatto è che tali fossili si sonotrovati non solo in Francia e in Africaorientale ma anche in varie località di-stribuite su una vasta area del VecchioMondo: il deserto occidentale dell'Egit-to, la regione di Barcellona, la valle delReno, la regione di Vienna, le zone mon-tuose nordorientali dell'Ungheria, la Ma-cedonia, la Grecia, l'Asia Minore, l'alto-piano di Potwar nel Pakistan, le SiwalikHills dell'India, le regioni carbonifere diYunnan nella Cina occidentale e la Cinacentromeridionale.
Nel corso della loro ampia e prolunga-ta radiazione evolutiva queste scimmieantropomorfe pare abbiano incontratoambienti sempre più freddi, dove le fore-ste tropicali e subtropicali venivano so-stituite da ambienti temperati con aperteforeste e savane boscose. I fossili trovatiin Europa e in Asia dopo il 1970 fannosuppore che tra 10 e 15 milioni di anni faDryopithecus e Gigantopithecus eranoprimati con uno scheletro facciale di di-mensioni uguali a quelle di un modernoscimpanzé o gorilla. Il terzo genere, Ra-mapithecus, aveva una faccia di piccoledimensioni.
Le relazioni tra i tre primati del tardoMiocene si potranno chiarire solo quan-do saremo in possesso di crani completidi neurocranio e di altre ossa degli arti.Sivapithecus è stato spesso classificatocome un membro della famiglia evoluti-va propria dell'uomo. Tuttavia sia Siva-pithecus, sia Gigantopithecus mostranoalcune caratteristiche proprie degli orni-nidi. Per esempio, tutti e tre i generihanno molari con smalto spesso. Talianalogie sono presumibilmente dovute alfatto che i tre generi sono tra loro impa-rentati, ma possono anche riflettere ri-sposte simili alle medesime variazioniambientali. Dei tre generi, Ramapithecusmostra chiaramente una somiglianza piùspiccata con i successivi ominidi. Ciono-nostante, la scoperta di altri reperti fos-sili di Sivapithecus e di Gigantopithecus,per non menzionare Ramapithecus, alte-rerà indubbiamente le ipotesi di comeanimali umanoidi si siano distaccati dal
Questi quattro profili di primati presentano caratteri contrastanti sianelle dimensioni sia nella struttura dello scheletro facciale. Il primo è
la forma gio. anile di Taung (a), un esemplare di Australopithecus de-scritto per primo da Raymond Dart nel 1925. Il secondo è una rico-struzione di Ramapithecus (b), un ominide molto più grosso dellaforma giovanile di Australopithecus. Il profilo è ricbstruito in base aesemplari scoperti recentemente (il colore indica le parti ricostruite
Ramapithecus a
Questo primate estinto è il più antico ominide di sicura appartenenzaall'albero evolutivo della famiglia dell'uomo. Il recente ritrovamentodi numerose nuove specie ha chiarito la sua collocazione evolutiva
di Elwyn L. Simons
18 19

GENERI
PONGO (ORANGO)GRANDIANTROPOMORFE
EVIVENTI PAN (SCIMPANZE)
GORILLA (GORILLA)
GRANDI SCIMMIEANTROPOMORFE ESTINT
DRYOPITHECUSSIVAPITHECUSGIGANTOPITHECUS
UOMINI ATTUALI {HOMO (UOMO)
FORMEPREUMANE ESTINTE
{RAMAPITHECUSAUSTRALOPITHECUS
I primati evoluti dell'Oligocene, Miocene e dei periodi più recenticomprendevano gruppi numerosi e cosmopoliti di grandi scimmieantropomorfe: le driopitecine (punti neri). La maggior parte di esse sipuò attribuire a due generi: Dryopithecus, rinvenuto principalmentein Africa ed Europa, e Sivapithecus, rinvenuto principalmente nellaregione orientale dell'Asia Minore. Queste grandi scimmie antropo-
morfe furono diffuse per più di 25 milioni di anni. Da esse originò lagrande scimmia antropomorfa Gigantopithecus (cerchi neri) e gli orni-nidi come Ramapithecus, un genere ora rinvenuto in cinque areedell'Eurasia e una dell'Africa orientale (crocette colorate). Come inRamapithecus, anche in Gigantopithecus sono riscontrabili cambia-menti anatomici delle mascelle e dei denti collegati con la dieta.
20 21
ramo delle scimmie antropomorfe. In o-gni caso, torniamo alle ramificazioni piùtarde dell'albero evolutivo della famigliadell'uomo, a partire da Ramapithecus.
Haritalyangar è il nome di un aggrega-to di villaggi in India situato circa 150chilometri a nord di Nuova Delhi nelleSiwalik Hills, un'area in cui affioranoletti fossiliferi del Miocene estendentisidall'India nordoccidentale al vicino Pa-kistan. Qui, nel 1932, G. Edward Lewis,un giovane studente diplomato alla YaleUniversity, che era giunto in questa re-gione accompagnato solo da un caval-lo da soma, scoprì alcuni fossili di quelleche egli chiamò «scimmie umanoidi».Egli attribuì uno dei fossili, una mascellasuperiore, a un nuovo genere e specieche chiamò Ramapithecus brevirostris. Ilnome del genere significa semplicemente«scimmia di Rama». Rama è un miticoprincipe, eroe di un poema epico india-no. Il nome della specie, scelto da Lewisera più significativo; il termine latino si-gnifica «dal muso corto», un caratteresomatico atipico per le scimmie. Lewisfu colpito da molti caratteri umanoididella mascella e dalla disposizione deidenti. Scrivendo la sua tesi di dottoratonel 1937, egli collocò il nuovo generenella famiglia Hominidae, la suddivisio-ne dell'ordine dei primati il cui unicorappresentante vivente è Homo sapiens.
T l successivo ritrovamento di un repertofossile di Ramapithecus non avvenne
fino al 1961. Scoperto da L.S.B. Leakeyvicino a Fort Ternan nel Kenya sudocci-dentale, includeva parti di entrambi i latidi una mascella superiore. Leakey, se-guendo la normale consuetudine, trattòla sua scoperta come se fosse un genere euna specie completamente nuova, dandoa esso il nome di Kenyapithecus wickeridal suo amico Fred Wicker, nella cuifattoria il fossile era stato rinvenuto.Sembra però probabile che Leakey rite-nesse che il ritrovamento di una mascellainferiore in diretta associazione sia conle mascelle superiori di Haritalyangar siacon quella di Fort Ternan o il confrontodiretto della sua scoperta con la specie diHaritalyangar, custodita a Yale, avreb-bero permesso di diminuire il numero digeneri provvisoriamente proposti.
Tuttavia, la successiva specie di Ra-mapithecus rinvenuta, dopo quella chefu portata alla luce negli scavi in Greciadurante la seconda guerra mondiale,ma che non fu formalmente descrittafino al 1972, fu attribuita a un nuovogenere e specie: Graecopithecus freybur-gi. Il nome della specie onorava Brunovon Freyburg, un geologo tedesco cheaveva estratto il fossile da uno strato delMiocene mentre era di guarnigione adAtene durante l'occupazione tedesca in
Grecia. Il reperto di von Freyburg eracostituito dalla parte dentale completadella mascella inferiore e, al tempo dellasua scoperta, vi erano anche tutti i denti.Era ritenuto relativamente di poca impor-tanza poiché un'altra scuola erroneamen-te identificava il fossile come una mandi-bola di Mesopithecus, una scimmia co-mune del Miocene. Prima che la speciefosse stata fotografata la maggior partedei denti erano saltati via e si erano persidurante un bombardamento su Berlinoin tempo di guerra.
Da aggiungere alle sempre più frequen-ti scoperte di fossili di Ramapithecus vifu una mandibola dissotterrata da un de-posito miocenico vicino a qandir, a circa65 chilometri a nord-est di Ankara in Tur-chia. Il ritrovamento avvenne nel 1973 ela specie fu descritta circa un anno dopo.A quel tempo fu attribuito a un altrogenere e a una specie completamentenuova: Sivapithecus alpani. Sivapithecus,o scimmia di Siva, era il nome che Lewisaveva dato a una delle specie da lui tro-vate in India; il nome della specie dellamandibola di Candir onorava il direttoredel Turkish Geological Survey.
Un gruppo più numeroso di fossili af-fini a Ramapithecus è stato inoltre sco-perto nei depositi carboniferi di età mio-cenica nei monti Rudabanya nell'Unghe-ria nordorientale. Le specie vennero alla
luce per un periodo di parecchi anni, manon furono descritte in dettaglio fino al1975. Sono state ascritte anch'esse a unnuovo genere e specie: il Rudapithecushungaricus.
Di tutti i reperti fossili di Ramapithe-cus recentemente scoperti e descritti, so-lo quelli rinvenuti in Pakistan a partiredal 1976 dal mio collega di Yale, DavidR. Pilbeam e dai suoi collaboratori, so-no stati riconosciuti sin dall'inizio comeappartenenti al genere Ramapithecus.Perfino un frammento di una mandibo-la, rinvenuta a Fort Ternan, riclassifica-to nel 1971 come Ramapithecus da P.Andrews del British Museum venne inizial-mente assegnato al genere Dryopithecus.
Non sempre questa disordinata deno-minazione dei reperti era dovuta esclusi-vamente a un'eccessiva tendenza alla sud-divisione. Ibrahim Tekkaya, il paleonto-logo turco che attribuì la mandibola diqandir al genere Sivapithecus, disponevadi poco materiale per il confronto. Laprincipale fonte d'informazione a sua di-sposizione era una discussione su unamandibola di Dryopithecus che era stataerroneamente attribuita al genere Rama-pithecus nel 1938. Questa erronea attri-buzione era stata ripetuta costantementenella letteratura scientifica e Tekkaya ri-teneva che la mancanza di somiglianzatra quella mandibola e il suo reperto laescludesse come specie di Ramapithecus.Egli e Andrews hanno pubblicato insie-me un rapporto che riclassifica corretta-mente il reperto come Ramapithecus.
Seguendo il medesimo presupposto,Miklos Kretzoi, il paleontologo unghere-se che chiamò i suoi nuovi reperti Ruda-pithecus, avrebbe potuto non usare que-sta denominazione, se la prima mandi-bola trovata in Ungheria nel 1969 nonfosse stata completamente erosa e incom-pleta. Quando venne in possesso dei suc-cessivi fossili di Rudabanya (che inclu-dono sia la mascella superiore sia l'infe-riore), la rassomiglianza con Ramapithe-cus risultò ben evidente. Kretzoi attual-mente riconosce questi fossili come fa-centi parte del gruppo delle ramapiteci-ne, anche se non possono essere ascritteal genere Ramapithecus.
Infine, l'eminente paleontologo G.H.R. von Koenigswald, che chiamò la man-dibola di Atene Graecopithecus, la sepa-rò dal genere Ramapithecus, perché man-dibole abbastanza complete di Ramapi-thecus erano allora ignote. Il primo buonesemplare ritrovato, la mandibola di
andir, non fu descritto fino al 1974.Perciò nel 1972 von Koenineswald nonpoteva aver scorto la stretta somiglianzache è evidente quando si confronta l'e-semplare di Atene con quelli di Fort Ter-nan e di Candir.
Vi sono ancora alstri resti di Ramapi-thecus «riabilitati». Per esempio, Pilbeame io, scrivendo nel 1965, seguimmo l'opi-nione universalmente accettata che gliominidi non hanno la «cresta scimmie-sca», che è caratteristica di tutte le scim-mie, antropomorfe comprese. La crestascimmiesca è un toro, o gibbosità oriz-zontale dell'osso, che si proietta verso
CATEGORIE PIÙ ELEVATE
SUPERFAMIGLIAOMINOIDEITUTTE LEGRANDI SCIMMIEANTROPOMORFEE TUTTE LEFORME UMANEE PREUMANE
l'interno a partire dalla faccia internadella mandibola. La sua presenza in unametà della mandibola proveniente daipressi di Domeli in Pakistan ci fece rite-nere sul momento che potesse essereesclusa come specie del genere Ramapi-thecus. Più tardi concludemmo che pro-tuberanze simili a creste sono presentisul lato interno della mandibola sia inRamapithecus sia nel più recente omini-de africano Australopithecus. Nel 1969Pilbeam riscattò la mandibola di Domelidalla sua immeritata oscurità, enumeròle sue caratteristiche da ominide e l'ag-giunse all'inventario dei resti fossili diRamapithecus.
Fino a questo momento sono state de-scritte due dozzine di mascelle assortite edi denti di Ramapithecus. Che cosa cidicono del più antico ominide antenatodell'uomo? La testimonianza ci è fornitada numerosi e chiari caratteri anatomici,che però conducono a un'inevitabile con-clusione: Ramapithecus si era adattato aun tipo di vita del tutto diverso da quelloseguito dalla maggior parte dei suoi pa-renti appartenenti al genere Dryopithe-cus che vivevano nelle foreste. Dove lemascelle e i denti riflettono quel tipo diadattamento, presentano somiglianze conl'ominide africano Australopithecus. Trai due, tuttavia, intercorre un grande las-so di tempo. Le specie di Ramapithecusnon risulta fossero diffuse in Eurasia inun periodo precedente a otto milioni dianni fa. I primi fossili di Australopithe-cus e di Homo appaiono essere quellitrovati a partire dal 1974 in Etiopia daDonald C. Johanson della Case WesternReserve University e in Tanzania da Ma-ry Leakey; di questi i più antichi risalgo-no a meno di quattro milioni di anni fa.
Per tornare ai dettagli anatomici, nel
I 1967, mentre stavo rivedendo la mor-fologia delle più note scimmie antropo-morfe fossili, trovai con mia sorpresache quelle vissute nel periodo miocenicoavevano un'arcata dentaria del tutto di-versa da quella tipica delle scimmie an-tropomorfe viventi. Tra le scimmie an-tropomorfe attuali l'arcata ha una formaa U, poiché i due lati della mascella so-no paralleli. Nelle scimmie antropomor-fe tipiche del Miocene l'arcata ha forma
a V, poiché i due lati della mascella di-vergono posteriormente.
Due anni dopo, E. Genet-Varcin, pre-cisò che l'arcata dentaria della maggiorparte delle mascelle di Autralopithecus èanch'essa conformata a V. (In entram-bi i gruppi l'apice della V è squadrataanteriormente.) Perciò il medesimo tipodi arcata è diffuso nei due gruppi sepa-rati tra loro da milioni di anni. L'arcatadentaria di Homo sapiens non è né con-formata a U né a V, ma piuttosto asemicerchio. Di conseguenza sia l'arcatasemicircolare dell'uomo moderno, siaquella a U delle scimmie antropomorfederivò presumibilmente da una origina-ria forma a V.
A che punto della variazione evolutivava posta la forma della mascella di Ra-mapithecus? La forma delle sue mascelleè intermedia: la fila di denti diverge di20 gradi dalla disposizione parallela.Quest'angolo è più ampio rispetto a quel-lo delle scimmie antropomorfe del Mio-cene (10 gradi) e più stretto rispetto aquello di Australopithecus (30 gradi).
Sia Ramapithecus sia Australopithecushanno una mandibola che è caratteristicaper quanto riguarda Io spessore; infatti,nella regione dei molari, Io spessore èuniforme dall'alto al basso. Questa ro-bustezza contrasta con quella della man-dibola delle attuali scimmie antropomor-fe, che è molto più sottile nella regionedei molari. I denti molari di Ramapithe-cus e di Australopithecus danno la stessaimpressione di robustezza. Sono grossi eappiattiti e, visti dall'alto, hanno i con-torni arrotondati. Diversamente dai mo-lari di scimpanzé e gorilla, e anche diDryopithecus in senso stretto (escluden-do Sivapithecus), hanno uno smalto spes-so e quindi sono più resistenti all'usura.I denti canini e incisivi sono più piccolidi quelli delle scimmie antropomorfe at-tuali. Inoltre la mandibola di Ramapi-thecus e di Australopithecus è rinforzatain modo da permettere una poderosamasticazione da ben due creste. Le cre-ste, note rispettivamente come il toro su-periore e il toro inferiore, erano presentianche nelle specie più robuste di Austra-lopithecus recente, risalenti a un milionedi anni fa.
Un'altra caratteristica insolita diffusa
FAMIGLIA PONGIDI(TUTTE LE GRANDISCIMMIE ANTROPOMORFE)
FAMIGLIA OMINIDI(TUTTE LE FORMEUMANE E PREUMANE)
La superfamiglia degli ominoidei, una grande suddivisione dell'ordine dei primati, comprende trefamiglie due delle quali, pongidi e ominidi, sono rappresentate nel grafico. I pongidi comprendo-no oltre ai tre generi di grandi scimmie antropomorfe viventi, tre generi fossili qui elencati.La famiglia ominidi include il genere vivente Homo e due generi estinti di forme preumane.
a
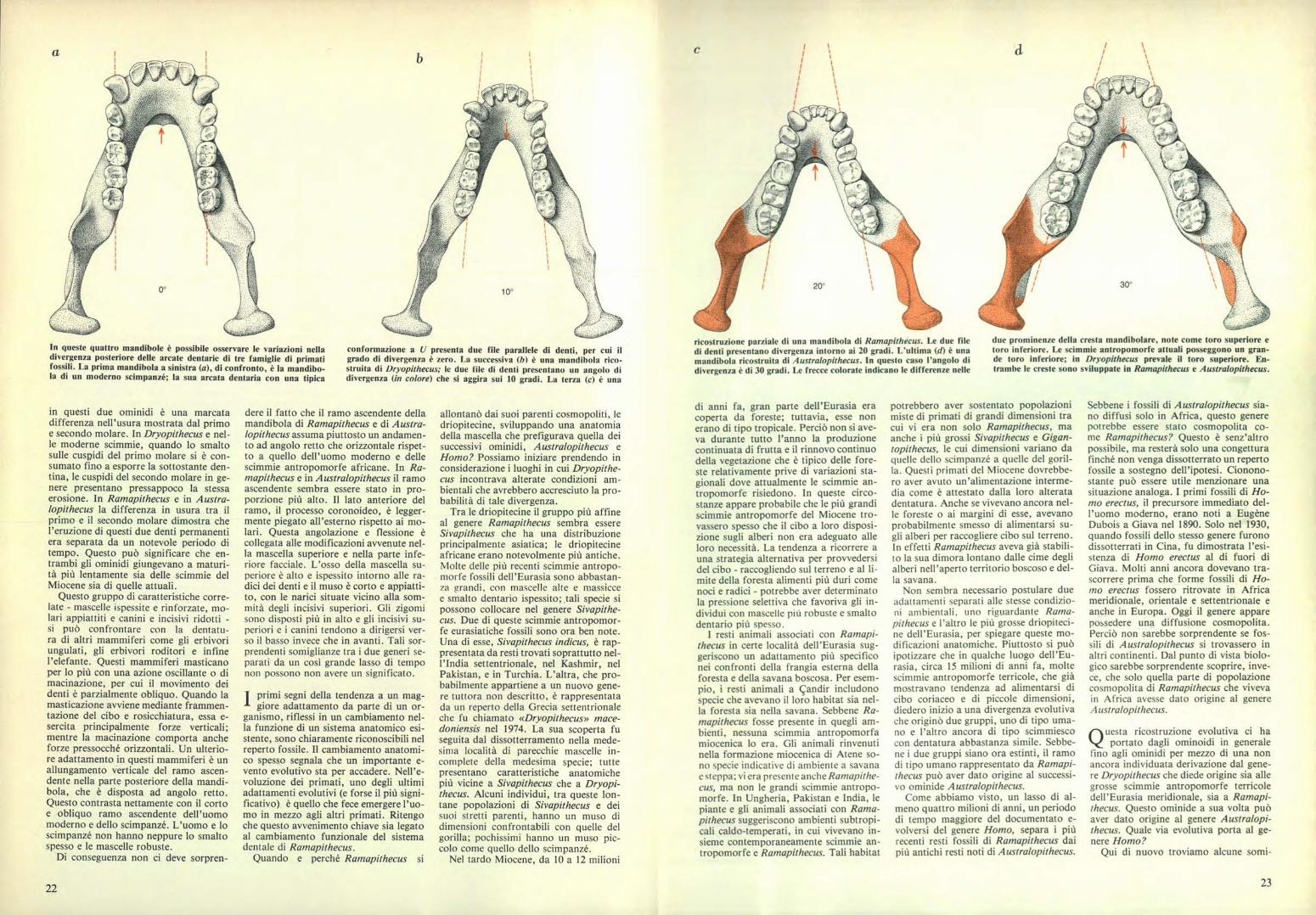
In queste quattro mandibole è possibile osservare le variazioni nelladivergenza posteriore delle arcate dentarie di tre famiglie di primatifossili. La prima mandibola a sinistra (a), di confronto, è la mandibo-la di un moderno scimpanzé; la sua arcata dentaria con una tipica
conformazione a U presenta due file parallele di denti, per cui ilgrado di divergenza è zero. La successiva (b) è una mandibola rico-struita di Dryopithecus; le due file di denti presentano un angolo didivergenza (in colore) che si aggira sui 10 gradi. La terza (c) è una
ricostruzione parziale di una mandibola di Ramapithecus. Le due filedi denti presentano divergenza intorno ai 20 gradi. L'ultima (d) è unamandibola ricostruita di Australopithecus. In questo caso l'angolo didivergenza è di 30 gradi. Le frecce colorate indicano le differenze nelle
ci
due prominenze della cresta mandibolare, note come toro superiore etoro inferiore. Le scimmie antropomorfe attuali posseggono un gran-de toro inferiore; in Dryopithecus prevale il toro superiore. En-trambe le creste sono sviluppate in Ramapithecus e Australopithecus.
in questi due ominidi è una marcatadifferenza nell'usura mostrata dal primoe secondo molare. In Dryopithecus e nel-le moderne scimmie, quando lo smaltosulle cuspidi del primo molare si è con-sumato fino a esporre la sottostante den-tina, le cuspidi del secondo molare in ge-nere presentano pressappoco la stessaerosione. In Ramapithecus e in Austra-lopithecus la differenza in usura tra ilprimo e il secondo molare dimostra chel'eruzione di questi due denti permanentiera separata da un notevole periodo ditempo. Questo può significare che en-trambi gli ominidi giungevano a maturi-tà più lentamente sia delle scimmie delMiocene sia di quelle attuali.
Questo gruppo di caratteristiche corre-late - mascelle ispessite e rinforzate, mo-lari appiattiti e canini e incisivi ridotti -si può confrontare con la dentatu-ra di altri mammiferi come gli erbivoriungulati, gli erbivori roditori e infinel'elefante. Questi mammiferi masticanoper lo più con una azione oscillante o dimacinazione, per cui il movimento deidenti è parzialmente obliquo. Quando lamasticazione avviene mediante frammen-tazione del cibo e rosicchiatura, essa e-sercita principalmente forze verticali;mentre la macinazione comporta ancheforze pressocché orizzontali. Un ulterio-re adattamento in questi mammiferi è unallungamento verticale del ramo ascen-dente nella parte posteriore della mandi-bola, che è disposta ad angolo retto.Questo contrasta nettamente con il cortoe obliquo ramo ascendente dell'uomomoderno e dello scimpanzé. L'uomo e loscimpanzé non hanno neppure lo smaltospesso e le mascelle robuste.
Di conseguenza non ci deve sorpren-
dere il fatto che il ramo ascendente dellamandibola di Ramapithecus e di Austra-lopithecus assuma piuttosto un andamen-to ad angolo retto che orizzontale rispet-to a quello dell'uomo moderno e dellescimmie antropomorfe africane. In Ra-mapithecus e in Australopithecus il ramoascendente sembra essere stato in pro-porzione più alto. Il lato anteriore delramo, il processo coronoideo, è legger-mente piegato all'esterno rispetto ai mo-lari. Questa angolazione e flessione ècollegata alle modificazioni avvenute nel-la mascella superiore e nella parte infe-riore facciale. L'osso della mascella su-periore è alto e ispessito intorno alle ra-dici dei denti e il muso è corto e appiatti-to, con le narici situate vicino alla som-mità degli incisivi superiori. Gli zigomisono disposti più in alto e gli incisivi su-periori e i canini tendono a dirigersi ver-so il basso invece che in avanti. Tali sor-prendenti somiglianze tra i due generi se-parati da un così grande lasso di temponon possono non avere un significato.
primi segni della tendenza a un mag-giore adattamento da parte di un or-
ganismo, riflessi in un cambiamento nel-la funzione di un sistema anatomico esi-stente, sono chiaramente riconoscibili nelreperto fossile. Il cambiamento anatomi-co spesso segnala che un importante e-vento evolutivo sta per accadere. Nell'e-voluzione dei primati, uno degli ultimiadattamenti evolutivi (e forse il più signi-ficativo) è quello che fece emergere l'uo-mo in mezzo agli altri primati. Ritengoche questo avvenimento chiave sia legatoal cambiamento funzionale del sistemadentale di Ratnapithecus.
Quando e perché Ramapithecus si
allontanò dai suoi parenti cosmopoliti, ledriopitecine, sviluppando una anatomiadella mascella che prefigurava quella deisuccessivi ominidi, Australopithecus eHomo? Possiamo iniziare prendendo inconsiderazione i luoghi in cui Dryopithe-cus incontrava alterate condizioni am-bientali che avrebbero accresciuto la pro-babilità di tale divergenza.
Tra le driopitecine il gruppo più affineal genere Ramapithecus sembra essereSivapithecus che ha una distribuzioneprincipalmente asiatica; le driopitecineafricane erano notevolmente più antiche.Molte delle più recenti scimmie antropo-morfe fossili dell'Eurasia sono abbastan-za grandi, con mascelle alte e massiccee smalto dentario ispessito; tali specie sipossono collocare nel genere Sivapithe-cus. Due di queste scimmie antropomor-fe eurasiatiche fossili sono ora ben note.Una di esse, Sivapithecus indicus, è rap-presentata da resti trovati soprattutto nel-l'India settentrionale, nel Kashmir, nelPakistan, e in Turchia. L'altra, che pro-babilmente appartiene a un nuovo gene-re tuttora non descritto, è rappresentatada un reperto della Grecia settentrionaleche fu chiamato «Dryopithecus» mace-doniensis nel 1974. La sua scoperta fuseguita dal dissotterramento nella mede-sima località di parecchie mascelle in-complete della medesima specie; tuttepresentano caratteristiche anatomichepiù vicine a Sivapithecus che a Dryopi-thecus. Alcuni individui, tra queste lon-tane popolazioni di Sivapithecus e deisuoi stretti parenti, hanno un muso didimensioni confrontabili con quelle delgorilla; pochissimi hanno un muso pic-colo come quello dello scimpanzé.
Nel tardo Miocene, da 10 a 12 milioni
di anni fa, gran parte dell'Eurasia eracoperta da foreste; tuttavia, esse nonerano di tipo tropicale. Perciò non si ave-va durante tutto l'anno la produzionecontinuata di frutta e il rinnovo continuodella vegetazione che è tipico delle fore-ste relativamente prive di variazioni sta-gionali dove attualmente le scimmie an-tropomorfe risiedono. In queste circo-stanze appare probabile che le più grandiscimmie antropomorfe del Miocene tro-vassero spesso che il cibo a loro disposi-zione sugli alberi non era adeguato alleloro necessità. La tendenza a ricorrere auna strategia alternativa per provvedersidel cibo - raccogliendo sul terreno e al li-mite della foresta alimenti più duri comenoci e radici - potrebbe aver determinatola pressione selettiva che favoriva gli in-dividui con mascelle più robuste e smaltodentario più spesso.
I resti animali associati con Ramapi-thecus in certe località dell'Eurasia sug-geriscono un adattamento più specificonei confronti della frangia esterna dellaforesta e della savana boscosa. Per esem-pio, i resti animali a Candir includonospecie che avevano il loro habitat sia nel-la foresta sia nella savana. Sebbene Ra-mapithecus fosse presente in quegli am-bienti, nessuna scimmia antropomorfamiocenica lo era. Gli animali rinvenutinella formazione miocenica di Atene so-no specie indicative di ambiente a savanae steppa; vi era presente anche Ramapithe-cus, ma non le grandi scimmie antropo-morfe. In Ungheria, Pakistan e India, lepiante e gli animali associati con Rama-pithecus suggeriscono ambienti subtropi-cali caldo-temperati, in cui vivevano in-sieme contemporaneamente scimmie an-tropomorfe e Ramapithecus. Tali habitat
potrebbero aver sostentato popolazionimiste di primati di grandi dimensioni tracui vi era non solo Ramapithecus, maanche i più grossi Sivapithecus e Gigan-topithecus, le cui dimensioni variano daquelle dello scimpanzé a quelle del goril-la. Questi primati del Miocene dovrebbe-ro aver avuto un'alimentazione interme-dia come è attestato dalla loro alteratadentatura. Anche se vivevano ancora nel-le foreste o ai margini di esse, avevanoprobabilmente smesso di alimentarsi su-gli alberi per raccogliere cibo sul terreno.In effetti Ramapithecus aveva già stabili-to la sua dimora lontano dalle cime deglialberi nell'aperto territorio boscoso e del-la savana.
Non sembra necessario postulare dueadattamenti separati alle stesse condizio-ni ambientali, uno riguardante Rama-pithecus e l'altro le più grosse driopiteci-ne dell'Eurasia, per spiegare queste mo-dificazioni anatomiche. Piuttosto si puòipotizzare che in qualche luogo dell'Eu-rasia, circa 15 milioni di anni fa, moltescimmie antropomorfe terricole, che giàmostravano tendenza ad alimentarsi dicibo coriaceo e di piccole dimensioni,diedero inizio a una divergenza evolutivache originò due gruppi, uno di tipo uma-no e l'altro ancora di tipo scimmiescocon dentatura abbastanza simile. Sebbe-ne i due gruppi siano ora estinti, il ramodi tipo umano rappresentato da Ramapi-thecus può aver dato origine al successi-vo ominide Australopithecus.
Come abbiamo visto, un lasso di al-meno quattro milioni di anni, un periododi tempo maggiore del documentato e-volversi del genere Homo, separa i piùrecenti resti fossili di Ramapithecus daipiù antichi resti noti di Australopithecus.
Sebbene i fossili di Australopithecus sia-no diffusi solo in Africa, questo generepotrebbe essere stato cosmopolita co-me Ramapithecus? Questo è senz'altropossibile, ma resterà solo una congetturafinché non venga dissotterrato un repertofossile a sostegno dell'ipotesi. Cionono-stante può essere utile menzionare unasituazione analoga. I primi fossili di Ho-mo erectus, il precursore immediato del-l'uomo moderno, erano noti a EugèneDubois a Giava nel 1890. Solo nel 1930,quando fossili dello stesso genere furonodissotterrati in Cina, fu dimostrata l'esi-stenza di Homo erectus al di fuori diGiava. Molti anni ancora dovevano tra-scorrere prima che forme fossili di Ho-mo erectus fossero ritrovate in Africameridionale, orientale e settentrionale eanche in Europa. Oggi il genere apparepossedere una diffusione cosmopolita.Perciò non sarebbe sorprendente se fos-sili di Australopithecus si trovassero inaltri continenti. Dal punto di vista biolo-gico sarebbe sorprendente scoprire, inve-ce, che solo quella parte di popolazionecosmopolita di Ramapithecus che vivevain Africa avesse dato origine al genereAustralopithecus.
uesta ricostruzione evolutiva ci haportato dagli ominoidi in generale
fino agli ominidi per mezzo di una nonancora individuata derivazione dal gene-re Dryopithecus che diede origine sia allegrosse scimmie antropomorfe terricoledell'Eurasia meridionale, sia a Ramapi-thecus. Questo ominide a sua volta puòaver dato origine al genere Australopi-thecus. Quale via evolutiva porta al ge-nere Homo?
Qui di nuovo troviamo alcune somi-
22 23
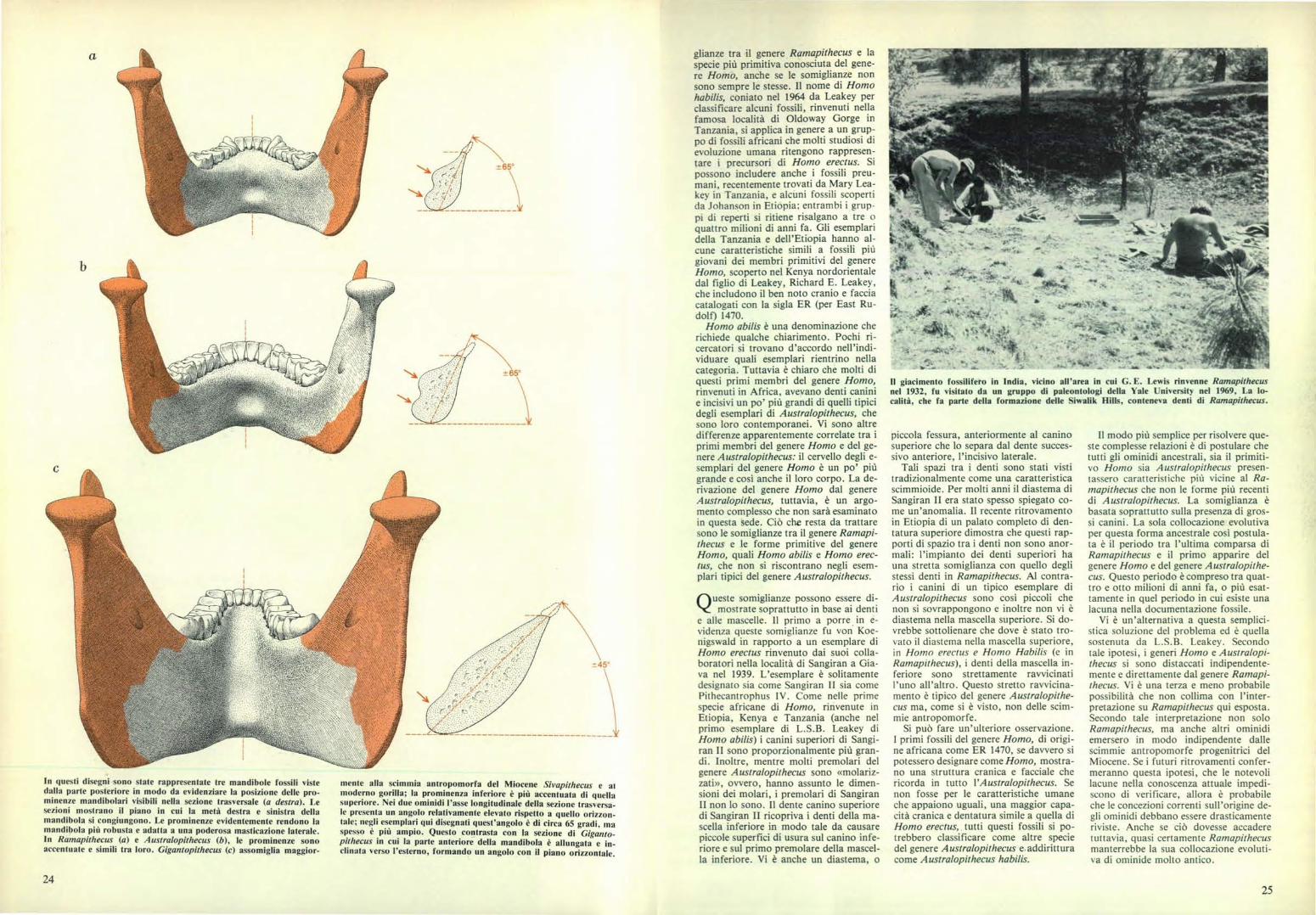
In questi disegni sono state rappresentate tre mandibole fossili vistedalla parte posteriore in modo da evidenziare la posizione delle pro-minenze mandibolari visibili nella sezione trasversale (a destra). Lesezioni mostrano il piano in cui la metà destra e sinistra dellamandibola si congiungono. Le prominenze evidentemente rendono lamandibola più robusta e adatta a una poderosa masticazione laterale.In Ramapithecus (a) e Australopithecus (b), le prominenze sonoaccentuate e simili tra loro. Gigantopithecus (c) assomiglia maggior-
-±-45°
mente alla scimmia antropomorfa del Miocene Sivapithecus e aimoderno gorilla; la prominenza inferiore è più accentuata di quellasuperiore. Nei due ominidi l'asse longitudinale della sezione trasversa-le presenta un angolo relativamente elevato rispetto a quello orizzon-tale; negli esemplari qui disegnati quest'angolo è di circa 65 gradi, maspesso è più ampio. Questo contrasta con la sezione di Giganto-pithecus in cui la parte anteriore della mandibola è allungata e in-clinata verso l'esterno, formando un angolo con il piano orizzontale.
a
-±65°
-±65°Il giacimento fossilifero in India, vicino all'area in cui G. E. Lewis rinvenne Ramapithecusnel 1932, fu visitato da un gruppo di paleontologi della Yale University nel 1%9, La lo-calità, che fa parte della formazione delle Sissalik Hills, conteneva denti di Ramapithecus.
glianze tra il genere Ramapithecus e laspecie più primitiva conosciuta del gene-re Homo, anche se le somiglianze nonsono sempre le stesse. Il nome di Homohabilis, coniato nel 1964 da Leakey perclassificare alcuni fossili, rinvenuti nellafamosa località di Oldoway Gorge inTanzania, si applica in genere a un grup-po di fossili africani che molti studiosi dievoluzione umana ritengono rappresen-tare i precursori di Homo erectus. Sipossono includere anche i fossili preu-mani, recentemente trovati da Mary Lea-key in Tanzania, e alcuni fossili scopertida Johanson in Etiopia: entrambi i grup-pi di reperti si ritiene risalgano a tre oquattro milioni di anni fa. Gli esemplaridella Tanzania e dell'Etiopia hanno al-cune caratteristiche simili a fossili piùgiovani dei membri primitivi del genereHomo, scoperto nel Kenya nordorientaledal figlio di Leakey, Richard E. Leakey,che includono il ben noto cranio e facciacatalogati con la sigla ER (per East Ru-dolf) 1470.
Homo abilis è una denominazione cherichiede qualche chiarimento. Pochi ri-cercatori si trovano d'accordo nell'indi-viduare quali esemplari rientrino nellacategoria. Tuttavia è chiaro che molti diquesti primi membri del genere Homo,rinvenuti in Africa, avevano denti caninie incisivi un po' più grandi di quelli tipicidegli esemplari di Australopithecus, chesono loro contemporanei. Vi sono altredifferenze apparentemente correlate tra iprimi membri del genere Homo e del ge-nere Australopithecus: il cervello degli e-semplari del genere Homo è un po' piùgrande e così anche il loro corpo. La de-rivazione del genere Homo dal genereAustralopithecus, tuttavia, è un argo-mento complesso che non sarà esaminatoin questa sede. Ciò che resta da trattaresono le somiglianze tra il genere Ramapi-thecus e le forme primitive del genereHomo, quali Homo abilis e Homo erec-tus, che non si riscontrano negli esem-plari tipici del genere Australopithecus.
Queste somiglianze possono essere di-mostrate soprattutto in base ai denti
e alle mascelle. il primo a porre in e-videnza queste somiglianze fu von Koe-nigswald in rapporto a un esemplare diHomo erectus rinvenuto dai suoi colla-boratori nella località di Sangiran a Gia-va nel 1939. L'esemplare è solitamentedesignato sia come Sangiran II sia comePithecantrophus IV. Come nelle primespecie africane di Homo, rinvenute inEtiopia, Kenya e Tanzania (anche nelprimo esemplare di L.S.B. Leakey diHomo abilis) i canini superiori di Sangi-ran II sono proporzionalmente più gran-di. Inoltre, mentre molti premolari delgenere Australopithecus sono «molariz-zati», ovvero, hanno assunto le dimen-sioni dei molari, i premolari di SangiranII non lo sono. Il dente canino superioredi Sangiran II ricopriva i denti della ma-scella inferiore in modo tale da causarepiccole superfici di usura sul canino infe-riore e sul primo premolare della mascel-la inferiore. Vi è anche un diastema, o
piccola fessura, anteriormente al caninosuperiore che lo separa dal dente succes-sivo anteriore, l'incisivo laterale.
Tali spazi tra i denti sono stati vistitradizionalmente come una caratteristicascimmioide. Per molti anni il diastema diSangiran II era stato spesso spiegato co-me un'anomalia. Il recente ritrovamentoin Etiopia di un palato completo di den-tatura superiore dimostra che questi rap-porti di spazio tra i denti non sono anor-mali: l'impianto dei denti superiori hauna stretta somiglianza con quello deglistessi denti in Ramapithecus. Al contra-rio i canini di un tipico esemplare diAustralopithecus sono così piccoli chenon si sovrappongono e inoltre non vi èdiastema nella mascella superiore. Si do-vrebbe sottolienare che dove è stato tro-vato il diastema nella mascella superiore,in Homo erectus e Homo Habilis (e inRamapithecus), i denti della mascella in-feriore sono strettamente ravvicinatil'uno all'altro. Questo stretto ravvicina-mento è tipico del genere Australopithe-cus ma, come si è visto, non delle scim-mie antropomorfe.
Si può fare un'ulteriore osservazione.I primi fossili del genere Homo, di origi-ne africana come ER 1470, se davvero sipotessero designare come Homo, mostra-no una struttura cranica e facciale chericorda in tutto l'Australopithecus. Senon fosse per le caratteristiche umaneche appaiono uguali, una maggior capa-cità cranica e dentatura simile a quella diHomo erectus, tutti questi fossili si po-trebbero classificare come altre speciedel genere Australopithecus e addiritturacome Australopithecus habilis.
Il modo più semplice per risolvere que-ste complesse relazioni è di postulare chetutti gli ominidi ancestrali, sia il primiti-vo Homo sia Australopithecus presen-tassero caratteristiche più vicine al Ra-mapithecus che non le forme più recentidi Australopithecus. La somiglianza èbasata soprattutto sulla presenza di gros-si canini. La sola collocazione evolutivaper questa forma ancestrale così postula-ta è il periodo tra l'ultima comparsa diRamapithecus e il primo apparire delgenere Homo e del genere Australopithe-cus. Questo periodo è compreso tra quat-tro e otto milioni di anni fa, o più esat-tamente in quel periodo in cui esiste unalacuna nella documentazione fossile.
Vi è un'alternativa a questa semplici-stica soluzione del problema ed è quellasostenuta da L.S.B. Leakey. Secondotale ipotesi, i generi Homo e Australopi-thecus si sono distaccati indipendente-mente e direttamente dal genere Ramapi-thecus. Vi è una terza e meno probabilepossibilità che non collima con l'inter-pretazione su Ramapithecus qui esposta.Secondo tale interpretazione non soloRamapithecus, ma anche altri ominidiemersero in modo indipendente dallescimmie antropomorfe progenitrici delMiocene. Se i futuri ritrovamenti confer-meranno questa ipotesi, che le notevolilacune nella conoscenza attuale impedi-scono di verificare, allora è probabileche le concezioni correnti sull'origine de-gli ominidi debbano essere drasticamenteriviste. Anche se ciò dovesse accaderetuttavia, quasi certamente Ramapithecusmanterrebbe la sua collocazione evoluti-va di ominide molto antico.
2425