
La funzione immunologica della pelle - Kataweb
5
Il coinvolgimento della pelle umana nel funzionamento del sistema immunitario è suggerito dalla correlazione strutturale che la pelle ha con il timo, organo del sistema immunitario in cui maturano i linfo- citi T. Le microfotografie di Tung Tien-Sun del Medical Center del- la New York University mostrano una sorprendente somiglianza tra i cheratinociti, principali cellule dell'epidermide (a sinistra), e le ceI- lule epiteliali del timo (a destra). Tutte queste cellule sono state incubate con un anticorpo contro la cheratina, proteina dei cherati- nociti; l'anticorpo legato alla cheratina è stato localizzato con un secondo anticorpo marcato con fluoresceina, un colorante fluore- scente giallo-verde. Sia nelle cellule del timo sia in quelle dell'epi- derrnide è riconoscibile un intreccio ondulato di fibrille di cheratina. di Richard L. Edelson e Joseph M. Fink lattia in organi che non siano la pelle. D'altra parte. il progredire delle lesioni non epidermotrope è correlato con la dif- fusione di cellule neoplastiche negli or- gani interni e con una prognosi decisa- mente più infausta. In altre parole, quelli che erano consi- derati tipi diversi di neoplasie linfociti- che T della pelle sono, invece, manife- stazioni cliniche di momenti diversi del- l'evoluzione di un singolo tipo di neopla- sia. Poiché la caratteristica biologica più marcata di questi linfociti T neoplastici è l'affinità iniziale per l'epidermide, ab- biamo chiamato la neoplasia «linfoma cutaneo di tipo T». In seguito al suo riconoscimento come singola entità pa- tologica. si è dimostrato che essa ha un'incidenza cumulativa superiore a quella del linfoma di Hodgkin ed è quin- di il linfoma più comune tra gli adulti. Un secondo denominatore comune tra i pazienti affetti da linfoma cutaneo di tipo T è la comparsa, come primo sintomo clinico, di una eruzione cutanea anche se la cellula neoplastica è qui un tipo di globulo bianco del sangue e non una cellula originaria della pelle. Questa scoperta è stata confermata da Henry Rappaport della Pritzer School of Medi- cine dell'Università di Chicago e da Louis B. Thomas del National Cancer Institute, i quali hanno esaminato cam- pioni autoptici prelevati da 45 soggetti affetti da linfoma cutaneo di tipo T. Essi hanno trovato numerosi casi in cui l'uni- ca manifestazione patologica era l'affe- zione cutanea, senza altre lesioni evi- denti a organi interni. Da ultimo, utilizzando un sistema spe- rimentale messo a punto da Samuel Broder e Thomas A. Waldman del Na- tional Cancer Institute, abbiamo stabili- to che il linfoma cutaneo di tipo T è di norma una neoplasia di una specifica classe di linfociti T: i linfociti T coadiu- vanti. Linfociti B normali, coltivati e purificati, anche in presenza di fitoe- moagglutinine stimolanti non produco- no anticorpi. L'aggiunta di linfociti T, sia normali sia provenienti da pazienti affet- ti da linfoma cutaneo di tipo T, ha indot- to i linfociti B a sintetizzare grandi quan- tità di anticorpi. Chiaramente, i linfociti T neoplastici mantengono spesso la tipi- ca capacità di stimolazione dei linfociti T coadiuvanti normali. na storia straordinaria si stava così delineando. Il linfoma cutaneo di tipo T era stato identificato come neo- plasia dei linfociti T coadiuvanti, che presentavano una spiccata preferenza La funzione immunologica della pelle Il più esteso organo corporeo non è solo un rivestimento passivo: è un elemento attivo del sistema immunitario con cellule specializzate che, interagendo l'una con l'altra, reagiscono all'invasione di sostanze estranee L a raffinata semplicità della pelle umana maschera la sua elevata complessità e la molteplicità delle sue funzioni. La pelle è spessa solo alcuni millimetri, ma è l'organo più esteso del nostro corpo e contiene una varietà di cellule altamente specializza- te, organizzate in intricate strutture e sottosistemi. Una delle sue funzioni più notevoli è stata riconosciuta solo di re- cente: la pelle è un elemento essenziale e attivo del sistema immunitario. In retrospettiva possiamo affermare che la scoperta di questo ruolo attivo nel sistema immunitario non sarebbe dovuta essere una sorpresa: la pelle è, infatti, l'interfaccia dell'organismo con l'ambien- te esterno e quella umana è particolar- mente vulnerabile perché è coperta solo in minima parte da peli. È quindi logico supporre che la nostra specie non sarebbe mai potuta sopravvivere alle infezioni de- rivanti dalle numerose ferite superficiali della pelle se questo rivestimento non fos- se stato in grado di mobilitare una potente e sofisticata reazione immunitaria. Spesso funzioni normali poco appari- scenti, ma importanti, vengono messe in luce dallo studio di disfunzioni patologi- che. Il fatto che la pelle svolga un ruolo non soltanto passivo in alcune malattie che la colpiscono è stato svelato per la prima volta da studi sulle neoplasie dei linfociti, i globuli bianchi del sangue che controllano il funzionamento del siste- ma immunitario. Già nel 1970 si sapeva, da studi effet- tuati sui topi, che i linfociti possono es- sere suddivisi in due classi principali, funzionalmente distinte: i linfociti B (che maturano principalmente nel mi- dollo osseo) e i linfociti T (che matura- no nel timo). I primi sono interessati nell'immunità umorale, cioè sintetizza- no gli anticorpi, che reagiscono con an- tigeni specifici, quali sono le molecole presenti sulla superficie dei microrgani- smi patogeni o delle cellule neoplasti- che. I secondi, invece, sono responsabili dell'immunità cellulare. Per esempio, distruggono le cellule infettate da virus, avviano la risposta cellulare alle infe- zioni batteriche e reagiscono contro i trapianti incompatibili di tessuti. Alcuni sottogruppi di linfociti T modulano la risposta immunitaria: i linfociti T «coa- diuvanti» (helper) contribuiscono alla maturazione dei linfociti B in altri linfo- citi che secernono anticorpi e provoca- no l'incremento di specifiche popola- zioni di linfociti T; i linfociti T «sop- pressori», invece, riducono l'attività dei linfociti B e limitano le dimensioni delle popolazioni di linfociti T. Nel 1972, per la prima volta, è stato possibile classificare anche i linfociti umani in B e T. Un certo numero di ricercatori si è impegnato a riclassifica- re le neoplasie linfocitiche come proli- ferazioni specifiche dell'uno o dell'altro tipo di linfociti. Queste neoplasie inclu- dono le leucemie, in cui grandi quantità di linfociti tumorali circolano nel san- gue, e i linfomi, in cui le cellule neopla- stiche si accumulano nei linfonodi e negli organi linfatici. Il risultato di que- sta riclassificazione è stato sorprenden- te. Sebbene nel sangue i linfociti T sia- no normalmente almeno tre volte più numerosi dei linfociti B, per la maggior parte le leucemie e i linfomi degli adulti sono risultati neoplasie di quest'ultimo tipo di linfociti. Ira Green, Philip S. Schein, Charles H. Kirkpatrick, Ethan M. Shevach, Marvin A. Lutzner e uno di noi (Edel- son) hanno tuttavia scoperto, lavorando ai National Institutes of Health, che un vasto gruppo di neoplasie, comprenden- te certe leucemie e certi linfomi in cui la pelle subisce significative infiltrazioni di cellule tumorali, poteva essere attribuito regolarmente ai linfociti T. La dicotomia era sorprendente: praticamente l'origi- ne di tutte le neoplasie linfocitiche in cui si riscontra un'estesa infiltrazione della pelle da parte di cellule tumorali è stata dimostrata nei linfociti T; quella della maggior parte delle neoplasie in cui la pelle non è coinvolta è stata dimostrata nei linfociti B. In altre parole, i linfociti T neoplastici sembravano avere una par- ticolare affinità per la pelle. uesta scoperta sottintendeva che tutte le neoplasie dei linfociti T che avevano affinità per la pelle potessero essere forme diverse di un'unica malat- tia. Era probabile, inoltre, che l'affinità per la pelle delle cellule T neoplastiche riflettesse un'analoga proprietà delle cellule T normali, da cui le altre erano derivate: una popolazione significativa di cellule T doveva risiedere abitual- mente nella pelle. Isolati linfociti «di passaggio» erano stati spesso osservati nella pelle umana normale, ma la loro presenza non era stata mai considerata significativa. Poteva darsi, quindi, che il fenomeno da noi osservato fosse un'am- plificazione maligna di un'interazione, normalmente non evidente, tra pelle e alcuni linfociti T in essa residenti. Trovammo altri tre denominatori comuni tra i pazienti con le varie manife- stazioni cliniche delle neoplasie linfociti- che T, riguardanti la pelle. In primo luo- go, in ciascun paziente, la malattia segui- va un'evoluzione naturale. Le primissime lesioni osservate tendono a essere di tipo «epidermotropo», cioè caratterizzate dalla migrazione di cellule neoplastiche nell'epidermide, lo strato più esterno del- la pelle. Queste lesioni precoci sono pro- gressivamente sostituite da lesioni «non epidermotrope», in cui l'epidermide è risparmiata, mentre le cellule neoplasti- che si accumulano in posizioni della pelle più profonde e nei tessuti sottocutanei. Se sono presenti solo lesioni epidermotrope, spesso non si manifestano sintomi di ma- 48 (-3 49
Transcript of La funzione immunologica della pelle - Kataweb
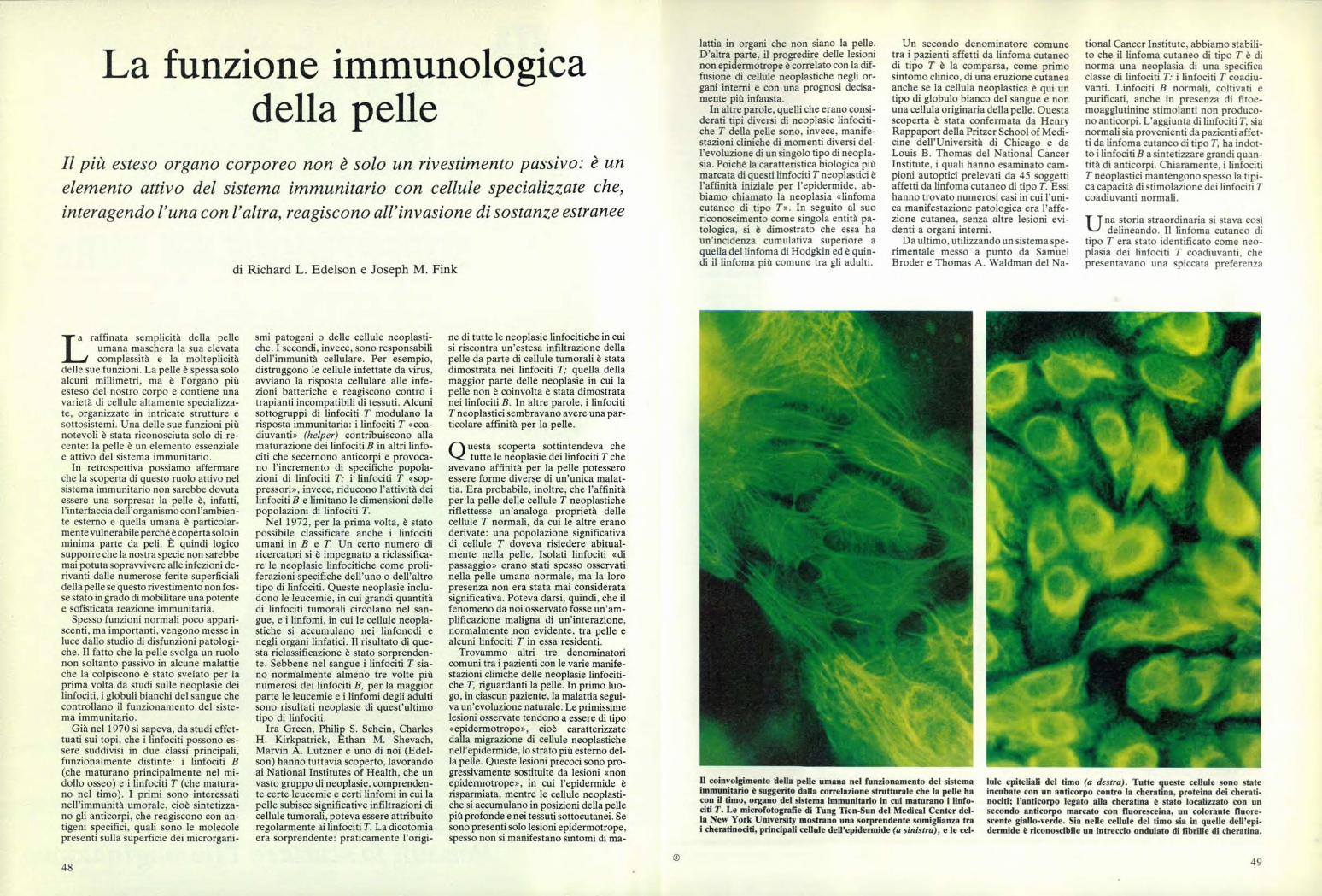
Il coinvolgimento della pelle umana nel funzionamento del sistemaimmunitario è suggerito dalla correlazione strutturale che la pelle hacon il timo, organo del sistema immunitario in cui maturano i linfo-citi T. Le microfotografie di Tung Tien-Sun del Medical Center del-la New York University mostrano una sorprendente somiglianza trai cheratinociti, principali cellule dell'epidermide (a sinistra), e le ceI-
lule epiteliali del timo (a destra). Tutte queste cellule sono stateincubate con un anticorpo contro la cheratina, proteina dei cherati-nociti; l'anticorpo legato alla cheratina è stato localizzato con unsecondo anticorpo marcato con fluoresceina, un colorante fluore-scente giallo-verde. Sia nelle cellule del timo sia in quelle dell'epi-derrnide è riconoscibile un intreccio ondulato di fibrille di cheratina.
di Richard L. Edelson e Joseph M. Fink
lattia in organi che non siano la pelle.D'altra parte. il progredire delle lesioninon epidermotrope è correlato con la dif-fusione di cellule neoplastiche negli or-gani interni e con una prognosi decisa-mente più infausta.
In altre parole, quelli che erano consi-derati tipi diversi di neoplasie linfociti-che T della pelle sono, invece, manife-stazioni cliniche di momenti diversi del-l'evoluzione di un singolo tipo di neopla-sia. Poiché la caratteristica biologica piùmarcata di questi linfociti T neoplastici èl'affinità iniziale per l'epidermide, ab-biamo chiamato la neoplasia «linfomacutaneo di tipo T». In seguito al suoriconoscimento come singola entità pa-tologica. si è dimostrato che essa haun'incidenza cumulativa superiore aquella del linfoma di Hodgkin ed è quin-di il linfoma più comune tra gli adulti.
Un secondo denominatore comunetra i pazienti affetti da linfoma cutaneodi tipo T è la comparsa, come primosintomo clinico, di una eruzione cutaneaanche se la cellula neoplastica è qui untipo di globulo bianco del sangue e nonuna cellula originaria della pelle. Questascoperta è stata confermata da HenryRappaport della Pritzer School of Medi-cine dell'Università di Chicago e daLouis B. Thomas del National CancerInstitute, i quali hanno esaminato cam-pioni autoptici prelevati da 45 soggettiaffetti da linfoma cutaneo di tipo T. Essihanno trovato numerosi casi in cui l'uni-ca manifestazione patologica era l'affe-zione cutanea, senza altre lesioni evi-denti a organi interni.
Da ultimo, utilizzando un sistema spe-rimentale messo a punto da SamuelBroder e Thomas A. Waldman del Na-
tional Cancer Institute, abbiamo stabili-to che il linfoma cutaneo di tipo T è dinorma una neoplasia di una specificaclasse di linfociti T: i linfociti T coadiu-vanti. Linfociti B normali, coltivati epurificati, anche in presenza di fitoe-moagglutinine stimolanti non produco-no anticorpi. L'aggiunta di linfociti T, sianormali sia provenienti da pazienti affet-ti da linfoma cutaneo di tipo T, ha indot-to i linfociti B a sintetizzare grandi quan-tità di anticorpi. Chiaramente, i linfocitiT neoplastici mantengono spesso la tipi-ca capacità di stimolazione dei linfociti Tcoadiuvanti normali.
na storia straordinaria si stava cosìdelineando. Il linfoma cutaneo di
tipo T era stato identificato come neo-plasia dei linfociti T coadiuvanti, chepresentavano una spiccata preferenza
La funzione immunologicadella pelle
Il più esteso organo corporeo non è solo un rivestimento passivo: è unelemento attivo del sistema immunitario con cellule specializzate che,interagendo l'una con l'altra, reagiscono all'invasione di sostanze estranee
L
a raffinata semplicità della pelleumana maschera la sua elevatacomplessità e la molteplicità
delle sue funzioni. La pelle è spessa soloalcuni millimetri, ma è l'organo piùesteso del nostro corpo e contiene unavarietà di cellule altamente specializza-te, organizzate in intricate strutture esottosistemi. Una delle sue funzioni piùnotevoli è stata riconosciuta solo di re-cente: la pelle è un elemento essenzialee attivo del sistema immunitario.
In retrospettiva possiamo affermareche la scoperta di questo ruolo attivo nelsistema immunitario non sarebbe dovutaessere una sorpresa: la pelle è, infatti,l'interfaccia dell'organismo con l'ambien-te esterno e quella umana è particolar-mente vulnerabile perché è coperta solo inminima parte da peli. È quindi logicosupporre che la nostra specie non sarebbemai potuta sopravvivere alle infezioni de-rivanti dalle numerose ferite superficialidella pelle se questo rivestimento non fos-se stato in grado di mobilitare una potentee sofisticata reazione immunitaria.
Spesso funzioni normali poco appari-scenti, ma importanti, vengono messe inluce dallo studio di disfunzioni patologi-che. Il fatto che la pelle svolga un ruolonon soltanto passivo in alcune malattieche la colpiscono è stato svelato per laprima volta da studi sulle neoplasie deilinfociti, i globuli bianchi del sangue checontrollano il funzionamento del siste-ma immunitario.
Già nel 1970 si sapeva, da studi effet-tuati sui topi, che i linfociti possono es-sere suddivisi in due classi principali,funzionalmente distinte: i linfociti B(che maturano principalmente nel mi-dollo osseo) e i linfociti T (che matura-no nel timo). I primi sono interessatinell'immunità umorale, cioè sintetizza-no gli anticorpi, che reagiscono con an-tigeni specifici, quali sono le molecolepresenti sulla superficie dei microrgani-
smi patogeni o delle cellule neoplasti-che. I secondi, invece, sono responsabilidell'immunità cellulare. Per esempio,distruggono le cellule infettate da virus,avviano la risposta cellulare alle infe-zioni batteriche e reagiscono contro itrapianti incompatibili di tessuti. Alcunisottogruppi di linfociti T modulano larisposta immunitaria: i linfociti T «coa-diuvanti» (helper) contribuiscono allamaturazione dei linfociti B in altri linfo-citi che secernono anticorpi e provoca-no l'incremento di specifiche popola-zioni di linfociti T; i linfociti T «sop-pressori», invece, riducono l'attività deilinfociti B e limitano le dimensioni dellepopolazioni di linfociti T.
Nel 1972, per la prima volta, è statopossibile classificare anche i linfocitiumani in B e T. Un certo numero diricercatori si è impegnato a riclassifica-re le neoplasie linfocitiche come proli-ferazioni specifiche dell'uno o dell'altrotipo di linfociti. Queste neoplasie inclu-dono le leucemie, in cui grandi quantitàdi linfociti tumorali circolano nel san-gue, e i linfomi, in cui le cellule neopla-stiche si accumulano nei linfonodi enegli organi linfatici. Il risultato di que-sta riclassificazione è stato sorprenden-te. Sebbene nel sangue i linfociti T sia-no normalmente almeno tre volte piùnumerosi dei linfociti B, per la maggiorparte le leucemie e i linfomi degli adultisono risultati neoplasie di quest'ultimotipo di linfociti.
Ira Green, Philip S. Schein, CharlesH. Kirkpatrick, Ethan M. Shevach,Marvin A. Lutzner e uno di noi (Edel-son) hanno tuttavia scoperto, lavorandoai National Institutes of Health, che unvasto gruppo di neoplasie, comprenden-te certe leucemie e certi linfomi in cui lapelle subisce significative infiltrazioni dicellule tumorali, poteva essere attribuitoregolarmente ai linfociti T. La dicotomiaera sorprendente: praticamente l'origi-
ne di tutte le neoplasie linfocitiche in cuisi riscontra un'estesa infiltrazione dellapelle da parte di cellule tumorali è statadimostrata nei linfociti T; quella dellamaggior parte delle neoplasie in cui lapelle non è coinvolta è stata dimostratanei linfociti B. In altre parole, i linfocitiT neoplastici sembravano avere una par-ticolare affinità per la pelle.
uesta scoperta sottintendeva chetutte le neoplasie dei linfociti T che
avevano affinità per la pelle potesseroessere forme diverse di un'unica malat-tia. Era probabile, inoltre, che l'affinitàper la pelle delle cellule T neoplasticheriflettesse un'analoga proprietà dellecellule T normali, da cui le altre eranoderivate: una popolazione significativadi cellule T doveva risiedere abitual-mente nella pelle. Isolati linfociti «dipassaggio» erano stati spesso osservatinella pelle umana normale, ma la loropresenza non era stata mai consideratasignificativa. Poteva darsi, quindi, che ilfenomeno da noi osservato fosse un'am-plificazione maligna di un'interazione,normalmente non evidente, tra pelle ealcuni linfociti T in essa residenti.
Trovammo altri tre denominatoricomuni tra i pazienti con le varie manife-stazioni cliniche delle neoplasie linfociti-che T, riguardanti la pelle. In primo luo-go, in ciascun paziente, la malattia segui-va un'evoluzione naturale. Le primissimelesioni osservate tendono a essere di tipo«epidermotropo», cioè caratterizzatedalla migrazione di cellule neoplastichenell'epidermide, lo strato più esterno del-la pelle. Queste lesioni precoci sono pro-gressivamente sostituite da lesioni «nonepidermotrope», in cui l'epidermide èrisparmiata, mentre le cellule neoplasti-che si accumulano in posizioni della pellepiù profonde e nei tessuti sottocutanei. Sesono presenti solo lesioni epidermotrope,spesso non si manifestano sintomi di ma-
48 (-3 49
TIMO
LINFOCITO T LINFOCITO B
SOPPRESSIONE >-
AZIONE COADIUVANTE
FASE EPIDERMOTROPA:INVASIONE SUPERFICIALE
DELL'EPIDERMIDE
ANTICORPO
MUSCOLOLISCIOGHIANDOLA
ECCRINA
STRATO DICELLULE VIVE
STRATOCORNEO
,CHERATINOCITO M E LANP9i T.Q.
CELLULA DI LANGERHA- -
•
LINFOCITO
GHIANDOLASEBACEA
cr <L.0
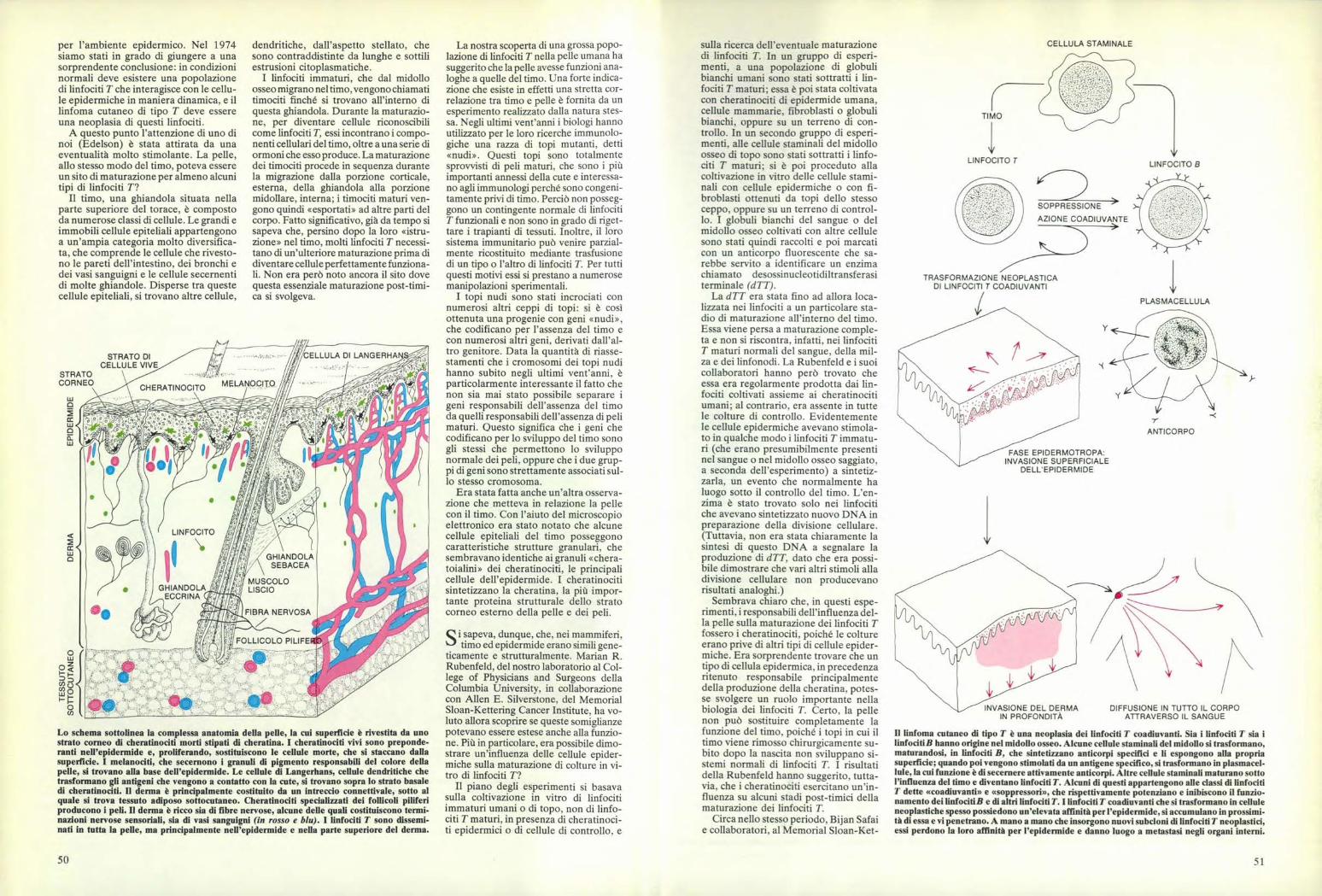
Lo schema sottolinea la complessa anatomia della pelle, la cui superficie è rivestita da unostrato corneo di cheratinociti morti stipati di cheratina. I cheratinociti vivi sono preponde-ranti nell'epidermide e, proliferando, sostituiscono le cellule morte, che si staccano dallasuperficie. I melanociti, che secernono i granuli di pigmento responsabili del colore dellapelle, si trovano alla base dell'epidermide. Le cellule di Langerhans, cellule dendritiche chetrasformano gli antigeni che vengono a contatto con la cute, si trovano sopra lo strato basaledi cheratinociti. Il derma è principalmente costituito da un intreccio connettivale, sotto alquale si trova tessuto adiposo sottocutaneo. Cheratinociti specializzati dei follicoli pifferiproducono i peli. Il derma è ricco sia di fibre nervose, alcune delle quali costituiscono termi-nazioni nervose sensoriali, sia di vasi sanguigni (in rosso e blu). I linfociti T sono dissemi-nati in tutta la pelle, ma principalmente nell'epidermide e nella parte superiore del derma.
per l'ambiente epidermico. Nel 1974siamo stati in grado di giungere a unasorprendente conclusione: in condizioninormali deve esistere una popolazionedi linfociti T che interagisce con le cellu-le epidermiche in maniera dinamica, e illinfoma cutaneo di tipo T deve essereuna neoplasia di questi linfociti.
A questo punto l'attenzione di uno dinoi (Edelson) è stata attirata da unaeventualità molto stimolante. La pelle,allo stesso modo del timo, poteva essereun sito di maturazione per almeno alcunitipi di linfociti T?
Il timo, una ghiandola situata nellaparte superiore del torace, è compostoda numerose classi di cellule. Le grandi eimmobili cellule epiteliali appartengonoa un'ampia categoria molto diversifica-ta, che comprende le cellule che rivesto-no le pareti dell'intestino, dei bronchi edei vasi sanguigni e le cellule secernentidi molte ghiandole. Disperse tra questecellule epiteliali, si trovano altre cellule,
dendritiche, dall'aspetto stellato, chesono contraddistinte da lunghe e sottiliestrusioni citoplasmatiche.
I linfociti immaturi, che dal midolloosseo migrano nel timo, vengono chiamatitimociti finché si trovano all'interno diquesta ghiandola. Durante la maturazio-ne, per diventare cellule riconoscibilicome linfociti T, essi incontrano i compo-nenti cellulari del timo, oltre a una serie diormoni che esso produce. La maturazionedei timociti procede in sequenza durantela migrazione dalla porzione corticale,esterna, della ghiandola alla porzionemidollare, interna; i timociti maturi ven-gono quindi «esportati» ad altre parti delcorpo. Fatto significativo, già da tempo sisapeva che, persino dopo la loro «istru-zione» nel timo, molti linfociti T necessi-tano di un'ulteriore maturazione prima didiventare cellule perfettamente funziona-li. Non era però noto ancora il sito dovequesta essenziale maturazione post-timi-ca si svolgeva.
La nostra scoperta di una grossa popo-lazione di linfociti T nella pelle umana hasuggerito che la pelle avesse funzioni ana-loghe a quelle del timo. Una forte indica-zione che esiste in effetti una stretta cor-relazione tra timo e pelle è fornita da unesperimento realizzato dalla natura stes-sa. Negli ultimi vent'anni i biologi hannoutilizzato per le loro ricerche immunolo-giche una razza di topi mutanti, detti«nudi». Questi topi sono totalmentesprovvisti di peli maturi, che sono i piùimportanti annessi della cute e interessa-no agli immunologi perché sono congeni-tamente privi di timo. Perciò non posseg-gono un contingente normale di linfocitiT funzionali e non sono in grado di riget-tare i trapianti di tessuti. Inoltre, il lorosistema immunitario può venire parzial-mente ricostituito mediante trasfusionedi un tipo o l'altro di linfociti T. Per tuttiquesti motivi essi si prestano a numerosemanipolazioni sperimentali.
I topi nudi sono stati incrociati connumerosi altri ceppi di topi: si è cosìottenuta una progenie con geni «nudi»,che codificano per l'assenza del timo econ numerosi altri geni, derivati dall'al-tro genitore. Data la quantità di riasse-stamenti che i cromosomi dei topi nudihanno subito negli ultimi vent'anni, èparticolarmente interessante il fatto chenon sia mai stato possibile separare igeni responsabili dell'assenza del timoda quelli responsabili dell'assenza di pelimaturi. Questo significa che i geni checodificano per lo sviluppo del timo sonogli stessi che permettono lo svilupponormale dei peli, oppure che i due grup-pi di geni sono strettamente associati sul-lo stesso cromosoma.
Era stata fatta anche un'altra osserva-zione che metteva in relazione la pellecon il timo. Con l'aiuto del microscopioelettronico era stato notato che alcunecellule epiteliali del timo posseggonocaratteristiche strutture granulari, chesembravano identiche ai granuli «chera-toialini» dei cheratinociti, le principalicellule dell'epidermide. I cheratinocitisintetizzano la cheratina, la più impor-tante proteina strutturale dello stratocorneo esterno della pelle e dei peli.
Q i sapeva, dunque, che, nei mammiferi,timo ed epidermide erano simili gene-
ticamente e strutturalmente. Marian R.Rubenfeld, del nostro laboratorio al Col-lege of Physicians and Surgeons dellaColumbia University, in collaborazionecon Allen E. Silverstone, del MemorialSloan-Kettering Cancer Institute, ha vo-luto allora scoprire se queste somiglianzepotevano essere estese anche alla funzio-ne. Più in particolare, era possibile dimo-strare un'influenza delle cellule epider-miche sulla maturazione di colture in vi-tro di linfociti T?
Il piano degli esperimenti si basavasulla coltivazione in vitro di linfocitiimmaturi umani o di topo, non di linfo-citi T maturi, in presenza di cheratinoci-ti epidermici o di cellule di controllo, e
sulla ricerca dell'eventuale maturazionedi linfociti T. In un gruppo di esperi-menti, a una popolazione di globulibianchi umani sono stati sottratti i lin-fociti T maturi; essa è poi stata coltivatacon cheratinociti di epidermide umana,cellule mammarie, fibroblasti o globulibianchi, oppure su un terreno di con-trollo. In un secondo gruppo di esperi-menti, alle cellule staminali del midolloosseo di topo sono stati sottratti i linfo-citi T maturi; si è poi proceduto allacoltivazione in vitro delle cellule stami-nali con cellule epidermiche o con fi-broblasti ottenuti da topi dello stessoceppo, oppure su un terreno di control-lo. I globuli bianchi del sangue o delmidollo osseo coltivati con altre cellulesono stati quindi raccolti e poi marcaticon un anticorpo fluorescente che sa-rebbe servito a identificare un enzimachiamato desossinucleotidiltransferasiterminale (dTT).
La dTT era stata fino ad allora loca-lizzata nei linfociti a un particolare sta-dio di maturazione all'interno del timo.Essa viene persa a maturazione comple-ta e non si riscontra, infatti, nei linfocitiT maturi normali del sangue, della mil-za e dei linfonodi. La Rubenfeld e i suoicollaboratori hanno però trovato cheessa era regolarmente prodotta dai lin-fociti coltivati assieme ai cheratinocitiumani; al contrario, era assente in tuttele colture di controllo. Evidentementele cellule epidermiche avevano stimola-to in qualche modo i linfociti T immatu-ri (che erano presumibilmente presentinel sangue o nel midollo osseo saggiato,a seconda dell'esperimento) a sintetiz-zarla, un evento che normalmente haluogo sotto il controllo del timo. L'en-zima è stato trovato solo nei linfocitiche avevano sintetizzato nuovo DNA inpreparazione della divisione cellulare.(Tuttavia, non era stata chiaramente lasintesi di questo DNA a segnalare laproduzione di dTT, dato che era possi-bile dimostrare che vari altri stimoli alladivisione cellulare non producevanorisultati analoghi.)
Sembrava chiaro che, in questi espe-rimenti, i responsabili dell'influenza del-la pelle sulla maturazione dei linfociti Tfossero i cheratinociti, poiché le coltureerano prive di altri tipi di cellule epider-miche. Era sorprendente trovare che untipo di cellula epidermica, in precedenzaritenuto responsabile principalmentedella produzione della cheratina, potes-se svolgere un ruolo importante nellabiologia dei linfociti T. Certo, la pellenon può sostituire completamente lafunzione del timo, poiché i topi in cui iltimo viene rimosso chirurgicamente su-bito dopo la nascita non sviluppano si-stemi normali di linfociti T. I risultatidella Rubenfeld hanno suggerito, tutta-via, che i cheratinociti esercitano un'in-fluenza su alcuni stadi post-timici dellamaturazione dei linfociti T.
Circa nello stesso periodo, Bijan Safaie collaboratori, al Memorial Sloan-Ket-
CELLULA STAMINALE
TRASFORMAZIONE NEOPLASTICADI LINFOCITI T COADIUVANTI
PLASMACELLULA
,RMA DIFFUSIONE IN TUTTO IL CORPOIN PROFONDITÀ ATTRAVERSO IL SANGUE
Il linfoma cutaneo di tipo T è una neoplasia dei linfociti T coadiuvanti. Sia i linfociti T sia ilinfocitiB hanno origine nel midollo osseo. Alcune cellule staminali del midollo si trasformano,maturandosi, in linfociti B, che sintetizzano anticorpi specifici e li espongono alla propriasuperficie; quando poi vengono stimolati da un antigene specifico, si trasformano in plasmacel-lule, la cui funzione è di secernere attivamente anticorpi. Altre cellule staminali maturano sottol'influenza del timo e diventano linfociti T. Alcuni di questi appartengono alle classi di linfocitiT dette «coadiuvanti» e «soppressori», che rispettivamente potenziano e inibiscono il funzio-namento dei linfocitiB e di altri linfociti T. I linfociti T coadiuvanti che si trasformano in celluleneoplastiche spesso possiedono un'elevata affinità per l'epidermide, si accumulano in prossimi-tà di essa e vi penetrano. A mano a mano che insorgono nuovi subcloni di linfociti T neoplastici,essi perdono la loro affinità per l'epidermide e danno luogo a metastasi negli organi interni.
50 51
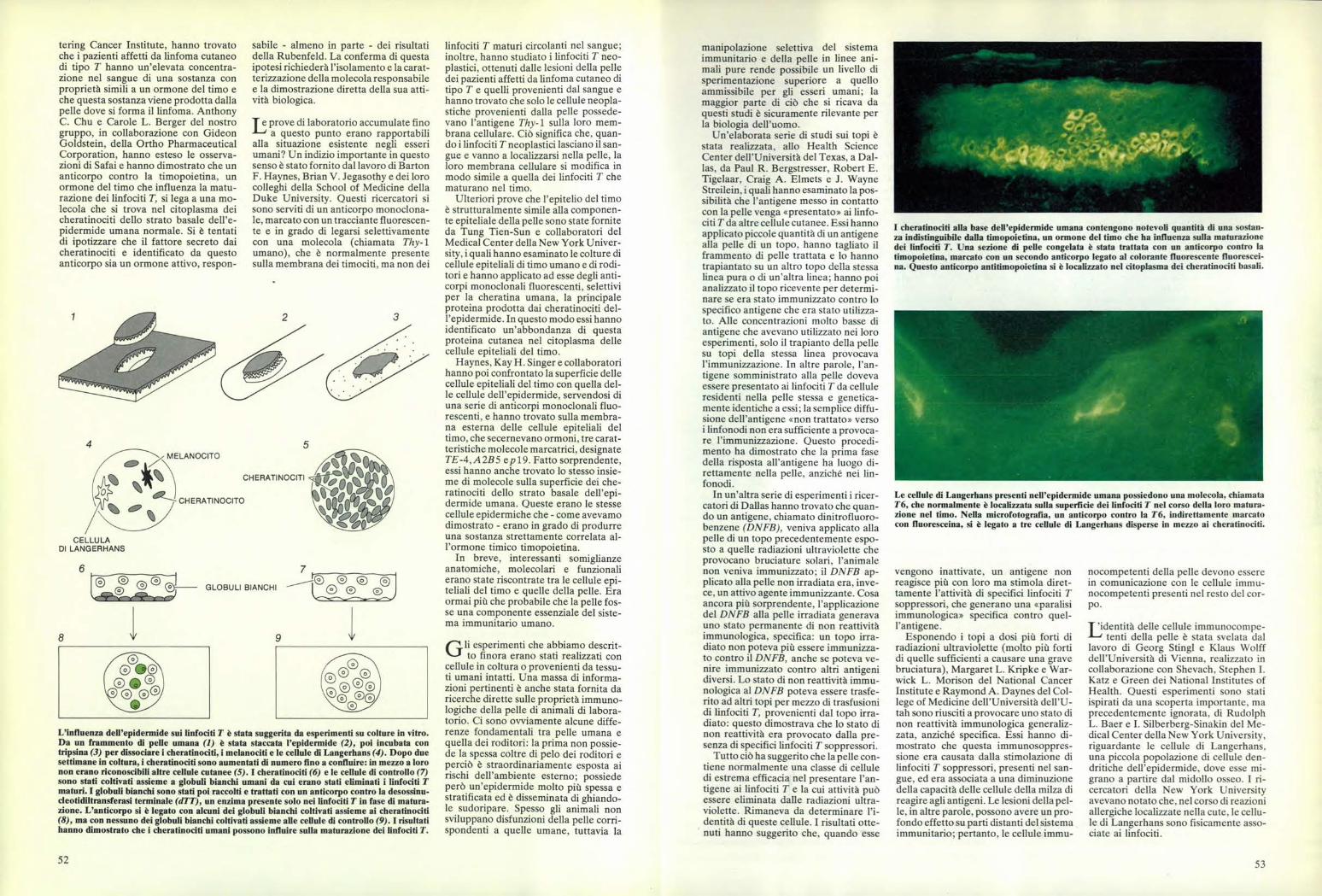
I cheratinociti alla base dell'epidermide umana contengono notevoli quantità di una sostan-za indistinguibile dalla timopoietina. un ormone del timo che ha influenza sulla maturazionedei linfociti T. Una sezione di pelle congelata è stata trattata con un anticorpo contro latimopoietina. marcato con un secondo anticorpo legato al colorante fluorescente fluorescei-na. Questo anticorpo antitimopoietina si è localizzato nel citoplasma dei cheratinociti basali.
Le cellule di Langerhans presenti nell'epidermide umana possiedono una molecola, chiamata76, che normalmente è localizzata sulla superficie dei linfociti T nel corso della loro matura-zione nel timo. Nella microfotografia, un anticorpo contro la 76, indirettamente marcatocon fluoresceina, si è legato a tre cellule di Langerhans disperse in mezzo ai cheratinociti.
4 5
2
MELANOCITO
CHERATINOCITI
CHERATINOCITO
CELLULADI LANGERHANS
6 7
GLOBULI BIANCHI
tering Cancer Institute, hanno trovatoche i pazienti affetti da linfoma cutaneodi tipo T hanno un'elevata concentra-zione nel sangue di una sostanza conproprietà simili a un ormone del timo eche questa sostanza viene prodotta dallapelle dove si forma il linfoma. AnthonyC. Chu e Carole L. Berger del nostrogruppo, in collaborazione con GideonGoldstein, della Ortho PharmaceuticalCorporation, hanno esteso le osserva-zioni di Safai e hanno dimostrato che unanticorpo contro la timopoietina, unormone del timo che influenza la matu-razione dei linfociti T, si lega a una mo-lecola che si trova nel citoplasma deicheratinociti dello strato basale dell'e-pidermide umana normale. Si è tentatidi ipotizzare che il fattore secreto daicheratinociti e identificato da questoanticorpo sia un ormone attivo, respon-
8
sabile - almeno in parte - dei risultatidella Rubenfeld. La conferma di questaipotesi richiederà l'isolamento e la carat-terizzazione della molecola responsabilee la dimostrazione diretta della sua atti-vità biologica.
e prove di laboratorio accumulate fino a questo punto erano rapportabilialla situazione esistente negli esseriumani? Un indizio importante in questosenso è stato fornito dal lavoro di BartonF. Haynes, Brian V. Jegasothy e dei lorocolleghi della School of Medicine dellaDuke University. Questi ricercatori sisono serviti di un anticorpo monoclona-le, marcato con un tracciante fluorescen-te e in grado di legarsi selettivamentecon una molecola (chiamata Thy-1umano), che è normalmente presentesulla membrana dei timociti, ma non dei
9
linfociti T maturi circolanti nel sangue;inoltre, hanno studiato i linfociti T neo-plastici, ottenuti dalle lesioni della pelledei pazienti affetti da linfoma cutaneo ditipo T e quelli provenienti dal sangue ehanno trovato che solo le cellule neopla-stiche provenienti dalla pelle possede-vano l'antigene Thy-1 sulla loro mem-brana cellulare. Ciò significa che, quan-do i linfociti T neoplastici lasciano il san-gue e vanno a localizzarsi nella pelle, laloro membrana cellulare si modifica inmodo simile a quella dei linfociti T chematurano nel timo.
Ulteriori prove che l'epitelio del timoè strutturalmente simile alla componen-te epiteliale della pelle sono state forniteda Tung Tien-Sun e collaboratori delMedical Center della New York Univer-sity, i quali hanno esaminato le colture dicellule epiteliali di timo umano e di rodi-tori e hanno applicato ad esse degli anti-corpi monoclonali fluorescenti, selettiviper la cheratina umana, la principaleproteina prodotta dai cheratinociti del-l'epidermide. In questo modo essi hannoidentificato un'abbondanza di questaproteina cutanea nel citoplasma dellecellule epiteliali del timo.
Haynes, Kay H. Singer e collaboratorihanno poi confrontato la superficie dellecellule epiteliali del timo con quella del-le cellule dell'epidermide, servendosi diuna serie di anticorpi monoclonali fluo-rescenti, e hanno trovato sulla membra-na esterna delle cellule epiteliali deltimo, che secernevano ormoni, tre carat-teristiche molecole marcatrici, designateTE-4,A 2B5 e p19. Fatto sorprendente,essi hanno anche trovato lo stesso insie-me di molecole sulla superficie dei che-ratinociti dello strato basale dell'epi-dermide umana. Queste erano le stessecellule epidermiche che - come avevamodimostrato - erano in grado di produrreuna sostanza strettamente correlata al-l'ormone timico timopoietina.
In breve, interessanti somiglianzeanatomiche, molecolari e funzionalierano state riscontrate tra le cellule epi-teliali del timo e quelle della pelle. Eraormai più che probabile che la pelle fos-se una componente essenziale del siste-ma immunitario umano.
Gli esperimenti che abbiamo descrit-to finora erano stati realizzati con
cellule in coltura o provenienti da tessu-ti umani intatti. Una massa di informa-zioni pertinenti è anche stata fornita daricerche dirette sulle proprietà immuno-logiche della pelle di animali di labora-torio. Ci sono ovviamente alcune diffe-renze fondamentali tra pelle umana equella dei roditori: la prima non possie-de la spessa coltre di pelo dei roditori eperciò è straordinariamente esposta airischi dell'ambiente esterno; possiedeperò un'epidermide molto più spessa estratificata ed è disseminata di ghiando-le sudoripare. Spesso gli animali nonsviluppano disfunzioni della pelle corri-spondenti a quelle umane, tuttavia la
manipolazione selettiva del sistemaimmunitario e della pelle in linee ani-mali pure rende possibile un livello disperimentazione superiore a quelloammissibile per gli esseri umani; lamaggior parte di ciò che si ricava daquesti studi è sicuramente rilevante perla biologia dell'uomo.
Un'elaborata serie di studi sui topi èstata realizzata, allo Health ScienceCenter dell'Università del Texas, a Dal-las, da Paul R. Bergstresser, Robert E.Tigelaar, Craig A. Elmets e J. WayneStreilein, i quali hanno esaminato la pos-sibilità che l'antigene messo in contattocon la pelle venga «presentato» ai linfo-citi T da altre cellule cutanee. Essi hannoapplicato piccole quantità di un antigenealla pelle di un topo, hanno tagliato ilframmento di pelle trattata e lo hannotrapiantato su un altro topo della stessalinea pura o di un'altra linea; hanno poianalizzato il topo ricevente per determi-nare se era stato immunizzato contro lospecifico antigene che era stato utilizza-to. Alle concentrazioni molto basse diantigene che avevano utilizzato nei loroesperimenti, solo il trapianto della pellesu topi della stessa linea provocaval'immunizzazione. In altre parole, l'an-tigene somministrato alla pelle dovevaessere presentato ai linfociti T da celluleresidenti nella pelle stessa e genetica-mente identiche a essi; la semplice diffu-sione dell'antigene «non trattato» versoi linfonodi non era sufficiente a provoca-re l'immunizzazione. Questo procedi-mento ha dimostrato che la prima fasedella risposta all'antigene ha luogo di-rettamente nella pelle, anziché nei lin-fonodi.
In un'altra serie di esperimenti i ricer-catori di Dallas hanno trovato che quan-do un antigene, chiamato dinitrofluoro-benzene (DNFB), veniva applicato allapelle di un topo precedentemente espo-sto a quelle radiazioni ultraviolette cheprovocano bruciature solari, l'animalenon veniva immunizzato; il DNFB ap-plicato alla pelle non irradiata era, inve-ce, un attivo agente immunizzante. Cosaancora più sorprendente, l'applicazionedel DNFB alla pelle irradiata generavauno stato permanente di non reattivitàimmunologica, specifica: un topo irra-diato non poteva più essere immunizza-to contro il DNFB, anche se poteva ve-nire immunizzato contro altri antigenidiversi. Lo stato di non reattività immu-nologica al DNFB poteva essere trasfe-rito ad altri topi per mezzo di trasfusionidi linfociti T, provenienti dal topo irra-diato: questo dimostrava che lo stato dinon reattività era provocato dalla pre-senza di specifici linfociti T soppressori.
Tutto ciò ha suggerito che la pelle con-tiene normalmente una classe di celluledi estrema efficacia nel presentare l'an-tigene ai linfociti T e la cui attività puòessere eliminata dalle radiazioni ultra-violette. Rimaneva da determinare l'i-dentità di queste cellule. I risultati otte-nuti hanno suggerito che, quando esse
vengono inattivate, un antigene nonreagisce più con loro ma stimola diret-tamente l'attività di specifici linfociti Tsoppressori, che generano una «paralisiimmunologica» specifica contro quel-l'antigene.
Esponendo i topi a dosi più forti diradiazioni ultraviolette (molto più fortidi quelle sufficienti a causare una gravebruciatura), Margaret L. Kripke e War-wick L. Morison del National CancerInstitute e Raymond A. Daynes del Col-lege of Medicine dell'Università dell'U-tah sono riusciti a provocare uno stato dinon reattività immunologica generaliz-zata, anziché specifica. Essi hanno di-mostrato che questa immunosoppres-sione era causata dalla stimolazione dilinfociti T soppressori, presenti nel san-gue, ed era associata a una diminuzionedella capacità delle cellule della milza direagire agli antigeni. Le lesioni della pel-le, in altre parole, possono avere un pro-fondo effetto su parti distanti del sistemaimmunitario; pertanto, le cellule immu-
nocompetenti della pelle devono esserein comunicazione con le cellule immu-nocompetenti presenti nel resto del cor-po.
J',identità delle cellule immunocompe-tenti della pelle è stata svelata dal
lavoro di Georg Stingi e Klaus Wolffdell'Università di Vienna, realizzato incollaborazione con Shevach, Stephen I.Katz e Green dei National Institutes ofHealth. Questi esperimenti sono statiispirati da una scoperta importante, maprecedentemente ignorata, di RudolphL. Baer e I. Silberberg-Sinakin del Me-dical Center della New York University,riguardante le cellule di Langerhans,una piccola popolazione di cellule den-dritiche dell'epidermide, dove esse mi-grano a partire dal midollo osseo. I ri-cercatori della New York Universityavevano notato che, nel corso di reazioniallergiche localizzate nella cute, le cellu-le di Langerhans sono fisicamente asso-ciate ai linfociti.
L'influenza dell'epidermide sui linfociti T è stata suggerita da esperimenti su colture in vitro.Da un frammento di pelle umana (1) è stata staccata l'epidermide (2), poi incubata contripsina (3) per dissociare i cheratinociti, i melanociti e le cellule di Langerhans (4). Dopo duesettimane in coltura, i cheratinociti sono aumentati di numero fino a confluire: in mezzo a loronon erano riconoscibili altre cellule cutanee (5). I cheratinociti (6) e le cellule di controllo (7)sono stati coltivati assieme a globuli bianchi umani da cui erano stati eliminati i linfociti Tmaturi. I globuli bianchi sono stati poi raccolti e trattati con un anticorpo contro la desossinu-cleotidiltransferasi terminale (dTT), un enzima presente solo nei linfociti T in fase di matura-zione. L'anticorpo si è legato con alcuni dei globuli bianchi coltivati assieme ai cheratinociti(8), ma con nessuno dei globuli bianchi coltivati assieme alle cellule di controllo (9). I risultatihanno dimostrato che i cheratinociti umani possono influire sulla maturazione dei linfociti T.
52 53
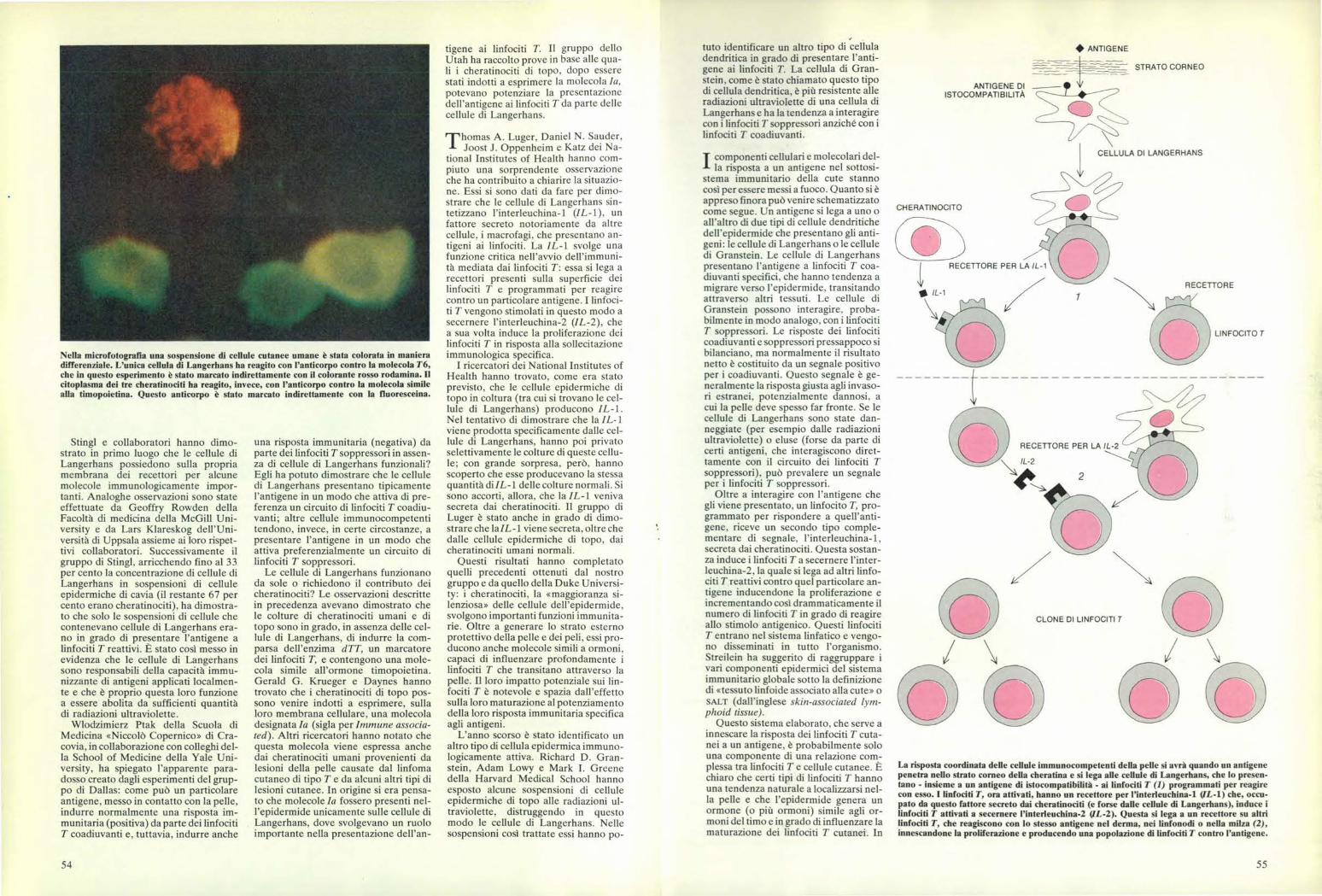
Nella microfotografia una sospensione di cellule cutanee umane è stata colorata in manieradifferenziale. L'unica cellula di Langerhans ha reagito con l'anticorpo contro la molecola T6,che in questo esperimento è stato marcato indirettamente con il colorante rosso rodamina. Ilcitoplasma dei tre cheratinociti ha reagito, invece, con l'anticorpo contro la molecola similealla timopoietina. Questo anticorpo è stato marcato indirettamente con la fluoresceina.
RECETTORE PER LA IL-2
2
CLONE DI LINFOCITI T
CHERATINOCITO
o RECETTORE PER LA IL-1
1RECETTOR E
LINFOCITO T
Stingi e collaboratori hanno dimo-strato in primo luogo che le cellule diLangerhans possiedono sulla propriamembrana dei recettori per alcunemolecole immunologicamente impor-tanti. Analoghe osservazioni sono stateeffettuate da Geoffry Rowden dellaFacoltà di medicina della McGill Uni-versity e da Lars Klareskog dell'Uni-versità di Uppsala assieme ai loro rispet-tivi collaboratori. Successivamente ilgruppo di Stingl, arricchendo fino al 33per cento la concentrazione di cellule diLangerhans in sospensioni di celluleepidermiche di cavia (il restante 67 percento erano cheratinociti), ha dimostra-to che solo le sospensioni di cellule checontenevano cellule di Langerhans era-no in grado di presentare l'antigene alinfociti T reattivi. È stato così messo inevidenza che le cellule di Langerhanssono responsabili della capacità immu-nizzante di antigeni applicati localmen-te e che è proprio questa loro funzionea essere abolita da sufficienti quantitàdi radiazioni ultraviolette.
Wlodzimierz Ptak della Scuola diMedicina «Niccolò Copernico» di Cra-covia, in collaborazione con colleghi del-la School of Medicine della Yale Uni-versity, ha spiegato l'apparente para-dosso creato dagli esperimenti del grup-po di Dallas: come può un particolareantigene, messo in contatto con la pelle,indurre normalmente una risposta im-munitaria (positiva) da parte dei linfocitiT coadiuvanti e, tuttavia, indurre anche
una risposta immunitaria (negativa) daparte dei linfociti T soppressori in assen-za di cellule di Langerhans funzionali?Egli ha potuto dimostrare che le celluledi Langerhans presentano tipicamentel'antigene in un modo che attiva di pre-ferenza un circuito di linfociti T coadiu-vanti; altre cellule immunocompetentitendono, invece, in certe circostanze, apresentare l'antigene in un modo cheattiva preferenzialmente un circuito dilinfociti T soppressori.
Le cellule di Langerhans funzionanoda sole o richiedono il contributo deicheratinociti? Le osservazioni descrittein precedenza avevano dimostrato chele colture di cheratinociti umani e ditopo sono in grado, in assenza delle cel-lule di Langerhans, di indurre la com-parsa dell'enzima dTT, un marcatoredei linfociti T, e contengono una mole-cola simile all'ormone timopoietina.Gerald G. Krueger e Daynes hannotrovato che i cheratinociti di topo pos-sono venire indotti a esprimere, sullaloro membrana cellulare, una molecoladesignata la (sigla per Immune associa-ted). Altri ricercatori hanno notato chequesta molecola viene espressa anchedai cheratinociti umani provenienti dalesioni della pelle causate dal linfomacutaneo di tipo T e da alcuni altri tipi dilesioni cutanee. In origine si era pensa-to che molecole la fossero presenti nel-l'epidermide unicamente sulle cellule diLangerhans, dove svolgevano un ruoloimportante nella presentazione dell'an-
tigene ai linfociti T. Il gruppo delloUtah ha raccolto prove in base alle qua-li i cheratinociti di topo, dopo esserestati indotti a esprimere la molecola la,potevano potenziare la presentazionedell'antigene ai linfociti T da parte dellecellule di Langerhans.
Thomas A. Luger, Daniel N. Sauder,
Joost J. Oppenheim e Katz dei Na-tional Institutes of Health hanno com-piuto una sorprendente osservazioneche ha contribuito a chiarire la situazio-ne. Essi si sono dati da fare per dimo-strare che le cellule di Langerhans sin-tetizzano l'interleuchina-1 (IL-1), unfattore secreto notoriamente da altrecellule, i macrofagi, che presentano an-tigeni ai linfociti. La IL-1 svolge unafunzione critica nell'avvio dell'immuni-tà mediata dai linfociti T: essa si lega arecettori presenti sulla superficie deilinfociti T e programmati per reagirecontro un particolare antigene. I linfoci-ti T vengono stimolati in questo modo asecernere l'interleuchina-2 (IL-2), chea sua volta induce la proliferazione deilinfociti T in risposta alla sollecitazioneimmunologica specifica.
I ricercatori dei National Institutes ofHealth hanno trovato, come era statoprevisto, che le cellule epidermiche ditopo in coltura (tra cui si trovano le cel-lule di Langerhans) producono IL-1.Nel tentativo di dimostrare che la IL-1viene prodotta specificamente dalle cel-lule di Langerhans, hanno poi privatoselettivamente le colture di queste cellu-le; con grande sorpresa, però, hannoscoperto che esse producevano la stessaquantità di IL- I delle colture normali. Sisono accorti, allora, che la IL-1 venivasecreta dai cheratinociti. Il gruppo diLuger è stato anche in grado di dimo-strare che la IL-1 viene secreta, oltre chedalle cellule epidermiche di topo, daicheratinociti umani normali.
Questi risultati hanno completatoquelli precedenti ottenuti dal nostrogruppo e da quello della Duke Universi-ty: i cheratinociti, la «maggioranza si-lenziosa» delle cellule dell'epidermide,svolgono importanti funzioni immunita-rie. Oltre a generare lo strato esternoprotettivo della pelle e dei peli, essi pro-ducono anche molecole simili a ormoni,capaci di influenzare profondamente ilinfociti T che transitano attraverso lapelle. Il loro impatto potenziale sui lin-fociti T è notevole e spazia dall'effettosulla loro maturazione al potenziamentodella loro risposta immunitaria specificaagli antigeni.
L'anno scorso è stato identificato unaltro tipo di cellula epidermica immuno-logicamente attiva. Richard D. Gran-stein, Adam Lowy e Mark I. Greenedella Harvard Medical School hannoesposto alcune sospensioni di celluleepidermiche di topo alle radiazioni ul-traviolette, distruggendo in questomodo le cellule di Langerhans. Nellesospensioni così trattate essi hanno po-
tuto identificare un altro tipo di celluladendritica in grado di presentare l'anti-gene ai linfociti T. La cellula di Gran-stein, come è stato chiamato questo tipodi cellula dendritica, è più resistente alleradiazioni ultraviolette di una cellula diLangerhans e ha la tendenza a interagirecon i linfociti T soppressori anziché con ilinfociti T coadiuvanti.
T componenti cellulari e molecolari del-la risposta a un antigene nel sottosi-
stema immunitario della cute stannocosì per essere messi a fuoco. Quanto si èappreso finora può venire schematizzatocome segue. Un antigene si lega a uno oall'altro di due tipi di cellule dendritichedell'epidermide che presentano gli anti-geni: le cellule di Langerhans o le celluledi Granstein. Le cellule di Langerhanspresentano l'antigene a linfociti T coa-diuvanti specifici, che hanno tendenza amigrare verso l'epidermide, transitandoattraverso altri tessuti. Le cellule diGranstein possono interagire, proba-bilmente in modo analogo, con i linfocitiT soppressori. Le risposte dei linfociticoadiuvanti e soppressori pressappoco sibilanciano, ma normalmente il risultatonetto è costituito da un segnale positivoper i coadiuvanti. Questo segnale è ge-neralmente la risposta giusta agli invaso-ri estranei, potenzialmente dannosi, acui la pelle deve spesso far fronte. Se lecellule di Langerhans sono state dan-neggiate (per esempio dalle radiazioniultraviolette) o eluse (forse da parte dicerti antigeni, che interagiscono diret-tamente con il circuito dei linfociti Tsoppressori), può prevalere un segnaleper i linfociti T soppressori.
Oltre a interagire con l'antigene chegli viene presentato, un linfocito T, pro-grammato per rispondere a quell'anti-gene, riceve un secondo tipo comple-mentare di segnale, l'interleuchina-1,secreta dai cheratinociti. Questa sostan-za induce i linfociti T a secernere l'inter-leuchina-2, la quale si lega ad altri linfo-citi T reattivi contro quel particolare an-tigene inducendone la proliferazione eincrementando così drammaticamente ilnumero di linfociti T in grado di reagireallo stimolo antigenico. Questi linfocitiT entrano nel sistema linfatico e vengo-no disseminati in tutto l'organismo.Streilein ha suggerito di raggruppare ivari componenti epidermici del sistemaimmunitario globale sotto la definizionedi «tessuto linfoide associato alla cute» oSALT (dall'inglese skin-associated lym-phoid tissue).
Questo sistema elaborato, che serve ainnescare la risposta dei linfociti T cuta-nei a un antigene, è probabilmente solouna componente di una relazione com-plessa tra linfociti T e cellule cutanee. Èchiaro che certi tipi di linfociti T hannouna tendenza naturale a localizzarsi nel-la pelle e che l'epidermide genera unormone (o più ormoni) simile agli or-moni del timo e in grado di influenzare lamaturazione dei linfociti T cutanei. In
• ANTIGENE
_----71---_-r,,-- _ — STRATO CORNEO
ANTIGENE DIISTOCOMPATIBILITA
CELLULA DI LANGERHANS
La risposta coordinata delle cellule immunocompetenti della pelle si avrà quando un antigenepenetra nello strato corneo della cheratina e si lega alle cellule di Langerhans, che lo presen-tano - insieme a un antigene di istocompatibilità - ai linfociti T (I) programmati per reagirecon esso. I linfociti T, ora attivati, hanno un recettore per l'interleuchina-1 (IL-1) che, occu-pato da questo fattore secreto dai cheratinociti (e forse dalle cellule di Langerhans), induce ilinfociti T attivati a secernere l'interleuchina-2 (IL-2). Questa si lega a un recettore su altrilinfociti T, che reagiscono con lo stesso antigene nel derma, nei linfonodi o nella milza (2),innescandone la proliferazione e producendo una popolazione di linfociti T contro l'antigene.
54
55

I raccoglitori per 11 1985Questi raccoglitori corrispondono ai volumi
XXXIV e XXXV de LE SCIENZE, e rispettivamenteai fascicoli da gennaio (n. 197) a giugno (n. 202)
e da luglio (n. 203) a dicembre (n. 208).Sono ancora disponibili i raccoglitori dal Vol. XXVIII
al XXXII! e dei raccoglitori non numerati appositamenteapprontati per sostituire i raccoglitori esauriti.
I raccoglitori si possono richiedere direttamenteall'editore usando l'apposita cartolina allegata allarivista e unendo il relativo importo; gli ordini infatti
vengono evasi solo a pagamento avvenuto.
Ogni raccoglitore L. 4.500
I raccoglitori si trovano anche presso i seguenti punti di vendita:BOLOGNA: Libreria Parolini - Via U. Bassi 14
FIRENZE: Libreria Marzocco - Via de' Martelli 22/RGENOVA: Libreria Int. Di Stefano - Via R. Ceccardi 40/R
MILANO: Le Scienze S.p.A. - Via del Lauro 14TORINO: Libreria Zanaboni - C.so Vittorio Emanuele 41
NAPOLI: Libreria Guida A. - Via Port'Alba 20/21PADOVA: Libreria Cortina - Via F. Marzolo 4
PALERMO: Libreria Dante - Quattro Canti di CittàROMA: Claudio Aranci - Viale Europa 319 (EUR)
tutto il mondo i ricercatori stanno oggiutilizzando sistemi sperimentali animaliper decifrare ulteriormente la relazionedinamica tra epidermide e linfociti T.Una scoperta casuale avvenuta nel no-stro laboratorio potrebbe forse offrirel'opportunità di estendere questi studiagli esseri umani.
Stavamo esaminando campioni dipelle umana intatta per scoprire unamolecola chiamata T6, che si trovanormalmente sulla superficie dei linfo-citi T in via di maturazione nello stratocorticale del timo; quando questi linfo-citi abbandonano il timo non posseggo-no più la T6. Avevamo avanzato l'ipo-tesi che i linfociti T neoplastici espri-messero nuovamente la T6, la qualeservirebbe allora come marcatore e fa-ciliterebbe la diagnosi precoce del lin-foma cutaneo di tipo T. Ellen Fithian hamesso le cellule epidermiche umanenormali in presenza di un anticorpomonoclonale contro la T6, aspettandosidi trovare che esse non la esprimevanopiù. Con nostra grande sorpresa, inve-ce, ha trovato che l'anticorpo reagivacon cellule epidermiche sparse, chemancavano delle altre caratteristichedei linfociti T. La forma dendritica e ladistribuzione di queste cellule T6-posi-tive hanno suggerito che si trattasse dicellule di Langerhans. Questa identifi-cazione è stata confermata quando si èdimostrato che esse esprimevano laproteina di membrana la, caratteristicadelle cellule di Langerhans normali.Shinichero Takezaki e Sherie L. Morri-son, del nostro gruppo, hanno dimo-strato successivamente che la molecolaT6, presente sulle cellule di Langer-hans, è chimicamente indistinguibile daquella che si trova sui timociti.
La T6 sulle cellule di Langerhans(notata anche da George F. Murphy ecollaboratori a Harvard, all'incirca nellostesso periodo) costituisce un comodomarcatore per identificare rapidamentequeste cellule. Ma, fatto ancora piùimportante, essa dà luogo ad alcuni in-terrogativi provocatori. Perché le celluledi Langerhans umane esprimono unmarcatore di membrana caratteristicodei linfociti T immaturi? Le cellule diLangerhans migrano nella pelle a partiredal midollo osseo attraverso il circolosanguigno, ma il sangue degli adultinormali non contiene, di fatto, un nume-ro significativo di cellule T6-positive: lecellule di Langerhans verrebbero forseindotte a sintetizzare la molecola T6 dacellule presenti nell'epidermide? Oppu-re sono i linfociti T, sotto l'influenza del-la pelle, che sintetizzano e secernono lamolecola T6, la quale successivamenteaderirebbe alle cellule di Langerhans?La T6 (che è presente in concentrazionemolto elevata sulla superficie delle cellu-le di Langerhans) può svolgere qualchefunzione nel «trattamento» dell'antige-ne da parte della cellula? Con le tecno-logie attuali si dovrebbero facilmenteottenere delle risposte.


















