CARATTERIZZAZIONE DELL'INFILTRATO INFIAMMATORIO … · stimati nell'ordine di 14,5 cani e 3,5 gatti...
89
1 Università degli Studi di Pisa Dipartimento di Scienze Veterinarie Tesi Magistrale in Medicina Veterinaria CARATTERIZZAZIONE DELL'INFILTRATO INFIAMMATORIO NEI MENINGIOMI DEL GATTO Tesi di Laurea Magistrale di: Guido Rocchigiani Relatore: Controrelatore: Prof. Carlo Cantile Prof.sa Veronica Marchetti Co-relatore: Prof. Alessandro Poli Anno Accademico 2011/2012
Transcript of CARATTERIZZAZIONE DELL'INFILTRATO INFIAMMATORIO … · stimati nell'ordine di 14,5 cani e 3,5 gatti...

1
Università degli Studi di Pisa
Dipartimento di Scienze Veterinarie
Tesi Magistrale in Medicina Veterinaria
CARATTERIZZAZIONE DELL'INFILTRATO
INFIAMMATORIO NEI MENINGIOMI DEL GATTO
Tesi di Laurea Magistrale di:
Guido Rocchigiani
Relatore: Controrelatore:
Prof. Carlo Cantile Prof.sa Veronica Marchetti
Co-relatore:
Prof. Alessandro Poli
Anno Accademico 2011/2012

2
I meningiomi sono i tumori intracranici più frequenti nel gatto. E' stato osservato
che alcune cellule infiammatorie infiltrano questi tumori nell'uomo e nel cane, però
poco si conosce riguardo alle interazioni tra sistema immunitario e meningioma nel
gatto. Lo scopo di questo studio era quello di individuare quale tipo cellulare fosse
presente all'interno del tumore e di scoprire se fossero state presenti delle
correlazioni tra il quadro clinico del paziente e l'istotipo e il grado del tumore.
Ventisette campioni di tumore, fissati in formalina e inclusi in paraffina sono stati
analizzati utilizzando l'immunoistochimica con anticorpi specifici per CD 3, CD 79a,
CD 20 e FoxP3. I risultati rivelano la complessa fisiopatologia della risposta
immunitaria e la sua relazione con il comportamento biologico dei meningiomi felini.
Parole chiave: Cervello, Gatto, Immunoistochimica, Linfocita, Meningioma, FoxP3,
Tumore
-------------------------------------------------------------------
Abstract: Meningiomas are the most frequent intracranial tumors in cats. A variety
of inflammatory cells have been shown to infiltrate these tumors in humans and
dogs, but little is known about interactions between the immune system and
naturally occurred brain tumors in cats. The purpose of this study was to individuate
the infiltrating immune cells subset within the tumor and find out if there was any
relationship with the clinical picture of the patient and histotype and grade of the
tumor. Twenty-seven formalin fixed, paraffin embedded tumor samples evaluated
using immune-histochemistry with antibodies specific for
CD 3, CD 79a, CD 20 and FoxP3 . The Findings reveal the complex pathophysiology
of the immune response and its relationship to the biological behavior of feline
meningiomas .
Key words: Brain, Cat, Immunehistochemistry , Lymphocyte , Meningioma , FoxP3 ,
Tumor

3
Sommario Generalità: ................................................................................................................................ 4
Aspetti clinici: ........................................................................................................................... 7
Diagnosi: ................................................................................................................................. 11
Rilievi Anatomo-Patologici: .................................................................................................... 13
Classificazione dei meningiomi: ......................................................................................... 13
Genetica dei meningiomi in veterinaria: ............................................................................ 17
Caratterizzazione molecolare dei meningiomi:.................................................................. 18
Marker linfocitari : ................................................................................................................. 25
FoxP3: ..................................................................................................................................... 25
Generalità: .......................................................................................................................... 25
Natural T Regulatory lymphocytes (nTreg): ....................................................................... 26
Induced T regulatory lymphocytes (iTreg) : ....................................................................... 29
Attività immunodepressiva dei Treg: ................................................................................. 40
Treg e FoxP3 negli animali domestici: ................................................................................ 42
Tumori e linfociti T : ........................................................................................................... 43
Treg e immunità tumorale: ................................................................................................ 47
FoxP3 ed immunoterapia ................................................................................................... 49
Materiali e metodi: ................................................................................................................. 52
Campioni: ........................................................................................................................... 52
Immunoistochimica (IH): .................................................................................................... 53
Risultati: ................................................................................................................................. 55
Discussione: ............................................................................................................................ 64
Conclusioni: ............................................................................................................................ 68
Bibliografia ............................................................................................................................. 68
Ringraziamenti: ...................................................................................................................... 86

4
Generalità:
Il meningioma è la forma tumorale più comune riscontrabile a livello del sistema
nervoso centrale (SNC) sia nel cane sia nel gatto (1) (2) (3), mentre sono riportati
sporadici casi nel cavallo,vacca e pecora (4). I tumori intracranici del SNC vengono
stimati nell'ordine di 14,5 cani e 3,5 gatti ogni 100.000 soggetti . E' stato dimostrato,
in recenti analisi retrospettive, che i meningiomi costituiscono il 45% (nel cane) (5) e
l'85% (nel gatto) (6) dei tumori primari a livello intracranico, ed il 22,3% (cane) ed il
59% (gatto) dei tumori dell'intero sistema nervoso. Non è possibile parlare di
meningioma senza ripassare le strutture anatomiche da cui origina questa neoplasia
ossia le meningi.
Sono rivestimenti che coprono interamente sia l'encefalo che il midollo spinale, e
ne assicurano protezione meccanica, immunitaria e contribuiscono anche alla
regolazione della sua irrorazione sanguigna. Sono sostanzialmente divisibili in 3
componenti, ognuna delle quali possiede delle piccole differenze tra la loro
porzione spinale e quella encefalica :
1. Dura madre: la più esterna, è un tessuto connettivo fibroso, i cui fasci di fibre
collagene, intrecciati in vari sensi, ospitano fibrociti e fibre elastiche. Lungo la
colonna vertebrale, è separata dal periostio vertebrale e lo spazio che si
interpone tra l'osso e questa meninge si chiama spazio epidurale, in cui
vengono praticate iniezioni per l' anestesia locale ; nella parte encefalica invece,
la dura madre unisce il cranio con la meninge sottostante. Qua , in punti ben
precisi, sono presenti i seni venosi della dura madre ,strutture simili a vene
piuttosto grandi, intrappolate nella matrice connettivale della meninge, che
ricevono il sangue refluo da tutto quanto l'encefalo . La dura madre inoltre
presenta dei fori, attraverso cui fuoriescono sia i nervi spinali che i vasi destinati
al midollo spinale e alle meningi più profonde e,rivestendole anche un po' più
internamente, formano come un manicotto attorno ai vasi, insieme al
perinevrio e a una piccola propaggine della 2° meninge ( spazio di Virchow-
Robin, trattato più avanti ). La dura madre è chiamata anche pachimeninge o
"meninge dura".

5
2. Aracnoide: contenuta dentro la dura madre, non è vascolarizzata, avvolge e
sostiene tutte le strutture vascolari e nervose che la attraversano. E' costituita
da 2 parti: la prima è costituita da una sottile lamina di collagene e fibre
elastiche addossata alla dura madre e presenta ,su entrambe le facce, alcuni
fibroblasti particolari, in grado di esercitare anche attività fagica; la seconda si
trova più internamente ed è chiamata spazio subaracnoideo, in cui trabecole e
setti di connettivo danno origine ad un reticolo di spazi comunicanti, in cui
scorre il liquido cefalo rachidiano (o CSF - Cerebro Spinal Fluid ). Sono
considerate componenti di questa meninge, delle strutture peculiari chiamate
granulazioni subaracnoidee: sono dei rilievi di forma rotondeggiante o
irregolare, che si proiettano nella dura madre, e, più precisamente, nei suoi seni
venosi, e sono costituite da minuscole villosità, che comportano altrettanti
diverticoli; ognuna di queste è rivestita da uno strato di cellule, che , formando
come un cappuccio sulla sommità della villosità, prende contatto con
l'endotelio venoso del seno. A parte questa piccola area di fusione, le
granulazioni subaracnoidee sono completamente separate dalla dura madre;
inoltre la cavità centrale della villosità presenta un fondo cieco che è in
comunicazione con lo spazio subaracnoideo. La funzione di queste strutture
non è ancora compresa, nonostante si ritenga che costituiscano delle stazioni di
filtrazione del liquido cefalo rachidiano, prima di immettersi nel sangue.
Insieme alla meninge successiva, costituisce la leptomeninge o "meninge
molle"
3. Pia madre: è l'ultima meninge, quella che avvolge direttamente tutto quanto il
SNC. Come la precedente, è priva di vascolarizzazione ed è nutrita per
diffusione dal sangue delle strutture più profonde e dal liquido cefalo
rachidiano. Oltre a rivestire il SNC, emana come dei processi attorno ai vasi e, al
di sopra di essi, l'aracnoide fa la stessa cosa, formando complessivamente un
manicotto ; questo persiste per un certo tratto, prima di unirsi alla parete
vascolare , e nella sua lunghezza, si trova un prolungamento della cavità
subaracnoidea, che avvolge il vaso. Questo spazio perivascolare , in cui sembra

6
penetrare il liquido cefalo rachidiano, è conosciuto come "spazio di Virchow -
Robin".
Approfonditi studi dimostrano come queste neoplasie presentino una manifesta
variabilità dal punto di vista istopatologico, mostrando dei "pattern" di tipo
mesenchimale e epiteliale: per esempio la natura mesenchimale si manifesta in
proliferazioni di cellule fusiformi e la produzione di collagene, che forma un
variabile stroma fibroso; invece la presenza di cellule poligonali, munite di diverse
giunzioni intercellulari e la presenza di specifiche proteine , sono le caratteristiche
epiteliali che si riscontrano in diverse forme di questi tumori. Tutto questo forse
rispecchia il fatto che sia il mesoderma che la cresta neurale (ectoderma)
concorrono alla formazione delle meningi. (7) Si crede che questi tumori derivino
da cellule appartenenti all'aracnoide, in particolare dalle granulazioni
subaracnoidee; per questo motivo sono , generalmente , attaccati alla dura madre e
si presentano con una localizzazione ricorrente, che varia da specie a specie, proprio
in corrispondenza con le suddette strutture . Le regioni più frequenti dove si
sviluppano sono ,nel cane, le regioni olfattive / frontali, il pavimento della cavità
cranica, il chiasma ottico ,le regioni sopra e parasellari e spinali; inoltre sono
riportati alcuni casi anche nella regione cerebello- ponto-midollare (8), nello spazio
retro bulbare (9) e nell'orecchio medio (10); inoltre è stata osservata una
prevalenza maggiore nei cani meso e dolicocefalici (11) (12). Nel gatto invece le
localizzazioni maggiormente osservate sono a livello delle meningi sopratentoriali,
della tela corioidea del 3° ventricolo, e più raramente, delle meningi cerebellari e
del midollo spinale (3) (13). Generalmente le razze a pelo corto domestiche
presentano una maggiore probabilità di sviluppare meningiomi, anche se sono
riportati casi in Siamesi, Persiani e Maine coon (14) (15) .Inoltre ,nei felini domestici,
non sono rari meningiomi multipli (16), presentandosi circa nel 17 % dei casi della
neoplasia in questione; in alcuni casi, si assiste anche alla contemporanea presenza
di forme benigne e maligne (17) mentre nel cane è molto rara la presenza di più di
un meningioma (18). Nel gatto è stata osservata una maggiore prevalenza in animali
di mezza età, sopra i 9 anni nonostante siano stati diagnosticati anche in gatti di

7
pochi mesi, spesso associati ad una mucopolisaccaridosi di tipo I (19); a differenza
dei cani c'è una ,se pur lieve, maggiore predispozione nel sesso maschile rispetto a
quello femminile (3:2 circa) (14). I meningiomi, sia felini che canini, sono stati
diagnosticati con concorrenti malattie nervose o anche extranervose: Lobetti, per
esempio, ha riportato un contemporaneo linfoma timico (20) oppure anche la
sopracitata mucopolisaccaridosi di tipo I insieme ad altri tipi di neoplasie; in
particolare è stato riscontrato che il 13,9% dei gatti e il 19% dei cani presentava,
insieme ad un meningioma, altre malattie neoplastiche intracraniche.
Aspetti clinici (21):
Come in tutti i tipi di neuropatologie, l'aspetto clinico dipende moltissimo dalla
localizzazione della lesione. Queste forme tumorali , nella maggior parte dei casi,
tendono ad avere un accrescimento espansivo ed è la compressione del
parenchima circostante a determinare i segni clinici della malattia. Proprio per
queste ragioni i meningiomi spinali, che costituiscono solo il 4% della casistica di
tutti questi tumori (22), presentano un quadro clinico molto diverso da quelli
intracranici. La massa tumorale infatti può presentarsi in qualsiasi punto delle
meningi e ,premendo sul midollo spinale, generare segni clinici diversi a seconda del
punto dove compare il tumore. Per esempio un meningioma (prendendo in
considerazione una massa tumorale che comprime molto il midollo spinale) che
compare a livello di T 7-8 genera deficit neurologici a livello degli arti posteriori ma
non a quelli anteriori; invece se il tumore si presenta a livello C4-C5, si
manifesteranno segni neurologici anche a quelli anteriori. In particolar modo, per
poter localizzare il tumore , approssimativamente, bisogna verificare la presenza di
deficit neurologici a livello degli arti e se questi corrispondono ad una sindrome da
motoneurone superiore o sindrome da motoneurone inferiore ( nella letteratura
anglosassone chiamate rispettivamente UMN o LMN syndrome). I segni clinici di
queste 2 sindromi sono riportati nella tabella seguente:

8
Riflessi
spinali
Atrofia
muscolare
Tono
muscolare
Paresi /
paralisi
UMNS Normali o
aumentati
Tardiva e
moderata
Normale o
aumentato
Possibili
entrambe
LMNS Diminuiti o
assenti
Precoce e
intensa
Diminuito Possibili
entrambe
Tabella 1 Schematizzazione dei segni clinici per gli UMN e LMN
Lo stato dei riflessi spinali, come quello patellare, è l'indice più importante per
differenziare se la lesione è presente nel motoneurone che innerva il muscolo(LMN)
o se è in un nervo situato a monte(UMN). Questo dipende dal fatto che l'intero
circuito responsabile di questi riflessi ( ossia il neurone sensitivo e quello motore)
sono situati nello stesso segmento midollare; se il riflesso manca, indica ,con ogni
probabilità, che la lesione è confinata in quella porzione di midollo spinale , che
innerva quell'arto ( per l'anteriore è C6 - T2, per il posteriore L4-S2 ). Se invece la
lesione è a monte l'arco riflesso sarà integro e l'intensità della risposta sarà normale
o addirittura aumentata, per l'interruzione di alcune connessioni con nuclei presenti
nell'encefalo,che modulano l'intensità di contrazione della muscolatura scheletrica.
La figura nella pagina seguente (Fig. 1) indica la localizzazione ( in rosso) del danno
( e quindi anche della massa tumorale) in base alla presentazione dei sopracitati
segni clinici. Quanto detto finora è uno schema applicabile a tutte le forme tumorali
che premono sul midollo spinale, dato che sono clinicamente molto difficili da
distinguere tra loro. La crescita lenta del tumore causa una progressiva
compressione sul midollo spinale, che porta ad un decorso generalmente cronico
dei segni clinici quali progressiva paresi / paralisi, atassia , incontinenza urinaria . Il
decorso e la gravità dei segni clinici varia da lieve a molto elevata, in base al tasso di
crescita del tumore, la sua localizzazione e il grado di compressione esercitato sul
midollo spinale (22).

9
Figura 1: Localizzazione della lesione spinale in base alla ai segni da motoneuroni a livello degli arti
I meningiomi intracranici ( difficilmente differenziabili anch'essi da altre neoplasie
intracraniche) sono caratterizzati invece da segni clinici molto variabili,anche per
intensità, a causa della moltitudine delle strutture nervose che possono
danneggiare; nonostante questo, nel 50% dei gatti sono asintomatici e costituiscono
dei reperti accidentali in sede necroscopica (14). La massa tumorale altera la
normale fisiologia nervosa a causa della compressione che esercita sul parenchima
circostante( che può portare anche alla distruzione dello stesso), ma può anche
interferire con la circolazione vascolare ( portando a edema cerebrale) e del liquido
cefalo rachidiano . Questi ultimi due effetti causano un aumento della pressione
intracranica, che può provocare l'erniazione del cervello oltre la lamina del tentorio
o del cerveletto oltre il foramen magnum.
Arto posteriore
UMNS
Arti toracici normali
T2 - L3
Arti toracici UMNS
Cervello o C1 - C5
Arti toracici LMNS
C6 - T2
LMNS
Arti toracici normali
L4-S2
Arti toracici UMNS
L4 - S2 e cervello o C1-C5
Arti toracici LMNS
L4 - S2 E C6 - T2
Normale
Arti toracici UMNS
raro ( cervello o C1 - C5)
Arti toracici LMNS
Plesso brachiale

10
I meningiomi possono diventare piuttosto grandi prima di dare una sintomatologia
clinica, che, molto spesso, esordisce semplicemente con una alterazione del
comportamento. Oltre a questo stato, che può protrarsi anche per diversi mesi
senza altri deficit neurologici, il primo sintomo di neoplasia intracranica, soprattutto
nel cane, è spesso una crisi convulsiva (23). Mano a mano che la neoplasia aumenta
di volume, possono comparire altri segni neurologici e le crisi convulsive si possono
presentare più frequenti e refrattarie al trattamento con anticonvulsivanti. Se il
meningioma è localizzato nel prosencefalo i sintomi più comuni sono ,oltre
all'alterazione comportamentale, presenza di "circling" ( continui di movimenti in
cerchio), cecità (o amaurosi) , deficit della risposta alla minaccia, con i riflessi
palpebrali e fotomotori normali , diminuita propriocezione controlaterale e deficit
sensitivi facciali (22). La causa del "circling" non è totalmente chiara e si pensa che
una disfunzione talamica sia la responsabile, mentre per le alterazioni della vista si
ritiene che i deficit visivi siano dovuti o alla pressione sulla radiazione ottica ( o
tratto genicolo-striato) o sulla corteccia visiva (occipitale) (14). Meningiomi nella
restante parte dell'encefalo presentano invece una sintomatologia piuttosto
differente: quelli a livello del tronco encefalico possono causare emi o tetraparesi e
deficit dei nervi cranici ( tranne del I e del II paio, dato che hanno le loro emergenze
nel prosencefalo ) mentre quelli che si localizzano nel cervelletto possono provocare
atassia, dismetria, tremori e "circling" (24) (25). I meningiomi che si formano
nell'angolo ponto-cerebellare sono invece spesso associati alla sindrome vestibolare
paradossa: con questo termine si intende un quadro neurologico sovrapponibile
con quello di danno vestibolare (con segni neurologici quali inclinazione della testa,
nistagmo) ,che però si presentano dal lato opposto rispetto alla forma classica (13).
In alcuni casi però, la precisa localizzazione neurologica non può essere rilevata, a
causa di segni clinici prodotti da complicazioni del tumore, quali edema cerebrale,
idrocefalo da ostruzione o erniazione cerebrale.

11
Diagnosi: Data la variabilità con cui si presenta al punto di vista clinico la malattia, la diagnosi
a volte può essere insidiosa. Sicuramente in qualsiasi gatto con più di 7 anni, con
deficit neurologici, che progrediscono lentamente e che non esordiscono in maniera
iperacuta/acuta, bisogna inserire tra i primi posti, nella diagnosi differenziale, una
neoplasia intracranica. Esami di laboratorio quali emogramma, biochimica clinica e
analisi delle urine purtroppo non ci danno alcun tipo di informazione sulla presenza
o meno della patologia in questione; nemmeno l'analisi del liquido cefalo rachidiano,
tecnica diagnostica tra le più importanti le malattie del sistema nervoso, sembra
essere d'ausilio per la diagnosi di meningioma. In primo luogo perchè il prelievo del
CSF è sconsigliato in tutti i casi dove è presente un aumento della pressione
intracranica, segno a volte presente in caso di meningioma: infatti il prelievo,
normalmente svolto a livello della cisterna cerebello-midollare, determina una
riduzione della pressione a valle dell' encefalo, e questo, in caso di aumentata
pressione intracranica, può portare ad una erniazione del cervelletto o del cervello;
inoltre gli unici reperti riscontrabili sono un aumento delle proteine ed una
lieve/moderata pleiocitosi, prevalentemente neutrofilica (3), reperti comuni in
diverse malattie. La diagnostica per immagini rappresenta uno strumento
diagnostico sicuramente più efficace, rispetto a quelli sopracitati. Nel caso di
meningiomi spinali, la radiografia semplice (anche se effettuata correttamente ) non
sempre permette di rivelare la presenza di un meningioma, anche se l'espansione
del tumore può allargare uno o più spazi intervertebrali e ridurre lievemente lo
spessore dell'osso che lo contiene, mentre la mielografia a contrasto è senza dubbio
più affidabile: è una tecnica che permette di localizzare abbastanza precisamente la
massa, valutare la sua grandezza e verificare se il tumore è extradurale, intradurale
e extramidollare, o intramidollare. I tumori intradurali e extramidollari, come il
meningioma, sono i tumori delle guaine nervose, il sarcoma, il neuroepitelioma, il
linfoma o altri tumori metastatici ed appaiono alla radiografia con un "pattern" a
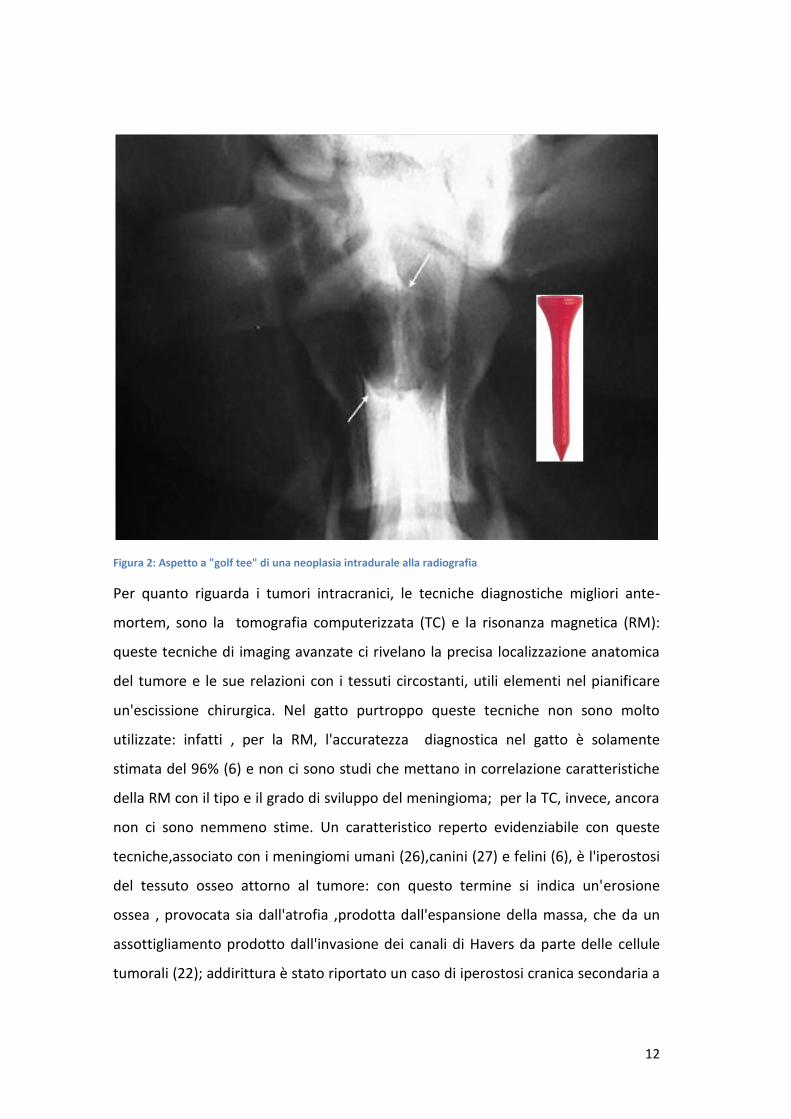
golf tee ( come indicato nella foto seguente) .
12
Figura 2: Aspetto a "golf tee" di una neoplasia intradurale alla radiografia
Per quanto riguarda i tumori intracranici, le tecniche diagnostiche migliori ante-
mortem, sono la tomografia computerizzata (TC) e la risonanza magnetica (RM):
queste tecniche di imaging avanzate ci rivelano la precisa localizzazione anatomica
del tumore e le sue relazioni con i tessuti circostanti, utili elementi nel pianificare
un'escissione chirurgica. Nel gatto purtroppo queste tecniche non sono molto
utilizzate: infatti , per la RM, l'accuratezza diagnostica nel gatto è solamente
stimata del 96% (6) e non ci sono studi che mettano in correlazione caratteristiche
della RM con il tipo e il grado di sviluppo del meningioma; per la TC, invece, ancora
non ci sono nemmeno stime. Un caratteristico reperto evidenziabile con queste
tecniche,associato con i meningiomi umani (26),canini (27) e felini (6), è l'iperostosi
del tessuto osseo attorno al tumore: con questo termine si indica un'erosione
ossea , provocata sia dall'atrofia ,prodotta dall'espansione della massa, che da un
assottigliamento prodotto dall'invasione dei canali di Havers da parte delle cellule
tumorali (22); addirittura è stato riportato un caso di iperostosi cranica secondaria a

13
meningioma, generato da una vera e propria invasione di tessuto tumorale, in un
gatto (28). Da recenti studi è emerso che le caratteristiche del meningioma rilevabili
con la RM possono essere "imitate" da altre patologie, quali forme cerebrali di
toxoplasmosi (29) e criptococcosi (30). Comunque, nonostante tutti i progressi fatti
in queste tecniche di imaging avanzate, la diagnosi definitiva può avvenire solo
tramite biopsia che può essere eseguita prendendo il campione direttamente dopo
una craniotomia, oppure mediante un complesso sistema guidato dalla TC (31).
Rilievi Anatomo-Patologici:
Classificazione dei meningiomi (32):
Macroscopicamente, nel gatto appaiono come masse globulari, con margini netti,
di un colore da grigio a giallo, solide e consistenti, a sede intradurale ed
extramidollare. La loro base d'attacco, con la dura madre o con le leptomeningi, può
essere peduncolata, ampia o totale ( in questo caso si parla di meningioma "en
plaque" ); hanno la tendenza a comprimere i tessuti circostanti,senza infiltrarli, e
possono presentarsi, come già detto, multipli. Dal punto di visto
istopatologico,possono essere classificati in diversi stadi, secondo il sistema WHO
( World Health Organization) dei meningiomi umani: questa tipizzazione prende in
considerazione le caratteristiche istopatologiche e genetiche di queste neoplasie,
che vengono raggruppate in 3 categorie, sostanzialmente in base al grado di
malignità. All'interno di questi gruppi sono presenti ulteriori sottotipi ( per un
totale di 16 ), i cui più importanti in veterinaria sono riportati nell'elenco qui
riportato:
1) Meningiomi di grado I: questo gruppo contiene la maggior parte dei tumori
benigni. Questi sono caratterizzati dal fatto di non invadere il tessuto nervoso e
di non presentare caratteristiche citologiche atipiche o anaplastiche.
a) Meningoteliale: fini fibre di collagene delineano nidi e lobuli di cellule
neoplastiche meningoteliali, a forma poligonale o epiteliode,nucleo

14
ovoidale vescicolare, spesso con un nucleolo prominente e abbondante
citoplasma; é una delle forme più comuni nel gatto.
b) Fibroblastico: sono presenti fasci di cellule sottili, con nucleo allungato,
immerse in una matrice extracellulare di collagene, che, a volte, ostacola
la visione delle cellule stesse.
c) Transizionale: presenta caratteristiche morfologiche a metà tra le 2
categorie sopracitate. Un elemento abbastanza tipico è la presenza di
cellule disposte in vortici ed è possibile la presenza di corpi
psammomatosi (concrezioni calcaree concentriche)
d) Angioblastico: la componente dominante è quella vascolare, che occupa
più del 50 % della massa tumorale; in genere i vasi sanguigni
presentano un elevato grado di ialinosi
e) Psammomatoso: è presente un elevato numero di corpi psammomasi
presenti nella massa tumorale; talvolta è talmente alto da coprire
addirittura le scarse cellule neoplastiche, in genere meningoteliali.
f) Microcistico: presenta un aspetto cribroide, dovuto alla presenza di
cellule con varie digitazioni (ricordano proprio l'aracnoide) ed un certo
grado di spongiosi cellulare.
g) Mixoide: ricco di matrice alcianofila e cellule vacuolate.
2) Meningiomi di grado II: mentre quelli di grado I sono facilmente diagnosticabili
al microscopio, questo gruppo di meningiomi vengono spesso confusi con altri
tipi di tumori. Nella recente revisione, tra le loro caratteristiche, è stata inserita
la capacità invasiva del tumore, nonostante in diversi casi non si presenti
questa proprietà.
a) Meningioma atipico: le caratteristiche di questi sono la presenza di un
indice mitotico più elevato ( 4 o più mitosi su 10 campi microscopici, a
40X) e almeno 3 delle seguenti caratteristiche (33):
i) crescita a tappeto
ii) necrosi

15
iii) elevata cellularità
iv) nucleolo prominente
v) cellule con elevato rapporto nucleo/citoplasma
Il termine "atipico" può essere fuorviante, dato che l'atipia cellulare non
è necessaria per la diagnosi e nemmeno così comune nei preparati,
mentre molto più significativo è l'indice mitotico. Per necrosi, si intende
un fenomeno spontaneo, non mediato da altri fattori come
embolizzazioni o radiazioni.
b) Meningioma cordoide: presenta cordoni o trabecole di cellule di aspetto
epiteliale eosinofiliche, uno stroma ricco di mucine, con vacuoli. In
genere, questa struttura è intervallata ad aree tumorali meningoteliali o
di transizione.
3) Meningiomi di grado III: sono considerati i tumori maligni, fortunatamente
molto più rari dei precendenti.
a) Meningioma anaplastico: presenta diverse caratteristiche, tipiche del
tumore maligno, come un elevato indice mitotico ( 20 o più mitosi per
10 HPF) o una completa anaplasia, cioè un elevato grado di
sdifferenziazione in cui risulta abbastanza complicato diagnosticare se si
tratti effettivamente di un meningioma o di un altro tumore, primario o
metastatico.
b) Meningioma papillare: variante della prima, caratterizzato da un
"pattern" di crescita papillare o pseudopapillare a livello perivascolare,
una parziale perdita di coesione cellulare e di strutture , simili alle
rosette presenti in alcune forme di ependimomi. Di solito sono presenti
porzioni di tumore che mantengono caratteristiche meningoteliali e si
assiste ad un marcato ispessimento della parete vascolare.
c) Meningioma Rabdoide: altra variante del primo, qui sono presenti
cordoni di cellule rabdoidi (ricordano dei bastoni), che posseggono
nucleo eccentrico e nucleolo prominente e un citoplasma eosinofilico
con spesso inclusioni globulari paranucleari, costituiti da fasci di

16
filamenti intermedi, avvolti su se stessi. Ovviamente sono spesso
presenti altre caratteristiche tipiche di un tumore maligno quali elevato
indice mitotico, atipia cellulare e necrosi. Come nel precedente, il
sottotipo rabdoide è associato ad incrementata mortalità e possibili
comparsa di metastasi extra-craniche.
Nonostante questa classificazione, stilata dalla WHO, sia destinata alla medicina
umana, viene utilizzata anche in veterinaria: infatti l'aspetto istopatologico di
queste forme tumorali è praticamente uguale a quelle dei nostri animali, mentre
cambiano le frequenze di distribuzioni dei vari tipi di tumore. Per esempio, da alcuni
studi (34), è emerso che la prevalenza del meningioma atipico(grado II) è molto più
elevata nel cane (40%) che nell'uomo ( 8%), mentre quella dei meningiomi benigni
(grado I) è minore ( 40-57% del cane contro l' 80% dell'uomo). Inoltre, sempre nel
cane, è stato osservato che può essere sufficiente un indice mitotico > 4 x HPF e
l'invasione del tessuto nervoso per diagnosticare un meningioma di grado grado II.
Nel gatto invece non sono stati riportati casi di meningiomi grado grado III ( tranne
un sospetto caso di meningioma papillare, localizzato al cervelletto) (35) ,
confermando l'ipotesi che i meningiomi felini abbiano un comportamento meno
aggressivo; per queste ragioni, il sistema di "grading" in questione, dovrebbe , nel
gatto, subire alcune modificazioni. Le metastasi sono rari reperti nei meningiomi di
cane ( prevalentemente a cuore e polmoni ) mentre è stato riportato solo uno nel
gatto (36) ( a livello dei reni e dell'utero ). Ci sono varie teorie che tentano di
spiegare la bassa prevalenza di metastasi di questi tumori. I meningiomi possiedono
diverse interdigitazioni e giunzioni intercellulari e questo potrebbe ridurre
l'esfoliazione di cellule tumorali e la loro diffusione nel torrente ematico;
inoltre ,dato che la maggior parte sono istologicamente benigni, le poche cellule che
si staccano dalla massa potrebbero non essere in grado di insiedarsi in altri tessuti e
colonizzarli (37).

17
Genetica dei meningiomi in veterinaria:
La correlazione che si osserva tra alcune razze di cani e meningiomi, ci suggerisce
della forte componente genetica nello sviluppo e nella progressione di questi
tumori. Nella specie canina sono stati fatti diversi studi, intenzionati a caratterizzare
la principali alterazioni genetiche correlate ai meningiomi, tra cui riporto uno del
2009, ad opera di Dickinson et al (38): questo lavoro si propone di verificare se
fosse presente una correlazione tra la perdita di alcuni oncosoppressori e
l'insorgenza di meningiomi. Con l'RT-PCR è stato ricercata la presenza del gene NF2,
e con Western blotting sono stati misurati i livelli d'espressione delle proteine,
tradotte da NF 2, 4.1B e TSLC1:
1) NF 2: oncosoppressore presente sull'autosoma 22 umano e responsabile
dell'espressione di una proteina, la Neurofibromina (o schwannomina o
merlina) , che possiede funzioni regolatorie sul ciclo cellulare. Individui che
presentano alterazioni a questo gene sono predisposte a sviluppo di
meningiomi e schwannomi multipli (39); la perdita d'espressione di NF 2 è stata
correlata al 30-80% di meningiomi umani, ed in particolar modo, è stato
osservato che questa modifica genetica è molto meno presente nei meningiomi
di tipo meningoteliale (40).
2) 4.1B: anche la proteina codificata dal suo gene (chiamato anche DAL, che , in
inglese, vuol dire "proteina differentemente espressa nel adenocarcinoma
polmonare) possiede proprietà regolatrici e una sua alterazione è stata
riscontrata nel 76% dei meningiomi umani (41).
3) TLSC: codifica per una proteina trans-membrana che interagisce con quella
precedente e una sua perdita d'espressione, nell'uomo, è stata riscontrata nel
30-85% del tumore in questione (42).
I meningiomi presi in esame erano 30, ed erano presenti sia di grado WHO I che e II,
e comprendevano forme sia intracraniche che spinali e ,da questi, sono emersi dati
interessanti: infatti non è venuta alla luce nessuna associazione tra l'espressione
delle proteine sopracitate e il grado del tumore, contrariamente a quanto avviene
nell'uomo; inoltre , sempre contravvenendo a quanto avviene in medicina umana,
non è stata osservata una minore associazione tra meningiomi meningoteliali e la

18
perdita d'espressione di NF 2. Dallo studio quindi emerge che la disfunzione di
oncosoppressori come 4.1B e TLSC1 sono presenti in alcuni meningiomi canini, e
che alterazioni genetiche di NF2 non seguono il "pattern" umano. Ciò nonostante,
secondo molti studiosi, il cane rappresenta un modello ottimale, dato che
avvengono spontaneamente e con un'incidenza simile (se non superiore) a quella
della popolazione umana (43). Uno studio del 2009 è volto proprio a verificare le
caratteristiche genetiche in comune tra tumori intracranici nell'uomo e nel cane
(11): tramite un'analisi dell'ibridazione genomica comparativa basata su micro-
array di DNA, sono stati studiati i profili citogenetici e individuare le alterazioni più
frequenti associate ai meningiomi . Ne è scaturito che le aneuploidie (alterazioni nel
numero di cromosomi) più comuni nei meningiomi erano una perdita di CFA
(cromosoma canino) 17 (66,7%,) e CFA 27 (40%), mentre l'acquisizione di
cromosomi era più rara, di cui, i più comuni erano CFA 19 e 26 ; purtroppo però non
è stata osservata un'associazione tra un aberrazione cromosomica specifica e un
grado o un istotipo di meningioma.
Caratterizzazione molecolare dei meningiomi:
Da quanto detto finora, I meningiomi sono delle forme tumorali, che, se pur ben
conosciute, possono risultare talvolta difficili da diagnosticare, anche con le
tecniche sopracitate. Per questo motivo, la metodica che permette con maggiore
sicurezza caratterizzare tale neoplasia è l' Immunoistochimica (IHC). Sono stati
eseguiti molti studi basandosi su IHC per verificare quali erano le proteine
maggiormente espresse in questo tipo di neoplasie. Nel 1990 fu dimostrata la
presenza di recettori per estrogeni e progesterone in meningiomi del cane e del
gatto (44) , ipotizzata da tempo dato che era già nota la presenza nei meningiomi
umani. E' stato osservato ,in più studi, che , in queste neoplasie, il riscontro di
recettori del progesterone (PR) è più elevato di quello per gli estrogeni (ER),
nonostante ci siano delle discrepanze tra vari studi sull'oggettiva prevalenza di
meningiomi PR positivi. Adamo , Cantile e Steinberg (45), nel loro studio, avevano
notato che l'immunopositività per il PR era presente in 14 su 15 casi di meningioma,
tra cane e gatto( di questi 15, 2 erano campioni di cervello sano), ed un valore

19
analogo emergeva da uno studio del Mandara (46); altri due autori, Theon e Ramos
Vara,hanno riscontrato ,nei loro studi (47), dei valori differenti, e cioè,
rispettivamente, 70% ( questo lavoro però prendeva in esame solamente
meningiomi di cane) e 44% dei casi totali. Opinione condivisa da tutti invece è la
rarissima espressione ,in questi tumori, di ER. Le ricerche su queste molecole, oltre
ad avere un interesse diagnostico non indifferente, possono avere anche dei risvolti
clinici interessanti : infatti è plausibile che trattamenti anti-progestinici possano
costituire un trattamento, soprattutto in quei tumori inoperabili o ricorrenti che
esprimono questi recettori. Uno degli studi più recenti riguardante l'analisi dei
marker tumorali specifici per i meningiomi risale a 3 anni fa ed è stato condotto da
Ramos Vara e colleghi (48): nel loro articolo, hanno valutato con quale percentuale,
nuove molecole si presentassero in queste neoplasie mediante IHC, confrontandole
con i marker tradizionali. Nei meningiomi umani, l'antigene ritrovato nella
stragrande maggioranza, tra il 75 e il 100% dei meningiomi (49) e considerato il
marker più affidabile di queste neoplasie, è l' EMA ( "Epithelial Membrane Antigen);
sfortunatamente gli anticorpi anti -EMA non cross-reagiscono con i tessuti felini o
canini (50). Le molecole ricercate nello studio con l'IHC sono:
Citocheratine: numeroso gruppo di molecole prodotto dalle cellule epiteliali.
Costituiscono il marker aspecifico ma caratteristico per quasi tutti i tumori di
natura epiteliale
Vimentina : Presente praticamente in tutte le neoplasie di origine
mesenchimale.
GFAP ( "Glial Fibrillar Acidic Protein" ): Proteina fibrillare prodotta in alcune
cellule del sistema nervoso tra cui astrociti e cellule ependimali (51).
Ki67: proteina situata a livello nucleare nell'interfase, mentre nelle altre fasi del
ciclo cellulare si trova sulla superficie dei cromosomi. Il fatto di essere presente
in tutte le fasi attive di replicazione cellulare (G1 , S, G2 e mitosi) ed assente nel
G0, la rende un ottimo marker per vedere se le cellule sono in fase di
replicazione oppure no. La sua identificazione viene usata per verificare la

20
correlazione tra l'indice di proliferazione e il comportamento biologico e clinico
di alcune neoplasie (52).
S 100: sotto questo nome sono comprese un grande numero di proteine che
condividono la stessa struttura molecolare di base, che possiedono funzioni
fisiologiche tra le più disparate e rivestono un ruolo in malattie come
cardiomiopatie, disordini neurologici e tumori (53).
Enolasi specifica neuronale: molecola che deve il suo nome al fatto di essere
stata scoperta inizialmente solo nelle cellule nervose e neuroendocrine. In
realtà questo enzima è espresso anche in altre cellule normali ed in alcuni tipi di
tumore.
PCNA(Proliferating Cellular Nuclear Antigen): Proteina responsabile della
polimerasi del DNA, possiede praticamente le stesse caratteristiche della
proteina Ki67.
Sinaptofisina: è una proteina presente nelle cellule neuroendocrine e in tutte le
cellule che posseggono sinapsi; essendo specifica dalle cellule nervose , è un
marker ottimale per i tumori che sorgono da questi tessuti (54) . Il meningioma
non esprime questo marker.
H - telomerasi: è un complesso ribonucleico che previene accorciamenti delle
estremità cromosomiche ( chiamate telomeri) nelle cellule eucariote.Questi
enzimi sintetizzano a livello telomerico sequenze ricche di guanosina e proteine
associate. La presenza di questi enzimi in un tumore rendono le sue cellule
capaci di un'attività replicativa maggiore e quindi indice di maggiore malignità.
Recettori per gli estrogeni e progesterone.
Questi costituiscono comuni marker ricercati nei meningiomi delle specie
domestiche (55) (56) . Lo studio di Ramos Vara si prefigge di valutare la presenza di
nuovi marker nei meningiomi, sia canini che felini. Quelli considerati nello studio
sono :
CD 34: chiamato anche sialomucina, è una glicoproteina espressa nelle cellule
endoteliali , e funge da ligando per le integrine.

21
E caderine e Claudina 1: entrambe sono proteine che compongono alcune
giunzioni intercellulari. L' assenza di espressione di queste proteine è un segno
spesso associato a malignità (57)
GLUT 1( proteine trasportatrice di glucosio 1) :è una proteina di membrana
fortemente espressa a livello di cellule endoteliali che regola il transito oltre il
vaso di glucosio (58). La molecola in questione è stata riscontrata in molti casi
di meningioma giovanile umano (59).
Laminine: sono una vasta famiglia di proteine che ,insieme ad altre,
compongono le lamine basali di tutti quanti gli organi e prendono parte a
diversi processi , tra cui differenziazione, migrazione ed adesione cellulare (60) ;
sembra che queste svolgano un ruolo nella carcinogenesi (61).
PGP 9,5 ( Protein Gene Product): proteina che si presenta in elevate
concentrazioni, circa 50 volte che negli altri organi nelle cellule nervose
encefaliche; costituisce un marker per tutte le malattie che si manifestano in
queste cellule (62).
Tutti questi studi di IHC sono state eseguite prima su meningi sane, per verificare
l'espressione di queste molecole in assenza di tumore. I risultati ottenuti , molto
simili tra cane e gatto, sono i seguenti: ( in parentesi sono indicate le regioni in cui si
riscontra immunopositività) :
o Fortemente espressi : S-100 e PGP 9,5 ( leptomeninge) ;
CD 34 ( lo strato più superficiale della leptomeninge,alcune cellule sparse negli
strati più profondi e le cellule endoteliali) ; E caderine ( anche loro nello strato
più esterno mentre in quelli interni una presentano un moderata positività);
laminina ( soprattutto gli strati esterni )
o Leggermente espressi: Claudina - l ( solo in alcune cellule più interne) ; GLUT 1
( moderata positività citoplasmatica granulare in alcune cellule nella membrana
più esterna ed in scarse cellule più interne ) ; vimentina ( dappertutto) ;
o Non espressi: Citocheratine ; GFAP.

22
I risultati provenienti invece dai meningiomi, con le relative frequenze sono i
seguenti :
Ramos-Vara , a partire da questi dati, propone come marker per i meningiomi canini
e felini vimentina, CD 34 ed E-caderina, a cui è possibile associare la claudina-l, che ,
pur avendo una bassa - moderata specificità per questa tipologia di tumore,
possiede elevata specificità; questa proteina ,inoltre , in medicina umana, permette
di differenziare meningiomi dagli schwannomi (63), elemento che sarebbe
opportuno indagare anche in veterinaria. Dallo studio infine emerge che manca una
correllazione tra la positività al Ki67 e l'indice mitotico : questo, potrebbe derivare
dal fatto che ci siano delle differenze nelle fasi di moltiplicazione delle cellule
tumorali oppure che ci siano delle difficoltà nel distinguere cellule in mitosi con
nuclei picnotici (64).
Altri due recenti studi italiani sono orientati a caratterizzare i tumori in questione,
con lo scopo di trovare dei marker prognostici relativi ai meningiomi e/o possibili
bersagli farmacologici .Il primo risale al 2007 e prende in considerazione le già citate
telomerasi (65): l'immunopositività è stata osservata prevalentemente a livello
nucleare e nucleolare ( anche con rari reperti citoplasmatici) mentre non è stata
riscontrata nessuna correlazione tra la positività e istotipo,sesso o età, esclusa una
0% 20% 40% 60% 80% 100%
GFAP
Citocheratine
PR
PGP 9.5
claudina-l
Laminina
S 100
GLUT-1
CD 34
Vimentina
9%
39%
44%
55%
60%
72%
75%
86%
94%
100%
Risultati dello studio di Ramos-Vara
Positività nei meningiomi campionati

23
maggiore espressione nei meningiomi felini che in quelli canini. Questi dati sono
stati messi a confronto con indici di replicazione cellulare, ossia con il Ki67
( precedentemente già menzionato) e il conteggio delle mitosi con una colorazione
con blu di toluidina. L'IHC del Ki67 non ha dato nessun risultato positivo, forse a
causa del prolungato tempo di stoccaggio nei blocchetti di paraffina o in formalina,
che ha portato a modificazioni subcellulari tali da impedire il legame tra anticorpo e
proteina in questione. il conteggio mitotico eseguito con la colorazione con blu di
toluidina permette di distinguere molto bene le cellule in mitosi (66); nello studio è
stato riscontrato un basso indice mitotico nell' 84 % delle neoplasie totali e questo
dato non è correlato con l'immunopositività alla h-telomerasi. Una possibile
spiegazione per questo elemento forse risiede nel fatto che l'attivazione della
telomerasi ,un parametro che comunque rientra nelle caratteristiche dei tumori
maligni , possa avvenire indipendentemente da una incrementata proliferazione.
L'espressione del suddetto enzima infatti conferisce alle cellule una capacità infinita
di replicazione (67). Questi dati fanno presupporre che la telomerasi possa rivestire
un ruolo nella prognosi del tumore, indipendente da parametri cinetici e che abbia
un valore diverso tra gatto e cane, data la diversa prevalenza riscontrata.
L'altro studio invece si occupa dell'espressione nel tumore delle metalloproteasi di
matrice (MMP) . Queste fanno parte di una famiglia di più di 9 endoproteasi, capaci
di degradare macromolecole della matrice extracellulare del tessuto connettivo .
Possiedono una struttura abbastanza omologa, con alcune piccole differenze che si
riflettono su una maggiore specificità per alcuni substrati: Le MMP 2 e 9 per
esempio vengono chiamate rispettivamente gelatinasi A e B perchè degradano fibre
collagene già parzialmente degradate (gelatina). La malignità di un tumore è stata
da molto tempo correlata alle MMP (68) : infatti questi enzimi , degradando le
membrane basali e le proteine extracellulari, aprono dei passaggi per le cellule
tumorali e ampliano di fatto le capacità invasive della neoplasia. Le più importanti,
in questo ruolo, sono proprio le gelatinasi A e B, che sono quelle che degradano
meglio il collagene IV, costituente più diffuso nelle membrane basali di tutti i tessuti.
Inoltre è emerso da diversi studi che le MMP rivestono un determinati ruolo
24
nell'angiogenesi e nell'iniziazione del tumore (69). Proprio sulla scia di studi del
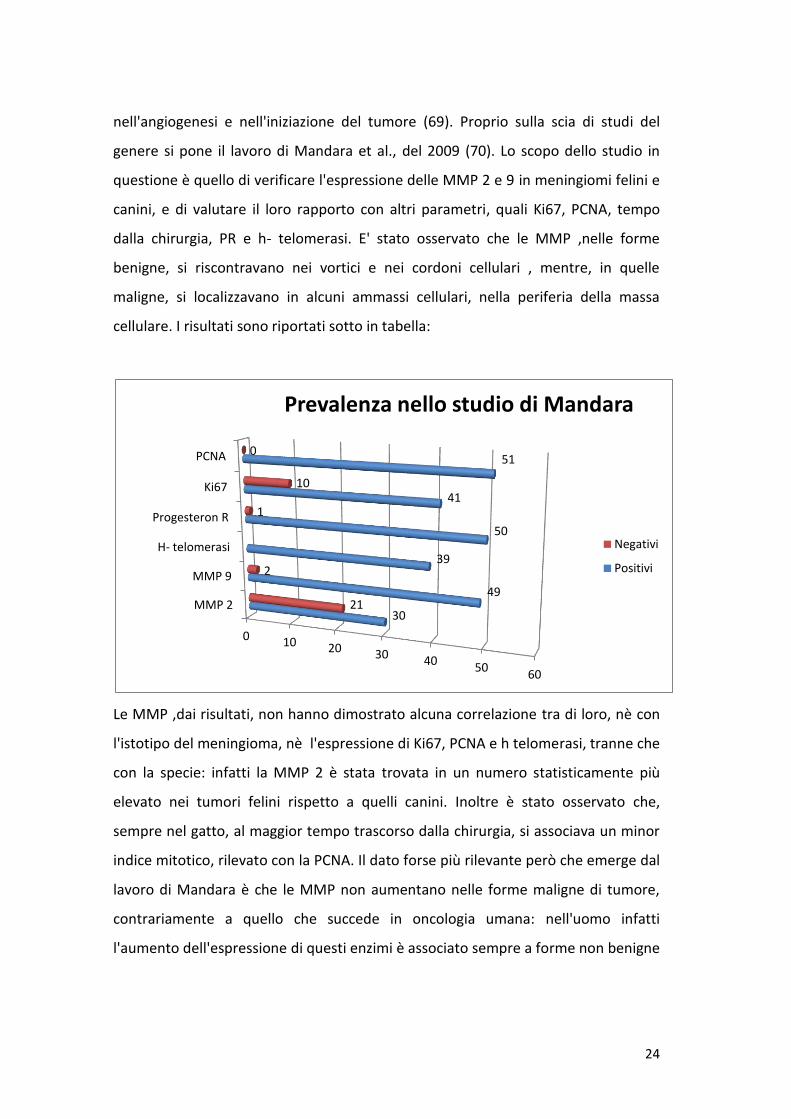
genere si pone il lavoro di Mandara et al., del 2009 (70). Lo scopo dello studio in
questione è quello di verificare l'espressione delle MMP 2 e 9 in meningiomi felini e
canini, e di valutare il loro rapporto con altri parametri, quali Ki67, PCNA, tempo
dalla chirurgia, PR e h- telomerasi. E' stato osservato che le MMP ,nelle forme
benigne, si riscontravano nei vortici e nei cordoni cellulari , mentre, in quelle
maligne, si localizzavano in alcuni ammassi cellulari, nella periferia della massa
cellulare. I risultati sono riportati sotto in tabella:
Le MMP ,dai risultati, non hanno dimostrato alcuna correlazione tra di loro, nè con
l'istotipo del meningioma, nè l'espressione di Ki67, PCNA e h telomerasi, tranne che
con la specie: infatti la MMP 2 è stata trovata in un numero statisticamente più
elevato nei tumori felini rispetto a quelli canini. Inoltre è stato osservato che,
sempre nel gatto, al maggior tempo trascorso dalla chirurgia, si associava un minor
indice mitotico, rilevato con la PCNA. Il dato forse più rilevante però che emerge dal
lavoro di Mandara è che le MMP non aumentano nelle forme maligne di tumore,
contrariamente a quello che succede in oncologia umana: nell'uomo infatti
l'aumento dell'espressione di questi enzimi è associato sempre a forme non benigne
0 10 20 30 40 50
60
MMP 2
MMP 9
H- telomerasi
Progesteron R
Ki67
PCNA
30
49
39
50
41
51
21
2
1
10
0
Prevalenza nello studio di Mandara
Negativi
Positivi

25
di tumore (71), anche se ci sono delle incongruenze ,in letteratura, tra la presenza
delle MMP e il sistema di "grading" WHO.
Marker linfocitari :
Da quanto emerso finora quindi, in veterinaria, mentre ci sono diversi marker che
permettono di distinguere se un tumore sia un meningioma o meno, poche sono le
molecole che ci danno informazioni sulla prognosi della neoplasia e, proprio in
questo ambito, si inserisce il mio lavoro. La ricerca del sottoscritto è volta infatti
alla caratterizzazione dell'infiltrato infiammatorio,soprattutto nei confronti delle
sottopopolazioni linfocitiche. I marker di membrana utilizzati per la
caratterizzazione dell'infiltrato infiammatorio sono stati il CD 3,CD 79a,CD 20 e
FoxP3:
CD 3: chiamato anche TCR (Recettore cellulare T), riconosce gli antigeni e
traduce questo in segnali intracellulari che determinano l'attivazione del
linfocita T (72).
CD 79: Eterodimero composto dalla subunità CD79a e CD79b presente sulla
membrana dei linfociti B, costituisce il BCR (Recettore Cellulare B) insieme a
immunoglobuline di membrana, come le IgD o IgM (73).
CD 20: è un antigene di membrana che consiste di 4 regioni transmembrana ed
un dominio citoplasmatico che contiene più diversi siti ad attività catalitica.
Viene espresso principalmente sui linfociti B, ma è stato individuato anche su
linfociti T normali e neoplastici (74).
FoxP3:
Generalità:
questa fa parte di una vasta famiglia di proteine, chiamate Forkhead box, che sono
coinvolte in un ampia varietà di processi biologici, agendo da fattori di trascrizione:
con questo termine si indicano molecole che ,legandosi in siti specifici del DNA ,
favoriscono o inibiscono la trascrizione e, di conseguenza, modulano l'espressione
di proteine. Il nome di questa famiglia deriva dalle strutture a due teste appuntite,

26
simili a una forchetta, ritrovate negli embrioni di Drosophila fork head , un
mutante che mancava di intestino anteriore e posteriore; al suo interno è stata
scoperta la Fox, con dominio di 110 amminoacidi capace di legarsi al DNA,
altamente conservato in tutte le molecole della famiglia (75). Questo è costituito da
un cuore di 3 α-eliche , affiancato da due anse ,chiamate anche "ali" ( infatti ,nella
letteratura anglosassone, sono conosciute come "winged helix protein") ; sono
capaci di legarsi al DNA tramite la loro caratteristica struttura a "elica-ripiegamento-
elica" delle 3 catene α, permettendogli di regolare la trascrizione con questo
legame. L'attuale nomenclatura prevede che con Fox, si indichi la famiglia, con una
lettera ( da A a Q ) la sotto famiglia e con un numero arabo il gene/ la proteina (il
gene si distingue dalla proteina perchè scritto in corsivo) in particolare .La FoxP3
viene chiamata anche scurfina perchè venne scoperta in una linea di topi mutanti
chiamata scurfy, in cui gli esemplari venivano a morte per un iperproliferazione di T
helper e massiva produzione di citochine infiammatorie (76), simile alla IPEX
(immunodisfunzioni, poliendocrinopatia, enteropatia relativa al cromosoma X) (77).
E' sicuramente la molecola più studiata all'interno della famiglia , soprattutto per il
suo ruolo in immunologia. Viene fortemente espressa in una classe di linfociti
chiamati T regolatori ( Treg ) caratterizzati dalla positività al CD4 CD25 e alla FoxP3:
la presenza del CD 4 è tipica anche dei linfociti T helper ( essendo uno specifico
recettore per gli MHC di classe II, a cui si lega insieme al TCR ), mentre il CD 25 è la
catena α del recettore per l'IL 2, presenta anche su linfociti attivati,monociti e
linfociti B. Pur essendo state scoperte in tempi relativamente recenti, costituiscono
un importante classe linfocitica, che riveste un ruolo essenziale nell'immuno-
omeostasi della risposta immunitaria (78). Da molti studi effettuati sull'uomo e sul
topo , queste cellule hanno marcate capacità immunosoppressive, conferite proprio
dalla FoxP3, e sono divisibili principalmente in cellule regolatorie T naturali e
indotte.
Natural T Regulatory lymphocytes (nTreg) (79):
Sono prodotti nel timo, preferenzialmente ma non esclusivamente a livello della
midollare, dove avviene la selezione negativa dei linfociti T (80). Successivamente lo

27
abbandonano per costituire circa il 5% di tutti i linfociti CD4+ presenti nella periferia
dell'organismo. Esprimono CD4, alti livelli di CD25 e anche FoxP3 e sono essenziali
per la prevenzione di fenomeni di autoimmunità e per mantenere sotto controllo il
sistema immunitario. Sono state individuate diverse molecole chiave per lo sviluppo
e il mantenimento delle nTreg:
TCR ( CD3 ): all'unanimità è stato accettato che la stimolazione di questa
molecola, presente su tutti i vari tipi di linfociti, è essenziale nella formazione
delle Treg; diversa è la situazione riguardo a quali antigeni siano implicati in
questa presentazione, per cui sono stati proposti modelli differenti. Quello
attualmente più accreditato afferma che i nTreg derivino da timociti con un TCR
che abbia un'affinità verso self-antigeni più elevata di quella richiesta per una la
selezione positiva, ma generalmente più bassa per subire una selezione
negativa (81).
CD 28: molecola presente anch'essa sulla membrana della stragrande
maggioranza di linfociti, e capace di legarsi ad una coppia di proteine di
membrana presente sulle APC,la B7-1 (o CD 80) e la B7-2 (o CD 86). Questo
legame, come quello dell'antigene con il TCR, risulta essenziale per lo sviluppo
del linfocita e, per questo motivo, suddetto passaggio viene chiamato
costimolazione e le molecole che rientrano in questo processo, che possono
essere diverse, costimolatorie . In realtà questo doppio legame è vitale non solo
per i nTreg ma anche per l'attivazione di tutti quanti i linfociti (82). Da alcuni
esperimenti, è stato osservato che topi mancanti di CD 28 e B7 presentavano
una riduzione del numero di nTreg nel timo e nei tessuti periferici; questo fatto
fu inizialmente interpretato come se difetti a livello della costimolazione
abbassassero i livelli di IL-2, un'altra molecola fondamentale per i nTreg.
Questa prima tesi però fu smentita grazie ad uno studio, in cui precursori che
mancavano di CD 28, non riuscivano a differenziarsi in nTreg ,nonostante fosse
reso disponibile ai linfociti T IL-2 (83). Questo lavoro ha dimostrato come la
costimolazione, mediata dal CD 28 linfocitico e il B7 dell'APC, risulti di primaria
importanza per queste cellule.
28
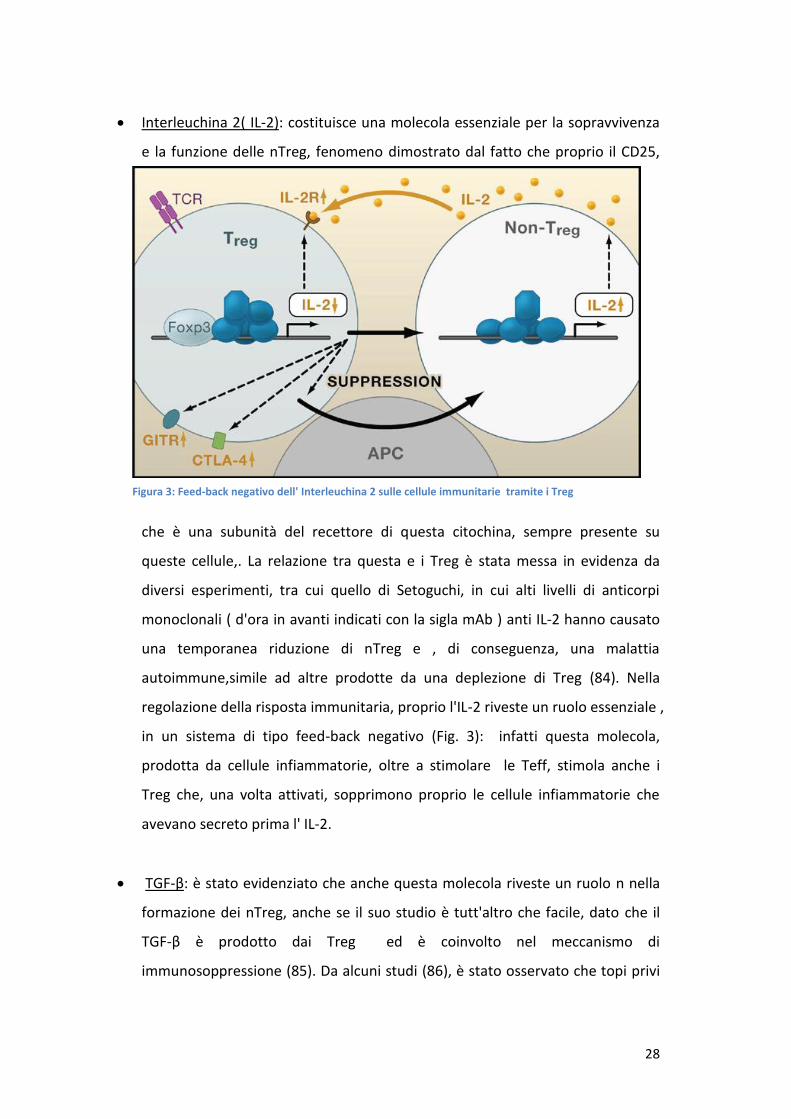
Interleuchina 2( IL-2): costituisce una molecola essenziale per la sopravvivenza
e la funzione delle nTreg, fenomeno dimostrato dal fatto che proprio il CD25,
che è una subunità del recettore di questa citochina, sempre presente su
queste cellule,. La relazione tra questa e i Treg è stata messa in evidenza da
diversi esperimenti, tra cui quello di Setoguchi, in cui alti livelli di anticorpi
monoclonali ( d'ora in avanti indicati con la sigla mAb ) anti IL-2 hanno causato
una temporanea riduzione di nTreg e , di conseguenza, una malattia
autoimmune,simile ad altre prodotte da una deplezione di Treg (84). Nella
regolazione della risposta immunitaria, proprio l'IL-2 riveste un ruolo essenziale ,
in un sistema di tipo feed-back negativo (Fig. 3): infatti questa molecola,
prodotta da cellule infiammatorie, oltre a stimolare le Teff, stimola anche i
Treg che, una volta attivati, sopprimono proprio le cellule infiammatorie che
avevano secreto prima l' IL-2.
TGF-β: è stato evidenziato che anche questa molecola riveste un ruolo n nella
formazione dei nTreg, anche se il suo studio è tutt'altro che facile, dato che il
TGF-β è prodotto dai Treg ed è coinvolto nel meccanismo di
immunosoppressione (85). Da alcuni studi (86), è stato osservato che topi privi
Figura 3: Feed-back negativo dell' Interleuchina 2 sulle cellule immunitarie tramite i Treg

29
di TGF-β avevano, nei primi giorni di vita, una ridotta quantità di nTreg, mentre,
nei giorni successivi il numero di queste cellule tornava normale. Da questo
dato si è dedotto che la molecola in questione è implicata nella iniziale
proliferazione di nTreg, ma non costituisce affatto un elemento essenziale nella
loro genesi.
Induced T regulatory lymphocytes (iTreg) (81) : sono Treg che condividono praticamente le stesse caratteristiche antigeniche dei
nTreg (FoxP3+ ; CD 4+ ; CD 25+ ), ma ,diversamente da loro, si formano da linfociti T
"vergini" ( o naive), dimostrando una maggiore versatilità rispetto ai primi. Le cellule
iTreg sono attualmente oggetto di studi approfonditi, dato che si crede siano
coinvolte nella tolleranza agli antigeni alimentari e dei batteri commensali e per il
fatto che sono le Treg maggiormente riscontrate a livello di trapianti, tessuti in
preda a infiammazioni croniche (87) e microambienti tumorali. Sono diversi i
meccanismi e le molecole chiave implicate nel fenomeno della loro induzione e
sono affrontate nell'elenco sottostante :
TCR : tutte le APC ( cellule che presentano l'antigene) sono in grado di indurle ,
insieme a linfociti B, cellule staminali mesenchimali e cellule soppressorie di
derivazione mieloide ( MDSC). E' stato osservato che una bassa dose di antigeni
favorisce una trasformazione in Treg mentre una elevata porta alla conversione
in cellule T effettrici ( Teff ) (88). Inoltre emerge da uno studio che una piccola
concentrazione di un buon agonista TCR induce molto meglio la formazione di
iTreg rispetto ad un'alta concentrazione di uno debole. Da ciò si desume che il
destino di un linfocita naive dipenda dalla quantità di TCR impegnati nel legame,
e cioè se elevata si convertirà in un Teff, se bassa in un iTreg.
TGF-β: Il suo ruolo nell'induzione dei iTreg in vivo e in vitro è confermata da
molti lavori a tal riguardo (89), e, addirittura, è in grado di trasformare T naive
in iTreg in presenza di una elevata quantità di TCR stimolata,contrariamente a
quanto avviene in sua assenza (90). Ciò nonostante, sono state individuate delle
cellule precursori CD4+CD25+FoxP3- che, dopo aver lasciato il timo, si sono
trasformate in iTreg dopo l'incontro con self-antigeni e il legame con l'IL2,

30
dimostrando un modello d'induzione indipendente dal TGF-β. La presenza di
suddette cellule meriterebbe ulteriori approfondimenti (81).
CTLA 4 ( CD 152): ha una struttura omologa al CD 28, menzionato prima, ed è
infatti capace di legarsi alle molecole B7-1 e B7-2 presenti sulle APC, rivestendo
un ruolo però diverso nella costimolazione dei Treg e delle Teff. Infatti la
stimolazione di questo recettore o del CD 28 ha effetti diversi in base alla
cellula T in esame. Per le Teff il legame con il CD 28 è essenziale per
l'attivazione e la proliferazione dei linfociti effettori attivati, mentre il legame
con CTLA 4 ha un effetto inibitorio (82). i nTreg possiedono un comportamento
analogo per quanto riguarda il CD 28 (vedi sopra ), mentre le iTreg possiedono
proprio un rapporto inverso con queste molecole costimolatorie: infatti il
legame tra CTLA 4 e B7 costituisce un processo fondamentale nell'induzione dei
iTreg da parte del TGF-β, mentre la costimolazione B7-CD28 sembra essere
importante solo per la produzione di IL-2 e, ad alti livelli , esercita addirittura
l'inibizione, nell'induzione TGF-β dipendente. Questo è stato dimostrato in un
esperimento (91), in cui topi che avevano problemi nel meccanismo di
costimolazione CD28-B7 sviluppavano un maggior numero di iTreg. Questo
complesso ruolo della costimolazione nella fisiologia linfocitica è schematizzata
nella figura seguente (Fig.4)
Figura 4: Schematizzazione della costimolazione nelle diverse sottopopolazioni di linfociti T
Costimolazione CD28/B7
Essenziale nelle Teff
Essenziale nelle nTreg
Ad alti livelli,
inibisce le iTreg
Costimolazione CTLA4/B7
Inibisce le Teff
Essenziale nelle nTreg
Essenziale nelle iTreg

31
IL 2: non è ancora chiaro il ruolo che questa molecola rivesta nell'induzione dei
Treg in vivo ma sicuramente è essenziale nell'induzione TGF-β dipendente (92).
Acido Retinoico e batteri commensali: sembra che sia l'acido retinoico ( o più
precisamente un suo metabolita, ossia "all-trans retinoic acid" o ATRA ) sia i
batteri commensali enterici regolino il fenomeno della tolleranza immunitaria
a livello intestinale, agendo proprio sulla formazione dei iTreg: infatti da diversi
studi è emerso che la mucosa enterica esprime alti livelli di ATRA , che potenzia
la conversione dei T naive e determina una infiltrazione intestinale, anche in
presenza di forti segnali di costimolazione CD28-B7 (93). Inoltre alcune cellule
dendritiche CD 103+ presenti proprio nella lamina basale ,soprattutto del
piccolo intestino, aumentano la conversione in iTreg , secernendo ATRA e TGF-β
(94) ; proprio queste ultime 2 molecole ,secondo recenti studi,
condividono ,parzialmente, il meccanismo con cui inducono la conversione in
iTreg dei linfociti T naive . I batteri della microflora intestinale invece hanno
sui Treg un impatto variabile , che varia in base alla loro complessa
composizione : per esempio, il trattamento con la vancomicina(un antibiotico )
di alcuni topi, aumenta la quantità di Treg nella lamina propria mentre abbassa
quella dei linfociti T helper (95). La proliferazione di iTreg è stata osservata
anche in altri esperimenti, come quello svolto da Atarashi , in cui ha iniettato
via os alcune specie di Clostridium in topi neonati (96); un risultato analogo è
stato ottenuto attraverso la somministrazione del polisaccaride A , prodotto da
Bacteroides fragilis, in cui la moltiplicazione avveniva a livello intestinale, e più
precisamente nel GALT ( tessuto linfoide associato all'intestino ) (97). E' stato
osservato invece che la somministrazione di DNA di batteri della microflora
nell'intestino viene riconosciuto dal TLR 9 e porta alla produzione di
interleuchine pro infiammatorie che inibiscono la conversione dei iTreg (98). Da
questa serie di esperimenti, si capisce come la flora batterica, perennemente
presente nell'intestino, metta in comunicazione l'immunità innata ed acquisita
e costituisca un elemento chiave nella regolazione delle risposte immunitarie
locali, esplicando azioni positive o negative proprio sui nTreg e iTreg (81).

32
Alcuni farmaci possono infine influenzare l'induzione delle Treg ,come per
esempio la rapamicina: questo farmaco immunodepressore inibisce i linfociti T
effettori, bloccando la proteina chinasica B ( conosciuta come PKB o Akt, vedi
più avanti), senza impedire la moltiplicazione dei nTreg e stimolando allo stesso
tempo la trasformazione dei T naive in iTreg, con un meccanismo sinergico con
quello del TGF-β (99). La ciclosporina invece, pur essendo sempre un farmaco
immunosoppressore, inibisce la formazione dei iTreg e delle Teff sia in vitro
che in vivo (100).
Inoltre, da alcuni studi, è emerso che anche le nTreg hanno un ruolo in questo
processo di induzione: infatti loro possono convertire cellule T naive in iTreg, che, a
loro volta, vanno ad inibire l'attività dei T helper vicini, con una regolazione
omeostatica tale da mantenere un equilibrio bilanciato che i Treg e le Teff. Kendal
et al. (101) hanno dimostrato però che la presenza di nTreg è essenziale per la
continua immunosoppressione locale, mentre i iTreg possono mantenerla,
convertendo i linfociti T naive locali in iTreg. E' stata osservata anche la formazione
di iTreg in sedi di trapianti , dove assumono un ruolo cruciale le cellule dendritiche
plasmocitoidi (102). Ma quali sono i meccanismi con cui questi mediatori chimici
inducono questa trasformazione a livello dei linfociti T naive? La risposta a questa
domanda risiede nell'espressione della proteina chiave delle Treg, ossia la FoxP3.
Questo processo avviene grazie alla trascrizione del suo gene Foxp3 , che comincia
quando la RNA polimerasi si lega alla promotore, ed è regolata anche da alcune
sequenze non codificanti conservate (CNS) : per l'esattezza sono 3 le CNS nel locus
Foxp3 che modulano, tramite varie interazioni con altre molecole, l'espressione
della proteina in questione e, più precisamente :
1. CNS 1: sembra non essere un elemento di vitale importanza nella
differenziazione dei Treg ,anche se possiede un ruolo prominente
nell'induzione dei iTreg nel GALT
2. CNS 2: è coinvolto nell'espressione di Foxp3 nelle cellule figlie di Treg in
divisione e costituisce un elemento indispensabile per la stabilità d'espressione
della proteina in questione in queste

33
3. CNS 3: è responsabile di un imponente aumento della frequenza di Treg
generati sia nel timo che a livello periferico (103).
Vediamo adesso più in dettaglio come viene regolata l'espressione di questo
essenziale fattore di trascrizione nei linfociti:
Sistema di attivazione TCR-dipendente: la stimolazione antigenica mediata dalle
APC e la costimolazione porta all'attivazione intracellulare di alcuni fattori di
trascrizione come NF-kB ( fattore nucleare kB), NFAT (Fattore nucleare delle
cellule T attivate ) e AP1 ( Proteina di attivazione 1).
o NF-kB : dopo la stimolazione antigenica, per la produzione di questo fattore di
trascrizione, è essenziale un complesso di molecole intracellullari ossia il
complesso CBM (CARMA1 / Bcl 10 / Malt1) , come evidenziato da alcuni
esperimenti (104); inoltre è stato osservato che questo complesso è a monte
del NF-kB per il fatto che topi, con difetti sull'elemento CARMA1,
sviluppavano normali Treg , in presenza di una forma attiva di NF-kB (105) .
NF-kB agisce sull'espressione della FoxP3 grazie ad un'altra molecola, la c-rel,
che si lega direttamente al promotore del FoxP3 e al suo CNS 3.
o NFAT e AP1: anche questi fattori di trascrizione, prodotti dalla stimolazione
antigenica del TCR, si legano al promotore del FoxP3, favorendo così la sua
espressione.
o Questi fattori di trascrizione non sono esclusivi dei Treg, ma sono presenti
anche nella regolazione dell'espressione di tutti i linfociti ed un difetto in un
"pathway" comune ad altri linfociti si ripercuote sull'intero sistema
immunitario. Un esperimento che dimostra questo è quello svolto nel 2010 da
Fu G. et al. (106), in cui, una mutazione in un enzima coinvolto nel
segnalamento del TCR, la PLCγ1 ( fosfolipasi C gamma 1 ) , si ripercuote sulla
normale fisiologia sia dei Treg e delle Teff.
Complesso IL 2 - STAT5: L'interazione tra l'IL 2 ed il suo recettore è alla base di
una vasta serie di meccanismi intracellulari, tra cui quello della MAPK ( MAP
chinasi ) o del PI3K ( fosfoinositide 3 chinasi), ma il più importante ,nel sistema
immunitario, è sicuramente quello dello STAT5 ( o trasduttore del segnale ed

34
attivatore della trascrizione 5 ): questo infatti va a legarsi al promotore e al
CNS2 del FoxP3, favorendo la sua espressione in una maniera analoga ai fattori
di trascrizione sopracitati.
Asse IP3K - Akt - mTOR: queste 3 molecole e cioè ,rispettivamente IP3K,
proteina chinasica B (Akt) e il bersaglio della rapamicina nei mammiferi (mTOR)
sono presenti in tutte le cellule linfocitiche e vengono attivate anch'esse dal
TCR, CD 28 e dall'IL2. In questo meccanismo vediamo però come le Treg si
comportino in maniera differente dai Teff convenzionali: in quest'ultime infatti
l'attivazione di quest'asse porta non solo ad attivazione delle Teff, ma anche ad
un blocco dello sviluppo delle Treg,fenomeno osservato in diversi esperimenti
(107). La ragione di tutto questo risiede nel fatto che una bassa concentrazione
di queste proteine mantiene elevato il numero di 2 fattori di trascrizione, la
Foxo1 e la Foxo3: quest'ultime molecole, appartenenti alla stessa famiglia della
FoxP3, si legano al promotore e alla CNS 2 e facilitano l'espressione di
quest'ultima proteina (108).
Complesso TGF-β - Smad e Acido retinoico: Nonostante non sia stato ancora
compreso a pieno il ruolo che il TGF-β abbia nella formazione delle Treg in vivo,
è chiaro che questa molecola faccia parte del complesso meccanismo all'origine
delle iTreg e non solo,come vedremo più avanti. Il legame con il suo recettore
determina la produzione di una serie di fattori di trascrizione, le Smad (XXX) ed
è proprio la Smad 3 che va a legarsi al CNS 1 del FoxP3 ,facilitando l'espressione
del gene in questione (109); a ciò si aggiunge il fatto che il TGF-β esercita
un'azione inibitoria su un fattore di repressione, il Dnmt 1 (110). Ciò nonostante,
è stato visto che un blocco delle Smad solo parzialmente riduce l'induzione in
vitro delle iTreg, e questo fenomeno indica che l'espressione del FoxP3 dipende
anche da altri fattori TGF-β indipendenti, come ERK e JNK/MAPK, il cui ruolo in
queste cellule però non è stato ancora studiato a fondo (111).
Il meccanismo con cui ATRA agisce, tramite il legame con il suo recettore, è
legato al suo effetto inibitore su un altro fattore di trascrizione, il GATA-3:
questa molecola, che riveste un importante ruolo nella normale fisiologia dei

35
Th2 (82), sembra infatti inibire la trascrizione del FoxP3, mediante il legame col
promotore di questo gene (112). Con il complesso TGF-β, ATRA condivide
proprio il bersaglio molecolare, ossia il GATA-3: infatti le Smad 3 e 4 inducono
l'espressione di una importante proteina, ossia la Id3, che promuove
l'induzione delle Treg mediante un doppio meccanismo. In primo luogo ,
sinergicamente con ATRA, blocca il GATA-3, impedendo il suo legame con il
promotore del FoxP3 ,lasciandolo sgombro per la fase successivo; qua, un altro
fattore di trascrizione, E2A , trova spazio per legarsi a questo segmento di DNA
e, tramite il suo legame, facilita la espressione di FoxP3. Ciò nonostante, il TGF-
β non sembra in grado di convertire, da solo, T naive in Treg senza la presenza
di IL 2 e la stimolazione antigenica/ costimolazione. Tutti questi complessi
meccanismi, alcuni dei quali sono tuttora ancora oggetti di studio, sono
riassunti nell'illustrazione seguente (Fig. 5)
Figura 5: Principali "pathway" molecolari coinvolti nella regolazione dell'espressione di FoxP3

36
Quanto detto finora dà un'idea di come le molecole finora descritte costituiscano
degli elementi essenziali nella differenziazione dei linfociti T naive in Treg, anche se
una di queste è in realtà "fa il doppio gioco": con questo termine voglio indicare che
la" molecola incriminata", ossia il TGF-β, è coinvolto non solo nella conversione del
linfocita T naive in Treg ma anche in Th17,una linea di linfociti helper scoperta
recentemente che esula dalla accademica divisione in Th1 e Th2 e che presenta un
rapporto molto complesso con i Treg. Queste cellule sono in grado di secernere
diversi tipi di interleuchine, come IL 17 e 21, e sembra che siano coinvolte nella
patogenesi di diverse malattie autoimmuni (113) . Quindi ,se il TGF-β è presente in
entrambe queste differenziazioni, chi è il responsabile della conversione dei T naive
in Treg o in Th17 ? La risposta a questa domanda è stata trovata in una serie di
esperimenti, risalenti al 2011. Alcune T naive sono state infatti convertite in Th17
utilizzando TGF-β associato a IL 6 o altre interleuchine proinfiammatorie (114),
mentre altre in Treg , con TGF-β in associazione a IL 2 o acido retinoico (115).
Sembra inoltre che questo meccanismo molecolare permetta la formazione o di
Treg o di Th17, dato che alcune molecole espresse durante la trasformazione in un
tipo cellulare inibiscono mediatori dell'altro tipo: per esempio l'espressione della
FoxP3 esercita un'interferenza su alcuni fattori di trascrizione essenziali per lo
sviluppo dei Th17, ossia RORγt e RORα. La chiave di volta però, che determina il
futuro di un T naive, è legato al rapporto tra la concentrazione di STAT5 (molecola
espressa dall' IL2, che facilita l'espressione della FoxP3) / STAT3 (fattore di
trascrizione prodotto dall'IL 6, capace di inibire la conversione in Treg) (116).
Oltre a questa linea specifica di T helper, interleuchine differenti possono indirizzare
la risposta immunitaria più verso una di tipo T helper 1 o 2, dimostrando la grande
plasticità e variabilità del sistema immunitario. Nella foto in basso sono riportati i
principali tipi cellulari in cui i T naive possono differenziarsi e le molecole chiave in
questi processi (Fig. 6).

37
Figura 6: Differenziazione del linfocita T "naive" in Treg o Teff e le molecole coinvolte.
Nell'illustrazione precedente però non sono considerate altre cellule, scoperte
recentemente, con proprietà immunosoppressive, quali per esempio i Tr1 : queste
sono prodotte in vitro a partire da linfociti T naive in presenza di IL 10 e secernono
sia IL 10 che TGF-β (117); pur non presentando FoxP3, i Tr1 possiedono
caratteristiche simili a quelle dei Treg regolari, come il fatto di presentare una
ridotta proliferazione dopo stimolazione antigenica, deprimere per contatto cellule
adiacenti, produrre una modesta quantità di IL 2 e dimostrare un fenotipo di
membrana attivato (118). Sembra quindi che queste cellule in vitro condividano
parzialmente le stesse funzioni dei Treg, anche se il loro ruolo in vivo è ancora da
scoprire.
A rendere ancora più complesso e variabile la risposta immunitaria ci pensano
ancora una volta i Treg, per il fatto che questi infatti possono , in presenza di
determinati stimoli, dedifferenziarsi e convertirsi in altri tipi linfocitari, tra cui ,per
esempio, anche nei già menzionati Th17. In uno studio infatti è stato osservato che

38
Treg, FoxP3+, si sono convertite in Th17 in un ambiente infiammatorio, con un alta
concentrazione di IL6 (119). Inoltre sono state individuate in vivo delle cellule
FoxP3+che presentavano incredibilmente anche RORγt, mettendo in evidenza
l'intricato ma parallelo rapporto che queste 2 linee linfocitarie possiedono. Queste
cellule però, producevano una minore quantità di IL 17, probabilmente per il fatto
che FoxP3 inibisce l'espressione di questa citochina, inibendo direttamente sul
RORγt (120). Questa transizione sembra mediata dall'attività delle istone deacetilasi,
un gruppo di enzimi che sottraggono gruppi acetili dagli istoni del DNA, regolando di
fatto la sua trascrizione: è stato evidenziato che l'utilizzo di un inibitore di questi
enzimi, la tricostatina A, determina un marcato calo di questa trasformazione,
suggerendo che modifiche epigenetiche sono essenziali in questo processo (121).
Una maggiore trasformazione delle Treg in Th17 è stata invece osservata in un
esperimento, in cui erano state inserite in colture cellulari cellule dendritiche
attivate da un recettore di tipo C per le lectine ( dectina-1), implicato in meccanismi
nel riconoscimento di cellule fungine (122). Oltre a Th17, i Treg possono
differenziarsi in vere e proprie Teff: per esempio, in uno studio alcuni
nTreg ,prelevati dalle placche di Peyer e messi in un ambiente linfopenico, hanno
dimostrato una perdita d'espressione di FoxP3 ed hanno avviato un'attività da T
helper, volta a promuovere la produzione di IgA (123). Zhou, nel suo esperimento,
ha osservato che alcuni Treg, in vivo, perdono l'espressione della FoxP3 ed
assumono il fenotipo di cellule T della memoria: queste "ex FoxP3" derivano sia da
nTreg che da iTreg ed sono in grado di produrre grandi quantità di citochine
infiammatorie, soprattutto interferone γ (INF γ) (124) Quest'ultime cellule meritano
molte attenzioni dato che sono state ritrovate in diverse patologie autoimmuni,
come la sclerosi multipla (125), e questo fa presupporre che siano coinvolte nella
patogenesi di queste malattie. Da quanto detto finora, si può intendere quanto
possa essere complesso classificare ed individuare questa nuova versatile linea
cellulare. Per fortuna la FoxP3 ci aiuta nel riconoscimento di queste cellule, per il
fatto che rappresenta il marker migliore per queste cellule, nonostante molte altre
proteine vengano espresse: per esempio il CD 25 è frequentemente espresso sulla

39
loro membrana però è espresso anche su altri linfociti T attivati ed, inoltre, in
occasione di infezioni batteriche, alcune Treg hanno dimostrato un fenotipo FoxP3+
CD 25- (126). Una caratteristica tipica di questi Treg, utilizzabile in alternativa alla
FoxP3 ma più complicata, è una contemporanea up-regulation dei CD 25 ed una
down-regulation del CD 127, una subunità del recettore per l'IL7 (127). FoxP3 è un
marker ideale, non solo per il fatto che viene espresso esclusivamente dalle Treg,
ma che proprio costituisce un elemento essenziale nella loro attività
immunosoppressiva: infatti, è stato osservato che FoxP3 agisce da fattore di
trascrizione, riducendo l'espressione di IL 2 e 4 e IFN γ, mentre favorisce la sintesi di
CTLA 4 e CD 25 e GITR (proteina indotta dai glucocorticoidi correlata al TNFR) (128).
Dato che molti dei geni con cui interagisce la FoxP3, sono bersaglio di un altro
fattore di trascrizione, la NFAT insieme alla AP 1, e che sono stati osservati diversi
domini in grado di legarsi alla FoxP3 adiacenti a NFAT, si ipotizza che questi due
fattori di trascrizione siano correlati. La teoria più recente è che la FoxP3 inibisca
l'assemblaggio dei complessi NFAT:AP1:DNA, sostituendosi all'AP 1 e quindi
formando complessi NFAT :FoxP3:DNA (129). La FoxP3 riduce l'espressione di
queste IL interagendo anche con un altro fattore di trascrizione, la NF-kB (130).
Oltre a quest'azione cooperativa con NFAT e NF-kB, altri aspetti della FoxP3 come
fattore di trascrizione devono essere ancora approfonditi. Da due recenti studi , uno
di Marson et al (131) ed uno di Zheng et al. (132), condotti tramite tecniche di
immunoprecipitazione della cromatina ( ChIP), sono emersi dei risultati
interessanti . Nel suo studio, Marson, dopo aver visto che la FoxP3 era in grado di
legarsi con promotori di più di mille geni, ha confrontato i geni che venivano
espressi in cellule di ibridomi FoxP3 positivi e negativi, dopo una stimolazione
antigenica: da questo sono stati individuati per l'esattezza 125 geni che venivano
down-regolati nella popolazione FoxP3+ e non in quella Foxp3-, e la maggior parte di
questi sono coinvolti nella stimolazione del TCR e delle cellule T attivate. Risultati
analoghi sono emersi dallo studio dal lavoro di Zheng, che, oltre a confermare
l'ipotesi di Marson, ha rilevato la presenza di alcune modificazioni istoniche a livello
delle regioni a cui si lega la FoxP3, suggerendo che ,anche in questo ambito,

40
modifiche epigenetiche concorrono alla funzione di questo fattore di trascrizione.
Infine è stato riscontrato che la FoxP3 inibisce la CREB ( cAMP response element
binding ), un altro fattore di trascrizione, mediante il legame con la p300, una
proteina co-attivatrice di questa molecola (133). Un altro marker che potrebbe
essere utilizzato per differenziare nTreg dalle iTreg è Helios,un fattore di
trascrizione appartenente alla famiglia delle Ikaros (134): questa comprende una
serie di proteine capaci di interagire col DNA grazie ad due domini "zinc-finger" N-
terminali ed uno in grado di legarsi alle proteine C-terminale, struttura condivisa da
tutti i membri della famiglia (135). In realtà il loro utilizzo per distinguere nTreg
dalle altre recentemente è stato confutato da uno studio in cui sono stati
riscontrati Helios proprio in iTreg in vivo. Ciò nonostante, il loro utilizzo come
marker può essere sempre valido, dato che sono direttamente correlati alla loro
funzione rispetto che al grado di differenziazione delle Treg (136): infatti è stato
osservato che Helios determina una maggiore espressione di FoxP3, legandosi al
suo promotore e facilitando la sua trascrizione e che modificazioni di Helios possono
presagire ulteriori modifiche sulla FoxP3. Inoltre sembra che una iper espressione di
Helios induca apoptosi nei linfociti T CD 4 e questo fenomeno potrebbe essere
utilizzato per combattere malattie neoplastiche (137).
Attività immunodepressiva dei Treg:
Dopo aver trattato in lungo e in largo come viene regolata l'espressione e la
funzione della FoxP3, ed delle Treg, vediamo come le Treg esercitano la loro azione
immunodepressiva sulle altre cellule del sistema immunitario, la quale può essere
riassunta in 2 differenti meccanismi:
Meccanismo dipendente da contatto: come indicato nella definizione, la
soppressione della risposta immunitaria avviene grazie al contatto tra Treg e
cellule immunitarie, o , più precisamente grazie all'interazione tra le varie
proteine di membrana che possiedono. La più conosciuta è sicuramente il CTLA
4: questa proteina ( già discussa anteriormente), è in grado di legarsi alle
proteine B7-1 e B7-2, presenti sulle APC, e questo legame stimola l'attivazione
delle Treg ( e quindi gli altri meccanismi soppressivi) e riduce l'espressione del

41
CD 28, il cui legame con le B7 potrebbe inibire lo sviluppo delle Treg (138). A ciò
si aggiunge il fatto che il legame tra CTLA 4 e B7, determina l'attivazione di un
enzima all'interno delle cellule dendritiche, e cioè la indoloammina-pirrolo 2,3
diossigenasi,chiamata anche IDO: questa catalizza una reazione che porta alla
conversione del triptofano in chinurenine e altri metaboliti che esercitano
effetti immunodepressivi nell'ambiente circostante (139). Questi infatti
inducono citotossicità sulle APC oppure convertono T naive in Treg, oltre a
ridurre la concentrazione di triptofano, che determina una riduzione
dell'attivazione delle cellule T (140). A ciò, si aggiunge la capacità che hanno
queste Treg di indurre apoptosi, e lo fanno in maniere differenti: è stato
osservato per esempio che iTreg umani esprimono granzimi B e sono in grado
di eliminare cellule tumorali utilizzando perforine (141); inoltre , sia nTreg che
iTreg , possiedono capacità citotossiche mediate da perforine contro diverse
cellule autologhe del sistema immunitario, come linfociti T helper, citossici,
macrofagi CD 14+ e cellule dendritiche, eliminandole senza l'intervento del FasL
(142) . Infine è presente una molecola d'adesione, il CD 223 ( chiamato anche
LAG 3, ossia gene per l'attivazione del linfocita) ed è associato al CD 4 , con cui
riesca ad interagire con l'MHC di classe II. Il ruolo di questa proteina di
membrana nella immunosoppressione non è ancora chiara, ma sembra che ne
prenda parte, dato che anticorpi anti-LAG3 eliminano l'effetto depressore delle
Treg in vivo (143).
Meccanismo mediato da citochine: questo costituisce un importante mezzo
con cui le Treg riescono a provocare nelle loro vicinanze immunodepressione,
in cui assumono un ruolo chiave le citochine IL 10, TGF-β e IL 2.
o IL 10: Per quanto riguarda l'IL 10 sono stati fatti diversi studi in vivo: in uno di
questi , era stato osservato che l'utilizzo di Treg in trapianti allogenici di cute
induceva tolleranza, mentre l'aggiunta a questi di anticorpi anti-recettore dell'IL
10, determinava un rapido rigetto (144). In vitro è stato notato che l'IL 10
induce l'espressione nelle APC di una molecola costimolatoria di superficie, la
B7-H4, e questa rendeva l'APC stessa immunodepressiva (145).

42
o TGF-β: questa proteina oltre ad inibire la proliferazione e la differenziazione
delle Teff e l'attivazione dei macrofagi (82), è coinvolto nell'inattivazione dei
natural killer ( NK ) (146) e nella tolleranza ad alcuni parassiti. Riguardo a
quest'ultimo aspetto , è stato condotto uno studio in cui un parassita,
Heligmosomoides polygirus¸ aumenta la popolazione di Treg a livello intestinale
e si ritiene che questo fenomeno costituisca una valida strategia contro il
sistema immunitario dell'ospite del parassita. Alla base di questo processo è
stato individuato un antigene parassitario chiamato HES ( H.polygirus excretory-
secretory antigen), che ha la capacità d'indurre Treg e di sopprimere la
replicazione delle Teff in vitro: si presume che questo HES possa legarsi ai
recettori del TGF-β e mimare la sua azione per il fatto che il suoeffetto
immunodepressivo mancava se venivano utilizzati degli inibitori del TGF-β (147).
Oltre a questi effetti, dobbiamo ricordare l'importanza di questo mediatore nel
mantenimento delle nTreg e nell'induzione di nuove iTreg.
o IL 2: oltre a costituire un importante fattore di proliferazione e differenziazione
dei linfociti attivati dall'antigene, funge da feed back negativo sulla risposta
immunitaria (79).
Treg e FoxP3 negli animali domestici:
Tutto ciò che abbiamo finora riportato, sono i dati di innumerevoli esperimenti e
lavori su Treg umane e di topo, ma cosa sappiamo di più specifico nei nostri animali
domestici ? Sicuramente nella specie canina, le nostre conoscenza sono più vaste: il
concetto di "tolleranza periferica" non è affatto nuovo dato che Weiden, già nel
1976 aveva notato che una popolazione di cellule T, trasferita in alcuni cani trattati,
era responsabile dell'assenza di rigetto al trapianto (148). I Treg, in questa specie,
sono state scoperte in uno studio di Sacchini et al (149) nel 2007 e da allora sono
stati fatti molti altri lavori che prendevano in esame questa nuova linea
cellulare ,relativamente a malattie infiammatorie, parassitarie, autoimmuni e
tumorali; proprio su quest'ultimo ambito , riporto uno studio del 2009 ad opera di
O'neil (150), in cui comparava la quantità Treg in cani sani e in cani che
presentavano diversi tipi di tumori presenti, utilizzando come campioni, sangue e

43
linfonodi che drenavano il tumore: gli animali con tumori mostravano valori più alti
della popolazione sana, e, presi individualmente, nei carcinomi si rilevava il numero
più elevato di Treg. C'è da dire però che non tutti i lavori ,svolti da molti ricercatori
differenti, hanno dato i medesimi risultati: per esempio Rissetto et al (151). non
hanno riscontrato differenze nella quantità di Treg nel sangue o nei linfonodi, sia
vicini che lontani, tra cani sani e quelli che presentavano osteosarcoma.
Nel gatto, la scoperta di " cellule soppressorie" sono già note dai primi anni ottanta,
grazie ad un lavoro di Langweiler (152), ma si inizia a parlare di Treg dopo una serie
di lavori "cardine" sul rapporto di queste cellule con il FIV (Virus dell'
immunodeficienza felina) ad opera di Wayne e Mary Tompkin: da questi lavori è
stato scoperto che:
1. Queste cellule esprimevano FoxP3 (153)
2. Il contatto tra queste e altre cellule immunitarie determinava un arresto della
proliferazione e della produzione di IL 2 in vitro (154)
3. La presenza di queste cellule e la loro attivazione durante le prime fasi
dell'infezione sembrava che riducesse la risposta immunitaria contro il virus e
quindi permettesse l'instaurarsi dell'infezione cronica (155)
4. il legame tra TGF-β e il suo recettore influenzava l'attività di queste cellule e la
progressione dell'infezione. (156)
5. La deplezione dei Treg che poteva indurre questa malattia (157)
6. l'impatto che queste cellule avevano sulla risposta anti-virale di soggetti
cronicamente infetti (158).
Tumori e linfociti T :
Dopo aver parlato della presenza di queste cellule nei nostri animali domestici,
dobbiamo ancora chiarire un aspetto. Qual' è di preciso il rapporto tra le malattie
neoplastiche e la risposta T e Treg? E' stato osservato che la morte di cellule
tumorali , rilascia antigeni tumorali e altri segnali di pericolo che vengono captati
dalle cellule dendritiche locali, le quali , una volta attivate, sono capaci di reclutare
diversi linfociti T tumore -specifici nel sito d'infiammazione, chiamati TIL ( tumor-
infiltrating lymphocytes) ; i TIL dimostrano di saper reagire specificamente agli

44
antigeni delle cellule neoplastiche e , la loro presenza in alcuni tumori è stata
associata ad una maggiore probabilità di sopravvivenza del paziente (159) (160). La
presenza di TIL ( in cui possono presentarsi anche i Treg) non indica ,di per se, una
migliore prognosi o meno, dato che le cellule tumorali possiedono un vasto e
complesso "arsenale" molecolare, che le permette di sfuggire al sistema
immunitario.
I meccanismi con cui lo fanno sono principalmente:
Alterazione del riconoscimento antigenico: uno delle maniere con cui i tumori
riescono meglio a sfuggire alla risposta immunitaria è proprio quello di evadere
il meccanismo di riconoscimento antigenico, come una minor espressione o la
perdita completa dell' MHC di classe I: infatti è questa la molecola attraverso
cui i linfociti T citotossici ( CD 8+) monitorano le nostre cellule, eliminandole in
caso riscontrino infezioni virali o trasformazioni tumorali. Le cause che possono
portare ad alterazioni del MHC I sono mutazioni genetiche, alterazione della β-2
microglobulina, implicata nel trasporto del MHC I sulla superficie cellulare (161),
o altre modifiche epigenetiche , come una ipermetilazione istonica, che
determinano una riduzione dell'espressione di questa molecola (162). Difetti in
altre molecole coinvolte nella presentazione dell'antigene, come LMP 2 e 7,
TAP 1 e 2 o chaperoni come la tapsina, possono causare una perdita, parziale o
totale, di MHC di classe I sulla membrana cellulare (163). Anche difetti nel IFN γ
o in molecole del suo "pathway" possono dare luogo ad uno scenario simile,
dato che l'IFN γ regola l'espressione di TAP 1 e LMN 5 (164). Insieme a questi
fenomeni, un tumore può sfuggire al confronto con linfociti T citotossici
riducendo l'essudazione di questi dai vasi sanguigni: ciò viene raggiunto grazie
alla riduzione dell'espressione di alcune molecole d'adesione, come ICAM 1 o
altre selectine sui vasi che attraversano alcune neoplasie (165).
Resistenza al meccanismo di eliminazione mediato dai T citotossici: sono
sostanzialmente due i "pathway" con cui i CD 8+ possono indurre una cellula in
apoptosi, e sono quello delle perforine / granzimi e quello del recettore della
morte. Al primo, alcuni tumori possono sfuggire, grazie alla produzione di un

45
enzima che inibisce l'azione del granzima B, ossia la PI 9/ SPI 6, in particolare
alcuni melanomi, carcinomi cervicali e mammari (166). Il funzionamento
dell'altro sistema di "killing" è un po' più complesso: il legame tra recettori della
morte come CD 95 (o Fas) o TRAILR 1 e 2, e il loro ligando naturale, e
cioè,rispettivamente FasL e TRAIL, scatena un complesso sistema di trasduzione
del segnale, che porta alla distruzione guidata della cellula intera. Questo
processo però presenta diverse fasi in cui può essere bloccato: per esempio è
stato osservato che c-FLIP può opporsi all'apoptosi mediata da TRAIL ed
un'espressione elevata di questa proteina anti-apoptotica in tumori intestinali è
stata associata ad prognosi peggiori (167). Le cellule tumorali possono sfuggire
all'apoptosi anche esprimendo una minor quantità di recettori della morte
come accade, per esempio, in alcune forme di cancro al colon, dove si ha una
riduzione di CD95 (Fas); ciò nonostante è stato osservato che i livelli di questo
recettore sono aumentati in presenza di alcune citochine, come TNF-α e IFN γ
(168). Un'altra strategia adottabile dai tumori per scampare alle attacco delle
cellule immunitarie, è la produzione di recettori della morte transmembrana e
solubili non funzionali("decoy"): è stato evidenziato infatti che, in alcuni tumori,
queste molecole competono con i recettori funzionali per i loro ligandi,
aumentando di fatto le possibilità di sopravvivenza delle cellule tumorali (169).
Disattivazione del linfocita T attivato : grande importanza viene rivestita da una
molecola già ampliamente trattata, ossia il CTLA 4. Da uno studio è emerso
infatti che le cellule tumorali ,insieme ad altre immunitarie, sono in grado di
determinare una up-regulation di queste proteine di membrana e, quindi ,
ridurre la risposta immunitaria nel micro ambiente del tumore (170). Alcuni
tumori sono in grado di produrre invece TGF-β , che, oltre a essere implicata
nell'induzione delle Treg (vedi sopra), influenza anche l'attività dei Teff: infatti
sembra che la citochina in questione ostacoli la cascata di reazioni intracellulari
alla base dell'attivazione dei linfociti T (171) e riduca la citotossicità dei CD 8+,
andando a ridurre l'espressione di proteine quali perforine e granzimi (172); per
questi motivi si ritiene che tumori in grado di secernere TGF-β presentino una

46
prognosi peggiore (173). In altri tumori invece, elevate concentrazioni di
ciclossigenasi-2 (COX 2) e suoi prodotti ( PGE2) sono stati individuati da tempo :
molti concordano sul fatto che la presenza di queste molecole faccia virare la
risposta immunitaria verso il tipo Th2, meno efficace nell'immunità tumorale
(174). Infine, un altro meccanismo utilizzato dalle cellule neoplastiche per
ridurre l'attivazione dei T citotossici consiste nell'utilizzo delle galectine : queste
sono delle proteine capaci di glucidi, in particolar modo β-galattosidi, e sono
coinvolte in diversi processi cellulari quali, chemiotassi,adesione , apoptosi etc.
(175); le galectine sono state recentemente considerate dei fattori
immunosoppressivi, e la loro presenza è stata riscontrata in diversi tipi di
tumori umani (176). La galectina 1 , in particolar modo, sembra possedere
proprietà immunodepressive , inducendo apoptosi nei linfociti T (177) e
ostacolando l'attivazione del TCR (178).
Meccanismi di "killing" mediati dal tumore : alcuni tumori sono in grado di
adottare dei meccanismi con cui uccidere le cellule del sistema immunitario. Un
esempio, un po' contradditorio riguarda proprio una proteina già menzionata,
ossia il FasL: questa molecola viene secreta in grande quantità dai linfociti T che
subiscono una stimolazione al TCR,ma non una costimolazione e questa grande
quantità di FasL rilasciata, va ad indurre apoptosi in altre cellule T ( linfocita
"fratricida" ), comprese i linfociti che la secernono (linfocita "suicida"). Questa
molecola dunque , espressa in diversi tipi di tumore differenti (179) , sembra
possedere un effetto inibitorio sulla risposta immunitaria (180). Ciò
nonostante ,alcuni studi affermano che la FasL possegga proprietà pro
infiammatorie e quindi antitumorali in vivo, come evidenziato in alcuni
esperimenti. In questi il trapianto di tumori geneticamente modificati in modo
da esprimere sulla membrana FasL , inducevano un rigetto più
rapido ,accompagnato da una abbondante infiltrazione neutrofilica (181).
Ulteriori complicazioni sono sorte quando recenti studi hanno messo in
evidenza alcune microvescicole contenenti FasL presenti in alcune cellule di un
melanoma, la cui degranulazione determinava la morte di diversi precursori

47
della linea linfoide ed effetti immunodepressori sistemici (182); oltre agli effetti
mediati dal legame di questo col suo recettore della morte, si ritiene che le
microvescicole contengano anche molecole con proprietà immunosopressive
(183). Un altro sistema di difesa adottato dalle cellule neoplastiche utilizza un
enzima la cui azione è già stata trattata, ossia il IDO: infatti ,in uno studio, è
stato osservato che alcune tumori esprimono questa molecola e che l'utilizzo di
levo-1 metiltriptofano blocca il suddetto enzima (184). Da questo lavoro si
pensa che molecole in grado di ridurre la concentrazione di questo enzima,
possano essere utilizzate per migliorare l'immunoterapia.
Reclutamento di cellule ad azione immunosoppressiva : oltre ai già discussi Treg
è stata descritta un altra categoria cellulare ,responsabile di modulare la
risposta immunitaria nei tumori, e, cioè, le MDSC (o cellule soppressorie di
derivazione mieloide): queste cellule vengono prodotte in risposta ad alcune
citochine provenienti dal tumore e sono costituite da una serie di cellule
mieloidi a vari stadi di differenziazione. In stadi avanzati di cancro, sono stati
ritrovati MSDC, probabilmente attirati da GM-CSF (fattore di stimolazione delle
colonie di granulociti e macrofagi) secreto da cellule tumorali (185), hanno
dimostrato di indurre una riduzione dell'IFN γ secreto da parte di linfociti T CD
8+. Infine Le MSDC sembra che inibiscano i linfociti T grazie alla produzione di
enzimi in grado di metabolizzare amminoacidi, quali arginasi, ossido nitrico
sintasi e altri (186).
Treg e immunità tumorale:
Da quanto riportato in questa parte, possiamo notare quanto può essere complessa
ed articolata la difesa che oppone un tumore alla sua eliminazione, ma che ruolo
hanno i Treg in tutto questo ? Il primo studio che ha messo in evidenza un rapporto
tra Treg e immunità tumorale, risale al 1999 : qui l'utilizzo di anticorpi monoclonali
anti CD 25 (recettore dell' IL 2) prima dell'inoculazione di un tumore, conferiva
protezione nei confronti della malattia neoplastica (187). Seguenti studi hanno
rivelato infatti la presenza di Treg in diversi tipi di tumori e l'anticorpo che blocca il
suddetto recettore determina una riduzione del numero di queste cellule ; è stato

48
osservato in un altro esperimento che la deplezione di Treg mediata dalla
"Denileuchina Difitox", un'immuno-tossina coniugata con l'IL 2, provoca un
potenziamento della risposta immunitaria e la riduzione della crescita del tumore
(188). Un'altra dimostrazione delle loro proprietà immunodepressive nei tumori, è
data da un esperimento in cui trasferimenti di T helper senza Treg risultano molto
più protettivi rispetto a trasferimenti con questi ultimi presenti (189). Come la
deplezione di Treg aumenta la risposta antitumorale, così un aumento di questi
linfociti riduce la protezione che il sistema immunitario conferisce nei confronti di
queste malattie. Postulando che i Treg sono in grado di riconoscere alcuni antigeni
self associati al tumore, identificati dal SEREX (acronimo per "SErological
identifaction of antigens by Recombinant EXpression cloning" ), un innovativo
metodo per rilevare antigeni tumorali, riporto un lavoro del 2003: qui
l'immunizzazione con plasmidi codificanti per questi antigeni self immunogeni
facilitavano la metastatizzazione di cellule tumorali iniettate via endovenosa. Il
trasferimento di Treg e non di T helper, da questi soggetti, potenziava la metastasi,
indicando che i primi sono ,almeno in parte, responsabili dell'immunodepressione in
questi soggetti (190).
Nell'uomo il coinvolgimento dei Treg nell'immunità tumorale è illustrato , per
esempio, dalla risposta a NY-ESO-1, un antigene espresso in diversi tipi di neoplasie
umane. E' stato osservato che la risposta umorale verso questo antigene è
direttamente proporzionale all'attività dei linfociti T citotossici specifici per NY-ESO-
1, e ciò indica che anche i Linfociti helper sono coinvolti in questa reazione,
aumentando i livelli di immunità cellulo-mediata e anticorpale contro l'antigene in
questione. La risposta Th1 è rilevabile in pazienti affetti da cancro,in cui
spontaneamente vengono prodotti (191) ; straordinariamente, T helper prelevati
da individui sani, possono, in vitro, reagire contro NY-ESO-1 dopo l'eliminazione di
Treg. Da questo dato si intuisce che linfociti T CD 4+ NY-ESO-1 specifici sono
presenti in tutti gli individui, ma la loro attivazione è regolata proprio dai Treg.
Analogamente, anche altri T helper "naive" contro diversi antigeni tumorali, come la
tirosinasi nel melanoma, sono presenti in soggetti sani e mantenuti quiescenti dai

49
Treg (192). Questi sono stati riscontrati in masse tumorali e linfonodi drenanti di
diversi tipi di neoplasie ed il loro numero e, in particolar modo, il rapporto tra T CD
8+ e FoxP3+ sono stati correlati a prognosi infauste nel tumore mammario (193) ,
gastrico (194) e ovarico (195). Il meccanismo di reclutamento, si ritiene sia mediato
dai macrofagi infiltranti il tumore ( o TIM), tramite la loro secrezione di chemochina
CCL 22, capace di attrarre Treg per la loro espressione superficiale di CCR 4 (196);
sembra che anche uno stato infiammatorio sia in grado di attirare Treg (197) nel sito
tumorale. Inoltre c'è da considerare che alcune cellule neoplastiche e/o cellule
dendritiche all'interno del tumore producono una grande quantità di TGF-β,
responsabile dell'induzione di Treg in vitro (198). Si ipotizza che gli antigeni rilasciati
dalle cellule tumorali in attiva mitosi o mentre muoiono vengano riconosciuti dai
Treg , e che ciò induca una loro moltiplicazione (199). Nell'illustrazione sottostante
è raffigurato schematicamente come viene attuata l'immunodepressione a livello di
un tumore, i cui meccanismi molecolari sono stati ampiamente trattati
precedentemente.
Figura 7 Rappresentazione del ruolo dei Treg nell'immunità tumorale
FoxP3 ed immunoterapia
La conoscenza di tutti questi complessi meccanismi molecolari alla base della
fisiologia dei Treg sono di importanza essenziale, dato che ,da queste nozioni, è
possibile individuare possibili bersagli per farmaci immunoterapici, come
dimostrato da diversi studi a riguardo. Da questi, sono stati elencati una serie di

50
molecole che possono entrare nel novero di possibili bersagli ed i più interessanti
sono :
IL 2 e CD 25: sicuramente l'utilizzo di anticorpi anti-CD 25 o immunotossine
unite all' IL 2 ,come dimostrato da alcuni esperimenti(riferimenti bibliografici
prima), hanno come effetto quello di ridurre la popolazione di Treg attivati; Ciò
nonostante, dato che il Cd 25 è comunque espresso sulla superficie di linfociti T
attivati e l' IL 2 è importante anche nell'attivazione dei T citotossici, il blocco di
queste molecole sembra ostacolare parzialmente anche la risposta
antitumorale (200). A questo riguardo però bisogna osservare che , se
l'anticorpo monoclonale contro il CD 25 viene somministrato prima del tumore,
la risposta immunitaria è comunque sempre forte, mentre se avviene dopo
l'inoculazione della massa neoplastica risulta molto più debole (201). Da ciò si
deduce che l'utilizzo di IL 2 possa avere degli effetti opposti, in base alla fase e
allo stato della risposta immunitaria contro il cancro.
GITR: già menzionata prima, è una molecola costimolatoria espressa in linfociti
T helper e citotossici "naive", ed up-regolata durantela loro attivazione (202).
Inoltre è espresso ad alti livelli nelle Treg e la sua attivazione tramite un
anticorpo monoclonale agonista o il suo ligando, determina un calo dell'attività
immunodepressiva di questo tipo cellulare (203); trattamenti con le 2 molecole
sopracitati hanno dimostrato un aumento della risposta tumore -specifica, sia
di tipo T helper che citotossica (204). Inoltre è stato osservato che il
trattamento anti-GITR era più efficace se il tumore era di maggiori dimensioni:
questo rispecchia probabilmente il fatto questo trattamento potenzia l'attività
di linfociti T citotossici a cui è già stato presentato l'antigene e ,quindi, attivati
(205). Infine la stimolazione di questo recettore insieme a quella di antigeni
tumorali, ha dimostrato non solo un incremento delle Teff tumore-specifiche,
ma anche una refrattarietà all'azione soppressiva delle Treg (206). I risultati qui
riportati fanno del GITR uno dei bersagli terapeutici più interessanti al
momento

51
CTLA-4: abbiamo già discusso abbondantemente sul ruolo di questa molecola
costimolatoria nella fisiologia dei linfociti, sia Teff che Treg. Alcuni studi hanno
messo in evidenza che il blocco selettivo di questa proteina di membrana
determini un'inibizione sulla crescita del tumore, riducendo gli effetti depressivi
dei Treg e potenziando quelli infiammatori delle Teff (207). Oltre a questo però,
è stato osservato che l'inibizione della CTLA 4 determina non solo una riduzione
di Treg nella massa neoplastica,ma anche di T citossici (208). Pertanto,
l'associazione di mAb anti-CTLA 4 con altre immunoterapici sembra una
promettente via per stimolare efficaci risposte antitumorali.
OX 40: anche questa è una molecola costimolatoria, della famiglia dei recettori
del TNF ed è espressa transitoriamente dai linfociti T attivati, mentre è
costitutiva nei Treg. Alcuni studi avevano dimostrato che la stimolazione
tramite mAb anti-OX 40 riduce la loro attività immunodepressiva (209). In un
altro studio, l'uso di Treg e Teff, entrambi privati di OX 40, ha dimostrato che la
stimolazione di questa molecola non ha solo effetto sulle Treg, ma anche sulle
Teff, le quali vengano potenziate dal suddetto stimolo. Inoltre In uno studio è
stato osservato che l'uso di OX40L o anticorpi con azione agonista aumentasse
la protezione contro forme tumorali, quali melanoma, carcinoma mammario,
sarcoma e carcinoma al colon, in alcuni topi (210).
Recettore per l'acido folico (FR) 4: i roditori hanno le Treg che esprimono RF 4
ad un livello maggiore delle T naive e, su stimolo del TCR, sono in grado di
esprimerlo ancora di più (211). Pertanto l'utilizzo di mAb anti FR 4 sembra
aumentare la risposta immunitaria, riducendo la popolazione di Treg attivati,
risparmiando le altre popolazioni linfocitarie. Rimane da dimostrare se nei
nostri animali anticorpi anti FR 4 hanno la stessa azione.
In tutte queste, il traguardo che si è cercato di raggiungere è quello di spostare
l'equilibrio della risposta immunitaria dalla parte delle Teff, come si può vedere
nella figura 8.

52
Materiali e metodi:
Campioni:
Ventisette meningiomi che presentavano all'istologia un infiltrato infiammatorio
sono stati prelevati da altrettanti gatti, le cui caratteristiche sono riportate nella
pagina sottostante (tabella 2). I campioni, fissati in formalina di Policard al 10 % al
momento del prelievo, sono stati inclusi in blocchetti di paraffina e tagliati al
microtomo in sezioni di 5 micrometri e posti su alcuni vetrini, per poi essere
sparaffinati tramite un passaggio in xilolo ed infine idratati grazie all'uso
di ,rispettivamente, alcol 100° e alcol a 96°.
Figura 8: Potenziali bersagli utilizzabili per alterare l'equilibrio tra Treg e Teff ed effetto sulla risposta immunitaria antitumorale
53
Numero
istologiaSesso Età Localizzazione della massa
12852 Maschio castrato 8 Fronto parietale
12451 Femmina castrata 13 Multifocale
12321 Femmina castrata 8 Parieto occipitale
12166 Femmina 15 Temporo parietale
12103 Maschio castrato 11 Temporale
12100 femmina 13 Frontale
11815 Maschio castrato 13 Parieto occipitale
11796 Maschio castrato 16 fronto parietale
11772 Maschio 14 Prosencefalo
11748 Maschio 13 Fronto parietale
11427 Femmina castrata 14 Parietale
11319 Maschio castrato 13 Parietale
11183 Femmina castrata 13 Emisfero sx
11134 Femmina 15 Spinale (T1- 2)
11019 Femmina n.d. n.d.
10923 Femmina castrata 11 Fronto temporale
10867 Femmina castrata 11 Spinale (C3-4)
10845 Femmina 18 mesi Tela corioidea 3° ventricolo
10360 Femmina castrata 12 Parieto occipitale
10301 Maschio castrato 17 Prosencefalo
10296 Maschio 15 Temporale
10105 Maschio 15 Prosencefalo
10104 Maschio 14 Prosencefalo
10039 Femmina 11 Spinale (T13-L1)
10037 Maschio 11 Tela corioidea 3° ventricolo
10005 Maschio 9 Della falce
9857 Maschio castrato 13 Multifocale
Tabella 2 Sesso, età (anni) e localizzazione del meningioma, dai pazienti analizzati nello studio
Immunoistochimica (IH):
Come prima fase, è stato effettuato un "antigen -retrieval" su tutti i
campioni,tranne quelli destinati ad essere incubati con gli anticorpi anti-CD 20, con
Tampone Citrato pH 6 (Bio-Optica, Milano), portata ad una temperatura vicina al
punto di ebollizione per circa 13 minuti,grazie ad un forno a microonde. Una volta
raffreddati i vetrini, sono stati montati su sheldon coverplate. Ho bloccato le

54
perosidasi endogene impiegando un bloccatore delle perossidasi (Peroxidase
Blocking Solution ,Dako REALTM , Carpinteria, CA ), e dopo il lavaggio con una
soluzione "madre"(costuita da 500 ml di TBS 1X + 250 µl di tween), ho aggiunto 2
gocce di Ultra V-block ( Ultra V Block ,Thermo Scientific, Fremont, CA ) .Dopo aver
eseguito ancora lavaggi, ho somministrato nel serbatoio del vetrino,100 µl di
anticorpo primario diluito, utilizzando la soluzione madre, a concentrazioni
specifiche per anticorpo:
Anti CD 3 : mouse monoclonal IgG1 , Santa Cruz Biotechnology - diluizione 1:50
Anti CD 79a: mouse monoclonal IgG1 ,Santa Cruz Biotechnology - diluizione 1:50
Anti CD 20: Thermo Scientific, Fremont, CA - diluizione 1:400
Anti Foxp3: Anti-human/mouse Foxp3 purified, eBioscience - diluizione 1:100
Ho lasciato gli anticorpi primari a incubare per un'ora a temperatura ambiente,
tranne per i quelli anti FoxP3, per i quali li ho lasciati in una overnight a temperatura
di refrigerazione. Dopo aver eseguito un lavaggio con la soluzione madre, è stato
impiegato ,per tutti, l'anticorpo secondario (Biotynilated goat polyvalent scondary,
Thermo Scientific Fremont, CA ) e, dopo 10 minuti di attesa, un ennesimo lavaggio,
è stato inserito il complesso streptavidina-perossidasi ( Streptavidin - Peroxidase,
Thermo Scientific Fremont, CA). Per la rivelazione, si è ricorsi alla soluzione DAB
( ImmPACT TM DAB Peroxidase Substrate Kit, Vector, Burlingame, CA ) e,
successivamente il vetrino ha subito un lavaggio con acqua distillata e smontato dal
coverplate. Successivamente, il vetrino è stato messo 50 secondi in ematossilina,
per poi sciaquato e essere disidratato ed infine montato su un vetrino normale, con
sopra un vetrino coprioggetti. Un linfonodo di gatto è stato utilizzato come
controllo positivo mentre i risultati sono stati calcolati mediante dieci conteggi
"random" su un'area standardizzata di 8,25 x 10-2 mm2 per vetrino ; le analisi
statistiche dei risultati sono state effettuate con il software SPSS 13 (Statistical
Package for Social Science, IBM ,New York, USA) dove: L'analisi della varianza è
stata effettuata mediante un ANOVA, mentre Le differenze tra i gruppi presi in
esame con il test di Tuckey e Bonferroni Le correlazioni tra le diverse popolazioni
con il test di Pearson

55
Risultati:
I campioni sono stati prelevati per la maggiora parte da necroscopie, e provengono
da 27 gatti domestici. La quasi totalità (89,2%) è rappresentata da tumori
intracranici, mentre la restante parte (10,8%) da neoplasie spinali; all'interno dei
primi la stragrande maggioranza delle neoplasie si presentava nella regione
sopratentoriale (87,5%) Tutti i meningiomi presi in esame sono di grado WHO I e
sono costituiti da diversi istotipi, tra cui transizionale (48,1%), meningoteliale
(33,3%), psammomatoso (11,1%) e fibromatoso ( 7,4%). I gatti da cui è stata
effettuato il prelievo erano per il 51,8% femmine, e 48,1 maschi, mentre la
percentuale di pazienti sterilizzati era di 51,8 % contro il 48,1% di pazienti interi. I
controlli positivi sono venuti positivi per CD 3 e CD 20, dimostrando cellule positive
per i primi nella zona perifollicolare,mentre per i secondi nel follicolo linfatico (Fig. 9)
Figura 9: A: Linfociti T presenti nella sezione del linfonodo felino B: Linfociti B nella medesima sezione
Il controllo positivo per il CD 79a non ha avuto successo, al contrario di quello per la
FoxP3,in cui si poteva vedere la disposizione nucleare della proteina cercata(Fig. 10 )
Figura 10: A: Linfociti Treg nella sezione di linfonodo felino, 20X B: Linfociti Treg nella stessa sezione , 40 X

56
Tabella 3: Sono indicati i l'istotipo, localizzazione, media di linfociti CD 3+,CD 20
+ , FoxP3
+ intra e peritumorali e
grado di necrosi singolarmente, caso per caso . * = media delle cellule immunopositive calcolata mediante dieci conteggi "random" su un'area standardizzata di 8,25 x 10
-2 mm
2 |n.d. = non determinata |
- = assente | +/- = microfocolai isolati | + = microfocolai confluenti
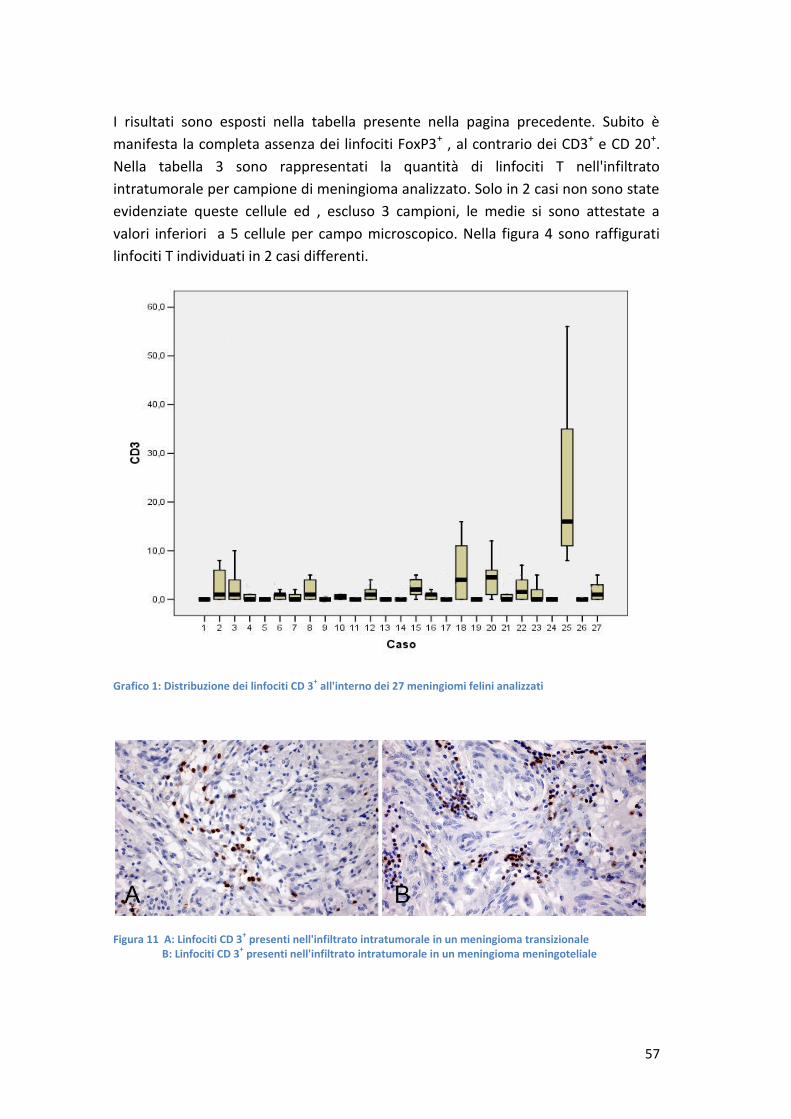
57
I risultati sono esposti nella tabella presente nella pagina precedente. Subito è
manifesta la completa assenza dei linfociti FoxP3+ , al contrario dei CD3+ e CD 20+.
Nella tabella 3 sono rappresentati la quantità di linfociti T nell'infiltrato
intratumorale per campione di meningioma analizzato. Solo in 2 casi non sono state
evidenziate queste cellule ed , escluso 3 campioni, le medie si sono attestate a
valori inferiori a 5 cellule per campo microscopico. Nella figura 4 sono raffigurati
linfociti T individuati in 2 casi differenti.
Grafico 1: Distribuzione dei linfociti CD 3+ all'interno dei 27 meningiomi felini analizzati
Figura 11 A: Linfociti CD 3+ presenti nell'infiltrato intratumorale in un meningioma transizionale
B: Linfociti CD 3+ presenti nell'infiltrato intratumorale in un meningioma meningoteliale
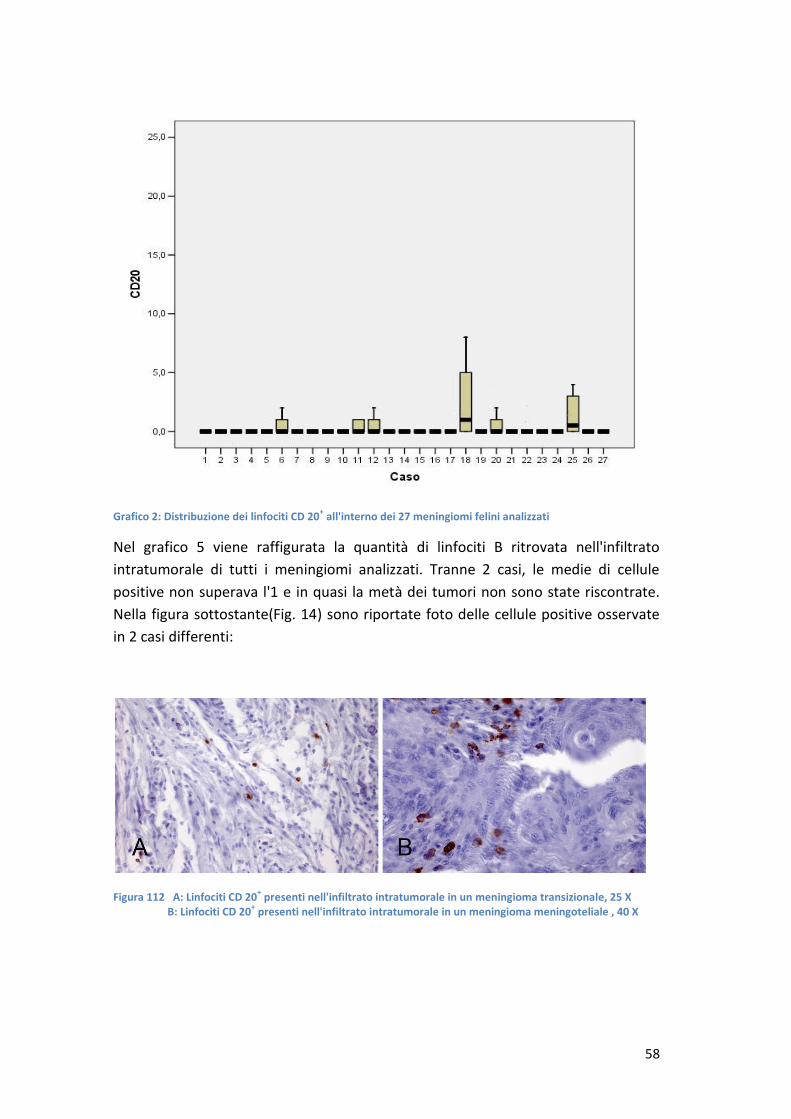
58
Grafico 2: Distribuzione dei linfociti CD 20+ all'interno dei 27 meningiomi felini analizzati
Nel grafico 5 viene raffigurata la quantità di linfociti B ritrovata nell'infiltrato
intratumorale di tutti i meningiomi analizzati. Tranne 2 casi, le medie di cellule
positive non superava l'1 e in quasi la metà dei tumori non sono state riscontrate.
Nella figura sottostante(Fig. 14) sono riportate foto delle cellule positive osservate
in 2 casi differenti:
Figura 112 A: Linfociti CD 20+ presenti nell'infiltrato intratumorale in un meningioma transizionale, 25 X
B: Linfociti CD 20+ presenti nell'infiltrato intratumorale in un meningioma meningoteliale , 40 X
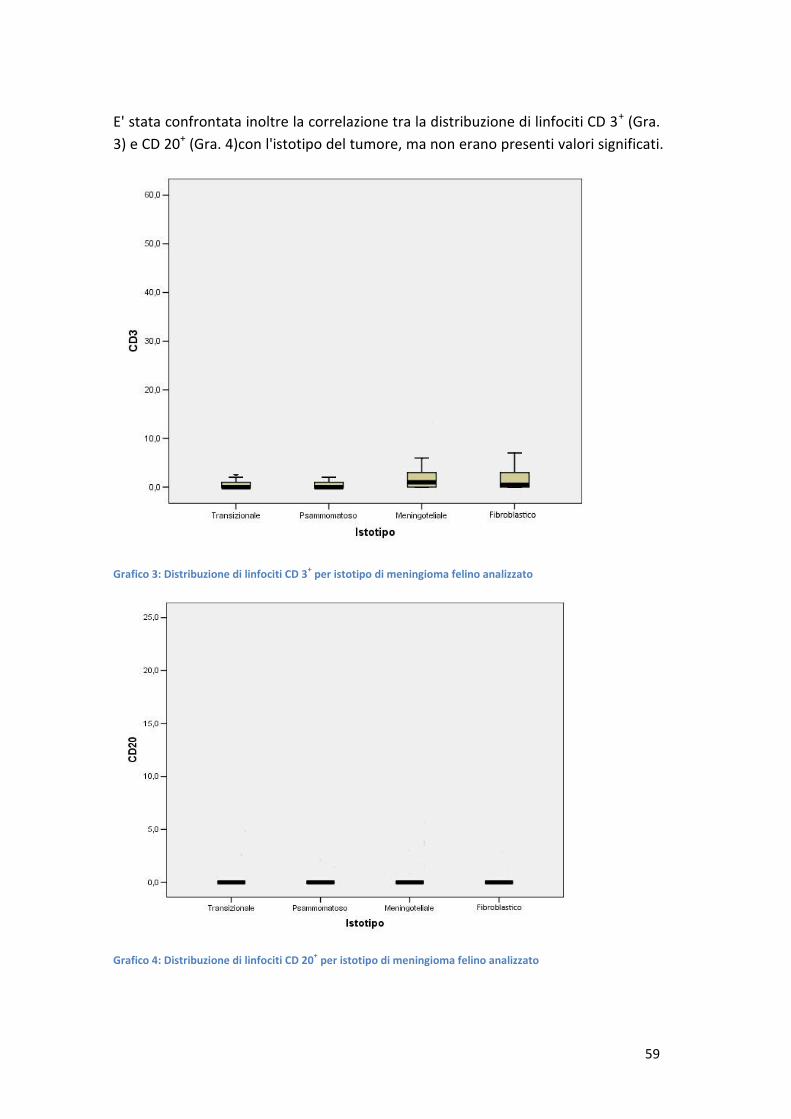
59
E' stata confrontata inoltre la correlazione tra la distribuzione di linfociti CD 3+ (Gra.
3) e CD 20+ (Gra. 4)con l'istotipo del tumore, ma non erano presenti valori significati.
Grafico 3: Distribuzione di linfociti CD 3+ per istotipo di meningioma felino analizzato
Grafico 4: Distribuzione di linfociti CD 20+ per istotipo di meningioma felino analizzato
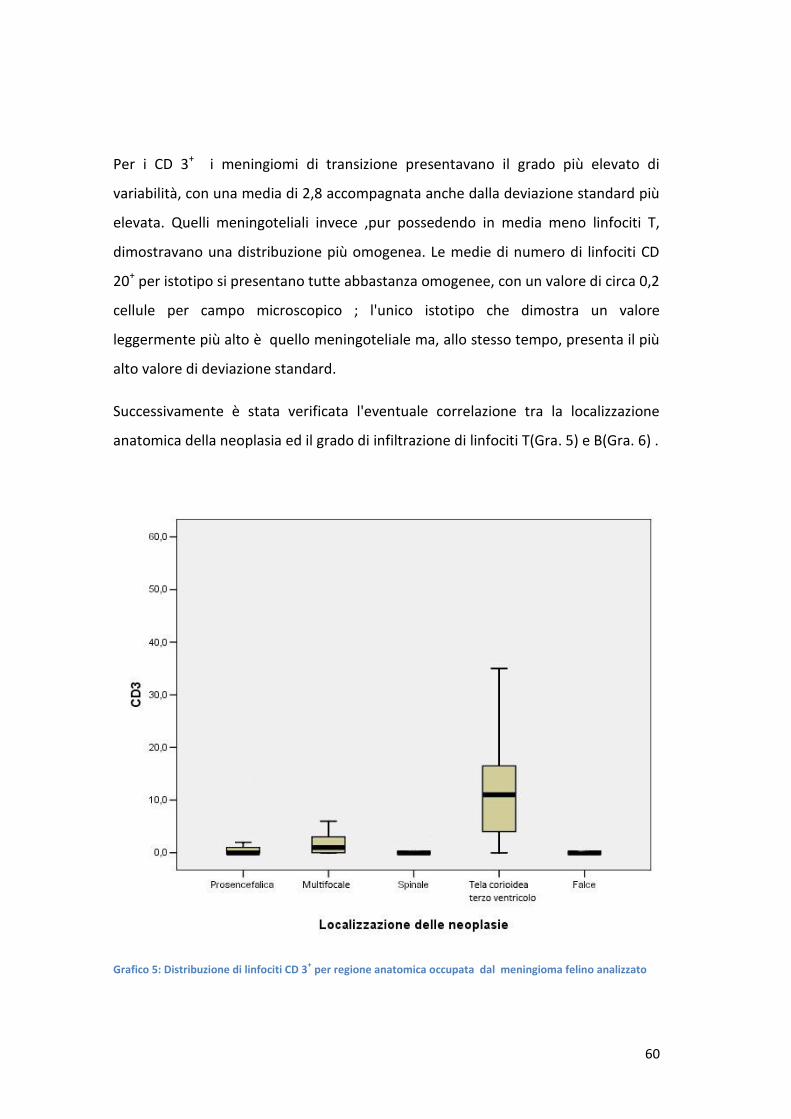
60
Per i CD 3+ i meningiomi di transizione presentavano il grado più elevato di
variabilità, con una media di 2,8 accompagnata anche dalla deviazione standard più
elevata. Quelli meningoteliali invece ,pur possedendo in media meno linfociti T,
dimostravano una distribuzione più omogenea. Le medie di numero di linfociti CD
20+ per istotipo si presentano tutte abbastanza omogenee, con un valore di circa 0,2
cellule per campo microscopico ; l'unico istotipo che dimostra un valore
leggermente più alto è quello meningoteliale ma, allo stesso tempo, presenta il più
alto valore di deviazione standard.
Successivamente è stata verificata l'eventuale correlazione tra la localizzazione
anatomica della neoplasia ed il grado di infiltrazione di linfociti T(Gra. 5) e B(Gra. 6) .
Grafico 5: Distribuzione di linfociti CD 3+ per regione anatomica occupata dal meningioma felino analizzato

61
Grafico 6: Distribuzione di linfociti CD 20+ per regione occupata dal meningioma felino analizzato
In entrambi i linfociti T e B è stata osservata una correlazione statisticamente
significativa tra l'infiltrazione linfocitica e l'insorgenza del tumore a partire dalla tela
corioidea del 3° ventricolo; le altre localizzazioni anatomica non hanno dimostrato
una simile associazione, sia per le cellule CD3+ che per quelle CD 20+.
Inoltre è stato ricercata una correlazione tra linfociti che infiltravano la periferia, sia
T (Fig 11) che B con l'istotipo (Fig. 12 ; Fig 13) e la localizzazione anatomica (Fig 14 ;
15) :

62
Figura 123 A: Linfociti CD 3+ che infiltrano la periferia di un meningioma transizionale
B: Linfociti CD 3+ che infiltrano la periferia di un meningioma meningioteliale
Grafico 7: Distribuzione del numero di cellule CD 3 presenti nella periferia del tumore , in base all'istotipo

63
Grafico 8: Distribuzione del numero di cellule CD 20+
presenti nella periferia del tumore , in base all'istotipo
Grafico 9: Distribuzione del numero di cellule CD 3+ presenti nella periferia del tumore
in base alla
localizzazione

64
Grafico 10: Distribuzione del numero di cellule CD 20+ presenti nella periferia del tumore
in base alla
localizzazione
Dai dati qui riportati non sono state verificate correlazioni statisticamente
significative tra i linfociti presenti nell'infiltrato peritumorale ed istotipo e
localizzazione anatomica. E' stato valutato anche una associazione tra questo e la
presenza di necrosi all'interno della massa, ma anche questa si è dimostrata
insignificante dal punto di vista statistico.
Discussione: Le cellule infiammatorie ricercate in questo lavoro sono state linfociti T , B e Treg. I
CD 3+ hanno dimostrato una discreta tendenza a presentarsi nell'infiltrato
infiammatorio nei meningiomi analizzati. Un simile risultato è emerso da alcuni
studi volti a caratterizzare l'infiltrato infiammatorio in meningiomi umani e canini:
infatti in uno studio sull'uomo (212) sono stati individuati nel 94% dei casi, valore
molto vicino a quello ottenuto da un analogo studio sui tumori meningei del cane
(213) . Questo trend non è affatto eccezionale, dato che i linfociti T sono
notoriamente preposti alla sorveglianza dell'organismo, riconoscendo ed

65
eliminando cellule neoplastiche potenzialmente nocive. Nell'uomo sembra che la
maggior parte di questi siano linfociti T citotossici CD 8 + mentre i T helper CD 4+
non erano praticamente presenti nell'infiltrato infiammatorio dei tumori.
Sfortunatamente non sono disponibili anticorpi rivolti contro questi 2 antigeni in
grado reagire con tessuti di cane o gatto , fissati in formalina e inclusi in paraffina .
Per quanto riguarda i B sono stati incontrati delle difficoltà con la loro rivelazione
tramite anticorpi anti-CD 79a e, per questo motivo, è stato utilizzato un altro
antigene di membrana, ossia il CD 20. Nel gatto, la media di queste cellule si è
attestata ad un livello molto basso ,sia per campioni positivi (il 51,8% dei casi non ne
presentava nemmeno uno) sia per quantità; risultati analoghi provengono da analisi
sui meningiomi umani e canini, suggerendo che i linfociti T e B si comportino nella
stessa maniera nel cane e nel gatto. Ciò nonostante, nell'uomo è riportata una
rarissima variante di meningioma , caratterizzato da un imponente infiltrato di
linfociti, plasmacellule e pochi istiociti, che costituisce un eccezione a questi risultati
(214). Dall'analisi dei campioni, i pochi linfociti B che sono stati riscontrati non
hanno dimostrato nessuna correlazione con i T. Il significato della loro presenza
all'interno dei TIL è ancora oggetto di approfonditi studi ,anche se si crede che le
cellule qua presenti esplichino attività di APC (215). Il risultato forse più interessante
che emerge da questo lavoro è la completa assenza di Treg nei meningiomi felini. Il
fatto che queste neoplasie non presentino nessuna cellula FoxP3+ nell' infiltrato
infiammatorio fa sorgere non pochi interrogativi, considerando che sono state
ritrovate in molte altre neoplasie, anche a livello encefalico (216): Treg sono stati
individuati infatti anche in meningiomi canini, se pur con una distribuzione non
correlata con il grado del tumore (213). Una possibile ragione per la completa
assenza di queste cellule immunitarie nei meningiomi felini può risiedere in 2
proteine, la CCL22 e la CCL2: queste sono delle chemochine che presentano
proprietà chemiotattiche nei confronti dei Treg (196), ed è stato osservato che la
prima nei meningiomi umani è scarsamente prodotta, mentre la seconda non viene
espressa in quelli di grado I . Inoltre è stato osservato che i Treg presenti nel tumore
esprimevano alti livelli di CCR 4, il principale recettore per la CL22, mentre non

66
esprimevano CCR 7, responsabile della convenzionale infiltrazione linfocitaria:
questa discrepanza potrebbe spiegare il motivo per cui non è stata riscontrata una
correlazione tra cellule T effettrici e Treg (217). E' plausibile quindi pensare che il
gatto possa reclutare in questi tumori Treg in una maniera analoga, giustificando
quindi la totale assenza di linfociti FoxP3+ nei meningiomi analizzati,per il fatto di
appartenere tutti al grado WHO I; il cane invece sembra non condividere queste
caratteristiche, per il fatto di presentare Treg nell'infiltrato di tumori meningei
benigni, facendo supporre che all'origine di questa differenza ci siano fattori non
considerati finora. Un altro dato che è stato raccolto con questo lavoro, è che
l'istotipo del meningioma non sembra influenzare la qualità e la quantità
dell'infiltrato infiammatorio. Bisogna premettere che nel gatto questi tipi di tumore
si presentano spesso in una forma stereotipata, in cui cioè coesistono più parti che
presentano caratteristiche morfologiche di istotipi differenti e, classificato come
"transizionale" ; a volte il meningioma può essere classificato in maniera diversa se
è diffuso prevalentemente un istotipo, ma questo tipo di distinzione non è affatto
netta. Risultati analoghi provengono da uno studio sui meningiomi canini (213).
L'unica correlazione statisticamente significativa, è stata quella della localizzazione
anatomica con il numero di linfociti infiltrati, sia T sia B : infatti i meningiomi che
originavano dalla tela corioidea del 3° ventricolo, hanno mostrato la media di
linfociti B e T più elevata. Questo risultato indica forse che la ragione di questa forte
correlazione, risiede nell'anatomia cerebrale: infatti la tela corioidea, che fa parte
della pia madre, si addossa ai plessi coroidei, ed i tumori che originano da questo
strato protrudono nel sistema ventricolare. Infatti è possibile che il microambiente
in cui si sviluppa il tumore sia differente rispetto a tutte le altre localizzazioni
anatomiche del meningioma, e questo possa influenzare di conseguenza il tipo di
cellule che lo infiltrano. In questa sede è ipotizzabile che possano essere coinvolte
anche un particolare tipo cellulare, ossia i macrofagi intraventricolari, termine che
comprende 3 tipi cellulari differenti, ossia le cellule sopraependimali, le c. "free
floating" e le c. dell'epiplesso: le prime stanno addossate all'ependima dei ventricoli,
le seconde si situano in posizione più distante dall'epitelio, mentre le ultime, che

67
costituiscono il gruppo più rappresentato dei macrofagi intraventricolari, stanno
strettamente associate all'epitelio dei plessi coriodei. Pur presentando una
localizzazione differente tra loro, queste cellule condividono le caratteristiche
immunofenotipiche, come l'immunopositività all' MHC I, MHC II , antigene comune
leucocitario ed altri antigeni, tipici di una cellula macrofagica (218). Si può ipotizzare
che una neoplasia in questa sede, possa determinare un'attivazione dei macrofagi
intraventricolari, e che, una volta attivati,siano in grado di attirare un numero
maggiore di cellule infiammatorie in grado di infiltrare la massa neoplastica. Un
altro aspetto che è stato valutato è la presenza di linfociti nell'immediata periferia
del tumore : a tal proposito, bisogna premettere che non è stato possibile
effettuare l' analisi dei linfociti periferici in tutti i campioni (solo nel 26 % dei casi
totali), dato che il tessuto nervoso periferico alla neoplasia era presente solo nei
casi di provenienza necroscopica e non chirurgica. Ciò nonostante non è stata
osservata alcuna correlazione tra il numero di linfociti peritumorali ed il numero di
quelli all'interno della massa, istotipo o localizzazione. Questo si è rivelato
sicuramente un fenomeno alquanto peculiare che ci ha fatto interrogare su quale
fosse la causa all'origine di questa discrepanza ed è stato il motivo che ha richiesto
la verifica della presenza di necrosi all'interno del tumore: infatti cellule in stato di
necrosi sono in grado di rilasciare diverse molecole ad azione flogogena, ed attrarre
quindi cellule infiammatorie. Nonostante fossero presenti alcuni microfocolai di
necrosi all'interno della neoplasia, questi non hanno dimostrato alcuna relazione
con il tipo e la quantità di linfociti infiltrati, sia all'interno della massa che nella sua
immediata periferia. Inoltre all'esame microscopico erano riconoscibili ,tra le cellule
infiltrate, neutrofili e macrofagi , fenomeno già documentato in meningiomi umani
(219) . I macrofagi, insieme alle cellule della microglia, potrebbero rivestire il ruolo
di APC ma il loro ruolo in queste malattie, è ancora non molto chiaro ed oggetto di
dibattito (220)

68
Conclusioni:
Questo studio costituisce uno dei primi tentativi di caratterizzazione i subset
linfocitari nell'infiltrato dei meningiomi felini: questi hanno dimostrato una discreta
infiltrazione di linfociti T, una esigua di B ed una totale assenza di Treg. Inoltre le
popolazioni prese in considerazione non hanno dimostrato alcuna correlazione con
linfociti peritumorali, istotipo, grado di necrosi a parte la localizzazione a livello
della tela corioidea del 3° ventricolo in cui il tumore presenta un maggior numero di
linfociti sia T che B. Il fatto che non fossero presenti cellule Treg immunopositive
comporta diversi aspetti da considerare. In primo luogo che il numero di linfociti T
regolatori non possa costituire un valore pronostico / predittivo per la malignità del
tumore; inoltre l'introduzione potenziale di un protocollo immunoterapeutico, volto
a ridurre il numero di Treg all'interno della massa neoplastica, nel meningioma
felino, non pare essere apllicabile. Questi risultati iniziano a rivelare la complessa
fisiopatologia del sistema immunitario e della sua relazione con il comportamento
biologico delle neoplasie del SNC.
Bibliografia
1. RA, Le courter. Current concepts in the diagnosis and treatment of brain tumours in dogs
and cats. 1999, 40.
2. Snyder JM, Shofer FS, Van Winkle TJ, et al. Canine intracranial primary neoplasia: 173
cases (1986-2003). Journal of veterinary internal medicine. 2006, 20, pp. 669 -675.

69
3. Troxel MT, Vite CH, Van Winkle TJ, et al. Feline intracranial neoplasia: retrospective
review of 160 cases (1985-2001). Journal of veterinaruy internal medicine. 2003, 17, pp.
850–859.
4. Josephson GK, Little PB. Four bovine meningeal tumors. The Canadian veterinary journal.
1990, 31, pp. 700-703.
5. Snyder, J.M., Lipitz, L., Skorupski, K.A., Shofer, F.S., Van Winkle, T.J. Secondary
intracranial neoplasia in the dog: 177 cases (1986–2003). Journal of Veterinary Internal
Medicine. 2008, 22, pp. 172- 177.
6. Troxel, M.T., Vite, C.H., Massicotte, C., McLear, R.C., Van Winkle, T.J., Glass, E.N., Tiches,
D., Dayrell-Hart, B. Magnetic resonance imaging features of feline intracranial neoplasia:
Retrospective analysis of 46 cats. Journal of Veterinary Internal Medicine. 2004, Vol. 18, pp.
176–189.
7. O'rahilly R, Müller G. The meninges in human development. J neuropathol Exp Neurol.
1986, Vol. 45, pp. 588-608.
8. Bagley, R.S., Silver, G.M., Gavin, P.R. Cerebellar cystic meningioma in a dog. Journal of
the American Animal Hospital Association. 2000, 36, pp. 413–415.
9. Pérez, V., Vidal, E., González, N., Benavides, J., Ferreras, M.C., Villagrasa, M., Pumarola,
M. Orbital meningioma with a granular cell component in a dog, with extracranial
metastasis. Journal of Comparative Pathology. 2005, 133, pp. 212– 217.
10. Owen, M.C., Lamb, C.R., Lu, D., Targett, M.P. Material in the middle ear of dogs having
magnetic resonance imaging for investigation of neurologic signs. Veterinary Radiology and
Ultrasound. 2004, 45, pp. 149–155.
11. Thomas R, Duke SE, Wang HJ, Breen TE, Higgins RJ, Linder KE, Ellis P, Langford CF,
Dickinson PJ, Olby NJ. et al. 'Putting our heads together': insights into genomic
conservation between human and canine intracranial tumors. J Neurooncol. 2009, 94, pp.
333–349.
12. M, Carlsson P. and Mahlapuu. Forkhead transcription factors: key players in
development and metabolism. Dev. Biol. 2002, 250, pp. 1- 23.
13. Quesnel, A.D., Parent, J.M. Paradoxical vestibular syndrome in a cat with a cerebellar
meningioma. The Canadian Veterinary Journal. 1995, 36, pp. 230–232.
14. LA, Nafe. Meningiomas in cats: a retrospective clinical study of 36 cases. J Am Vet Med
Assoc. 1979, 174, pp. 1224-1227.
15. Gordon L, Thacher C, Matthiessen D, et al. Results of craniotomy for the treatment of
cerebral meningioma in 42 cats. Vet Surg. 1994, pp. 94-100.

70
16. Forterre, F., Matis, U., Schrell, U., Geier, M., Gutmannsbauer, B., Schmahl, W.
Intracranial meningiomas e findings, therapy and results in nine cats and one dog.
Tierärztliche Praxis. 200, 28, pp. 170–177.
17. Lu, D., Pocknell, A., Lamb, C.R., Targett, M.P. Concurrent benign and malignant
multiple meningiomas in a cat: Clinical, MRI and pathological findings. Veterinary Record.
2003, 152, pp. 780–782.
18. Sturges, B.K., Dickinson, P.J., Bollen, A.W., Koblik, P.D., Kass, P.H., Kortz, G.D., Vernau,.
Magnetic resonance imaging and histological classification of intracranial meningiomas in
112 dogs. Journal of Veterinary Internal Medicine. 2008, 22, pp. 86–95.
19. Haskins ME, McGrath JT. Meningiomas in young cats with mucopolysaccharidosis I. J
neuropathol Exp Neurol.
20. Lobetti, R.G., Nesbit, J.W., Miller, D.B. Multiple malignant meningiomas in a young cat.
Journal of the South African Veterinary Association. 1997, 68, pp. 62–65.
21. Michael D. Lorenz, Joe N. Kornegay. Handbook of veterinary neurology. s.l. : Saunders,
2004. pp. 131- 154; 219 - 235 ; 368 - 375.
22. Adamo PF, Forrest L, Dubielzig R. Canine and feline meningiomas: diagnosis, treatment
and prognosis. Compend Contin Educ Pract Vet. 2004, 26, pp. 951–965.
23. RA, Le Courter. Brain tumors in dogs and cats: Diagnosis and management. Vet Med
Report. 1990, 2, pp. 332-342.
24. Braund K, Ribas J. Central nervous system meningiomas. Compend ContinEduc Pract Vet.
1986, 8, pp. 241–248.
25. P, Luttgen. Common Neurologic Problems. Philadelphia : Saunders, 1988. pp. 713–715.
26. Pieper, D.R., Al-Mefty, O., Hanada, Y., Buechner, D. Hyperostosis associated with
meningioma of the cranial base: Secondary changes or tumor invasion. Neurosurgery. 1999,
44, pp. 742–746.
27. Mercier, M., Heller, HL., Bischoff, MG., Looper, J., Bacmeister, C.X. Imaging diagnosis—
Hyperostosis associated with meningioma in a dog. Veterinary Radiology and Ultrasound.
2007, 48, pp. 421–423.
28. Gutierrez-Quintana, R., Gunn-Moore, D.A., Lamm, C.G., Penderis, J. Feline intracranial
meningioma with skull erosion and tumour extension into an area of skull hyperostosis.
Journal of Feline Medicine and Surgery. 2011, 13, pp. 296–299.
29. Falzone, C., Baroni, M., De Lorenzi, D., Mandara, M.T. Toxoplasma gondii brain
granuloma in a cat: Diagnosis using cytology from an intraoperative sample and sequential
magnetic resonance imaging. Journal of Small Animal Practice. 2008, 49, pp. 95–99.

71
30. Sykes, J.E., Sturges, B.K., Cannon, M.S., Gericota, B., Higgins, R.J., Trivedi, S.RDickinson,
P.J., Vernau, K.M., Meyer, W., Wisner. Clinical signs, imaging features, neuropathology,
and outcome in cats and dogs with central nervous system cryptococcosis from California.
Journal of Veterinary Internal Medicine. 2010, 24, pp. 1427–1438.
31. Moissonier P, Blot S, Beuvon F, et al. Steretactic CT-guided brain biopsy in the dog. J
Small Anim Pract. 2002, pp. 115–123.
32. Mawrin C, Perry A. Pathological classification and molecular genetics of meningiomas. J
Neurooncol. 2010, 99, pp. 379-91.
33. Perry A, Stafford SL, Scheithauer BW, Suman VJ, Lohse CM. Meningioma grading: an
analysis of histologic parameters. Am J Surg Pathol. 1997, 21, pp. 1455–1465.
34. Burger, P.C., Scheithauer, B.W., Vogel, F.S. Surgical Pathology of the Nervous System
and its Coverings. New York : s.n., 2002. pp. 49–71.
35. E. Kaldrymidou, Z. S. Polizopoulou. Papillary Meningioma in the Cerebellum of a Cat. J.
Comp. Path. 2000, 123, pp. 222–225 .
36. A comparative study of canine and feline meningioma classification based on the WHO
histological classification system in humans. Mandara, M.T., Pavone, S., Brunetti, B.,
Mandrioli, L. Bologna : s.n., 2009. Proceedings of the 22nd Symposium ESVNECVN.
37. Kepes, J.J. residential address: The histopathology of meningiomas. A reflection of
origins and expected behavior? Journal of Neuropathology and Experimental Neurology.
1986, 45, pp. 95–107.
38. Dickinson PJ, Surace EI, Cambell M, et al. Expression of the tumor suppressor genes
NF2, 4.1B, and TSLC1 in canine meningiomas. Vet Pathol. 2009, 46, pp. 884–892.
39. Louis DN, Ramesh V, Gusella JF. Neuropathology and molecular genetics of
neurofibromatosis 2 and related tumors. Brain Pathol. 1995, 5, pp. 163–172.
40. Kros J, de Greve K, van Tilborg A, Hop W,Pieterman H, Avezaat C, Lekanne Dit Deprez
R, Zwarthoff E. NF2 status of meningiomas is associated with tumour localization and
histology. J Pathol. 2001, 194, pp. 367–372.
41. Perry A, Cai DX, Scheithauer BW, Swanson PE, Lohse CM, Newsham IF, Weaver A,
Gutmann DH. Merlin, DAL-1, and progesterone receptor expression in clinicopathologic
subsets of meningioma: a correlative immunohistochemical study of 175 cases. J
Neuropathol Exp Neurol. 2000, 59, pp. 872–879.
42. Surace EI, Lusis E, Murakami Y, Scheithauer BW, Perry A, Gutmann DH. Loss of tumor
suppressor in lung cancer-1 (TSLC1) expression in meningioma correlates with increased
malignancy grade and reduced patient survival. J Neuropathol Exp Neurol. 2004, 63, pp.
1015–1027.

72
43. Dobson JM, Samuel S, Milstein H, Rogers K, Wood JL. Canine neoplasia in the UK:
estimates of incidence rates from a population of insured dogs. J Small Anim Pract. 2002, 43,
pp. 240–246.
44. Speciale, J., Koffman, B., Bashirelahi, N. and Steinberg, S. Identification of gonadal
steroid receptors in meningiomas from dogs and cats. American Journal of Veterinary
Research. 1990, 51, pp. 833-835.
45. Adamo, P.F., Cantile, C., Steinberg, H. Evaluation of progesterone and estrogen
receptor expression in 15 meningiomas of dogs and cats. American Journal of Veterinary
Research. 2003, 64, pp. 1310–1318.
46. Mandara, M.T., Ricci, G., Rinaldi, L., Sarli, G., Vitellozzi, G. Immunohistochemical
identification and image analysis quantification of oestrogen and progesterone receptors in
canine and feline meningioma. Journal of Comparative Pathology. 2002, 127, pp. 214–218.
47. Theon, A. P., Lecouter, R. A., Carr, E. A. and Griffey, S. M. Influence of tumour cell
proliferation and sexhormone receptors on effectiveness of radiation therapy for dogs with
incompletely resected meningiomas. Journal of the American Veterinary Medical
Association. 2000, 216, pp. 701-707.
48. Ramos-Vara JA, Miller MA, Gilbreath E, Patterson JS. Immunohistochemical detection
of CD34, E-cadherin, claudin-1, glucose transporter 1, laminin, and protein gene product 9.5
in 28 canine and 8 feline meningiomas. Vet Pathol. 2010. 2010, 47, pp. 725-37.
49. Theaker JM, Gatter KC, Esiri MM, Fleming KA. Epithelial membrane antigen and
cytokeratin expression by meningiomas: an immunohistochemical study. J Clin Pathol. 1986,
39, pp. 435–439.
50. Koestner A, Higgins RJ. Tumors in Domestic Animals. Ames : Meuten DJ, 2002.
51. Jacque CM, Vinner C, Kujas M, Raoul M, Racadot J, Baumann NA. Determination of
glial fibrillary acidic protein (GFAP) in human brain tumors. J. Neurol. Sci. 1978, 35, pp. 147–
55.
52. Scholzen T, Gerdes J. The Ki-67 protein: from the known and the unknown. J. Cell.
Physiol. 2000, 182, pp. 311–22.
53. Wolf R, Voscopoulos CJ, FitzGerald PC, et al. he mouse S100A15 ortholog parallels
genomic organization, structure, gene expression, and protein-processing pattern of the
human S100A7/A15 subfamily during epidermal maturation. J. Invest. Dermatol. 2006, 126,
pp. 1600–8.
54. Leong, Anthony S-Y, Cooper, Kumarason and Leong, F Joel W-M. Manual of Diagnostic
Cytology. s.l. : Greenwich Medical Media, 2003.

73
55. Artlich A, Scmidt D. Immunohistochemical profile of meningiomas and their histological
subtypes. Hum Pathol. 1990, 21, pp. 843–849.
56. Barnhart KF, Wojcieszyn J, Storts RW. Immunohistochemical staining patterns of canine
meningiomas and correlation with published immunophenotypes. Vet Pathol. 2002, 39, pp.
311–321.
57. Swisshelm K, Macek R, Kubbies M. Role of claudins in tumorigenesis. Adv. Drug Deliv.
Rev. 2005, 57, pp. 919–28.
58. ruz PW, Mueckler MM. Structural analysis of the GLUT1 facilitative glucose transporter.
Mol. Membr. Biol. 2002, 18, pp. 183–93.
59. North PE, Waner M, Mizeracki A, Mihm MC Jr. GLUT1: a newly discovered
immunohistochemical marker for juvenile hemangiomas. Hum. Pathol. 2000, 31, pp. 11–22.
60. al., Timpl R et. Laminin – a glycoprotein from basement membranes. J Biol Chem. 1979,
254, pp. 9933–7.
61. Petz M, Them N, Huber H, Beug H, Mikulits W. La enhances IRES-mediated translation
of laminin B1 during malignant epithelial to mesenchymal transition. Nucleic Acids Research.
2011, 39, pp. 01–13.
62. Doran JF, Jackson P, Kynoch PA, Thompson RJ. Isolation of PGP 9.5, a new human
neurone-specific protein detected by high-resolution two-dimensional electrophoresis. J
Neurochem. 1983, 40, pp. 1542-7.
63. Bhattacharjee M, Adesina AM, Goodman C, Powell S. Claudin-1 expression in
meningiomas and schwannomas: Possible role in differential diagnosis. J Neuropathol Exp
Neurol. 2003, 62, p. 581.
64. G, Brugal. Interpretation of proliferation markers. 1994. pp. 234–239.
65. Mandrioli L, Panarese S, Cesari A, Mandara MT, Marcato PS, Bettini G.
Immunohisitochemical expression of h-telomerasereverse transcriptase in canine and feline
meningiomas. J Vet Sci. 2007, 8, pp. 111–115.
66. Chieco P, Pagnoni M, Romagnoli E, Melchiorri C. A rapid and simple staining method,
using toluidine blue, for analysing mitotic figures in tissue sections. Histochem J. 1993, 25,
pp. 569-577.
67. Maes L, Kalala JPO, Cornelissen R, De Ridder L. Progression of astrocytomas and
meningiomas: an evaluation in vitro. Cell Prolif. 2007, 40, pp. 14-23.
68. Liotta LA, Thorgeirsson UP, Garbisa S. Role of collagenases in tumor cell invasion.
Cancer and metastis review. 1982, 1, pp. 277 - 288.

74
69. Stamenkovic, Ivan. Matrix metalloproteinases in tumor invasion and metastasis. Cancer
biology. 2000, 10, pp. 415 - 433.
70. Mandara MT, Pavone S, Mandrioli L, Bettini G, Falzone C, Baroni M. Matrix
metalloproteinase-2 and matrix metalloproteinase-9 expression in canine and feline
meningioma. Vet Pathol. 2009, 46, pp. 836-45.
71. Okada M, Miyake K, Matsumoto Y, Kawai N. Matrix metalloproteinase-2 and matrix
metalloproteinase-9 expression correlate with the recurrence of intracranial meningiomas.
Journal of neuro-oncology. 2004, 66, pp. 29 - 37.
72. Biotechnology, Santa Cruz. CD 3 antibody / CD 3 antibodies | Santa Cruz Biotech.
[Online] http://www.scbt.com/table-cd3.html.
73. —. CD 79 antibody / CD 79 antibodies | Santa Cruz Biotech. [Online]
http://www.scbt.com/table-cd79.html.
74. —. CD 20 antibody / CD 20 antibodies | Santa Cruz Biotech. [Online]
http://www.scbt.com/table-cd20.html.
75. Weigel, D., Jurgens, G., Kuttner, F., Seifert, E., and Jackle, H. The homeotic gene fork
head encodes a nuclear protein and is expressed in the terminal regions of the Drosophila
embryo. Cell. 1989, 57, pp. 645 - 658.
76. Brunkow, M.E., Jeffery, E.W., Hjerrild, K.A., Paeper, B., Clark, L.B., Yasayko, S.A.,
Wilkinson, J.E., Galas, D., Ziegler, S.F., and Ramsdell, F. Disruption of a new
forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of
the scurfy mouse. Nat. Genet. 2001, 27, pp. 68–73.
77. Ochs, H.D., Ziegler, S.F., and Torgerson, T.R. FOXP3 acts as a rheostat of the immune
response. Immunol. Rev. 2005, 203, pp. 156–164.
78. Sakaguchi, S. Regulatory T cells: key controllers of immunologic self-tolerance. Cell.
2000, 5, pp. 455–458.
79. Sakaguchi S, Ono M, Setoguchi R, Yagi H, Hori S, Fehervari Z, Shimizu J, Takahashi T,
Nomura T. FoxP3+ CD25+ CD4+ natural regulatory T-cells in dominant self-tolerance and
autoimmune disease. Immunol Rev. 2006, 212, pp. 8–27.
80. Fontenot JD, Dooley JL, Farr AG, Rudensky AY. Developmental regulation of Foxp3
expression during ontogeny. J. Exp. Med. 2005, 202, pp. 901–6.
81. Xiaomei Yuan, Thomas R. Malek. Cellular and molecular determinants for the
development of natural. Human Immunology. 2012, 73, pp. 773–782.
82. Abbas, Lichtmann. Cellular and Molecular Immunology. Philadelphia : saunders, 2003.

75
83. Tai X, Cowan M, Feigenbaum L, Singer A. CD28 costimulation of developing thymocytes
induces Foxp3 expression and regulatory T cell differentiation independently of interleukin
2. Nat. Immunol. 2005, 6, pp. 152–62.
84. Setoguchi, R., Hori, S., Takahashi, T., and Sakaguchi, S. Homeostatic maintenance of
natural Foxp3+ CD25+ CD4+ regulatory T cells by interleukin (IL)-2 and induction of
autoimmune disease by IL-2 neutralization. J. Exp. Med. 2005, 201, pp. 723–735.
85. Marie JC, Letterio JJ, Gavin M, Rudensky AY. TGF-b1 maintains suppressor function and
Foxp3 expression in CD4+CD25+ regulatory T cells. J. Exp. Med. 2005, 201, pp. 1061–7.
86. Liu Y, Zhang P, Li J, Kulkarni AB, Perruche S, Chen W. A critical function for TGF-b
signaling in the development of natural CD4+CD25+Foxp3+ regulatory T cells. Nat. Immunol.
2008, 09, pp. 632–40.
87. Curotto de Lafaille MA, Kutchukhidze N, Shen S, Ding Y, Yee H, Lafaille JJ. Adaptive
Foxp3+ regulatory T cell-dependent and -independent control of allergic inflammation.
Immunity. 2008, 29, pp. 114–26.
88. Turner MS, Kane LP, Morel PA. Dominant role of antigen dose in CD4+Foxp3+
regulatory T cell induction and expansion. J. Immunol. 2009, 183, pp. 4895–903.
89. Kretschmer K, Apostolou I, Hawiger D, Khazaie K, Nussenzweig MC, von Boehmer H.
Inducing and expanding regulatory T cell populations by foreign antigen. Nat. Immunol.
2005, 6, pp. 1219–27.
90. Gottschalk RA, Corse E, Allison JP. TCR ligand density and affinity determine peripheral
induction of Foxp3 in vivo. J. Exp. Med. 2010, 207, pp. 1701–11.
91. Semple K, Nguyen A, Yu Y, Wang H, Anasetti C, Yu XZ. le K, Nguyen A, Yu Y, Wang H,
Anasetti C, Yu XZ. Strong CD28 costimulation suppresses induction of regulatory T cells from
naive precursors through Lck signaling. Blood. 2011, 117, pp. 3096–10.
92. Davidson TS, DiPaolo RJ, Andersson J, Shevach EM. Cutting Edge: IL-2 is essential for
TGF-b-mediated induction of Foxp3+ T regulatory cells. J. Immunol. 2007, 178, pp. 4022–6.
93. Benson MJ, Pino-Lagos K, Rosemblatt M, Noelle RJ. All-trans retinoic acid mediates
enhanced T reg cell growth, differentiation, and gut homing in the face of high levels of co-
stimulation. J. Exp. Med. 2007, 204, pp. 1765–74.
94. Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, et al. Small intestine
lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic
acid. J. Exp. Med. 2007, 204, pp. 1775–85.
95. Ivanov II. Frutos Rde L, Manel N, Yoshinaga K, Rifkin DB, Sartor RB, et al. Specific
microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the
small intestine. Cell Host Microbe. 2008, 4, pp. 337–49.

76
96. Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, et al. Induction of
colonic regulatory T cells by indigenous Clostridium species. Science. 2011, 331, pp. 337–41.
97. Round JL, Mazmanian SK. Inducible Foxp3+ regulatory T-cell development by a
commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. 2010, 107, pp.
12204–9.
98. Hall JA, Bouladoux N, Sun CM, Wohlfert EA, Blank RB, Zhu Q, et al. Commensal DNA
limits regulatory T cell conversion and is a natural adjuvant of intestinal immune responses.
Immunity. 2008, 29, pp. 637–49.
99. Francisco LM, Salinas VH, Brown KE, Vanguri VK, Freeman GJ, Kuchroo VK, et al. PD-L1
regulates the development, maintenance, and function of induced regulatory T cells. J Exp
Med. 2009, 206, pp. 3015–29.
100. Gao W, Lu Y, El Essawy B, Oukka M, Kuchroo VK, Strom TB. Contrasting effects of
cyclosporine and rapamycin in de novo generation of alloantigen-specific regulatory T cells.
Am J Transplant. 2007, 7, pp. 1722–32.
101. Kendal AR, Chen Y, Regateiro FS, Ma J, Adams E, Cobbold SP, et al. Sustained
suppression by Foxp3_regulatory T cells is vital for infectious transplantation tolerance. J
Exp Med. 2011, 208, pp. 2043–53.
102. Ochando JC, Homma C, Yang Y, Hidalgo A, Garin A, Tacke F, et al.
Alloantigenpresenting plasmacytoid dendritic cells mediate tolerance to vascularized grafts.
Nat Immuno. 2006, 07, pp. 652–62.
103. Ye Zheng Steven Josefowicz Ashutosh Chaudhry Xiao P. Peng Katherine Forbush,
Alexander Y. Rudensky,. Role of conserved non-coding DNA elements in the Foxp3 gene in
regulatory T-cell fate. Nature. 2010, 463, pp. 808–812.
104. Molinero LL, Yang J, Gajewski T, Abraham C, Farrar MA, Alegre ML. CARMA1 controls
an early checkpoint in the thymic development of FoxP3+ regulatory T cells. J. Immunol.
2009, 182, pp. 6736–43.
105. Long M, Park SG, Strickland I, Hayden MS, Ghosh S. Nuclear factor-jB modulates
regulatory T cell development by directly regulating expression of Foxp3 transcription
factor. Immunity. 2009, 31, pp. 921–31.
106. Fu G, Chen Y, Yu M, Podd A, Schuman J, He Y, et al. Phospholipase C{c}1 is essential
for T cell development, activation, and tolerance. J. Exp. Med. 2010, 207, pp. 309–18.
107. Sauer S, Bruno L, Hertweck A, Finlay D, Leleu M, Spivakov M, et al. T cell receptor
signaling controls Foxp3 expression via PI3K, Akt, and mTOR. Proc. Natl. Acad. Sci. 2008,
105, pp. 7797–802.

77
108. Ouyang W, Beckett O, Ma Q, Paik JH, DePinho RA, Li MO. Foxo proteins cooperatively
control the differentiation of Foxp3+ regulatory T cells. Nat. Immunol. 2010, 11, pp. 618–27.
109. Tone Y, Furuuchi K, Kojima Y, Tykocinski ML, Greene MI, Tone M. Smad3 and NFAT
cooperate to induce Foxp3 expression through its enhancer. Nat. Immunol. 2008, 9, pp.
194–202.
110. Josefowicz SZ, Wilson CB, Rudensky AY. Cutting edge: TCR stimulation is sufficient for
induction of Foxp3 expression in the absence of DNA methyltransferase 1. J. Immunol. 2009,
182, pp. 6648–52.
111. Lu L, Wang J, Zhang F, Chai Y, Brand D, Wang X, et al. Role of SMAD and non- SMAD
signals in the development of Th17 and regulatory T cells. J. Immunol. 2010, 184, pp. 4295–
306.
112. Mantel PY, Kuipers H, Boyman O, Rhyner C, Ouaked N, Ruckert B, et al. GATA3-driven
Th2 responses inhibit TGF-b1-induced FOXP3 expression and the formation of regulatory T
cells. PLoS Biol. 2007, 5, p. 329.
113. Bi Y, Yang R. Direct and indirect regulatory mechanisms in TH17 cell differentiation
and functions. Scand J Immunol. 2012, 75, pp. 543-52.
114. Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFb in the context of
an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells.
Immunity. 2006, 24, pp. 179–89.
115. Coombes JL, Siddiqui KR, Arancibia-Carcamo CV, Hall J, Sun CM, Belkaid Y, et al. A
functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T
cells via a TGF-b and retinoic acid-dependent mechanism. J. Exp. Med. 2007, 204, pp. 1757–
64.
116. Yang XP, Ghoreschi K, Steward-Tharp SM, Rodriguez-Canales J, Zhu J,Grainger JR, et
al. Opposing regulation of the locus encoding IL-17 through direct, reciprocal actions of
STAT3 and STAT5. Nat. Immunol. 2011, 12, pp. 247–54.
117. Groux, H., O’Garra, A., Bigler, M., Rouleau, M., Antonenko, S., de Vries, J.E., and
Roncarolo, M.G. A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents
colitis. Nature. 1997, 389, pp. 737–742.
118. Vieira, P.L., Christensen, J.R., Minaee, S., O’Neill, E.J., Barrat, F.J., Boonstra, A.,
Barthlott, T., Stockinger, B., Wraith, D.C., and O’Garra, A. IL- 10-secreting regulatory T cells
do not express Foxp3 but have comparable regulatory function to naturally occurring
CD4+CD25+ regulatory T cells. J. Immunol. 2004, 172, pp. 5986–5993.
119. Xu L, Kitani A, Fuss I, Strober W. utting edge: regulatory T cells induce CD4+
CD25−Foxp3− T cells or are self-induced to become Th17 cells in the absence of exogenous

78
TGF-beta. utting edge: regulatory T cells induce CD4+ CD25−Foxp3− T cells or are self-
induced to become Th17 cells in the absence of exogenous TGF-beta. 2007, 178, pp. 6725–9.
120. Zhou L, Lopes JE, Chong MM, Ivanov II, Min R, Victora GD, et al. TGF-beta-induced
Foxp3 inhibits T(H)17 cell differentiation by antagonizing RORgammat function. Nature.
2008, 453, pp. 236–40.
121. Koenen HJ, Smeets RL, Vink PM, van Rijssen E, Boots AM, Joosten I. Human
CD25highFoxp3pos regulatory T cells differentiate into IL-17-producing cells. Blood. 2008,
112, pp. 2340–52.
122. Osorio F, LeibundGut-Landmann S, LochnerM, Lahl K, Sparwasser T, Eberl G, et al. DC
activated via dectin-1 convert Treg into IL-17 producers. Eur J Immunol. 2008, 38, pp. 3274–
81.
123. Tsuji M, Komatsu N, Kawamoto S, Suzuki K, Kanagawa O, Honjo T, et al. Preferential
generation of follicular B helper T cells from Foxp3+ T cells in gut Peyer’s patches. Science.
2009, 323, pp. 1488–92.
124. Zhou X, Bailey-Bucktrout SL, Jeker LT, Penaranda C, Martinez-Llordella M, Ashby M,
et al. Instability of the transcription factor Foxp3 leads to the generation of pathogenic
memory T cells in vivo. Nat Immunol . 2009, 10.
125. Viglietta V, Baecher-Allan C, Weiner HL, Hafler DA. Loss of functional suppression by
CD4+CD25+ regulatory T cells in patients with multiple sclerosis. J Exp Med. 2004, 199, pp.
971–9.
126. Fontenot JD, Rasmussen JP, Williams LM, Dooley JL, Farr AG, Rudensky AY.
Regulatory T-cell lineage specification by the forkhead transcription factor FoxP3. Immunity.
2005, 22, pp. 329–341.
127. Liu W, Putnam AL, Xu-Yu Z, Szot GL, Lee MR, Zhu S, Gottlieb PA, Kapranov P,
Gingeras TR, Fazekas de St Groth B, Clayberger C, Soper DM, Ziegler SF, Bluestone JA.
CD127 expression inversely correlates with FoxP3 and suppressive function of human CD4+
Treg cells. J Exp Med. 2006, 203, pp. 1701–1711.
128. Hori, S. et al. Control of regulatory T cell development by the transcription factor
Foxp3. Science. 2003, 299, pp. 1057–1061.
129. Wu Y, Borde M, Heissmeyer V, et al. FOXP3 controls regulatory T cell function through
cooperation with NFAT. Cell. 2006, 126, pp. 375-387.
130. Bettelli, E. et al. oxp3 interacts with nuclear factor of activated T cells and NF-kB to
repress cytokine gene expression and effector functions of T helper cells. Proc. Natl. Acad.
Sci. 2005, 102, pp. 5138– 5143.

79
131. Marson A, Kretschmer K, Frampton GM, et al. Foxp3 occupancy and regulation of key
target genes during T-cell stimulation. Nature. 2007, 445, pp. 931-935.
132. Zheng Y, Josefowicz SZ, Kas A, et al. Genome-wide analysis of Foxp3 target genes in
developing and mature regulatory T cells. Nature. 2007, 445, pp. 936-940.
133. Grant, C. et al. Foxp3 represses retroviral transcription by targeting both NF-kB and
CREB pathways. PLoS Pathog. 2006, 2.
134. Thornton AM, Korty PE, Tran DQ,Wohlfert EA,Murray PE, Belkaid Y, et al. Expression
of Helios, an Ikaros transcription factor family member, differentiates thymic-derived from
peripherally induced Foxp3+T regulatory cells. J Immunol. 2010, 184, pp. 3433–41.
135. Georgopoulos K, Moore DD, Derfler B. Ikaros, an early lymphoid-specific transcription
factor and a putative mediator for T cell commitment. Science. 1992, 258, pp. 808–12.
136. Gottschalk RA, Corse E, Allison JP. Expression of Helios in peripherally induced Foxp3+
regulatory T cells. J Immunol. 2012, 188, pp. 976–80.
137. Getnet D, Grosso JF, Goldberg MV, Harris TJ, Yen HR, Bruno TC, et al. A role for the
transcription factor Helios in human CD4(+)CD25(+) regulatory T cells. Mol Immunol. 2010,
47, pp. 1595–600.
138. Takahashi, T. et al. Immunologic self-tolerance maintained by CD25+CD4+ naturally
anergic and suppressive T cells: induction of autoimmune disease by breaking their
anergic/suppressive state. Int. Immunol. 1998, 10, pp. 1969-1980.
139. Sakaguchi, Miyara and S. Natural regulatory T cells: mechanisms of suppression.
Trends in Molecular Medicine. 2007, 3, pp. 108–116.
140. Munn, A. L. Mellor and D. H. IDO expression by dendritic cells: tolerance and
tryptophan catabolism. Nature Reviews Immunology. 2004, 10, pp. 762–774.
141. W. J. Grossman, J. W. Verbsky, B. L. Tollefsen, C. Kemper, J. P. Atkinson, and T. J. Ley.
Differential expression of granzymes A and B in human cytotoxic lymphocyte subsets and T
regulatory cells. Blood. 2004, 104, pp. 2840–2848.
142. W. J. Grossman, J. W. Verbsky, W. Barchet, M. Colonna, J. P. Atkinson, and T. J. Ley.
Human T regulatory cells can use the perforin pathway to cause autologous target cell
death. Immunity. 2004, 21, pp. 589–601.
143. Huang, C.T. et al. Role of LAG-3 in regulatory T cells. Immunity. 2004, 21, pp. 503–513.
144. Kingsley, C.I. et al. CD25+CD4+ regulatory T cells prevent graft rejection: CTLA-4- and
IL-10-dependent immunoregulation of alloresponses. J. Immunol. 2002, 168, pp. 1080–1086.
145. Kryczek, I. et al. Cutting edge: induction of B7-H4 on APCs through IL-10: novel
suppressive mode for regulatory T cells. J. Immunol. 2006, 177, pp. 40–44.

80
146. Ghiringhelli, F. et al. CD4+CD25+ regulatory T cells inhibit natural killer cell functions in
a transforming growth factor-bdependent manner. J. Exp. Med. 2005, 202, pp. 1075–1085.
147. J. R. Grainger, K. A. Smith, J. P. Hewitson et al. Helminth secretions induce de novo T
cell Foxp3 expression and regulatory function through the TGF-β pathway. Journal of
Experimental Medicine. 2010, 11, pp. 2331–2341.
148. Weiden PL, Storb R, Tsoi MS, Graham TC, Lerner KG, Thomas ED. Infusion of donor
lymphocytes into stable canine radiation chimeras: implications for mechanism of
transplantation tolerance. J Immunol. 1976, 116, pp. 1212–9.
149. Sacchini F, Pinheiro DY, Singh Y, Tisato V, Armitage-Chan E, Cunningham F, et al.
Characterising regulatory T cells in healthy dogs: preliminary studies. Immunology. 2007.
150. O'Neill K, Guth A, Biller B, Elmslie R, Dow S. Changes in regulatory T cells in dogs with
cancer and associations with tumor type. J Vet Intern Med. 2009, 23, pp. 875–81.
151. Rissetto KC, Rindt H, Selting KA, Villamil JA, Henry CJ, Reinero CR. loning and
expression of canine CD25 for validation of an anti-human CD25 antibody to compare T
regulatory lymphocytes in healthy dogs and dogs with osteosarcoma. Vet Immunol
Immunopathol. 2010, 135, pp. 137–45.
152. Langweiler M, Cockerell GL. Generation of concanavalin A-induced suppressor cells in
the cat. Int Arch Allergy Appl Immunol. 1982, 69, pp. 148–55.
153. Lankford S, Petty C, LaVoy A, Reckling S, Tompkins W, Dean GA. Cloning of feline
FOXP3 and detection of expression in CD4+CD25+ regulatory T cells. Vet Immunol
Immunopathol. 2008, 122, pp. 159–66.
154. Vahlenkamp TW, Tompkins MB, Tompkins WA. Feline immunodeficiency virus
infection phenotypically and functionally activates immunosuppressive CD4+ CD25+ T
regulatory cells. J Immunol. 2004, 172, pp. 4752–61.
155. Mexas AM, Fogle JE, Tompkins WA, Tompkins MB. CD4+CD25+ regulatory T cells are
infected and activated during acute FIV infection. Vet Immunol Immunopathol. 2008, 126,
pp. 263–72.
156. Petty CS, Tompkins MB, Tompkins WA. Transforming growth factor-beta/
transforming growth factor-betaRII signaling may regulate CD4+CD25+ Tregulatory cell
homeostasis and suppressor function in feline AIDS lentivirus infection. J Acquir Immune
Defic Syndr. 2008, 47, pp. 148–60.
157. Smithberg SR, Fogle JE, Mexas AM, Reckling SK, Lankford SM, Tompkins MB, et al. In
vivo depletion of CD4+CD25+ regulatory T cells in cats. J Immunol Methods. 2008, 329, pp.
81–91.

81
158. Mikkelsen SR, Reckling SK, Egan EA, Dean GA. In vivo depletion of CD4+CD25hi
regulatory T cells is associated with improved antiviral responses in cats chronically infected
with feline immunodeficiency virus. Virology. 2010, 403, pp. 163–72.
159. M. Stumpf, A.Hasenburg,M.O. Riener et al. Intraepithelial CD8−positive T
lymphocytes predict survival for patients with serous stage III ovarian carcinomas:
relevance of clonal selection of T lymphocytes. British Journal of Cancer. 2009, 9, pp. 1513–
1521.
160. H. T. Khong, Q. J.Wang, and S. A. Rosenberg. Identification of multiple antigens
recognized by tumor-infiltrating lymphocytes from a single patient: tumor escape by
antigen loss and loss ofMHC expression. Journal of Immunotherapy. 2004, 3, pp. 184–190.
161. D. J. Hicklin, Z. Wang, F. Arienti, L. Rivoltini, G. Parmiani, and S. Ferrone. 2-
Microglobulin mutations, HLA class I antigen loss, and tumor progression in melanoma.
Journal of Clinical Investigation. 1998, 12, pp. 2720–2729.
162. E. Fonsatti, L. Sigalotti, S. Coral, F. Colizzi, M. Altomonte, and M. Maio. Methylation-
regulated expression of HLA class I antigens in melanoma. International Journal of Cancer.
2003, 3, pp. 430–431.
163. Seliger, B. Molecular mechanisms of MHC class I abnormalities and APM components
in human tumors. Cancer Immunology, Immunotherapy. 2008, 11, pp. 1719–1726.
164. T. Rodrìguez, R. Mèndez, A. Del Campo et al. Distinct mechanisms of loss of IFN-
gamma mediated HLA class I inducibility in two melanoma cell lines. BMC Cancer. 2007, Vol.
7.
165. C. Weishaupt, K. N. Munoz, E. Buzney, T. S. Kupper, and R. C. Fuhlbrigge. T-cell
distribution and adhesion receptor expression in metastatic melanoma. Clinical Cancer
Research. 2007, 9, pp. 2549–2556.
166. J. P. Medema, J. de Jong, L. T. C. Peltenburg et al. Blockade of the granzyme
B/perforin pathway through overexpression of the serine protease inhibitor PI-9/SPI-6
constitutes a mechanism for immune escape by tumors. Proceedings of the National
Academy of Sciences of the U.S.A. 2001, 20, pp. 11515–11520.
167. G. J. Ullenhag, A. Mukherjee, N. F. S. Watson, A. H. Al- Attar, J. H. Scholefield, and L.
G. Durrant. Overexpression of FLIPL is an independent marker of poor prognosis in
colorectal cancer patients. Clinical Cancer Research. 2007, 17, pp. 5070–5075.
168. P. Moller, K. Koretz, F. Leithauser et al. Expression of APO- 1 (CD95), a member of the
NGF/TNF receptor superfamily, in normal and neoplastic colon epithelium. International
Journal of Cancer. 1994, 3, pp. 371–377.
169. J. Cheng, T. Zhou, C. Liu et al. Protection from Fasmediated apoptosis by a soluble
form of the Fas molecule. Science. 1994, 5154, pp. 1759–1762.

82
170. Chen, Zou and L. Inhibitory B7-family molecules in the tumour microenvironment.
Nature Reviews Immunology. 2008, 6, pp. 467–477.
171. M. G. di Bari, M. E. Lutsiak, S. Takai et al.,. TGF-beta modulates the functionality of
tumor-infiltrating CD8+ T cells through effects on TCR signaling and Spred1 expression.
ancer Immunology, Immunotherapy. 2009, 11, pp. 1809–1818.
172. Massaguè, D. A. Thomas and J. TGF-β directly targets cytotoxic T cell functions during
tumor evasion of immune surveillance. Cancer Cell. 2005, 5, pp. 369–380.
173. B. H. A. von Rahden, H. J. Stein, M. Feith et al. Overexpression of TGF-β1 in
esophageal (Barrett’s) adenocarcinoma is associated with advanced stage of disease and
poor prognosis. Molecular Carcinogenesis. 2006, 10, pp. 786– 794.
174. B. A. Pockaj, G. D. Basu, L. B. Pathangey et al. Reduced Tcell and dendritic cell
function is related to cyclooxygenase- 2 overexpression and prostaglandin E2 secretion in
patients with breast cancer. Annals of Surgical Oncology. 2004, Vol. 11, 3, pp. 328–339.
175. L. Adams, G. K. Scott, and C. S. Weinberg. Biphasic modulation of cell growth by
recombinant human galectin-1. Biochimica et Biophysica Acta—Molecular Cell Research.
1996, 2, pp. 137–144.
176. S. Rorive, N. Belot, C. Decaestecker et al. Galectin-1 is highly expressed in human
gliomas with relevance for modulation of invasion of tumor astrocytes into the brain
parenchyma. GLIA. 2001, 3, pp. 241–255.
177. N. L. Perillo, K. E. Pace, J. J. Seilhamer, and L. G. Baum. Apoptosis of T cells mediated
by galectin-1. Nature. 1995, 6558, pp. 736–739.
178. D. Chung, V. P. Patel, M. Moran, L. A. Lewis, and M. C. Miceli. Galectin-1 induces
partial TCR ζ-chain phosphorylation and antagonizes processive TCR signal transduction.
Journal of Immunology. 2000, 7, pp. 3722–3729.
179. G. A. Niehans, T. Brunner, S. P. Frizelle et al. Human lung carcinomas express Fas
ligand. Cancer Research. 1997, 6, pp. 1007–1012.
180. M. Hahne, D. Rimoldi, M. Schroter et al. Melanoma cell expression of Fas(Apo-1/CD95)
ligand: implications for tumor immune escape. Science. 1996, 5291, pp. 1363– 1366.
181. H. Arai, D. Gordon, E. G. Nabel, and G. J. Nabel. Gene transfer of Fas ligand induces
tumor regression in vivo. Proceedings of the National Academy of Sciences of the United
States of America. 1997, 95, pp. 13862–13867.
182. G. Andreola, L. Rivoltini, C. Castelli et al. Induction of lymphocyte apoptosis by tumor
cell secretion of FasLbearing microvesicles. Journal of ExperimentalMedicine. 2002, 10, pp.
1303–1316.

83
183. M. Iero, R. Valenti, V. Huber et al. Tumour-released exosomes and their implications
in cancer immunity. Cell Death and Differentiation. 2008, 1, pp. 80–88.
184. S. L¨ob, A. K¨onigsrainer, D. Zieker et al. IDO1 and IDO2 are expressed in human
tumors: levo- but not dextro-1- methyl tryptophan inhibits tryptophan catabolism. Cancer
Immunology, Immunotherapy. 2009, 1, pp. 153–157.
185. Fusenig, M. M. Mueller and N. E. Constitutive expression of G-CSF and GM-CSF in
human skin carcinoma cells with functional consequence for tumor progression.
International Journal of Cancer. 1999, 6, pp. 780–789.
186. Finn, J. Schmielau and O. J. Activated granulocytes and granulocyte-derived hydrogen
peroxide are the underlying mechanism of suppression of T-cell function in advanced
cancer patients. Cancer Research. 2001, 12, pp. 4756– 4760.
187. Onizuka S, Tawara I, Shimizu J, Sakaguchi S, Fujita T, Nakayama E. Onizuka S, Tawara I,
Shimizu J, Sakaguchi S, Fujita T, Nakayama Ey. Cancer Res. 1999, 59, pp. 3128–33.
188. Dannull J, Su Z, Rizzieri D, Yang BK, Coleman D, Yancey D, Zhang A, Dahm P, Chao N,
Gilboa E, Vieweg J. Enhancement of vaccine-mediated antitumor immunity in cancer
patients after depletion of regulatory T cells. J Clin Invest. 2005, 115, pp. 3623–33.
189. Antony PA, Piccirillo CA, Akpinarli A, Finkelstein SE, Speiss PJ, Surman DR,Palmer DC,
Chan CC, Klebanoff CA, Overwijk WW, Rosenberg SA, Restifo NP. CD8þ T cell immunity
against a tumor/ self-antigen is augmented by CD4þ T helper cells and hindered by
naturally occurring T regulatory cells. J Immunol. 2005, 174, pp. 2591–601.
190. Nishikawa H, Kato T, Tanida K, Hiasa A, Tawara I, Ikeda H, Ikarashi Y, Wakasugi H,
Kronenberg M, Nakayama T, Taniguchi M, Kuribayashi K, et al. CD4þ CD25þ T cells
responding to serologically defined autoantigens suppress antitumor immune responses.
Proc Natl Acad Sci USA. 2003, 100, pp. 10902–6.
191. Gnjatic S, Nishikawa H, Jungbluth AA, Gure AO, Ritter G, Jager E, Knuth A, Chen YT,
Old LJ. NY-ESO-1: review of an immunogenic tumor antigen. Adv Cancer Res. 2006, 95, pp.
1–30.
192. Danke NA, Koelle DM, Yee C, Beheray S, Kwok WW. Autoreactive T cells in healthy
individuals. J Immunol. 2004, 172, pp. 5967–72.
193. Bates GJ, Fox SB, Han C, Leek RD, Garcia JF, Harris AL, Banham AH. Quantification of
regulatory T cells enables the identification of high-risk breast cancer patients and those at
risk of late relapse. J Clin Oncol. 2006, 24, pp. 5373–80.
194. Sasada T, Kimura M, Yoshida Y, Kanai M, Takabayashi A. CD4þCD25þ regulatory T
cells in patients with gastrointestinal malignancies: possible involvement of regulatory T
cells in disease progression. Cancer. 2003, 98, pp. 1089–99.

84
195. Curiel TJ, Coukos G, Zou L, Alvarez X, Cheng P, Mottram P, Evdemon-Hogan M,
Conejo-Garcia JR, Zhang L, Burow M, Zhu Y, Wei S, et al. Specific recruitment of regulatory
T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat
Med. 2004, 10, pp. 942–9.
196. Ishida T, Ishii T, Inagaki A, Yano H, Komatsu H, Iida S, Inagaki H, Ueda R. Specific
recruitment of CC chemokine receptor 4-positive regulatory T cells in Hodgkin lymphoma
fosters immune privilege. Cancer Res. 2006, 66, pp. 5716–22.
197. D., Pardoll. Does the immune system see tumors as foreign or self? Annu Rev Immunol.
2003, 21, pp. 807–39.
198. Liu VC, Wong LY, Jang T, Shah AH, Park I, Yang X, Zhang Q, Lonning S, Teicher BA, Lee
C. Tumor evasion of the immune system by converting CD4þCD25_ T cells into CD4þCD25þ
T regulatory cells: role of tumor-derived TGF-b. J Immunol. 2007, 178, pp. 2883–92.
199. Nishikawa H, Kato T, Tawara I, Saito K,Ikeda H, Kuribayashi K, Allen PM, Schreiber RD,
Sakaguchi S, Old LJ, Shiku H. Definition of target antigens for naturally occurring CD4þ
CD25þ regulatory T cells. J Exp Med. 2005, 201, pp. 681–6.
200. Onizuka S, Tawara I, Shimizu J, Sakaguchi S, Fujita T, Nakayama E. Tumor rejection by
in vivo administration of anti-CD25 (interleukin-2 receptor a) monoclonal antibody. Cancer
Res. 1999, 59, pp. 3128–33.
201. Shimizu J, Yamazaki S, Sakaguchi S. Induction of tumor immunity by removing
CD25þCD4þ T cells: a common basis between tumor immunity and autoimmunity. J
Immunol. 1999, 163, pp. 5211–8.
202. Shevach EM, Stephens GL. The GITR-GITRL interaction: co-stimulation or
contrasuppression of regulatory activity? Nat Rev Immunol. 2006, 6, pp. 613–8.
203. Kanamaru F, Youngnak P, Hashiguchi M, Nishioka T, Takahashi T, Sakaguchi S,
Ishikawa I, Azuma M. Costimulation via glucocorticoid-induced TNF receptor in both
conventional and CD25þ regulatory CD4þ T cells. J Immunol. 2004, 172, pp. 7306–14.
204. Cohen AD, Diab A, Perales MA, Wolchok JD, Rizzuto G, Merghoub T, Huggins D, Liu C,
Turk MJ, Restifo NP, Sakaguchi S, Houghton AN. Agonist anti-GITR antibody enhances
vaccine-induced CD8þ T-cell responses and tumor immunity. Cancer Res. 2006, 66, pp.
4904–12.
205. Ko K, Yamazaki S, Nakamura K, Nishioka T, Hirota K, Yamaguchi T, Shimizu J,Nomura
T, Chiba T, Sakaguchi S. Treatment of advanced tumors with agonistic anti-GITR mAb and
its effects on tumor-infiltrating Foxp3þCD25þCD4þ regulatory T cells. J Exp Med. 2005, 202,
pp. 885–91.

85
206. Nishikawa H, Kato T, Hirayama M, Orito Y, Sato E, Harada N, Gnjatic S, Old LJ, Shiku H.
Regulatory T cell-resistant CD8þ T cells induced by glucocorticoid-induced tumor necrosis
factor receptor signaling. Cancer Res. 2008, 68, pp. 5948–54.
207. Peggs KS, Quezada SA, Chambers CA, Korman AJ, Allison JP. Blockade of CTLA- 4 on
both effector and regulatory T cell compartments contributes to the antitumor activity of
anti-CTLA-4 antibodies. J Exp Med. 2009, 206, pp. 1717–25.
208. Kavanagh B, O’Brien S, Lee D, Hou Y, Weinberg V, Rini B, Allison JP, Small EJ, Fong L.
CTLA4 blockade expands FoxP3þ regulatory and activated effector CD4þ T cells in a dose-
dependent fashion. Blood. 2008, 112, pp. 1175–83.
209. Valzasina B, Guiducci C, Dislich H, Killeen N, Weinberg AD, Colombo MP. Triggering of
OX40 (CD134) on CD4þCD25þ T cells blocks their inhibitory activity: a novel regulatory role
for OX40 and its comparison with GITR. Blood. 2005, 105, pp. 2845–51.
210. A. D.Weinberg, M. M. Rivera, R. Prell et al. Engagement of the OX-40 receptor in vivo
enhances antitumor immunity. Journal of Immunology. 2000, 168, pp. 2160–2169.
211. Yamaguchi T, Hirota K, Nagahama K, Ohkawa K, Takahashi T, Nomura T, Sakaguchi S.
Control of immune responses by antigen-specific regulatory T cells expressing the folate
receptor. Immunity. 2007, 27, pp. 145–59.
212. M L ROSSI, F CRUZ SANCHEZ, J T HUGHES, M M ESIRI, H B COAKHAM.
Immunocytochemical study of the cellular immune. J Clin Pathol. 1988, 41, pp. 314-319.
213. L. B. Boozer, T. W. Davis, L. B. Borst, K. M. Zseltvay, N. J. Olby and C. L. Mariani.
Characterization of Immune Cell Infiltration Into Canine Intracranial Meningiomas. Vet
Pathol. 2012, 49, pp. 784-795.
214. Kaushik Majumdar, Ravindra K. Saran , Pramod K. Chaurasia , Ila Tyagi , Daljit Singh.
Sphenoid wing lymphoplasmacyte-rich meningioma with occasional emperipolesis closely
simulating an intracranial Rosai-Dorfman disease: a diagnostic dilemma. Clin Neuropathol.
2013, 32, pp. 122-7.
215. Nielsen JS, Sahota RA, Milne K, Kost SE, Nesslinger NJ, Watson PH, Nelson BH. CD20+
tumor-infiltrating lymphocytes have an atypical CD27- memory phenotype and together
with CD8+ T cells promote favorable prognosis in ovarian cancer. Clin Cancer Res. 2012, 18,
pp. 3281-92.
216. Heimberger AB, Abou-Ghazal M, Reina-Ortiz C, et al. Incidence and prognostic impact
of FoxP3þ regulatory T cells in human gliomas. Clin Cancer Res. 2008, 14, pp. 5166–5172.
217. Joannes F.M. Jacobs, Albert J. Idema , Kalijn F. Bol , J. André Grotenhuis , I. Jolanda M.
de Vries, Pieter Wesseling , Gosse J. Adema a. Prognostic significance and mechanism of
Treg infiltration in human brain tumors. Journal of Neuroimmunology. 2010, 225, pp. 195–
199.

86
218. Ling EA, Kaur C, Lu J. Origin, nature, and some functional considerations of
intraventricular macrophages, with special reference to the epiplexus cells. Microsc Res
Tech. 1988, 41, pp. 43-56.
219. Bo L, Mork SJ, Nyland H. An immunohistochemical study of mononuclear cells in
meningiomas. Neuropathol Appl Neurobiol. 1992, 18, pp. 548–558.
220. Streit WJ, Conde JR, Fendrick SE, et al. Role of microglia in the central nervous
system’s immune response. Neurol Res. 2005, 27, pp. 685–691.
221. Ronald J. deLeeuw, Sara E. Kost , Juzer A. Kakal and Brad H. Nelson. The Prognostic
Value of FoxP3þ Tumor-Infiltrating Lymphocytes in Cancer: A Critical Review of the
Literature. Clin Cancer Res. 2012, 18, pp. 3022-3029.
222. Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, Lagorce-Pages C, et al.
Type, density, and location of immune cells within human colorectal tumors predict clinical
outcome. Science. 2006, 313, pp. 1960-4.
223. Dunn GP, Bruce AT, Ikeda H, et al. Cancer immunoediting: from immunosurveillance
to tumor escape. Nat Immunol. 2002, 3, pp. 991–998.
224. FM., Burnet. Cancer—a biological approach. Br Med J. 1957, 1, pp. 841–847.
Ringraziamenti: Premettendo che sarebbe veramente troppo difficile ringraziare adeguatamente
tutte le persone che mi hanno aiutato in questo mio lungo cammino, il mio primo
ringraziamento va alla mia famiglia, che mi ha sostenuto,spronato,accudito anche
nei momenti più sconfortevoli; allo stesso modo ringrazio anche i miei nonni e zii
che hanno contribuito non poco fino all'arrivo di questo importante giorno.
Ringrazio moltissimo anche tutto lo "staff tecnico" che mi ha insegnato e guidato in
questo lavoro ,a partire da Pietro, che non solo mi ha assistito in ogni fase della mia
tesi, ma ha regalato autentici momenti di ilarità in laboratorio; medesimi
ringraziamenti li rivolgo ai miei professori, Carlo e Alessandro, che oltre ad essere
estremamente competenti nel loro campo , si sono dimostrati comprensivi ,pazienti
e premurosi con il sottoscritto. Ringrazio molto anche Lisa, che mi ha trasmesso
l'arte del "taglio al microtomo" come un artigiano fa al proprio discepolo.
Ringrazio poi Giuliosky e Ferdi i miei due coinquilini, nonchè amici da sempre, che

87
hanno dovuto sopportare molteplici fisime personali nonchè angherie di qualsiasi
natura, ma con cui ho condiviso veramente una grossa fetta della mia PESante
personalità . Ringraziamenti senesi vanno anche a tutti i membri del Jah Parks (Fero,
Dieghito, Zio, Nicco, Jerry, Imps) ,ai mostri storici del liceo (Faze, Bubbolo, Mido,
Vivio, Boa ), agli amici di vecchia data come Filippo,Ozzy, Gigio, Breaks e Giulia.
Grazie anche agli innumerevoli mostri minori, che allietano ogni ritorno nella città
natale, sia con reggae funesto, cialde che durano notti intere o serate passate a
ricordare sketch passati. Un ringraziamento lo meritano sicuramente anche tutti i
miei vecchi compagni di classe (Rosinò,Vala,Bibons,Numba, Paris, Elisa, Vitty, Susò,
Elena,Marta,Marghe, Silvia, Pia, Carla, Virgy, Rizzo, Aitio Cate, Aly ) per aver
condiviso momenti tra i più cari che tengo dentro. Ringrazio anche i vecchi troiai
dei Cappuccini United ( Mone, Trek, Shorts, Grinch, Daddo,Lucche, e il Duca ) che ,
pur essendo passato molto tempo da quei momenti, custodisco ancora
gelosamente il numero 0,5. Mi sento poi di dover ringraziare tutti i miei amici
"pisani", che mi hanno fatto sentire a casa per tutti questi anni, e che mi
mancheranno terribilmente, una volta tornato in terra natia e più precisamente:
Agre, Moglie, Vio,Donni, Chiara,Della,Dome,Elisa,Arianna, Eleonora tutti grandi
prototipi ideali della M.V. moderna |Astra ragazza dall'estro formidabile
|Bomber|Cencio: accanito frequentatore della biblio |Chiara e Giulia grandissime
amiche e compagne di studio con cui ho condiviso moltissimi momenti di felicità, tra
esami e cene|Chiara una testona dal cuore d'oro che mi ha sopportato in queste
lunghe giornate|Cinzia e Lore coppia eterna di questo percorso|Clarks sempre
fissato con le zucchine e le meduse|Corso compagno di infinite notti in vetto|Diego,
diventato babbo da poco, ma amico da molto, insieme a tutto il magico staff della
Tana, sponsor di innumerevoli gioviali convivi, passati e ,spero, futuri |Domz, a cui
l'unica cosa che posso dire è minore tre|Edo che ci tira sempre peso nell'arte del
Subeba|Elettra amica dal cuore grande e color giallo blu|Fede compaÑero y oficial
heredor de mi erammù espaÑol|Fynetz che nonostante le avversità riesce sempre a
tirare fuori il meglio di sè|Giovà e Dani grandi festaioli da disco |Giulia e Alice le
uniche vere pisane conosciute il 1° anno|Jacopo grande appassionato di basket e

88
caccia| Jack, Giulia,Alessandra, Elettra, Giacomo, Luca, Silvia, Valeria, Valeria,
Costanza, Giovanna, Camilla, Valentina, Sandy, Gaia, Sonia,Costy, Valeria, Luca,
Sara Compagni di moltissimi momenti in facoltà e cene||Laura Marta Alessia
Roberta Marta eterne amiche che mi sono state al lato dai momenti più felici a
quelli più tristi e duri || Lottaccio che beve sempre di meno, come Elena può
confermare |Magna Tola e Cristiano dediziosi compagni di biblioteca|Manuela
Arianna Roby Carlotta bimbe dall'originalità smisurata|Marrux , con cui non vedo
l'ora di rifare la prossima scomessa|Matteo Ciccio e David tra le menti più svitate di
tutta facoltà
Matthews un vero collega di trash nonchè primo compagno di Paci|Michele e
Matteo : colleghi di laurea (spero)|Monica che mi ha permesso di fare una delle
esperienze più belle della mia vita, l'erasmus|Nicola il cecio col dito esploratore più
esplosivo che abbia mai conosciuto |Paolo Successore e sostituto ufficiale, nonchè
vicino da tempi immemori| Patri carissima persona che mi ha aiutato e confortato
in svariati momenti |Peppe sempre iperattivo, tamponato dalla bontà di Giulia |
Pons Jennifer Marta e Paesans compaÑeros con la sangre mas caliente que nà
|Samu a cui non so quante glie ne devo per aiuti di tipo logistico, ma che ha un
piccolo difetto : non si ricorda mai a che età il cavallo ha la bocca fatta | Susan
Chiara e Clara uno splendido trio di bimbe |Uarons un amico dalla rocciosità unica,
FAX|Tazio un fvustone d'eccellenza| Vale, Ludo e Fede instancabili pausare sempre
pronte a rallegrarti | Zinnaccio, Macciò e Marrius mostri di giurisprudenza che
hanno tenuto a freno il cane dei cani
e poi tutti gli altri colleghi di veterinaria che non ho citato finora.
Ringraziamenti speciali vanno anche ai migliori / peggiori elementi che hanno
caratterizzato il mio erasmus e mi riferisco a |Rocco e Umberto due dei migliori
tegami che potessi trovare a giro, in grado di farti sorridere con un sol gesto | Fede
il capo-cambusa tra i più molesti e forti mai conosciuti |Claudio e Jacopo Gli
inseparabili stanlio e onlio dello schifo| il Proca il più grande esperto di tecnologia
mai conosciuto, nonostante i suoi problemi| Adrian il pazzo per eccellenza, che
andrò a visitare presto | Tuttifrutti l'irrefrenabile viaggiatore che viaggia a barcelò|

89
Clement companero de piso che adorava i russian torpedo | il Boyler sempre caldo
a dovere |Turrins che si è sempre battuto contro la schiavitù dei suoi amici |
Giovanni Startac che non ha mai apprezzato il giouco del pallone| i Dottori Fazio,
Nania e Capilli che mi hanno fatto consocere Messina stando a Cordoba | Baffo |
Marione Massa l'uomo dalla chiacchera più convincente al mondo. Ringrazio poi i
carissimi colleghi veterinari conosciuti nella mia esperienza spagnola : Giulia da Pisa,
le mitiche torinesi Costy, Betta, Betta, Silvia (CLEMOOOOOO), le mie beddazze che
hanno vegliato su di me molte volte Irene e Roby, i diligenti, Nassi e Fabio, e tutti gli
altri miei amici erasmus, in primis le mitiche limonere,Laura MondAino, Monica,
Aidish e Manon, poi Francesco, Danilo , Diogo, Filipa,Anaconda-Filip, Helen,
Sballantaines . Ringrazio infine tutti i miei migliori amici spagnoli, ossia Alex, fratello
spirituale che mi manca troppo, le mie mamme amorose spagnole Ester e Celeste,
insieme alla mitica famiglia senesa Enrique,Cristina,Javi e Frassino. PQLP.