
Membrana Plasmatica Rafts di membrana. Falchetto, Dottorato 2013.

25/03/2012
1
Membrana Plasmatica
Rafts lipidici
http://en.wikipedia.org/wiki/Lipid_raft

25/03/2012
2

25/03/2012
3
BUSB 9
“LIPID RAFTS”(zattere lipidiche)
Un sempre maggiore numero di prove sperimentalisuggerisce che la membrana plasmatica contenga“rafts lipidici” arricchiti in sfingolipidi, colesterolo edalcune proteine di membrana
BUSB 10
Rafts lipidici (2)
La maggior parte delle molecole lipidiche nelle membrane cellulari sono disposte in modo casuale nel monostrato lipidico in cui risiedono.
Le forze di van der Waals di attrazione fra le code di acidi grassi vicini non sono sufficientemente selettive da tenere insieme gruppi di molecole di questo tipo.
Tuttavia, per alcune molecole lipidiche, come gli sfingolipidi, che tendono ad avere catene di acidi grassi lunghe e sature, le forze di attrazione possono essere sufficientemente forti da trattenere transitoriamente molecole adiacenti vicine formando piccoli microdomini. Tali microdomini ‐ “rafts” lipidici ‐ possono essere considerati come fasi di separazione transitorie nel doppio strato lipidico in cui si concentrano gli sfingolipidi.
11
Importanza dei legami doppi in posizione cisdelle catene idrocarburiche
I legami doppi rendono più difficile il compattamento reciproco delle catene, così rendendo più difficile il congelamento del ”bilayer” lipidico.Inoltre, dato che le catene degli acidi grassi dei lipidi insaturi
sono più sparpagliate, i bilayers lipidici che contengono lipidi insaturi sono più sottili dei “bilayers” formati esclusivemente dai lipidi saturi.
http://www.ncbi.nlm.nih.gov/books/NBK26871/figure/A1875/
Un “raft” (zattera) lipidico. I “rafts” lipidici sono piccole zone specializzate delle membrane dove alcuni lipidi (soprattutto sfingolipidi e colesterolo) e proteine (verde scuro) si concentrano. Dato che il bilayer lipidico è un può più spesso nei “rafts”, alcune proteine di membrana vi si accumulano.
http://www.ncbi.nlm.nih.gov/books/NBK26871/figure/A1881/

25/03/2012
4
BUSB 13
Struttura chimica generale degli sfingolipidi. Diversi sostituenti (R) danno:
H: ceramide
Fosfocolina: sfingomielina
Zuchero/i: glicosfingolipidi
STRUTTURA GENERALE DEGLI SFINGOLIPIDI
BUSB 14
CERAMIDII ceramidi sono una famiglia di molecole lipidiche.
Un ceramide è composto da sfingosina e di un acido grasso
Si trovano in elevata concentrazione nella membrana plasmatica come uno dei componenti della sfingomielina, che è uno dei principali lipidi del doppio strato lipidico.
Per anni si è presunto che i ceramidi e altri sfingolipidi fossero soltanto elementi strutturali ma oggi si sa che sono molto di più.
Forse uno degli aspetti più affascinanti del ceramide è che esso può agire da molecola di segnalamento.
Le funzioni più note dei ceramidi come segnalatori cellulari includono la regolazione del differenziamento, proliferazione e morte cellulare programmata (apoptosi)
BUSB 15
GLICOSFINGOLIPIDINella classe dei glicolipidi la testa polare é legata alla sfingosina mediante legame glicosidico di una molecola di zucchero, invece che mediante un legame fosfoesterico, come nel caso dei fosfolipidi.
(A) Il galattocerebroside è chiamato un glicolipide neutro perchè lo zucchero che forma la sua testa non è carico.
(B) Un ganglioside contiene sempre uno o più residui di acido sialico (noti anche come acido N-acetilneuraminico,NANA) carichi la cui strutturà è illustrata in (C).
Gal = galattosio; Glc = glucosio, GalNAc = N-acetilgalattosamina (questi 3 zuccheri non sono carichi)
BUSB 16
Sfingolipidi
I principali glicolipidi sono:
-cerebrosidi:(singolo zucchero)
-globosidis (oligosaccaridi
-sulfatidi (singolo zucchero acido)
-gangliosidi (diversi zuccheri acidi)
Senza residuo di zucchero, fosfolipide

25/03/2012
5
BUSB 17
Rafts lipidici (3)Si pensa che la membrana plasmatica delle cellule animali contenga molti di questi “rafts” lipidici (~70 nm di diametro), che sono ricchi sia di sfingolipidi che di colesterolo.
Dato che le catene di idrocarburi dei lipidi ivi concentrati sono più lunghe e più dritte delle catene di acidi grassi della maggior parte dei lipidi di membrana, i “rafts” sono più spessi delle altre zone del “bilayer” e sono meglio in grado di accomodare alcune proteine di membrana, che tendono quindi ad accumularsi in quelle regioni.
In questo modo, si pensa che i “rafts” lipidici organizzino tali proteine – sia concentrandole per il trasporto in piccole vescicole, sia permettendo alle proteine di funzionare in modo integrato ‐ come fanno quando convertono segnali extracellulari in segnali intracellulari.
18
Importanza dei legami doppi in posizione cisdelle catene idrocarburiche
I legami doppi rendono più difficile il compattamento reciproco delle catene, così rendendo più difficile il congelamento del ”bilayer” lipidico.Inoltre, dato che le catene degli acidi grassi dei lipidi insaturi
sono più sparpagliate, i bilayers lipidici che contengono lipidi insaturi sono più sottili dei “bilayers” formati esclusivemente dai lipidi saturi.
http://www.ncbi.nlm.nih.gov/books/NBK26871/figure/A1875/
Un “raft” (zattera) lipidico. I “rafts” lipidici sono piccole zone specializzate delle membrane dove alcuni lipidi (soprattutto sfingolipidi e colesterolo) e proteine (verde scuro) si concentrano. Dato che il bilayer lipidico è un può più spesso nei “rafts”, alcune proteine di membrana vi si accumulano.
http://www.ncbi.nlm.nih.gov/books/NBK26871/figure/A1881/
BUSB 20
Struttura chimica generale degli sfingolipidi. Diversi sostituenti (R) danno:
H: ceramide
Fosfocolina: sfingomielina
Zuchero/i: glicosfingolipidi
STRUTTURA GENERALE DEGLI SFINGOLIPIDI

25/03/2012
6
BUSB 21
CERAMIDII ceramidi sono una famiglia di molecole lipidiche.
Un ceramide è composto da sfingosina e di un acido grasso
Si trovano in elevata concentrazione nella membrana plasmatica come uno dei componenti della sfingomielina, che è uno dei principali lipidi del doppio strato lipidico.
Per anni si è presunto che i ceramidi e altri sfingolipidi fossero soltanto elementi strutturali ma oggi si sa che sono molto di più.
Forse uno degli aspetti più affascinanti del ceramide è che esso può agire da molecola di segnalamento.
Le funzioni più note dei ceramidi come segnalatori cellulari includono la regolazione del differenziamento, proliferazione e morte cellulare programmata (apoptosi)
BUSB 22
GLICOSFINGOLIPIDINella classe dei glicolipidi la testa polare é legata alla sfingosina mediante legame glicosidico di una molecola di zucchero, invece che mediante un legame fosfoesterico, come nel caso dei fosfolipidi.
(A) Il galattocerebroside è chiamato un glicolipide neutro perchè lo zucchero che forma la sua testa non è carico.
(B) Un ganglioside contiene sempre uno o più residui di acido sialico (noti anche come acido N-acetilneuraminico,NANA) carichi la cui strutturà è illustrata in (C).
Gal = galattosio; Glc = glucosio, GalNAc = N-acetilgalattosamina (questi 3 zuccheri non sono carichi)
BUSB 23
Sfingolipidi
I principali glicolipidi sono:
-cerebrosidi:(singolo zucchero)
-globosidis (oligosaccaridi
-sulfatidi (singolo zucchero acido)
-gangliosidi (diversi zuccheri acidi)
Senza residuo di zucchero, fosfolipide
BUSB 24
Rafts lipidici (3)Si pensa che la membrana plasmatica delle cellule animali contenga molti di questi “rafts” lipidici (~70 nm di diametro), che sono ricchi sia di sfingolipidi che di colesterolo.
Dato che le catene di idrocarburi dei lipidi ivi concentrati sono più lunghe e più dritte delle catene di acidi grassi della maggior parte dei lipidi di membrana, i “rafts” sono più spessi delle altre zone del “bilayer” e sono meglio in grado di accomodare alcune proteine di membrana, che tendono quindi ad accumularsi in quelle regioni.
In questo modo, si pensa che i “rafts” lipidici organizzino tali proteine – sia concentrandole per il trasporto in piccole vescicole, sia permettendo alle proteine di funzionare in modo integrato ‐ come fanno quando convertono segnali extracellulari in segnali intracellulari.

25/03/2012
7
Rafts lipidici (4)
Per la maggior parte, le molecole lipidiche in un monostrato si muovono in torno indipendentemente da quelle dell’altro monostrato. Tuttavia, nei “rafts” lipidici, le lunghe catene idrocarburiche degli sfingolipidi di un monostrato interagiscono con quelle dell’altro monostrato.Perciò, i due monostrati di un “raft” lipidico interagiscono mediante le loro code lipidiche.
BUSB 25
BUSB 26
Thomas Harder and Kai Simons: Caveolae, DIGS, and the dynamics of sphingolipid-cholesterol microdomains. Current Opinion in Cell Biology 1997, 9:534-542
Modello ipotetico dell’organizzazione lipidica nei microdomini “raft”.Gli sfingolipidi (figure in rosso con due gambe) e il colesterolo intercalato (figure più piccole arancione) formano un microdominio altamente impacchettato in un ambiente ricco di fosfatidilcolina (PC; figure blu, due gambe).Le regioni ricche di PC sono impacchettate meno strettamente e più fluide dei domini ricchi in sfingolipidi e colesterolo.
BUSB 27
BUSB 28

25/03/2012
8
Rafts lipidici (5)
I rafts lipidici pottrebbero mediare lo smistamento (“sorting”) di glicosfingolipidi e proteine ancorate mediante GPI alla membrana plasmatica apicale (Alberts et al.: Molecular Biology of the Cell. Garland Pub; 4th edition (March
2002), pp. 589‐590 pp. 763‐764)
La membrana plasmatica apicale di molte cellule è enormemente arricchita di glicosfingolipidi che aiutano a proteggere questa superficie esposta dal danno ad esempio provocato dagli enzimi digestivi e pH acido in siti come lo stomaco o il lume dell’intestino.Anche proteine della membrana plasmatica che sono legate al bilayer lipidico da un àncora di glicosilfosfatidilinositolo (GPI), si trovano esclusivamente nella membrana plasmatica apicale. Se sui usano tecniche di DNA ricombinante per legare un’àncora di GPI ad una proteina che normalmente sarebbe consegnata alla superficie basolaterale, la proteine viene invece consegnata alla membrana apicale.
Rafts lipidici (6)
Si ritiene che le proteine con àncora di GPI vengano dirette alla membrana apicale perché sono associate ai glicosfingolipidi nei “rafts” lipidici che si formano nella membrana della rete trans del Golgi. Anche le proteine di membrana con domini trans‐membrana insolitamente lunghi si accumulano nei “rafts”. Inoltre, i “rafts” preferenzialmente contengono proteine GPI‐ancorate e alcune proteine che legano carboidrati (lectine) che possono aiutare a stabilizzare le membrane. Una volta che hanno selezionato il loro carico caratteristico di molecole, i rafts gemmano dalla rete trans del Golgi in vescicole di trasporto destinate alla membrana plasmatica apicale.
Modello di “rafts” lipidici nella rete trans del Golgi. Si pensa che gli glicosfingolipidi e il colesterolo formino “rafts” nel “bilayer lipidico”. Proteine di membrana con segmenti trans‐membrana sufficientemente lunghi si distribuiscono preferenzialmente nei “rafts” lipidi e quindi vengono smistate in vescicole di trasporto. Questi “rafts” vengono in seguito inglobati in vescicole di trasporto che li trasportano al dominio apicale della membrana plasmatica. Proteine che legano i carboidrati (lectine) nel lume della rete trans‐ del Golgi possono aiutare a stabilizzare i “rafts”, come illustrato.
http://www.ncbi.nlm.nih.gov/books/NBK26892/figure/A2430/
Esempi di proteine ancorate alla membrana plasmatica mediante lipidi e glicolipidi.Alcune proteine (ad es., la proteina dei linfociti Thy‐1) sono ancorate al foglietto esterno della membrana plasmatica mediante ancore a glicosil fosfatidil inositolo (GPI) che vengono legate covalentemente al loro C‐terminale nel reticolo endoplasmatico. Queste proteine sono glicosilate ed esposte sulla superficie cellulare. Altre proteine sono ancorate al foglietto interno della membrana plasmatica in seguito alla loro traduzione su ribosomi liberi nel citosol. La proteina Ras illustrata è ancorate mediante un gruppo prenilico legato covalentemente alla catena laterale di una cisteina del C‐terminale e da un gruppo palmitoilo legato ad una cisteina localizzata 5 aminoacidi più a monte. La protein Src è ancorata mediante un gruppo miristoil legato al suo N‐terminale. Anche una regione carica positivamente della Src gioca un ruolo nell’associazione alla membrana, forse mediante l’interazione con le teste cariche negativamente della fosfatidilserina.http://www.ncbi.nlm.nih.gov/books/NBK9898/figure/A1979/?report=objectonly

25/03/2012
9
GLICOLIPIDI E TRASDUZIONE DEL SEGNALE
Rafts
GLICOLIPIDI E TRASDUZIONE DI SEGNALEKohji Kasahara (Tokyo Metropolitan Institute of Medical Science)
Hakomori et al. hanno dimostrato che i glicosfingolipidi (GSLs) sono modulatori della trasduzione di segnale.Studi recenti mostrano che l’aggregazione dinamica di GSLs e colesterolo per dare origine a microdomini nell’ambito del bilayer lipidico e l’associazione di una gran varietà di molecole di segnalamento con questo dominio. I GSLs sono relativamente ricchi di acidi grassi con catene aciliche sature, che permettono uno stretto impachettamento e conferiscono temperature di fusione alte. Viceversa, i fosfolipidi sono relativamente ricchi di acidi grassi con catene aciliche cis‐insature (stutture a gomito), che impediscono l’impachetamento stretto, e conferiscono temperature di fusione basse.
http://www.glycoforum.gr.jp/science/word/glycolipid/GLB04E.html
Proteine ancorate tramite GPI, proteine acilate come la famiglia src delle tirosina chinasi e le proteine trimerica G, sono asssociate ai microdomini di GSLs.Le proteine GPI‐ancorate hanno di solito catene aciliche sature che probabilmente si inseriscono preferenzialmente nei microdomini GSLs. La famiglia Src delle proteina chinasi vengono modificate da lipidi a catena satura (palmitoilazione e miristilazione) che probabilmente si inseriscono preferenzilamente nei microdomini GSL.

25/03/2012
10
Il cross‐linking mediato da anticorpi (o ligandi) delle proteine GPI‐ancorate induce l’attivazione della famiglia delle src‐chinasi e un transiente aumento della fosforilazione a livello della tirosina di diversi substrati (Fig. 2a). Anche il cross‐linking degli GSLs mediato da anticorpi induce l’attivazione della famiglia src delle proteina chinasi e un transitorio aumento di fosforilazione della tirosina (Fig.2b).
Ciò dimostra che il cross‐linking dei GSLs mediato da anticorpi può simulare il segnalamento mediato da proteine GPI‐ancorate. Anticorpi contro le proteine GPI‐ancorate fanno immunoprecipitare la famiglia delle src‐chinasi, e anticorpi contro i GSL co‐immunoprecipitano le src‐chinasi e le proteine GPI‐ancorate. Inoltre, la rimozione enzimatica della frazione di carboidrati dei GSLs della superficie cellulare ostacola l’attivazione della famiglia delle src‐chinasi mediante anticorpo‐mediato cross‐linking delle proteine GPI‐ancorate. Queste osservazioni suggeriscono che i GSLs sono coinvolti nel signalamento mediato da proteine GPI‐ancorate.Nonostante non ci sia ancora accordo sul fatto che le proteine GPI‐ancorate si associno stazionariamente ai microdomini GSLs, si pensa che il cross‐linking mediato da anticorpi delle proteine GPI‐ancorate induca la traslocazione verso i microdomini GSL o stabilizzino la loro associazione con i microdomini GSL.

25/03/2012
11
I microdomini GSL (“rafts”) sono coinvolti nel segnalamento mediato da immunorecettori e recettori dei fattori di crescita. Un’attivazione efficiente delle cellule T richiede un segnale da un “T‐cell antigen receptor” e un secondo segnale da una molecola co‐stimolatoria. La co‐stimolazione porta al reclutamento di microdomini GSL verso il sito di contatto cellula‐cellula fra la cellula T e la cellula che presenta l’antigene. La concentrazione di chinasi della famiglia src e di molecole “downstream”, e l’esclusione della tirosina fosfatasi CD45, permette una forte e prolungata fosforilazione di diversi substrati.Inoltre, i recettore per l’”Epidermal Growth Factor, EGF” e Rassono presenti nei microdomini GSL e il trattamento con EGF induce la traslocazione di Raf‐1 e MAPKK chinasi dal citosol ai microdomini GSL. Ciò suggerisce che i microdomini GSL possono essere siti di inizio per la cascata delle MAP chinasi.
BUSB 42
BUSB 43
BUSB 44

25/03/2012
12
BUSB 45
BUSB 46
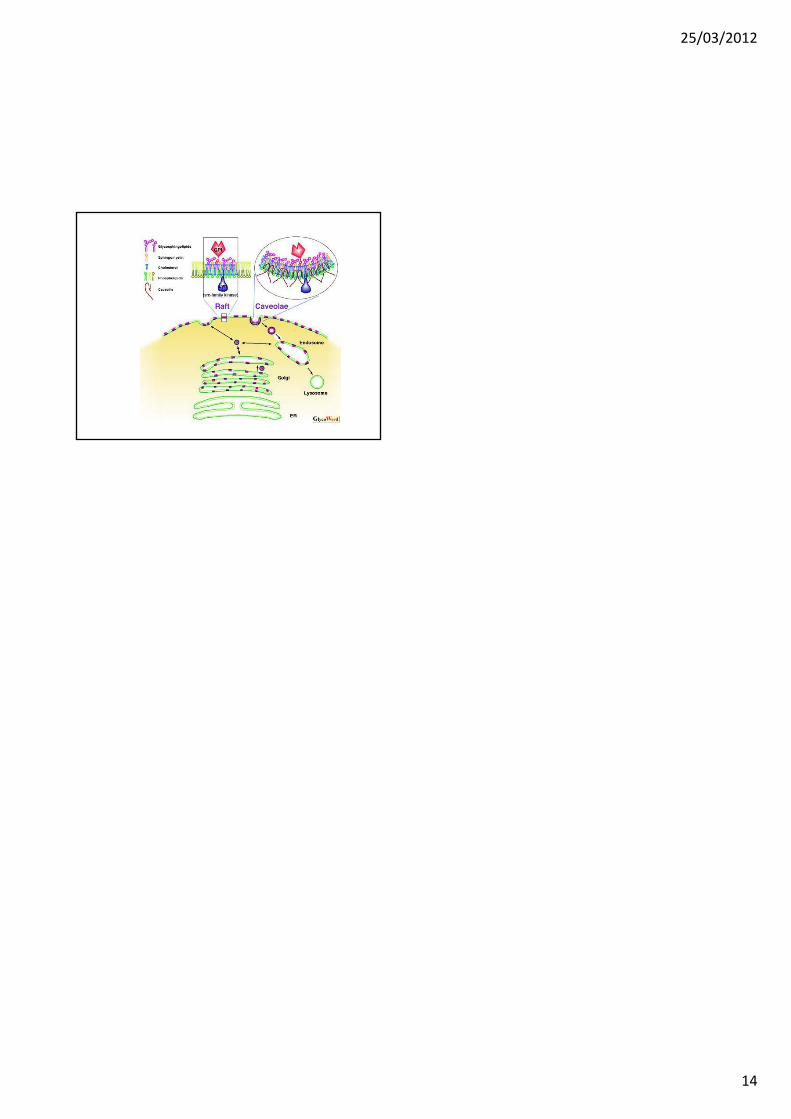
CAVEOLAE
CAVEOLAE (1)
(2005) Ci sono almeno due tipi di membrane a bassa densità,resistenti ai detergenti: caveolae e rafts lipidici.Caveolae: temine usato per descrive invaginazioni a forma di fiascoche sono arricchite in colesterolo e circondate da un rivestimentodella proteina strutturale caveolina-1.I rafts lipidici sono domini piatti della membrana, che sono privi dicaveolina-1 ma ciò nonostante sono arricchiti di colesterolo e sonoresistenti all’estrazione con detergenti.
Le caveolae sono state identificate per prima mediante esame conmicroscopia elettronica a metà degli anni 50 da due ricercatori(Palade, 1953; Yamada, 1955), come invaginazioni a forma difiaschette di 50-100 nm della membrana plasmatica.

25/03/2012
13
Alberts, 3rd ed: Figura 13.48. Caveolae sulla membrana plasmatica di un fibroblasto umano.(A) ME a trasmissione di un fibroblasto in sezione trasversale che mostra caveolae comeprofonde invaginazioni della membrana plasmatica. (B) ME a scansione di una replica“deep‐etch”, che mostra numerose caveolae sul versante citoplasmatico della membranaplasmatica. Il loro rivestimento sembra essere fatto da filamenti disposti concentricamenteche contengono la proteina transmembrana caveolina. Notare che le caveolae differisconosia in dimensioni che in struttura dai pozzetti rivestiti da clatrina, uno dei quali è visibile inaltro a destra di (B).
CAVEOLAE (2)
• Le caveolae sono state osservate in una grandiversità di tipi cellulari, in particolare nelle celluleendoteliali, ma non nei tessuti neuronali.
• Nelle caveolae sono particolarmente concentrate molte proteine e lipidi (Tabella 1); tuttavia, la marcatura delle cellule con un marcatore del dominio “Pleckstrin Homology, PH” specifico per il PIP2 indica che questo lipide non è concentrato nelle caveolae.
• Il principale marcatore delle caveolae è la proteina caveolina.
CAVEOLAE (3)
Caveolae. I glicosfingolipidi, e altri lipidi con catene aciliche lunghe e dritte sono indicati in arancione, i lipidi normali in giallo-verde. Le proteine transmembrana caveoline sono in blu. "I rubini rossi" rappresentano enzimi e recettori ancorati a GPI. Le sfere verdi sono ”Src-like kinases” palmitolate, e i recettori transmembranosi grigio e arancione rappresentano recettori di segnalamento associati alle caveolae.
http://www.bms.ed.ac.uk/research/others/smaciver/Cyto‐Topics/caveolae.htm
25/03/2012
14















![Cellular and molecular comparison of redifferentiation of … · 2010-01-26 · proliferative-competent progeny cells [9] that possess ability to redifferentiate into lipid-assimilating](https://static.fdocumenti.com/doc/165x107/5f612d6b67aa1f530602a4b5/cellular-and-molecular-comparison-of-redifferentiation-of-2010-01-26-proliferative-competent.jpg)


