2 Lipidi e membrane BUSB 2aparte stud...14/03/2014 2 Forme gelificate e fluide del Effettobilayer...
22
14/03/2014 1 Lipidi & Membrane Laurea Magistrale Biologia Sperimentale e Applicata 2° PARTE Lipidi e membrane EFFETTO DEL TIPO DI LIPIDE SULLA FLUIDITA’ E SPESSORE DELLA MEMBRANA (SEGUE) Membrane Influenza dei legami doppi in posizione cis delle catene idrocarburiche I legami doppi rendono più difficile il compattamento reciproco delle catene, così rendendo più difficile il congelamento del ”bilayer” lipidico. Inoltre, dato che le catene degli acidi grassi dei lipidi insaturi sono più sparpagliate, i bilayers lipidici che contengono lipidi insaturi sono più sottili dei “bilayers” formati esclusivemente dai lipidi saturi.
Transcript of 2 Lipidi e membrane BUSB 2aparte stud...14/03/2014 2 Forme gelificate e fluide del Effettobilayer...

14/03/2014
1
Lipidi & Membrane
Laurea Magistrale
Biologia Sperimentale e Applicata 2° PARTE
Lipidi e membrane
EFFETTO DEL TIPO DI LIPIDE SULLA FLUIDITA’ E SPESSORE DELLA MEMBRANA (SEGUE)
Membrane
Influenza dei legami doppi in posizione cisdelle catene idrocarburiche
I legami doppi rendono più difficile il compattamento reciproco delle catene, così rendendo più difficile il congelamento del ”bilayer” lipidico.
Inoltre, dato che le catene degli acidi grassi dei lipidi insaturi sono più sparpagliate, i bilayers lipidici che contengono lipidi insaturi sono più sottili dei “bilayers” formati esclusivemente dai lipidi saturi.

14/03/2014
2
Forme gelificate e fluide del bilayer lipidico
(Sopra): Schema della transizione gel-fluido. I fosfolipidi con lunghe catene di acidi grassi saturi tendono ad assemblarsi in un doppio strato altamente ordinato, tipo gel, in cui vi è poca sovrapposizione delle code apolari nei due foglietti. Il calore disordina le code apolari ed induce una transizione gel-fluido entro una gamma di temperature di soli pochi gradi. Mentre le catene si disordinano, anche il bilayer diminuisce di spessore.
(Sotto). Modelli molecolari dei monostrati fosfolipidici.Lodish et al., 7° ed., 2013
Effetto della composizione lipidica sullo spessore e curvatura del bilayer (1)
a) Un bilayer puro di sfingomielina (SM) è più spesso di uno formato da un fosfogliceride tipo fosfatidilcolina (PC). Il colesterolo promuove l’ordine sui bilayers di fosfogliceridi, ma non influenza lo spessore del bilayer più ordinato di SM.
b) I fosfolipidi come la PC hanno una forma cilindrica e formano monostrati essenzialmente piatti, mentri quelli con gruppi di testa più piccoli, come la fosfatidiletanolamina (PE) hanno forma conica.
Lodish et al., 7° ed., 2013
Effetto della composizione lipidica sullo spessore e curvatura del bilayer (2)
c) Un bilayer arricchito con fosfatidilcolina (PC) nel foglietto exoplasmico e con fosfatidiletanolamina (PE) nel foglietto citosolico, come avviene nella maggior parte delle membrane plasmatiche, avrebbe una curvatura naturale.
Esempi di formazione di vescicoleEndocitosi
http://upload.wikimedia.org/wikipedia/commons/thumb/1/1a/Endocytosis_types.svg/672px‐Endocytosis_types.svg.png

14/03/2014
3
Esempi di formazione di vescicoleTraffico di vescicole nel Golgi (1)
http://www.cytochemistry.net/cell‐biology/gol1.jpg
Esempi di formazione di vescicoleTraffico di vescicole nel Golgi (2)
http://www.hhmi.org/sites/default/files/Our%20Scientists/Investigators/FerroNovick‐fig2.jpg
Domini lipidici (1)
In una miscela di sfingolipidi e di glicerofosfolipidi, il colesterolo può indurre immiscibilità fluido‐fluido che provoca una segregazione laterale in due o più fasi fluide.
Il colesterolo interagisce con gli sfingolipidi del foglietto esterno e con la fosfatidilserina disatura nel foglietto citoplasmatico.
Le fasi arricchite in colesterolo hanno una struttura più ordinata e sono state designate “liquido ordinato” in opposizione a “liquido disordinato).
Due domini di composizioni differenti possono essere distanziati da 1 µm, oppure un dominio può essere sovrapposto all’altro, oppure formare un cerchio attorno ad esso.
Sprong H, van der Sluijs P, van Meer G. How proteins move lipids and lipids move proteins. Nat RevMol Cell Biol. 2001 Jul;2(7):504‐13.
http://www.nature.com/nrm/journal/v2/n7/fig_tab/nrm0701_504a_F3.html#figure‐titleSprong H, van der Sluijs P, van Meer G. How proteins move lipids and lipidsmove proteins. Nat Rev Mol Cell Biol. 2001
Jul;2(7):504‐13.
Il colesterolo aumenta la lunghezza della fosfatidilcolina (PC) ma non quella della sfingomielina (SM). La sfingomielina con o senza colesterolo forma doppi strati con uno spessore di 46-47 Å per la sfingomielina con un acido grasso saturo con 18 atomi di carbono (C18:0) e di 52-56 Å per la sfingomielina C24:0. Viceversa, lo spessore di un bilayer di PC C16:o/C18:1 è di 35 Å, che viene aumentato a 40 Å dal colesterolo. Lo spessore della regione idrofobica del bilayer è aumentato da 26 a 30 Å.
14/03/2014
4
http://www.nature.com/nrm/journal/v2/n7/fig_tab/nrm0701_504a_F3.html#figure‐titleSprong H, van der Sluijs P, van Meer G. How proteins move lipids and lipidsmove proteins. Nat Rev Mol Cell Biol. 2001
Jul;2(7):504‐13.
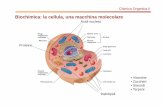
A seconda che i domini su entrambe le superficie colocalizzano o meno, possono essere presenti bilayers con quattro diversi spessori in queste membrane.
http://www.nature.com/nrm/journal/v2/n7/images/nrm0701_504a_f5.gifSprong H, van der Sluijs P, van Meer G. How proteins move lipids and lipidsmove proteins. Nat Rev Mol Cell Biol. 2001
Jul;2(7):504‐13.
La conformazione molecolare dei lipidi determina le proprietà fisiche delle membrane (1)
La conformazione molecolare dei lipidi determina le proprietà fisiche delle membrane (2)
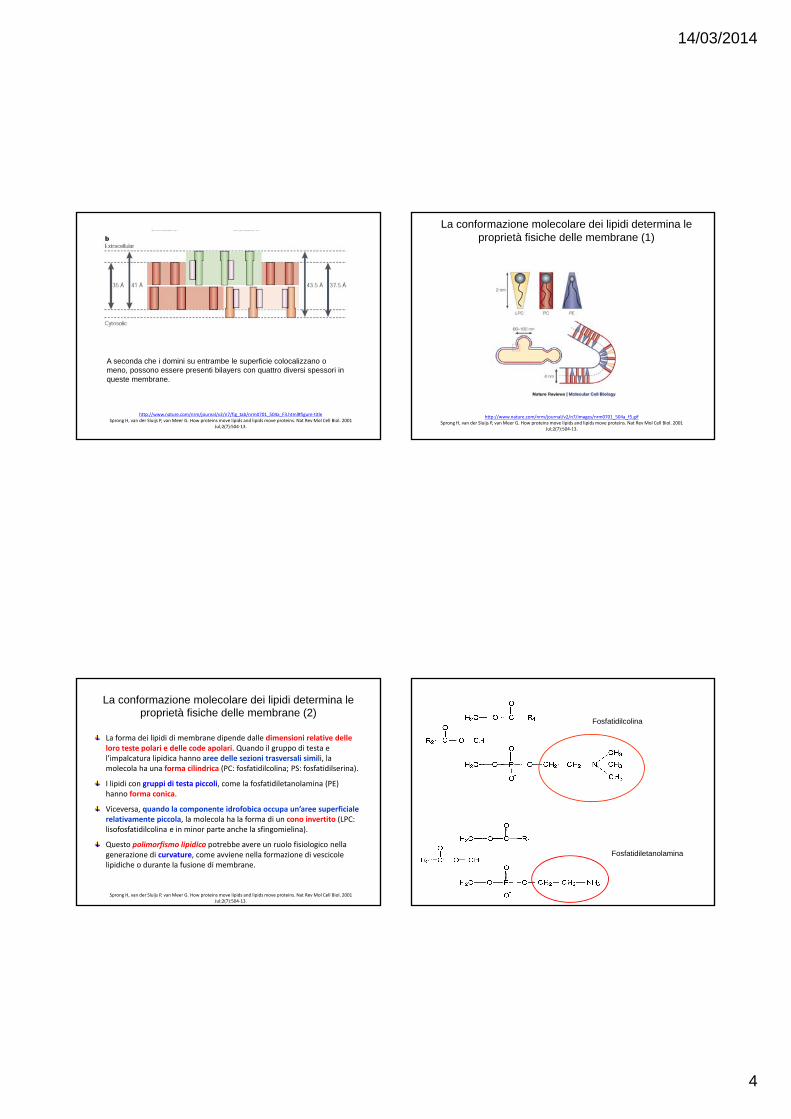
La forma dei lipidi di membrane dipende dalle dimensioni relative delle loro teste polari e delle code apolari. Quando il gruppo di testa e l’impalcatura lipidica hanno aree delle sezioni trasversali simili, la molecola ha una forma cilindrica (PC: fosfatidilcolina; PS: fosfatidilserina).
I lipidi con gruppi di testa piccoli, come la fosfatidiletanolamina (PE) hanno forma conica.
Viceversa, quando la componente idrofobica occupa un’aree superficiale relativamente piccola, la molecola ha la forma di un cono invertito (LPC: lisofosfatidilcolina e in minor parte anche la sfingomielina).
Questo polimorfismo lipidico potrebbe avere un ruolo fisiologico nella generazione di curvature, come avviene nella formazione di vescicole lipidiche o durante la fusione di membrane.
Sprong H, van der Sluijs P, van Meer G. How proteins move lipids and lipidsmove proteins. Nat Rev Mol Cell Biol. 2001 Jul;2(7):504‐13.
Fosfatidiletanolamina
Fosfatidilcolina

14/03/2014
5
La conformazione molecolare dei lipidi determina le proprietà fisiche delle membrane (3)
La superficie citosolica della membrane plasmatica contiene 40% di
fosfatidiletanolamina (PE), 60% di fosfatidilserina più fosfatidilcolina
(PS+PC) mentre il foglietto rivolto vero l’ambiente esterno contiene 60% di
PC, 30% di SM e 10% di PE. La PE di per se adotta una fase esagonale, e
questa tendenza probabilmente favorisce l’invaginazione della membrana.
La gemmazione in direzione opposta, verso il lume degli endosomi, può
richiedere acido lisobisfosfatidico, un cono invertito, sulla superficie
luminale e fosfatidilinositolo‐3‐fosfato.
Il colesterolo è richiesto per la gemmazione delle vescicole sinaptiche che
hanno un’elevata curvatura (40‐50 nm di diametro).
Il colesterolo + la sfingomielina sono inoltre importanti per stabilizzare le
membrane durante la fusione, e la fosfatidiletanolamina stimola
moltissimo l’efficienza di fusione.
Quando le fosfolipasi agiscono sui fosfolipidi producono lisofosfolipidi, che mancano da una delle due catene aciliche.
I lisofosfolipidi sono non solo importanti molecole di segnalamento, rilasciate dalle cellule e riconosciute da recettori specifici, ma possono anche influenzare le proprietà fisiche delle membrane dove si trovano.
Acido lisobisfosfatidico
http://www.cyberlipid.org/phlip/pgly05.htm
E’ un curioso fosfolipide con una configurazione diversa da tutti gli altri noti fosfolipidi dei mammiferi. E’ stato identificato per primo nei polmoni dei mammiferi. E’ molto abbondate nel compartimento lisosomiale del fegato di ratto, soprattutto in alcune disfunzioni di accumulo lisosomiale (ad es, malattie di Niemann-Pick, lipofuscinosi ceroidi neuronali). Ci sono dati preliminari sul suo coinvolgimento sia nella dinamica che nelle funzioni di smistamento lipidi/proteine degli endosomi tardivi, soprattutto per il controllo della distribuzione di colesterolo.Inoltre, i gruppi R1 e R2 sono frequentemente acido arachidonico (macrofagi alveolari) o acido docosaesanoicoin alcune altre cellule con acido oleico come altro gruppo acilico. Più recentemente è stato dimostrato che l’acido lisobisfosfatidico è il principale fosfolipide, insieme alla fosfatidilcolina, degli endosomi tardivi. L’acido lisobisfosfatidico esibisce proprietà fusogeniche dipendenti dal pH in modo unico e caratteristico e quindi è il lipide ideale per regolare le proprietà dinamiche di queste membrane cellulari.
PROTEINE DI MEMBRANA
Membrane

14/03/2014
6
CATEGORIE
Aminoacidi
Ionizzazione degli AA polari previsti di carica
A pH fisiologico praticamente tutti i residui di ac. glutammico sono carichi negativamente
A pH fisiologico praticamente tutti i residui di lisina sono carichi positivamente
Ac. glutammico
Lisina

14/03/2014
7
Polari carichi (1) Polari ma privi di carica
Non Polari (Idrofobici) (1) Catene laterali con proprietà particolari
GLICINA: la catena laterale è formata solo da un atomo di H e può adattarsi sia ad un ambiente idrofilo che idrofobico. Spesso si trova in siti dove due polipeptidi sono a stretto contatto
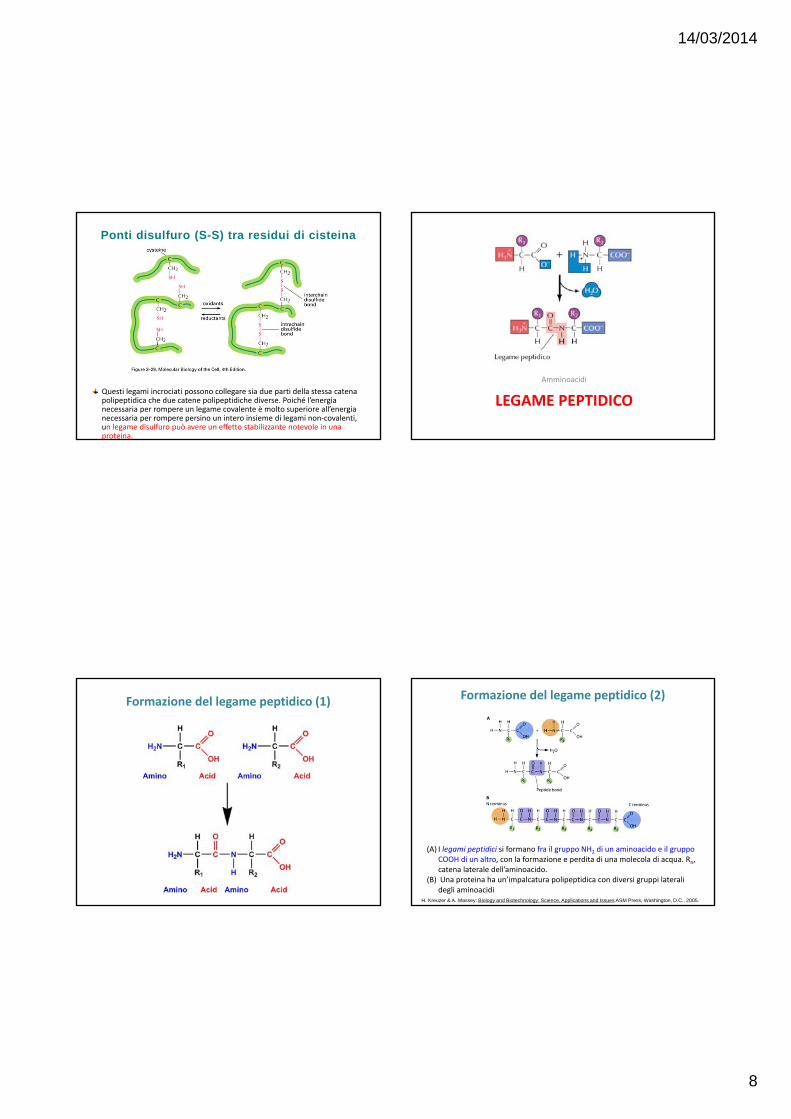
CISTEINA: Sebbene la catena laterale abbia un carattere polare non carico, ha la particolarità di costitutire un legame covalente con un’altra cisteina, per formare ponti disolfuro (S‐S), che irrigidiscono la catena.
PROLINA. Sebbene la catena laterale abbia carattere polare non carico, ha la particolarità di creare snodi nelle catene polipeptidiche ed interrompere la struttura secondaria ordinata
14/03/2014
8
Ponti disulfuro (S-S) tra residui di cisteina
Questi legami incrociati possono collegare sia due parti della stessa catena polipeptidica che due catene polipeptidiche diverse. Poiché l’energia necessaria per rompere un legame covalente è molto superiore all’energia necessaria per rompere persino un intero insieme di legami non‐covalenti, un legame disulfuro può avere un effetto stabilizzante notevole in una proteina.
LEGAME PEPTIDICO
Amminoacidi
Formazione del legame peptidico (1) Formazione del legame peptidico (2)
(A) I legami peptidici si formano fra il gruppo NH2 di un aminoacido e il gruppo COOH di un altro, con la formazione e perdita di una molecola di acqua. Rn, catena laterale dell’aminoacido.
(B) Una proteina ha un’impalcatura polipeptidica con diversi gruppi laterali degli aminoacidi
H. Kreuzer & A. Massey: Biology and Biotechnology: Science, Applications and Issues ASM Press, Washington, D.C., 2005.

14/03/2014
9
Gli amminoacidi hanno un’impalcatura idrofilica che può formare catene e uno fra i 20 diversi tipi di catene laterali, che possono essere idrofiliche o idrofobiche.
Una proteina è una catena di aminoacidi che si piega in una conformazione tridimensionale specifica.
H. Kreuzer & A. Massey: Biology and Biotechnology: Science, Applications and Issues ASM Press, Washington, D.C., 2005.
STRUTTURA TRIDIMENSIONALE DELLE
PROTEINE
STRUTTURA PROTEINE (3)
http://en.wikipedia.org/wiki/Protein_structure
STRUTTURA PRIMARIA DELLE
PROTEINE (1)
E’ la sequenza lineare specifica degli AA che compongono la catena.Determina da sola il ripiegamento della proteina.Con 20 diversi AA il n° di differenti polipeptidi che si possono formare è di 20n
dove n è il n° di AA della catena.
http://w3.hwdsb.on.ca/hillpark/Departments/Science/Watts/SBI3U/Class_Summary/class_summary_spring_2009_sbi3u.html

14/03/2014
10
Struttura secondariaα‐elica & foglietto β
La struttura secondaria rappresenta la conformazione ordinata che alcuni tratti di proteina possono assumere, sulla base della struttura primaria, cioè della sequenza aminoacidica.
La struttura secondaria è caratterizzata dalla presenza di ponti idrogeno fra i gruppi del legame peptidico di residui non adiacenti, mentre non sono direttamente coinvolte le catene laterali degli aminoacidi.
All'interno della stessa proteina, diversi tratti possono assumere la medesima struttura secondaria o strutture secondarie differenti. Le principali forme di strutture secondarie presenti nelle proteine sono l'α‐elica e le strutture β foglietto. http://www.unisr.it/biotechbook/view.asp?id=250
IMPORTANZA DEI PONTI DI IDROGENO PER LA FORMAZIONE DI UN’ELICA E DI ALTRE STRUTTURE
ORDINATE
Una elica si forma quando una serie di subunità si legano una all’altra in modo regolare
α‐elica (1)
α‐elica (2)

14/03/2014
11
β‐foglietto (2) β‐foglietto (3)
Note sul ripiegamento delle proteine (1)
L’acqua contiene due legami polari ossigeno‐idrogeno ed è una molecola estremamente polare.Perciò si associa “confortevolmente” con altre molecole polari o cariche elettricamente.Per questa ragione, le molecole che sono elettrostaticamente cariche o polari sono IDROFILICHE.Poichè le molecole non polari non si associano “confortevolmente” con l’acqua, esse sono IDROFOBICHE.Le catene laterali idrofobiche (non polari) degli amminoacidi non si associano stabilmente con il fluido intracellulare (o extracellulare).
Note sul ripiegamento delle proteine (2)
Viceversa, le catene laterali idrofiliche degli amminoacidi (cariche o polari) si possono associare stabilmente con il fluido perchè le loro cariche, o cariche parziali possono essere neutralizzate dalle cariche parziali complementari delle molecole polari dell’acqua.
Una regola basilare che determina la struttura delle proteine in ambiente acquoso è, per quanto possibile, il ripiegamento dei gruppi laterali idrofobici concentrandoli all’interno della proteina, così creando un ambiente idrofobico privo di acqua.
Le catene laterali idrofiliche sono invece stabili quando esposte al citoplasma sulla superficie della proteina.

14/03/2014
12
Note sul ripiegamento delle proteine (3)
Si dice perciò che una proteina in un ambiente acquoso contiene una zona centrale (“core”; nocciolo) idrofobica e stabile.La struttura tridimensionale di ogni singola proteina (STRUTTURA TERZIARIA) può essere vista come la migliore soluzione al problema di creare la zona centrale idrofobica per ogni struttura primaria.Questo presenta un ulteriore problema: l’impalcatura/asse comune (sequenza di legami peptidici) contiene un gran numero di legami NH e CO, che sono altamente polari.
Note sul ripiegamento delle proteine (4)
Alla superficie della proteina questi legami parzialmente carichi possono essere prontamente neutralizzati mediante legami di idrogeno con l’acqua.
Tuttavia, perchè una struttura proteica sia stabile le cariche parziali dell’impalcatura polipeptidica debbono essere neutralizzate anche all’interno della proteina, dove l’acqua non è presente.
Note sul ripiegamento delle proteine (5)
La soluzione di questo problema è un fattore di importanza fondamentale che determina la struttura della proteina:
L’asse della proteina deve neutralizzare le sue stesse cariche parziali.
I gruppi NH possono formare legami d’idrogeno con i gruppi CO, neutralizzandosi a vicenda.
Per costrizioni geometriche, i gruppi CO e NH dello stesso amminoacido non sono in posizione tale da poter formare ponti d’idrogeno l’uno con l’altro.
Viceversa, l’asse polipeptidico deve essere disposto accuratamente in posizione tale che gruppi NH e CO lungo l’asse siano in posizione da potere formare ponti d’idrogeno con gruppi complementari in altre posizioni lungo l’asse.
L’‐elica e il foglietto (STRUTTURE SECONDARIE) sono le due disposizioni più comunemente riscontrate nelle proteine che permettono la formazione dei legami d’idrogeno.
Struttura terziaria

14/03/2014
13
Struttura terziaria
Struttura terziaria
Three‐dimensional structure of proteins
Tertiary structure
Quaternary structure

14/03/2014
14
Struttura terziaria
http://chemistry.umeche.maine.edu/CHY431/Proteins8.html
Gli elementi strutturali (spirali e tornanti, eliche, filamenti e stratti) si combinano a formare «motivi».I motivi a loro volta si combinano a formare «domini». Le proteine di piccole dimensioni possono formare un solo dominio.Le proteine di maggiori dimensioni sono combinazione di domini. La struttura prodotta dall’organizzazione di elementi strutturali in domini nella struttura globale viene chiamata struttura terziaria della proteina.In genere i domini si comportano come se potessero avere esistenza e stabilità indipendenti.
Fosfoglicerato chinasi
Karp – Biologia cellulare e molecolare – Concetti ed esperimenti
Struttura quaternaria
http://www.answers.com/topic/hemoglobin
Struttura quaternaria (1)
Molte proteine contengono più di una catena polipeptidica.
L’interazione tra queste catene sta alla base della struttura quaternaria.
Le interazioni sono esattamente le stesse che determinano la struttura terziaria (ponti S‐S, ponti di idrogeno, interazioni ioniche e interazioni idrofobiche) solo con l’eccezione che hanno luogo fra una o più catene polipeptidiche, dette «subunità».
D. Whitford: «Proteins: Structure and Function». Wiley, 2005.
Struttura quaternaria (2)
Può essere basata su subunità identiche o subunità diverse:
Omodimeri: es. triosifosfato isomerasi (enzima coinvolto nella glicolisi), HIV proteasi, molti fattori di trascrizione.
Trimero: es. proteina MS2 del capside virale
Tetramero: es. emoglobina, con due diverse subunità: 2 subunità α e 2 subunità β.

14/03/2014
15
Struttura quaternaria (3)
La corretta attività funzionale della proteine richiede la formazione della struttura quaternaria e la specifica associazione della subunità.
Nonostante singolarmente le forze siano deboli, esse sono numerose e portano all’assemblaggio delle subunità ed ad aumentata stabilità.
La struttura quaternaria permette la formazione di siti di catalisi o di legame nell’interfaccia fra le unità; tali siti sono impossibili da trovare nelle proteine monomeriche.
Struttura quaternaria (4)
Ulteriori vantaggi derivano dal fatto che il legame con il substrato della reazione catalitica o con il ligando provoca alterazioni conformazionali all’interno di tutto il complesso e offre la possibilità di regolazione dell’attività biologica – BASE PER LA REGOLAZIONE ALLOSTERICA DELLE PROTEINE.
Permette quindi una grande versatilità di funzioni.
HIV proteasi
http://upload.wikimedia.org/wikipedia/en/1/1d/HIV_protease_1EBY.png
Complesso fra fattore di trascrizione T‐box /DNA
http://www.embl‐grenoble.fr/groups/dna/t.gif
Triosifosfato isomerasi
http://en.wikipedia.org/wiki/Triosephosphate_isomerase
Dimero della Triosifosfato isomerasi

14/03/2014
16
Altre proteine con struttura quaternaria (1)
Integrina
Recettore ad attività chinasica (cross‐fosforila residui di tirosina o serina/treonina)
Mioglobina e emoglobina
Mioglobina
Proteine di membrana
http://www.nfsdsystems.com/w3bio315/; http://www.ncbi.nlm.nih.gov/books/NBK28193/figure/A105/?report=objectonly

14/03/2014
17
Proteine di membrana Tipica proteina con diversi legami S‐S (1)
Tipica proteina transmembrana a passaggio singolo “single‐pass”. Si noti che la catena lipidica attraversa il doppio strato lipidico come α‐elica destrogira e che le catene oligosaccaridiche e i legami disolfuro sono tutti sulla superficie non citosolica della membrana. I legami disolfuro non si formano fra i gruppi sulfidrilici nel dominio citoplasmatico della proteina, perchè l’ambiente riducente del citosol mantiene questi gruppi nella loro forma ridotta (‐SH).
Tipica proteina con diversi legami S‐S (2)
Una molecola di anticorpo. (A) Una tipica molecola di anticorpo ha la forma a Y e ha due siti di legame identici per il suo antigene, uno in ciascuna delle braccia della Y. La proteina è composta da quattro catene polipeptidiche (due catene pesanti identiche e due catene leggere identiche e più piccole) tenute insieme da legami disulfuro. Ogni catena è fatta da diversi domini di tipo immunoglobulina, qui ombreggiati sia in azzurro che in grigio. Il sito di legame con l’antigene si forma laddove un dominio variabile della catena pesante (VH) e un dominio variabile della catena leggera (VL) vengono a contatto. Questi sono i domini che differiscono di più in sequenza e struttura nei diversi anticorpi.
Le proteine interagiscono con le membranein tre modi diversi
• In base alla loro posizione rispetto alla membrana:
– Integrali
– Ancorate a lipidi
– Periferiche

14/03/2014
18
http://www.ncbi.nlm.nih.gov/books/NBK28193/figure/A103/?report=objectonly
Questa struttura induce le catene laterali degli aminoacidi a proiettarsi radialmente.
Quando diverse eliche α sono impacchettate strettamente le loro catene laterali possono essere interconnette oppure delle costrizioni stereochimiche possono provocare la formazione di canali all’interno delle catene.
I residui che si proiettano all’esterno devono essere predominantemente idrofobici per interagire con le catene di acidi grassi dei bilayers lipidici.
Il bilayer ha uno spessore di circa 3 nm. Ogni residuo peptidico si estende all’interno dell’ α elica per 1.5 Å. Perciò, nonostante modificazioni locali del bilayer o interazioni con altri polipeptidi di membrana possano alterare questo requisito, i segmenti transmembrana di solito richiedono circa 20 residui aminoacidici per attraversare totalmente il bilayer.
Le proteine integrali di membrana sono caratterizzate dalla presenza di segmenti idrofobici con approssimativamente questa lunghezza.
I domini transmembrana delle proteine integrali di membrane sono predominantemente delle eliche α
Nella maggior parte delle proteine transmembrana la catena polipeptidica attraversa il doppio strato lipidico
in conformazione ad α‐elica (1)
Una proteina transmembrana ha sempre un orientamento
caratteristico nella membrana.
Questo riflette il modo asimmetrico con cui è sintetizzato ed
inserito nel doppio strato nel Reticolo Endoplasmatico ruvido e le
diverse funzioni dei suoi domini citosolici e non‐citosolici.
Questi domini sono separati da segmenti della catena polipeptidica
che attraversano la membrana e che sono in contatto con
l’ambiente idrofobico del doppio strato lipidico e sono composti in
gran parte di residui di aminoacidi con catene laterali non polari.

14/03/2014
19
Proteine transmembrana (2)
Poichè i legami peptidici stessi sono polari e dato che l’acqua è assente, tutti i legami peptidici nell’ambito del doppio strato sono portati a formare legami di idrogeno gli uni con gli altri.
Il legame di idrogeno fra i legami peptidici viene massimizzato se la catena polipeptidica forma una ‐elica regolare nell’attraversamento; si ritiene che sia in questo modo che la grande maggioranza dei segmenti che attraversano la membrana delle catene polipeptidiche attraversino il doppio strato. ‐eliche transmembrana
… ma anche i ‐foglietti si adattano all’attraversamento della membrana
Proteine ancorate alla membrana da lipidi
http://www.ncbi.nlm.nih.gov/books/NBK9898/figure/A1979/?report=objectonly

14/03/2014
20
Ancore lipidiche Associazione proteine con lipidi di membranaAlcune proteine citosoliche hanno domini che si legano alle teste polari di lipidi che occorrono transientemente nella membrana.
Gli enzimi che creano o degradano questi lipidi sono soggetti a regolazione mediata da segnali, fornendo un meccanismo per modulare l’affinità di una proteina verso la superficie di una membrana:
Ad es. i domini “pleckstrin homology”, (PH)” sono in grado di legare il fosfatidilinosiltolo.
Alcuni domini PH si legano al PIP2 (PI‐4,5‐P2).
Altri domini PH riconoscono e si legano a derivati del foafatidilinositolo con gruppi Pi esterificati con il gruppo 3’‐OH dell’inositolo.
ES: PI‐3‐P, PI‐3,4‐P2, e PI‐3,4,5‐P3.
http://www.rpi.edu/dept/bcbp/molbiochem/MBWeb/mb1/part2/lipid.htm#animat1
Le associazioni alla membranapossono aver luogo mediante legame selettivo
delle proteine a gruppi polari della testa dei lipidi
Un esempio è la spettrina, che si lega al fosfatidilinositolo‐4,5‐bisfosfato rivolto verso il citoplasma mediante un dominio “pleckstrin homology”, (PH)” .Diversi enzimi e proteine strutturali si legano alla membrana in seguito ad attivazione da Ca2+. Es: proteina chinasi C (PKC), fosfolipasi A2 e sinaptotagmina.Una regolazione allosterica dell’idrofobicità delle superficie di legame delle proteine ha spesso luogo. Uno dei casi meglio noti è il legame Ca2+‐dipendente della calmodulina ad altre proteine.
Le annessine sono una famiglia di proteine che formano associazioni Ca2+‐dipendenti con la membrana cellularemediante interazione diretta con fosfolipidi; vice‐versa, le interazioni con i fosfolipidi aumentano la loro affinità verso il Ca2+.
http://www.ncbi.nlm.nih.gov/books/NBK28193/
http://www.geneticaebiologia.unito.it/membrana/globulo_rosso.jpg
spettrina

14/03/2014
21
Ancore lipidiche
http://www.ncbi.nlm.nih.gov/books/NBK21570/#A611
Glicosilfosfatidil inositolo (GPI)
Ancore lipidiche (a)
Alcune proteine si legano alla membrana mediante un’ancora lipidica legata covalentemente, che si inserisce nel bilayer.
Una proteina può legarsi al versante citosolico della membrana plasmatica mediante un acido grasso legato covalentemente (ad es. palmitato o miristato) oppure un gruppo isoprenoide.
Il palmitato di solito si collega mediante un legame tioestere al gruppo –SH di un residuo di cisteina. Una proteine può essere rilasciata dalla membrana mediante idrolisi del legame esterico – depalmitoilazione.
Un gruppo isoprenoide, come ad es. un residuo farnesilico, è legato ad alcune proteine mediante un legame tioesterico ad un gruppo tiolico di una cisteina.

14/03/2014
22
Ancore lipidiche (b)
Il miristato può essere aggiunto co‐traduzionalmente alla glicina N‐terminale di un gran numero di proteine periferiche, participando perciò al loro legame con la superficie citoplasmatica della membrane.
La subunità catalitica della protein chinasi c‐AMP dipendente, la calcineurina B la NADH‐citocromo b5 reduttasi sono proteine miristoilate.
Gli acidi grassi, soprattutto il palmitato, possono legarsi sotto forma di tioesteri ad un residuo di cisteina che di solito è localizzato vicino ad un dominio di legame con la membrana.
Sia proteine integrali di membrana (es. rodopsina o recettore per la transferrina) e proteine associate alla membrana (es. anchirina e vinculina) possono essere acilate.
Un gran numero di proteine può essere prenilata post‐traduzionalmente.
Una via biosintetica per le proteine prenilate coinvolge proteine precursore con una sequenza C‐terminale CXXX (C: cisteina). Un gruppo C20 di un geranilgeranil pirofosfato viene aggiunto al gruppo sulfidrilico della cisteina. I tre AA terminali vengono ins eguito rescissi e infine un gruppo metilico viene aggiunto al gruppo cisteina‐α‐carbossile esposto.
Le proteine prenilate includono diversi trasduttori di segnale della classe delle piccole proteine G e subunità γ di proteine G trimeriche.
http://www.ncbi.nlm.nih.gov/books/NBK28193/
Ancore lipidiche (c)
I glicosilfosfatidil inositoli (GPI) sono glicolipidi complessi che si legano ad alcune proteine presenti sulla superficie esterna della membrana plasmatica. Il loro legame è simile a quello indicato sotto, nonostante la composizione dell’oligosaccaride possa variare:
Proteina (C‐terminale) ‐ fosfoetanolamina – mannosio ‐mannosio ‐mannosio ‐ N‐acetilglucosamina ‐ inositolo (di fosfatidilinositolo inserito nella membrana)
La proteina è ancorata ad una certa distanza all’esterno della membrana dalla lunga catena oligosaccaridica.
Le proteine legate a GPI possono essere rilasciate dalla superficie esterna delle cellula dalle fosfolipasi.
Ancore lipidiche (d)
Le proteine possono essere ancorate al foglietto esterno mediante legame covalente con fosfoinositidi glicosilati complessi.
Le proteine ancorate a glicosilfosfatidilinositolo (GPI) includono la fosfatasi alcalina, la 5’‐nucleotidasi, un’isoforma dell’acetilcolinesterasi e un’isoforma della N‐CAM.
Lodish et al., 7° ed., 2013