
ERGOCINETICHE E CHEMIO-BIO CINETICHE - unive.it · studio s'innesta nella linea dell'ecotossicolgia...
55
G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.1 Cap.VIII ERGOCINETICHE E CHEMIO-BIO CINETICHE
Transcript of ERGOCINETICHE E CHEMIO-BIO CINETICHE - unive.it · studio s'innesta nella linea dell'ecotossicolgia...

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.1
Cap.VIII
ERGOCINETICHE E
CHEMIO-BIO CINETICHE

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.2
8.0.0.0.- Ergo-cinetiche e Chemio-bio-cinetiche1. Xenobiotici. Inquinamento e bilanci energetici. Il trade-off
L’ergo-cinetica é lo studio dell’assunzione, dell’accumulo, della distribuzione, della trasformazione
e dell’eliminazione dell’energia negli organismi viventi.
La tossico-cinetica é lo studio delle velocita’ (cinetiche) delle reazioni ambientali con particolare
riguardo ai processi d'assunzione, accumulo, distribuzione e di release (eliminazione) dei composti
chimici xenobiotici (e non) immessi negli ecosistemi da parte degli organismi viventi.1
Un composto xenobiotico è definito come un composto estraneo ad un organismo ossia che non
ha un ruolo essenziale nei processi biochimici di quell’organismo. Da cio’ discende che un composto
chimico normale per un organismo può essere xenobiotico per un altro. Così composti xenobiotici
possono esistere anche d’origine antropogenica originati forse fin dall’inizio della vita evolutiva nel
nostro mondo terrestre.
Da un punto di vista evolutivo l’esistenza naturale di composti xenobiotici che potremmo
considerare aggressivi chimici, è di considerevole interesse. Per esempio si conosce l'evoluzione dei
meccanismi di detossificazione che gli animali hanno sviluppato nei confronti di xenobiotici prodotti da
piante.
É chiaro, peraltro, che noi ci occuperemo prevalentemente dei composti xenobiotici non
naturalmente prodotti ma di quelli che, originati dall’attivita’ industriale (seppur connessa con l'attivita’
antropica della vita comune), si sono immessi negli ecosistemi pervenendo, come target finale,
all’uomo. É comunque importante ricordare che composti xenobiotici esistenti naturalmente negli
ecosistemi come, ad esempio, le piretrine, la nicotina, vari tipi di micotossine ecc., seguono, nelle
strutture biologiche, gli stessi processi tossicocinetici dei composti xenobiotici prodotti dai processi di
sintesi industriale sia come materie prime che come cataboliti della stessa attivita’ processuale.
LA CHEMIO-BIO-CINETICA é una parte fondamentale dell’ecotossicologia perchè permette la
comprensione e la previsione del comportamento di sostanza inquinanti negli organismi viventi. Il suo
1. Viene usato spesso anche il termine tossicocinetica ma il termine tossico non é dei migliori in quanto indicherebbe un’azione tossica (e non solo chimica). Invece i processi studiati in queste cinetiche sono validi anche

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.3
studio s'innesta nella linea dell'ecotossicolgia che, come abbiamo visto nei capitoli precedenti, studia
la dinamica ambientale ossia il movimento degli xenobiotici nell’ecosistema globale (modello LI (f) ER),
nell’aria, nell’acqua e nel suolo.
Anche se il processo che chemio-bio-cinetico é prevalentemente d’interesse per i composti organici
lipofili (in grado quindi di ripartirsi prevalentemente nella fase lipidica) chiaramente i cammini di tutti i
composti chimici sono gli stessi; varieranno solo le velocita’ dei processi d’assunzione, di distribuzione
e di rilascio.
L’ERGO-CINETICA, a livello cellulare, é di pari importanza della tossicocinetica se non in misura
superiore. Infatti come un composto chimico si ripartisce nelle varie componenti dell’organismo, così
fa l’energia condizionandone la vita e la sopravvivenza.
Anche l’energia, assunta tramite l’alimentazione ed eliminata nella forma piu’ degradata (calore)
come catabolita, segue i principi della termodinamica (pare ovvio!) che regolano il comportamento
della tossicocinetica.
In ergocinetica un parametro importante da sviluppare é quello dei bilanci d’energia negli organismi
espresso dal concetto di trade-off. Come vedremo tale concetto fornira’ un’ottica nuova per valutare e definire i processi
d’inquinamento e l’inquinamento medesimo attraverso l’inferenza e l'interferenza con i processi vitali
d'energia degli organismi.
É un concetto nuovo e forse poco comprensibile di primo acchito ma che s’inserisce perfettamente nei principi dell’energia che abbiamo sviluppato e sottolineato fin dai primi capitoli di questo trattato e di cui, qui di seguito, parleremo più estesamente.
Il termine trade-off, mutuato dalle scienze economiche, in Ecotossicologia intende il bilancio globale di materia e quindi d'energia che un sistema biologico realizza durante la sua esistenza
e che corrisponde ad un dare-avere tra l'energia assunta attraverso l'alimentazione (o in alcuni casi
attraverso i raggi solari) e l'energia consumata nei processi biochimici essenziali: metabolismo basale, sintesi e accrescimento, manutenzione cellulare e tissutale, escrezioni e produzione (riproduzione), come riportato nello schema successivo.
BOX 8.1 per composti “benefici” nei confronti dell’organismo bersaglio (es.: farmaci).

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.4
Metabolismo basale
Costi energetici per la detossificazione
Energia assorbita Costi metabolici di sintesi
Riproduzione = Scope for growth
Escrezioni
I processi d’inquinamento vanno visti in termini ecotossicologici come interferenti nell’equilibrio di
trade-off del sistema biologico interessato.
L’inquinamento si pone come processo competitivo dei processi base energetici e, quindi,
puo’ essere considerato come antagonista nell’uso delle energie destinate agli obiettivi piu’ nobili dei
processi vitali.
In sostanza, come vedremo, la cellula vitale considera il composto inquinante (o, in termini piu’
generali, lo stressore chimico o fisico o psicologico che sia) come una molecola estranea alla sua
funzionalita’ biologica, corpo estraneo che si oppone alla stessa sua sopravvivenza. E l’istinto, a
quest’ultima legato, lo induce ad usare quanta piu’ energia possibile per mettere in essere tutte le
difese biochimiche (e quindi energeticamente depauperanti) per eliminare l’agente xenobiotico ostile.
Ovviamente le energie utilizzate e consumate nel processo di difesa non sono piu’ disponibili ( o
lo sono i misura ridotta) per gli altri processi vitali con rallentamento se non con inibizione totale degli
stessi.
Tutti sono ben coscienti che una malattia infettiva ci lascia molto deboli e debilitati: abbiamo una
prova evidente che uno stressore (il biochimismo dell’agente patogeno) ha consumato parte delle
nostre energie (anche se aiutato da un “contributo energetico” dei farmaci) lasciando l’organismo in
carenza d'energia. Una febbre violenta, che passi da sola senza l’aiuto di terapie, lascia ancora di piu’
l’organismo a livello di disponibilita’ energetiche assai basso.2
L’interferenza dello stressore puo’ essere così elevata ed intensa da provocare non solo
rallentamenti nello sviluppo somatico e cerebrale ma anche impedire la normale riproduzione. E se
l'inquinamento si traduce in una perdita di riproduzione, danneggia anche il meccanismo
d’acquisizione delle risorse e d'accumulo fino alla negazione della possibilità di produrre energia per la
detossificazione. In questo secondo caso si riducono le possibilità di morte pagando il costo in termini
di perdita di riproduzione. In pratica tutte le risorse energetiche, incluse una parte di quelle destinate
alla riproduzione (ma non la parte pregiata d'energia, gelosamente stoccata e tenuta di riserva per il
2 Basti pensare al pacchetto energetico di “calore” che la febbre provoca e che é dato dai processi di difesa dell’organismo che, aumentando la temperatura, tentano di rendere condizioni inibitorie per lo stressore patogenico

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.5
processo finale della riproduzione stessa, (che non viene e non puo' essere utilizzata se non per lo
specifico obiettivo), vengono indirizzate contro la minaccia alla propria sopravvivenza rappresentata
dallo stressore.
In tale processo tutti i "soldati" d'energia vengono inviati al “fronte” e le riserve, "in caserma" non
sono in grado di attivare i processi di riproduzione per i quali esisterebbero, comunque, le risorse
energetiche.
In tal modo l'organismo tenta di mantenere la sua propria esistenza a discapito della
procreazione di nuovi individui che, come sappiano, é il destino entropico del sistema biologico.
Suo malgrado, ma in una logica stringente in termini d'energia, l'organismo scambia la perdita di produzione (riproduzione) con la riduzione del tasso di mortalità.
In un certo modo si potrebbe pensare a questo scambio tra tasso di riproduzione e tasso di
mortalità come ad un affare commerciale coatto (trade-off) che si sviluppa in una nazione in
condizioni di crisi (guerre ecc.) tra l'industria della produzione e quella della difesa, perchè i
meccanismi che servono a difendere la sopravvivenza della nazione stessa attingono alle stesse
risorse necessarie per un armonico sviluppo economico e tecnologico della nazione che, di
conseguenza, si ferma (morbilita' economica ed ambientale) quand'anche non arrivi alla completa
distruzione (mortalita' economica ed ambientale).
Questo modo d'intendere l'inquinamento ben s’inquadra nel concetto di "Scope for Growth" (SFG) definito come la differenza tra l'accumulo d'energia e le perdite metaboliche totali.
Studi in campo e in laboratorio hanno confermato come le conseguenze a lungo termine sulla
crescita e sulla sopravvivenza degli individui possano essere predette da effetti misurati sul bilancio
d’energia a livello d’individuo.
Ad esempio Widdows e Donidri (1991) hanno rilevato come la causa della riduzione nell' SFG in
Mytilus edulis in luoghi contaminati possa essere assegnata ad inquinanti specifici.
Il modo con cui gli animali si difendono dagli inquinanti (tossici od interferenti) è di varia natura: il
principale è quello di copularli con una struttura chimica eliminabile con i liquidi organici, e quindi
permetterne la fuoriuscita dal corpo cellulare.
Un secondo sistema è quello di bloccarli in situ dopo averli resi innocui, o attraverso una forma di
riduzione (precipitazione di granuli di metallo nella cellula o in siti intracellulari), o, inglobandoli in
proteine di difesa (metallotioneine).
Ambedue i processi possono coesistere e, come sempre, prevarrà quello che energicamente sarà
più favorito. Tutti questi processi sono naturalmente a costo energetico più o meno elevato a seconda
dell'intensità del processo stesso depauperando il budget energetico di base di cui abbiamo
precedentemente parlato.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.6
8.1.0.0.- Processi ergo-cinetici: I modelli energetici
Il trade-off si puo’ studiare attraverso vari modelli energetici. Nella nostra trattazione considereremo
due classi d'animali: i molluschi ed i pesci, cui applicheremo le considerazioni bio-energetiche.
In particolare ci occuperemo dei processi d'assunzione/eliminazione che saranno oggetto della
seconda parte del capitolo relativa alle tossicocinetiche dei nutrienti.
Nel caso dei molluschi il processo d’assunzione/eliminazione in termini energetici è stato
proposto in maniera ottimale da Kooijman3 che ha tentato di spiegare numericamente le modifiche
delle condizioni fisiologiche dell'animale. Tale modello, noto come Dynamic Energy Budget (DEB),
descrive le dinamiche energetiche in funzione della crescita e della riproduzione dell'animale.
In tale modello si considera l'assunzione del cibo direttamente dall'ambiente e l'eliminazione del
tossico attraverso la riproduzione tramite l'emissione dei gameti nell'ambiente, oltre alle vie principali
d’accumulo ed eliminazione
Un fattore che appare importante nel modello è il contenuto in lipidi nel tessuto che sembra
condizionare la velocità d’assunzione e di rilascio.
Poichè il contenuto in lipidi nell'animale varia nel tempo, è presumibile pensare che essi influenzino
la concentrazione del tossico nell'animale anche quando il tossico permane costante nell'ambiente nel
quale l'animale vive.
Ovviamente l'azione dei lipidi sarà svolta principalmente a livello del doppio strato lipidico della
membrana, per cui l'influenza si avrà a livello del flusso di materia attraverso la membrana in direzione
del citoplasma.
Un punto relativamente critico del modello DEB, è che le condizioni fisiologiche dell'individuo
utilizzato per lo studio non vengano alterate dal composto utilizzato nell'esperimento.
É ovvio pensare che la presenza di un tossico possa altresì influenzare le risposte fisiologiche
dell'animale e quindi è importante sapere qual è il limite critico al quale questo processo ha luogo. Nel
caso dei mitili, secondo la sperimentazione valori dell'ordine dei 100 µg l-1 non influenzano
negativamente i processi fisiologici della respirazione, della crescita e della ingestione del tossico.
L’interpretazione di dati ecotossicologici e di monitoraggio ambientale è stata perfezionata dalla
conoscenza della relazione tra la concentrazione degli elementi xenobiotici nell’ambiente e nei tessuti
dei biota.
La maggior parte degli studi sulle cinetiche di assunzione e di eliminazione ammettono
implicitamente delle condizioni di stato stazionario per gli altri processi fisiologici nell’organismo. I dati
3 Kooijman et al., 1990

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.7
di monitoraggio in campo e sperimentali sono quindi il più delle volte analizzati con modelli di
assunzione e rilascio monocompartimentali.
Questi modelli però non si adattano sempre bene ai dati ottenuti, specialmente quando l’organismo
cambia condizioni fisiologiche con un andamento paragonabile all’assunzione/eliminazione degli
elementi xenobiotici come nel caso del ciclo riproduttivo o del cambiamento di taglia o di riserve
d’energia4.
Il modello d’assunzione/eliminazione proposto da Kooijman e van Haren5 vuole spiegare i
cambiamenti nelle condizioni fisiologiche (alimentazione/lipidi) dell’organismo.
É basato sul modello Dynamic Energy Budget (DEB)6 che descrive crescita, dinamiche
energetiche e riproduzione come funzione della taglia corporea.
L’assunzione è via cibo e/o direttamente dall’ambiente e l’eliminazione è via riproduzione
(gameti) e/o direttamente nell’ambiente. Le velocità d’assunzione e di rilascio dipendono dal contenuto
in lipidi, che cambia nel tempo, influenzando quindi il carico corporeo anche quando la concentrazione
nell’ambiente è costante.
Nel caso dei molluschi, l’animale, scelto come esempio, é il Mytilus galloprovincialis (o aedulis) - su
cui Ca’Foscari ha sviluppato numerose ricerche - di cui si assume, per semplicità, che le condizioni
fisiologiche stesse non vengano influenzate dai composti inquinanti in esame.
Il carico di xenobiotico in M. edulis è influenzato da molte variabili fisiologiche tra cui temperatura,
taglia7 e ciclo riproduttivo: sono state riscontrate correlazioni tra mitili della stessa taglia e quantità di
cibo8. La concentrazione di xenobiotico viene di solito espressa come peso secco del tessuto e dato
che i pesi dei vari tessuti variano con la stagione e la temperatura, le variazioni di concentrazione del
cadmio sono difficili da interpretare.
Infatti il peso secco riflette in parte la condizione in cui si trovano le gonadi e le riserve di energia,
in parte la taglia del mitilo. Interessanti studi sui PCB hanno dimostrato, ad esempio, come il carico di
PCB aumenti all’aumentare della lunghezza della conchiglia e diminuisca con l’emissione dei gameti.
Lo sviluppo del modello tiene conto soltanto della parte dell’alimentazione dividendo uno stadio
giovanile pre-riproduttivo ed uno stadio adulto riproduttivo. Vengono distinte quattro frazioni separate
del corpo: la frazione acquosa (p.e. sangue), la componente strutturale, le riserve accumulate di
energia, e una componente a parte per la riproduzione (i mitili perdono 40-70 % del peso umido
durante la fecondazione, che recuperano durante tutto il resto dell’anno).
4 McDowell Capuzzo et al., 1989 5 Kooijman et al., 1990 6 Kooijman S.A.L.M., 1986, 1988 e 1993 7 Simpson R.D.,1979; Boalch et al., 1981 8 Jansen et al., 1979; Borchardt et al., 1983 e 1985; Riisgard et al., 1987

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.8
8.1.1.0.- Processi ergo-fisiologici
L’assunzione del cibo è proporzionale all’area di superficie (dell’apparato filtratore e dell’intestino)
del mitilo e segue un responso funzionale del tipo II di Holling
f =X
K + X (8.1)
dove f indica il responso funzionale scalare, X la quantità ambientale di cibo e K la costante di
saturazione. Si considera costante la conversione cibo-energia dell’intestino. L’energia entrante si
aggiunge a quella delle riserve, che segue un processo del primo ordine:
ddt
e = vV −1/ 3( f − e) (8.2)
dove e è la densità pesata di immagazzinamento, V è il volume strutturale (lunghezza3) e v è la
conduttività elettrica (l t-1) (p.e.: una misura di resistenza per il flusso di energia dall’acquisizione
energetica all’utilizzazione dell’energia). L’energia utilizzata viene presa dalla parte immagazzinata di cui una frazione fissa k è usata per la crescita e il mantenimento, che è considerato
proporzionale al biovolume. Conseguentemente, la crescita è data da una differenza pesata tra l’area
di superficie e il volume:
ddt
V =V 2/ 3ev − Vgm
e + g (8.3)
dove g è un rapporto di investimento adimensionale (costi energetici per nuovo biovolume in
relazione al massimo di energia disponibile per la crescita più il mantenimento) ed m la costante del
contributo al mantenimento (t-1) (rapporto dei costi per il mantenimento e sintesi di biovolume). La crescita si blocca quando c’è un calo delle riserve d’energia:

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.9
e < V1/3mg / v (8.4)
Una frazione 1 - k dell’energia usata per l’immagazzinamento è usata nello sviluppo più la riproduzione. Si assume che il mantenimento allo stadio adulto sia pari a (1 - k ) gmVj dove Vj è la
taglia all’inizio dello stadio adulto (lunghezza ). Da questa regola di distribuzione segue che
l’investimento nella riproduzione è dato da:
ddt
G =(1− k)e
e+ g(vgV 2/ 3 + gmV) − (1− k)gmVj (8.5)
e, per organismi che non crescono,:
ddt
G = evV 2 /3 − kgmV − (1 − k)gmVj (8.6)
dove G = Ggo + Gga è il volume delle gonadi e dei gameti (l3). I gameti vengono conservati in un
organulo che viene svuotato una volta all’anno al momento della riproduzione. Ciò causa un
sostanziale cambiamento nel contenuto in lipidi attraverso le stagioni all’interno del mitilo. I gameti
accumulati, Gga, non sono metabolicamente disponibili per altri scopi. La densita' del volume
riproduttivo é r = G/V (8.7) ed il suo cambiamento:
drdt
= V −2 V dGdt
− G dVdt
⎛ ⎝
⎞ ⎠ = V −1 dG
dt− GV −2 dV
dt (8.8)
8.1.2.0. Cinetiche energy-related 8.1.2.1. Cinetiche energy-related - Molluschi
I composti xenobiotici, una volta accumulati dalla frazione acquosa, si ripartiscono nel corpo in

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.10
funzione della fugacita' del composto stesso. L’accumulo, come l’eliminazione, si considerano
proporzionali all’area di superficie dell’organismo V2/3. Quindi l’accumulo dall’acqua (p.e. dalle
branchie) e l’accumulo dal cibo (p.e. dall’intestino) sono entrambe proporzionali all’area della
superficie. L’equazione differenziale che ne risulta per la concentrazione, sulla base del peso umido
(#. massa-1 in cui # indica il numero o il peso delle molecole) è la seguente:
d C[ ]dt
=K1wcd + Kif fcp
ds(1 + α e
−1 + r)V 1/ 3 − C (K2
hV 1/ 3 + V −1 dVdt
+ (1 + α e
−1 + r)−1 drdt
) (8.10)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.11
ove C é la concentrazione dello xenobiotico nel corpo dell'animale (peso umido, K1w e K1f sono le
costanti di accumulo dall’acqua e dal cibo rispettivamente e rad è la costante di eliminazione (L t-1). Le
costanti ds e αe sono la densità specifica delle parti molli e il volume massimo delle riserve
energetiche come frazione della taglia corporea. Una variabile che dipende dal composto è h:
h = γ + (Kp − 1)e + Kpr (8.11)
in cui Kp è un coefficiente di ripartizione del composto xenobiotico tra la frazione acquosa ed il
compartimento corporeo della energia e γ è una costante propria del composto che comprende il
coefficiente di ripartizione tra la frazione acquosa e il compartimento strutturale. Quando Kp >> 1 :
h = γ + Kp (e + r) (8.12)
Le variabili cd e cp indicano rispettivamente le concentrazioni del composto xenobiotico in
soluzione e adsorbite al particolato (# . lunghezza-3 ).

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.12
8.1.2.2. Cinetiche energy-related - Pesci9
La crescita di un animale (nel caso dei sistemi acquatici, prevalentemente i pesci), pur nella sua
complessità, può essere descritta dal punto di vista termodinamico come un insieme di processi
medianti i quali un "sistema aperto" riesce ad aumentare la propria entalpia libera G = H - TS, attraverso la degradazione di una parte dell'entalpia libera "contenuta" nelle molecole dell'alimento.
Espellendo dal proprio organismo l'eccesso di entropia prodotta, attraverso le feci o mediante il
calore, l'organismo riesce a mantenere basso il proprio contenuto entropico, a spese di un aumento
dell'entropia dell'ambiente
In accordo con le leggi della termodinamica, applicate ai sistemi aperti, la crescita può quindi essere
globalmente descritta da un bilancio energetico, riferito ad un certo intervallo di tempo:
R = E + M + SfG (8.13)
Secondo tale relazione, l'entalpia libera R che entra nell'organismo con l'alimentazione in un
determinato intervallo di tempo si ripartisce in una frazione E, a contenuto entropico più elevato, che
rappresenta i prodotti di rifiuto dell'organismo quali sono le urine, le escrezioni branchiali e superficiali,
le feci, ed il calore; una quota M, costituita dall'entalpia libera utilizzata nei processi metabolici,
comprendente le spese per la digestione, la sintesi di molecole utili, attività metaboliche di base e altre
attività volontarie.
Ciò che rimane, viene detto SfG o Scope for Growth, e rappresenta la frazione di entalpia libera
che l'organismo può utilizzare per sintetizzare nuovi tessuti, proteine, grassi, carboidrati.
Occorre precisare che lo SfG non é utilizzato solo per la crescita in dimensioni: lo sviluppo delle
gonadi compete infatti con la crescita dell'individuo, detta crescita somatica, per l'uso delle risorse.
Come si è detto, l'entalpia libera che viene introdotta attraverso l'alimento (R), viene assimilata
solo in parte (RD): la restante frazione viene espulsa attraverso le feci (EF). L'assimilazione della frazione digerita necessita tuttavia processi di trasformazione dei composti
organici attraverso la rottura delle proteine e la formazione di grassi e altre proteine utili all'organismo,
con escrezione di ammoniaca e altri prodotti di rifiuto (urine (EU) e altre escrezioni (EB e ES)) e
produzione di calore denominato Heat lncrement (HI).
Quanto è stato assimilato (RA= R- RD) è utilizzato per il metabolismo basale (MB) con produzione di
9 Questa parte delle dispense é tratta da una tesi di laurea in Scienze Ambientali di cui é relatrice la Prof. Patrizia Torricelli che, cortesemente, ha autorizzato l'uso del testo sovra riportato.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.13
calore, per il movimento ed altre attività (MA): ciò che rimane è lo SfG, che può essere utilizzato per la
crescita somatica e lo sviluppo delle gonadi.
Le componenti di questo bilancio energetico, variano naturalmente da specie a specie, (differenze
interspecifiche), da individuo a individuo, (differenze intraspecifiche) in relazione a differenze del
genotipo e, per lo stesso individuo, possono variare anche in funzione dell'età.
I termini del bilancio dipendono inoltre da una serie di fattori ambientali, quali la temperatura
dell'acqua, la disponibilità e la qualità del cibo, l'ossigeno disciolto, il fotoperiodo, la qualità dell'acqua
e la salinità, che influenzano i processi fisiologici ed, in ultima analisi, la crescita.
Gli esempi di modelli di crescita applicati ai pesci sono numerosi in letteratura e possono essere
sommariamente divisi in due classi: la prima raggruppa i modelli che si possono definire empirici,
poichè descrivono la crescita senza entrare nel merito delle attività fisiologiche mentre i modelli
appartenenti alla seconda classe, definiti bioenergetici, si fondano sul calcolo dei termini del bilancio
3.1.
Un modello matematico bioenergetico dell'accrescimento si propone di quantificare il bilancio, e di
legare i vari termini a variabili osservabili, come ad, esempio, il peso o la taglia dell'animale, la cui
evoluzione temporale si ottiene poi integrando le variazioni istantanee.
La struttura del modello, cioè la forma delle equazioni differenziali che lo costituiscono, viene
definita essenzialmente sulla base della letteratura esistente riguardo la fisiologia e le abitudini
alimentari dell'animale.
L'analisi dei dati sperimentali raccolti in laboratorio o durante esperimenti condotti in campo ma in
condizioni almeno parzialmente controllate consente una prima stima dei parametri del modello, ad
esempio del tasso di respirazione. Successivamente, il modello viene calibrato ed infine validato
mediante il confronto con le osservazioni raccolte sul campo.
Questi due momenti del processo di costruzione del modello matematico sono molto importanti
poiché permettono di verificare quantitativamente le ipotesi utilizzate e di correggere eventualmente i
valori di alcuni parametri stimati in laboratorio mettendo in evidenza le ipotesi semplificative che
consentono di passare dal bilancio energetico ad un bilancio di biomassa.Le principali ipotesi sono:
1. La crescita è isometrica e quindi le proporzioni tra le varie parti del corpo rimangono
costanti; è l'ipotesi fondamentale che consente di considerare i parametri esponenti del peso,
caratteristici della morfologia della specie, costanti per tutta la vita dell'individuo. Significa anche che la
ripartizione dell'energia nell'organismo per metabolismo basale (fasting catabolism) e catabolismo
alimentare (feeding catabolism) nonché l'assimilazione (gross anabolism) sono regolate da funzioni
costanti: non intervengono cioè cambiamenti strutturali con l'età dell'organismo.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.14
2. L'alimento ha la stessa composizione del pesce; è una ipotesi semplificativa che
permette di evitare considerazioni sull'alimento e le sue influenze sulla appetibilità e sulla digeribilità; è
una ipotesi che nel modello viene superata utilizzando coefficienti di conversione: l'indice qualitativo di
trasformazione utilizzato sarà il rapporto tra contenuto energetico del pesce e dell'alimento.
Non c'è esplicita distinzione tra crescita somatica e crescita gonadica, e quindi, teoricamente il
modello proposto rappresenta l'accrescimento in termini di SfG, calcolato come differenza tra i
termini di anabolismo e catabolismo, entrambi quantificati.
Non considerando le spese sessuali, esemplificazione valida se i consumi energetici per l'apparato
riproduttivo sono limitati, lo Scope for Growth risulta approssimabile alla crescita somatica, G =
dw/dt. Per un animale che inizi ad essere maturo sessualmente al secondo anno di vita, la scelta di un
opportuno intervallo temporale di applicazione del modello, permette di accettare questa
semplificazione e di considerare lo SfG uguale alla crescita somatica.
L'accrescimento può essere espresso attraverso una riformulazione della relazione iniziale:
G= (R- Ef) - ( EU+EB+ES+HI+MA) - MB (8.14)
Ed é, quindi, rappresentato dalla somma algebrica dell'alimento digerito, delle perdite sotto forma
di escrezioni, di calore, di attività motorie legate all'alimentazione e del metabolismo basale.
• L'alimento digerito (RD=R-EF) è proposto come funzione del cibo ingerito nell'unità di tempo,
ΦdR/dt), che, a sua volta, dipende dal peso, dalla temperatura e dal tipo di alimento.
• Le escrezioni, l'aumento di calore e l'attività motoria (EU+EB+ES+HI+MA) vengono rappresentate
come funzione dell'alimento assimilato nel termine φ(w,T)detto catabolismo alimentare.
• Il metabolismo basale MB è posto come funzione di temperatura e peso, (γφ).

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.15
R
M A RA
SfG
M BRD
EF
EU
EB ES HI
R = Razione alimentare Rd = Razione digeribile Ra = Razione assimilata Ef = Feci Eu = Urine Eb = Escrezioni branchiali Es = Escrezioni superficiali HI = Calore Ma = Attivita' motorie Mb = Metabolismo basale SfG = Scope for Growth : energia utilizzata per la crescita somatica (G) e gonadica (Ms)
TRADE-OFF (TO) e SCOPE FOR GROWTH (SfG) BILANCIO DI ENERGIA IN UN SISTEMA BIOLOGICO
Fig.8.1 Il bilancio è completato ponendo G, ovvero la formazione di nuovi tessuti, pari a dw/dt, in virtù delle
ipotesi discusse in precedenza:
dwdt
= ΦdRdt
w,T, R( )⎛ ⎝ ⎜
⎞ ⎠ ⎟ − γ Φ
dRdt
w,T,R( )⎛ ⎝ ⎜
⎞ ⎠ ⎟
⎛ ⎝ ⎜
⎞ ⎠ ⎟ − ϕ w,T( ) (8.15)
dove:
• w è il peso fresco dell'animale e dw è il tasso di accrescimento;
• R è la razione alimentare; dR/dt è l'alimento consumato nell'unità di tempo;
• T è la temperatura;
• φdR/dt) è il termine anabolico (gross anabolism): indica la quantità assorbita dall'intestino nell'unità di
tempo; essa è definita come funzione dell'alimento consumato;

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.16
• γ(φ)−φ(w) é il termine catabolico ed indica le spese, nell'unità di tempo dell'organismo; il primo
termine γ(φ) é il catabolismo legato all'alimentazione (feeding catabolism), ed è espresso come
funzione della quantità digerita; il secondo addendo, φ(w) è il metabolismo basale o catabolismo a
digiuno (fasting catabolism), è indipendente dall'alimentazione ed è funzione della taglia del pesce,
espressa dal peso, e della temperatura. Tutti i termini sono influenzati dalla temperatura T.
(1-a) β dR/dt
k(T)wRD
(1-β) dR/dt dR/dt = Razione alimentare a = coefficiente del catabolismo alimentare k(T) = coefficiente del metabolismo basale n = esponente del peso per il metabolismo basale dw/dt = accrescimento in peso La razione alimentare e' definita in funzione del peso e della temperatura = dR/dt = h(T)w
MODELLO BIOENERGETICO SEC.URSIN (1967) CINETICHE DEL BILANCIO DI ENERGIA
dR/dt n
dw/dt
α β dR/dt
n
Fig.8.2 8.2.1.0.- Anabolismo, catabolismo, metabolismo Supponendo che l'animale trovi cibo a sufficienza, la quantità d’energia assorbita nell'unità di
tempo diviene funzione unicamente della taglia dell'animale e della temperatura. Infatti, la quantità di
cibo assorbito é proporzionale alla superficie dell'intestino, sede dell'assimilazione del cibo. D'altra
parte la superficie intestinale dipende dalla taglia, ovvero é proporzionale a wm: l'esponente 0<m<1
esprime la relazione esistente tra superfici ed il peso. Si ha cioè che:

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.17
dRdt
= h(T) × wm (8.16)
dove h(T) esprime la variazione delle necessità alimentari in funzione della temperatura.
Le esigenze nutrizionali possono tuttavia non essere soddisfatte completamente, o per scarsità di
prede o per somministrazioni di alimento in quantità inferiori alle necessità: se ne tiene conto
introducendo il parametro f, chiamato livello di alimentazione, che esprime la frazione di alimento
ingerito rispetto alla quantità necessaria per soddisfare le esigenze nutrizionali.
Il livello d’alimentazione, f, in generale è funzione della disponibilita’ dell'alimento: nel caso degli
allevamenti intensivi esso dipende dalle quantita’ somministrate di mangime e dalla competizione
intraspecifica.
La sua determinazione è invece estremamente complessa in ambiente naturale, in quanto la
razione di cibo che l'animale riesce a procurarsi dipende dalle dinamiche degli organismi predati e
dalla competizione con altre specie.
ll livello di alimentazione assume i valori compresi nell'intervallo 0≤f≤1; f = 0 significa che l'alimento
fornito non soddisfa nessuna esigenza alimentare e rappresenta la situazione di digiuno; f = 1
descrive lo scenario di massima alimentazione, ovvero il completo soddisfacimento dei fabbisogni
alimentari.
La quantità di alimento ingerito nell'unità di tempo diventa quindi:
dRdt
= h(T) × f × wm (8.17)
L'anabolismo è definito come la quantità d’alimento assimilato e dipende dalla quantità di alimento
ingerito e dalla sua qualità. Il coefficiente di assimilazione, β esprime la frazione assorbita dell'alimento
ingerito e può assumere valori compresi tra 0 (alimento non assimilabile) ed 1 (alimento
completamente assimilabile).
Esso dipende dalla composizione chimica dell'alimento ed è una funzione decrescente del livello di
alimentazione f nel caso di basse alimentazioni il coefficiente di assimilazione sarà massimo, mentre
nel caso di massima alimentazione (f = 1) il coefficiente di assimilazione sarà minimo. La frazione di
alimento pari ad (1-β) è quella non assimilata, che viene eliminata attraverso le feci: la misura di

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.18
queste escrezioni consente perciò di risalire al coefficiente di assimilazione.
Il livello di alimentazione, f ed il coefficiente di assimilazione, β, sono, in generale, funzioni del
tempo, poiché le disponibilità alimentari possono variare, soprattutto in natura: l'approssimazione qui
adottata è quella di considerarne il valore medio e porlo costante nel tempo.
Sotto queste ipotesi, il termine anabolico viene quindi espresso come:
ΦdRdt
⎛ ⎝ ⎜
⎞ ⎠ ⎟ = β ×
dRdt
= h(T) × f × β × wn (8.18)
Il catabolismo alimentare é rappresentato dall’insieme dei consumi energetici che l'organismo
deve sostenere per alimentarsi e rendere disponibili composti organici utili ad ogni sua cellula. Questa
parte del metabolismo comprende l'energia necessaria per il lavoro meccanico di ricerca, di cattura e
d’ingestione del cibo; include anche le spese che occorrono nei processi di usi delle molecole
organiche per ottenere energia e composti utili per la sintesi dei tessuti.
I processi descritti, legati all'alimentazione, avvengono con aumento di entropia, che viene espulsa
sotto forma di calore prodotto dall'organismo (Heat lncrement, HI) e mediante l'escrezione di
sostanze di scarto a basso contenuto di entalpia libera, derivanti dalla lisi e la ricostruzione delle
molecole, le urine: ne deriva che il catabolismo alimentare può essere misurato attraverso
quantificazione delle perdite energetiche in calore e urine.
É ragionevole supporre che il catabolismo alimentare sia una frazione della quantità d’energia
assimilata con l'alimento.
Il metabolismo basale costituisce l'insieme dei processi fisiologici con spesa energetica necessari
a mantenere in vita l'animale, esclusa l'alimentazione, e viene perciò chiamato anche "catabolismo a
digiuno" (fasting catabolism).
É rappresentabile con la perdita in peso del pesce dovuta alle spese energetiche, ricavate con la
respirazione, che l'organismo deve sostenere per la circolazione e la respirazione stessa. Il
metabolismo basale, proprio per la sua indipendenza dall'alimentazione, è espresso come funzione di
peso e temperatura.
Il tasso di respirazione viene quindi assunto proporzionale alla taglia dell'individuo espressa come
potenza del peso (w), e ad un fattore che dipende dalla temperatura.
La dipendenza funzionale dalla temperatura, descritta attraverso il parametro k(T), verrà discussa

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.19
approfonditamente nel paragrafo successivo. Sotto queste ipotesi, si ottiene l'equazione:
dO2
dt= −ω × k(T) × wn
(8.19)
dove ω è la costante di proporzionalità tra ossigeno consumato e peso perso dall'animale a dieta,
ω =dO2
dwe quindi:
−ϕ(w) = −k(T ) × wn (8.20)
L'esponente n è posto costante grazie all'ipotesi di crescita isometrica: ha valori positivi e nelle
diverse specie ittiche assume valori prossimi ad 1.
Introducendo queste ipotesi, la formulazione generale del bilancio metabolico dell'organismo
diventa:
dwdt
= (1− α ) × h(T) × f × β × wm − k(T ) × wn (8.21)
Questa equazione mette in evidenza che, nelle ipotesi fatte, il tasso di accrescimento dipende dalla
taglia dell'organismo e dalla temperatura, che influenzano i due termini dell’ anabolismo netto
(differenza tra anabolismo e catabolismo alimentare) e del metabolismo basale.
Il modello proposto, rappresenta quindi nella sua struttura funzionale le principali funzioni
fisiologiche del pesce: questo risulta anche dal confronto tra la Fig. 8.1, che descrive la ripartizione
dell'energia ingerita attraverso il cibo, e la Fig. 8.2, che rappresenta graficamente gli stessi termini
descritti con la formulazione proposta.
Le funzioni della temperatura h(T) e k(T), sono estremamente importanti per poter descrivere
realisticamente l'accrescimento di specie ittiche alle nostre latitudini, dove l'ampia escursione della
temperatura dell'acqua modula l'accrescimento su base stagionale.
Supponendo invece che la temperatura vari di poco, come accade nei mari tropicali, potremo ri
scrivere l’equazione (8.21), raggruppando i termini costanti:
dwdt
= H wm − k wn (8.22)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.20
8.3.0.0.- Processi chemo-bio-cinetici: I bilanci di materia 8.3.1.0.- Generalita’ La concentrazione che un tossico viene ad assumere in un organismo animale o vegetale
dipende da due processi antagonisti: il processo d’assunzione (Uptake) e quello di rilascio o di
depurazione. I due processi per ogni stadio o nella loro globalita' sono esprimibili con due costanti che
indicano la velocita' di accumulo e/o di clearance del tossico da quello specifico stadio biologico o dal
corpo completo dell'animale.
La prima costante (Ka) é chiamata la costante della velocita' d’assunzione (Uptake Rate Costant) ed é espressa in ml g-1 h-1o, assumendo la densita' del tessuto in un grammo per millilitro, in
h-1).
La seconda costante, indicata con Ke e la costante del primo ordine di eliminazione (depurazione)
dell'animale o di clearance (anch'essa in h-1). Una volta che il composto chimico è entrato attraverso la membrana cellulare nell’organismo,
potremmo individuare, in quest’ultimo, quattro siti importanti di reazione: i siti dell’azione tossica, i siti
del metabolismo, i siti di stoccaggio ed i siti di escrezione.
A) I siti dell’azione tossica. In tali siti la forma tossica della sostanza inquinante interagisce con
una macromolecula endogena (es.: una proteina o DNA) od una struttura biologica (es.: la membrana
cellulare) portando a manifestazioni tossiche nell’intero organismo.
B) I siti del metabolismo. Sono rappresentati da enzimi che metabolizzano i composti xenobiotici.
Di solito il processo metabolico porta alla detossificazione del composto. Attenzione, pero’, poiché in
alcuni casi il processo metabolico porta all’attivazione (aumento della tossicita’) del composto.
C) I siti di stoccaggio. In questi siti il composto xenobiotico viene inertizzato sotto forma di una
struttura inerte dal punto di vista della attivita’ tossicologia (granuli di metallo, per esempio).
D) I siti d’escrezione. L’escrezione può essere la stessa sostanza inquinante originale inalterata.
Prevalentemente, pero’, cio’ che viene escreto é il prodotto di biotrasformazione (metabolita o
composto coniugato).
Il modello piu’semplice di tossicinetica é rappresentabile da un modello monocompartimentale ove
si assume che il composto xenobiotico entri, si accumuli, e si metabolizzi in un solo “reattore”. In realtà
i compartimenti di un organismo sono molti e l’ubicazione del tossico puo’ essere molto varia.
Così, uno xenobiotico può essere immagazzinato sia in depositi grassi che in membrane inerti.
Anche una struttura bersaglio per un tossico o per un farmaco puo’ essere multipla e localizzata in
parti diverse dell’organismo.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.21
Ad esempio una potente neurotossina come la colinesterasi può incontrare siti bersaglio sia nel
sistema nervoso centrale che in quello periferico.
Dopo il processo di uptake la sostanza inquinante é trasportata ai diversi compartimenti del corpo
da sangue e linfa (vertebrati) o dall’emolinfa (insetti). Il passaggio negli organi e nei tessuti si realizza
per diffusione attraverso barriere di membrana o, per composti estremamente lipofili attraverso co-
trasporto con lipidi.
Molecole prive di carica, con pesi molecolari inferiori a 800 (impedimento sterico!) e con un discreto
coefficiente di ripartizione ottanolo/acqua, si muovono attraverso le membrane per diffusione passiva.
Anche le lipoproteine agiscono come carrier “dissolvendo” composti particolarmente lipofili.
I composti non metabolizzati vengono immagazzinati in strutture di deposito dei grassi od altre
strutture lipofile come le membrane o le lipoproteine.
Tali depositi di composti lipofili potenziamente tossici possono essere protettivi a breve termine. A
lungo termine, comunque, puo’ succedere che il processo si inverta e vi sia un rilascio del composto dalla struttura di accumulo. Questo processo é sempre legato al potenziale chimico
assunto dal composto nella struttura di storage (stoccaggio).
Quando il potenziale chimico o la fugacita’ eccedono le “resistenze” della struttura, si ha il rilascio
del tossico stoccato. Questo può condurre a improvvisi effetti tossici nella cellula interessata. Questo
fenomeno, chiamato anche tossicita’ ritardata (delayed toxicity) é stato osservato nel caso di alcuni
insetticidi clorurati come la dieldrina.
La bassa fugacita’ dei composti xenobiotici nelle strutture lipidiche espressa da alti valori di Kow,
rispetto alla fase acquosa porta al fatto che il composto non viene espulso direttamente nelle feci o
nelle urine degli organismi.
Riportiamo, nella figura che segue, i quattro processi di trasporto prevalenti attraverso la
membrana citoplasmatica dei metalli:
1. Processi di trasporto carrier-mediated. (Fig.8.3 - prima figura dall'alto).Sono quelli, mediati da
un trasportatore, in cui alcune proteine (L) formano, con il metallo, un complesso solubile nei lipidi,
ML; il complesso diffonde entro la membrana, ed il metallo può essere rilasciato nel citosol (il
contenuto acquoso della cellula vivente). La maggior parte dei metalli entra nella cellula attraverso
questo percorso.
2. Processi di trasporto attraverso i canali proteici. (Fig.8.3 - seconda figura dall'alto) Ioni del
metallo possono essere trasportati fra le proteine che si estende attraverso la membrana e che
presentano molti gruppi idrofilici.
3. Processi di trasporto per diffusione passiva. (Fig.8.3 - terza figura dall'alto).Specie del metallo
che sono solubili nei lipidi (non polari) si possono dissolvere nella membrana e rapidamente
attraversarla. Questo processo è invocato spesso per spiegare l’uptake rapido di metalli-alchili da

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.22
organismi unicellulari.
4. Processi di trasporto per endocitosi. (Fig.8.3 - quarta figura dall'alto). La membrana del plasma
è fluida. Una regione della membrana può invaginare ed avvolgere una particella contenente il metallo
e fondersi con essa per formare una vescicola intracellulare.
Fig.8.3
8.3.2.0.- Il fattore di bioconcentrazione e di bioaccumulo 8.3.2.1.- Il fattore di bioconcentrazione (BCF) Il parametro che da' un’indicazione importante dell'andamento del processo di assunzione é il
BCF o fattore di bioconcentrazione. Il BCF é il rapporto tra la concentrazione media dello xenobiotico (o di un qualsiasi composti
chimico ripartibile) nel tessuto dell'organismo esaminato in condizioni di stato stazionario e la
corrispondente concentrazione dello stesso tossico nell'acqua alla quale viene esposto l'organismo.
BCF =Corganismo
Cacqua
=CCw
(8.23)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.23
Il BCF é di norma correlato con il contenuto in grasso dell'organismo, e quindi varia entro e fra le
specie acquatiche in funzione del contenuto in grassi. Nel pesce, ad esempio il grasso puo' variare da
2 a 15 %.
Se un composto ha un valore di BCF superiore a 1000 allora é necessario proseguire nelle
ricerche in merito al rischio tossicologico che il prodotto puo' porre. Composti come il DDT,PCB e
dieldrina hanno valori di BCF di gran lunga superiori a 1000. I valori di BCF possono ancora variare
considerevolmente (da uno a due ordini di grandezza) fra le specie ed il periodo di vita, probabilmente
per la diversa velocita' delle reazioni di disintossicazione che sono, normalmente, minori a livello dei
primi stadi di vita.
Quindi un valore di BCF non deve essere utilizzato in assoluto quando il sistema biologico varia
nel tempo come caratteristiche dimensionali dell'individuo e come eta'. Cio’ non ostante si puo' dire
con buona certezza che un composto che abbia un BCF di 100 o meno non presenta problemi di
concentrazione ambientale e, quindi, di tossicologia ambientale.
Contrariamente a cio' che succede per gli animali acquatici per gli animali terrestri il contatto piu'
importante é quello della dieta. I valori di BCF nel tessuto grasso, valutati per 23 prodotti chimici di
interesse ambientale, si sono riscontrati piu' piccoli con fattori da tre a quattro ordini di grandezza
rispetto al pesce anche se tra le specie si riscontra una buona correlazione cio' che consente di
ottenere dati per una specie acquatica attraverso una regressione dei dati per una specie terrestre.
Un modo per predire la distribuzione dei composti chimici nell'ambiente si puo' ottenere
combinando le costanti Kow,Koc e la solubilita' del prodotto in acqua come riportato nelle relazioni che
seguono.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.24
BOX 8.2 Correlazione Eq.Regressione Coeff.Corr. r ______________________________________________________________ S/BCF log S= 2,531-0,916 log BCF -0,72 Koc/BCF log Koc = 1,963+0,681 log BCF +0,87
BCF/S log BCF= 2,791-0,564 log S -0,72 BCF/Koc log BCF = -1,579+1,119 log Koc +0,87
Solub/Koc log S = 5,09-1,28 log Koc -0,84
Koc/S log Koc= 3,64-0,55 log S -0,84
Come si puo' notare le prime quattro relazione includono direttamente il BCF mentre le altre due
consentono di calcolare parametri d'uso nelle prime quattro da altri fattori. Sul modo di calcolo del
BCF torneremo piu' avanti attraverso l'esame dei modelli d’assunzione di tossici in modo
monocompartimentale e bicompartimentale.
8.3.2.2.0.- Modelli compartimentali
Gli obbiettivi principali dello studio chemio-bio-cinetico sono quelli di determinare la quantita’, la
velocita’ e la natura dei processi di assorbimento, distribuzione, metabolismo, ed escrezione di un
composto chimico in una struttura animale.
Una premessa necessaria é che, poiché il fine delle ricerche della ecotossicologia e della che
chemio-bio-cinetica é quello di fare valutazioni previsionali del comportamento del composto chimico
nei confronti dell’essere umano, il soggetto animale da usarsi per le valutazioni chemo-bio-cinetiche
andra’ cercato in un animale dal comportamento uguale o molto simile all’uomo.
Ma é difficile individuare, senza dati precedente, una specie animale che metabolizzi un
composto xenobiotico in modo simile all’uomo.
Di solito gli studi iniziali si fanno nel ratto o nel cane o nella scimmia, nel tentativo di
determinare la variabilità della specie. Se ci sono differenze significative fra specie, è importante
determinare se le differenze nei parametri dei processi chemo-bio-cinetici si correlino con le differenze
nella tossicita’ o nell’attività farmacologica.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.25
Come abbiamo gia’ detto, la chemo-bio-cinetica mira a quantificare i processi gia’ discussi
precedentemente. Così, la chemo-bio-cinetica fornisce informazioni quantitativi sull'assorbimento,
distribuzione, biotransformatione, escrezione di composti chimici xenobiotici, (medicine e sostanze
endogene incluse), in funzione del tempo.
Una difficolta’ di molti tossicologhi e biologhi relativamente alla chemo-bio-cinetica è il concetto
di compartimenti. Un corpo è composto da un gran numero di organi, tessuti, cellule e fluidi, ed ad ognuno di essi
può essere definito, morfologicamente e funzionalmente, come un compartimento.
Comunque, in chemo-bio-cinetica, un compartimento si riferisce al complesso di quegli organi,
tessuti, celle, e fluidi per quale le velocita’ di uptake e di rimozione susseguente di un composto
chimico sono sufficientemente simili da escludere una differenziazione cinetica.
Il compartimento che si equilibria rapidamente viene considerato come il compartimento centrale, e può comprendere tutti quei tessuti dove il sangue é presente in rapida perfusione10 mentre
il i cosiddetti compartimenti lenti o periferici includono tessuti con una piu’ limitata perfusione di
sangue (ad esempio: il grasso e le ossa).
8.3.2.2.1.- Modello monocompartimentale aperto
L'assunzione e il rilascio di un composto chimico dal tessuto animale possono essere considerati
come parte di un processo omogeneo inteso come verificantesi in un organismo che si comporta
come un singolo compartimento, nel quale l'assunzione del tossico si verifica solo attraverso l'acqua
ed é direttamente proporzionale alla concentrazione nell'acqua del tossico stesso.
Inoltre tutte le molecole del tossico sono egualmente disponibili ad essere eliminate.
Ancora in questo caso, si assume che il processo di clearance o eliminazione sia una cinetica del
primo ordine e che, quindi, l'eliminazione del tossico sia direttamente proporzionale alla
concentrazione del tossico stesso nell'animale.
Infine va assunto che la dimensione ed il volume del compartimento non vari durante il periodo in
cui si effettua lo studio senza che vi siano, quindi, fenomeni di accrescimento nell'animale.
10 Per gli animali dotati di circolazione sanguigna.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.26
Il modello chemo-bio-cinetico più semplice è quello del modello aperto mono-compartimentale.
Fig. 8.4 Schema di modello mono-comportamentale con ka e ke le costanti di assunzione e
di clearance, rispettivamente.
Usando questo modello, si presume che il composto chimico xenobiotico si equilibri con tutti tessuti nei quali è distribuito in modo sufficientemente rapido da impedire differenziazioni cinetiche.
Ad esempio se sono richiesti 30 minuti per raggiungere l’equilibrio nel corpo dopo l’entrata dello
xenobiotico nello stream sanguigno, analisi di sangue, tessuti ed excreti fatte a 30 minuti di intervallo
mostreranno che non vi é differenza di concentrazioni tra le varie fasi e, quindi, che tutto l’assieme
(corpo) rappresenta un solo compartimento.
In sostanza il modello prevede un entrata di un composto xenobiotico nel substrato biologico
che, fino ad una certa concentrazione Ct, non mette in atto meccanismi di difesa in quanto la cellula o
la struttura biologica, in generale, non riconosce in tale concentrazione un rischio per la sua propria
sopravvivenza.
Oltre ad un certo valore soglia (threshold) si innescano meccanismi di allarme di tipo

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.27
biochimico che portano a processi di difesa. Essi possono essere processi di inglobamento e/o precipitazione dl tossico sotto forma di
strutture chimiche inerti (es.: precipitazione dei granuli di metallo elementare in siti para-cellulari,
formazione di metallotioenine ecc.) o veri e propri processi metabolici di aggressione dei siti piu’
reattivi della molecola dello xenobiotico.
In tal caso, di solito, avviene una prima azione di ossidazione ed idrolisi che porta ad una
struttura chimica piu’ facilmente metabolizzabile ed eliminabile come composto in grado, poi, di
essere escreta ed eliminata dalla struttura bersaglio.
Mentre, quindi, il processo di assunzione inizia immediatamente una volta che il substrato
biologico viene ad essere messo a contatto con lo xenobiotico infase liquida e/o gassosa, quello di
clearance o release o di eliminazione entra in funzione solo e soltanto dopo un certo tempo che é
funzione del tipo di substrato biologico, del tipo di composto chimico e della pre-esistenza o meno, nel
substrato biologico stesso, di una certa quantita’ del composto.
Infatti, come é ben noto, il flusso di composto, ad esempio, attraverso una membrana cellulare é
determinato (cfr.Tool per l’Ecotossicologia Cap.IIA) dal gradiente di concentrazione interno/esterno
della membrana cellulare che esprime, anche, la fugacita’od il potenziale chimico del composto nella
fase esterna e nella fase interna della cellula, rispettivamente.
Al tempo t = 0, il composto xenobiotico si trova solo all’esterno della struttura e, quindi, in
funzione della legge di Fick, la sua velocita’ di passaggio (e quindi il flusso) é massimo. Man mano che
la differenza di concentrazione interno/esterno tende a zero, in funzione delle forze di resistenza (Di) il
flusso si riduce fino a portarsi anch’esso a zero. Ma oltre ad una certa concentrazione inizia il processo
di eliminazione per cui, se la sorgente di contaminazione permane, si avra’ un processo costante di
assunzione e di clearance (stato stazionario o di equilibrio dinamico con ∆G=0)
Se ogni organismo si comporta come un solo compartimento la cinetica di bioconcentrazione di
un tossico nel tessuto di un animale (nel caso il pesce) é la combinazione tra i processi di assunzione,
regolati dalla costante ka, e quelli di clearance regolati dalla ke. La relazione generale della cinetica é
quindi:
CekwCakperditeassunzionedtdC −=−=
(8.24)
con

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.28
Cw = la concentrazione del composto chimico in acqua (mg l-1)
C = la concentrazione del composto chimico nell'animale
t = il tempo (in ore)
ka = la costante di assunzione (in ml g-1 h-1)
ke = la costante di clearance (in h-1)
In condizioni di stato stazionario le due velocita', di assunzione e di eliminazione, si equivalgono e,
pertanto:
ssCekwCakdtdC −== 0
(8.25)
dove con Css si indica la concentrazione del tossico nel tessuto del pesce allo stato stazionario.
risolvendo per Css si ha:
ekwCak
ssCwCakssCek ==
(8.26)
Il valore di BCF in condizioni di stato stazionario puo' essere dato da:
ekak
wCssCBCF ==
(8.27)
Riferendoci alla quantita' di tossico assunto per unita' di tempo, dalla relazione principale e
integrata si ottiene:
⎥⎦⎤
⎢⎣⎡
⎟⎠⎞⎜
⎝⎛−−= tekwC
ekakC exp1
(8.28)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.29
ovvero
⎥⎦⎤
⎢⎣⎡
⎟⎠⎞⎜
⎝⎛−= tekssCC exp1
8.29)
Come si puo' notare per la definizione di fattori di bioconcentrazione é indispensabile conoscere i
valori di ka e ke. Cio' puo' essere fatto sperimentalmente ovvero calcolato dai dati fisiologici
dell'animale da esperimento e dalla caratteristiche chimiche e chimico-fisiche del composto. In questo
secondo caso, due sono gli approcci fondamentali: quello monocompartimentale e quello a due
compartimenti.
Nel primo caso i tessuti dell'animale sono considerati come un unico pool in cui il tossico si
distribuisce in maniera omogenea; nel secondo caso, invece, i tessuti si comportano almeno con due
velocita' di assorbimento diverse.
Come si puo’ intuire, ai fini tossicologici ed ecotossicologici, il processo piu’ importante é,
certamente quello di clearance e/o di eliminazione. É, quindi, opportuno esaminare con maggior
dettaglio questo processo iniziando dal sistema aperto mono-compartimentale.
Assumendo che la velocita’ d’eliminazione del composto chimico sia proporzionale alla sua
concentrazione nel plasma, detta concentrazione puo’ essere descritta da una cinetica apparente del primo ordine ed espressa nella forma di una equazione lineare differenziale:
tet Ck
dtdC
−= (8.30)
dove C(t) è la concentrazione a tempo t, e ke, è la costante cinetica eliminazione. Soluzione di
questa equazione differenziale con le condizioni iniziali C(t) = C(0) al tempo zero danno le seguenti
forme esponenziale e logaritmiche:
( )tkCC et −= exp0 (8.31)
e logarimica
0lnln CtkC et +−= (8.32)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.30
0log303,2
log CtkC et +
−= (8.33)
In queste equazioni C(0) è la concentrazione del composto chimico nel plasma al tempo zero.
Plottando C(t) contro il tempo su carta semilogaritmica si otterra’ una linea diritta con slope - ke ed
intercetta C(0).
Calcolato ke, che é espresso in unita’ di tempo reciproco, il tempo richiesto per ridurre la concentrazione dello xenobiotico nel compartimento alla metà puo’ essere calcolato; questo
tempo è definito come t 1/ 2 o tempo di semi-vita e puo’ essere calcolato dall’equazione:
ekekt 693,02ln
2/1 == (8.34)
Quando il composto chimico non è assorbito instantaneamente le espressioni matematiche
necessarie per descrivere la concentrazione nel plasma in funzione del tempo, divengono più
complicate. Assumendo che le cinetiche d’assorbimento e di eliminazione siano ambedue del primo
ordine, la concentrazione C(t) nel plasma è data dall'espressione:
( ) ( ) ( )tkexptkexpkkV
kf.DC(t) aeead
a0 −−−−
= (8.35)
In questa espressione i termini che non sono stati finora menzionati sono D0 = la dose; f = la
frazione di dose assorbita; Vd, il volume apparente di distribuzione; e k1 = la costante cinetica
apparente di primo l'apparente ordine di assorbimento (uptake).
La costante d’eliminazione (clearance), k, viene determinata usando quello porzione della linea
solida che rappresenta la concentrazione del plasma dopo che l’assorbimento è completo. In Fig.8.5
cio’ si verifica quando la linea tratteggiata si fonde nella linea solida.
La costante della cinetica d’assorbimento, k1 può essere valutata proiettando la linea solida
all’indietro fino all'origine. Lo stesso diagramma può essere usato per il calcolo del t 1/ 2 per il processo
di assorbimento e di ka.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.31
Il volume di distribuzione, Vd, è un termine che descrive il volume apparente nel quale un
composto chimico è distribuito quando si possa ritenere che l’affinità del plasma e di tutti i tessuti per
quello specifico composto sia la stessa.
Un'analogia potrebbe essere quella di porre una nota quantita’ di colorante in un liquido
contenuto in un volume sconosciuto. All’equilibrio (e cioé quando il colorante si é distribuito
omogeneamente in tutta il volume), il volume del sistema (Vd) puo’ essere ottenuto dividendo la dose
(D0) per la concentrazione unitaria.
Nel plasma la concentrazione dei composti chimici si riduce a causa del processo di
eliminazione (clearance) e della distribuzione nei tessuti. Perciò per valutare Vd è necessario
proiettare il tratto di curva corrispondente al processo di eliminazione fino all’origine. Il valore ottenuto
dall’intercetta al tempo zero da questa proiezione, diviso da D0 da’ il volume di distribuzione, Vd, in ml
kg-1.
Il valore di Vd fornisce alcuni informazioni importanti sulla distribuzione del composto xenobiotico
nel corpo. Con l’aumento della distribuzione dello xenobiotico nei tessuti, per qualunque motivo,
(affinità chimico-fisica, attivo trasporto attivo nelle cellule ecc.), anche Vd aumenta.
Se la distribuzione di un chimico nel corpo umano si limita al plasma, al fluido extracellulare od
al liquido corporeo totale, i valori rispettivi di Vd sono circa 40, 170, e 580 ml kg-1. Se uno xenobiotico
ha un'alta affinità per un tessuto particolare, per esempio, l'affinità di un lipofilo per il grasso, Vd può
eccedere in modo significativo i 1000 ml kg-1.
Fino a ora si sono presi in considerazione solo i concetti relativi alla concentrazione dello
xenobiotico nel plasma. Ma questi concetti sono ugualmente applicabili ad altri tessuti come pure agli
escreti, all’aria ed ai gas emessi dal corpo, oppure ai liquidi e/o solidi come l’orina e le feci.
In aggiunta alla concentrazione, gli stessi concetti possono essere applicati se si desidera
calcolare la quantita’ totale di uno xenobiotico nel corpo, A(t), in funzione del tempo di esposizione.
Per esempio se una dose D0 è ingerita e si assume che il processo sia una cinetica apparente del
primo ordine, la quantita’ dello xenobiotico nel corpo é espresso da:
A(t) = D0 exp (-ket) (8.36)
che é equivalente a: C(t) = C(0) exp (-ket)
Una certa quantità (dose singola) di un composto xenobiotico entra in un compartimento
attraverso il fluido che bagna il compartimento stesso (esempio: il plasma). Ad un certo punto inizia il
processo d’eliminazione (clearance) controllato dalla costante di eliminazione Ke. A seguito del processo
di clearance, la concentrazione nel plasma diminuisce secondo un retta il cui esempio è riportato nella

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.32
figura seguente:
Fig.8.5 (a)
Il valore della concentrazione iniziale viene valutato per estrapolazione dei dati di analisi del
plasma che si ottengono analizzando a tempi successivi il plasma stesso. La scala delle concentrazioni
viene posta logaritmica e l’inclinazione della retta fornisce il valore di –ke. Data la seguente tabella di
valori sperimentali della concentrazione dello xenobiotico, calcolare il valore della concentrazione iniziale
sia graficamente che numericamente sapendo che:
tk0
eeCC −=
e, quindi, che:
tklnClnC e0 −=−
e
2,303tklogClogC e
0 −=−

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.33
Calcolare, infine, il tempo di dimezzamento dello xenobiotico nel plasma, ricordando che:
e1/2 k
ln2t =
Il processo di bioaccumulo in un “compartimento biologico” è descritto dall’espressione:
CkCkdtdC
eaa −=
ove C è la concentrazione internamente al compartimento e Ca quella nel sito di assorbimento esterno al
compartimento.
La curva sottostante riproduce l’andamento del processo di bioconcentrazione in un sistema
monocompartimentale per l’aggiunta di uno xenobiotico in una sola singola dose.. La prima parte della
curva espressa dai valori di concentrazione (pallini neri) rappresenta il progressivo accumulo dello
xenobiotico; come si nota la curva inizia con una rapida pendenza (fase logaritmica) cui segue un flesso
nel momento in cui entrano in funzione i sistemi di difesa da parte della cellula (compartimento) che viene
avvisata dai suoi sensori (di energia?) che un composto anomalo rispetto alla sua struttura biochimica, si
sta accumulando nel compartimento con potenziale azione ostile alla stessa sopravvivenza della cellula.
Inizia, quindi, il processo di detossificazione.
Tale processo, chiamato di clearance o di eliminazione, è in realtà un processo (o più processi)
metabolico che annulla, attraverso modifiche della struttura molecolare , la capacità tossica dello
xenobiotico. Il metabolismo può inserire nella molecola dello xenobiotico gruppi –OH, rendendo il
composto stesso più solubile ed eliminabile nei liquidi organici; può complessate lo xenobiotico con
formazione di composti stabili e non tossici (es.: le Metallotioneine per i metalli tossici), può bloccarlo
sotto forma di composto inerte in situ (ossia internamente al compartimento stesso (come è il caso dei
granuli di metalli pesanti).
In ogni caso il processo di clearance si oppone a quello d’accumulo ed é controllato da una
costante d’eliminazione ke. Dalla relazione sovra scritta, quindi, si capisce come il processo di
bioaccumulo sia direttamente proporzionale alla concentrazione dello xenobiotico nel mezzo in cui si
trova il compartimento e proporzionale in senso negativo alla concentrazione che lo xenobiotico ha
raggiunto nel compartimento.
G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.34
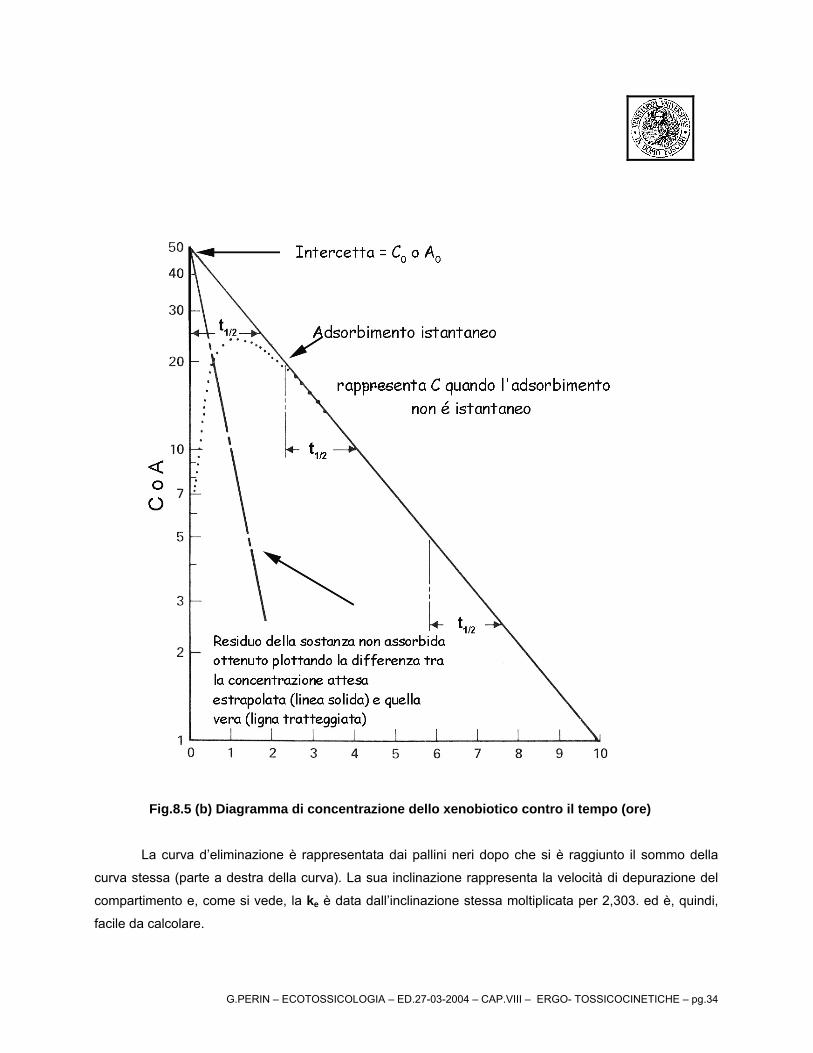
Fig.8.5 (b) Diagramma di concentrazione dello xenobiotico contro il tempo (ore)
La curva d’eliminazione è rappresentata dai pallini neri dopo che si è raggiunto il sommo della
curva stessa (parte a destra della curva). La sua inclinazione rappresenta la velocità di depurazione del
compartimento e, come si vede, la ke è data dall’inclinazione stessa moltiplicata per 2,303. ed è, quindi,
facile da calcolare.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.35
Infatti:
dalle:
(a)eCC tk0
e−=
tklnClnC e0 −=−
2,303tklogClogC e
0 −=−
si ricava che
(b)2,303
kttlogClogC e
12
12 −=−−
Ricavata la ke è importante calcolare la ka (costante di uptake o di assorbimento). Quest’ultima si
calcola con il cosiddetto metodo dei residui. La formula da applicare è analoga a quella riportata in (b) ma
inserendo i valori della nuova retta (pallini chiari – (c)), costruita con i valori delle differenze tra le
concentrazioni nel compartimento riportate sulla linea estrapolata [valori fino a circa 18 ore nel
diagramma riportato come esempio - Fig.8.5(c) – retta tratto x ] ed i valori delle concentrazioni nel
compartimento nella curva a sinistra – tratto y.
La formula, quindi, diventa:
)(c2,303
kttlogClogC a
12
12 −=−−
Nel diagramma d’esempio si può notare come la cinetica d’assorbimento (uptake) è maggiore di
quella della clearance come mostrano le diverse inclinazioni delle due rette. Questo fenomeno è generale
e solo in pochi casi il processo di clearance è più rapido di quello di uptake (alcuni composto polari).

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.36
Fig.8.5 (c)
La concentrazione massima nel compartimento (Cmax) ed il tempo che serve per raggiungere la
concentrazione massima possono essere calcolati dalle:
1ea
e
amax )k(k
kklnt −−=

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.37
)e(e)k(kV
DFkkk
VDFC maxamaxe
1eae
tktk
eaD
a
)k(kk
a
e
Dmax
−−
−
−−
=⎟⎟⎠
⎞⎜⎜⎝
⎛=
−
ove D è la dose istantanea, F è la frazione dello xenobiotico che entra nel compartimento non modificata
(e quindi dà una indicazione sulla bio-disponibilità del composto: per F=0, nessuna bio-disponibilità; per F=1, totale disponibilità) e VD è il volume apparente di distribuzione,.
8.3.2.2.2.- Modello bicompartimentale/multicompartimentale
Le considerazioni fatte per il modello monocompartimentale non sono sempre verificate nella
realta'. Anzi, nella maggior parte dei casi molti residui di tossico nell'animale non si comportano come
fossero parte di un tutt'uno omogeneo.
Certi tessuti, infatti, rilasceranno con una velocita' maggiore il tossico di altri per cui, in realta',
bisognera' tener conto che, una volta determinata la concentrazione di tossico rilasciato al tempo t, vi
saranno altre parti dell'animale che continueranno il processo di clearance. Indipendentemente dalle
pathways del processo é chiaro che dovremo, quindi, pensare di aver almeno come due diversi
compartimenti uno in cui i processi di clearance sono veloci ed uno in cui sono, relativamente, piu'
lenti.
Anche questa é peraltro una soluzione semplificante ma, almeno si avvicina di piu' alla
realta' di situazioni che certamente si verificano specialmente negli animali a complessita' strutturale
elevata. Un modo generico di esprimere il modello bicompartimentale é dato dalla relazione:
)exp()exp()( tttC βψαφ −+−= (8.37)
ove ψφ + rappresentano la concentrazione totale del composto chimico nel corpo dell'animale
all'inizio del processo di clearance. Come si puo' notare il processo di clearance é determinato dalla
somma dei due processi, uno lento e l'altro veloce espressi dalle due equazioni esponenziali in αt e
βt.
Analizzando in maggior dettaglio il modello bicompartimentale (o (multicompartimentale),
riscontreremo che compartimenti nei quali il composto chimico xenobiotico ha raggiunto equilibrio con
plasma prima che vengano prelevati i campioni di sangue apparirà cineticamente come un
compartimento unitario, mentre quello in cui l’equilibrio si raggiunge piu’ lentamente, (compartimento profondo), darà origine a una curva di concentrazione nel plasma che si presenta come bifasica. Il

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.38
modello usato per descrivere questo sistema è, piu’ propriamente, quello del modello aperto bicompartimentale. Nel disegno che segue si possono notare il modello bicompartimentale e quello
tri-compartimentale.
Fig.8.6
Nel bi-compartimentale, il sub-compartimento centrale e quelli rapidamente compartimenti
equilibratisi sono da considersi unitari.
Poiché i siti ove si realizza la maggiore quantita’di trasformazioni metaboliche e di escrezione
sono il fegato ed i reni.
Poiché questi organi sono per fusi col sangue, si puo’ ritenere che siano parte del
compartimento centrale e che il processo di eliminazione (clearance) si verifica nel compartimento
centrale.
In fig.8.8 viene simulata la curva di concentrazione nel plasma in un sistema a due
compartimenti dopo una rapida introduzione endovenosa di uno xenobiotico.
Il composto chimico viene come prima cosa rapidamente distribuito nei tessuti ben irrorati dal
sangue e poi, più lentamente, agli altri tessuti che formano il compartimento profondo.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.39
Fig.8.7 Interpretazione di un sistema tricompartimentale e dei valori delle costanti k1 e k2
Se, come precentemente fatto, assumiamo che tutte le cinetiche di trasferimento dello
xenobiotico siano del primo ordine, un sistema di lineare di equazioni differenziali descrive il modello
bi-compartimentale e cioé:
d
DDe V
tCVktCktCkdt
tdC )()()()( 2112 +−−= (8.38)
e
)()()(21
12 tCkV
tCVkdt
tdCD
ddD −== (8.39)
dove C(t) e CD(t) sono, rispettivamente, le concentrazioni del composto chimico xenobiotico nei
compartimenti centrali e profondi. I volumi apparenti di distribuzione per questi compartimenti sono Vd
per il compartimento centrale e VD per il compartimento a lento scambio. Se il flusso volumetrico
apparente tra i due compartimenti fosse lo stesso, ossia se k12 Vd = k21 VD il sistema di equazioni
differenziali può essere risolto con le condizioni iniziali C(0) = D0/ Vd e CD(0) = 0 al tempo zero dando
G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.40
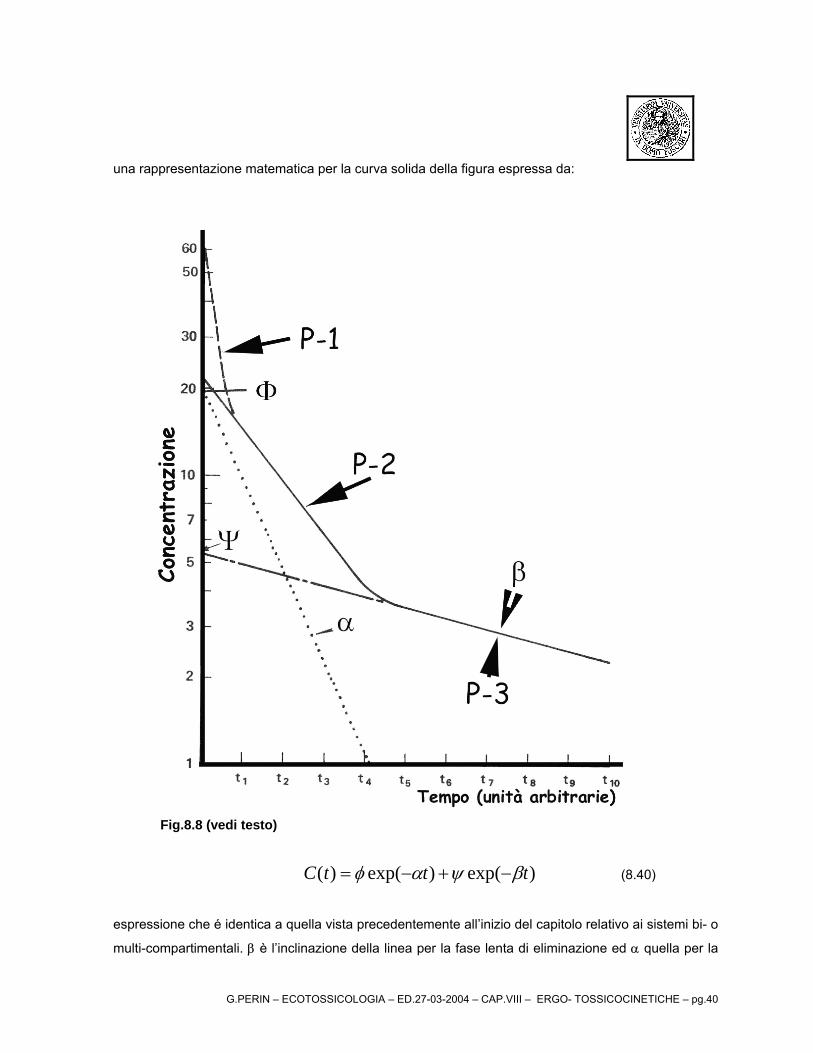
una rappresentazione matematica per la curva solida della figura espressa da:
Fig.8.8 (vedi testo)
)exp()exp()( tttC βψαφ −+−= (8.40)
espressione che é identica a quella vista precedentemente all’inizio del capitolo relativo ai sistemi bi- o
multi-compartimentali. β è l’inclinazione della linea per la fase lenta di eliminazione ed α quella per la

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.41
fase rapida di eliminazione.
Il valore di beta viene calcolato come prima descritto mentre, per il calcolo di α si ricorre ad una
tecnica chiamata “feathering”. Questa tecnica consiste nel proiettare la linea solida per la fase lenta
all’indietro fino all'origine (linea trattino-linea-trattino) e sottraendo i valori rispettivi proiettati dai valori
sperimentali usati per costruire la fase rapida di rimozione. Questi valori vengono riplottati (linea
punteggiata).
La slope di questa linea è α. I valori per φ e ψ sono l'intercetta all’ordinata delle rette
rappresentanti le fasi di eliminazione rapida e lenta, rispettivamente.
Le costanti cinetiche k12 k21 e ke possono essere determinate dalle seguenti relazioni:
ψφψαφβ
++
=21k (8.41)
21kke
αβ= (8.42)
( )ekkk +−+= 2112 βα (8.43)
k12 è di importanza particolare perchè da essa si puo’ facilmente calcolare la quantita’ del
composto chimico xenobiotico nel compartimento profondo (AD(t)) mediante la seguente equazione:
( ) ( )ttDktAD βααβ
−−−−
= expexp)( 012 (8.44)
La conoscenza della quantita’ di tossico presente nel compartimento profondo consente di
capire se vi sono relazioni tra l'effetto del tossico stesso, verificato sperimentalmente, e la sua
presenza in un compartimento profondo.
É inoltre, assai importante in ecotossicologia sapere se esiste una fase lenta di eliminazione del composto xenobiotico (rappresentata, come detto, dal (o dai) compartimento(i) profondo(i)) la cui
esistenza si pone come un indicatore di allarme che suggerisce che con ripetute somministrazioni
del tossico si puo’ andare incontro ad una fase di tossicita’ dovuta a bioaccumulo.
La concentrazione di uno xenobiotico nel plasma o nei tessuti o la sua quantita’ nel corpo totale
a seguito di ripetute esposizioni è illustrata nel seguente diagramma per un sistema aperto mono-
compartimentale.
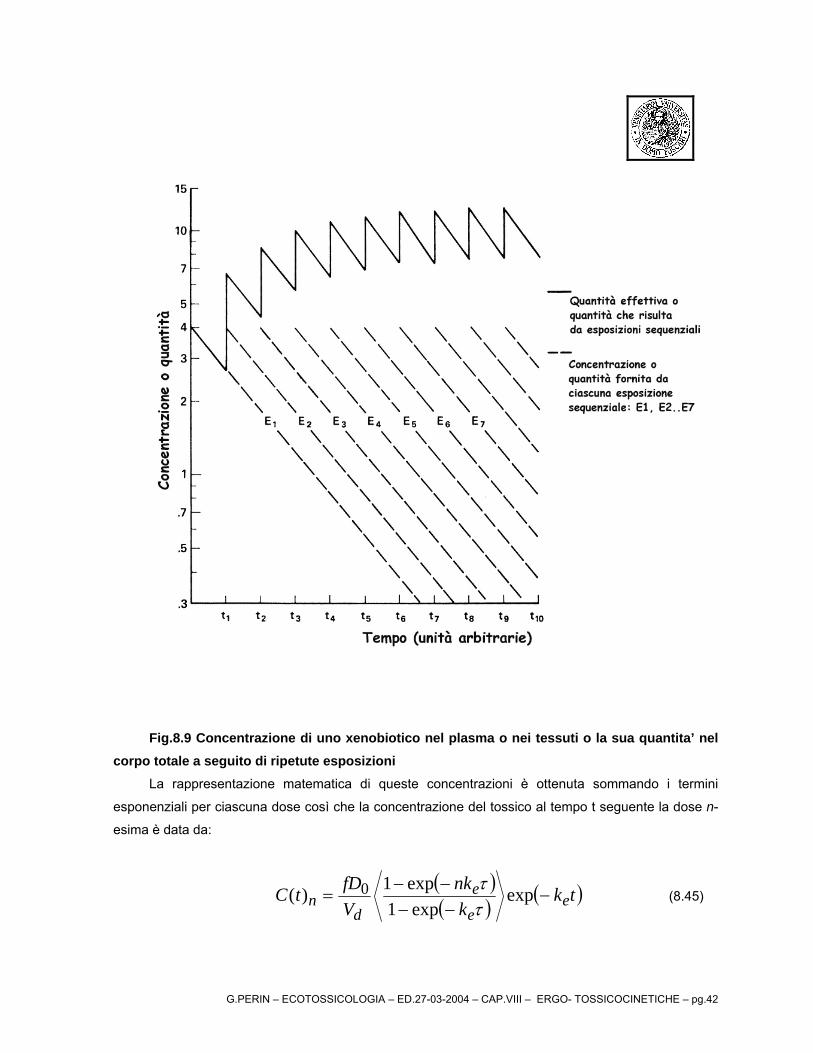
G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.42
Fig.8.9 Concentrazione di uno xenobiotico nel plasma o nei tessuti o la sua quantita’ nel corpo totale a seguito di ripetute esposizioni
La rappresentazione matematica di queste concentrazioni è ottenuta sommando i termini
esponenziali per ciascuna dose così che la concentrazione del tossico al tempo t seguente la dose n-
esima è data da:
( )( ) ( tk
knk
VfDtC e
e
e
dn −
−−−−
= expexp1
exp1)( 0ττ ) (8.45)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.43
dove τ è l'intervallo tra dosi, Dopo un gran numero di dosi, il termine exp(-nkeτ)
tende a zero per cui la concentrazione dello xenobiotico diviene:
( )( )τe
e
d ktk
VfDtC
−−−
=∞ exp1exp)( 0 (8.46)
Una volta che la concentrazione raggiunge il valore di plateau, ulteriori esposizioni alla stessa
dose ed alla stessa frequenza non porteranno a nuovi aumenti Al valore di plateau la concentrazione
massima che si verifica immediatamente dopo l’ultima esposizione sara’ data da:
( )τed kVfDC
−−=∞ exp1
1(max) 0 (8.47)
e la concentrazione minima si verifichera’ immediatamente prima della successiva esposizione e sara’
data da:
( )( )τ
τ
e
e
d kk
VfDC
−−−
=∞ exp1exp(min) 0 (8.48)
L'espressione che definisce la concentrazione media dopo che il plateau è stato raggiunto è:
τedkVfDavC 0)( =∞ (8.49)
Se l'esposizione è tale che deve essere considerata anche la cinetica di assorbimento (del primo
ordine con ka = costante cinetica), la concentrazione dello xenobiotico nel plasma dopo n dosi
somministrate ripetutamente con intervalli di tempo τ, la concentrazione é data dalla:
( )( )( ) ( ) ( )
( ) ( )−−⎟⎟⎠
⎞⎜⎜⎝
⎛−−
−−−−⎟⎟
⎠
⎞⎜⎜⎝
⎛−−
−−−
= τττ
ττ
aaa
eee
eada
n kknktk
knk
kkVkfDtC exp
exp1exp1exp
exp1exp1)( 0
(8.50)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.44
8.3.2.2.3 Cinetiche di sistemi non lineari o saturabili
Le curve dose-effetto che risultano dall’analisi delle reazioni di una struttura biologica a seguito
della somministrazione di un composto xenobiotico, mostrano di solito, un andamento log-normale.
Estrapolando si puo’ prevedere che che alcuni individui risponderanno a dosi infinitesime mentre altri
risponderanno mai, per quanto grande sia la dose.
L'assunzione che ne segue è che il profilo chemobiocinetico del composto sia indipendente dal
livello di dose somministrato.
Peraltro, molti processi metabolici ed escretori sono saturabili, e, quindi, la chemobiocinetica non lineare è dell'importanza massima in tossicologia.
Molti processi metabolici e di trasferimento attivo hanno una capacita’ limitata di reazione con un
composto xenobiotico. La cinetica di tali processi non-lineari è definita dalla ben nota equazione di
Michaelis-Menten.
)()()(tCK
tCVdt
tdC
mm
+=
− (8.51)
dove C(t) rappresenta concentrazione del chimico a tempo t, Vm è la velocita’ massima del
processo, e Km è la concentrazione dello xenobiotico alla quale la cinetica del processo è uguale alla
metà di Vm. Due importante limiti per questa equazione si hanno:
a) quando la concentrazione del composto è molto più piccola di Km (C (t)<< Km per cui
l’equazione si riduce a:
)()( tCKV
dttdC
mm=
− (8.52)
ed il rapporto di Vm/ Km si approssima ad una costante cinetica apparente del primo ordine,
b) quando la concentrazione dello xenobiotico è molto più più grande di Km (C (t)>> Km)
allora la cinetica é espressa da:
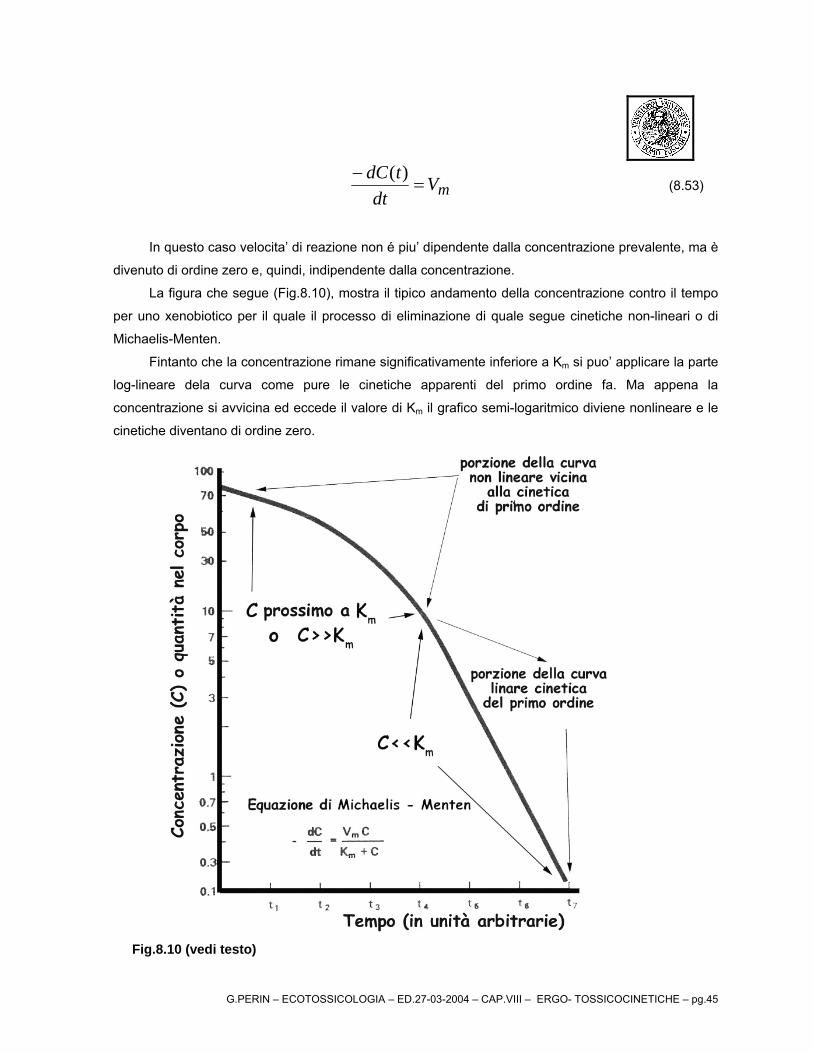
G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.45
mVdt
tdC=
− )( (8.53)
In questo caso velocita’ di reazione non é piu’ dipendente dalla concentrazione prevalente, ma è
divenuto di ordine zero e, quindi, indipendente dalla concentrazione.
La figura che segue (Fig.8.10), mostra il tipico andamento della concentrazione contro il tempo
per uno xenobiotico per il quale il processo di eliminazione di quale segue cinetiche non-lineari o di
Michaelis-Menten.
Fintanto che la concentrazione rimane significativamente inferiore a Km si puo’ applicare la parte
log-lineare dela curva come pure le cinetiche apparenti del primo ordine fa. Ma appena la
concentrazione si avvicina ed eccede il valore di Km il grafico semi-logaritmico diviene nonlineare e le
cinetiche diventano di ordine zero.
Fig.8.10 (vedi testo)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.46
8.3.2.3.0.- Modello monocompartimentale basato su dati stimati Per derivare i valori delle costanti ka e ke in assenza dei dati sperimentali si puo’ utilizzare un
modello basato sull'assorbimento fisiologico.
Qui di seguito considereremo, come esempio, un tipo di procedura applicata al pesce come
modello mono-compartimentale in cui la variazione della concentrazione dello xenobiotico nel tempo
puo’ essere espressa da una relazione del tipo:
dCdt
=E CwRv
F (8.56)
E = efficienza del composto chimico nel passare la membrana branchiale
C, Cw = concentrazioni del tossico nel pesce e nell'acqua, rispettivamente;
Rv = volume dell'acqua che passa attraverso le branchie per unita' di tempo;
F = peso del pesce
La relazione esprime la velocita' di assunzione del tossico come efficacia da parte del pesce
nell'estrarre il composto chimico dall'acqua che fluisce attraverso le branchie. Rv éuna funzione della
concentrazione d’ossigeno nell'acqua.
Poiché la quantita' d’ossigeno necessaria a mantenere il ritmo respiratorio normale nell'animale
acquatico é data dalla relazione:
Rv =Q
EoxCox (8.57)
dove Q = il metabolismo energetico totale del pesce in ml di O2 h-1)
Eox = efficienza del passaggio di O2 attraverso le branchie;
Cox = conc.ossigeno nell'acqua (ml ml-1),
e poiché Q é legato al peso del pesce attraverso la : Q = aFr dove:
r = esponente di crescita del pesce (sec. Norstrom et al. = 0,8)
a = coefficiente di velocita' metabolica in unita' di ml di ossigeno h-1 g-0,8 di pesce

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.47
F = peso del pesce,
con un valore di Eox calcolato in 0,75 (valore é influenzato dalla temperatura e dalla dimensione del
pesce), 11 si puo' ricavare il valore della ka, attraverso le seguenti relazioni:
wCakdtdC
FvRwCE
dtdC ==
(8.58) da cui:
FvER
akFvRwEC
wCak == (8.59)
e sostituendo per E si ha:
( )F
vRowKak 02,0log07,0 −=
(8.60)
ke puo' essere calcolata da
BCFak
ek =
(8.61)
BCF puo' essere ottenuta da una delle seguenti equazioni:
valutata per tutto il corpo del pesce log BCF = 0,76 log Kow - 0,23
valutata sul solo muscolo log BCF = 0,542 log Kow + 0,124
valutata sul solo muscolo log BCF = -0,508 log S + 3,41
11 Il calcolo di E é un po' complesso e richiede alcune considerazioni. La efficienza con cui un agente si muove attraverso le membrane branchiali é un processo di ripartizione e puo' essere valutata attraverso il valore di Kow. La seguente relazione é stata ottenuta da Neely (1979):
02,0log07,0 −= owKE

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.48
8.3.2.4.0.- Esempio di calcolo del modello monocompartimentale
BOX 8.4 Esempio Calcolare l'assunzione di TCDD nella trota di peso 35 g in un acqua alla temperatura di 13°C con
una concentrazione di ossigeno di 10,5 mg L-1. Il coefficiente di crescita del pesce é valutato in 0,8, il
coefficiente metabolico in 0.18 e l'efficienza di transfer dell'ossigeno attraverso le branchie in 0,75. La
solubilità del TCDD in acqua é di 200 ng L-1 e la Kow di 6,1.
Risposta:
Per calcolare ka, valutiamo innanzitutto Q. Q é uguale a Q = aFr
Q=0,18 x (35)0,8 = 3,09
Calcoliamo ora Rv dalla relazione
R v =Q
E ox C ox =
= 3,09/ 80,75) x (7,98.10-3 )
ricordando che 10,5 ppm di ossigeno corrispondono a 7,98.10-3 ml di O2 ml-1 di acqua.
Rv é quindi = 535 ml di acqua h-1
ka é quindi E x Rv/F = (0,07 log Kow-0,02) x 535 /35 = 6,3 ( ml g-1 h-1)
Per calcolare K2 ricorriamo alla BCF = ka/ke da cui ke = ka/BCF
ricordando che:
log BCF = 0,76 log Kow - 0,23 si avra'
log BCF = (0,76 x 6,18) - 0,23 = 4,45 da cui BCF = 28.139
ke é quindi = (6,3 -1)/28.183 = 2,23 x 10-4 (h-1)

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.49
Nell'esempio riportato I dati calcolati sono in eccesso (ka) ed in difetto (ke) rispetto a quelli
sperimentali (4,58 e 5.10-4 rispettivamente) ma forniscono, comunque un buon ordine di grandezza
delle due costanti.
Cio' é probabilmente dovuto alla semplificazione del modello che, nel nostro caso e
monocompartimentale.
Abbiamo cioé presunto che tutto il corpo dell'animale, indipendentemente dalle caratteristiche del
tessuto, si comporti nella stessa maniera.
Sia cioé come un bulk omogeneo. Cio' ovviamente é una semplificazione che in alcuni casi non é
fattibile. Tessuti diversi, in una struttura animale complessa, assumono con cinetiche diverse ed anche
il processo di rilascio o di clearance ha velocita' diverse a seconda del tessuto considerato.
Una valutazione piu' prossima alla realta' puo' essere fornita da un modello bicompartimentale
che implica due compartimenti tissutali uno a rapida ed un altro a lenta cinetica assunzione/clearance.
Nel primo comparto e solo in esso avvengono i processi di assunzione e di clearance mentre nel
secondo vi é un progressivo accumulo dei composti biotossici in funzione della concentrazioni.
8.3.3.1.0.- Il Fattore di Bioaccumulazione Abbiamo gia' visto l'importanza del fattore di Bioconcentrazione (BCF) ed il modo per calcolarlo.
Peraltro, sperimentalmente é strato possibile rilevare come in natura l'accumulo nel corpo animale sia
diverso (e qualche volta di un ordine di grandezza superiore) a quello che si ottiene in laboratorio.
Cio' perché l'accumulo attraverso l'assorbimento cutaneo o branchiale é decisamente, in questo
caso, inferiore a quello che si ha attraverso l'ingestione del tossico attraverso la dieta.
Bisogna quindi tener conto di ulteriori importanti parametri che coinvolgano l'assimilazione del
tossico dal cibo ingerito, il rapporto tra il peso corporeo dell'animale ed il peso del cibo ingerito e la
concentrazione del tossico nell'alimento che viene tratto dall'acqua attraverso l’ingestione di
componenti di livelli trofici inferiori alla specie animale considerata.
La relazione che riassume questi concetti é espressa dalla:
wCfC
KRaN
e
akk
⎟⎟
⎠
⎞
⎜⎜
⎝
⎛+=
3 (8.62)
ove:

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.50
N = Fattore di Bioaccumulazioneed
a = l'efficienza di assimilazione del composto chimico in esame (espressa in grammi di tossico
assorbiti per grammi di tossico ingeriti.
R = rapporto specifico del peso d’alimentazione (grammi di peso ingerito per grammo di peso
corporeo al giorno)
Cf = concentrazione del tossico nel cibo
Cw = concentrazione del tossico nell'acqua.
8.4.0.0.- Bioaccumulazione : Trasferimento attraverso la catena alimentare Un composto chimico, come abbiamo visto, si accumula nel corpo dell'animale con cinetiche piu'
o meno rapide e secondo equilibri di assorbimento e di clearance.
Una parte del tossico viene inoltre metabolizzata nello stesso tessuto animale e permane sotto
forma di metaboliti e cataboliti fino alla eventuale finale eliminazione.
Per parametri conservativi, ossia per composti che non vengono rapidamente metabolizzati lungo i
vari step della catena alimentare, si puo' valutare il progressivo bioaccumulo che porta a processo di
bioaccumulo lungo la food chain o la food web.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.51
La conoscenza degli steps di tale processo d’accumulo e quindi, la possibilità di conoscere la
concentrazione di un tossico in animali a livello trofico progressivamente superiore é di estremo
interesse in ecotossicologia.
La concentrazione di un tossico a ciascun livello trofico (i) é valutata per quel livello dal fattore di
Bioconcentrazione BCF corretto per il contributo dato dal livello trofico inferiore (i-1) conoscendo ,
inoltre, i valori di trasferimento della catena alimentare per la specie trofica superiore considerata, (fi). La relazione che collega i vari livelli é data dalla:
Ni = (BCF) i +ai Ri
K3 (i)Ni−1 (8.63)
così, ad esempio:
{ }
13
223
332
333
3
13
222
333
323
3333
)('''''
)('''
)(
'')(
''')(
''')(
BCFk
Rak
RaBCFk
RaBCF
Nk
RaBCFk
RaBCFNk
RaBCFN
++=
=++=+=
e ponendo:
iii fik
Ra=
)(3 (8.64)
{ }123233
122332333)()()(
)()()(BCFffBCFfBCF
NfBCFfBCFNfBCFN++=
=++=+= (8.65)
Con la stessa procedura e ponendo (BCF)i = Bi , le relazioni per i primi quattro livelli trofici sono:
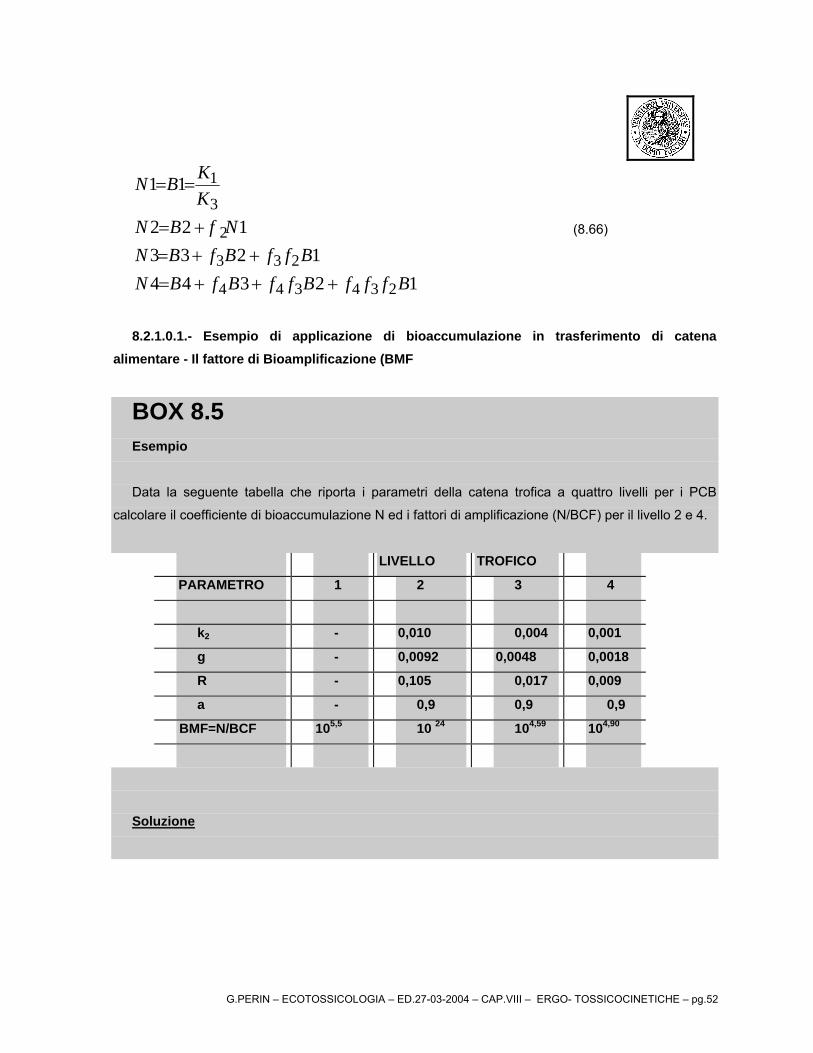
G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.52
123441233
122
11
234344
233
2
3
1
BfffBffBfBNBffBfBN
NfBNKKBN
+++=++=
+=
==
(8.66)
8.2.1.0.1.- Esempio di applicazione di bioaccumulazione in trasferimento di catena
alimentare - Il fattore di Bioamplificazione (BMF
BOX 8.5 Esempio
Data la seguente tabella che riporta i parametri della catena trofica a quattro livelli per i PCB
calcolare il coefficiente di bioaccumulazione N ed i fattori di amplificazione (N/BCF) per il livello 2 e 4.
LIVELLO TROFICO
PARAMETRO 1 2 3 4
k2 - 0,010 0,004 0,001
g - 0,0092 0,0048 0,0018
R - 0,105 0,017 0,009
a - 0,9 0,9 0,9
BMF=N/BCF 105,5 10 24 104,59 104,90
Soluzione

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.53
Il coefficente di bioaccumulazione a livello 2 (N2) é dato dalla somma del BCF del livello 2 (B2) piu’
il prodotto di f2 per il coefficiente di bioaccumulazione al livello 1 ossia N2 = B2 + f2B1. In maniera
analoga si calcola il valore del coefficiente a livello 4 dalla relazione N4 = B4+f4B3+ f4f3B2+ f4f3f2B1.
Calcoliamo quindi i valori dei vari f.
89,20018,0001,0
009,0*9,0''''''''
74,10048,0004,0
017,0*9,0''''''
5,30092,001,0105,0*9,0
''''
2
444
2
333
2
222
=+
=+
=
=+
=+
=
=+
=+
=
gkRaf
gkRaf
gkRaf
I valori dei coefficienti sono allora:
9,45,525,5
59,49,4434344
11,65,529,52
1010*89,2*74,1*5,310*74,1*89,2
10*89,21012344
1010*5,310122
=+
++=+++=
=+=+=
BfffBffBfBN
BfBN
8.5.0.0.- Il fattore MATissueC L'uso del BMF o fattore di biomagnificazione su descritto é usato, inoltre, nella valutazione
modellistica della catena alimentare per la definizione di rischio (environmental risk assessment).
Un approccio consiste nel valutare la concentrazione massima del tossico nel tessuto (MATissueC) senza che intervengano, per l'animale, fenomeni di mortalita'.
In questo caso il BMF consente di calcolare la concentrazione massima possibile nel mezzo
(acqua, sedimento, aria ecc.) chiamata anche criterio biotico probabilistico o PCB. Così ad esempio, in acqua:

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.54
PCBw =MATissueC
BMFw (8.67)
ove con w a pedice si intende che il parametro é calcolato nel mezzo acqua.
8.5.0.0.- La Tossicocinetica per la Definizione dell'Esposizione. La definizione dell'esposizione ad un tossico puo' essere fatta attraverso misure sperimentali che
forniscono il dato piu' obbiettivo atto a definire i fattori di bioconcentrazione necessari, come vedremo,
per correlare il processo di assunzione del tossico alla sua tossicita', estrapolandola, se possibile,
anche all'essere umano.
Sfortunatamente non sempre i dati sperimentali sono disponibili; anzi il piu' delle volte si hanno
solo alcune informazioni sulle caratteristiche del composto oltre a quelle del comportamento
dell'animale usato nell'esperimento.
É peraltro possibile ottenere da queste sole informazioni, dati utilizzabili per il prosieguo della
valutazione tossicologica; attraverso proprio le conoscenze della fisiologia dell'animale usato (in
questo caso il pesce) e le caratteristiche del composto chimico.
Vedremo, piu' avanti, quando esamineremo le tecniche del QSAR, come si possa arrivare a piu'
dettagliate valutazioni di correlazione tra alcune caratteristiche strutturali del composto chimico e la
sua azione tossica e/o il suo comportamento tossicocinetico.
Per ora esaminiamo come é possibile valutare le costanti K1 e K2 nonché il BCF da dati presunti
sia dell'animale sperimentato che del composto chimico esaminato.
L'esame tossicocinetico verra' fatto secondo due procedure: la prima che considera che il tossico si
assorba in maniera uniforme in tutto l'organismo dell'animale (modello ad un compartimento), la
seconda che i tessuti assorbano e rilascino in modo diverso tra loro il tossico considerato (modello a
due compartimenti)
8.6.0.0.- Il fattore di applicazione (AF e MATC) La determinazione della tossicita' cronica presenta notevoli difficolta' poiché pochi sono gli individui
che si prestano a tempi lunghi di sperimentazione.
Un altro fatto che crea non pochi problemi é la mancanza di relazioni significative tra la tossicita'
acuta e quella cronica di un composto chimico tra mammiferi, pesci e la stessa Daphnia.

G.PERIN – ECOTOSSICOLOGIA – ED.27-03-2004 – CAP.VIII – ERGO- TOSSICOCINETICHE – pg.55
Un processo molto usato per predire la tossicita' cronica da dati di tossicita' acuta, specialmente tra
specie molto simili, é quello di calcolare il rapporto tra LC50 e quella che é definita come la
concentrazione del tossico alla quale cronicamente non si riscontra alcun effetto (MATC = Maximum
Acceptable Toxicant Concentration) in stadi di vita dell'animale particolarmente sensibili o nell'arco
totale della vita.
Questo rapporto é conosciuto come il fattore di applicazione AF (Application Factor) e quindi:
AF =LC 50
MATC (8.68)
I valori per gli AF per 31 composti di varia struttura chimica e di interesse ambientale variano da 2
a 3725.
Non sempre é possibile avere i dati sperimentali necessari per calcolare AF; in questo caso si puo'
estrapolare da valori esistenti per composti simili. Utilizzando il criterio del ubi minor, maior cessat
ossia quel criterio di sicurezza che consiste nello scegliere (nell'impossibilità di avere dati sicuri) le
condizioni piu' favorevoli all'ambiente, si puo' avere una certa informazione in massima sicurezza
nell'attesa, comunque di ottenere sperimentalmente i dati necessari (che probabilmente riducono i
termini di rischio).
Per esempio, per il 1,1,1,-tricloroetano (TCE) non ci sono dati per la sua tossicita' cronica. D'altro
canto egli fa parte di una numerosa schiera di composti similari per i quali ci sono dati abbondanti. Gli
AF di nove cloroderivati organici simili variano da 1 a 35; su tale base si puo' scegliere un fattore
conservativo di 35 (AF=35); conoscendo il valore della tossicita' acuta (che esiste e, comunque
sarebbe facile ottenere sperimentalmente) che é di 5 mg l-1, si puo' calcolare il MATC di 145 µg L-1 in
acqua. Questo MATC fornisce almeno un fattore di sicurezza di 14 volte al di sopra della
concentrazione media di meno di 10 µg L-1 di TCE che si puo' riscontrare nell'ambiente (escludendo,
ovviamente, i casi di incidente grave).


















