
La giustizia presso i Sumeri e i Babilonesi ipertesto della prof.ssa M. G. Desogus.

- 1 -
CINETICHE DI CRESCITA BATTERICA
DI INTERESSE PER LA SICUREZZA
ALIMENTARE IN CAMPI IRRADIATI
CON MICROONDE A BASSA POTENZA
Francesco Desogus
DOTTORATO DI RICERCA IN INGEGNERIA INDUSTRIALE UNIVERSITÀ DEGLI STUDI DI CAGLIARI XXII CICLO

- 2 -

- 3 -

- 4 -
Questa Tesi può essere utilizzata nei limiti stabiliti dalla normativa vigente sul Diritto
d’Autore (Legge 22 aprile 1941 n. 633 e succ. modificazioni e articoli da 2575 a 2583
del Codice Civile) ed esclusivamente per scopi didattici e di ricerca; è vietato qualsiasi
utilizzo per fini commerciali. In ogni caso tutti gli utilizzi devono riportare la corretta
citazione delle fonti. La traduzione, l’adattamento totale e parziale sono riservati per
tutti i Paesi. I documenti depositati sono sottoposti alla legislazione italiana in vigore
nel rispetto del Diritto d’Autore, da qualunque luogo essi siano fruiti.

- 5 -
CINETICHE DI CRESCITA BATTERICA
DI INTERESSE PER LA SICUREZZA
ALIMENTARE IN CAMPI IRRADIATI
CON MICROONDE A BASSA POTENZA
FRANCESCO DESOGUS
Tutor: Prof. Ing. Renzo Carta
Dottorato di Ricerca in Ingegneria Industriale Università degli Studi di Cagliari XXII Ciclo

- 6 -

INDICE
- 7 -
INDICE
INTRODUZIONE ......................................................................................11
1. LE ORIGINI E I PRESUPPOSTI DEL LAVORO
1.1 La sicurezza alimentare........................................................................... 15
1.2 Il rischio microbiologico negli alimenti .................................................. 18
1.3 I trattamenti nell’industria alimentare .................................................. 23
1.4 L’irradiazione e l’utilizzo delle microonde in campo alimentare ........ 27
1.5 Prospettive e vantaggi dell’utilizzo di microonde a bassa potenza ...... 32
2. I FONDAMENTI SCIENTIFICI
2.1 La radiazione elettromagnetica e le microonde..................................... 37
2.2 L’interazione tra radiazioni elettromagnetiche e reazioni chimiche ... 40
2.3 Gli effetti biologici della radiazione elettromagnetica .......................... 44
2.4 Effetti termici versus effetti non termici delle microonde .................... 46
2.4.1 Effetti fisico-chimici e assorbimento delle microonde................................51
2.4.2 Effetti delle microonde sulle velocità di crescita e di morte cellulari .......53
2.4.3 Alterazioni cromosomiche ed effetti genetici delle microonde ..................58
2.4.4 Effetti delle microonde su tessuti eccitabili e membrane...........................59
2.4.5 Effetti sull’omeostasi degli ioni Ca2+............................................................63
2.4.6 Altri effetti delle microonde in vitro ............................................................65
2.4.7 Considerazioni sui meccanismi degli effetti non termici............................67
2.5 Le specie microbiche indagate ................................................................ 68
2.5.1 Il Bacillus clausii ...........................................................................................68
2.5.2 La Pseudomonas aeruginosa.........................................................................70
2.5.3 Lo Staphylococcus aureus .............................................................................74
2.5.4 La Listeria monocytogenes ............................................................................75

INDICE
- 8 -
3. LA SPERIMENTAZIONE
3.1 Il progetto sperimentale........................................................................... 81
3.2 Le necessità della sperimentazione ......................................................... 85
3.3 L’apparato sperimentale ......................................................................... 86
3.3.1 La “parte biologica” .....................................................................................86
3.3.2 La “parte elettronica”...................................................................................91
3.4 Le determinazioni analitiche ................................................................... 94
3.4.1 La determinazione dello “Specific Absorption Rate” ................................94
3.4.2 La misurazione della densità ottica .............................................................99
3.4.3 La conta in piastra ......................................................................................104
3.4.4 Densità ottica e conta in piastra a confronto ............................................107
3.5 La determinazione delle costanti di crescita ........................................ 110
4. I RISULTATI DELLA RICERCA
4.1 Le costanti di crescita sperimentali ...................................................... 117
4.2 Il confronto tra i dati a potenza variabile ............................................ 120
4.3 Il confronto tra i dati a frequenza variabile ........................................ 125
4.4 Commenti conclusivi e prospettive future ........................................... 130
BIBLIOGRAFIA ......................................................................................133
Ringraziamenti .................................................................................................... 153



INTRODUZIONE
- 11 -
INTRODUZIONE
Il presente lavoro di tesi è frutto dell’attività di ricerca triennale svolta
nell’ambito del corso di Dottorato di Ricerca, e si inserisce in un programma di
ricerca di più ampio respiro sugli effetti che l’irradiazione con microonde è in grado
di produrre sulla velocità di evoluzione di processi chimici e biologici.
I contenuti della trattazione sono costituiti dalla valutazione sperimentale degli
effetti non termici prodotti da microonde a bassissima potenza su quattro specie
batteriche, che sono il Bacillus clausii, la Pseudomonas aeruginosa, lo
Staphylococcus aureus e la Listeria monocytogenes, ed i risultati possono essere
ritenuti di interesse nell’ambito delle complesse problematiche relative alla sicurezza
alimentare.
Proprio per via di tale complessità, viene spiegato inizialmente il concetto stesso
di “sicurezza alimentare”, e si riportano delle informazioni di carattere generale sui
trattamenti più o meno diffusamente applicati nel vasto settore dell’industria
alimentare. Lo scopo di ciò, più che di effettuare una trattazione di per sé esaustiva,
è quello di fornire un inquadramento via via più specifico alla tematica principale, di
cui è certamente innegabile l’elevato grado di innovatività rispetto al panorama delle
attuali tecnologie, che sono basate essenzialmente su effetti di tipo termico.
Successivamente sono passati in rassegna (anche qui in modo forse non del tutto
esauriente, ma comunque in misura ampia) precedenti lavori e studi scientifici sul
riscontro (o meno) di effetti di varia natura (principalmente biologici) prodotti
dall’esposizione a campi elettromagnetici. Ciò sia perché si è ritenuto quantomeno
necessario fornire una sorta di “stato dell’arte”, sia perché è funzionale alla
discussione sulle relazioni tra effetti termici ed effetti non termici, e sia perché è
utile a dare perlomeno un’idea della complessità, sia teorica che pratico-
sperimentale, del problema affrontato. Solo a questo punto vengono presentati, nel
loro specifico, il progetto sperimentale, la struttura utilizzata e le tecniche analitiche
adottate.

INTRODUZIONE
- 12 -
I risultati ottenuti, frutto di un’intensissima attività, e la loro non facile
interpretazione, come si avrà modo di vedere, pur lasciando aperti degli
interrogativi, allo stesso tempo fungono indubbiamente da stimolo per la curiosità
scientifica che dovrà accompagnare gli ulteriori sviluppi a cui la ricerca andrà
incontro, e per i quali la definizione e la messa a punto delle strutture e delle
complesse metodologie che sono state compiute in questi anni costituiranno una
solida ed importante base di partenza.

CAPITOLO 1


CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 15 -
1.1 La sicurezza alimentare
Con l’espressione “sicurezza alimentare” si è tradizionalmente indicata la
possibilità di garantire, in modo costante e generalizzato, acqua ed alimenti in
quantità sufficienti al soddisfacimento del fabbisogno energetico dell’essere umano,
al fine di consentirne la sopravvivenza. A livello globale, poiché sono ancora
numerose le aree del pianeta dove il problema dell’accesso all’acqua e al cibo da
parte di intere popolazioni persiste, e dove questo è ben lungi da soluzioni di
carattere definitivo, tale definizione continua a mantenere il suo significato
originario.
Nei Paesi industrializzati le popolazioni, o perlomeno per una buona parte di
queste, hanno ormai raggiunto elevati livelli di benessere; in Italia, per esempio, a
partire dal secondo dopoguerra, grazie al forte sviluppo economico che ha
interessato più o meno tutte le classi sociali, il problema della fame può dirsi
debellato, e lo stesso è avvenuto, pur talvolta in tempi e con modalità differenti, in
altre società. Questa evoluzione si è riflessa, tra le altre cose, anche sul concetto
stesso di “sicurezza alimentare”, con il quale, perlomeno quando ci si riferisce ai
Paesi industrializzati, poiché si dà per scontata la disponibilità di cibo e acqua, si
intende piuttosto la possibilità di approvvigionamento di questi in condizioni
igieniche; pertanto la dicitura “sicurezza alimentare” può essere ormai, alla luce
delle considerazioni appena fatte, intesa come sinonimo di “igiene alimentare”, il cui
scopo deve essere quello di garantire la buona qualità di un cibo o di una bevanda
sotto il profilo igienico e sanitario.
E sono proprio i forti cambiamenti economici e sociali che hanno trasformato le
società durante il processo di industrializzazione a trovare una diretta
corrispondenza nelle profonde modificazioni che hanno parallelamente interessato il
sistema alimentare, passato da uno stato in cui il produttore dei generi alimentari ne
era, nella maggior parte dei casi, anche il consumatore, ad una situazione di sempre
meno stretto rapporto tra produzione e consumo, diventati entrambi di massa e

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 16 -
assoggettati quasi esclusivamente alle logiche economiche di mercato. Merci in
viaggio su distanze spesso lunghe, scambi commerciali, prodotti provenienti da
Paesi la cui legislazione in materia agricola e alimentare è meno stringente, ma
anche fenomeni come la grande distribuzione e la ristorazione di massa, con il
necessario largo uso di prodotti conservati, che devono spesso essere mantenuti
all’interno della catena del freddo, hanno aperto scenari con problemi e punti critici
sempre nuovi, cui le normative in materia, e di conseguenza la tecnologia, hanno
dovuto trovare delle soluzioni. Inoltre, il continuo lancio sul mercato di nuovi
prodotti alimentari fa sì che l’innovazione tecnologica in questo campo sia
notevolmente rapida e oggetto di ingenti (ma comunque remunerativi) sforzi
economici da parte dei produttori. Evidentemente questi ultimi non possono essere,
comunque, gli unici soggetti interessati, e l’adozione di prassi idonee alla
salvaguardia dei necessari requisiti di igiene e salubrità dei prodotti alimentari deve
interessare tutta la filiera e dunque, oltre agli stessi produttori, anche tutti coloro che
intervengono nei successivi passaggi che il prodotto subisce fino a giungere al
consumatore finale.
A livello mondiale, gli organismi maggiormente impegnati in materia di
sicurezza alimentare sono l’Organizzazione delle Nazioni Unite per l’Alimentazione
e l’Agricoltura (FAO) e l’Organizzazione Mondiale della Sanità (OMS); nel 1963 le
due istituzioni hanno dato vita al “Codex Alimentarius”, un programma creato al
fine di sviluppare standard e linee guida orientati alla protezione della salute dei
consumatori, promuovendo la lealtà delle pratiche eseguite nel commercio dei
prodotti alimentari e il coordinamento dei lavori in materia di normativa alimentare
svolti da organizzazioni internazionali governative e non governative (Joint
FAO/WHO Food Standards Programme, 2006). La Commissione preposta, istituita
allo scopo, ha la responsabilità di mettere a punto le norme e, dopo una valutazione
da parte dei governi nazionali, di pubblicarle come standard, linee guida o
raccomandazioni a livello regionale o mondiale, a seconda del caso; può trattarsi di
norme generali o specifiche, legate ad un particolare alimento o a un gruppo di

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 17 -
alimenti, o concernenti i processi produttivi o ancora le modalità di
regolamentazione nazionale per la sicurezza e la salute dei consumatori.
È più recente l’interessamento europeo in materia di sicurezza alimentare, o
quantomeno lo è l’elevazione della sicurezza alimentare a priorità: nel 2000 è stato
presentato il “Libro Bianco sulla Sicurezza Alimentare” della Commissione Europea
(Commissione delle Comunità Europee, 2000), che descriveva l’insieme delle azioni
necessarie al completamento e alla modernizzazione della legislazione dell’Unione
Europea in materia di alimentazione, al fine di renderla più coerente e di consentirne
al meglio l’applicazione. A seguito di ciò, e con lo scopo di adottare un piano
d’azione integrato che coniugasse qualità e sicurezza nel rispetto delle produzioni
tipiche, l’Unione Europea ha dato vita, nel 2002, all’Autorità Europea per la
Sicurezza Alimentare (EFSA, “European Food Safety Authority”), con sede a
Parma, che è dotata di una commissione di esperti e tecnici, indipendenti dai
rispettivi governi, con funzioni consultive (da parte della Commissione Europea) e
di indirizzo per le scelte di politica agroalimentare e sanitaria; nel 2004 in Italia è
stato creato il Comitato Nazionale per la Sicurezza Alimentare (CNSA), con
funzioni di interfaccia dell’EFSA.
In linea generale, gli strumenti utili a raggiungere gli obiettivi di sicurezza
alimentare, che sono molteplici, possono essere obbligatori, in quanto imposti dalla
Legge, o non obbligatori ma comunque opportuni. Tra quelli obbligatori vi è
l’HACCP (“Hazard Analysis and Critical Control Points”), che è un sistema di
autocontrollo che i produttori di alimenti devono mettere in atto al fine di valutare i
pericoli e stimare i rischi connessi col processo produttivo per poter stabilire misure
di controllo e prevenzione dell’insorgenza di problemi igienici e sanitari. La novità
introdotta dal sistema HACCP consiste nello spostamento dei controlli, prima
effettuati a valle del processo produttivo sul prodotto finito, verso tutte le fasi della
produzione, consentendo l’attuazione di misure preventive; i controlli sono anche
estesi alle fasi di distribuzione dell’alimento, ugualmente soggette alla presenza di
punti critici. Il sistema HACCP fu introdotto nell’Unione Europea con la Direttiva

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 18 -
43/93/CEE (recepita in Italia con il D. Lgs. 155/97), che prevede l’obbligo di
applicarlo per tutti gli operatori del settore alimentare; tale normativa è stata poi
sostituita dal Regolamento CE 852/2004, recepito in Italia con il D. Lgs. 193/07, che
ha esteso l’applicazione dell’HACCP anche al settore della produzione agricola e ne
ha reso alcuni aspetti maggiormente flessibili nell’applicazione alle piccole imprese.
Tra le misure invece non obbligatorie, e dunque applicate su base volontaria, vi sono
quelle riassunte e codificate nello Standard ISO 22000:2005 (allineato con le
precedenti norme ISO 9000 e ISO 14000), che norma la tracciabilità delle filiere
agroalimentari e armonizza gli standard nazionali e internazionali preesistenti in
materia di sicurezza alimentare e HACCP; in base a questo, ogni singolo anello delle
catene di produzione e distribuzione degli alimenti viene interessato, compresi gli
operatori coinvolti in modo indiretto, come per esempio i produttori degli
imballaggi, i servizi di pulizia e di disinfestazione, etc..
Quanto detto sinora mostra come vi sia la necessità di norme chiare e allo stesso
tempo sufficientemente dettagliate, in quanto la sicurezza alimentare può essere
garantita solo con l’applicazione di adeguate pratiche di prevenzione e controllo dei
rischi nella produzione e nella manipolazione degli alimenti. Se tali misure non
vengono applicate, o non sono applicate correttamente, gli alimenti possono divenire
oggetto di contaminazioni da parte di diversi agenti, di natura fisica, chimica e/o
microbiologica, e perciò causa di numerose patologie, talvolta non gravi, ma che in
alcuni casi possono anche rivelarsi letali.
1.2 Il rischio microbiologico negli alimenti
Tra i principali fattori di rischio (di natura fisica, chimica e microbiologica)
connessi con l’assunzione di alimenti vi sono prodotti fitosanitari, contaminanti
organici persistenti (come policlorobifenili, idrocarburi policiclici aromatici,
policloro-dibenzo-p-diossine, policloro-dibenzo-furani), metalli pesanti (come

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 19 -
cadmio, piombo, arsenico, mercurio), OGM (Organismi Geneticamente Modificati),
micotossine e organismi patogeni di vario genere (Tiecco, 2001); questi agenti
possono entrare in contatto con gli alimenti in quanto normalmente presenti
nell’acqua, nell’aria, nel suolo, oppure a causa di sistemi di produzione e
conservazione igienicamente non adeguati (quando non si tratti di manipolazioni
fraudolente). Ci si concentrerà, in questo paragrafo, sul solo rischio di natura
microbiologica, in quanto funzionale con il resto della trattazione e con gli obiettivi
del lavoro sperimentale descritto nella presente tesi.
Gran parte delle specie di microorganismi è non solo utile, ma spesso addirittura
necessaria per la vita umana, basti pensare ai batteri che colonizzano l’intestino e
che vivono in un vero e proprio rapporto simbiotico con l’uomo. La presenza di
microorganismi negli alimenti, che in linea generale può essere dovuta alla naturale
contaminazione di questi ultimi negli ambienti di produzione e lavorazione o
all’aggiunta volontaria di innesti (naturali o selezionati) da parte dell’uomo, è spesso
inevitabile, e altrettanto spesso necessaria, in quanto differenti specie e biotipi sono
parte integrante di numerosi alimenti fermentati, per la preparazione dei quali
svolgono un ruolo centrale (Bourgeois et al., 1990). Tra questi vi sono batteri, lieviti
e muffe che sono essenziali per la trasformazione di alcuni prodotti agricoli in
alimenti con caratteristiche di commestibilità e conservabilità enormemente
superiori, basti pensare alle applicazioni più antiche, come la panificazione e la
vinificazione (che hanno luogo grazie a saccaromiceti), e poi ad innumerevoli
prodotti caseari (ottenuti con lattofermenti e muffe), all’aceto, ai salumi, ai prodotti
da forno, alla birra, e a numerosissimi altri alimenti di normale uso quotidiano; vi
sono poi anche i batteri lattici (Lactobacillus, Bifidobacterium, Enterococcus, etc.),
che sono legati alla preparazione di latti acidi (come yogurt e simili) e che, se
ingeriti vivi in numero sufficientemente elevato, possono avere effetti positivi sulla
salute dell’organismo ospite, rafforzandone l’ecosistema intestinale, al punto che
sono considerati essi stessi un alimento e sono definiti probiotici per via dei loro
effetti benefici (Galli Volonterio, 2005). Molti microorganismi sono parte essenziale

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 20 -
di processi biochimici che consentono la trasformazione della materia prima in
numerosi derivati con composizione chimica, struttura e aroma anche parecchio
differenti, agendo sia come entità cellulari che tramite estratti enzimatici
appositamente rilasciati su zuccheri, proteine e lipidi; in questo senso, le diverse
tecnologie produttive, a seconda di come selezionano i biotipi microbici e ne
definiscono lo sviluppo, risultano determinanti sulla qualità percepita dal
consumatore (Ray, 2004).
Può tuttavia capitare, in particolar modo se non vengono applicate rigorose
regole igieniche nella preparazione, nella cottura o nella conservazione degli
alimenti, che le popolazioni microbiche eventualmente presenti nell’alimento
possano crescere oltre certi limiti, provocando trasformazioni indesiderate quali
ammuffimenti, marcimenti, formazione di mucillagini, insorgenza di odori e sapori
anomali, che si traducono nel degrado delle caratteristiche di qualità e di
commestibilità e dunque nella commerciabilità dell’alimento stesso (Galli
Volonterio, 2005). Può trattarsi, nella maggior parte dei casi, di microorganismi di
specie indesiderate che hanno contaminato l’alimento alla fonte (contaminazione
primaria) o durante la lavorazione (contaminazione secondaria): il primo caso è
quello relativo, per esempio, alle carni di animali portatori di alcune malattie o
batteri, o a verdure, frutta, pesci, uova o latte già contaminati in partenza; la seconda
eventualità è inerente agli ambienti, alle attrezzature, ai comportamenti degli
operatori e all’eventualità che l’uomo stesso sia portatore sano di malattie o batteri
(Bourgeois et al., 1990). Spesso però (e qui sta forse il vero problema) gli alimenti
contaminati non presentano un aspetto (colore, consistenza, etc.), odori o sapori
anomali, e dunque non destano sospetti e vengono ugualmente consumati: ecco che
l’ingestione di questi può causare malattie (tossinfezioni) alimentari da
microorganismi patogeni, che sono stati patologici conseguenti al consumo di
alimenti contenenti i microorganismi patogeni o anche solo una tossina di origine
batterica (Tiecco, 2001). Vi sono a proposito almeno due casi possibili: il primo è
quello delle intossicazioni, che sono causate dalla presenza, nell’alimento, di tossine

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 21 -
prodotte in quantità sufficienti dai microorganismi (detti perciò tossigeni) che sono
riusciti a moltiplicarsi nell’alimento prima della sua ingestione, per i quali l’alimento
ha rappresentato il substrato indispensabile alla crescita; essendo la tossina e non i
microorganismi la causa diretta della sintomatologia, questi ultimi possono anche
non essere più presenti nell’alimento. La seconda possibilità è quella delle
tossinfezioni in senso stretto che sono dovute, invece, all’attività combinata di
tossine e microorganismi che le producono, i quali possono continuare a
moltiplicarsi nell’intestino; in questa ultima eventualità non è strettamente
necessario che la popolazione batterica si sia moltiplicata già nell’alimento prima
della sua ingestione, e tali infezioni possono perciò svilupparsi anche con una carica
infettante relativamente modesta: si possono così avere infezioni enteroinvasive,
nelle quali si ha la semplice moltiplicazione dei microorganismi nel tratto
intestinale, e infezioni enterotossiche, nelle quali il microorganismo infettante
produce le enterotossine all’interno del tratto intestinale.
I fattori che possono favorire o meno la colonizzazione di un alimento e lo
sviluppo in esso di microorganismi sono essenzialmente, oltre alla composizione
chimica dell’alimento (presenza di sali, zuccheri, etc.), temperatura, umidità,
pressione parziale di ossigeno, potenziale di ossido-riduzione e acidità (Fellows,
2000); gli alimenti maggiormente soggetti al fenomeno sono carni e derivati,
prodotti a base di uova, latticini e derivati (come gelati e creme), prodotti ittici e
salse.
Le patologie forse più note e pericolose sono il botulismo (causato dal
Clostridium botulinum) e la salmonellosi (dovuta al batterio Salmonella
typhimurium), ma diversi e anche gravi disturbi possono essere arrecati da alimenti
contaminati da Escherichia coli, Clostridium perfringens, Staphylococcus aureus,
Streptococcus faecalis, Bacillus cereus, Listeria monocytogenes, Yersinia
enterocolitica, Campylobacter jejuni, Vibrio parahoemolyticus, Toxoplasma gondii
(Bourgeois et al., 1990). In linea generale tali microorganismi patogeni, oltre ad
ambienti prossimi alla neutralità, prediligono temperature prossime ai 37°C (che è la

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 22 -
temperatura corporea), considerazione sulla quale, come verrà meglio spiegato più
avanti (paragrafo 3.1), è stato impostato il lavoro sperimentale oggetto della presente
tesi.
Alla luce di tutto quanto detto sinora, risulta evidente la necessità di garantire
opportune condizioni di sicurezza adottando adeguate tecniche di confezionamento,
conservazione, distribuzione, commercializzazione e somministrazione, in modo tale
da evitare la creazione di condizioni per le quali nell’alimento, che spesso già di per
sé è un ottimo terreno di coltura, sia consentito lo sviluppo dei microorganismi che
sono di norma già presenti come fatto fisiologico e naturale: solo così è infatti
possibile ridurre al minimo il rischio di contaminazione degli alimenti e di
conseguenza proteggere la salute umana. Nei Paesi industrializzati ormai, grazie
all’utilizzo di appropriate tecnologie e al buon livello igienico-sanitario, i problemi
di contaminazione e sviluppo microbico negli alimenti possono dirsi forse limitati,
ma in alcuni casi specifici, o per taluni soggetti a rischio, come anziani,
immunodepressi o portatori di patologie debilitanti, possono rivelarsi anche piuttosto
gravi (Tiecco, 2001). E in ogni caso, nonostante la messa in atto di tutti i più
moderni sistemi di prevenzione e delle procedure di autocontrollo (come l’HACCP),
allo stato attuale non è possibile escludere che delle contaminazioni possano avere
luogo, seppure in una piccola o piccolissima percentuale dei casi, sia perché, come
detto, i microorganismi potenzialmente pericolosi pervadono qualsiasi ambiente e
sono diffusamente presenti nel mondo vegetale e animale, sia perché, per esigenze di
mercato, una fetta sempre più consistente della produzione alimentare si sta
spostando verso gli alimenti “minimamente trattati”, cioè il più possibile simili a
quelli freschi, e dunque sottoposti quanto meno possibile a trattamenti severi e
all’utilizzo di additivi e conservanti: l’obiettivo diventa quello di controllare la
moltiplicazione microbica in modo tale da evitare che la numerosità batterica superi
determinate soglie di nocività (Galli Volonterio, 2005). Dunque non si può, né forse
si potrà mai, parlare di eliminazione, bensì solo di minimizzazione, del rischio.

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 23 -
1.3 I trattamenti nell’industria alimentare
L’utilizzo delle tecnologie nel settore alimentare ha sempre avuto,
essenzialmente, un duplice scopo: migliorare le caratteristiche di appetibilità e di
digeribilità degli alimenti e garantirne la conservabilità fino al consumatore.
Oggigiorno, tra gli obiettivi perseguiti, vi sono sicuramente lo sviluppo di nuovi
prodotti, il conferimento all’alimento di determinate caratteristiche (magari più
vicine alle richieste del mercato), il miglioramento delle proprietà nutrizionali e
organolettiche, oltre naturalmente al garantimento delle condizioni di sicurezza. Uno
dei primi processi nati con il principale obiettivo della tutela della salute pubblica fu
la pastorizzazione del latte. Le tecnologie alimentari rivestono un ruolo centrale
nell’HACCP, in quanto hanno molto spesso un ruolo rilevante nel controllo del
rischio, e la comprensione dei meccanismi che le caratterizzano e dei parametri che
influenzano i processi è necessaria, se non fondamentale, per la corretta scelta delle
misure di controllo e per l’efficacia di queste, che spesso consistono
nell’applicazione di una combinazione, anche complessa, di trattamenti.
I trattamenti alimentari (che possono essere di tipo fisico o chimico), in linea del
tutto generale, possono essere indirizzati verso tre diversi aspetti: rendere sicuro un
determinato alimento, controllare la presenza di eventuali contaminanti (prevenendo
la moltiplicazione di microorganismi o la produzione di tossine), prevenire la
ricontaminazione dopo l’applicazione dei trattamenti (Fellows, 2000). Come già si è
avuto modo di spiegare, rendere sicuro (dal punto di vista microbiologico così come
da tutti gli altri) un alimento non significa annullare completamente il rischio, bensì
ridurlo ad un livello accettabile; tra i trattamenti volti a ciò vi sono i trattamenti
termici tradizionali, l’irradiazione, la disinfezione chimica, congelamento e
surgelazione (applicabili in questo caso solo contro i parassiti), l’uso di alte
pressioni. I trattamenti termici costituiscono sicuramente la categoria più comune (e
forse, allo stato attuale, più efficace) per la distruzione dei batteri patogeni; vi sono
cottura, arrostimento, bollitura, frittura, pastorizzazione e sterilizzazione, ottenibili, a
CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 24 -
seconda dei casi, per contatto con aria calda, vapore, acqua calda o con una
superficie riscaldata, o mediante riscaldamento con microonde; il principio
fondamentale sul quale si basano risiede nel comportamento dei microorganismi alle
differenti temperature. Tutte le specie di microorganismi, infatti, hanno un intervallo
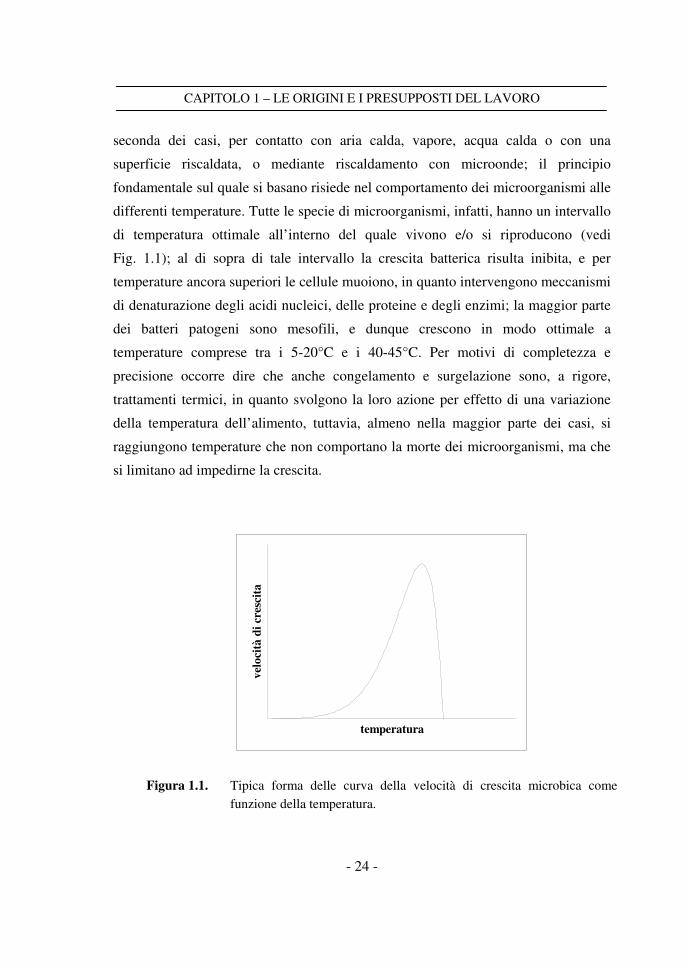
di temperatura ottimale all’interno del quale vivono e/o si riproducono (vedi
Fig. 1.1); al di sopra di tale intervallo la crescita batterica risulta inibita, e per
temperature ancora superiori le cellule muoiono, in quanto intervengono meccanismi
di denaturazione degli acidi nucleici, delle proteine e degli enzimi; la maggior parte
dei batteri patogeni sono mesofili, e dunque crescono in modo ottimale a
temperature comprese tra i 5-20°C e i 40-45°C. Per motivi di completezza e
precisione occorre dire che anche congelamento e surgelazione sono, a rigore,
trattamenti termici, in quanto svolgono la loro azione per effetto di una variazione
della temperatura dell’alimento, tuttavia, almeno nella maggior parte dei casi, si
raggiungono temperature che non comportano la morte dei microorganismi, ma che
si limitano ad impedirne la crescita.
temperatura
vel
oci
tà d
i cr
esci
ta
Figura 1.1. Tipica forma delle curva della velocità di crescita microbica come
funzione della temperatura.

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 25 -
Poiché il processo di morte delle cellule segue in linea di massima una cinetica
del primo ordine (Teixeira, 1992), indicando con N il numero di cellule presente in
un dato alimento al tempo t, con 0N il numero di cellule presente al tempo 0t , e con
Dk la costante cinetica del processo di morte, si ha:
( )[ ]0D0 t-tkexpNN ⋅−⋅= (1.1)
Nella pratica (Teixeira, 1992; Ramaswamy e Singh, 1997) ci si riferisce al cosiddetto
“tempo di riduzione decimale”, comunemente indicato con D, che è il tempo
necessario, ad una data temperatura, affinché la popolazione microbica si riduca del
90%; D rappresenta dunque una misura della resistenza termica di una determinata
specie di microorganismi a quella temperatura, ed è pari a:
=
N
NLog
t-tD
0
0 (1.2)
La relazione tra D e Dk , intervenendo un cambiamento di base del logaritmo, è la
seguente:
( )Dk
110lnD ⋅= (1.3)
Tra le specie che mostrano una maggiore resistenza al calore vi sono Staphylococcus
aureus e Listeria monocytogenes (Ramaswamy et al., 1989); le spore sono di norma
molto più resistenti, e sono distrutte solo da temperature superiori a 100°C, e tra le
spore più resistenti vi sono quelle del Clostridium botulinum. Il valore di D, oltre
che dipendere dalla specie microbica, è influenzato dalle caratteristiche chimico-
fisiche del mezzo (attività dell’acqua, acidità, composizione, etc.) e dallo stato
fisiologico delle cellule (età, stato di crescita, etc.) (Ramaswamy e Singh, 1997).
Ramaswamy et al. (1989) riportano una nutrita gamma di dati cinetici relativi a
diverse specie e condizioni ambientali.

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 26 -
I trattamenti termici hanno effetti non solo sulle popolazioni microbiche, ma
anche su caratteristiche e proprietà dell’alimento quali consistenza, colore, sapore,
enzimi, vitamine, etc.; per questo è sempre necessario ricercare il miglior
compromesso possibile tra l’abbattimento del rischio e i cambiamenti (degradativi)
di composizione, nutrizionali e organolettici. Da questa esigenza è nato un processo
come quello della pastorizzazione, che può essere condotto a diverse combinazioni
temperatura-tempo (Jay et al., 2009): a bassa (63°C per 30 minuti), ad alta (72°C per
15 secondi) e ad altissima (135°C per 1 secondo) temperatura; tuttavia permangono,
anche in casi come questo, i problemi legati al fatto che la temperatura non è
uniforme in tutto il corpo dell’alimento per via della naturale formazione di gradienti
che dipendono dalle caratteristiche (composizione, struttura, stato fisico) del
prodotto, dalle sue dimensioni, dalla forma e dalle modalità di trasferimento del
calore dal mezzo riscaldante al prodotto stesso. Per completezza si cita qui anche il
riscaldamento ohmico degli alimenti, ottenuto con il passaggio di una corrente
elettrica attraverso il materiale da trattare. L’utilizzo di campi elettrici per ottenere la
pastorizzazione di alimenti, infatti, è stato riportato fin dal 1935 da Getchell (1935),
che fece passare della corrente alternata direttamente attraverso del latte, generando
così del calore che aveva l’effetto di inattivare i microorganismi presenti.
Attualmente le tecnologie di conservazione degli alimenti basate sull’utilizzo di forti
campi elettrici pulsati vengono sviluppate con l’intento di incontrare gli standard
commerciali dei prodotti (Zhang et al., 1994), e sono tanto più promettenti quanto
più cresce la domanda di prodotti commerciali alimentari simili ai prodotti freschi,
senza perdite eccessive in termini di aromi, nutrienti e vitamine; Zhang et al. (1995)
hanno studiato un processo che combinasse un trattamento alimentare continuo con
campi elettrici pulsati e il confezionamento in condizioni asettiche.
Tra i trattamenti volti a controllare lo sviluppo di microorganismi vi sono
quelli basati sulla riduzione dell’acqua libera (essiccamento, cristallizzazione,
congelamento, utilizzo di agenti leganti, etc.), che ha un peso notevole sulla velocità
di crescita e sulla produzione di tossine in quanto condiziona le cinetiche delle

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 27 -
reazioni chimiche ed enzimatiche, anche se vi sono specie, come lo Staphylococcus
aureus, che possono crescere anche con livelli di attività dell’acqua piuttosto bassi;
vi sono inoltre i trattamenti volti alla modifica dell’acidità dell’alimento, per creare
livelli di pH sfavorevoli per i microorganismi (acidificazione, fermentazione, etc.), i
trattamenti basati sul controllo del potenziale di ossido-riduzione, soprattutto tramite
la limitazione della disponibilità di ossigeno (confezionamento sottovuoto,
confezionamento in atmosfera modificata con gas inerti) e quelli che prevedono
l’utilizzo di agenti chimici antimicrobici (sali conservanti, batteriostatici, acidi
organici, etc.) (Ramaswamy e Singh, 1997).
Tra i trattamenti il cui scopo è prevenire la contaminazione (o la
ricontaminazione) rientrano il confezionamento, nonché l’applicazione delle corrette
norme igieniche nella progettazione e nel mantenimento di strutture, linee di
lavorazione e apparecchiature. Vi possono essere, infine, trattamenti combinati,
come per esempio la pastorizzazione congiunta con il confezionamento asettico, la
salatura, l’affumicatura, e molti altri.
1.4 L’irradiazione e l’utilizzo delle microonde in campo alimentare
Nel paragrafo 1.3 si è solo accennato all’esistenza di trattamenti basati in
qualche modo sull’irradiazione; a questo punto pare necessario, in una trattazione
che non può essere, per ovvi motivi di spazio e di coerenza con gli obiettivi stessi
del lavoro di tesi, esaustiva di tutte le tecnologie applicate nell’industria alimentare,
dedicare loro uno spazio esclusivo che aiuti il lettore a focalizzare la sua attenzione
su ciò a cui maggiormente si coniuga il lavoro sperimentale che è stato svolto.
Le tecnologie che prevedono l’irradiazione degli alimenti, richiamando la
“classificazione” che è stata fatta nel paragrafo 1.3, possono essere inserite tra quelle
che hanno come scopo la riduzione del rischio microbiologico, andando ad incidere
(limitandola) sulla velocità di crescita microbica e sulla produzione di tossine; altri

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 28 -
obiettivi dell’irradiazione possono essere la distruzione o l’inattivazione di insetti,
oppure la prevenzione di alcune tipologie di modificazione dell’alimento (per
esempio l’eccessiva maturazione di alcuni tipi di frutta o la germogliazione di bulbi
e tuberi).
Se l’interesse verso l’irradiazione (con radiazioni ionizzanti) degli alimenti è
nato già nel 1921 (Josephson, 1983), come riportato da Fellows (2000), le radiazioni
attualmente più comunemente utilizzate sono i raggi gamma (capaci di una maggiore
profondità di penetrazione) e i raggi X (con profondità di penetrazione inferiori); il
dosaggio può essere:
• basso (fino a 1 kJ/kg): la radiazione in questo caso è utilizzata per la
disinfestazione da insetti, come antigermogliante o come ritardante dei
processi fisiologici (maturazione) della frutta;
• medio (tra 1 e 10 kJ/kg): questi dosaggi sono utilizzati per aumentare
la vita commerciale di alimenti facilmente deperibili come prodotti
ittici o alcuni tipi di frutta, per eliminare microorganismi patogeni da
pesce e carni e per migliorare le proprietà di alcuni alimenti (per
esempio nella preparazione di vegetali disidratati e succhi);
• alto (tra 10 e 50 kJ/kg): in quest’ultimo caso si ottiene la
sterilizzazione industriale di carni, prodotti ittici e altro, spesso in
combinazione con trattamenti termici leggeri.
Di norma, le dosi necessarie aumentano al decrescere delle dimensioni degli
organismi da eliminare, nell’ordine: parassiti, batteri Gram-negativi, batteri Gram-
positivi e muffe, spore e lieviti, virus; l’irradiazione non ha effetti su tossine
batteriche e micotossine. Organismi quali l’Agenzia Internazionale per l’Energia
Atomica (IAEA), la FAO e l’OMS hanno congiuntamente ritenuto (Organizzazione
Mondiale della Sanità, 1977; 1981) che dosi fino a 10 kJ/kg provochino alterazioni
minimali su macronutrienti e micronutrienti, o comunque paragonabili a quelle
provocate dai trattamenti termici.

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 29 -
I principali vantaggi dell’irradiazione degli alimenti, secondo Wilkinson e Gould
(1996), sono:
• minime variazioni organolettiche per via del minimo riscaldamento
indotto negli alimenti trattati;
• peggioramento delle caratteristiche nutrizionali paragonabile a quello
indotto dalle altre tecniche di conservazione;
• possibilità di trattare anche alimenti già confezionati o congelati;
• possibilità di preservare alimenti freschi dal deterioramento senza
l’uso di conservanti chimici;
• bassi costi energetici e operativi.
Dall’altra parte vi sono gli svantaggi messi in luce da Webb e Lang (1987) e da Welt
(1985), che sembrano essere il maggiore ostacolo alla diffusione di tali trattamenti:
• possibilità di “recuperare” alimenti con cariche batteriche troppo
elevate e dunque diversamente invendibili;
• impossibilità di distinguere l’insalubrità di un alimento nel caso in cui
vengano eliminati selettivamente i microorganismi che causano il
deterioramento apparente e non vengano eliminati i batteri patogeni;
• rischi derivanti dalla possibilità che vengano distrutti i batteri
produttori di tossine solo dopo che la contaminazione dell’alimento da
parte delle tossine ha avuto luogo;
• possibile sviluppo, da parte dei microorganismi, di meccanismi di
resistenza alla radiazione;
• perdita di valore nutrizionale;
• sostanziale inadeguatezza delle procedure analitiche per il
riconoscimento degli alimenti trattati (con possibilità di frodi);

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 30 -
• diffidenza da parte del consumatore dovuta a timori circa la
radioattività indotta e al pregiudizio nei confronti dell’industria
nucleare.
A proposito di quest’ultimo punto, Gaunt (1986) riporta come, nel caso dei raggi
gamma, l’energia emessa è assolutamente insufficiente per indurre radioattività negli
alimenti, mentre nel caso dei raggi X vi può essere una radioattività indotta, ma
comunque a livelli insignificanti (nella peggiore delle ipotesi non superiore al 2%
del livello di massima accettabilità). Per quanto attiene invece le procedure
analitiche per l’identificazione degli alimenti trattati, Stevenson (1994) e Delincée
(1998) passano in rassegna le metodologie a disposizione, basate essenzialmente su
analisi indirette di diversi effetti fisici, chimici e biologici indotti dalla radiazione.
Tauxe (2001) è sostanzialmente ottimista sulla futura diffusione dell’uso delle
radiazioni ionizzanti in campo alimentare in quanto i vantaggi sarebbero di gran
lunga maggiori rispetto agli svantaggi, ai quali peraltro sarebbe possibile trovare dei
rimedi efficaci.
È ancora Fellows (2000) ad indicare quale sia il meccanismo di azione dei raggi
X e dei raggi gamma, che si distinguono da altre forme di radiazione per via della
loro capacità ionizzante: i prodotti della ionizzazione, che possono essere sia
elettricamente carichi (ioni) che neutri (radicali liberi), reagiscono ulteriormente
provocando nel materiale irradiato delle modificazioni che vanno sotto il nome di
“radiolisi”; è quest’ultima il fenomeno responsabile della distruzione di
microorganismi, insetti e parassiti durante l’irradiazione dell’alimento che li
contiene. Inoltre, nei cibi ad elevato contenuto d’acqua, questa viene ionizzata, e si
producono poi idrogeno, perossido di idrogeno e i radicali ⋅H , ⋅OH e ⋅2HO ;
questi ultimi hanno una vita molto breve (inferiore a 10-5 s), ma sufficiente per
distruggere le cellule batteriche. L’azione distruttiva esercitata sui microorganismi si
manifesta attraverso modificazioni indotte a livello della membrana cellulare,
dell’attività enzimatica metabolica e soprattutto degli acidi nucleici. Componenti
liposolubili e acidi grassi essenziali con l’irradiazione vengono spesso perduti, e

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 31 -
alcuni alimenti, come per esempio i latticini, non sono idonei ad essere sottoposti a
questi trattamenti perche inducono lo sviluppo di un aroma rancido; la presenza di
ossigeno accelera questo processo, motivo per cui, in genere, l’irradiazione delle
carni avviene sottovuoto. Josephson et al. (1975) sostengono che, ai livelli di
dosaggio utilizzati commercialmente, la radiazione ha scarsissimi effetti sulla
digeribilità delle proteine e sulla composizione degli amminoacidi; Kraybill (1982),
evidenziando un certo disaccordo tra gli studiosi circa i danni nutrizionali dovuti
all’irradiazione, ha fornito un quadro dettagliato di tali aspetti.
Andando ad esaminare le tipologie di radiazioni non ionizzanti utilizzate
nell’industria alimentare, vi sono anche i raggi ultravioletti (radiazioni UV) e le
microonde. Le radiazioni UV, caratterizzate da una piccola profondità di
penetrazione (se paragonata a quella delle radiazioni ionizzanti), possono essere utili
per la distruzione di microorganismi presenti nell’ambiente, su superfici o in strati
liquidi sottili, ma hanno scarsa efficacia, per esempio, contro lieviti e spore, sia
batteriche che di muffe (Fellows, 2000). Per quanto riguarda l’utilizzo delle
microonde, invece, si tratta di una tecnologia di riscaldamento dielettrico, attraverso
la quale si ottiene il riscaldamento del materiale per il rapidissimo movimento
vibrazionale delle molecole dipolari (in particolare di quelle dell’acqua, ma sono
interessati anche lipidi, proteine e zuccheri) indotto dal campo magnetico alternato;
se le dimensioni del materiale sono modeste, il riscaldamento ottenuto è
sufficientemente uniforme (Buchachenko e Frankevich, 1993). Quindi, nelle
applicazioni industriali, le microonde presentano, rispetto ai sistemi tradizionali, il
vantaggio di consentire un riscaldamento volumetrico, in quanto si basano sulla
veicolazione, al materiale, di energia e non di calore: il riscaldamento che ne risulta
è più rapido e selettivo, ed è relativamente più rapido per quei materiali che non
sono buoni conduttori di calore; occorre più attenzione se la struttura dell’alimento è
eterogenea, in quanto la velocità di riscaldamento può assumere valori differenti tra
le diverse parti e possono così formarsi aree con temperature anche notevolmente
diverse tra loro. Le microonde, nei processi alimentari, sono utilizzate per operazioni

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 32 -
quali cottura, essiccamento, pastorizzazione e sterilizzazione. L’utilizzo delle
microonde comporta un maggior consumo di energia elettrica rispetto alle tecniche
di riscaldamento convenzionali, ma allo stesso tempo consente un minore utilizzo di
combustibili, e costituisce un risparmio in termini di energia primaria; la migliore
efficienza energetica si avrebbe probabilmente con l’accoppiamento della tecnologia
a microonde con le tecnologie tradizionali (Fellows, 2000).
1.5 Prospettive e vantaggi dell’utilizzo di microonde a bassa potenza
È opportuno, a questo punto, illustrare quali siano le motivazioni che stanno alla
base della scelta delle microonde a bassa potenza: l’elemento essenziale è
sicuramente che solo l’utilizzo di basse potenze può consentire la discrimiazione tra
effetti termici ed effetti non termici delle microonde. E l’eventuale esistenza di
effetti non termici, oltre ad essere un fatto di sicura e notevole rilevanza da un punto
di vista prettamente scientifico, potrebbe aprire la strada alla messa a punto di
trattamenti alimentari innovativi e non più basati sull’innalzamento termico
dell’alimento, con indubbi vantaggi in termini sia qualitativi che economici. In
prima istanza, infatti, operando a temperature notevolmente più basse rispetto a
quelle dei trattamenti termici convenzionali (ai quali si contrapporrebbe un
trattamento fondato sugli effetti non termici della radiazione sui microorganismi
patogeni), sarebbe possibile preservare ulteriormente le proprietà nutrizionali e
organolettiche degli alimenti trattati mantenendoli, da questo punto di vista,
maggiormente simili ai corrispondenti prodotti freschi: se anche con l’utilizzo di
radiazioni ionizzanti a bassi dosaggi si possono produrre gli stessi effetti della
pastorizzazione ottenuta mediante riscaldamento convenzionale, ma con minori
effetti su proprietà organolettiche (Steele e Engel, 1992), si può a buona ragione
ritenere che tali proprietà siano ancor meno intaccate dall’utilizzo di radiazioni non
ionizzanti. E non sarebbe affatto trascurabile l’impatto economico che avrebbe, sul
settore alimentare, l’introduzione di trattamenti non termici che, per loro stessa

CAPITOLO 1 – LE ORIGINI E I PRESUPPOSTI DEL LAVORO
- 33 -
natura, comporterebbero notevoli risparmi energetici. Se dunque, in linea di
massima, ad un aumento della qualità dei cibi (ma la regola vale anche per qualsiasi
altro prodotto industriale) corrisponde un aumento dei costi, in questo caso tale
assunto sembrerebbe violato da una tipologia di trattamento più economica e
caratterizzata da risultati qualitativamente migliori; ciò aprirebbe probabilmente la
strada verso trattamenti a freddo “paralleli” a quelli termici, e si potrebbero forse
utilizzare realmente diciture come “pastorizzazione a freddo”, che Crawford e Ruff
(1996), già più di dieci anni fa, proposero di sostituire alla dicitura di “irradiazione”.
Non è da escludere, tuttavia, anche il possibile sfruttamento sinergico di effetti
termici e non termici; fenomeni di sinergia tra effetti termici e altri effetti (di natura
non meglio precisata) sono infatti già presenti negli studi di letteratura: a tal
proposito è assai interessante (per quanto riguarda, in questo caso, l’efficacia del
processo di pastorizzazione del latte) il lavoro di Kamau et al. (1990), che hanno
sperimentato la distruzione per via termica delle specie Listeria monocytogenes e
Staphylococcus aureus immediatamente successiva all’attivazione della
lattoperossidasi (uno degli enzimi più abbondanti del latte), riscontrando una
sinergia tra le due azioni battericide e un conseguente aumento del margine di
sicurezza durante il processo di pastorizzazione rispetto ai due patogeni presenti nel
latte.


CAPITOLO 2


CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 37 -
2.1 La radiazione elettromagnetica e le microonde
Per “radiazione elettromagnetica” (Mazzoldi et al., 2002; Halliday et al., 2006)
si intende la propagazione nello spazio dell’energia (detta energia elettromagnetica)
associata a campi elettrici e magnetici variabili nel tempo, generati da cariche e
correnti oscillanti, intercorrelati tra loro secondo le equazioni di Maxwell. Le
radiazioni elettromagnetiche sono caratterizzate da tre principali grandezze fisiche
tra loro correlate, che sono:
• lunghezza d’onda (comunemente indicata col simbolo λ, è la distanza
tra punti equivalenti di una forma d’onda, per esempio tra due
massimi);
• frequenza (comunemente indicata con ν, è il numero di cicli della
forma d’onda che si ripetono nell’unità di tempo, ed è il reciproco del
periodo, che è a sua volta l’intervallo di tempo che intercorre tra gli
istanti corrispondenti a due punti equivalenti della forma d’onda);
• energia fotonica (indicata con E), ovvero l’energia trasmessa dalla
radiazione tra due punti nello spazio nella direzione di propagazione.
Lunghezza d’onda e frequenza sono legate tra loro dalla relazione:
cνλ =⋅ (2.1)
dove c è una costante fisica che rappresenta la velocità di propagazione delle onde
elettromagnetiche (ovvero della luce) nel vuoto, ed è pari a 299.792,458 km/s;
l’energia fotonica è invece legata alla frequenza dalla relazione:
νhnE ⋅⋅= (2.2)
dove h è la costante di Planck ed n è il numero quantico principale, ed è perciò un
intero positivo.

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 38 -
Le frequenze possibili delle onde elettromagnetiche coprono un vastissimo
intervallo di valori, detto spettro elettromagnetico; per motivi di praticità,
quest’ultimo è convenzionalmente suddiviso in regioni che assumono
denominazioni differenti a seconda dell’intervallo di frequenze (o di lunghezze
d’onda) al quale sono associate; vi sono delle distinzioni anche in relazione
all’energia associata e al tipo di utilizzo che ne viene fatto. Sono individuate le
seguenti tipologie di radiazione con frequenza crescente (e lunghezza d’onda
decrescente, in quanto le due grandezze sono inversamente proporzionali):
• onde radio ( MHz 300ν ≤ ; m 1λ ≥ );
• microonde ( GHz 300MHz 300ν ÷= ; m 1mm 1λ ÷= );
• radiazione infrarossa ( THz 284GHz 300ν ÷= ; mm 1nm 007λ ÷= );
• luce visibile ( THz 749THz 284ν ÷= ; nm 700nm 004λ ÷= );
• raggi ultravioletti ( PHz 30THz 749ν ÷= ; nm 400nm 10λ ÷= );
• raggi X ( EHz 300PHz 30ν ÷= ; nm 10pm 1λ ÷= );
• raggi gamma ( EHz 300ν ≥ ; pm 1λ ≤ ).
Restringendo lo spazio di analisi al campo di nostro interesse, le microonde sono
chiamate così in quanto effettivamente molto corte, e la loro distinzione rispetto alle
onde radio, che peraltro non è poi così netta, deriva essenzialmente dalle diverse
applicazioni tecnologiche; le microonde sono in realtà comprese nelle parti UHF
(“Ultra High Frequency”, con GHz 3MHz 300ν ÷= ), SHF (“Super High
Frequency”, con GHz 30GHz 3ν ÷= ) e EHF (“Extremely High Frequency”, con
GHz 300GHz 30ν ÷= ) dello spettro radio; il limite superiore di 300 GHz è invece
legato a motivazioni fisiche, in quanto oltre tale valore di frequenza l’assorbimento
della radiazione elettromagnetica da parte dell’atmosfera terrestre diventa
notevolmente intenso, al punto che questa può essere considerata opaca a quelle
frequenze.

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 39 -
La maggior parte delle applicazioni delle microonde ricadono nel campo
compreso tra 1 e 80 GHz. Tra i principali utilizzi vi sono:
• ponti radio, ovvero trasmissione di segnali analogici (come ad
esempio quelli televisivi) o digitali (con velocità di trasmissione fino a
qualche centinaia di megabit al secondo) tra antenne paraboliche
terrestri, su distanze nell’ordine delle decine o centinaia di chilometri
e all’interno di bande di frequenza specificamente fissate dagli
organismi regolatori nazionali e internazionali nell’intervallo tra 2 e
80 GHz, con potenze per ogni canale portante comprese tra una
frazione di watt a qualche watt al massimo;
• comunicazione, alla frequenza di 1,8 GHz, tra i telefoni cellulari GSM
e le stazioni radio base;
• forni a microonde, che utilizzano un generatore a magnetron di
microonde alla frequenza di 2,45 GHz per la cottura o il riscaldamento
dei cibi;
• comunicazione con i satelliti, in quanto a differenza delle onde radio
di frequenza inferiore non subiscono interferenze durante
l’attraversamento dell’atmosfera terrestre e offrono, inoltre, una
maggiore larghezza di banda, con conseguente maggiore velocità di
trasmissione dei dati;
• radar, che utilizzano le microonde per il rilevamento a distanza della
presenza e del movimento di oggetti;
• protocolli di comunicazione senza fili, quali bluetooth e IEEE 802.11,
che utilizza microonde a 2,4 GHz di frequenza nelle varianti g e b, e a
5 GHz nella variante a;
• servizi di accesso ad Internet a lunga distanza (anche 25 km) che si
stanno ultimamente diffondendo in alcuni paesi e che operano a
frequenze comprese tra 3,5 e 4 GHz;

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 40 -
• servizi di diffusione televisiva, di accesso ad Internet e di telefonia su
cavo coassiale che utilizzano microonde a bassa frequenza;
• utilizzi più di carattere scientifico e futuristico sono il maser (un
dispositivo simile al laser ma operante nello spettro delle microonde),
le cavità risonanti utilizzate negli acceleratori di particelle e il
trasferimento di energia a distanza, che oggi trova applicazione in
armamenti di nuova generazione, e che già dagli anni ’70 del secolo
scorso la NASA studiò pensando alla possibilità di trasferire sulla
Terra l’energia prodotta su satelliti dotati di ampi pannelli solari (vi
sono tutt’ora progetti di centrali solari orbitanti).
Le microonde (Mazzoldi et al., 2002; Halliday et al., 2006) possono essere
generate mediante due diversi tipi di tecnologia: dispositivi a stato solido e
dispositivi con tubi a vuoto. I primi si basano sull’utilizzo di semiconduttori (silicio
o arseniuro di gallio) e sono i transistor ad effetto campo (FET), i transistor a
giunzione bipolare (BJT), i diodi Gunn e IMPATT, e i dispositivi integrati a
microonde (MMIC, acronimo di “monolithic microwave integrated circuit”),
realizzati a partire da wafer di arseniuro di gallio, mentre versioni più particolari di
transistor vengono utilizzate per le frequenze più alte; i secondi funzionano grazie al
movimento balistico degli elettroni nel vuoto sotto il controllo di opportuni campi
elettrici o magnetici, e sono il magnetron, il klystron, il travelling wave tube (TWT)
e il gyrotron.
2.2 L’interazione tra radiazioni elettromagnetiche e reazioni chimiche
Qualsiasi processo chimico, la sua fattibilità e i risultati da esso ottenibili non
possono essere studiati se non in stretto legame con le implicazioni di carattere
energetico associate al processo stesso: la quantità di energia richiesta, le modalità di
veicolazione di tale energia verso le specie reagenti, l’efficienza con la quale

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 41 -
l’energia viene utilizzata nel processo. Una reazione chimica consiste infatti in un
processo fisico di ridisposizione di atomi ed elettroni avente come risultato la
trasformazione di molecole. Vale a questo punto parafrasare un concetto espresso
dal famoso fisico Feynman (1992), secondo il quale la chimica sarebbe la branca più
complessa della fisica, irraggiungibile da parte degli stessi fisici, in quanto sarebbero
troppe le interazioni, dipendenti dal tempo, tra elettroni ed elettroni e tra elettroni e
nuclei, che hanno luogo durante un evento chimico.
La trasformazione chimica di sostanze e materiali è un processo costoso in
termini energetici e dunque tutti i principi del controllo e della promozione delle
reazioni chimiche hanno origine dal dogma dell’energia, ovvero dal concetto
secondo il quale il disavanzo energetico può essere controbilanciato o riducendo i
costi energetici oppure assicurando efficienti modalità di trasmissione dell’energia
alle molecole reagenti. Tra l’ampia gamma di metodi utilizzabili per promuovere e
controllare le reazioni chimiche, basati sul principio sopra esposto, l’azione dei
campi magnetici è sempre stata la possibilità forse meno considerata (Sterna et al.,
1980), e ciò si spiega con l’ovvia motivazione che l’energia magnetica che può
essere fornita a specie reagenti, molecole, atomi, ioni, radicali, è di entità in fin dei
conti trascurabile persino con campi magnetici di intensità elevatissima; può essere
anche milioni di volte più piccola dell’energia termica, e centinaia di milioni di volte
più piccola di quella effettivamente necessaria per la reazione chimica
(Buchachenko, 1974). Tuttavia, oltre alla questione energetica, vi è una proprietà
fisica di primaria importanza nella chimica, il momento angolare (spin) degli
elettroni e dei nuclei dei reagenti. Infatti, tutte le reazioni chimiche sono selettive
rispetto allo spin, cioè sono possibili solo in alcune condizioni di spin e non lo sono
affatto in condizioni di spin differenti: ciò implica che reagenti chimicamente
identici possiedano reattività completamente differenti a seconda delle condizioni
dello spin, e che se le condizioni dello spin non sono favorevoli, la reazione non
possa avvenire neanche se sarebbe consentita dalle condizioni energetiche
(Carrington e McLachlan, 1967; Salikhov et al., 1984; Atkins e Friedman, 2004). Le

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 42 -
uniche interazioni che sono in grado di modificare lo spin dei reagenti e mutare uno
stato non reattivo in uno reattivo (e viceversa) sono proprio quelle magnetiche, che
non apportano nessun contributo in termini energetici, ma che possono invece
modificare lo spin (Geacintov et al., 1969; 1970). Nelle molecole diamagnetiche gli
spin elettronici sono orientanti secondo versi opposti, ed è possibile dunque un solo
stato, mentre nei radicali, che possiedono un elettrone spaiato, gli stati possibili sono
due (doppietto). Le specie radicaliche, dunque, per via dell’elettrone spaiato, sono le
più soggette al fenomeno (Deutch, 1972), insieme alle specie in possesso di due
elettroni spaiati, e a ioni inorganici paramagnetici quali per esempio Fe+++, Fe++ e
Cu++ (Harberkorn, 1977). Un atomo di ossigeno possiede due elettroni spaiati con
spin concordi, per cui a livello molecolare (O2) sono possibili tre diverse
combinazioni (tripletto), così come accade per altre molecole, e la complessità può
aumentare a quattro combinazioni (quartetto), cinque (quintetto), e così via
(Schulten e Wolynes, 1978). Le transizioni tra stati di spin elettronico sono
responsabili della Risonanza di Spin Elettronico (ESR), mentre le transizioni tra stati
di spin nucleare sono alla base della Risonanza Magnetica Nucleare (NMR); queste
transizioni sono indotte da un campo magnetico di microonde alla frequenza di
precessione elettronica o nucleare, e sono accompagnate da modificazioni nello spin;
il campo magnetico può essere imposto dall’esterno, come accade per la ESR e per
la NMR, oppure esso può essere generato dalle vibrazioni reticolari stocastiche che
hanno luogo a livello molecolare, e che producono campi magnetici fluttuanti e
rumorosi di differenti frequenze e ampiezze; la componente spettrale di questo
rumore con frequenza pari a quella della precessione elettronica o nucleare induce le
transizioni tra differenti stati di spin (Evans et al., 1973; Zientara e Freed, 1979).
Il principio fondamentale è che, in una reazione chimica elementare, lo spin
totale si conserva, cioè sia gli spin che le loro proiezioni non cambiano; per questo
motivo si può dire che le reazioni chimiche sono selettive rispetto allo spin, e sono
possibili solo se lo spin totale dei prodotti è uguale a quello dei reagenti, mentre non
lo sono affatto in caso contrario (Steiner e Ulrich, 1989).

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 43 -
Anche reazioni chimiche di elevata importanza biologica sono selettive rispetto
allo spin, e perciò magnetosensibili; un esempio è fornito dalle reazioni di
fotosintesi, che manifestano effetti indotti dalla componente magnetica delle
microonde (Buchachenko e Frankevich, 1993). A livello di biologia cellulare e
chimica degli enzimi vi sono numerose osservazioni di effetti indotti dal campo
magnetico, tra cui (solo per citarne alcune) quelle di Atkins e Lambert (1975),
Barnothy J. M. (1963), Barnothy J. M. et al. (1956), Barnothy M. F. (1964),
Barnothy M. F. e Barnothy J. M. (1958), Barnothy M. F. e Sumegi (1969), Grissom
(1992).
Da un punto di vista generale, sono due i requisiti che devono essere soddisfatti
affinché lo spin totale della coppia di reagenti sia adeguato ai prodotti della reazione:
da una parte devono sussistere interazioni magnetiche tra la coppia che possano
influire sullo spin, dall’altra l’accoppiamento deve durare un tempo sufficiente
perché le interazioni magnetiche possano modificare lo stato dello spin (Noyes,
1954); in via del tutto approssimata si può affermare che tale tempo è inversamente
proporzionale all’energia magnetica necessaria per la conversione dello spin (Razi
Naqvi et al., 1980). L’energia magnetica necessaria alla conversione dello spin può
essere anche da quattro a nove ordini di grandezza più piccola dell’energia dei
legami chimici, tuttavia la sua peculiarità di poter governare lo spin la rendono
indispensabile alla reazione (Mozumder, 1968).
La selettività delle reazioni chimiche rispetto allo spin e le dinamiche relative
allo spin sono alla base delle due modalità di generazione degli effetti magnetici, che
sono gli effetti connaturati indotti dalla reazione stessa e quelli indotti dalle onde
elettromagnetiche (Merrifield et al., 1969; Johnson e Merrifield, 1970).
L’influenza delle onde elettromagnetiche sull’evoluzione delle reazioni
chimiche è basata sull’interazione tra il campo magnetico dell’onda elettromagnetica
e i momenti magnetici elementari delle particelle che prendono parte alla reazione;
l’interazione può in linea di principio mutare lo stato di tali particelle, influenzando
così la probabilità che abbia luogo la reazione tra di esse (Boiden Pedersen e Freed,

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 44 -
1974; den Hollander, 1975). Secondo quest’ottica, l’influenza della potenza
elettromagnetica assorbita sarebbe simile a quella fotochimica prodotta dalla luce
(Buchachenko et al., 1987).
2.3 Gli effetti biologici della radiazione elettromagnetica
Lo studio delle interazioni tra onde elettromagnetiche non ionizzanti e sistemi
biologici complessi, a partire dai semplici aggregati molecolari fino alle cellule, agli
organi e agli interi organismi costituisce una parte assai rilevante della più generale
ricerca sulle interazioni tra campi elettromagnetici e materia. Oltre a questo,
importante stimolo è stata ed è l’opinione pubblica nei confronti dei possibili effetti
nocivi che i campi elettromagnetici potrebbero avere sulla salute, in particolare per
quanto riguarda le linee di trasmissione di corrente alternata in alta tensione, gli
schermi video, le procedure di diagnostica per immagini quale la Risonanza
Magnetica Nucleare, etc.. Particolare interesse è scaturito dai riscontri
epidemiologici che hanno messo in relazione l’esposizione di esseri umani a campi
elettromagnetici deboli e il rischio di sviluppare patologie. Negli ultimi decenni del
secolo scorso sono andati aumentando gli studi sperimentali sugli effetti biologici e
medici dei campi elettromagnetici, e sono stati di pari passo proposti ipotetici
meccanismi che potessero essere coinvolti nella generazione di quegli effetti
(Buchachenko e Frankevich, 1993).
In linea di principio entrambe le componenti del campo elettromagnetico,
ovvero quella elettrica e quella magnetica, possono essere responsabili di effetti
biologici. La componente elettrica interagisce con la parte dielettrica del sistema
biologico preso in considerazione, inducendo perdite dielettriche e di conseguenza
un incremento della temperatura che a sua volta crea modificazioni nei processi
biologici (attività enzimatica, trasduzione dei segnali attraverso la membrana
cellulare, sintesi di DNA e RNA, regolazione dell’omeostasi intracellulare degli ioni

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 45 -
Ca++, etc.), e di conseguenza effetti macroscopici che a questi sono legati, quali
velocità di crescita cellulare e risposte biologiche dell’organismo. La componente
magnetica, invece, interagirebbe con gli intermedi paramagnetici (quali paia di
radicali o paia di radicali ionici), producendo meccanismi mediati attraverso lo spin
(Salikhov et al., 1984; Steiner e Ulrich, 1989; Buchachenko e Frankevich, 1993).
Entrambe le componenti offrono insomma, almeno da un punto di vista teorico,
meccanismi che, anche in presenza di campi elettromagnetici relativamente deboli,
sono in grado di influenzare funzioni biologiche. Tuttavia parrebbe che nei processi
biochimici non vi siano paia di radicali o paia di radicali ionici dotate di
caratteristiche dinamiche e magnetiche tali da produrre effetti significativi e dunque
influenzabili e chimicamente modificabili da parte di un campo elettromagnetico; e
anche se esistono paia dotate di queste caratteristiche come intermedi, questi hanno
probabilmente tempi di vita o di rilassamento troppo brevi (per esempio, paia che
includono gli ioni Fe++ o Fe+++) perché il campo elettromagnetico possa interferire
con le dinamiche dello spin (Harberkorn, 1977; Buchachenko e Frankevich, 1993).
Quanto detto pocanzi porta alla conclusione che il principale interesse sia costituito
dalla componente elettrica del campo. Di importanza basilare è l’eterogeneità
microstrutturale che caratterizza i sistemi biologici a livello di cellule, organi e
organismi, che ha almeno due importanti conseguenze. La prima è che tale
eterogeneità produce una distribuzione non omogenea del campo elettromagnetico
nei vari elementi strutturali, per cui vi è la possibilità che l’ampiezza locale del
campo sia molto maggiore del suo valore medio, anche di diversi ordini di
grandezza; la seconda è che la non equivalenza degli elementi strutturali di cellule
ed organi rispetto alle proprietà dielettriche e di conducibilità si traduce in un
assorbimento del campo fortemente non omogeneo, e quindi anche la temperatura e
le correnti ioniche locali possono superare, anche di parecchio, i loro rispettivi valori
medi (Grissom, 1992). Come risultato di ciò, pur trattandosi di fenomeni locali e
aventi perciò luogo su una scala (microscopica) non rilevabile con le metodologie
standard di misura, essi possono tuttavia avere un peso considerevole sulle funzioni
cellulari; tali effetti possono poi ripercuotersi sulla stessa attività cellulare, sulle

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 46 -
membrane e, risalendo la scala di grandezza, sull’intero organismo, concetto che
pare in accordo con la maggior parte delle determinazioni sperimentali
(Buchachenko e Frankevich, 1993); per esempio, Akoev (1986) ha riscontrato effetti
biologici indotti dal campo elettromagnetico solo su sistemi caratterizzati da
eterogeneità a livello microscopico e che invece non si manifestavano in sistemi con
la stessa natura chimica ma strutturalmente omogenei (Sarvarov e Salikhov, 1976).
2.4 Effetti termici versus effetti non termici delle microonde
L’identificazione e la valutazione degli effetti biologici provocati dalle
microonde è materia complessa e controversa. Per via forse dell’effettiva scarsità di
informazioni sui meccanismi di interazione tra microonde e sistemi biologici, si è
andata costituendo, in parte della comunità scientifica, una forte opinione che i
campi di microonde sono incapaci di generare effetti biologici diversi dal
riscaldamento (Michaelson, 1991). Sono relativamente recente è l’interesse per gli
effetti non termici delle microonde sui tessuti (D’Andrea et al., 1979; Adey, 1981;
D’Andrea et al., 1986a; D’Andrea et al., 1986b), ma sono ormai in tanti ad avere
verificato l’esistenza di tali effetti. Una delle possibili spiegazioni di carattere
generale degli effetti biologici delle microonde è che queste ultime sono un
fenomeno praticamente assente nell’ambiente elettromagnetico naturale e ciò,
secondo Golant (1989) e Betzky (1992), potrebbe avere almeno due importanti
conseguenze: in primo luogo gli organismi viventi, durante il corso dell’evoluzione,
potrebbero non avere sviluppato un adattamento alle microonde; secondariamente,
alcune specifiche caratteristiche delle microonde e l’assenza di rumore ambientale
esterno potrebbero avere reso tale banda elettromagnetica adatta per la
comunicazione sia intracellulare che intercellulare; tali argomentazioni, per quanto
forse non adeguatamente dimostrate, sono tuttavia utilizzabili come possibile
spiegazione dell’elevata sensibilità che gli organismi sembrano mostrare nei
confronti delle microonde; e infatti, secondo gli autori, sono numerosi gli effetti

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 47 -
biologici delle microonde riportati dalla letteratura scientifica, molti dei quali
difficilmente conciliabili con la profondità di penetrazione (meno di un millimetro)
nei tessuti biologici. Sono diversi i modelli teorici sviluppati per spiegare le
peculiarità ed i meccanismi primari dell’azione biologica delle microonde (Fröhlich,
1980; 1988; Golant, 1989; Grundler e Kaiser, 1992; Belyaev et al., 1993a; Kaiser,
1995).
Vi sono prove che le microonde provochino differenti effetti biologici a seconda
della forza del campo, della frequenza, della forma d’onda, della modulazione e
della durata dell’esposizione (Rai et al., 1994a; 1994b). Alcuni di questi effetti erano
stati in precedenza principalmente attribuiti al riscaldamento apportato dalle
microonde (Gandhi, 1987), ma studi più recenti hanno mostrato o suggerito che vi
sono o vi possono essere effetti non termici, in termini di energia richiesta per
diversi tipi di trasformazioni molecolari e altre alterazioni, indotti dalle microonde.
Una delle maggiori difficoltà nella distinzione tra effetti termici ed effetti non
termici è quella di mantenere la temperatura costante durante l’irradiazione delle
microonde; il problema è stato affrontato da Kozempel et al. (1997), che hanno
messo a punto un processo discontinuo mediante il quale è stato possibile ridurre la
numerosità di popolazioni di Pediococcus sp. in soluzioni zuccherine mantenendo
una temperatura (nominale) di 35°C per tempi inferiori ai nove minuti per mezzo di
un efficiente sistema di rimozione del calore. Un altro metodo, a tal proposito, è
stato messo a punto da Sato (2001), che nello stesso lavoro, a 45, 47,5 e 50°C (ma
non a 35) ha osservato un incremento della velocità di morte dei batteri di
Escherichia coli esposti alle microonde rispetto alla velocità alle stesse temperature
ma senza microonde, proponendo come meccanismo controllante modificazioni
nella struttura secondaria e terziaria delle proteine dei microorganismi, indotte dalle
rapide oscillazioni del campo elettrico. Geveke et al. (2002) hanno sviluppato un
processo sperimentale continuo a microonde allo scopo di isolare gli effetti non
termici da quelli termici durante una ipotetica operazione di pastorizzazione non
termica di liquidi; il processo si basa sul rapido apporto di energia termica al sistema

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 48 -
alimentare mediante l’utilizzo delle microonde, combinato con un altrettanto rapido
sistema di rimozione mediante scambiatore termico; gli autori hanno riscontrato che,
alla temperatura di 35°C e con tempi di esposizione tra 3 e 8 minuti, non vi era una
significativa riduzione dei lieviti né della carica batterica se le microonde non
venivano abbinate a calore, a condizioni di pH sfavorevoli per i microorganismi o ad
antimicrobici. Culkin e Fung (1975) hanno constatato che batteri di Escherichia coli
e di Salmonella typhimurium venivano distrutti se esposti a microonde alla
frequenza di 915 MHz, e che la loro morte avveniva a temperature inferiori ed in
tempi minori rispetto ai metodi di riscaldamento convenzionali; essi hanno inoltre
notato effetti differenti a seconda dell’intensità del campo; i risultati ottenuti li
hanno condotti ad ipotizzare meccanismi sottesi ai fenomeni osservati che andassero
oltre il normale effetto termico associato alle microonde. Singh et al. (1994) hanno
studiato l’azione di microonde continue e modulate sull’alga Nostoc muscorum,
riportando che gli effetti fisiologici prodotti sull’alga sono diversi in funzione della
frequenza dell’irradiamento; gli effetti mediati dalla presenza dell’acqua, secondo
gli autori, sarebbero inoltre una prova della capacità dell’acqua di “ricordare” le
caratteristiche del campo elettromagnetico applicato per un periodo di tempo più
esteso di quello della stessa applicazione. L’effetto di microonde modulate con onde
quadre di varie frequenze sul comportamento fisiologico del cianobatterio Anabaena
doliolum è stato studiato da Samarketu et al. (1996), esponendo il microorganismo
con la soluzione nutriente per un’ora a microonde di frequenza 9,575 GHz e con
densità di potenza pari a 0,658 mW/cm2; lo studio ha rivelato effetti non termici
delle microonde consistenti in modificazioni dei flussi ionici, delle velocità e/o della
direzione di reazioni biochimiche. Moore et al. (1979) hanno osservato un calo
reversibile di virulenza di cellule di Agrobacterium tumefaciens dopo l’esposizione a
microonde alla frequenza di 10 GHz e con una densità di potenza di 0,58 mW/cm2
per tempi compresi tra 30 e 120 minuti, attribuendo la causa di ciò ad un
cambiamento (temporaneo) in uno o più processi metabolici direttamente connessi
con la virulenza come effetto non termico; la virulenza tornava ai livelli originari
dopo 12 ore. Dreyfuss e Chipley (1980) hanno provato a caratterizzare alcuni degli

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 49 -
effetti prodotti dall’irradiazione con microonde su cellule di Staphylococcus aureus,
registrando variazioni nell’attività di diversi enzimi metabolici sostanzialmente
diverse da quelle normalmente legate al riscaldamento convenzionale, e deducendo
da ciò la natura non termica degli effetti riscontrati. È stato riportato da Justeen
(1979) che microonde a bassa intensità possono generare effetti comportamentali
senza modificare la temperatura interna dei soggetti esposti; sono stati inoltre
riportati numerosi effetti di natura sia fisiologica (Ray e Behari, 1990) che
biochimica (Kunjilwar e Behari, 1993) su ratti anche dopo esposizione a microonde
a bassa potenza. Banik et al. (2002a; 2002b) hanno tentato di studiare l’effetto
dell’esposizione alle microonde sulla biometanazione mediante l’utilizzo di un
ceppo batterico metanogenico (Methanosarcina barkeri DSM 804); colture
anaerobiche di questo microorganismo sono state irradiate con microonde
(frequenza compresa tra 13,5 e 36,5 GHz) per 2 ore al fine di valutarne gli effetti
sulla crescita batterica e sul processo di biometanazione; si è appurato che, alle
frequenze utilizzate, la generazione di biogas è risultata più rapida, ed è aumentato il
contenuto di metano nel biogas stesso; anche la velocità di crescita batterica è
risultata incrementata, e la maggior produzione di biogas si è avuta per 31,5 GHz di
frequenza.
I limiti di esposizione per l’uomo alle onde elettromagnetiche imposti dalle varie
legislazioni nazionali (normati in Italia dal D. M. 381/98, “Regolamento recante
norme per la determinazione dei tetti di radiofrequenza compatibili con la salute
umana”) o indicati da organismi sovranazionali tra cui in primis l’ICNIRP
(International Commission on Non-Ionizing Radiation Protection), al quale fa
riferimento la stessa Organizzazione Mondiale della Sanità, si basano
essenzialmente sul concetto che occorra stare al di sotto della soglia oltre la quale si
manifestano significativi effetti termici, e non tengono dunque conto della
possibilità che l’esposizione ai campi elettromagnetici possa causare effetti biologici
(intendendo con tale dicitura modificazioni a livello di cellula, tessuto o intero
organismo di tipo fisiologico, biochimico e comportamentale) e sulla salute umana

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 50 -
(intendendo con ciò modificazioni che siano nocive per l’organismo) di natura non
termica anche ben al di sotto di tale soglia. Certo non è semplice operare una netta
distinzione tra zone in cui si ha un tipo di effetti e zone in cui si ha l’altro tipo, infatti
il livello energetico necessario a causare effetti termici può dipendere da diversi
aspetti, quali il tipo di sistema biologico esposto (coltura cellulare, animale piccolo o
grande, corpo umano), la frequenza della radiazione, la polarizzazione del campo, il
controllo della temperatura ambiente intorno al sistema biologico. Per questo
motivo, volendo addivenire ad una definizione di effetto termico e di effetto non
termico, si può dire che per effetto termico si intende un effetto che si manifesta
quando viene assorbita una quantità di energia elettromagnetica sufficiente a
provocare, nel campione, un incremento di temperatura misurabile (dunque
dell’ordine di grandezza di almeno 0,1°C), mentre per effetto non termico può
intendersi, al contrario, qualunque effetto associato con l’assorbimento di un
quantitativo di energia inferiore a quello associabile alle normali fluttuazioni
termiche del sistema biologico esaminato.
Quando ci si riferisce alla quantità di energia assorbita da un sistema biologico,
nella letteratura si utilizza in via preferenziale il SAR, acronimo di “Specific
Absorption Rate”, che dimensionalmente corrisponde ad una potenza (assorbita) per
unità di massa di tessuto biologico esposto alla radiazione; è evidente che,
trattandosi di una grandezza che tiene conto della potenza effettivamente assorbita
dal tessuto, e che è quindi differente rispetto a quella del campo, il SAR non è
misurabile direttamente, ma deve essere ricavato per via indiretta attraverso
specifiche prove sperimentali o stimato mediante modelli matematici, come per
esempio è stato fatto da Durney et al. (1986) per diversi tipi di substrato.
Per quanto riguarda gli effetti termici, esistono in letteratura moltissimi studi e
dati che consentono di valutarli, derivati sia da prove in vitro (su sistemi cellulari o
componenti subcellulari) che in vivo (su animali esposti alla radiazione per tempi
variabili), che sono alla base delle linee guida sui livelli di esposizione tollerabili
dall’essere umano, consigliati da enti sovranazionali quali l’NCRP (U. S. National

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 51 -
Council on Radiation Protection and Measurement, 1986), l’IRPA (International
Radiation Protection Association, 1988) e l’OMS (Organizzazione Mondiale della
Sanità, 1993); si tratta, chiaramente, di prove di laboratorio in cui la causa del
riscaldamento è stata tale da poter provocare una risposta misurabile da parte del
sistema biologico in oggetto; mutazioni genetiche provocate da microonde, almeno
stando alle recensioni di Blackman (1984), Elder (1987) e Michaelson e Lin (1987),
sono state riscontrate in studi sperimentali solo a fronte di incrementi di temperatura
sostanziali. Ciò sarebbe in accordo con l’osservazione che, alla frequenza delle
microonde, l’energia fotonica è ancora molto bassa, e dunque non in grado di
provocare danni diretti al DNA; vi sono inoltre conferme di carattere sperimentale di
colture cellulari esposte a microonde che hanno visto variazioni del genoma solo
oltre una certa soglia di incremento termico (Elder e Cahill, 1984; Cleary, 1990a;
Liburdy, 1992). Nel caso di studi su animali, gli effetti osservati di radiazioni nella
frequenza delle onde radio e delle microonde sono vari e comprendono variazioni
nel sistema corporeo di termoregolazione, nelle funzioni endocrine, nel sistema
cardiovascolare, nel sistema immunitario, nell’attività del sistema nervoso e nel
comportamento (Roberts et al., 1986; Elder, 1987; Cleary, 1990b); tali
modificazioni sono quasi sempre reversibili, e cessano al cessare dell’esposizione, a
meno che la potenza assorbita non diventi abbastanza elevata (oltre 100 W/kg) da
provocare effetti irreversibili nell’animale quali problemi nello sviluppo fetale,
insorgenza di cateratta, ustioni, cauterizzazione di ferite.
2.4.1 Effetti fisico-chimici e assorbimento delle microonde
Alcuni studi hanno mostrato effetti specifici delle microonde su sistemi
biochimici ma in assenza di organismi viventi, per esempio in soluzioni di
biomolecole e persino in acqua pura. Fesenko e Gluvstein (1995) hanno analizzato
gli effetti delle microonde sulle oscillazioni periodiche della tensione durante la
scarica di un condensatore ad acqua, costituito da un campione di acqua distillata in

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 52 -
un capillare di un millimetro, esposto a microonde mediante una guida d’onda aperta
all’estremità; gli stessi hanno constatato differenze nel comportamento dell’acqua
relativamente alla scarica al variare delle condizioni di irradiamento, ipotizzando che
l’acqua fosse caratterizzata da una sorta di “memoria” a lungo termine, non
spiegabile con le conoscenze disponibili, che sottintenderebbe ad alcuni degli effetti
biologici ascritti alle microonde. Gli effetti diretti delle microonde sulle proprietà
dell’acqua pura sono stati osservati anche da Berezhinskiĭ et al. (1993) e Litvinov et
al. (1994), mediante interferometria olografica: i campioni di fluido sono stati
attraversati da raggi laser (alla lunghezza d’onda di 630 nm), e sono state analizzate
ampiezza e numero delle bande di interferenza registrate; sono stati riscontrati effetti
alla frequenza di irradiazione di 51,5 GHz (ma non a 41,5 GHz), tra l’altro
maggiormente accentuati nel caso che i campioni fossero costituiti da soluzioni al
2% di plasma umano, non giustificabili con l’esiguo aumento di temperatura
prodotto (pari a circa 1°C) se paragonati a quelli registrati inducendo lo stesso
aumento di temperatura con modalità di riscaldamento convenzionali. Secondo
Belyakov et al. (1989) anche altre proprietà del plasma, quali la costante dielettrica e
il coefficiente di assorbimento, possono essere alterate dall’irradiazione con
microonde, in misura differente a seconda del particolare soggetto donatore del
sangue. Khizhnyak e Ziskin (1996) hanno analizzato le peculiarità dei fenomeni di
riscaldamento con microonde e convezione in soluzioni acquose: oltre all’atteso
graduale innalzamento termico, i due ricercatori hanno constatato sia oscillazioni di
temperatura che un decremento della temperatura media, risultati attribuiti a
fenomeni di tipo convettivo, come per esempio la formazione di un vortice toroidale;
quando quest’ultimo diventa stabile, pur mantenendo costante la potenza irradiata, la
temperatura, dopo essere cresciuta, incomincia a diminuire; all’aumentare della
densità di potenza la temperatura locale, in modo del tutto inaspettato, decresce,
producendo un quasi paradossale effetto opposto al riscaldamento. Il supposto ruolo
dell’acqua come bersaglio primario della radiazione a microonde ha probabilmente
spinto Zavizion et al. (1994) e Kudryashova et al. (1995) verso lo studio di come
l’assorbimento in acqua delle microonde (alle lunghezze d’onda di 2,0, 5,84 e

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 53 -
7,12 mm) sia influenzato dalla presenza di altre sostanze quali gli amminoacidi (in
concentrazioni di 0,25 e di 2,5 mol/l); dal momento che l’assorbimento da parte
delle molecole amminoacidiche sarebbe di per sé trascurabile, si arrivò alla
conclusione che, nella maggior parte dei casi, l’assorbimento da parte delle soluzioni
decresceva in maniera proporzionale alla concentrazione degli amminoacidi, e in
misura tanto maggiore quanto maggiore era la lunghezza del radicale idrofobo; in
alcuni casi più limitati (sarcosina e glicina), al contrario, l’assorbimento da parte
dell’acqua sarebbe stato incrementato. Una dettagliata analisi teorica
dell’assorbimento delle microonde su strutture piane ad elevato contenuto di acqua
era stata precedentemente compiuta da Ryakovskaya e Shtemler (1983); i due autori
hanno fornito delle relazioni di dipendenza del SAR dalla frequenza della
radiazione, dalla temperatura, dallo spessore del mezzo assorbente e dalla presenza o
meno di strati di materiale dielettrico al di sopra e/o al di sotto del mezzo; hanno
inoltre modellato la risposta all’irradiazione di semplici e comuni strutture
biologiche da laboratorio come sospensioni cellulari in piastre di Petri e cuvette;
hanno anche posto l’accento sul fatto che la presenza di uno strato di materiale
dielettrico adiacente ad un film sottile di mezzo assorbente (situazione che si verifica
per esempio nella piastra di Petri) può aumentare il SAR anche di venti volte rispetto
a quello verificabile sulla superficie di uno strato spesso dello stesso mezzo, e ciò
potrebbe indurre, in alcune circostanze, a sottostimare notevolmente il SAR.
2.4.2 Effetti delle microonde sulle velocità di crescita e di morte cellulari
Una delle priorità della ricerca sugli effetti ottenibili in vitro, come identificata
da Repacholi (1998), è la determinazione della soglia di campo alle radiofrequenze
oltre la quale vengono alterate le cinetiche relative al ciclo cellulare e quelle di
proliferazione cellulare. L’influenza dell’esposizione alle onde radio sulla
moltiplicazione cellulare è stata studiata da diversi ricercatori, ottenendo differenti
risultati. Il dibattito sugli effetti delle microonde sulla velocità di crescita ha avuto

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 54 -
inizio tra gli anni ’70 e gli anni ’80 del secolo scorso, quando Grundler et al. (1977;
1982; 1988) hanno riportato come la velocità di crescita del lievito Saccharomyces
cerevisiae possa essere sia incrementata (fino al 15%) che diminuita (fino al 29%)
da microonde variandone la frequenza nel’intervallo compreso tra 41,8 e 42,0 GHz
(le prove condotte hanno riguardato sia cellule in sospensione che disposte in
monostrato). In accordo con le osservazioni di Grundler e Kaiser (1992), effetti di
entità simile sono prodotti da densità di potenza comprese tra 5 pW/cm2 e
10 mW/cm2 (con modulazione di 8 kHz); tuttavia il fatto che i successivi tentativi di
riprodurre i medesimi risultati di Furia et al. (1986) e di Gos et al. (1997) non siano
andati a buon fine suggerisce la possibilità che tali effetti dipendano da particolari
condizioni non ancora identificate o non controllabili. Dardanoni et al. (1985) hanno
studiato l’effetto della frequenza e della modulazione sulla crescita del lievito
Candida albicans; l’irradiazione con microonde modulate ad 1 kHz provocava una
riduzione della velocità di crescita del 15% alla frequenza di 72 GHz, ma non a 71,8
o a 72,2 GHz; a quest’ultima frequenza, dopo 3 ore di esposizione continua, la
velocità di crescita aumentava del 25%. Golant et al. (1994) hanno riportato che
l’irradiazione di una cultura di Saccharomyces cerevisiae per 50 minuti con
microonde alla frequenza di 46 GHz e con densità di potenza pari a
0,03 mW/cm2 produceva fluttuazioni periodiche marcatamente sincronizzate della
velocità di crescita e di gemmazione, che persistevano fino anche a venti generazioni
successive; la periodicità della gemmazione è un fenomeno che era stato osservato
anche nei campioni di controllo, dove tuttavia si era presentato in maniera molto
meno marcata e con periodi più brevi. Effetti di sincronizzazione delle microonde
sono stati osservati anche in esemplari di piante superiori da Shestopalova et al.,
(1995), che hanno esposto semi di orzo per 20 minuti a microonde con frequenza di
61,5 GHz e densità di potenza di 0,1 mW/cm2. Levina et al. (1989) hanno studiato
l’effetto delle microonde (30 minuti di esposizione a 1,5 mW/cm2 e alla lunghezza
d’onda di 7,1 mm) sullo sviluppo di popolazioni del protozoo Spirostum sp. in un
mezzo salino con lievito di birra (550 mg/l) come nutriente; lo studio ha evidenziato
come l’irradiazione abbia avuto effetti sui meccanismi di controllo della crescita

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 55 -
della popolazione, secondo modalità dipendenti dallo stadio di sviluppo della
popolazione e da altre caratteristiche proprie di quest’ultima. Tambiev et al. (1989)
hanno riscontrato che l’esposizione, per un tempo pari a 30 minuti, a radiazioni con
una densità di potenza di 2,2 mW/cm2 e con 7,1 mm di lunghezza d’onda,
incrementava la velocità di crescita dell’alga Spirulina platensis del 50%; poiché si
tratta di un’alga utilizzata per l’ottenimento, a livello industriale, di proteine
alimentari e di prodotti biologicamente attivi, gli autori hanno ritenuto che le loro
osservazioni potessero avere un notevole impatto dal punto di vista biotecnologico.
Altri studi, come quelli di Tambiev e Kirikova (1992), Rebrova (1992) e Shub et al.
(1995), hanno riportato effetti delle microonde sulla velocità di crescita di numerose
specie di batteri, cianobatteri, alghe, lieviti e piante superiori (finocchio, lattuga,
pomodoro); per esempio, per i due lieviti S. cerevisiae e S. carlsbergensis, le
microonde sono state in grado di abbreviare le fasi di crescita della coltura da 2,3
fino a 6 volte, e di incrementare la velocità di crescita della biomassa del 253%;
anche sul batterio Escherichia coli l’effetto sulla crescita si è rivelato essere sia
stimolante che di inibizione a seconda delle caratteristiche dell’irradiamento
(lunghezza d’onda compresa tra 6,0 e 6,7 mm, densità di potenza di 1 mW/cm2,
tempo di esposizione di 30 minuti); i tre studi, tuttavia, riportano solo una
descrizione sommaria di diverse esperienze degli autori, e perciò non forniscono
sufficienti dettagli per una piena valutazione degli effetti prodotti o per la possibile
ripetizione degli esperimenti condotti.
Webb e Dodds (1968) e Webb e Booth (1969) hanno riportato come cellule di
Escherichia coli fatte crescere in un brodo nutriente ed esposte a microonde
manifestassero una divisione cellulare più lenta ed anticipassero l’inibizione dei
processi metabolici rispetto al tempo di vita cellulare. Olsen (1965) osservò che il
numero di spore vitali di Aspergillus niger, Penicillum sp. e Rhizopus nigricans
veniva drasticamente ridotto dalle microonde durante il processo di panificazione.
Diverse teorie sono state proposte per spiegare come l’energia elettromagnetica
possa uccidere i microorganismi senza fare ricorso al calore; alcune di queste sono

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 56 -
contenute nella recensione di Knorr et al. (1994). Un’altra, detta “teoria della rottura
dielettrica” (Zimmermann et al., 1974), ipotizza che l’imposizione di un campo
elettrico esterno induca un potenziale elettrico transmembrana addizionale maggiore
del normale potenziale della cellula; superato un certo limite, la membrana cellulare
si rompe, dando luogo alla formazione di pori, con conseguente aumento della
permeabilità e perdita dell’integrità cellulare. Kinosita e Tsong (1977) hanno
proposto un meccanismo per lo sviluppo dei pori dovuto ai campi elettrici pulsanti,
presentando prove della migrazione di soluti intracellulari attraverso la membrana
cellulare con conseguente distruzione della cellula stessa. Pothakamury et al. (1993),
alla ricerca di una spiegazione alla base degli effetti non termici prodotti
dall’interazione tra campi elettromagnetici e microorganismi, conclusero che
l’effetto di inibizione dei campi magnetici sia statici che oscillanti sulla crescita
microbica racchiuderebbe in sé la possibilità di inattivare i microorganismi
eventualmente presenti negli alimenti. Il brevetto di Hofmann (1985) spiega che un
campo magnetico oscillante accoppia l’energia nelle parti magneticamente attive
delle macromolecole biologiche con le oscillazioni; in questo modo, quando un certo
numero di dipoli magnetici sono presenti in un’unica molecola, l’energia trasferita
alla molecola può essere sufficiente per la rottura di un legame covalente; sono
soggette a questo fenomeno molecole vitali per un microorganismo quali DNA o
proteine, per cui il microorganismo può essere distrutto dal trattamento, o anche
semplicemente reso inattivo dal punto di vista delle capacità riproduttive. Sulla base
di questo brevetto, Mertens e Knorr (1992) si sono proposti di valutare l’utilità
dell’energia veicolata dalle microonde per l’uccisione dei batteri in alimenti liquidi a
basse temperature e di mettere a punto un processo che rendesse al contempo minimi
i danni termici all’alimento; per far questo, i due ricercatori hanno studiato l’effetto
delle microonde sul microorganismo Pediococcus sp. in un sistema a flusso con
completo riciclo.
Pothakamury et al. (1995), ritenendo che l’inattivazione di microorganismi
mediante esposizione a campi elettrici pulsati ad alta tensione sia un obiettivo

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 57 -
piuttosto promettente nel campo delle tecnologie di preservazione degli alimenti non
basate sul riscaldamento, hanno dimostrato l’inattivazione dell’Escherichia coli (un
batterio Gram-negativo) e dello Staphylococcus aureus, mantenendo la temperatura,
durante le prove sperimentali, sempre al di sotto di quella letale per le due specie.
Cleary et al. (1990a) hanno condotto degli studi in merito esponendo in modo
continuo differenti tipologie di cellule ad onde radio alle frequenze di 27 MHz e di
2,45 GHz, e riportando una incrementata proliferazione di cellule di glioma
registrata a distanza di uno, tre e cinque giorni dall’esposizione delle cellule al
campo per due ore, con livelli di SAR compresi tra 5 e 50 W/kg; le prove sono state
condotte utilizzando un sistema di controllo termico che ha permesso di mantenere
le variazioni di temperatura al di sotto di 0,1°C; non è stata individuata una soglia
minima in quanto effetti sono stati riscontrati anche al minimo valore di SAR
applicato. Un altro studio di Cleary et al. (1990b), nelle medesime condizioni
sperimentali, e portato a termine con l’utilizzo dello stesso sistema, ha riscontrato
effetti simili anche su linfociti periferici umani; successivi studi (Cleary, 1995;
Cleary et al., 1996) hanno riscontrato, ancora nelle stesse condizioni sperimentali,
alterazioni nelle cinetiche del ciclo cellulare di cellule ovariche di criceto. Stagg et
al. (1997) hanno esposto cellule di glioma e colture primarie di cellule gliali di ratto
a onde radio identiche a quelle dei segnali dei telefoni cellulari, per periodi di tempo
di 24 ore, ma con livelli di esposizione molto inferiori rispetto a quelli degli studi
precedenti, osservando incrementi nell’assorbimento da parte di acido nucleico
marcato radioattivamente durante la sintesi del DNA in corrispondenza al valore di
SAR di 5,9 mW/kg, ma non a valori superiori (59 mW/kg), né inferiori (0,59
mW/kg); nello stesso lavoro, inoltre, sono presentate le curve di crescita per
entrambe le tipologie cellulari, ottenute mediante conta dopo tempi di esposizione
fino a dodici giorni, mostrando che non vi sono variazioni legate al campo
elettromagnetico applicato. In uno studio in cui hanno utilizzato onde
elettromagnetiche simili a quelle dei segnali dei telefoni cellulari GSM, Kwee e
Raskmark (1998) hanno valutato la moltiplicazione di cellule di epitelio amniotico

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 58 -
umano esposte a radiazione di frequenza 960 MHz, con valori di SAR pari a 0,021,
0,21 e 2,1 mW/kg, per tempi di esposizione pari a 20, 30 e 40 minuti: è stata
riscontrata una diminuzione della velocità di crescita, ma solo per tempi pari o
superiori a 30 minuti. Altri studi sulla crescita cellulare con esposizione a onde
radio, come quelli di Antonopoulos et al. (1997), Donnellan et al. (1997) e French et
al. (1997), forniscono ulteriori informazioni circa gli effetti dell’esposizione alle
onde radio sulla proliferazione cellulare, tuttavia dai dati forniti non è chiaro se, e in
quali condizioni, vi sono alterazioni nella velocità di crescita, né emerge un chiaro
legame tra la potenza assorbita e le risposte riscontrate.
2.4.3 Alterazioni cromosomiche ed effetti genetici delle microonde
L’assenza di effetti mutagenici o di ricombinazione genetica delle microonde è
stata chiaramente dimostrata a cavallo degli anni ’70 e ’80 del secolo scorso da
Dardalhon et al. (1980; 1981). Allo stesso tempo, tuttavia, altri studi hanno indicato
che le microonde influenzerebbero la struttura microscopica e la funzione dei
cromosomi, oltre che la tolleranza cellulare ai comuni mutageni e le funzioni di
riparazione delle lesioni; tra questi spiccano quelli di Belyaev et al. (1993a; 1993b;
1994; 1996), che rilevarono con un elevato grado di precisione le frequenze di
risonanza mediante una tecnica, detta AVTD (acronimo di “Anomalous Viscosity
Time Dependence”), che consente di rispecchiare i cambiamenti microstrutturali del
DNA e dei legami tra DNA e proteine: in corrispondenza di una frequenza di
risonanza, infatti, effetti biologici possono manifestarsi anche con densità di potenza
basse come 10-19 W/cm2, pur aumentando di intensità al crescere di quest’ultimo
parametro; l’intensità degli effetti non aumenta più oltre valori compresi tra 10-17 e
10-8 W/cm2, a seconda della densità cellulare nei campioni irradiati; per l’E. coli
sono stati individuati picchi di risonanza a 51,76 e a 41,34 GHz. Va detto tuttavia
che, come riportato da Pakhomov et al. (1998), la tecnica AVTD non rientra tra
quelle convenzionalmente accettate ed utilizzate nel campo della biologia cellulare,

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 59 -
e l’interpretazione dei risultati che essa fornisce è incerta; il problema è stato persino
oggetto di accesa disputa, con i commenti di Osepchuk e Petersen (1997) sul lavoro
di Belyaev et al. (1996) e la successiva replica di Belyaev et al. (1997); secondo
Pakhomov et al. (1998) è presumibile che parte della potenza ad una frequenza
armonica possa essere trasmessa ad un campione nonostante vi sia invece una forte
attenuazione alla frequenza fondamentale, e quantunque non fosse quello poco sopra
esposto il caso, le osservazioni effettuate sugli effetti della risonanza rappresentano
in ogni caso un fatto importante che meriterebbe di essere ripreso da altri studiosi
indipendenti. Rojavin e Ziskin (1995) hanno trovato che la preventiva esposizione di
colture di E. coli a microonde alla frequenza di 61 ± 2,1 GHz e con intensità di
potenza di 1 mW/cm2 ne aumentava il grado di sopravvivenza in seguito
all’esposizione a raggi UV; il meccanismo più probabile, secondo gli autori, sarebbe
l’attivazione, diretta o indiretta, di un sistema di riparo dalla luce. Pakhomova et al.
(1997), dopo aver studiato gli effetti genetici dell’irradiazione a frequenze comprese
tra 61,02 e 61,42 GHz sul ceppo D7 del lievito S. cerevisiae (densità di potenza di
0,13 mW/cm2, tempo di esposizione di 30 minuti), seguita da trattamento con raggi
UV (lunghezza d’onda di 254 nm, tempo di esposizione di 60 minuti, dose di 100
J/m2), giunsero alla conclusione che le microonde non alteravano la mutagenesi
indotta dai raggi UV, mentre potevano favorire i processi di ricombinazione genetica
indotti anch’essi dai raggi UV; inoltre, pur non potendo affermarlo con assoluta
certezza, gli autori ritennero che un meccanismo di tipo termico, per questo effetto,
fosse alquanto improbabile.
2.4.4 Effetti delle microonde su tessuti eccitabili e membrane
Oltre al patrimonio genetico, la membrana cellulare è considerata un altro
possibile bersaglio primario delle microonde. Diversi lavori affermano l’esistenza di
effetti anche profondi, tuttavia sono stati pochi i tentativi di riprodurli nuovamente.

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 60 -
Le membrane cellulari sono composte da proteine contenute in una matrice
lipidica, per cui la valutazione degli effetti prodotti da campi elettromagnetici alle
radiofrequenze deve tenere conto di entrambi i costituenti. Tra le proteine della
membrana, quelle di maggior interesse sono le proteine che costituiscono i canali
ionici: trattasi di complessi multiproteici che consentono il passaggio, attraverso
l’instaurazione di un gradiente elettrochimico, degli ioni inorganici, in maniera di
norma selettiva, attraverso la membrana. I canali ionici sono preposti al passaggio
transmembranaceo di ioni quali Na+, K+ e Ca2+. Questo, a sua volta, influenza
l’eccitabilità elettrica della membrana come pure alcune funzioni biologiche della
cellula, tra cui il rilascio di neurotrasmettitori. È stato riportato, a tal proposito, che i
campi elettromagnetici alle radiofrequenze, sia continui che pulsati, a varie potenze,
influenzano diverse proprietà dei canali ionici, possono causare un rallentamento del
processo di formazione delle proteine canale, possono diminuire la frequenza di
apertura dei singoli canali, possono incrementare notevolmente la velocità di scarica
rendendola “a scoppio” (Repacholi, 1998).
Per quanto riguarda l’influenza dell’esposizione alle microonde sul rilascio degli
ioni Ca2+, come sarà meglio analizzato nel paragrafo 2.4.5, vi sono stati studi (Bawin
et al., 1975; Dutta et al., 1984) che hanno riportato un incremento nel rilascio ionico,
mentre altri ricercatori (Merritt et al., 1982), con diverse caratteristiche di
modulazione, non hanno riscontrato variazioni rispetto alla norma. Brovkovich et al.
(1991) hanno riportato che la radiazione a 61 GHz di frequenza e con densità di
potenza pari a 4 mW/cm2 attivava in misura significativa la pompa Ca2+ nel reticolo
sarcoplasmico dei muscoli scheletrici e cardiaci del ratto; la velocità di assorbimento
degli ioni Ca2+ da parte delle membrane del reticolo sarcoplasmico è stata misurata
mediante un elettrodo selettivo in un mezzo contenente ATP; un trattamento
intermittente con microonde (5 minuti di esposizione, 15 minuti di pausa, 3 cicli),
effettuato sul reticolo sarcoplasmico di muscoli scheletrici, ha incrementato la
velocità di assorbimento del 23%, e tale velocità è stata mantenuta per un’ora oltre il
termine dell’esposizione; l’irradiazione ininterrotta, invece, non ha avuto effetti in

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 61 -
10 minuti, ma ha incrementato l’assorbimento del 27% dopo 20 minuti, e l’effetto ha
raggiunto il massimo al 48% dopo 40 minuti; per quanto riguarda i muscoli cardiaci,
un’esposizione di 5 minuti è stata sufficiente a produrre un incremento
dell’assorbimento del 18%. Geletyuk et al. (1995) hanno mostrato come la
radiazione a 42,25 GHz (tempo di esposizione di 20-30 minuti, densità di potenza di
0,1 mW/cm2) modifichi le caratteristiche di attivazione dei canali Ca2+-K+ di colture
di cellule renali; in particolare, il campo ha incrementato l’attività dei canali con una
bassa attività iniziale, mentre ha inibito i canali con una elevata attività iniziale. In
un successivo studio di Fesenko et al. (1995) gli effetti sopra riportati sono stati
riprodotti senza irradiare direttamente la membrana, solo mediante bagno in
soluzioni precedentemente esposte per 30 minuti a radiazione alla frequenza di
42,25 GHz e con densità di potenza di 2 mW/cm2; né il pH né la concentrazione di
Ca2+ della soluzione sono risultati alterati, dunque non è stato possibile comprendere
il meccanismo di alterazione dello stato di attivazione dei canali; le soluzioni
mantenevano la loro efficacia per almeno 10-20 minuti dopo il termine
dell’esposizione.
Vi sono studi scientifici che riguardano anche gli effetti di campi alle
radiofrequenze o di microonde sul trasporto attraverso le membrane dei cationi Na+
e K+; tra questi, quello di Cleary (1995) riporta effetti all’interno di intervalli di SAR
(tra 0,2 e 200 W/kg) e di frequenza (tra 27 MHz e 10 GHz) piuttosto ampi.
Kataev et. al (1993) hanno studiato le correnti della membrana in cellule di alga
gigante (Nitellopsis obtusa, Characea), riscontrando come l’irradiazione per 30-60
minuti a 41 GHz di frequenza (5 mW/cm2) fosse in grado di annullare la corrente dei
cloruri fino a 10-14 ore dopo il termine dell’esposizione; effetti inibitori marcati
sono stati registrati anche a 50 e a 71 GHz, mentre la maggior parte delle frequenze
applicate, comprese nell’intervallo 38-78 GHz, al contrario incrementavano (con
effetto reversibile nell’arco di 30-40 minuti) la corrente del 200-400%; il massimo
incremento di temperatura è stato di 1°C, e secondo gli autori né un effetto né l’altro
potevano essere giustificati con questo; anche la corrente del calcio è cambiata

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 62 -
durante l’esposizione al campo, ma l’effetto non è stato dipendente dalla frequenza e
poteva tutto sommato essere spiegato con il riscaldamento. Esperimenti su
membrane artificiali e neuroni di lumaca non hanno mostrato nessun effetto
specifico della frequenza delle microonde (Alekseev e Ziskin, 1995). Burachas e
Mascoliunas (1989) hanno studiato gli effetti delle microonde sul potenziale di
azione composto (CAP) di nervo sciatico (isolato) di rana; il potenziale è diminuito
esponenzialmente fino a ridursi di dieci volte dopo 50-110 minuti di esposizione a
77,7 GHz di frequenza (densità di potenza di 10 mW/cm2), per poi tornare sui livelli
originari al termine dell’esposizione; tuttavia, dopo ciò, il nervo diventava più
sensibile alle microonde, e durante la successiva esposizione il decadimento si
mostrava ancora più rapido, e si completava in solo 10-15 minuti. Un differente
effetto sulla stessa tipologia di tessuto è stato descritto da Chernyakov et al. (1989), i
quali lo esposero per 2-3 ore a microonde (con densità di potenza di 0,1-0,2
mW/cm2), sia con cambiamenti regolari di 1 GHz di frequenza ogni 8-9 minuti, sia
con cambiamenti di frequenza casuali (all’interno della banda 53-78 GHz) ogni 1-4
minuti; in particolare la seconda tipologia di trattamento, in 11 casi su 12, ha indotto
un brusco “riarrangiamento” del CAP secondo modalità, a detta degli autori, del
tutto imprevedibili. Nessuno di questi effetti è stato tuttavia riscontrato da Pakhomov
et al. (1997a; 1997b), secondo i quali l’irradiazione per 10-60 minuti con densità di
potenza tra 0,2 e 1 mW/cm2, sia a diverse frequenze mantenute costanti che a
frequenze variate a gradino, non ha alterato il CAP; con densità di potenza compresa
tra 2,0 e 2,8 mW/cm2 si sono avuti dei modesti effetti, indipendenti però dalla
frequenza e attribuibili probabilmente all’innalzamento termico. Secondo
Chernyakov et al. (1989) microonde a bassa potenza possono mutare le funzioni
della membrana di cellule di muscoli striati e pace-maker cardiaco: l’esposizione per
90 secondi o meno a densità di potenza di 0,1-0,15 mW/cm2 (a frequenze comprese
tra 54 e 78 GHz) ha rallentato la perdita naturale del potenziale transmembranaceo
nei miociti, o addirittura lo ha incrementato di 5-20 mV; l’effetto è stato riscontrato
nell’80% dei casi, senza tuttavia una chiara dipendenza dalla frequenza della
radiazione; gli stessi autori, per quanto riguarda l’effetto delle microonde

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 63 -
sull’attività di pace-maker, hanno trovato che su cellule di rana prelevate dalla zona
senoatriale, nella maggior parte dei casi, l’esposizione alle microonde produceva un
immediato (spesso in meno di 2 secondi) restringimento dell’intervallo tra gli
impulsi; l’entità dei cambiamenti aumentava linearmente con l’incremento della
potenza incidente da 20-30 sino a 500 mW/cm2; rispetto alla frequenza, le variazioni
prodotte non hanno mostrato un andamento monotono, bensì picchi ben localizzati;
il riscaldamento massimo dopo 2 secondi di esposizione è stato calcolato (per
1 mW/cm2) in 0,005°C, e considerata anche la rapida risposta fisiologica, gli autori
hanno escluso che l’effetto potesse essere di tipo termico.
Benché dunque sembri confermato il fatto che i campi elettromagnetici possono
modificare il meccanismo di scambio ionico delle membrane, tuttavia non sembra
essere allo stesso tempo emerso quali siano le specifiche modalità di interazione, e
rimangono sconosciuti i meccanismi attraverso i quali i campi elettromagnetici
agiscono effettivamente sulla struttura molecolare delle proteine. Ugualmente
irrisolto sembra rimanere, sia a livello meccanicistico che dal punto di vista degli
effetti apparenti, il ruolo dei campi elettromagnetici alle radiofrequenze e delle
microonde rispetto ai lipidi della membrana. Phelan et al. (1992) hanno ipotizzato
che l’esposizione alle radiofrequenze induca transizioni di fase nelle vescicole
lipidiche (liposomi) esposte a campi continui e con un SAR pari a 0,2 W/kg,
tuttavia, come anche ammette a tal proposito Repacholi (1998), sono forse troppo
pochi i dati a disposizione per poter stabilire qualcosa di maggiormente concreto.
2.4.5 Effetti sull’omeostasi degli ioni Ca2+
Il lavoro pioniere su tale argomento fu quello di Bawin et al. (1975), in cui fu
riportato che la radiazione in modulazione di frequenza a 147 MHz incrementava in
vitro l’efflusso degli ioni Ca2+ da cellule neurali di pulcino, tuttavia non vi erano
effetti con la sola frequenza portante; il picco di effetti si ottenne invece tra gli 11 e i
16 Hz di modulazione; la potenza incidente applicata era compresa tra 10 e 20

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 64 -
W/m2, ma gli effetti non si mostrarono funzione della potenza. Pochi anni più tardi
Bawin et al. (1978), utilizzando un’onda portante a 450 MHz modulata a 16 Hz, con
una densità di potenza incidente di 1 W/m2, riscontrarono gli stessi effetti di
incremento anche con una densità di potenza di 10 W/m2, mentre nessun effetto si
manifestava con densità inferiori (0,05 W/m2) o superiori (20 W/m2). Un effetto
biologico che si manifesta all’interno di un intervallo relativo ad una certa grandezza
fisica, ma non in corrispondenza degli estremi dello stesso intervallo, è quello che
nella letteratura specializzata si trova indicato spesso come “window phenomenon”,
ovvero “fenomeno finestra”, e proprio gli effetti descritti da Bawin et al. (1978)
furono citati così, relativamente alla densità di potenza, da Postow e Swicord (1996).
“Effetti finestra” furono trovati anche da Blackman et al. (1979; 1980a), che
utilizzarono una radiazione a 147 MHz di frequenza e con una potenza incidente di
7,5 W/m2, riscontrando effetti che non si manifestavano invece né a 5 W/m2, né a 10
W/m2; un ulteriore studio di Blackman et al. (1980b) individuò degli effetti in una
finestra della frequenza di modulazione a 16 Hz con una frequenza portante di
50 MHz e in una finestra della densità di potenza intorno a 17 W/m2. In cellule
nervose coltivate in un campo alla frequenza di 915 MHz, fu riscontrato da parte di
Dutta et al. (1984) un incremento nell’efflusso di calcio per SAR pari a 0,05 e
1,0 W/kg, ma non per valori inferiori, superiori o intermedi. Le prove di Shelton e
Merritt (1981), condotte alla frequenza portante di 1000 MHz modulata a 16 Hz e a
32 Hz e con densità di potenza incidente di 5, 10, 20 e 150 W/m2 su tessuti cerebrali
di ratto, d’altra parte, non hanno rivelato alcun effetto, così come è accaduto anche
nelle prove condotte da Merritt et al. (1982) irradiando con microonde (1000 MHz,
0,29 e 2,9 W/kg; 2450 MHz, 3 W/kg; 2060 MHz, 0,12 e 2,4 W/kg) tessuti cerebrali
di ratto preventivamente caricati con ioni calcio mediante iniezione
intraventricolare.
I lavori di Bawin et al. (1978) e di Blackman et al. (1979; 1980a) sembrerebbero
indicare che la finestra relativa alla densità di potenza riduca la sua ampiezza al
crescere della frequenza delle onde elettromagnetiche, e ciò potrebbe essere

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 65 -
confermato, seppur indirettamente, dalle misure effettuate da Bawin e Adey (1976) e
da Blackman et al. (1991), che hanno mostrato come l’efflusso di ioni calcio da
cellule cerebrali di pulcino possa essere alterato dall’esposizione a campi di
frequenza estremamente bassa e con densità di potenza inferiori di diversi ordini di
grandezza; a tale proposito, Prato et al. (1996) hanno messo in luce come, dal punto
di vista concettuale, l’esposizione a frequenze bassissime corrisponda
all’esposizione ad un campo con frequenza portante infinita e modulazione di
frequenza piccolissima, per cui, se i meccanismi associabili sono simili,
diventerebbe importante considerare il campo statico ambientale presente durante
l’esposizione.
Riassumendo, finestre relative alla densità di potenza sono state osservate per
modulazioni con frequenze molto basse, sebbene le prove che non vi siano effetti
oltre i 1000 MHz non siano del tutto convincenti dal momento che non sono stati
condotti esperimenti a quelle frequenze e con piccoli valori di SAR o di densità di
potenza.
2.4.6 Altri effetti delle microonde in vitro
Bulgakova et al. (1996) hanno studiato come l’esposizione dello Staphylococcus
aureus alle microonde influenzi la sua sensibilità ad antibiotici con differenti
meccanismi di azione; i tempi di esposizione sono andati da 1,5 a 60 minuti, con
frequenza fissa di 42,195 o 54 GHz oppure con frequenza variata a gradino tra 66 e
78 GHz con passo di 1 GHz, e con densità di potenza di 10 mW/cm2; il
riscaldamento imputabile alle microonde non ha mai superato 1,5°C, e sono state
condotte oltre un migliaio di prove sperimentali con 14 differenti antibiotici, tuttavia
gli effetti riscontrati, per quanto molto vari (la sensibilità è risultata aumentata o
diminuita a seconda della concentrazione dell’antibiotico), non hanno mostrato una
chiara dipendenza dalla potenza della radiazione né dalla frequenza; gli autori, tra
l’altro, hanno fatto notare come il trattamento con microonde potrebbe rivelare (o

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 66 -
anche indurre) una eterogeneità nella sensibilità ad alcuni antibiotici di una
popolazione cellulare.
Shub et al. (1995) hanno mostrato cambiamenti nella trasmissione di plasmidi R
in diversi ceppi di Escherichia coli e di Staphylococcus aureus dopo 30 minuti di
esposizione a microonde di lunghezza d’onda compresa tra 6,0 e 6,7 mm.
Berzhanskaya et al. (1995) hanno misurato una diminuzione della bioluminescenza
del Photobacterium leiognathi durante l’irradiazione con microonde a frequenze
comprese tra 36,2 e 55,9 GHz. Diversi risultati di studi su Enterobacter aerogenes
ed Escherichia coli hanno infine mostrato che l’irradiazione con microonde, a
seconda dei casi, inibiva o stimolava la sintesi di proteine, DNA e RNA, e la crescita
cellulare (Pakhomov et al., 1998).
Goldblith e Wang (1967) non hanno registrato differenze esponendo sospensioni
di Escherichia coli e di spore di Bacillus subtilis a riscaldamento mediante
microonde e a riscaldamento convenzionale; Welt et al. (1994) hanno indagato
l’effetto del trattamento con microonde sottoponendovi spore di Clostridium
sporogenes, senza trovare un incremento dell’effetto letale che non fosse
giustificabile con il rapido riscaldamento prodotto dalle microonde stesse.
Rebrova (1992) ha passato in rassegna diversi effetti delle microonde sul
metabolismo cellulare, sulla sintesi enzimatica e su altri processi in organismi
unicellulari, come l’incremento e l’annullamento della sintesi delle colicine nel
batterio Escherichia coli, la stimolazione della sintesi di enzimi fibrinolitici nel
Bacillus firmus, l’aumento del contenuto di peptidi, DNA e RNA nel Bacillus
mucilaginous, l’abbattimento del grado di tolleranza agli antibiotici dello
Staphylococcus aureus; l’autrice ha riportato che la massima variazione prodotta
dalle microonde andava dal 20 al 90%, a seconda della lunghezza d’onda e dello
stato iniziale del ceppo batterico; al contrario dei batteri, la velocità di riproduzione
e le proprietà biosintetiche dei funghi Aspergillus sp., Endomyces fibuliger e
Dacthilyum dendraides cambiavano solo in seguito ad esposizioni ripetute (anche
dieci volte); in alcuni lieviti le microonde acceleravano la formazione del maltosio

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 67 -
del 73%, mentre la sintesi del diacetile e delle aldeidi diminuiva del 20%; in
generale, la stimolazione selettiva della produzione di alcuni enzimi a discapito di
altri potrebbe costituire un importante obiettivo nel campo delle biotecnologie.
2.4.7 Considerazioni sui meccanismi degli effetti non termici
Come spiegato nei paragrafi precedenti, è stato provato, seppure in modo non
del tutto univoco, che esistono diversi effetti biologici che non dipendono da un
incremento di temperatura del campione. Alle radiofrequenze, sia il campo elettrico
che quello magnetico penetrano nei tessuti (Postow e Swicord, 1996), generando al
contempo calore, tuttavia gli effetti dei due campi sui tessuti tenderebbero a
distinguersi se il tessuto è dotato di elementi sensibili, all’uno o all’altro campo
piuttosto che a differenze di potenziale, effettivamente relazionati ad una particolare
funzione fisiologica o biochimica. Inoltre, l’effetto dipenderebbe anche dalla forma
d’onda e non solo dalla corrente indotta; onde continue e onde alle radiofrequenze in
modulazione di frequenza estremamente bassa danno luogo, anche se non
necessariamente, ad effetti non termici differenti. Se esistono, per esempio, elementi
sensibili al campo elettrico, è improbabile che essi possano rispondere abbastanza
rapidamente alla frequenza portante, tuttavia questa può costituire il mezzo
attraverso il quale può essere veicolata al tessuto una frequenza pulsata senza che vi
sia una significativa attenuazione del campo. Questo presupporrebbe, naturalmente,
l’esistenza di meccanismi di demodulazione dipendenti dall’ampiezza che
“separino” la frequenza di modulazione da quella portante, che sembrerebbero in
maniera quasi sorprendente confermati dai risultati riportati in letteratura circa gli
effetti della radiofrequenza portante e della modulazione in frequenza estremamente
bassa sull’efflusso degli ioni Ca2+ (vedi paragrafo 2.4.5), sull’attività dell’ornitina
decarbossilasi (Litovitz et al., 1991; Litovitz et al., 1993; Penafiel et al., 1997) e sul
comportamento associato ai recettori oppioidei (Kavaliers et al., 1994; Del Seppia et
al., 1995; Rojavin et al., 1998). Per quanto riguarda l’esposizione a campi di

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 68 -
frequenza estremamente bassa, solo il campo magnetico penetra il tessuto, e ciò può
generare un campo elettromagnetico indotto e una corrente indotta, secondo la legge
di Faraday. Un campo di radiofrequenze modulato con frequenze estremamente
basse può trasmettere lo stesso campo elettromagnetico ed indurre corrente senza
che il campo elettrico penetri nel tessuto, dunque la comprensione dei meccanismi
associati agli effetti delle frequenze estremamente basse potrebbe fornire importanti
indicazioni per la comprensione dei meccanismi attraverso i quali i campi alle
radiofrequenze in modulazione di frequenza estremamente bassa provocano effetti
non termici.
Un interessante aspetto è costituito dal fatto che buona parte dei risultati mostrati
suggeriscono con forza che l’entità degli effetti non termici è indipendente (o quasi)
dalla potenza assorbita, e ciò è dimostrato anche dalla presenza di effetti finestra,
come quelli individuati da Stagg et al. (1997) per quanto riguarda la velocità di
crescita e, per quanto riguarda l’efflusso degli ioni Ca2+, da Bawin et al. (1975;
1978) e da Blackman et al. (1979; 1980a; 1980b); questi ultimi, in particolare, hanno
suggerito come la modulazione di frequenza possa produrre effetti biologici diversi
rispetto all’irradiazione con onde continue, anche se l’esposizione viene
normalizzata rispetto alla medesima densità di potenza.
2.5 Le specie microbiche indagate
2.5.1 Il Bacillus clausii
Il Bacillus clausii è un batterio Gram-positivo (dotato cioè di una spessa parete
cellulare), capace di movimento, sporigeno e, come la maggior parte dei bacilli, ha
la forma di un bastoncello; la parete cellulare è costituita da peptidoglicani
(soprattutto mureina). È un’endospora che produce batteri che a loro volta danno
origine a spore di forma ellissoidale particolarmente resistenti a diversi antibiotici

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 69 -
inclusi eritromicina, lincomicina e cefalosporine (Green et al., 1999). Le sue cellule
tendono ad allinearsi in formazioni tipo catena, osservabili come filamenti. Il B.
clausii è alcalofilo, e questa sua natura si è rivelata utile nella prevenzione e nel
trattamento di diversi disturbi gastrointestinali come forma di batterio-terapia orale
(Senesi et al., 2001), oltre al fatto che rende la specie tutt’oggi interessante per
diverse possibili applicazioni biotecnologiche, e oggetto di studi anche recenti, come
quello di Kobayashi et al. (2005). Può trovarsi in diversi ambienti alcalini, inclusi
suoli e habitat marino. Kobayashi et al. (1995), studiando il ceppo KSM-K16, hanno
osservato la sua crescita tra 15 e 50°C e nel campo di pH compreso tra 7 e 10,5,
individuando nella temperatura di 40°C e nel pH pari a 9,0 le condizioni ottimali per
la sua crescita. Benché la respirazione aerobica sia più efficiente dal punto di vista
energetico, poiché il B. clausii vive comunemente in ambienti poveri di ossigeno
(come il suolo), esso è in grado di ridurre i nitrati utilizzandoli come gli accettori
terminali degli elettroni durante la respirazione anaerobica; la riduzione operata dal
B. clausii dei nitrati a nitriti può essere completata da altri batteri quali per esempio
la Pseudomonas aeruginosa (Paustian, 2000).
Le spore del B. clausii sono utilizzate nel probiotico Enterogermina®, che
stimola le funzioni immunitarie (soprattutto durante l’uso di antibiotici) del tratto
gastrointestinale incrementando la produzione di immunoglobulina A, e agendo così
indirettamente come antagonista di altri batteri patogeni; la sua capacità di resistere
a molti antibiotici, che potrebbe farlo apparire pericoloso per l’essere umano, gli
consente invece di svolgere al meglio questa sua attività (Green et al., 1999). La
specie è inoltre di interesse biotecnologico, in quanto i ceppi di B. clausii producono
enzimi quali xilanasi, cellulasi, amilasi e proteasi particolarmente utili per l’industria
(Senesi et al., 2001). Il ceppo KSM-K16, per esempio, produce proteasi (M, H e N-
proteasi) che spaccano i polipeptidi in porzioni più piccole di amminoacidi,
secernendoli direttamente nel brodo di coltura, in particolare durante i periodi di
scarsa nutrizione (quando contemporaneamente ha luogo anche il processo di
sporulazione), e ciò è oggetto di studi per l’implementazione di processi produttivi

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 70 -
di massa a livello industriale (Christiansen e Nielsen, 2002). Tra le possibili
applicazioni, Denizci et al. (2003) hanno individuato la possibilità di utilizzare le
proteasi così prodotte per ottenere biomasse utilizzabili a partire da proteine fibrose
come corna, piumaggio e pelame, per l’idrolisi di gelatine o in processi di recupero
dell’argento.
Il probiotico, resistente agli antibiotici, Enterogermina®, consiste in quattro
ceppi batterici (O/C, N/R, SIN e T), tutti recentemente riclassificati da B. subtilis a
B. clausii (Duc et al., 2004); è utilizzato in particolare nel trattamento della diarrea e
nella prevenzione dei disturbi gastrointestinali; la resistenza delle spore agli
antibiotici le rende particolarmente utili per l’utilizzo congiunto con trattamenti
antibiotici per altri patogeni; Duc et al. (2004) sostengono che l’acquisizione di
ulteriori conoscenze su tale attività del B. clausii consentirà di ottimizzare in campo
medico l’utilizzo del batterio e di individuarne, eventualmente, ulteriori
applicazioni.
I meccanismi attraverso i quali il B. clausii agisce alleviando i disturbi
gastrointestinali non sono stati ancora compresi del tutto. Casula e Cutting (2002),
sviluppando un metodo per lo studio delle caratteristiche della colonizzazione del B.
clausii nell’intestino di topi, hanno mostrato come questo è in grado di colonizzare
per brevi periodi di tempo la parete intestinale e provocare nei topi una risposta
immunitaria che uccide i batteri patogeni.
2.5.2 La Pseudomonas aeruginosa
La Pseudomonas aeruginosa è un batterio Gram-negativo, aerobio/anaerobio
facoltativo, capace di movimento (grazie alla presenza di 1-3 flagelli unipolari),
asporigeno, di piccole dimensioni (tra 0,5 – 1,0 µm per 1,5 – 5 µm), non
fermentativo, con semplici richieste nutrizionali, dotato di una spessa capsula,
largamente diffuso in ogni tipologia di ambiente (suolo, acqua, ambiente

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 71 -
ospedaliero, etc.) (La Placa, 2005). Il campo di temperature all’interno del quale la
crescita della P. aeruginosa è ottimale è compreso tra 4 e 42°C (Murray e
Rosenthal, 2008), ma è in grado di crescere anche a temperature superiori
(Palleroni, 1984). È considerata un patogeno opportunista per l’uomo, e trattasi di
un batterio molto virulento, che non riesce tuttavia a provocare seri quadri patologici
in soggetti immunocompetenti, fatta eccezione per i casi di infezione del meato
urinario, che può estendersi causando la necrosi dei tessuti e, come estrema
conseguenza, il decesso del paziente per setticemia (La Placa, 2005). I soggetti
maggiormente a rischio sono dunque principalmente quelli immunodepressi, ma per
motivi di maggiore esposizione anche in generale quelli con respirazione assistita e i
dializzati (il livello di trasmissibilità di apparecchi respiratori e di emodialisi è
particolarmente elevato), gli ustionati, quelli affetti da fibrosi cistica, i diabetici, i
tossicodipendenti; benché la P. aeruginosa sia in grado di infettare tutti i distretti
corporei, le tipologie di infezioni più diffuse sono quelle polmonari, quelle cutanee,
quelle delle vie urinarie, quelle dell’orecchio, quelle dell’occhio (Murray e
Rosenthal, 2008). Le terapie con antibiotici sono difficili per l’elevato grado di
resistenza della P. aeruginosa a molti di questi, e la situazione è complicata dal fatto
che i soggetti con difese immunitarie compromesse rispondono già di per sé
scarsamente alla terapia antibiotica. Essendo la monoterapia del tutto inefficace, si
utilizzano sempre combinazioni di antibiotici; tuttavia occorre sottolineare come
l’utilizzo di antibiotici ad ampio spettro, oltre a danneggiare eccessivamente la flora
batterica competitiva, può avere l’effetto di selezionare ceppi del batterio ad elevata
resistenza (La Placa, 2005). Come riportato da Martin et al. (2003), per quanto il
decorso clinico sia un fatto tutto sommato ben noto, sono al contrario piuttosto
scarse le conoscenze sulle cause di tali infezioni, e ciò nonostante si sia trattato,
negli Stati Uniti, negli anni che sono stati interessati dall’indagine (1979 – 2000)
della decima causa di morte. La resistenza della P. aeruginosa agli antibiotici è
dovuta, da una parte, alla produzione di una β-lattamasi in grado di inattivare le
penicilline e le cefalosporine, e dall’altra alla capacità di alterare la permeabilità dei
canali di membrana; la specie è inoltre resistente a diversi tipi di disinfettanti

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 72 -
(Murray e Rosenthal, 2008). Nei soggetti sani la P. aeruginosa è comunque presente
e costituisce all’incirca il 6% della flora batterica intestinale, mentre può
raggiungere il 38% in pazienti ospedalizzati e il 78% in soggetti
immunocompromessi (La Placa, 2005).
Il nome “Pseudomonas” significa letteralmente “falsa unità”, in quanto deriva
dalla composizione delle due parole greche “pseudo” (ovvero “falso”) e “monas”
(ovvero “singola unità”), e sta ad indicare la propensione del batterio in questione ad
aggregarsi insieme ad altri per formare una struttura complessa detta biofilm
(McArthur, 2006). La formazione del biofilm è considerata un fattore di virulenza in
quanto ha l’effetto di accrescere le caratteristiche di resistenza dei batteri sia agli
antibiotici che alla pulitura e alla fagocitosi (Parsek e Singh, 2003). Una delle prime
osservazioni del biofilm, definibile come una comunità strutturata di batteri
racchiusi in una matrice extracellulare di natura polimerica prodotta dalle cellule
stesse, fu quella di Henrici (1933); ma nonostante l’esistenza dei biofilm sia nota da
quasi un secolo, il processo di formazione ha iniziato ad essere studiato a livello
genetico e molecolare solo di recente. Costerton (2005) ha rilevato che una
percentuale compresa tra il 65 e l’80% delle infezioni batteriche trattate nei Paesi
sviluppati è imputabile a batteri che crescono in biofilm, e che alla stessa categoria
di batteri sono ascrivibili le infezioni batteriche associate all’uso di dispositivi
medici impiantabili; inoltre, al contrario della maggior parte dei biofilm, che sono
eterogenei, ovvero costituiti da batteri di più specie, quello generato dalla
P. aeruginosa è in genere monospecie. La matrice extracellulare del biofilm,
composta principalmente da esopolisaccaridi (la cui percentuale tende ad aumentare
con l’età del biofilm), proteine, fimbie, pili e DNA extracellulare, è in realtà
considerata la materia prima del biofilm, in quanto ne costituisce una percentuale
che va dal 50 al 90%; tale matrice può essere anche fortemente idrata (l’acqua viene
incorporata nella struttura mediante legami a idrogeno), e ciò ne previene
l’essiccamento, oltre a costituire una vera e propria barriera fisica che limita
fortemente la diffusione degli antibiotici al suo interno (De Beer e Stoodley, 2006).

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 73 -
Sauer et al. (2002) hanno fornito una accurata descrizione del processo di
formazione del biofilm, poi ripresa anche da Caiazza e O’Toole (2004),
individuando sei fasi, che sono nell’ordine: forma planctonica (nella quale il batterio
si muove liberamente nel liquido per mezzo del flagello di cui è dotato), attacco
reversibile (durante il quale il batterio si lega reversibilmente alla superficie
mediante deboli interazioni idrofobiche e forze di Van Der Waals che si instaurano
tra alcuni componenti della parete cellulare e la superficie), attacco irreversibile
(durante il quale comincia la formazione di un biofilm monostrato in quanto
comincia la produzione degli esopolisaccaridi), formazione delle microcolonie,
maturazione (durante la quale aumenta la produzione di esopolisaccaridi e di
conseguenza lo spessore del biofilm), dispersione (durante la quale cellule situate in
zone interne del biofilm si staccano e ritornano alla forma planctonica, con un
migliore accesso ai nutrienti sia per le cellule staccatesi, sia per quelle rimaste nel
biofilm, grazie alle cavità così formatesi). Vi sono dei metodi di comunicazione
cellulare che sovrintendono a tali fenomeni, e che sono in grado di guidare il
comportamento dell’intera popolazione batterica, che vanno sotto il nome di
Quorum Sensing; secondo quest’ottica, i batteri in grado di organizzarsi in tal modo
non possono essere considerati come semplici organismi individuali, bensì come
organismi sociali in grado di formare complesse strutture multicellulari, possibili
però solo a condizione che essi possano interagire tra di loro in qualche modo; da un
punto di vista evoluzionistico l’aggregazione in strutture complesse si è certamente
rivelata una scelta vincente, e lo dimostra il fatto che non solo gli eucarioti (che sono
per la maggior parte già di per sé organismi pluricellulari), ma anche il 99% dei
procarioti tende in realtà ad organizzarsi in ammassi cellulari più o meno complessi,
tra cui appunto i biofilm (Costerton et al., 1995).

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 74 -
2.5.3 Lo Staphylococcus aureus
Lo Staphylococcus aureus è un batterio Gram-positivo, aerobio/anaerobio
facoltativo, incapace di movimento, asporigeno, di forma sferica, che non presenta
una capsula evidente; è un batterio fortemente alofilo, essendo in grado di
svilupparsi anche in ambienti caratterizzati da un’elevata concentrazione (il 7,5%) di
NaCl (Ryan e Ray, 2004). Il nome della specie (aureus) deriva dal tipico pigmento
giallo oro caratteristico delle colture del batterio; lo S. aureus è presente sulla cute,
nel naso e nella faringe della maggior parte degli esseri umani adulti, perciò esso è
in grado di causare infezioni in qualsiasi momento (Kluytmans et al., 1997). La
capsula è di natura polisaccaridica ed è dotata di potere antifagocitario, ovvero è
inattaccabile da parte dei granulociti neutrofili, che costituiscono fino al 70% dei
leucociti (globuli bianchi) presenti nell’organismo umano; inoltre sulla superficie
della cellula batterica sono presenti diverse proteine che sono in grado di interagire
con altre strutture proteiche dell’organismo umano (Lowy, 1998). Lo S. aureus è
responsabile di infezioni acute che possono interessare distretti corporei quali cute,
apparato scheletrico, apparato respiratorio, apparato urinario, sistema nervoso
centrale; alcuni ceppi, inoltre, producono e rilasciano esotossine che possono
provocare intossicazioni e manifestazioni morbose di diverso tipo, tra cui
l’enterotossina, che provoca gastroenteriti in seguito ad eventi di intossicazione
alimentare causati da assunzione di cibi ricchi di lipidi (come per esempio crema o
panna), che favoriscono la crescita batterica, in cui sia presente una certa quantità di
enterotossina (Davis et al., 1973).
Lo S. aureus è dotato di una elevata capacità di resistenza agli antibiotici, in
particolare nelle infezioni nosocomiali; tale capacità è andata aumentando nel corso
degli anni (Tenover et al., 2001), rendendo antibiotici quali cefalosporine e
penicilline ormai non più efficaci (Chambers, 2001); pare, come affermato da Cui et
al. (2003), che la crescente resistenza alla vancomicina dipenda dall’inspessimento
della parete cellulare. Lo S. aureus, inoltre, a differenza di molte altre specie, è in

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 75 -
grado di resistere all’NO, una sorta di “antibiotico naturale” prodotto dall’organismo
umano che impedisce ai microorganismi sia la respirazione che la fermentazione, e
ciò grazie alla produzione di acido lattico che, attraverso il mantenimento di una
particolare situazione di equilibrio chimico, gli consente di vivere in un ambiente
altrimenti ostile; un enzima prodotto dallo stesso batterio promuove inoltre la
conversione dell’NO in prodotti non tossici; si ritiene che la stessa sostanza
pigmentosa che caratterizza il batterio sia anche responsabile dell’azione
antiossidante che consente al batterio di sopravvivere in tali condizioni (Clauditz et
al., 2006), e che possa addirittura avere un ruolo nella resistenza all’attacco da parte
dei granulociti neutrofili (Liu et al., 2005).
2.5.4 La Listeria monocytogenes
La Listeria monocytogenes è un batterio Gram-positivo, asporigeno, aerobio
facoltativo, a forma di bastoncello, capace di movimento grazie a dei flagelli a
temperature fino a 30°C, e non più a 37°C, ma può invece spostarsi tra cellule
eucarioti grazie alla formazione, mediante polimerizzazione, di filamenti di una
sostanza proteica, l’actina (Gründling et al., 2004).
La Listeria monocytogenes è l’agente patogeno causa della listeriosi, uno dei più
virulenti di origine alimentare con percentuali di mortalità tra gli individui colpiti
dall’infezione comprese tra il 20 e il 30% (Ramaswamy et al., 2007). Responsabile
negli Stati Uniti di circa 2.500 casi di infezione e di circa 500 decessi all’anno, la
listeriosi è la principale causa di morte tra le infezioni provocate da patogeni
batterici di origine alimentare, con mortalità che superano anche la Salmonella e il
Clostridium botulinum (Dharmarha, 2008). Frequente veicolo di infezione,
soprattutto nel caso delle meningiti nei neonati (ai quali l’infezione viene trasmessa
dalla madre per via transvaginale), di cui la L. monocytogenes è la terza causa, sono
i formaggi a pasta molle, che possono essere contaminati e consentono la crescita
batterica (Genigeorgis, 1991).

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 76 -
La L. monocytogenes fu descritta per la prima volta da Murray et al. (1926), che
le attribuirono il nome di Bacterium monocytogenes, poi cambiato nella forma
attuale da Harvey Pirie (1940); si dovette tuttavia attendere il secondo dopoguerra
per identificare la specie come importante causa di sepsi e meningiti neonatali
(Potel, 1952). Negli adulti, invece, solo più tardi la listeriosi fu associata ai pazienti
con sistema immunitario compromesso, come coloro che hanno fatto uso di sostanze
immunosopprimenti e corticosteroidi nel trattamento tumorale o i trapiantati
d’organo, oltre quelli infettati da virus HIV (Schlech, 2001); fu solo nel 1981, come
riportato da Schlech et al. (1983), che la L. monocytogenes fu identificata quale
causa di patologie di origine alimentare, quando a Halifax, in Nuova Scozia, 41 casi
di listeriosi (di cui 18 conclusi col decesso del paziente) in donne in stato di
gravidanza e neonati furono collegati, da un punto di vista epidemiologico, al
consumo di “coleslaw” (un’insalata di cavolo tritato, carote, cipolle e maionese)
contenente cavoli che erano stati concimati con letame ovino contaminato dalla
L. monocytogenes. Da allora sono numerosi i casi documentati di listeriosi di origine
alimentare, e oggi la L. monocytogenes è largamente considerata un consistente
pericolo nell’industria alimentare (Ryser e Marth, 2007).
L’infezione da L. monocytogenes è causa, come detto, della malattia detta
listeriosi, tra le cui manifestazioni vi sono, oltre alle infezioni intrauterine e della
cervice nelle donne in stato di gravidanza (che possono condurre all’aborto
spontaneo), setticemia e meningiti (Gray e Killinger, 1966), encefaliti (Armstrong e
Fung, 1993), ulcera corneale (Holland et al., 1987), polmonite (Whitelock-Jones et
al., 1989). I neonati sopravvissuti alla listeriosi materna possono soffrire di
granulomatosi infantisettica, che consiste nella diffusione in tutto il corpo di
granulomi piogeni, e di un ritardo dello sviluppo (U. S. Food and Drug
Administration, 2001). L’insorgenza delle patologie sopra menzionate è solitamente
preceduta da sintomi simili a quelli influenzali, inclusa febbre persistente; i sintomi
gastrointestinali quali nausea, vomito e diarrea possono precedere forme più gravi di
listeriosi o possono anche essere gli unici sintomi espressi; il tempo di insorgenza

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 77 -
dei sintomi gastrointestinali non è noto con esattezza, ma con ogni probabilità
supera le 12 ore (U. S. Food and Drug Administration, 2001). Uno studio di Wexler
e Oppenheim (1979) suggerì che la L. monocytogenes fosse l’unico, tra i batteri
Gram-positivi, a possedere dei lipopolisaccaridi che fungerebbero da endotossina,
ma ciò fu contraddetto da un successivo lavoro di Maitra et al. (1986).
La dose infettiva di L. monocytogenes varia a seconda del ceppo e della
sensibilità della vittima. Dai casi di patologia contratta attraverso l’assunzione di
latte crudo o in apparenza pastorizzato si potrebbe dedurre che, nei soggetti
maggiormente sensibili, anche solo un migliaio di batteri siano in grado di causarla
(U. S. Food and Drug Administration, 2001). La colonizzazione dell’organismo
umano inizia con l’invasione dell’epitelio gastrointestinale, da cui i microorganismi
penetrano nei monociti, nei macrofagi e nei leucociti polimorfonucleari dell’ospite,
colonizzando così il sangue (setticemia) e sviluppandosi; la loro presenza nei
fagociti consente inoltre l’accesso al cervello e probabilmente anche la migrazione
transplacentare verso il feto nelle donne in stato di gravidanza; la patogenesi si basa
insomma essenzialmente sulla capacità della L. monocytogenes di sopravvivere e
moltiplicarsi nelle cellule fagocitarie dell’organismo ospite. L’insorgenza di
meningiti listeriosiche può essere mortale fin nel 70% dei casi, la setticemia nel
50%, le infezioni perinatali e neonatali in oltre l’80%; nelle infezioni contratte
durante la gravidanza, la madre in genere sopravvive (U. S. Food and Drug
Administration, 2001).
Il fatto che la L. monocytogenes sia un parassita intracellulare la rendono
potenzialmente interessante come vettore di geni; per esempio, è recente lo studio
sull’utilizzo come vaccino vivo attenuato (detto “Lovaxin C”) contro il carcinoma
della cervice (Lowry, 2008).
La L. monocytogenes è un batterio particolarmente tenace e resiste benissimo al
congelamento, all’essiccamento e al calore, condizioni di norma deleterie per batteri
asporigeni (U. S. Food and Drug Administration, 2001). Esso è stato associato ad
alimenti quali il latte crudo, il latte fluido pastorizzato (Fleming et al., 1985), i

CAPITOLO 2 – I FONDAMENTI SCIENTIFICI
- 78 -
formaggi (in particolare quelli molli o freschi), il gelato, le verdure crude, le salsicce
preparate con carne cruda, il pollame sia crudo che cotto, le carni crude in genere, il
pesce sia crudo che affumicato; la sua capacità di crescere a temperature basse anche
fino a 0°C rende vana o comunque poco utile la refrigerazione dei cibi (U. S. Food
and Drug Administration, 2001). Addirittura a temperature prossime ai 4°C la
presenza di ioni Fe+++ pare favorire la crescita della L. monocytogenes (Dykes e
Dworaczek, 2002).
I metodi di analisi degli alimenti rispetto alla contaminazione da
L. monocytogenes sono lunghi (possono durare anche una settimana) e complessi,
anche se ultimamente si stanno sviluppando nuove metodologie che consentono di
accorciare notevolmente i tempi (BIO-RAD, 2009).

CAPITOLO 3


CAPITOLO 3 – LA SPERIMENTAZIONE
- 81 -
3.1 Il progetto sperimentale
Se si osserva il fenomeno della crescita batterica in un ambiente chiuso (tipo
“batch”) caratterizzato dalla presenza di idonee quantità degli opportuni nutrienti e
da una temperatura appropriata, si rileva come l’andamento nel tempo della
numerosità batterica (relativa ai batteri vivi) presenti un andamento caratteristico,
che si ripete in maniera similare per le diverse specie (Madigan e Martinko, 2007).
Tale tipico andamento, che è schematizzato in Fig. 3.1, viene comunemente
chiamato “curva di crescita”, ed è la rappresentazione grafica del processo cinetico
globale di crescita della popolazione, che è risultante delle due cinetiche
contrapposte di moltiplicazione cellulare e di morte; come si evince dalla Fig. 3.1, la
curva di crescita presenta quattro fasi fondamentali, tra loro sequenziali e facilmente
distinguibili, che sono descritte di seguito.
a) Fase di latenza (o “fase lag”) (1 in Fig. 3.1). Ha inizio con la semina, e
prelude alla moltiplicazione batterica: infatti, durante tale periodo, i
microorganismi non si moltiplicano, ma attivano i meccanismi che
consentono loro di adattarsi alle condizioni del nuovo ambiente e ai
principi nutritivi in esso presenti; si sviluppano i componenti cellulari
necessari per la divisione cellulare; la durata della fase di latenza
dipende da diversi fattori, quali la composizione del substrato, la
temperatura, le caratteristiche dell’atmosfera di incubazione, lo stato
fisiologico delle cellule inoculate; in linea generale, trattandosi di una
fase di adattamento, la sua durata è tanto inferiore quanto più le
condizioni del nuovo ambiente sono simili a quelle dell’ambiente di
provenienza delle cellule.
b) Fase di crescita esponenziale (o logaritmica o illimitata o “fase log”) (2
in Fig. 3.1). Durante questa fase le cellule cominciano a moltiplicarsi;
poiché lo fanno mediante un processo di divisione binaria, ad ogni
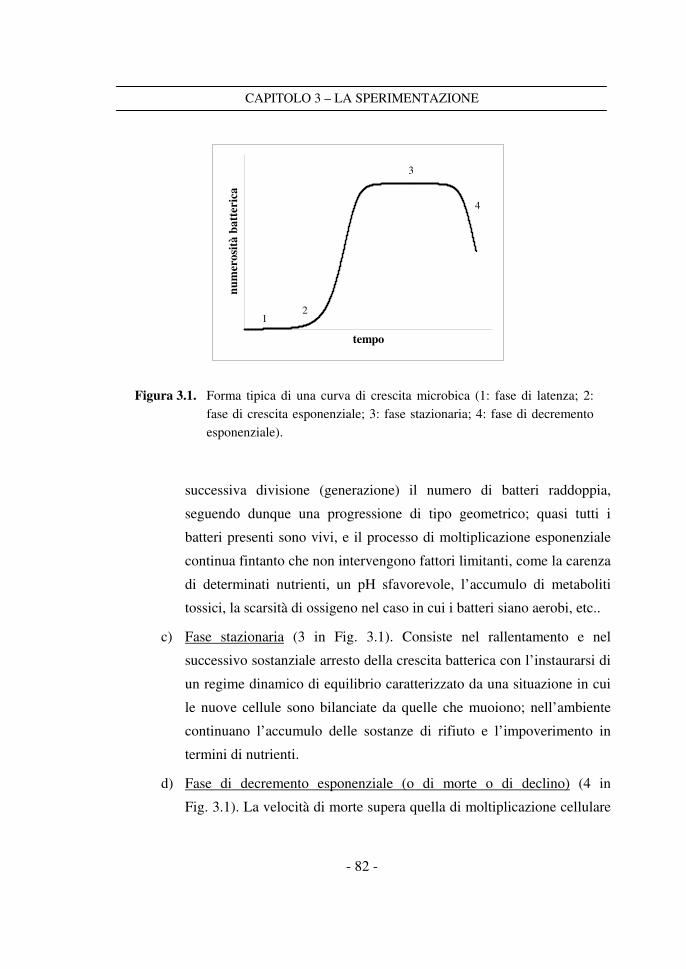
CAPITOLO 3 – LA SPERIMENTAZIONE
- 82 -
successiva divisione (generazione) il numero di batteri raddoppia,
seguendo dunque una progressione di tipo geometrico; quasi tutti i
batteri presenti sono vivi, e il processo di moltiplicazione esponenziale
continua fintanto che non intervengono fattori limitanti, come la carenza
di determinati nutrienti, un pH sfavorevole, l’accumulo di metaboliti
tossici, la scarsità di ossigeno nel caso in cui i batteri siano aerobi, etc..
c) Fase stazionaria (3 in Fig. 3.1). Consiste nel rallentamento e nel
successivo sostanziale arresto della crescita batterica con l’instaurarsi di
un regime dinamico di equilibrio caratterizzato da una situazione in cui
le nuove cellule sono bilanciate da quelle che muoiono; nell’ambiente
continuano l’accumulo delle sostanze di rifiuto e l’impoverimento in
termini di nutrienti.
d) Fase di decremento esponenziale (o di morte o di declino) (4 in
Fig. 3.1). La velocità di morte supera quella di moltiplicazione cellulare
tempo
nu
mer
osi
tà b
att
eric
a
Figura 3.1. Forma tipica di una curva di crescita microbica (1: fase di latenza; 2: fase di crescita esponenziale; 3: fase stazionaria; 4: fase di decremento esponenziale).
1 2
3
4

CAPITOLO 3 – LA SPERIMENTAZIONE
- 83 -
e la popolazione decresce rapidamente fino alla sua completa estinzione,
che sopraggiunge o per il completo esaurimento dei nutrienti e per il
consecutivo esaurimento dell’energia cellulare di riserva, oppure per
l’instaurarsi di condizioni ambientali quali un pH sfavorevole o
un’eccessiva concentrazione di prodotti (cataboliti) tossici.
A parte la forma simile della curva di crescita caratteristica delle diverse specie
di microorganismi, la variabile che genera le distinzioni più marcate tra le specie e,
per una stessa specie, tra le differenti condizioni ambientali, è il tempo, in quanto
questo può variare, per il completamento delle quattro fasi, da poche ore a diverse
settimane (Madigan e Martinko, 2007).
L’idea centrale sulla quale si fonda il progetto sperimentale oggetto del presente
lavoro di tesi è quella di valutare se, e in quale misura, vi possa essere un’influenza
delle microonde a bassa potenza, in generale, sulla crescita microbica, e in
particolare su quella di specie interessate alle problematiche relative alla sicurezza
alimentare, di cui si è parlato nel paragrafo 1.2. Infatti, come è stato lì ampiamente
discusso, la riduzione della carica batterica presente negli alimenti e il controllo
della crescita batterica sono due punti cardine dei trattamenti dell’industria
alimentare, le cui principali finalità sono, in linea generale, l’estensione della vita
commerciale dei prodotti e la minimizzazione del rischio per i consumatori. La
motivazione del ricorso a basse potenze, invece, risiede nella specifica intenzione di
concentrare l’indagine sperimentale sugli effetti non termici indotti dalle microonde,
la cui possibile esistenza è, per quanto in maniera forse non del tutto univoca,
ampiamente documentata in letteratura (si veda a tal proposito il paragrafo 2.4 e i
relativi sottoparagrafi): solo l’utilizzo di potenze sufficientemente basse,
congiuntamente ad un efficace sistema di controllo della temperatura, può infatti
consentire la migliore possibilità di discriminazione tra gli effetti non termici
ipotizzati e quelli termici, che sono invece ben noti; e si è già discusso (paragrafo
1.5) di quali potrebbero essere, potenzialmente, le ricadute future dei trattamenti non
termici, per cui non vi si tornerà nuovamente ora.

CAPITOLO 3 – LA SPERIMENTAZIONE
- 84 -
Come temperatura alla quale condurre la sperimentazione programmata è stata
scelta quella di 37°C in quanto, come detto al paragrafo 1.2, si tratta di una
temperatura prossima a quella ottimale di crescita per la maggior parte delle specie
microbiche patogene che possono dar luogo a contaminazioni alimentari, nonché per
quelle effettivamente esaminate; questo elemento sortisce anche l’effetto, dal punto
di vista sperimentale, di esaltare le eventuali differenze tra la velocità di crescita in
presenza ed in assenza delle microonde, in quanto la temperatura non è fattore
limitante della crescita stessa, e di contenere, allo stesso tempo, l’influenza di
eventuali errori dovuti al controllo della temperatura mediante termostatazione, in
quanto la temperatura ottimale di crescita individua anche la zona nella quale la
sensitività della velocità di crescita rispetto alla temperatura è minima (vedi Fig.
1.1). Inoltre, altro fatto non trascurabile, essendo 37°C la temperatura media
corporea dell’essere umano, essa è la massima alla quale è soggetto l’alimento nel
suo percorso dalla produzione alla digestione e assimilazione, qualora non sia
sottoposto a trattamenti termici: ipotizzare dunque un trattamento a 37°C non
comporterebbe nell’alimento perdite di proprietà nutrizionali superiori a quelle che
comporta la sua stessa ingestione. La temperatura qui fissata è la stessa dei lavori di
Carta e Desogus (2006; 2007; 2008b; 2008c; 2009) e di Carta et al. (2006a; 2006b).
Occorre, in conclusione, precisare che poco o nulla si potrà dire sugli eventuali
meccanismi di azione delle microonde, viste la varietà e la complessità delle
possibili interazioni con gli organismi biologici (si vedano a tal proposito i paragrafi
2.3 e 2.4), solo a partire da informazioni di tipo cinetico, né, molto probabilmente, si
potrebbe avere la certezza che l’eventuale azione non termica delle microonde sia
svolta direttamente sulle strutture cellulari in senso stretto, piuttosto che su enzimi o
ioni.

CAPITOLO 3 – LA SPERIMENTAZIONE
- 85 -
3.2 Le necessità della sperimentazione
In base a quanto detto nel precedente paragrafo 3.1, nella predisposizione della
struttura sperimentale è stato necessario risolvere alcuni problemi per poter
rispettare altrettante precise necessità. Il primo di questi è stato sicuramente quello di
come mettere in contatto tra loro microorganismi e microonde in modo controllato, e
ciò ha indirizzato la scelta verso un sistema circolante che potesse in qualche modo
consentire tempi di esposizione simili per le cellule presenti nella coltura. Il tutto
doveva chiaramente poter avvenire per tempi sufficienti almeno al completamento
della fase di crescita esponenziale, e dunque nell’ordine delle diverse ore (da 5 a 10),
mantenendo la temperatura, per tutta la durata della prova, costante al valore
prescelto, e garantendo condizioni di “sterilità” rispetto all’ambiente in entrambi i
sensi, cioè evitando contemporaneamente la penetrazione all’interno dell’ambiente
di reazione di microorganismi presenti all’esterno e lo spargimento nell’ambiente
dei microorganismi presenti nella miscela reagente; quest’ultima condizione è stata
ottenuta predisponendo una struttura di circolazione che potesse essere
preliminarmente ed interamente sottoposta a sterilizzazione, per poi rimanere
completamente chiusa sia prima che durante la prova (ad eccezione del momento
dell’inoculazione). Il problema delle determinazioni analitiche, poi, è strettamente
legato al fatto che il sistema reagente dovrebbe avere il minor numero possibile di
contatti con l’esterno, per cui si è preferito propendere per l’analisi in continuo della
miscela, senza l’effettuazione di campionamenti.
L’apparato sperimentale e le procedure connesse con il suo utilizzo sono state
dunque effettivamente concepite e realizzate per essere in grado di operare nelle
condizioni precedentemente esposte; i particolari tecnici sono contenuti nei paragrafi
successivi, e sono in parte i medesimi già presentati da Carta e Desogus (2006;
2007; 2008b; 2008c; 2009) e da Carta et al. (2006b).

CAPITOLO 3 – LA SPERIMENTAZIONE
- 86 -
3.3 L’apparato sperimentale
3.3.1 La “parte biologica”
La struttura sperimentale adottata, esemplificata in Fig. 3.2, prevede la
circolazione in continuo di sospensioni batteriche opportunamente preparate e
sottoposte a differenti condizioni di irradiazione. Il punto focale della struttura è
costituito da un reattore biologico posizionato sulla parte interna di una piastra di
acciaio a sua volta fissata, tramite una flangia, sulla parte terminale di una guida
d’onda; il reattore così concepito costituisce il “punto di contatto” tra il materiale
vivente e il campo elettromagnetico, contatto che può avvenire dunque in condizioni
sperimentali controllate e in sicurezza per gli operatori; la struttura costituita dalla
piastra e dal reattore è schematizzata in Fig. 3.3.
7
4
5
1 2
3
6
8
9
10
11
12
Figura 3.2. Schema dell’apparato sperimentale proposto (1: aria esterna; 2: filtro ceramico per l’aria esterna; 3: pompa peristaltica; 4: tappo; 5: bottiglia contenente la sospensione da irradiare; 6: reattore dipo PFR dove la sospensione batterica viene irradiata; 7: guida d’onda; 8: radiazione incidente; 9: piastra di supporto del reattore posta a chiusura della guida d’onda; 10: prelievo della sospensione batterica dalla bottiglia; 11: ingresso della sospensione batterica nella bottiglia; 12: sistema di analisi spettrofotometrica).

CAPITOLO 3 – LA SPERIMENTAZIONE
- 87 -
La sospensione batterica è contenuta in una bottiglia di pirex (5 in Fig. 3.2),
avente la funzione di serbatoio polmone, che viene riempita con la soluzione
nutriente per poi essere chiusa ermeticamente con un tappo in PVC termicamente
resistente (4 in Fig. 3.2) e sterilizzata. Il tappo è fornito di quattro passaggi con le
seguenti funzioni:
• insufflazione di aria esterna (1 in Fig. 3.2) passante attraverso un filtro
ceramico di porosità pari a 22 µm (2 in Fig. 3.2);
• uscita dalla bottiglia della sospensione reagente (10 in Fig. 3.2) per
essere inviata al reattore irradiato (6 in Fig. 3.2);
• ingresso della sospensione reagente (11 in Fig. 3.2) proveniente dal
reattore e dal sistema di analisi spettrofotometrica (12 in Fig. 3.2);
Figura 3.3. Schema costruttivo della piastra di chiusura della guida d’onda e di supporto del reattore (a: vista dall’alto; b: vista frontale).
a
b

CAPITOLO 3 – LA SPERIMENTAZIONE
- 88 -
• uscita dell’aria in eccesso utilizzata per l’ossigenazione della
sospensione reagente, fornita di un tubicino ad “U” chiuso
all’estremità con un tappo in cotone; tale sistema è stato preferito al
filtro ceramico in quanto questo sarebbe stato facilmente ostruito dalla
condensazione, sulla sua superficie filtrante, dell’umidità; la
particolare configurazione ad “U”, invece, oltre ad impedire che aria
esterna eventualmente richiamata all’interno della bottiglia possa
causare contaminazioni, permette di evitare la ricaduta, nella
sospensione, sia di filamenti di cotone che di goccioline di condensa.
Il fluido reagente contenuto nella bottiglia, qui mantenuto a temperatura costante
mediante termostato ad acqua, viene aspirato e messo in circolo mediante una
pompa peristaltica (3 in Fig. 3.2) con una portata di 40 ml min-1. Il circuito è stato
realizzato con tubi in silicone (ID 2 mm; OD 4 mm) e, in corrispondenza delle
congiunzioni, di acciaio inox (ID 2 mm). Il fluido giunge così nel reattore tubolare,
di lunghezza pari a 7,6 cm, realizzato anch’esso in silicone (ID 4 mm; OD 6 mm), e
posizionato, come detto, nella parte irradiata di una piastra in acciaio inox (9 in
Fig. 3.2) che chiude una guida d’onda (7 in Fig. 3.2); le dimensioni del reattore e la
portata hanno fatto sì che il tempo di permanenza della sospensione all’interno del
reattore sia stato, per tutte le prove effettuate, di circa 1,4 s. Il silicone è stato scelto
per le sue caratteristiche di flessibilità e di trasparenza alle microonde (von Hippel,
1954), mentre i raccordi (metallici) sono stati realizzati utilizzando prodotti
Swagelok che hanno consentito il completo isolamento dall’ambiente esterno e la
sicurezza delle connessioni.
I terreni di coltura utilizzati sono stati:
• “Mueller Hinton Broth” (MH) della ditta Microbiol®, infuso
disidratato ottenuto da cuore di bue con addizione di idrolizzato acido
di caseina e amido, che veniva disciolto in acqua distillata nella

CAPITOLO 3 – LA SPERIMENTAZIONE
- 89 -
misura di 22 g per litro di acqua, e che è stato utilizzato per le specie
Bacillus clausii e Pseudomonas aeruginosa;
• “Brain Hearth Infusion Broth” (BHI) della ditta Microbiol®, infuso
disidratato ottenuto da cuore di bue e cervello di vitello in condizioni
controllate, che veniva disciolto in acqua distillata nella misura di 37 g
per litro di acqua, e che è stato utilizzato per le specie Staphylococcus
aureus e Listeria monocytogenes.
La procedura sperimentale adottata prevede le seguenti fasi:
• introduzione di 200 ml di soluzione nutriente nel contenitore in pirex,
chiusura di questo e collegamento del circuito;
• sterilizzazione dell’intero circuito attraversato dal fluido (costituito
dall’insieme degli elementi 4, 5, 6, 9, 10 e 11 di Fig. 3.2) in autoclave
per 20 minuti alla temperatura di 121°C;
• raffreddamento della soluzione e dell’intera struttura fino a
temperatura ambiente;
• inoculazione nella bottiglia, mediante siringa sterile, attraverso il foro
di ingresso dell’aria esterna, privato all’uopo del filtro ceramico in
condizioni asettiche per impedire contaminazioni esterne, di piccole
quantità (2-3 ml) della sospensione batterica precedentemente
preparata per lo scopo;
• azionamento della pompa di ricircolo ed, eventualmente, dell’apparato
di generazione delle microonde;
• misurazione della densità ottica ad intervalli di tempo prefissati,
eventuale prelievo di campioni in condizioni asettiche mediante
rubinetto inserito allo scopo nel circuito, registrazione dei dati
provenienti dalle misure e monitoraggio dell’evoluzione della prova

CAPITOLO 3 – LA SPERIMENTAZIONE
- 90 -
con controllo circa il mantenimento dei valori impostati delle variabili
di processo e circa l’andamento delle misure stesse di densità ottica;
• interruzione della circolazione del fluido, della generazione delle
microonde e della registrazione dei dati, risterilizzazione e lavaggio
del circuito.
Le sospensioni batteriche preparate per l’inoculazione sono state ottenute
secondo le seguenti modalità:
• i microorganismi di Bacillus clausii dal prodotto commerciale
Enterogermina® (Sanofi-synthelabo®), costituito da una sospensione
di spore del batterio in acqua purificata con un contenuto di 4.108
spore per millilitro di sospensione; 1 ml di tale sospensione, in seguito
ad agitazione meccanica, veniva addizionato a 4 ml di soluzione
nutriente, quindi i microorganismi venivano fatti sviluppare per 24 h a
37°;
• la Pseudomonas aeruginosa da un ceppo locale coltivato nei
laboratori del Dipartimento di Scienze e Tecnologie Biomediche
(sezione di Microbiologia Applicata e Tecnologie Biomediche)
dell’Università degli Studi di Cagliari; i microorganismi, prima
dell’inoculazione, sono stati fatti adattare in MH per tempi compresi
tra 24 e 48 h alla temperatura di 37°C, fino al raggiungimento di un
sufficiente grado di torbidità;
• i batteri di Staphylococcus aureus utilizzati sono del ceppo ATCC
25923; la fase di adattamento a 37°C, anche in questo caso, ha avuto
durate comprese tra 24 e 48 h, fino al raggiungimento di una
sufficiente torbidità;
• la Listeria monocytogenes da un ceppo locale coltivato nei laboratori
del Dipartimento di Scienze e Tecnologie Biomediche (sezione di

CAPITOLO 3 – LA SPERIMENTAZIONE
- 91 -
Microbiologia Applicata e Tecnologie Biomediche) dell’Università
degli Studi di Cagliari; i microorganismi, prima dell’inoculazione,
sono stati fatti adattare in BHI per tempi ancora compresi tra 24 e 48 h
e sempre alla temperatura di 37°C, fino al raggiungimento di un
sufficiente grado di torbidità.
Per ogni coltura, prima di procedere con l’inoculazione dei microorganismi nel
sistema reagente, è stata misurata la concentrazione batterica mediante conta
delle colonie su terreno solidificato “Mueller Hinton Agar”, anche allo scopo di
accertare l’assenza di contaminazioni da parte di specie estranee.
3.3.2 La “parte elettronica”
L’apparato di generazione delle microonde a bassa potenza, già presentato da Carta
e Desogus (2008b; 2008c; 2009), è costituito da diversi componenti elettronici. Il
principale di questi è un oscillatore del tipo “miniature permanent magnet YIG-
Tuned (Yttrium Iron Garnet)”, modello “Micro Lambda Wireless, Inc. MLOM-
0204”. Tale apparecchio produce un segnale in uscita caratterizzato da una potenza
di 14 dBm (250 mW), la cui frequenza può essere regolata nel campo compreso tra 2
e 4 GHz. L’oscillatore fa uso di un transistor bipolare accoppiato con una sfera YIG
in un circuito a film sottile. La regolazione della frequenza operativa è ottenuta
agendo su un segnale di corrente in ingresso all’oscillatore nel campo compreso tra
-100 e +100 mA; tale segnale viene prodotto da un circuito “driver” che converte un
segnale di tensione in uno di corrente, e che è provvisto, oltre al principale, anche di
un piedino secondario (con una sensibilità inferiore rispetto al principale),
utilizzabile per operare una regolazione più fine; il circuito “driver”, al fine di
regolare la frequenza tra 2 e 4 GHz, accetta segnali di input compresi tra 0 e 5 V di
corrente continua; la tensione in ingresso, a sua volta, viene generata per mezzo di

CAPITOLO 3 – LA SPERIMENTAZIONE
- 92 -
una scheda di acquisizione dati della National Instruments (modello PCI-MIO 16-
E4), gestita dal software Virtual Brench.
L’espressione che consente di ottenere la frequenza operativa ν in funzione della
tensione in ingresso V è stata ottenuta con misure della frequenza in uscita effettuate
mediante un analizzatore di spettro portatile (modello “Agilent 8563E option
006”, con campo di frequenza misurabile compreso tra 30 Hz e 26,5 GHz, massimo
livello di potenza di 1 W); si è determinato che tale funzione può essere espressa
dalla seguente relazione lineare (con un coefficiente di correlazione pari a 0,9999):
V0,42,0ν ⋅+= (3.1)
Continuando a seguire la direzione di propagazione del segnale, dopo
l’oscillatore è connesso un “DC Block” (Suhner 1100.01.A), seguito a sua volta da
un attenuatore fisso da 6 dB (JFW Industries 4AH-06), e poi da un attenuatore
rotante (JFW Industries 50R-248), che consente una ulteriore attenuazione della
potenza in uscita di una quantità variabile tra 0 e 10 dB, con un’ampiezza di passo di
1 dB.
Il segnale generato, una volta trattato per mezzo dei componenti appena
descritti, viene trasferito ad un amplificatore di potenza (Herotek P/N AP271135),
segnale di corrente per la regolazione della frequenza
1 2 3 4 5 6
7 8
alimentazione
alimentazione
Figura 3.4. Schema dell’apparato di generazione delle microonde (1: oscillatore; 2: attenuatore fisso; 3: attenuatore rotante; 4: amplificatore; 5: isolatore; 6: accoppiatore cavo – guida d’onda; 7: guida d’onda; 8: piastra di chiusura della guida d’onda supportante il reattore).

CAPITOLO 3 – LA SPERIMENTAZIONE
- 93 -
alimentato con una tensione di 12 V di corrente continua; l’amplificazione prodotta
è pari, in condizioni ottimali, a 23 dB, tuttavia l’efficienza diminuisce con
l’aumentare della temperatura del dispositivo, per cui si è reso necessario dotarlo di
un sistema di raffreddamento abbastanza efficiente, dotato di una sufficiente
superficie di scambio posta a contatto con aria movimentata per mezzo di una
ventola. Successivamente il segnale incontra un isolatore (RF & NC CI-200-172),
che ha la funzione di consentire il passaggio del segnale incidente impedendo al
contempo il passaggio del segnale riflesso, che danneggerebbe l’amplificatore e gli
altri componenti del circuito.
Il segnale proveniente dall’amplificatore e uscente dall’isolatore viene a questo
punto veicolato, attraverso un cavo coassiale, ad un accoppiatore cavo – guida
d’onda (MEC P/N LA 40-30-CH WR430), che converte il segnale in ingresso via
cavo e, tramite un’antenna, lo invia alla guida d’onda, del tipo WR430 (MEC P/N
LA 160-30N), con dimensioni della sezione trasversale interna di 109 × 55 mm, e
quindi, tramite questa, al reattore biologico: la parte terminale della guida d’onda è
infatti chiusa da una piastra in acciaio inossidabile sulla quale è posizionato il
reattore stesso (vedi paragrafo 3.3.1). Le misure della potenza incidente possono
essere effettuate utilizzando un misuratore “power meter” della casa Hewlett
Packard, modello 436A, che misura la potenza del segnale proveniente da un
accoppiatore direzionale (coefficiente di accoppiamento di 10-3) collegato alla guida
d’onda mediante un sensore di potenza della stessa Hewlett Packard, modello
8481A; quest’ultimo è in grado di misurare livelli di potenza compresi tra -30 dBm e
+20 dBm (corrispondenti rispettivamente a 1 µW e a 100 mW), a frequenze
comprese tra 10 MHz e 18 GHz; per quanto riguarda la misura della frazione di
potenza delle microonde effettivamente assorbita dal sistema reagente, necessaria
per la valutazione del SAR della sospensione irradiata, si rimanda al paragrafo 3.4.1.
In linea teorica la massima potenza del segnale trasmesso lungo la guida d’onda
potrebbe essere di 30 dBm (1000 mW) ma, per via dell’efficienza globale
dell’apparato, della compressione del segnale per opera dell’amplificatore, nonché

CAPITOLO 3 – LA SPERIMENTAZIONE
- 94 -
della resa dei singoli componenti, la massima potenza misurata è stata di circa 26
dBm (400 mW). Tutte le connessioni tra i componenti e fino all’adattatore coassiale
sono state realizzate mediante cavi del tipo RG316 con connettori del tipo SMA. Le
misure di potenza effettuate tramite il “power meter” possono essere registrate ad
intervalli di tempo prestabiliti grazie alla stessa scheda di acquisizione dati della
National Instruments (modello PCI-MIO 16-E4) utilizzata per la generazione del
segnale inviato al circuito “driver”, e mediante il software di gestione Virtual
Brench: questo consente di verificare, anche a posteriori, il mantenimento, per tutto
il tempo richiesto, del livello di potenza impostato.
3.4 Le determinazioni analitiche
3.4.1 La determinazione dello “Specific Absorption Rate”
Si è spiegato, nel paragrafo 3.3.2, il funzionamento dell’apparato di generazione
delle microonde, e come sia possibile regolare la potenza irradiata lungo la guida
d’onda; quest’ultima è comunque diversa da quella effettivamente assorbita dalle
sospensioni batteriche circolanti all’interno del reattore, in quanto vi è una parte di
potenza che viene dissipata come conseguenza essenzialmente della natura dei
materiali coinvolti e della particolare configurazione geometrica del sistema. La
potenza effettivamente assorbita dal sistema reagente altro non è, in realtà, che lo
“Specific Absorption Rate” (SAR), di cui si è già parlato nel paragrafo 2.4; se,
dunque, per ragioni di comodità, si farà riferimento nel seguito del presente lavoro
alla sola potenza irradiata, per questioni di completezza delle informazioni fornite,
oltre che per correttezza scientifica nel confronto con le informazioni presenti in
letteratura, si è ritenuto di determinare, in via preventiva, quale fosse la frazione
della potenza irradiata che viene effettivamente assorbita dal sistema.

CAPITOLO 3 – LA SPERIMENTAZIONE
- 95 -
Tale determinazione è stata effettuata partendo dalle considerazioni che
seguono. La variazione della temperatura del fluido circolante è dovuta a:
• potenza trasmessa dalla radiazione a microonde (di seguito indicata
con RQ& );
• potenza termica generata dalle forze di attrito dovute alla circolazione
del fluido (di seguito indicata con AQ& );
• calore scambiato dal sistema con l’esterno (di seguito indicato con
EQ& ).
Indicando dunque con m la massa totale di fluido contenuta all’interno del sistema,
con pc il suo calore specifico a pressione costante, con T la temperatura del fluido e
con t il tempo si può scrivere il seguente bilancio termico:
EARp QQQdt
dTmc &&& −+=⋅⋅ (3.2)
La relazione 3.2, se il sistema può essere considerato adiabatico ( 0QE =& ), diviene:
mc
dt
dT
p
AR
⋅
+=
&&
(3.3)
Ora, poiché m rimane costante, e tutte le altre grandezze che compaiono a destra
nella 3.3 possono essere ritenute costanti al passare del tempo (se le variazioni di
temperatura sono modeste), fintantoché il sistema continua ad essere all’incirca
adiabatico è da ritenersi che l’andamento della temperatura rispetto al tempo debba
essere di tipo lineare; integrando la 3.3 (avendo posto 0TT = in corrispondenza del
tempo 0tt = ) si ottiene:
( )0p
AR0 tt
mc
QQTT −⋅
⋅
++=
&&
(3.4)

CAPITOLO 3 – LA SPERIMENTAZIONE
- 96 -
Poiché si può a buona ragione ritenere che il termine AQ& , che dipende dalle
caratteristiche interne del circuito e da quelle del moto del fluido, non sia influenzato
dall’applicazione o meno del campo elettromagnetico, la relazione 3.4, nel caso
particolare di assenza di campo ( 0QR =& ), può essere riscritta nella forma che segue:
( )0p
A0NR tt
mc
QTT −⋅
⋅+=
&
(3.5)
Conoscendo dunque l’andamento della temperatura in assenza e in presenza di
irradiazione, se si indica con NRδ la pendenza della retta temperatura vs. tempo nel
primo caso, e con Rδ la pendenza della stessa retta nel secondo caso, è possibile
determinare quale sia l’effettiva potenza trasmessa dalle microonde al sistema;
infatti, attraverso alcuni semplici passaggi, si può giungere alla seguente relazione:
( ) mcδδQ pNRRR ⋅⋅−=& (3.6)
A questo punto, dal confronto tra il valore calcolato di RQ& e la potenza trasmessa
lungo la guida d’onda, è possibile sapere quale sia, in corrispondenza della potenza
di irradiazione, la frazione di quest’ultima che viene effettivamente assorbita dal
sistema reagente.
La procedura sperimentale adottata per la determinazione del SAR come
frazione della potenza irradiata è stata la seguente:
• rimozione dal circuito della parte dedicata all’analisi
spettrofotometrica (12 in Fig. 3.2) e riassemblaggio dello stesso;
• coibentazione di tutte le rimanenti parti del circuito mediante tubo
isolante commerciale, ad unica esclusione delle parti tecnicamente
non rivestibili (la porzione di tubo posizionata all’interno della pompa
peristaltica e la parte interna alla guida d’onda);
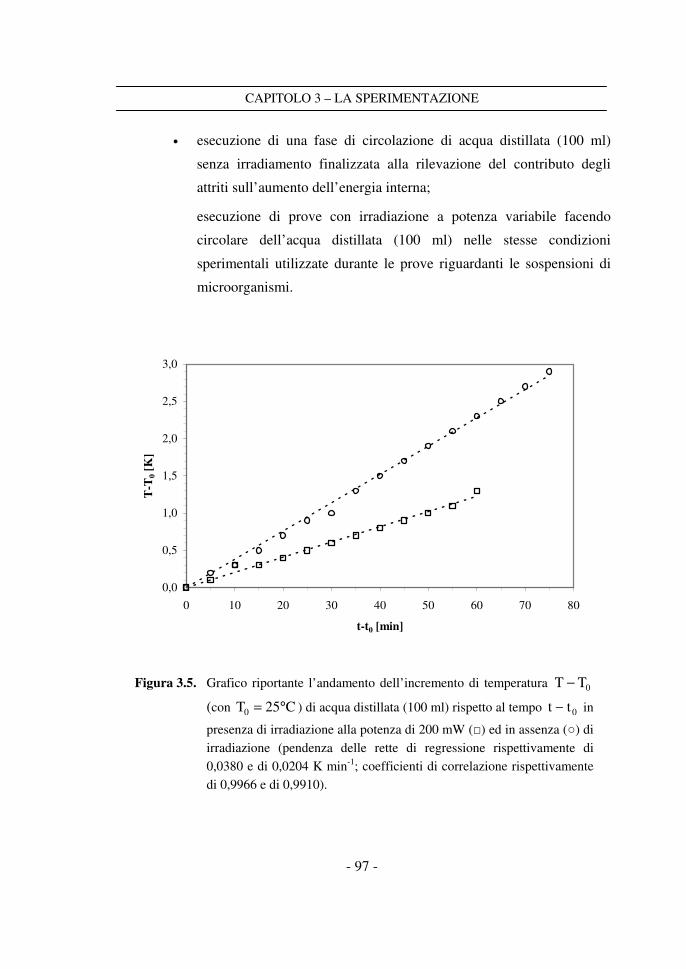
CAPITOLO 3 – LA SPERIMENTAZIONE
- 97 -
• esecuzione di una fase di circolazione di acqua distillata (100 ml)
senza irradiamento finalizzata alla rilevazione del contributo degli
attriti sull’aumento dell’energia interna;
esecuzione di prove con irradiazione a potenza variabile facendo
circolare dell’acqua distillata (100 ml) nelle stesse condizioni
sperimentali utilizzate durante le prove riguardanti le sospensioni di
microorganismi.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
0 10 20 30 40 50 60 70 80
t-t0 [min]
T-T
0 [
K]
Figura 3.5. Grafico riportante l’andamento dell’incremento di temperatura 0TT −
(con C25T0 °= ) di acqua distillata (100 ml) rispetto al tempo 0tt − in
presenza di irradiazione alla potenza di 200 mW (□) ed in assenza (○) di irradiazione (pendenza delle rette di regressione rispettivamente di 0,0380 e di 0,0204 K min-1; coefficienti di correlazione rispettivamente di 0,9966 e di 0,9910).

CAPITOLO 3 – LA SPERIMENTAZIONE
- 98 -
Per mezzo di un sensore di termometro digitale (Hanna Instruments HI 92710C)
introdotto nel contenitore (5 in Fig. 3.2) attraverso uno dei fori presenti nel tappo (4
in Fig. 3.2) è stato possibile rilevare e registrare la temperatura del fluido all’interno
della bottiglia al trascorrere del tempo.
In linea generale si è riscontrato l’atteso andamento lineare della temperatura in
funzione del tempo; a puro titolo di esempio si riporta in Fig. 3.5 il grafico che
delinea l’andamento dell’incremento di temperatura ( 0TT − ) rispetto al tempo
( 0tt − ) ottenuto con una potenza nominale di 200 mW a confronto con il caso in
assenza di irradiazione, mentre in Tabella 3.1 sono contenute le pendenze delle rette
temperatura vs. tempo ricavate sperimentalmente ( δ ) come media di tre prove per
ogni potenza nominale, e la percentuale di potenza effettivamente assorbita rispetto
a quella nominale. A conti fatti, si può dire che le condizioni sperimentali adottate
durante le prove oggetto del presente lavoro di tesi hanno visto valori di SAR
compresi all’incirca tra 3.10-4 e 10-3 W/g.
P [mW] δδδδ [K min-1] P%
0 0,0220 -
100 0,0264 61,3
200 0,0309 62,0
300 0,0338 54,8
400 0,0363 49,9
Tabella 3.1. Risultati sperimentali delle prove condotte per la determinazione della
frazione di potenza assorbita dal sistema (P: potenza nominale trasmessa
lungo la guida d’onda; δ : valore medio della pendenza delle rette
temperatura vs. tempo; P%: percentuale della potenza effettivamente assorbita rispetto a quella nominale).

CAPITOLO 3 – LA SPERIMENTAZIONE
- 99 -
3.4.2 La misurazione della densità ottica
Dato un campione attraversato da un fascio di luce monocromatica con
lunghezza d’onda fissata, si definisce trasmittanza (Tr) il seguente rapporto:
0
1
I
ITr = (3.7)
dove 0I rappresenta l’intensità della luce incidente, mentre 1I l’intensità della luce
trasmessa che emerge dal campione. Secondo la legge di Beer–Lambert (o di Beer–
Lambert–Bouguer), la trasmittanza è legata alla natura chimica e allo spessore del
mezzo attraversato secondo la relazione:
l-kλ10Tr ⋅= (3.8)
dove λk è una costante caratteristica del mezzo, valida alla lunghezza d’onda λ ,
detta coefficiente di estinzione, mentre l rappresenta il cammino ottico. La densità
ottica (OD), detta anche assorbanza, corrisponde al logaritmo decimale dell’inverso
della trasmittanza:
=
=
1
0
I
ILog
Tr
1LogOD (3.9)
Dunque, considerando l’espressione della trasmittanza secondo la relazione 3.8, la
densità ottica può essere espressa dalla relazione lineare:
lkOD λ ⋅= (3.10)
Nel caso in cui il mezzo attraversato sia costituito da una soluzione omogenea, di
norma la relazione 3.10 viene scritta nella forma seguente:
ClεOD λ ⋅⋅= (3.11)
dove C è la concentrazione molare del soluto e λε il coefficiente di assorbimento
molare; quest’ultimo è considerato una costante che dipende ancora dalla natura del

CAPITOLO 3 – LA SPERIMENTAZIONE
- 100 -
mezzo e dalla lunghezza d’onda della luce incidente, benché in effetti possa subire
lievi variazioni in funzione della temperatura; sulla relazione 3.11 si basa l’analisi
chimica quantitativa per spettrofotometria. In effetti tale relazione è valida solo in
determinati intervalli di concentrazione, in quanto al di fuori di questi la linearità tra
densità ottica e concentrazione può essere inficiata da particolari fenomeni chimico-
fisici, come per esempio la precipitazione del soluto.
Nel caso in cui si intenda utilizzare l’analisi spettrofotometrica per miscele
(sarebbe a questo punto improprio parlare di soluzioni) eterogenee (che è il caso
delle sospensioni batteriche in un terreno di coltura liquido), come anche messo in
luce da Ageno (1992), la tecnica (almeno entro certi limiti) conserva la sua validità
(è sufficiente intendere con C la concentrazione delle particelle disperse), ma
occorre tenere presente che la densità ottica rilevata è affetta da un errore che è
proporzionale al rapporto tra la dimensione delle particelle che assorbono la
radiazione incidente (nel nostro si tratta di batteri, che hanno dimensioni
caratteristiche nell’ordine di qualche micrometro) e la lunghezza d’onda della
radiazione (che è in genere nell’ordine di qualche decimo di micrometro, e che nelle
prove sperimentali condotte è stata fissata al valore di 0,600 µm). Questo fatto si
spiega considerando che la differenza tra 0I e 1I è costituita dall’intensità luminosa
non trasmessa e dunque diffusa, per cui il rapporto tra l’intensità della luce diffusa e
l’intensità della luce incidente rappresenta la frazione di intensità luminosa diffusa
rispetto a quella incidente. Poiché la luce diffusa non è direzionale, una frazione α di
questa raggiungerà comunque il sensore dello strumento nonostante non sia luce
trasmessa; se dunque si indica con appOD il valore della densità ottica
effettivamente rilevato dallo strumento, in luogo della relazione 3.9, si ha che:
( )
−⋅+=
Tr1αTr
1LogODapp (3.12)
Il termine destro della 3.12 può evidentemente essere scomposto ottenendo:

CAPITOLO 3 – LA SPERIMENTAZIONE
- 101 -
( )
−⋅++
=
Tr1αTr
TrLog
Tr
1LogODapp (3.13)
Considerando a questo punto la 3.9, si può esprimere lo scostamento tra OD e
appOD come:
( )
( )
−⋅+=
−⋅+=
Tr
Tr1α1Log
Tr1αTr
Tr-LogOD-OD app (3.14)
Dalla relazione 3.14 si evince come l’errore, che si configura come una sottostima
della densità ottica in quanto non può essere negativo, sia piccolo se la trasmittanza
è grande, e dunque per bassi valori della densità ottica; al crescere di quest’ultima,
invece, diminuisce Tr, e di conseguenza l’errore cresce. Tuttavia, non essendo il
coefficiente α determinabile a priori (esso può dipendere da diversi fattori
difficilmente individuabili), si è ritenuto di procedere preliminarmente, per ogni
specie batterica utilizzata, alla quantificazione empirica dell’errore commesso
all’interno del campo di valori di densità ottica misurati durante le successive prove
sperimentali. Per fare questo, per ogni specie batterica, partendo da una soluzione
molto concentrata, ottenuta alla fine di un processo di crescita (e dunque alle
massime concentrazioni ottenibili durante le prove sperimentali), si è proceduto con
diluizioni successive, misurando volta per volta la densità ottica; si è così riscontrato
come, in tutti i casi esaminati, la differenza tra densità ottica effettiva ed apparente
fosse del tutto irrilevante, inferiore all’1%. Questo passaggio, fondamentale per la
validazione delle misurazioni di densità ottica che sono state poi effettuate, ha
consentito di appurare come all’interno del campo di valori di densità ottica propri
delle prove effettuate per il presente lavoro di tesi l’errore esplicitato dalla relazione
3.14 sia stato del tutto trascurabile; tale constatazione è inoltre giunta a conferma di
precedenti rilevazioni, pur su specie batteriche differenti, dello stesso Ageno (1992).
Nello specifico, le analisi sono state condotte utilizzando uno spettrofotometro
(Varian Cary 50) inserito in linea nel circuito; ciò è stato ottenuto mediante una

CAPITOLO 3 – LA SPERIMENTAZIONE
- 102 -
cuvette a flusso in pirex (dunque sterilizzabile) appositamente realizzata per lo
scopo, di cui in Fig. 3.6 è riportato lo schema realizzativo. Il fluido, introdotto
attraverso l’apposito tubicino di ingresso (1 in Fig. 3.6) direttamente sul fondo, è
così costretto a percorrere la zona interessata dalla lettura spettrofotometrica dal
basso verso l’alto, per poi imboccare l’uscita (2 in Fig 3.6); tale configurazione
consente così di sfavorire la sedimentazione degli aggregati cellulari presenti nella
miscela circolante, e di favorire invece l’omogeneizzazione della stessa; l’aver
dotato inoltre la cuvette di un tappo apribile (3 in Fig. 3.6) consente l’effettuazione
di una pulizia accurata al termine di ciascuna prova.
Lo spettrofotometro viene gestito tramite l’apposito software fornito dal
costruttore che consente, oltre che di fissare la lunghezza d’onda del fascio di luce
1 2
3
Figura 3.6. Schema costruttivo della cuvette a flusso utilizzata per le analisi spettrofotometriche (1: ingresso del fluido; 2: uscita del fluido; 3: tappo ermetico sigillante).
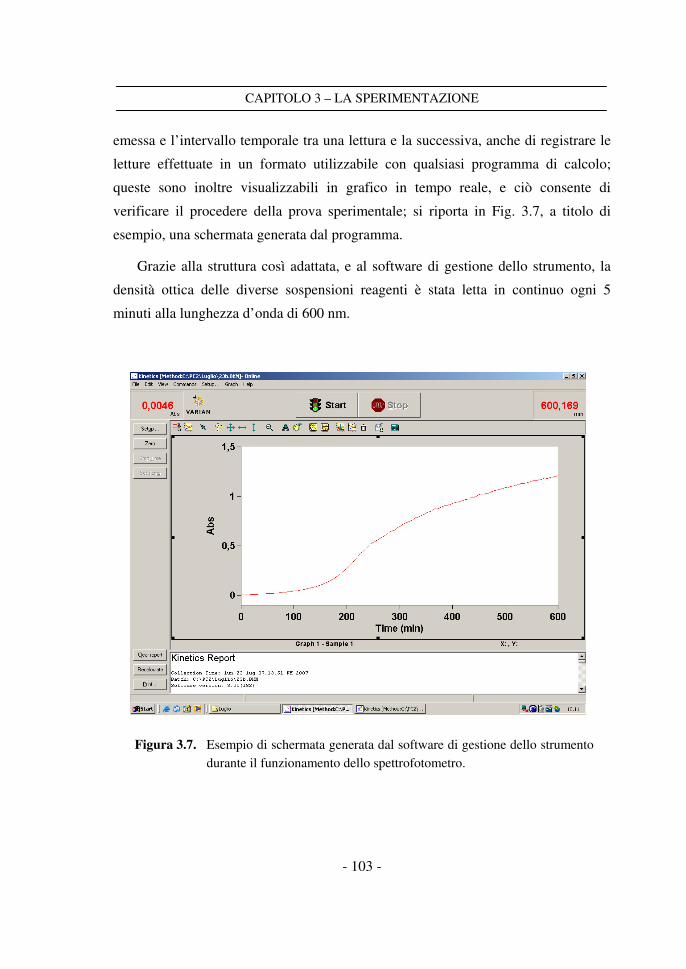
CAPITOLO 3 – LA SPERIMENTAZIONE
- 103 -
emessa e l’intervallo temporale tra una lettura e la successiva, anche di registrare le
letture effettuate in un formato utilizzabile con qualsiasi programma di calcolo;
queste sono inoltre visualizzabili in grafico in tempo reale, e ciò consente di
verificare il procedere della prova sperimentale; si riporta in Fig. 3.7, a titolo di
esempio, una schermata generata dal programma.
Grazie alla struttura così adattata, e al software di gestione dello strumento, la
densità ottica delle diverse sospensioni reagenti è stata letta in continuo ogni 5
minuti alla lunghezza d’onda di 600 nm.
Figura 3.7. Esempio di schermata generata dal software di gestione dello strumento durante il funzionamento dello spettrofotometro.

CAPITOLO 3 – LA SPERIMENTAZIONE
- 104 -
3.4.3 La conta in piastra
La crescita di una data popolazione batterica può essere misurata seguendo nel
tempo la variazione della massa cellulare totale (come avviene nel caso della misura
della densità ottica) o del numero di cellule; esistono a tal proposito parecchi metodi
per determinare la numerosità cellulare e la scelta del metodo da utilizzare dipende
essenzialmente dal tipo di microorganismo e dal problema che si intende affrontare
(Madigan e Martinko, 2007).
Tralasciando tutti i metodi di stima indiretta (applicabili solo in specifici casi e
con l’ausilio di strumentazioni spesso costose, e dunque non di interesse per il
presente lavoro sperimentale), la tecnica forse più immediata è quella della conta
diretta, che viene effettuata contando al microscopio le cellule presenti in un
campione essiccato su un vetrino o, in alternativa, in un campione liquido; tale
tecnica presenta tuttavia alcuni limiti, che consistono essenzialmente
nell’impossibilità di distinguere tra cellule vive e cellule morte (si tratta di un
metodo di conta totale), nella difficoltà di individuare cellule molto piccole, che
potrebbero pertanto non essere conteggiate, e nella oggettiva difficoltà di utilizzarla
per sospensioni a bassa densità cellulare. In molti casi, tra cui il nostro, perciò, si
preferiscono tecniche differenti, dette di conta vitale, che consentono di contare tutte
e solo le cellule vive. È vitale una cellula capace di dividersi e dare progenie, e il
metodo più usuale per ottenere una conta vitale di un campione consiste nel
determinare il numero di cellule presenti che sono capaci di formare colonie su un
opportuno terreno agarizzato; per questo motivo la conta vitale viene anche chiamata
conta in piastra o conta delle colonie, in quanto il presupposto di questo tipo di
procedura è che ogni colonia sia originata da una singola cellula vitale. Vi sono due
metodi per effettuare la conta in piastra: il piastramento in superficie e il
piastramento per inclusione. Nel piastramento in superficie, un volume noto
(solitamente 0,1 ml o anche meno) del campione opportunamente diluito viene
distribuito sulla superficie di una piastra di terreno agarizzato con una spatola di

CAPITOLO 3 – LA SPERIMENTAZIONE
- 105 -
vetro sterile; la piastra viene poi tenuta in incubazione fino alla comparsa delle
colonie, che possono così essere contate; il volume del campione distribuito sulla
piastra deve essere molto piccolo per evitare che il liquido in eccesso non si adsorba
sulla superficie della piastra e possa provocare la fusione di colonie diverse
rendendo difficile il conteggio. Nel piastramento per inclusione un volume noto (di
solito compreso tra 0,1 e 1 ml) di campione viene inoculato mediante pipetta in una
capsula Petri sterile; viene poi aggiunto il terreno agarizzato fuso e il tutto viene
miscelato facendo ruotare delicatamente la piastra; dato che il campione viene
mescolato con il terreno agarizzato è possibile utilizzare un volume maggiore di
quello usato nel piastramento in superficie, ma è tuttavia necessario essere certi che i
microorganismi che devono essere inclusi nell’agar siano in grado di sopportare per
breve tempo la temperatura dell’agar fuso, che è di 45ºC. In entrambe le tecniche è
importante che il numero di colonie che si sviluppano sulla piastra non sia troppo
elevato, in quanto l’eccessivo affollamento impedisce di fatto ad alcune cellule di
formare colonie, e il conteggio rischierebbe quindi di sottostimare la numerosità
batterica; è però allo stesso tempo essenziale che il numero di colonie non sia troppo
piccolo, perché ciò inficierebbe la significatività statistica della conta; la pratica
corrente consiste perciò nel contare le colonie solo nelle piastre che contengono un
numero di colonie compreso tra 30 e 300. Poiché, dunque, per ottenere un numero di
colonie appropriato per il conteggio, il campione deve quasi sempre essere
preventivamente diluito, e visto che, se non in rari casi, si conosce in anticipo, anche
approssimativamente, il numero delle cellule vitali presenti nel campione, la prassi
vuole che si effettui, per lo stesso campione, più di una diluizione. I risultati della
conta vitale vengono solitamente espressi come numero di UFC (acronimo di Unità
Formanti Colonia, in inglese CFU, che sta per “Colony-Forming Units”) per unità di
volume del campione.
Nonostante le imprecisioni intrinseche alla metodologia della conta vitale,
questa tecnica permette di ottenere, relativamente al numero di cellule vitali, le
migliori informazioni possibili, ed è per questo ampiamente utilizzata nell’ambito

CAPITOLO 3 – LA SPERIMENTAZIONE
- 106 -
della microbiologia, e in particolare nei settori della microbiologia alimentare,
casearia, medica e delle acque: il metodo, infatti, ha comunque il vantaggio di essere
molto sensibile, considerato che può essere applicato anche a campioni contenenti
pochissime cellule (al fine, per esempio, di individuare una contaminazione
microbica di piccola entità nei materiali in esame); inoltre, utilizzando terreni
colturali selettivi è possibile contare solo particolari gruppi di microorganismi
all’interno di una popolazione mista.
Per quanto riguarda il presente lavoro di tesi, la conta in piastra è stata effettuata
solo per una parte delle prove effettuate, al fine di verificare la corrispondenza tra le
informazioni ottenute dalla misurazione della concentrazione della biomassa totale
(ottenuta mediante analisi spettrofotometrica) e quelle ottenibili dalla misurazione
della concentrazione di microorganismi vivi; ma di questo si tratterà nel paragrafo
3.4.4; altro importante fatto è che la conta in piastra ha consentito di verificare
l’assenza di contaminazioni da parte di specie microbiche estranee all’interno delle
miscele reagenti. La conta batterica in piastra è stata effettuata, con l’ausilio delle
strutture presenti nei laboratori del Dipartimento di Scienze e Tecnologie
Biomediche (sezione di Microbiologia Applicata e Tecnologie Biomediche)
dell’Università degli Studi di Cagliari, seguendo il seguente protocollo:
• prelievo dal sistema reagente, in condizioni sterili, di campioni di 1 ml
ad intervalli di tempo prestabiliti mediante rubinetto inserito nel
circuito tra il reattore e la bottiglia raccoglitrice;
• diluizione dei campioni prelevati in terreno MH di un fattore
compreso tra 10-2 e 10-7 (a seconda della concentrazione batterica
“attesa”) e coltivazione in “CLED Agar medium” con incubazione per
24 ore alla temperatura di 37°C, al termine delle quali veniva
effettuata una conta preliminare;
• esecuzione della conta finale dopo 48 ore dalla semina, con
determinazione dell’effettivo numero di batteri presenti in ciascun
campione esaminato.

CAPITOLO 3 – LA SPERIMENTAZIONE
- 107 -
3.4.4 Densità ottica e conta in piastra a confronto
Un punto fondamentale del presente lavoro di tesi è stata la validazione della
metodologia analitica della misurazione della densità ottica applicata alla
determinazione delle costanti di crescita delle popolazioni batteriche che sono state
prese in esame. La marcata preferenza accordata a tale metodologia in luogo di
quella della conta in piastra, infatti, è motivata dai seguenti importanti fatti:
• assenza di contatto tra l’ambiente reagente di crescita dei
microorganismi e l’ambiente esterno, che ha come conseguenza la
totale eliminazione del rischio di contaminazioni, o perlomeno di
quelle imputabili al prelievo di campioni;
• possibilità di eseguire le determinazioni analitiche senza dover
disporre delle strutture e delle condizioni ambientali particolarmente
restrittive caratteristiche di un laboratorio di microbiologia, che sono
invece necessarie qualora si debbano maneggiare dei campioni;
• possibilità di compiere un numero notevolmente più elevato di
misurazioni in quanto, con la struttura disponibile, queste possono
essere effettuate in modo del tutto automatico ad intervalli di tempo
prestabiliti, anche particolarmente brevi (nell’ordine del minuto);
• possibilità di effettuare una quantità notevolmente maggiore di prove
sperimentali in quanto, grazie alla completa automazione delle
operazioni di misura, non è necessaria la presenza continuativa di un
operatore, e ciò consente l’utilizzo di tempi normalmente “morti”
come le ore notturne o le giornate festive: ciò si è rivelato di cruciale
importanza in quanto l’incidenza degli eventuali errori sperimentali è
tanto più piccola quanto maggiore è il numero di prove condotte per
ciascun gruppo di parametri testati.

CAPITOLO 3 – LA SPERIMENTAZIONE
- 108 -
In virtù di quanto detto sopra, il problema che ci si è posti è stato quello di stabilire
se, e in quale modo, fosse possibile, ai nostri fini, utilizzare misure di densità ottica,
effettuate in continuo sulla miscela reagente, in luogo della conta in piastra con
determinazione delle Unità Formanti Colonie; pare necessario sottolineare il fatto
che, con l’espressione “ai nostri fini” si intende ai fini della comparazione tra loro di
costanti di crescita comunque derivate, al solo scopo di individuare effetti relativi
imputabili alle condizioni del campo elettromagnetico applicato, ben lungi dunque
dalla ricerca di una validazione in senso generale, anche perché la cosa avrebbe poco
senso di per sé, in quanto le due tecniche misurano grandezze sostanzialmente
differenti. Si è dunque proceduto con la conta in piastra per almeno una parte delle
prove effettuate, al fine di verificare la corrispondenza tra le informazioni derivanti
dall’analisi spettrofotometrica (concentrazione di biomassa totale) e quelle derivanti
dalla conta in piastra (concentrazione di microorganismi vivi). Per motivi di brevità
si riporta il solo caso, già contenuto in un precedente lavoro di Carta e Desogus
(2008a), del confronto tra le costanti di crescita del microorganismo Listeria
monocytogenes derivate utilizzando parallelamente i dati derivati dalle due
metodologie analitiche e relative alla frequenza di 2,40 GHz e a livelli di potenza
incidente compresi tra 0 e 400 mW, con intervalli di 50 mW; in Fig. 3.8 sono
riportate le informazioni di cui sopra; per una descrizione più accurata della
metodologia di derivazione delle costanti di crescita si rimanda al successivo
paragrafo 3.5. Se le due tecniche analitiche fossero, ai nostri fini (secondo dunque
l’accezione già specificata), tra di loro intercambiabili, le costanti di crescita derivate
dalle due metodologie analitiche dovrebbero essere, per analoghe condizioni
sperimentali, uguali tra loro, e nel grafico di Fig. 3.8 dovrebbero essere
rappresentate da punti giacenti su una retta passante per l’origine degli assi e con
pendenza unitaria; in effetti, la retta di regressione (forzata per l’origine) dei punti
contenuti nel grafico in Fig. 3.8 ha una pendenza pari a circa 0,96, che può essere
considerato un valore caratterizzato da discreta bontà, considerata l’elevata
incidenza degli errori sperimentali. Risultati simili, generalmente compresi nel
campo 0,90-1,10, e dunque del tutto accettabili, sono stati ottenuti per tutte le specie
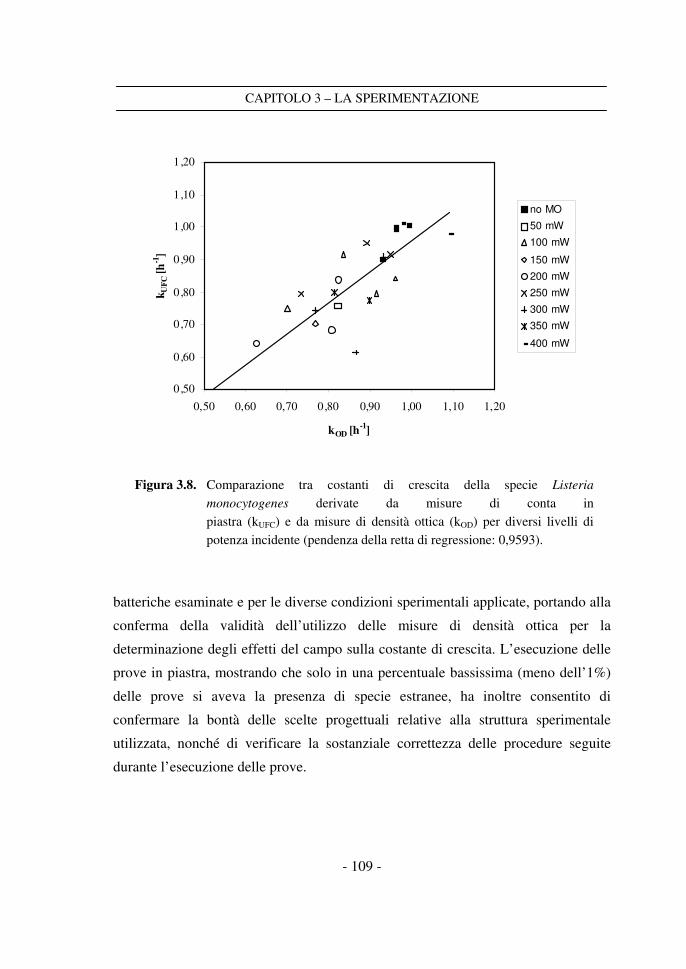
CAPITOLO 3 – LA SPERIMENTAZIONE
- 109 -
batteriche esaminate e per le diverse condizioni sperimentali applicate, portando alla
conferma della validità dell’utilizzo delle misure di densità ottica per la
determinazione degli effetti del campo sulla costante di crescita. L’esecuzione delle
prove in piastra, mostrando che solo in una percentuale bassissima (meno dell’1%)
delle prove si aveva la presenza di specie estranee, ha inoltre consentito di
confermare la bontà delle scelte progettuali relative alla struttura sperimentale
utilizzata, nonché di verificare la sostanziale correttezza delle procedure seguite
durante l’esecuzione delle prove.
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
0,50 0,60 0,70 0,80 0,90 1,00 1,10 1,20
kOD [h-1
]
kU
FC
[h
-1]
no MO
50 mW
100 mW
150 mW
200 mW
250 mW
300 mW
350 mW
400 mW
Figura 3.8. Comparazione tra costanti di crescita della specie Listeria
monocytogenes derivate da misure di conta in piastra (kUFC) e da misure di densità ottica (kOD) per diversi livelli di potenza incidente (pendenza della retta di regressione: 0,9593).

CAPITOLO 3 – LA SPERIMENTAZIONE
- 110 -
3.5 La determinazione delle costanti di crescita
La velocità di crescita di una popolazione di microorganismi, con particolare
riferimento alla fase di crescita esponenziale e alla fase stazionaria, può essere
espressa facendo ricorso a diverse tipologie di relazioni matematiche, tra le quali
quella più comunemente utilizzata, per via della sua semplicità formale, è
l’equazione di Monod; in questa sede tale espressione è stata scelta in quanto, pur
nella sua semplicità, ha mostrato di coniugarsi perfettamente con i dati sperimentali
che sono stati ottenuti, interpretandoli in maniera ottima.
L’equazione di Monod (Monod, 1949) esprime la velocità di crescita ( MR ) in
funzione della concentrazione di microorganismi ( CC ) e della concentrazione di
substrato ( SC ) per condizioni fissate di temperatura (da cui la costante di crescita
dipende in misura significativa), pH, aerazione, fonti di carbonio, fonti dei diversi
nutrienti; essa può essere scritta nella forma:
SM
SCM CK
CCkR
+
⋅⋅= (3.15)
dove k è la costante di crescita, mentre MK è detta “costante di Monod” ed è pari al
valore di CC in corrispondenza del quale si ha una velocità di crescita pari alla metà
di quella massima. La velocità di crescita massima (illimitata) si manifesta
allorquando si è nella condizione di eccesso di substrato che, utilizzando i termini
contenuti nell’equazione 3.15, equivale alla seguente condizione:
MS KC >> (3.16)
Se la 3.16 è verificata, allora la relazione 3.15 può essere approssimata con la
seguente:
CM CkR ⋅= (3.17)

CAPITOLO 3 – LA SPERIMENTAZIONE
- 111 -
cioè con un’equazione cinetica del primo ordine rispetto alla concentrazione di
microorganismi. Considerando la definizione stessa di MR , ed indicando con t la
variabile temporale, si ha che:
td
C dR C
M = (3.18)
Di conseguenza è possibile, scrivere la seguente forma linearizzata dell’equazione
3.17:
( )0C,0
C t-tkC
CLn ⋅=
(3.19)
dove C,0C è il valore della concentrazione batterica che corrisponde al tempo 0t ,
ovvero all’istante con il quale può farsi coincidere l’inizio della fase di crescita
esponenziale. Tale istante deve essere determinato empiricamente volta per volta, in
quanto fortemente variabile tra una prova e l’altra: l’intervallo di tempo che
intercorre tra l’inizio effettivo della prova e l’istante 0t , infatti, corrisponde alla fase
di latenza, la durata della quale è caratterizzata da una scarsa riproducibilità e può
difficilmente essere determinata a priori se non, come ha tentato di fare Baranyi
(1998; 2002), attraverso l’utilizzo di modelli stocastici. È forse banale, ma allo
stesso tempo importante ai fini del lavoro che e stato svolto, sottolineare il fatto che,
poiché nel termine di sinistra della 3.19 compare un rapporto di concentrazioni, è del
tutto ininfluente quale sia l’unità di misura adottata, quantunque non
“convenzionale”, come nel caso delle Unità Formanti Colonie per unità di volume e
in quello della densità ottica; con riferimento a quest’ultima eventualità, si rimanda
alla relazione 3.11, secondo la quale la densità ottica è direttamente proporzionale
alla concentrazione (e con intercetta nulla), e alle considerazioni già fatte a
proposito. A questo punto, con un’operazione di regressione lineare dei dati
sperimentali a disposizione per ogni prova, è possibile determinare la costante di
crescita corrispondente come pendenza della retta di regressione.

CAPITOLO 3 – LA SPERIMENTAZIONE
- 112 -
Una questione di importanza non secondaria, ai fini della trattazione sopra
esposta, è certamente quella definita pocanzi come “condizione di eccesso di
substrato”. Poiché, infatti, tutte le determinazioni della costante di crescita k che
sono state condotte si basano sulla condizione matematica rappresentata dalla 3.16,
si è ritenuto necessario quanto opportuno verificare la sussistenza di tale condizione
anche in termini fisici, non limitandosi quindi a prendere per buone in modo acritico
le concentrazioni di substrato consigliate dal produttore; tale verifica è stata
condotta, per ciascuna delle quattro specie prese in esame, secondo le modalità di
seguito descritte. Seguendo tutte le medesime indicazioni già fornite in questo
paragrafo e nei precedenti, sono state eseguite delle prove partendo da
concentrazioni iniziali di substrato differenti rispetto a quelle indicate nel paragrafo
3.3.1, sia maggiori sia minori (queste ultime fino a raggiungere condizioni di elevata
diluizione), e ricavando volta per volta la costante di crescita secondo la
linearizzazione riportata nella relazione 3.19. In realtà, nel caso più generale in cui
non si operi la semplificazione prodotta dalla condizione 3.16, l’applicazione della
3.19 non conduce alla determinazione di k, bensì della grandezza che definiamo con
k′ :
kβk ⋅=′ (3.20)
avendo posto:
SM
S
CK
Cβ
+= (3.21)
Devono essere effettuate ora alcune brevi considerazioni. Innanzitutto, anche quando
la condizione 3.16 non è verificata, la cosiddetta “fase di crescita esponenziale”
(così come definita nel paragrafo 3.1) esiste ancora, almeno fintanto che la
variazione di SC , dovuta al consumo del substrato da parte dei microorganismi, è
sufficientemente piccola, e per quanto questa fase possa essere più o meno breve,
sarà comunque individuabile e dunque sarà determinabile il valore di k′ ; inoltre, se
la variazione di SC è sufficientemente piccola, il suo valore, durante la fase di

CAPITOLO 3 – LA SPERIMENTAZIONE
- 113 -
crescita esponenziale, rimarrà prossimo al suo valore iniziale, che indichiamo con
S,0C , per cui la 3.21 può essere riscritta nella forma:
S,0M
S,0
CK
Cβ
+= (3.22)
È altresì evidente come β possa assumere solo valori compresi tra 0 (quando la
concentrazione iniziale del substrato è nulla) e 1 (quando il substrato è in eccesso,
cioè è verificata la condizione 3.16); k e k′ si discosteranno tra loro tanto più quanto
minore è S,0C rispetto a MK , mentre coincideranno quando sussiste l’ipotesi di
eccesso di substrato. È parimenti evidente che la 3.22 rappresenta una funzione
crescente monotona di S,0C , con raggiungimento di un livello di “plateau” quando k
e k′ vengono a coincidere; in quest’ultimo caso, e solo in questo, la crescita potrà
perciò dirsi, oltre che esponenziale (in senso matematico), anche illimitata, in quanto
non più limitata in qualche modo dalla scarsità di substrato. In base a tutte le
considerazioni fatte sinora è stata costruita, per ogni specie microbica esaminata, una
curva sperimentale di k′ (alla medesima temperatura di 37°C) in funzione di S,0C ,
verificando così in tutti e quattro i casi che la concentrazione di substrato utilizzata
durante la campagna sperimentale garantiva la sussistenza della condizione di
eccesso di substrato; tale verifica è, tra l’altro, conforme a quanto riportato da
Levenspiel (1984). Per ragioni di brevità (le curve sono tra loro analoghe), in
Fig. 3.9 si riporta la sola curva relativa alla specie Staphylococcus aureus (il
substrato utilizzato è il BHI); dall’esame di quest’ultima si evince come il “plateau”
possa dirsi raggiunto già per valori di S,0C intorno a 15 g/l (la concentrazione
utilizzata durante le prove principali è stata di 37 g/l, dunque più che doppia), e
come il valore di MK , in questo caso, dovrebbe aggirarsi intorno a 0,5÷1 g/l (valore
di S,0C corrispondente a valori di k′ posizionati all’incirca a metà altezza del
“plateau”). In Fig. 3.10 si riportano alcune curve di crescita della specie
Staphylococcus aureus ottenute per diversi valori di S,0C .
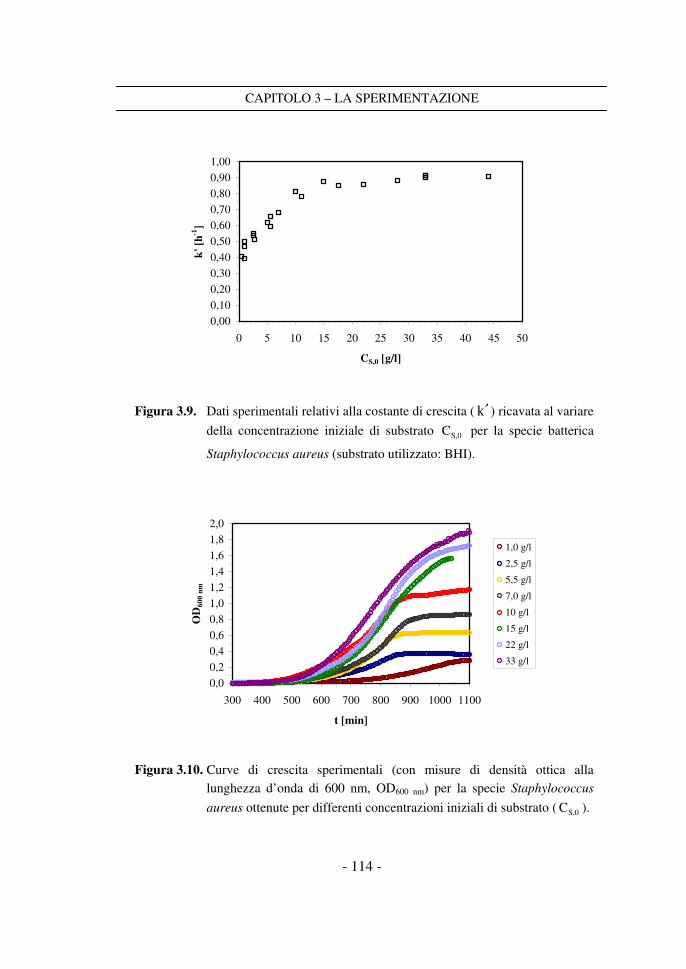
CAPITOLO 3 – LA SPERIMENTAZIONE
- 114 -
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
0 5 10 15 20 25 30 35 40 45 50
CS,0 [g/l]
k'
[h-1
]
Figura 3.9. Dati sperimentali relativi alla costante di crescita ( k′ ) ricavata al variare
della concentrazione iniziale di substrato S,0C per la specie batterica
Staphylococcus aureus (substrato utilizzato: BHI).
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
300 400 500 600 700 800 900 1000 1100
t [min]
OD
60
0 n
m
1,0 g/l
2,5 g/l
5,5 g/l
7,0 g/l
10 g/l
15 g/l
22 g/l
33 g/l
Figura 3.10. Curve di crescita sperimentali (con misure di densità ottica alla lunghezza d’onda di 600 nm, OD600 nm) per la specie Staphylococcus
aureus ottenute per differenti concentrazioni iniziali di substrato ( S,0C ).

CAPITOLO 4


CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 117 -
4.1 Le costanti di crescita sperimentali
Si presentano ora le informazioni sperimentali relative alle costanti di crescita
per le specie batteriche esaminate, ricavate secondo le modalità esposte nel capitolo
precedente e utilizzando dati di densità ottica ottenuti con letture alla lunghezza
d’onda di 600 nm effettuate ad intervalli di 5 (e, sporadicamente, di 2,5) minuti. Per
quanto riguarda il Bacillus clausii, le prove che lo hanno riguardato sono state
condotte tutte alla frequenza di 2,45 GHz e con potenze incidenti pari a 100, 200,
300 e 400 mW, oltre naturalmente alle prove condotte in assenza di irradiazione.
Tale fase sperimentale, condotta sul Bacillus clausii, ha avuto anche lo scopo di
coadiuvare la messa a punto dell’apparato e delle metodologie sperimentali,
trattandosi di un microorganismo non pericoloso per la salute e facilmente reperibile
nel prodotto commerciale Enterogermina® (Sanofi-synthelabo®), e proprio per
questo è stato interessato da un numero di prove superiore rispetto alle altre specie.
Sono state condotte, infatti, per ogni livello di potenza, un numero di prove tale da
garantire che almeno ventuno di queste fornissero costanti di crescita
sufficientemente vicine tra loro; le prove che hanno fornito un esito considerato
positivo sono state all’incirca l’80% del totale delle prove effettuate, intendendo per
“esito positivo” l’ottenimento di una costante di crescita che si discostasse di non
più del 5% dal valore medio e con un coefficiente di regressione (regressione lineare
effettuata ricorrendo alla relazione 3.19) non inferiore a 0,999. L’assunzione di un
intervallo di errore accettato nella misura di ± 5% rispetto al valore medio è dovuta
alla necessità di accettare elevati livelli di incertezza su numerose variabili (potenza
e frequenza della radiazione, SAR, velocità di circolazione della miscela, quantità
della miscela, concentrazione dei nutrienti, stato fisiologico iniziale delle cellule,
etc.), oltre alla possibilità di contaminazioni esterne, e si è rivelata la scelta più
idonea a considerare positive un numero (per l’appunto circa l’80% del totale)
sufficientemente elevato di prove. I valori medi delle costanti di crescita ottenute per
il Bacillus clausii sono riportati in Tabella 4.1; la loro rilevanza, visto che non si

CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 118 -
tratta di un patogeno, è spiegata con il fatto che questa specie appartiene allo stesso
genere di altre specie patogene di interesse per la sicurezza alimentare (come per
esempio il Bacillus cereus), alle quali è dunque morfologicamente simile.
Lo stesso intervallo di errore (± 5% rispetto al valore medio) è stato accettato
per le altre tre specie microbiche, per le quali sono state tuttavia eseguite prove fino
ad averne quindici (anziché ventuno) con esito positivo per ogni coppia di valori di
frequenza e di potenza; la percentuale di prove, rispetto al totale, concluse con esito
positivo è stata dell’85-90% (con punte del 95%), probabilmente per via del fatto
che, con il procedere della sperimentazione, sono stati limitati, almeno in parte, gli
errori commessi nella manipolazione delle cellule durante l’inoculazione: la bontà
dei risultati ottenuti è stata accertata con l’esecuzione, a scopo di controllo, di prove
di conta in piastra che hanno rivelato la diminuzione dei casi di contaminazione
della miscela reagente da parte di specie batteriche estranee. Nelle seguenti Tabelle
4.2, 4.3 e 4.4 sono contenuti i valori medi delle costanti di crescita ottenuti
rispettivamente per le specie Pseudomonas aeruginosa, Staphylococcus aureus e
Listeria monocytogenes.
P [mW] k [h-1]
0 0,43
100 0,40
200 0,39
300 0,39
400 0,40
Tabella 4.1. Costanti di crescita (k) della specie Bacillus clausii al variare della
potenza incidente (P) e alla frequenza di 2,45 GHz (valori medi, errore ± 5%).

CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 119 -
f [GHz]
P [mW] 2,20 2,30 2,40 2,50
0 1,06
100 0,84 0,78 0,85 0,94
200 0,76 0,89 0,84 0,88
300 0,81 0,94 0,67 0,91
400 0,77 0,69 0,80 0,88
Tabella 4.3. Costanti di crescita (k) della specie Staphylococcus aureus al variare
della potenza incidente (P) e della frequenza (f) (valori medi, errore ± 5%).
f [GHz]
P [mW] 2,20 2,30 2,40 2,50
0 1,06
100 1,08 1,06 1,06 1,10
200 1,06 1,10 1,11 1,13
300 1,04 1,05 1,06 1,03
400 1,20 1,05 1,02 1,12
Tabella 4.2. Costanti di crescita (k) della specie Pseudomonas aeruginosa al variare
della potenza incidente (P) e della frequenza (f) (valori medi, errore ± 5%).

CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 120 -
4.2 Il confronto tra i dati a potenza variabile
Per cercare di fornire una rappresentazione quanto migliore possibile dei dati
sperimentali che sono stati ottenuti, si sono riportati in forma grafica i valori
contenuti nelle tabelle precedenti, in modo da mettere maggiormente in evidenza le
variazioni registrate al variare della potenza (in questo paragrafo) e della frequenza
(nel paragrafo 4.3); nelle Figg. 4.1, 4.2, 4.3 e 4.4 sono riportati i grafici relativi, in
f [GHz]
P [mW] 2,30 2,40 2,50 2,60
0 0,84
50 0,77 0,75 0,73 0,78
100 0,77 0,81 0,78 0,83
150 0,83 0,85 0,79 0,80
200 0,76 0,84 0,79 0,88
250 0,86 0,86 0,76 0,83
300 0,75 0,85 0,83 0,94
350 0,81 0,85 0,86 0,84
400 0,89 0,92 0,85 0,86
Tabella 4.4. Costanti di crescita (k) della specie Listeria monocytogenes al variare
della potenza incidente (P) e della frequenza (f) (valori medi, errore ± 5%).
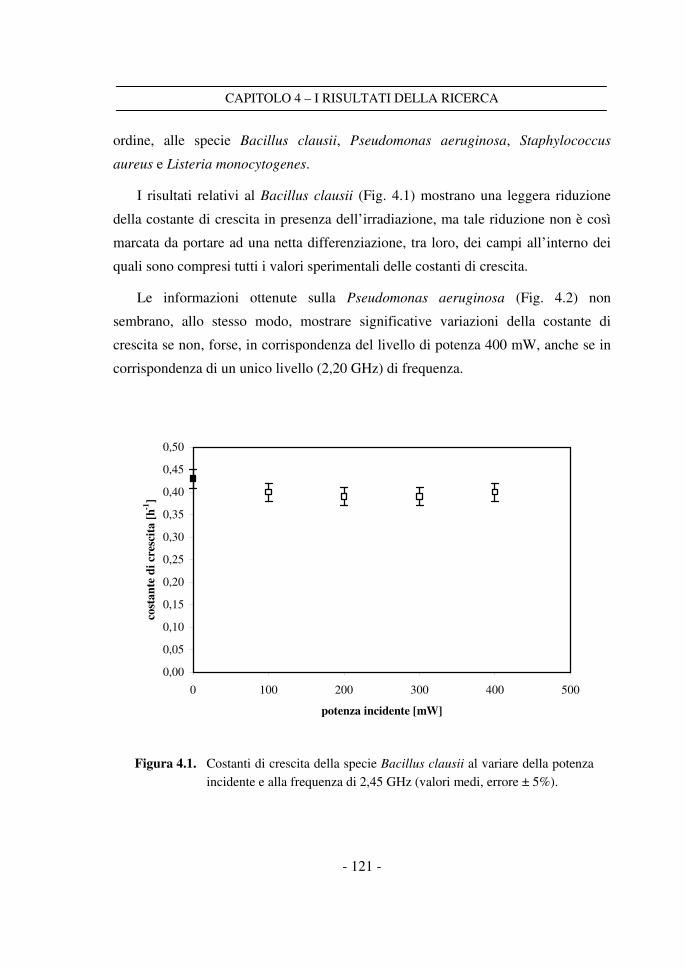
CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 121 -
ordine, alle specie Bacillus clausii, Pseudomonas aeruginosa, Staphylococcus
aureus e Listeria monocytogenes.
I risultati relativi al Bacillus clausii (Fig. 4.1) mostrano una leggera riduzione
della costante di crescita in presenza dell’irradiazione, ma tale riduzione non è così
marcata da portare ad una netta differenziazione, tra loro, dei campi all’interno dei
quali sono compresi tutti i valori sperimentali delle costanti di crescita.
Le informazioni ottenute sulla Pseudomonas aeruginosa (Fig. 4.2) non
sembrano, allo stesso modo, mostrare significative variazioni della costante di
crescita se non, forse, in corrispondenza del livello di potenza 400 mW, anche se in
corrispondenza di un unico livello (2,20 GHz) di frequenza.
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
0,45
0,50
0 100 200 300 400 500
potenza incidente [mW]
cost
an
te d
i cr
esci
ta [
h-1
]
Figura 4.1. Costanti di crescita della specie Bacillus clausii al variare della potenza
incidente e alla frequenza di 2,45 GHz (valori medi, errore ± 5%).
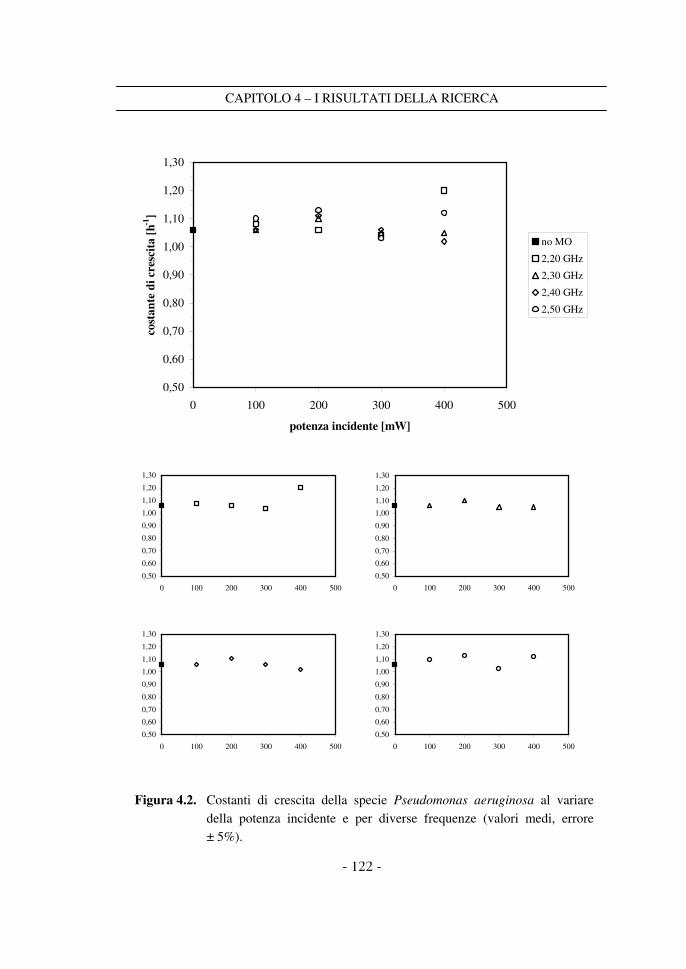
CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 122 -
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
potenza incidente [mW]
cost
an
te d
i cr
esci
ta [
h-1
]
no MO
2,20 GHz
2,30 GHz
2,40 GHz
2,50 GHz
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
Figura 4.2. Costanti di crescita della specie Pseudomonas aeruginosa al variare
della potenza incidente e per diverse frequenze (valori medi, errore ± 5%).
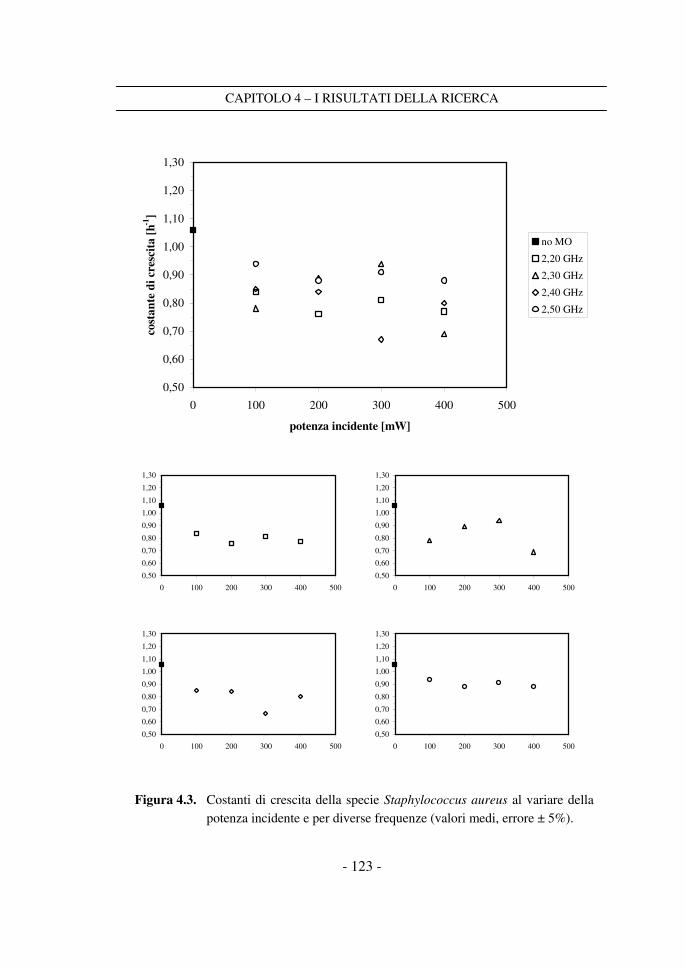
CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 123 -
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
potenza incidente [mW]
cost
an
te d
i cr
esci
ta [
h-1
]
no MO
2,20 GHz
2,30 GHz
2,40 GHz
2,50 GHz
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
Figura 4.3. Costanti di crescita della specie Staphylococcus aureus al variare della
potenza incidente e per diverse frequenze (valori medi, errore ± 5%).
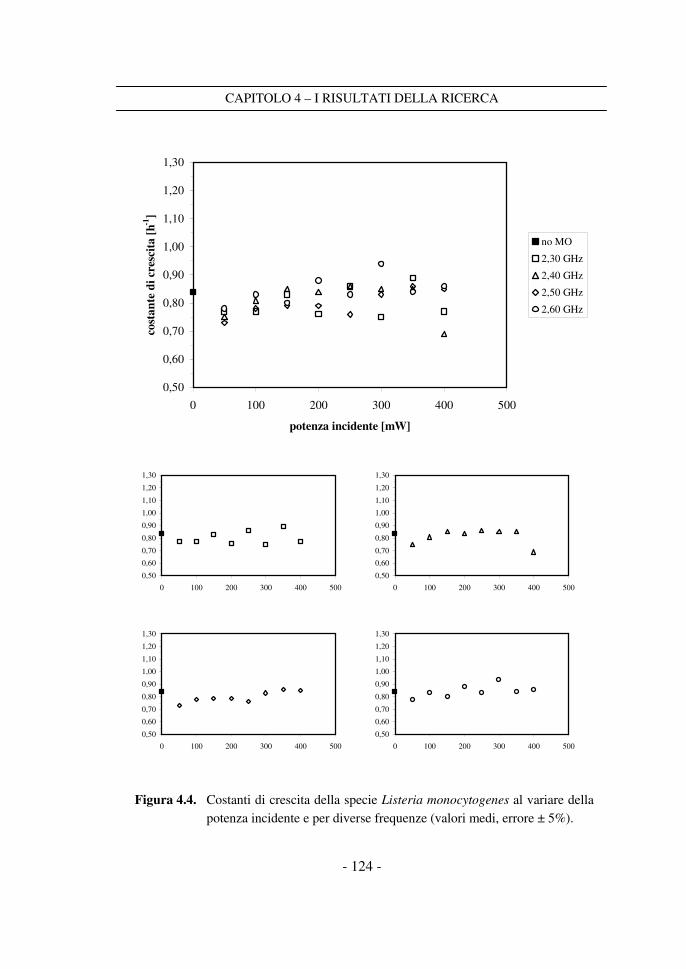
CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 124 -
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
potenza incidente [mW]
cost
an
te d
i cr
esci
ta [
h-1
]
no MO
2,30 GHz
2,40 GHz
2,50 GHz
2,60 GHz
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
0 100 200 300 400 500
Figura 4.4. Costanti di crescita della specie Listeria monocytogenes al variare della
potenza incidente e per diverse frequenze (valori medi, errore ± 5%).

CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 125 -
Per quanto riguarda lo Staphylococcus aureus (Fig. 4.3), invece, le variazioni
riscontrate tra l’assenza e la presenza di irradiazione sembrano essere più marcate, e
pressoché tutte nella stessa direzione della diminuzione; le variazioni indotte dal
livello di potenza non sembrano viceversa particolarmente significative, se non in
corrispondenza della frequenza 2,30 GHz.
La specie Listeria monocytogenes (Fig. 4.4) sembra ugualmente mostrare,
almeno nella maggior parte dei casi, variazioni nel senso della diminuzione, ma
comunque più contenute rispetto a quelle riscontrabili per lo Staphylococcus aureus;
vi è una certa variabilità rispetto alla potenza incidente, ma senza tendenze
unidirezionali.
4.3 Il confronto tra i dati a frequenza variabile
Analogamente a quanto fatto nel paragrafo 4.2, si riportano ora i grafici relativi
ai dati contenuti nelle Tabelle 4.2, 4.3 e 4.4, in modo da evidenziare l’andamento
delle costanti di crescita al variare della frequenza delle microonde; nelle Figg. 4.5,
4.6 e 4.7 sono riportati i grafici relativi, nell’ordine, alle specie Pseudomonas
aeruginosa, Staphylococcus aureus e Listeria monocytogenes (non vi è alcun grafico
per la specie Bacillus clausii in quanto le prove sono state condotte tutte all’unica
frequenza di 2,45 GHz).
La specie Pseudomonas aeruginosa (Fig. 4.5), oltre all’unico caso, già citato, in
corrispondenza del valore 2,20 GHz (per il livello di potenza 400 mW), non mostra
significative variazioni della costante di crescita al variare della frequenza.
Per ciò che riguarda la specie Staphylococcus aureus (Fig. 4.6), invece, le
variazioni già riscontrate nel senso della diminuzione sembrano essere più marcate
in corrispondenza delle frequenze 2,20 e 2,30 GHz, anche se il valore minimo è stato
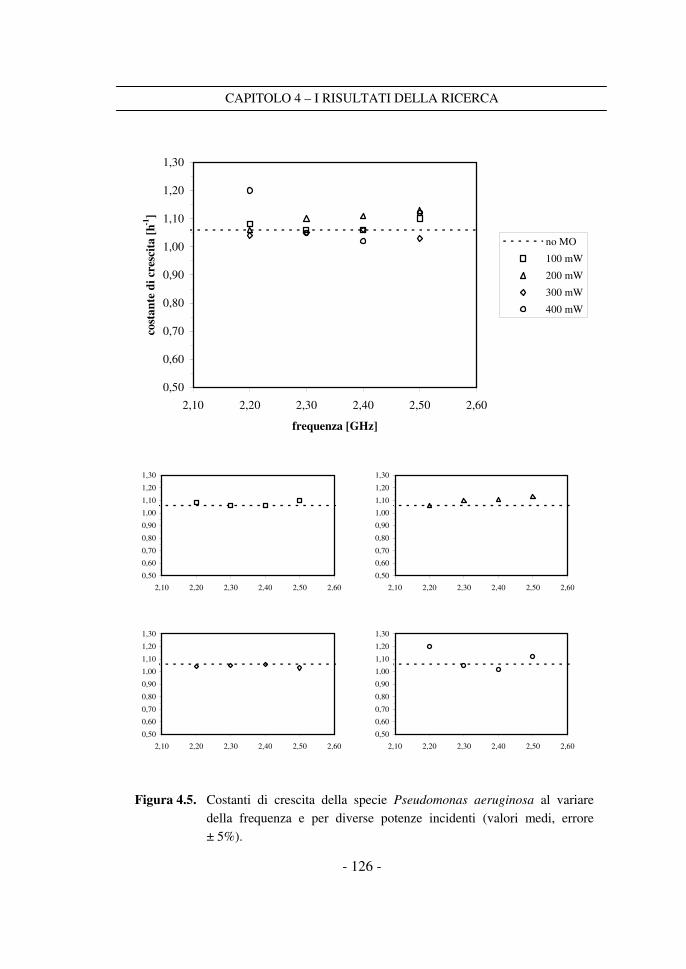
CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 126 -
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
frequenza [GHz]
cost
an
te d
i cr
esci
ta [
h-1
]
no MO
100 mW
200 mW
300 mW
400 mW
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
Figura 4.5. Costanti di crescita della specie Pseudomonas aeruginosa al variare
della frequenza e per diverse potenze incidenti (valori medi, errore ± 5%).

CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 127 -
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
frequenza [GHz]
cost
an
te d
i cr
esci
ta [
h-1
]
no MO
100 mW
200 mW
300 mW
400 mW
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,10 2,20 2,30 2,40 2,50 2,60
Figura 4.6. Costanti di crescita della specie Staphylococcus aureus al variare della
frequenza e per diverse potenze incidenti (valori medi, errore ± 5%).
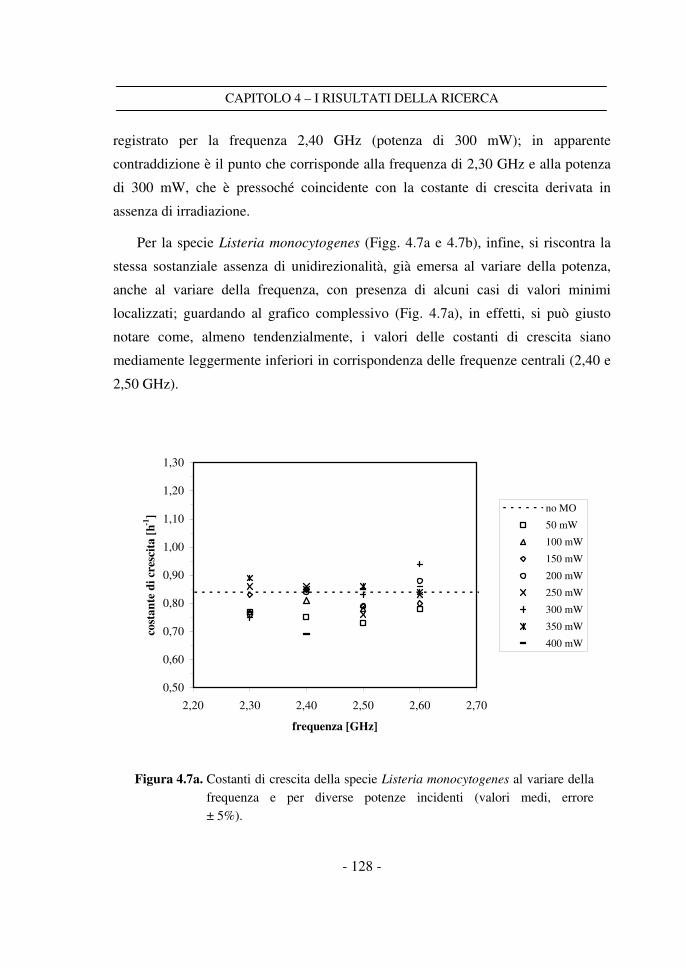
CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 128 -
registrato per la frequenza 2,40 GHz (potenza di 300 mW); in apparente
contraddizione è il punto che corrisponde alla frequenza di 2,30 GHz e alla potenza
di 300 mW, che è pressoché coincidente con la costante di crescita derivata in
assenza di irradiazione.
Per la specie Listeria monocytogenes (Figg. 4.7a e 4.7b), infine, si riscontra la
stessa sostanziale assenza di unidirezionalità, già emersa al variare della potenza,
anche al variare della frequenza, con presenza di alcuni casi di valori minimi
localizzati; guardando al grafico complessivo (Fig. 4.7a), in effetti, si può giusto
notare come, almeno tendenzialmente, i valori delle costanti di crescita siano
mediamente leggermente inferiori in corrispondenza delle frequenze centrali (2,40 e
2,50 GHz).
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
frequenza [GHz]
cost
an
te d
i cr
esci
ta [
h-1
]
no MO
50 mW
100 mW
150 mW
200 mW
250 mW
300 mW
350 mW
400 mW
Figura 4.7a. Costanti di crescita della specie Listeria monocytogenes al variare della
frequenza e per diverse potenze incidenti (valori medi, errore ± 5%).
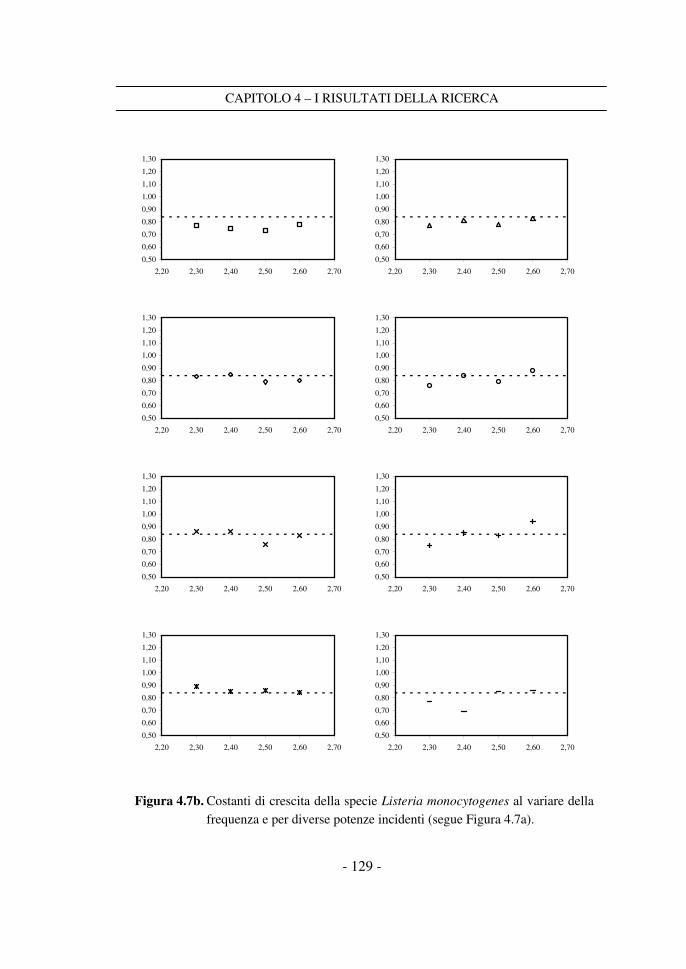
CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 129 -
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
0,50
0,60
0,70
0,80
0,90
1,00
1,10
1,20
1,30
2,20 2,30 2,40 2,50 2,60 2,70
Figura 4.7b. Costanti di crescita della specie Listeria monocytogenes al variare della
frequenza e per diverse potenze incidenti (segue Figura 4.7a).

CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 130 -
4.4 Commenti conclusivi e prospettive future
È innegabile, andando ad esaminare i risultati sperimentali presentati nei
paragrafi 4.1, 4.2 e 4.3, il fatto che manchi una precisa evidenza che possa condurre
a conclusioni nette nel senso dell’effettivo riscontro di effetti non termici prodotti
dalle microonde con potenze comprese tra 0 e 400 mW e alle frequenze indagate
sulle quattro specie batteriche che sono state oggetto della sperimentazione, ma
mancano altresì sufficienti elementi che possano condurre alla negazione degli
stessi. E proprio per questo, quel che pare altrettanto innegabile è che lo studio
richieda e meriti certamente di essere continuato e ampliato. È anche vero che, nelle
pur modeste variazioni delle costanti di crescita al variare delle condizioni di
esposizione alle microonde, manca il riscontro di una qualche correlazione tra
l’entità dell’effetto e le variabili potenza e frequenza, ma questo, almeno allo stato
attuale, può non essere considerato un fatto necessario, vista la presenza, in
letteratura, di alcuni casi analoghi e anche di diversi riscontri di cosiddetti “effetti
finestra” (vedi paragrafo 2.4 e relativi sottoparagrafi), che lasciano intendere,
almeno in linea teorica, la possibile manifestazione di effetti solo in particolari
condizioni sperimentali, e non per condizioni differenti, anche se solo di poco.
Elementi non secondari, che pare qui giusto sottolineare, sono l’accertata
affidabilità della struttura sperimentale così come è stata concepita e posta in opera e
la sostanziale correttezza delle procedure sperimentali e analitiche; si tratta di
constatazioni che, in quanto tutt’altro che scontate in fase di avvio del lavoro,
possono essere considerate una concreta e solida base di partenza per gli ulteriori
sviluppi dell’indagine sperimentale.
Tali ulteriori sviluppi potranno certamente concretarsi in un’estensione dei
campi di frequenza e di potenza indagati (anche se ciò comporterà alcune modifiche
nell’apparato di generazione delle microonde) così come nello studio di altre specie
microbiche; di interesse potrà essere anche l’utilizzo di differenti substrati,

CAPITOLO 4 – I RISULTATI DELLA RICERCA
- 131 -
possibilmente con caratteristiche più simili a quelle di alimenti reali, o addirittura di
alimenti in stato fluido veri e propri.


BIBLIOGRAFIA
- 133 -
BIBLIOGRAFIA
Adey W. R., 1981, “Tissue interactions with nonionizing electromagnetic fields”, Physiol
Rev 61(2): 435-514.
Akoev I. G. (ed.), 1986, “Biological effects of electromagnetic fields”, USSR Academy of Sciences, Pushchino (Russia).
Alekseev S. I., Ziskin M. C., 1995, “Millimeter microwave effect on ion transport across lipid bilayer membranes”, Bioelectromagnetics 16(2): 124-131.
Antonopoulos A., Eisenbrandt H., Obe G., 1997, “Effects of high-frequency electromagnetic fields on human lymphocytes in vitro”, Mutat Res Fundam Mol Mech Mutagen 395(2-3): 209-214.
Armstrong R. W., Fung P. C., 1993, “Brainstem encephalitis (Rhombencephalitis) due to Listeria monocytogenes: case report and review”, Clin Infect Dis 16: 689-702.
Atkins P. W., Friedman R. S., 2004, “Molecular quantum mechanics” (4a ed.), Oxford University Press, Oxford (Regno Unito).
Atkins P. W., Lambert T. P., 1975, “The effect of a magnetic field on chemical reactions”, Annu Rep Prog Chem (Sect A, Inorg and Phys Chem) 72: 67-88.
Banik S., Bandopadhyay S., Ganguly S., Dan D., 2002a, “Effect of inoculation of microwave irradiated methanogenic bacteria for improvement of biomethanation from jute caddis”, 4th National Symposium on Frontier Studies in Microbes and Host Microbes Interaction, University of Calcutta, Calcutta (India), 2-3 febbraio 2002.
Banik S., Bandopadhyay S., Ganguly S., Dan D., 2002b, “Application of microwave technology on methanogenic bacteria for improvement of its efficiency and endavour to increase yield of biogas”, 9th West Bengal State Science and Technology Congress, Visva-Bharati University, Santiniketan (India), 28 febbraio-2 marzo 2002.
Baranyi J. M., 1998, “Comparison of stochastic and deterministic concepts of bacterial lag phase”, J Theor Biol 192(3): 403-408.
Baranyi J. M., 2002, “Stochastic modelling of bacterial lag phase”, Int J Food Microbiol 73(2-3): 203-206.
Barnothy J. M., 1963, “Growth-rate of mice in stati magnetic fields”, Nature 200(4901): 86-87.
Barnothy J. M., Barnothy M. F., Boszormenyi-Nagy I., 1956, “Influence of a magnetic field upon the leucocytes of the mouse”, Nature 177(4508): 577-578.
Barnothy M. F. (ed.), 1964, “Biological effects of magnetic fields”, Plenum Press, New York (Stati Uniti).

BIBLIOGRAFIA
- 134 -
Barnothy M. F., Barnothy J. M., 1958, “Biological effect of a magnetic field and the radiation syndrome”, Nature 181(4626): 1785-1786.
Barnothy M. F., Sumegi I., 1969, “Abnormalities in organs of mice induced by a magnetic field”, Nature 221(5177): 270-271.
Bawin S. M., Adey W. R., 1976, “Sensitivity of calcium binding in cerebral tissue to weak environmental electric fields oscillating at low frequency”, Proc Natl Acad Sci USA 73(6): 1999-2003.
Bawin S. M., Kaczmarek L. K., Adey W. R., 1975, “Effects of modulated VHF fields on the central nervous system”, Ann N Y Acad Sci 247: 74-81.
Bawin S. M., Sheppard A., Adey W. R., 1978, “Possible mechanism of weak electromagnetic field coupling in brain tissue”, Bioelectrochem Bioenerg 5(1): 67-76.
Belyaev I. Y., Alipov Y. D., Polunin V. A., Shcheglov V. S., 1993a, “Evidence for dependence of resonant frequency of millimeter wave interaction with Escherichia coli K12 cells on haploid genome length”, Electromagn Biol Med 12(1): 39-49.
Belyaev I. Y., Shcheglov V. S., Alipov Y. D., Radko S. P., 1993b, “Regularities of separate and combined effects of circularly polarized millimeter waves on E. coli cells at different phases of culture growth”, Bioelectrochem Bioenerg 31(1): 49-63.
Belyaev I. Y., Alipov Y. D., Shcheglov V. S., Polunin V. A., Aizenberg O. A., 1994, “Cooperative response of Escherichia coli cells to the resonance effect of millimeter waves at super low intensity”, Electromagn Biol Med 13(1): 53-66.
Belyaev I. Y., Shcheglov V. S., Alipov Y. D., Polunin V. A., 1996, “Resonance effect of millimeter waves in the power range from 10-19 to 3 × 10-3 W/cm2 on Escherichia coli cells at different concentrations”, Bioelectromagnetics 17(4): 312-321.
Belyaev I. Y., Shcheglov V. S., Alipov Y. D., Ushakov V. L., 1997, “Reply to comments of Osepchuk and Petersen”, Bioelectromagnetics 18(7): 529-530.
Belyakov E. V., Kichaev V. A., Poslavskii M. V., Starshinina V. A., Soboleva E. S., 1989, “Use of blood radiophysical parameters in the extremely high-frequency range for diagnostic purposes”, in Devyatkov N. D. (ed.), Millimeter waves in medicine and biology, Radioelectronica, Mosca (Russia).
Berezhinskiĭ L. I., Gridina N. Y., Dovbeshko G. I., Lisitsa M. P., Litvinov G. S., 1993, “Visualization of the effects of millimeter radiation on blood plasma”, Biofizika 38(2): 378-384.
Berzhanskaya L. Y., Berzhanskii V. N., Beloplotova O. Y., 1995, “Effect of electromagnetic fields on the activity of bioluminescence in bacteria”, Biofizika 40: 974-977.
Betzky O. V., 1992, “Use of low-intensity electromagnetic millimeter waves in medicine”, Millimetr Volny Biol Med 1: 5-12.

BIBLIOGRAFIA
- 135 -
BIO-RAD, 2009, Bio-Rad Laboratories website, http://www3.bio-rad.com/cmc_upload/Technology_Spotlights/221138/16326_iqcheck.pdf
Blackman C. F., 1984, “Genetics and mutagenesis”, in Elder J. A., Cahill D. F. (ed.), Biological effects of radiofrequency radiation, U. S. Environmental Protection Agency, The Research Triangle Park (Stati Uniti).
Blackman C. F., Benane S. G., Elder J. A., House D. E., Lampe J. A., Faulk J. M., 1980a, “Induction of calcium-ion efflux from brain tissue by radiofrequency radiation: effect of sample number and modulation frequency on the power-density window”, Bioelectromagnetics 1(1): 35-43.
Blackman C. F., Benane S. G., Joines W. T., Hollis M. A., House D. E., 1980b, “Calcium-ion efflux from brain tissue: power-density versus internal field-intensity dependencies at 50 MHz RF radiation”, Bioelectromagnetics 1(3): 277-283.
Blackman C. F., Elder J. A., Weil C. M., Benane S. G., Eichinger D. C., House D. E., 1979, “Induction of calcium-ion efflux from brain tissue by radio-frequency radiation: effects of modulation frequency and field strenght”, Rad Res 14: 93-98.
Blackman C. F., Benane S. G., House D. E., 1991, “The influence of temperature during electric- and magnetic-field-induced alteration of calcium-ion release from in vitro brain tissue”, Bioelectromagnetics 12(3): 173-182.
Boiden Pedersen J., Freed J. H., 1974, “Some theoretical aspects of chemically induced dynamic nuclear polarization”, J Chem Phys 61(4): 1517-1525.
Brovkovich V. M., Kurilo N. B., Barishpol’ts V. L., 1991, “Action of millimeter-range electromagnetic radiation on the Ca pump of sarcoplasmic reticulum”, Radiobiologiia 31(2): 268-271.
Buchachenko A. L., 1974, “Chemically induced polarization of electrons and nuclei”, Nauka, Moscow (Russia).
Buchachenko A. L., Frankevich E. L., 1993, “Chemical generation and reception of radio- and microwaves”, VCH Publishers Inc., New York (Stati Uniti).
Buchachenko A. L., Molin Y. N., Sagdeev R. Z., Salikhov K. M., Frankevich E. L., 1987, “Magneto-spin effects in chemical reactions”, Sov Phys Usp 30(1): 79-80.
Bulgakova V. G., Grushina V. A., Orlova T. I., Petrykina Z. M., Polin A. N., Noks P. P., Kononenko A. A., Rubin A. B., 1996, “Effect of millimeter-band radiation of nonthermal intensity on the sensitivity of staphylococcus to various antibiotics”, Biofizika 41: 1289-1293.
Burachas G., Mascoliunas R., 1989, “Suppression of nerve action potential under the effect of millimeter waves”, in Devyatkov N. D. (ed.), Millimeter waves in medicine and biology, Radioelectronica, Mosca (Russia).

BIBLIOGRAFIA
- 136 -
Bourgeois C. M., Mescle J. F., Zucca J., 1990, “Microbiologia alimentare”, Tecniche Nuove, Milano.
Caiazza N. C., O’Toole G. A., 2004, “SadB is required for the transition from reversible to irreversible attachment during biofilm formation by Pseudomonas aeruginosa PA14”, J
Bacteriol 186(14): 4476-4485.
Carrington A., McLachlan A. D., 1967, “Introduction to magnetic resonance”, Harper & Row, Hagerstown (Stati Uniti).
Carta R., Desogus F., 2006, “Bacterial growth in microwave radiated reactors at low temperature”, 33rd International Conference of Slovak Society of Chemical Engineering, Tatranské Matliare (Slovacchia), 22-26 maggio 2006.
Carta R., Desogus F., 2007, “Effect of microwave radiation on the growth rate of Pseudomonas aeruginosa and Staphylococcus aureus at 37°C”, 34th International Conference of Slovak Society of Chemical Engineering, Tatranské Matliare (Slovacchia), 21-25 maggio 2007.
Carta R., Desogus F., 2008a, “Comparison between Listeria monocytogenes growth rate constants derived from optical density and UFC measurements”, MISA 2008 – 1° Incontro Bilaterale su Microonde nell’Ingegneria e nelle Scienze Applicate, Salerno, 21-23 maggio 2008.
Carta R., Desogus F., 2008b, “Influence of microwave frequency and power on the growth rate of Listeria monocytogenes at 37°C”, Convegno GR.I.C.U. 2008, Le Castella (KR), 14-17 settembre 2008.
Carta R., Desogus F., 2008c, “Pseudomonas aeruginosa growth in microwave irradiated environment”, in Marino G., Bardone E., Viglia A. (ed.), IBIC 2008 – 1st Industrial Biotechnology International Conference, Chemical Engineering Transactions (vol. 14), AIDIC Servizi S.r.l., Milano.
Carta R., Desogus F., 2009, “The effect of low-power microwaves on the growth of bacterial populations in a plug flow reactor”, AIChE J, in corso di pubblicazione, DOI: 10.1002/aic.12034.
Carta R., Desogus F., Errico M., 2006a, “Effect of microwave radiation on the growth rate of Bacillus clausii at 37°C”, 17th International Congress of Chemical and Process Engineering, Prague (Repubblica Ceca), 27-31 agosto 2006.
Carta R., Desogus F., Laconi S., 2006b, “Bacterial growth in microwave radiated reactors running at low temperature”, International Workshop on Finite Element Modelling in Chemical and Food Engineering, Fisciano (SA), 16-17 novembre 2006.
Casula G., Cutting M., 2002, “Bacillus probiotics: spore germination in the gastrointestinal tract”, Appl Environ Microbiol 68(5): 2344-2352.
Chambers H. F., 2001, “The changing epidemiology of Staphylococcus aureus?”, Emerg
Infect Dis 7(2): 178-182.

BIBLIOGRAFIA
- 137 -
Chernyakov G. M., Korochkin V. L., Babenko A. P., Bigdai E. V., 1989, “Reactions of biological systems of various complexity to the action of low-level EHF radiation”, in Devyatkov N. D. (ed.), Millimeter waves in medicine and biology, Radioelectronica, Mosca (Russia).
Christiansen T., Nielsen J., 2002, “Production of extracellular protease and glucose uptake in Bacillus clausii in steady-state and transient continuous cultures”, J Biotechnol 97(3): 265-273.
Clauditz A., Resch A., Wieland K. P., Peschel A., Götz F., 2006, “Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress”, Infect Immun 74(8): 4950-4953.
Cleary S. F., 1990a, “Cellular effects of radiofrequency electromagnetic fields”, in Gandhi O. P. (ed.), Biological effects and medical applications of electromagnetic energy, Prentice Hall, Englewood Cliffs (Stati Uniti).
Cleary S. F., 1990b, “Biological effects of radiofrequency electromagnetic fields”, in Gandhi O. P. (ed.), Biological effects and medical applications of electromagnetic energy, Prentice Hall, Englewood Cliffs (Stati Uniti).
Cleary S. F., 1995, “Effects of radiofrequency radiation on mammalian cells and biomolecules in vitro”, in Blank M. (ed.), Electromagnetic fields: biological interactions and mechanisms, Advances in Chemistry (vol. 250), Washington (Stati Uniti).
Cleary S. F., Cao G., Liu L.-M., 1996, “Effects of isothermal 2450 MHz microwave radiation on the mammalian cell cycle: comparison with effects of isothermal 27 MHz radiofrequency radiation exposure”, Bioelectrochem Bioenerg 39(2): 167-173.
Cleary S. F., Liu L.-M., Merchant R. E., 1990a, “Glioma proliferation modulated in vitro by isothermal radiofrequency radiation exposure”, Radiat Res 121(1): 38-45.
Cleary S. F., Liu L.-M., Merchant R. E., 1990b, “In vitro lymphocyte proliferation induced by radio-frequency electromagnetic radiation under isothermal conditions”, Bioelectromagnetics 11(1): 47-56.
Commissione delle Comunità Europee, 2000, “Libro bianco sulla sicurezza alimentare”, Bruxelles (Belgio).
Costerton L. W., 2005, “Il ruolo dei biofilm in sistemi naturali, ingegnerizzati e patologici”, Biofilm Microbici 2005, I Workshop Nazionale, Istituto Superiore di Sanità, Roma, 20-21 giugno 2005.
Costerton J. W., Lewandowski Z., Caldwell D. E., Korber D. R., Lappin-Scott H. M., 1995, “Microbial biofilms”, Annu Rev Microbiol 49: 711-745.
Crawford L. M., Ruff E. H., 1996, “A review of the safety of cold pasteurization through irradiation”, Food Contr 7(2): 87-97.

BIBLIOGRAFIA
- 138 -
Cui L., Ma X., Sato K., Okuma K., Tenover F. C., Mamizuka E. M., Gemmell C. G., Kim M. N., Ploy M. C., El Sohl N., Ferraz V., Hiramatsu K., 2003, “Cell wall thickening is a common feature of Vancomycin resistance in Staphylococcus aureus”, J Clin Microbiol 41(1): 5-14.
Culkin K. A., Fung D. Y. C., 1975, “Destruction of Escherichia coli and Salmonella
typhimurium in microwave cooked soups”, J Milk Food Technol 38(1): 8-15.
D’Andrea J. A., De Witt J. R., Emmerson R. Y., Bailey C., Strensaas S., Gandhi O. P., 1986a, “Intermittent exposure of rats to 2450 MHz microwaves at 2.5 mW/cm2: behavioral and physiological effects”, Bioelectromagnetics 7(3): 315-328
D’Andrea J. A., De Witt J. R., Gandhi O. P., Strensaas S., Lords J. L., Nielson H. C., 1986b, “Behavioral and physiological effects of chronic 2,450-MHz microwave irradiation of the rat at 0.5 mW/cm2”, Bioelectromagnetics 7(1): 45-56.
D’Andrea J. A., Gandhi O. P., Lords J. L., Durney C. H., Johnson C. C., Astle L., 1979, “Physiological and behavioral effects of chronic exposure to 2450-MHz microwaves”, J
Microwave Power Electromagn Energy 14(4): 351-362.
Dardalhon M., Averbeck D., Berteaud A. J., 1980, “Determination of a thermal equivalent of millimeter microwaves in living cells”, J Microw Power 14(4): 307-312.
Dardalhon M., Averbeck D., Berteaud A. J., 1981, “Studies on possible genetic effects of microwaves in procaryotic and eucaryotic cells”, Radiat Environ Biophys 20(1): 37-51.
Dardanoni L., Torregrossa M. V., Zanforlin L., 1985, “Millimeter wave effects on Candida
albicans cells”, J Bioelectricity 4: 171-176.
Davis B. D., Dulbecco R., Eisen H. N., Ginsberg H. S., Wood W. B., 1973, “Microbiology” (6a ed.), Harper & Row, Hagerstown (Stati Uniti).
De Beer D., Stoodley P., 2006, “Microbial biofilms”, in Dworkin M., Falkow S., Rosenberg E., Schleifer K. H., Stackebrandt E. (ed.), The procaryotes (vol. 1: Symbiotic associations, biotechnology, applied microbiology), Springer, New York (Stati Uniti).
Del Seppia C., Ghione S., Luschi P., Papi F., 1995, “Exposure to oscillating magnetic fields influences sensitivity to electrical stimuli. I. Experiments on pigeons”, Bioelectromagnetics 16(5): 290-294.
den Hollander J. A., 1975, “Radical pair substitution in chemically induced dynamic nuclear polarization. Co-operative effects”, Chem Phys 10(1): 167-184.
Delincée H., 1998, “Detection of food treated with ionizing radiation”, Trends Food Sci
Technol 9(2): 73-82.
Denizci A. A., Kazan D., Abeln E. C. A., Erarslan A., 2003, “Newly isolated Bacillus clausii GMBAE 42: an alkaline protease producer capable to grow under higly alkaline conditions”, J Appl Microbiol 96(2): 320-327.

BIBLIOGRAFIA
- 139 -
Deutch J. M., 1972, “Theory of chemically induced dynamic polarization in thin films”, J
Chem Phys 56(12): 6076-6081.
Dharmarha V., 2008, “The majority of deaths from Listeria food poisoning are in individuals with compromised immune systems: pregnant women, newborns, the elderly, and the immunosupressed”, in Brandriff K. M., A focus on Listeria monocytogenes, Food Safety Research Information Office, Beltsville (Stati Uniti).
Donnellan M., McKenzie D. R., French P. W., 1997, “Effects of exposure to electromagnetic radiation at 835 MHz on growth, morphology and secretory characteristics of a mast cell analogue, RBL-2H3”, Cell Biol Int 21(7): 427-439.
Dreyfuss M. S., Chipley J. R., 1980, “Comparison of effects of sublethal microwave radiation and conventional heating on the metabolic activity of Staphylococcus aureus”, Appl Environ Microbiol 39(1): 13-16.
Duc L. H., Hong H. A., Barbosa T. M., Henriques A. O., Cutting S. M., 2004, “Characterization of bacillus probiotics available for human use”, Appl Environ
Microbiol 70(4): 2161-2171.
Durney C. H., Massoudi H., Iskander M. F., 1986, “Radiofrequency radiation dosimetry handbook” (4a ed.), U. S. Air Force School of Aerospace Medicine, San Antonio (Stati Uniti).
Dutta S. K., Subramanian A., Ghosh B., Parshad R., 1984, “Microwave radiation-induced calcium efflux from human neuroblastoma cells in culture”, Bioelectromagnetics 5(1): 71-78.
Dykes G. A., Dworaczek M., 2002, “Influence of interactions between temperature, ferric ammonium citrate and glycine betaine on the growth of Listeria monocytogenes in a defined medium”, Lett Appl Microbiol 35(6): 538-542.
Elder J. A., 1987, “Radiofrequency radiation acrivities and issues: a 1986 perspective”, Health Phys 53(6): 607-611.
Elder J. A., Cahill D. F. (ed.), 1984, Biological effects of radiofrequency radiation, U. S. Environmental Protection Agency, The Research Triangle Park (Stati Uniti).
Evans G. T., Fleming P. D. III, Lawler R. G., 1973, “Hydrodynamic theory of CIDEP and CIDNP”, J Chem Phys 58(5): 2071-2078.
Fellows P. J., 2000, “Food processing technology – Principles and practice” (2a ed.), CRC Press, Boca Raton (Stati Uniti).
Fesenko E. E., Gluvstein A. Y., 1995, “Changes in the state of water, induced by radiofrequency electromagnetic fields”, FEBS Lett 367(1): 53-55.
Fesenko E. E., Geletyuk V. I., Kazachenko V. N., Chemeris N. K., 1995, “Preliminary microwave irradiation of water solutions changes their channel-modifying activity”, FEBS Lett 366(1): 49-52.

BIBLIOGRAFIA
- 140 -
Feynman R. P., 1992, “There’s plenty of room at the bottom”, J Microelectromech Syst 1(1): 60-66.
Fleming D. W., Cochi S. L., MacDonald K. L., Brondum J., Hayes P. S., Plikaytis B. D., Holmes M. B., Audurier A., Broome C. V., Reingold A. L., 1985, “Pasteurized milk as a vehicle of infection in an outbreak of listeriosis”, N Engl J Med 312(7): 404-407.
French P. W., Donnellan M., McKenzie D. R., 1997, “Electromagnetic radiation at 835 MHz changes the morphology and inhibits proliferation of a human astrocytoma cell line”, Bioelectrochem Bioenerg 43(1): 13-18.
Fröhlich H., 1980, “The biological effects of microwaves and related questions”, Adv
Electron Electron Phys 53: 85-152.
Fröhlich H. (ed.), 1988, “Biological coherence and response to external stimuli”, Springer-Verlag, Berlino (Germania).
Furia L., Hill D. W., Gandhi O. P., 1986, “Effect of millimeter-wave irradiation on growth of Saccharomyces cerevisiae”, IEEE Trans Biomed Eng 33(11): 993-999.
Galli Volonterio A., 2005, “Microbiologia degli alimenti”, Casa Editrice Ambrosiana, Milano.
Gandhi O. P., 1987, “The ANSI radiofrequency safety standard: its rationale and some problems”, IEEE Eng Med Biol Mag 6(1): 22-25.
Gaunt I. F., 1986, “Food irradiations – safety aspects”, Proc Inst Food Sci Technol 19(4): 171-174.
Geacintov N. E., Pope M., Fox S., 1970, “Magnetic field effects on photo-enhanced currents in organic crystals”, J Phys Chem Solids 31(6): 1375-1379.
Geacintov N. E., Pope M., Vogel F., 1969, “Effect of magnetic field on the fluorescence of tetracene crystals: exciton fission”, Phys Rev Lett 22(12): 593-596.
Geletyuk V. I., Kazachenko V. N., Chemeris N. K., Fesenko E. E., 1995, “Dual effects of microwaves on single Ca2+-activated K+ channels in cultured kidney cells Vero ”, FEBS
Lett 359(1): 85-88.
Genigeorgis C., Carniciu M., Dutulescu D., Farver T. B., 1991, “Growth and survival of Listeria monocytogenes in market cheeses at 4 to 30 degrees C”, J Food Prot 54(9): 662-668.
Getchell B. E., 1935, “Electric pasteurization of milk”, Agric Eng 16: 408-410.
Geveke D. J., Kozempel M., Scullen O. J., Brunkhorst C., 2002, “Radio frequency energy effects on microorganisms in foods”, Innovat Food Sci Emerg Technol 3(2): 133-138.
Golant M. B., 1989, “Resonance effect of coherent electromagnetic radiations of millimeter range of waves on the living organisms”, Biofizika 34(6): 1004-1014.

BIBLIOGRAFIA
- 141 -
Golant M. B., Kuznetsov A. P., Boszhanova T. P., 1994, “Mechanisms of synchronization of the yeast cell culture by the action of EHF radiation”, Biofizika 39: 490-495.
Goldblith S. A., Wang D. I. C., 1967, “Effect of microwaves on Escherichia coli and Bacillus subtilis”, Appl Environ Microbiol 15(6): 1371-1375.
Gos P., Eicher B., Kohli J., Heyer W.-D., 1997, “Extremely high frequency electromagnetic fields at low power density do not affect the division of exponential phase Saccharomyces cerevisiae cells”, Bioelectromagnetics 18(2): 142-155.
Gray M. L., Killinger A. H., 1966, “Listeria monocytogenes and listeric infection”, Bacteriol
Rev 30: 309-382.
Green D. H., Wakeley P. R., Page A., Barnes A., Baccigalupi L., Ricca E., Cutting S. M., 1999, “Characterization of two bacillus probiotics”, Appl Environ Microbiol 65(9): 4288-4291.
Grissom C. B., 1992, “Magnetic field effects on enzymatic reactions”, Symposium on Magnetic Field and Spin Effects in Chemistry, University of Konstanz, Konstanz (Germania), 26-31 luglio 1992.
Grundler W., Kaiser F., 1992, “Experimental evidence for coherent excitations correlated with cell growth”, Nanobiology 1: 163-176.
Grundler W., Keilmann F., Fröhlich H., 1977, “Resonant growth rate response of yeast cells irradiated by weak microwaves”, Phys Lett A 62(6): 463-466.
Grundler W., Keilmann F., Putterlik V., Strube D., 1982, “Resonance like dependence of yeast growth rate on microwave frequencies”, Br J Cancer 45: 206-208.
Grundler W., Jentzsch U., Keilmann F., Putterlik V., 1988, “Resonant cellular effects of low intensity microwave”, in Fröhlich H. (ed.), Biological coherence and response to external stimuli, Springer-Verlag, Berlino (Germania).
Gründling A., Burrack L.S., Bouwer H. G. A., Higgins D. E., 2004, “Listeria monocytogenes regulates flagellar motility gene expression through MogR, a transcriptional repressor required for virulence”, Proc Natl Acad Sci USA 101(33): 12318-12323.
Haberkorn R., 1977, “Theory of magnetic field modulation of radical recombination reactions. 1”, Chem Phys 19(2): 165-179.
Halliday D., Resnick R., Walker J., 2006, “Fondamenti di fisica” (6a ed.), Casa Editrice Ambrosiana, Milano.
Harvey Pirie J. H., 1940, “Listeria: change of name of a genus of bacteria”, Nature 145(3368): 264.
Henrici A. T., 1933, “Studies of freshwater bacteria. I. A direct microscopic technique”, J
Bacteriol 25: 277-287.
Hofmann G. A., 1985, “Deactivation of microorganisms by an oscillating magnetic field”, Brevetto mondiale WO 85/02094, Maxwell Laboratories Inc., San Diego (Stati Uniti).

BIBLIOGRAFIA
- 142 -
Holland S., Alfonso E., Gelender H., Heidemann D., Mendelsohn A., Ullman S., Darlene Miller M. A., 1987, “Corneal ulcer due to Listeria monocytogenes”, Cornea 6(2): 144-146.
International Radiation Protection Association, 1988, “Guidelines on limits of exposure to radiofrequency electromagnetic fields in the frequency renge from 100 kHz to 300 GHz. International Non-Ionizing Radiation Committee of the International Radiation Protection Association”, Health Phys 54(1): 115-123.
Jay J. M., Loessner M. J., Golden D. A., 2009, “Microbiologia degli alimenti”, Springer-Verlag, Milano.
Johnson R. C., Merrifield R. E., 1970, “Effects of magnetic fields on the mutual annihilation of triplet excitons in anthracene crystals”, Phys Rev B 1(2): 896-902.
Joint FAO/WHO Food Standars Programme, 2006, “Understanding the Codex Alimentarius”, World Hearth Organization, Food and Agriculture Organization of the United Nations, Roma.
Josephson E. S., 1983, “An historical review of food irradiation”, J Food Saf 5(4): 161-189.
Josephson E. S., Thomas M. H., Calhoun W. K., 1975, “Effects of treatment of foods with ionizing radiation”, in Harris R. S., Karmas E. (ed.), Nutritional evaluation of food processing, Avi Publishing Co., Westport (Stati Uniti).
Justeen D. R., 1979, “Behavioural and psychological effects of microwave radiation”, Bull N
Y Acad Med 55: 1058-1078.
Kaiser F., 1995, “Coherent oscillations – their role in the interaction of weak ELM-fields with cellular systems”, Neural Netw World 5(5): 751-762.
Kamau D. N., Doores S., Pruitt K. M., 1990, “Enhanced thermal destruction of Listeria
monocytogenes and Staphylococcus aureus by the lactoperoxidase system”, Appl
Environ Microbiol 56(9): 2711-2716.
Kataev A. A., Alexandrov A. A., Tikhonova L. L., Berestovsky G. N., 1993, “Frequency-dependent effects of the electromagnetic millimeter waves on the ion currents in the cell membrane of Nitellopsis: nonthermal action”, Biofizika 38(3): 446-462.
Kavaliers M., Ossenkopp K.-P., Prato F. S., Carson J. J. L., 1994, “Opioid systems and the biological effects of magnetic fields”, in Frey A. (ed.), On the nature of electromagnetic field interactions with biological systems, CRC Press, Boca Raton (Stati Uniti).
Khizhnyak E. P., Ziskin M. C., 1996, “Temperature oscillations in liquid media caused by continuous (nonmodulated) millimeter wavelength electromagnetic irradiation”, Bioelectromagnetics 17(3): 223-229.
Kinosita K. Jr, Tsong T. Y., 1977, “Formation and resealing of pores of controlled sizes in human erythrocite membrane”, Nature 268(5619): 438-441.

BIBLIOGRAFIA
- 143 -
Kluytmans J., Van Belkum A., Verbrugh H., 1997, “Nasal carriage of Staphylococcus
aureus: epidemiology, underlying mechanisms, and associated risks”, Clin Microbiol
Rev 10(3): 505-520.
Knorr D., Geulen M., Grahl T., Sitzmann W., 1994, “Food application of high electric field pulses”, Trends Food Sci Technol 5(3): 71-75.
Kobayashi T., Kageyama Y., Sumitomo N., Saeki K., Shirai T., Ito S., 2005, “Contribution of a salt bridge triad to the thermostability of a highly alkaline protease from an alkaliphilic bacillus strain”, World J Microbiol Biotechnol 21(6-7): 961-967.
Kozempel M., Scullen O. J., Cook R., Whiting R., 1997, “Preliminary investigation using a batch flow process to determine bacteria destruction by microwave energy at low temperature”, Lebensm Wiss U Technol 30: 691-696.
Kraybill H. F., 1982, “Effect of processing on nutritive value of food: irradiation”, in Rechcigl M. (ed.), Handbook of nutritive value of processed food (vol. 1), CRC Press, Boca Raton (Stati Uniti).
Kudryashova V. A., Zavizion V. A., Khurgin Y. I., 1995, “Effects of stabilization and destruction of water structure by amino acids”, 10th Russian Symposium “Millimeter Waves in Medicine and Biology, Mosca (Russia), 24-26 aprile 1995.
Kunjilwar K. K., Behari J., 1993, “Effect of amplitude-modulated radio frequency radiation on cholinergic system of developing rats”, Brain Res 601(1-2): 321-324.
Kwee S., Raskmark P., 1998, “Changes in cell proliferation due to environmental non-ionizing radiation: 2. Microwave radiation”, Bioelectrochem Bioenerg 44(2): 251-255.
La Placa M., 2005, “Principi di microbiologia medica” (10a ed.), Esculapio, Bologna.
Levenspiel O., 1984, “Chemical reactor omnibook”, Oregon OSU Book Stores, Corvallis (Stati Uniti).
Levina M. Z., Veselago I. A., Belaya T. I., Gapochka L. D., Mantrova G. M., Yakovleva M. N., 1989, “Influence of low-intensity VHF irradiation on growth and development of protozoa cultures”, in Devyatkov N. D. (ed.), Millimeter waves in medicine and biology, Radioelectronica, Mosca (Russia).
Liburdy R. P., 1992, “Biological interactions of cellular systems with time-varying magnetic fields”, in Magin R. L., Liburdy R. P., Persson B. (ed.), Biological effects and safety aspects of nuclear magnetic resonance imaging and spectroscopy, Annals of the New York Academy of Sciences (vol. 649), New York (Stati Uniti).
Litovitz T. A., Krause D., Mullins J. M., 1991, “Effect of coherence time of the applied magnetic field on ornithine decarboxylase activity”, Biochem Biophys Res Commun 178(3): 862-865.

BIBLIOGRAFIA
- 144 -
Litovitz T. A., Krause D., Penafiel M., Elson E. C., Mullins J. M., 1993, “The role of coherence time in the effect of microwaves on ornithine decarboxylase activity”, Bioelectromagnetics 14(5): 395-403.
Litvinov G. S., Gridina N. Y., Dovbeshko G. I., Berezhinskiĭ L. I., Lisitsa M. P., 1994, “Millimeter wave effect on blood plasma solution”, Electro- Magnetobiol 13(2): 167-174.
Liu G. Y., Essex A., Buchanan J. T., Datta V., Hoffman H. M., Bastian J. F., Fierer J., Nizet V., 2005, “Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity”, J Exp Med 202(2): 209-215.
Lowry F., 2008, “Live Listeria vaccine proves safe against end-stage cervical ca in human trial”, Ob Gyn News 43(10): 2.
Lowy F. D., 1998, “Staphylococcus aureus infections”, N Engl J Med 339(8): 520-532.
Madigan M. T., Martinko J. M., 2007, “Brock. Biologia dei microrganismi” (vol. 1: Microbiologia generale), Casa Editrice Ambrosiana, Milano.
Maitra S. K., Nachum R., Pearson F. C., 1986, “Establishment of beta-hydroxy fatty acids as chemical marker molecules for bacterial endotoxin by gas chromatography-mass spectrometry”, Appl Environ Microbiol 52(3): 510-514.
Martin G. S., Mannino D. M., Eaton S., Moss M., 2003, “The epidemiology of sepsis in the United States from 1979 through 2000”, N Engl J Med 348: 1546-1554.
Mazzoldi P., Nigro M., Voci C., 2002, “Elementi di fisica. Elettromagnetismo, onde”, Edises, Napoli.
McArthur J. V., 2006, “Microbial ecology: an evolutionary approach”, Elsevier, Amsterdam (Paesi Bassi).
Merrifield R. E., Avakian P., Groff R. P., 1969, “Fission of singlet excitons into pairs of triplet excitons in tetracene crystals”, Chem Phys Lett 3(3): 155-157.
Merritt J. H., Chamness A. F., Shelton W. W., 1982, “Attempts to alter 45Ca2+ binding to brain tissue with pulse-modulated microwave energy”, Bioelectromagnetics 3(4): 475-478.
Mertens B., Knorr D., 1992, “Developments of non-thermal processes for food preservation”, Food Technol 46(5): 124-133.
Michaelson S. M., 1991, “Biological effects of radiofrequency radiation: concepts and criteria”, Health Phys 61(1): 3-14.
Michaelson S. M., Lin J. C., 1987, “Biological effects and health implications of radiofrequency radiation”, Plenum Publishing Corporation, New York (Stati Uniti).
Monod J., 1949, “The growth of bacterial cultures”, Annu Rev Microbiol 3: 371-394.

BIBLIOGRAFIA
- 145 -
Moore H. A., Raymond R., Fox M., Galsky A. G., 1979, “Low-intensity microwave radiation and the virulence of Agrobacterium tumefaciens strain B6”, Appl Environ
Microbiol 37(1): 127-130.
Mozumder A., 1968, “Theory of neutralization of an isolated ion pair: application of the method of prescribed diffusion to random walk in a Coulomb field”, J Chem Phys 48(4): 1659-1665.
Murray E. G. D., Webb R. E., Swann M. B. R., 1926, “A disease of rabbits characterized by a large mononuclear leucocytosis, caused by a hitherto undescribed bacillus Bacterium
monocytogenes (n. sp.)”, J Pathol Bacteriol 29: 407-439.
Murray P. R., Rosenthal K. S., 2008, “Medical microbiology” (6a ed.), Mosby, St. Louis (Stati Uniti).
Noyes R. M., 1954, “A treatment of chemical kinetics with special applicability to diffusion controlled reactions”, J Chem Phys 22(8): 1349-1359.
Olsen C. M., 1965, “Microwaves inhibit bread mold”, Food Eng 37(7): 51-53.
Organizzazione Mondiale della Sanità, 1977, “Wholesomeness of irradiated foods”, WHO Technical Report Series (vol. 604), Ginevra (Svizzera).
Organizzazione Mondiale della Sanità, 1981, “Wholesomeness of irradiated foods”, WHO Technical Report Series (vol. 659), Ginevra (Svizzera).
Organizzazione Mondiale della Sanità, 1993, “Environmental Health Criteria 137 (1993): electromagnetic fields (300 Hz – 300 GHz)”, Ginevra (Svizzera).
Osepchuk J. M., Petersen R. C., 1997, “Comments on “Resonance effect of millimeter waves in the power range from 10(-19) to 3 X 10(-3) W/cm2 on Escherichia coli cells at different concentrations”, Belyaev et al., Bioelectromagnetics, 17:312-321 (1996), Bioelectromagnetics 18(7): 527-528, discussion 529.
Pakhomov A. G., Prol H. K., Mathur S. P., Akyel Y., Campbell C. B. G., 1997a, “Search for frequency-specific effects of millimeter-wave radiation on isolated nerve function”, Bioelectromagnetics 18(4): 324-334.
Pakhomov A. G., Prol H. K., Mathur S. P., Akyel Y., Campbell C. B. G., 1997b, “Frequency-specific effects of millimeter-wavelength electromagnetic radiation in isolated nerve”, Electromagn Biol Med 16(1): 43-57.
Pakhomov A. G., Akyel Y., Pakhomova O. N., Stuck B. E., Murphy M. R., 1998, “Current state and implications of research on biological effects of millimeter waves: a review of the literature”, Bioelectromagnetics 19(7): 393-413.
Pakhomova O. N., Pakhomov A. G., Akyel Y., 1997, “Effect of millimeter waves on UV-induced recombination and mutagenesis in yeast”, Bioelectrochem Bioenerg 43(2): 227-232.

BIBLIOGRAFIA
- 146 -
Palleroni N. J., 1984, “Genus I. Pseudomonas migula 237 AL”, in Krieg N. R., Holt J. G. (ed.), Bergey’s manual of systematic bacteriology, William & Wilkins Co., Baltimora (Stati Uniti).
Parsek M. R., Singh P. K., 2003, “Bacterial biofilms: an emerging link to disease pathogenesis”, Ann Rev Microbiol 57: 677-701.
Paustian T., 2000, “Nitrogen assimilation”, University of Wisconsin-Madison website, http://dwb.unl.edu/Teacher/NSF/C11/C11Links/www.bact.wisc.edu/microtextbook/metabolism/NitrogenAssim.html.
Penafiel L. M., Litovitz T., Krause D., Desta A., Mullins J. M., 1997, “Role of modulation on the effect of microwaves on ornithine decarboxylase activity in L929 cells”, Bioelectromagnetics 18(2): 132-141.
Phelan A. M., Lange D. G., Kues H. A., Lutty G. A., 1992, “Modification of membrane fluidity in melanin-containing cells by low-level microwave radiation”, Bioelectromagnetics 13(2): 131-146.
Potel J., 1952, “Zur granulomatosis infantiseptica”, Zentr Bakteriol Parasitenk 158: 329-331.
Postow E., Swicord M. L., 1996, “Modulated fields and “window” effects”, in Polk C., Postow E. (ed.), Handbook of biological effects of electromagnetic fields (2a ed.), CRC Press, New York (Stati Uniti).
Pothakamury U. R., Barbosa-Canovas G. V., Swanson B. G., 1993, “Magnetic-field inactivation of microorganisms and generation of biological changes”, Food Technol 47(12): 85-93.
Pothakamury U. R., Monsalve-Gonzalez A., Barbosa-Canovas G. V., Swanson B. G., 1995, “Inactivation of Escherichia coli and Staphylococcus aureus in model foods by pulsed electric field technology”, Food Res Int 28(2): 167-171.
Prato F. S., Kavaliers M., Carson J. J. L., 1996, “Behavioural evidence that magnetic field effects in the land snail, Cepaea nemoralis, might not depend on magnetite or induced electric currents”, Bioelectromagnetics 17(2): 123-130.
Rai S., Singh U. P., Mishra G. D., Singh S. P., Samarketu S. P., 1994a, “Effect of water’s microwaves power density memory on fungal spore germination”, Electro- Magnetobiol 13(3): 247-252.
Rai S., Singh U. P., Mishra G. D., Singh S. P., Samarketu S. P., 1994b, “Additional evidence of stable EMF-induced changes in water revealed by fungal spore germination”, Electro-
Magnetobiol 13(3): 253-259.
Ramaswamy H. S., Singh R. P., 1997, “Sterilization process engineering”, in Valentas K. J., Rotstein E., Singh R. P. (ed.), Handbook of food engineering practice, CRC Press, Boca Raton (Stati Uniti).

BIBLIOGRAFIA
- 147 -
Ramaswamy H. S., Voort F. R., Ghazala S., 1989, “An analysis of TDT and Arrhenius methods for handling process and kinetic data”, J Food Sci 54(5): 1322-1326.
Ramaswamy V., Cresence V. M., Rejitha J. S., Lekshmi M. U., Dharsana K. S., Prasad S. P., Vijila H. M., 2007, “Listeria – review of epidemiology and pathogenesis”, J Microbiol
Immunol Infect 40(1): 4-13.
Ray B., 2004, “Fundamental of food microbiology”, CRC Press, London (Regno Unito).
Ray S., Behari J., 1990, “Physiological changes in rats after exposure to low levels of microwave”, Radiat Res 123: 199-202.
Razi Naqvi K., Mork K. J., Waldenstrøm S., 1980, “Diffusion-controlled reaction kinetics. Equivalence of the particle pair approach of Noyes and the concentration gradient approach of Collins and Kimball”, J Phys Chem 84(11): 1315-1319.
Rebrova T. B., 1992, “Effect of millimeter-range electromagnetic radiation on the vital activity of microrganisms”, Millimetr Volny Biol Med 1: 37-47.
Repacholi M. H., 1998, “Low-level exposure to radiofrequency electromagnetic fields: health effects and research needs”, Bioelectromagnetics 19(1): 1-19.
Roberts N. J. Jr, Michaelson S. M., Lu S.-T., 1986, “The biological effects of radiofrequency radiation: a critical review and recommendations”, Int J Radiat Biol 50(3): 379-420.
Rojavin M. A., Cowan A., Radzievsky A. A., Ziskin M. C., 1998, “Antipruritic effect of millimeter waves in mice: evidence for opioid involvement”, Life Sci 63(18): PL251-PL257.
Rojavin M. A., Ziskin M. C., 1995, “Effect of millimeter waves on survival of UVC-exposed Escherichia coli”, Bioelectromagnetics 16(3): 188-196.
Ryakovskaya M. L., Shtemler V. M., 1983, “Absorption of electromagnetic waves of millimeter range in biological preparations with plane-layer structure”, in Devyatkov N. D. (ed.), Effect of nonthermal action of millimeter radiation on biological subjects, USSR Academy of Sciences, Mosca (Russia).
Ryan K. J., Ray C. G. (ed.), 2004, “Sherris Medical Microbiology” (4a ed.), Mc-Graw Hill, New York (Stati Uniti).
Ryser E. T., Marth E. H. (ed.), 2007, “Listeria, listeriosis, and food safety” (3a ed.), CRC Press, Boca Raton (Stati Uniti).
Salikhov K. M., Molin Y. N., Sagdeev R. Z., Buchachenko A. L., 1984, “Spin polarization and magnetic effects in radical reactions”, Elsevier, Amsterdam (Paesi Bassi).
Samarketu S. P., Singh S. P., Jha R. K., 1996, “Effect of direct modulated microwave modulation frequencies exposure on physiology of Cyanobacterium Anabaena
doliolum”, 1996 Asia-Pacific Microwave Conference, New Delhi (India), 17-20 dicembre 1996.

BIBLIOGRAFIA
- 148 -
Sarvarov F. S., Salikhov K. M., 1976, “Theory of spin-dependent recombination of radicals in homogeneous solution”, React Kinet Catal Lett 4(1): 33-41.
Sauer K., Camper A. K., Ehrlich J. W., Costerton J. W., Davies D. G., 2002, “Pseudomonas
aeruginosa displays multiple phenotypes during development as a biofilm”, J Bacteriol 184(4): 1140-1154.
Schlech W. F., 2001, “Foodborne listeriosis”, Clin Infect Dis 31(3): 770-775.
Schlech W. F., Lavigne P. M., Bortolussi R. A., Allen A. C., Haldane E. V., Wort A. J., Hightower A. W., Johnson S. E., King S. H., Nicholls E. S., Broome C. V., 1983, “Epidemic listeriosis – evidence for transmission by food”, New Engl J Med 308(4): 203-206.
Schulten K., Wolynes P. G., 1978, “Semiclassical description of electron spin motion in radicals including the effect of electron hopping”, J Chem Phys 68(7): 3292-3297.
Senesi S., Celandroni F., Tavanti A., Ghelardi E., 2001, “Molecular characterization and identification of Bacillus clausii strains marketed for use in oral bacteriotherapy”, Appl
Environ Microbiol 67(2): 834-839.
Shelton W. W. Jr, Merritt J. H., 1981, “In vitro study of microwave effects on calcium efflux in rat brain tissue”, Bioelectromagnetics 2(2): 161-167.
Shestopalova N. G., Makarenko B. I., Golovina L. N., Timoshenko Y. P., Baeva T. I., Vinokurova L. V., Miroshnichenko V. S., 1995, “Modification of synchronizing effect of millimeter waves on first mitoses by different temperature regimens of germination”, 10th Russian Symposium “Millimeter Waves in Medicine and Biology, Mosca (Russia), 24-26 aprile 1995.
Shub G. M., Luniova I. O., Denisova S. G., Ostrovskii N. V., 1995, “Effects of millimeter waves on bacteria in in vitro and in vivo experiments”, 10th Russian Symposium “Millimeter Waves in Medicine and Biology, Mosca (Russia), 24-26 aprile 1995.
Singh S. P., Rai S., Rai A. K., Tiwari S. P., Singh S. S., Samarketu, Abraham J., 1994, “Athermal physiological effects of microwaves on a cynobacterium Nostoc muscorum: evidence for EM-memory bits in water”, Med Biol Eng Comput 32(2): 175-180.
Stagg R. B., Thomas W. J., Jones R. A., Adey W. R., 1997, “DNA synthesis and cell proliferation in C6 glioma and primary glial cells exposed to a 836.55 MHz modulated radiofrequency field”, Bioelectromagnetics 18(3): 230-236.
Steele J. H., Engel R. E., 1992, “Radiation processing of food”, J Am Vet Med Assoc 201(10): 1522-1529.
Steiner U. E., Ulrich T., 1989, “Magnetic field effects in chemical kinetics and related phenomena”, Chem Rev 89(1): 51-147.
Sterna L., Ronis D., Wolfe S., Pines A., 1980, “Viscosity and temperature dependence of the magnetic isotope effect”, J Chem Phys 73(11): 5493-5499.

BIBLIOGRAFIA
- 149 -
Stevenson M. H., 1994, “Identification of irradiated foods”, Food Technol 48(5): 141-144.
Tambiev A. K., Kirikova N. N., 1992, “Perspectives on the application of millimeter-range electromagnetic radiation in photobiotechnology”, Millimetr Volny Biol Med 1: 48-54.
Tambiev A. K., Kirikova N. N., Lapshin O. M., Betzkii O V., Novskova T. A., Nechaev V. M., Petrov I. Y., 1989, “The combined effect of exposure to EMF of millimeter and centimeter wavelength ranges on productivity of microalgae”, in Devyatkov N. D. (ed.), Millimeter waves in medicine and biology, Radioelectronica, Mosca (Russia).
Tauxe R. V., 2001, “Food safety and irradiation: protecting the public from foodborne infections”, Emerg Infect Dis 7(3): 516-521.
Teixeira A, 1992, “Thermal process calculations”, in Heldman D. R., Lund D. B. (ed.), Handbook of food engineering, Marcel Dekker Inc., New York (Stati Uniti).
Tenover F. C., Biddle J. W., Lancaster M. V., 2001, “Increasing resistance to Vancomycin and other glycopeptides in Staphylococcus aureus”, Emerg Infect Dis 7(2): 327-332.
Tiecco G., 2001, “Igiene e tecnologia alimentare”, Il Sole 24 Ore Edagricole, Bologna.
U. S. Food and Drug Administration, 2001, “Foodborne pathogenic microorganisms and natural toxins handbook”, Center for Food Safety and Applied Nutrition, College Park (Stati Uniti).
U. S. National Council on Radiation Protection and Measurement, 1986, “Biological effects and exposure criteria for radiofrequency electromagnetic fields”, Report N. 86, Bethesda (Stati Uniti).
von Hippel A. R., 1954, “Dielectric materials and applications”, Technology Press of MIT, Cambridge (Stati Uniti).
Webb S. J., Booth A. D., 1969, “Absorption of microwaves by microorganisms”, Nature 222(5199): 1199-1200.
Webb S. J., Dodds D. D., 1968, “Inhibition ob bacterial cell growth by 136 gc microwaves”, Nature 218(5139): 374-375.
Webb T., Lang T., 1987, “Food irradiation: the facts”, Thorsons Publishing, Wellingborough (Regno Unito).
Welt M. A., 1985, “Barriers to widespread approval of food irradiation”, J Ind Irradiat
Technol 3(1): 75-86.
Welt B. A., Tong C. H., Rossen J. L., Lund D. B., 1994, “Effect of microwave radiation on inactivation of Clostridium sporogenes (PA 3679) spores”, Appl Environ Microbiol 60(2): 482-488.
Wexler H., Oppenheim J. D., 1979, “Isolation, characterization, and biological properties of an endotoxin-like material from the gram-positive organism Listeria monocytogenes”, Infect Immun 23: 845-857.

BIBLIOGRAFIA
- 150 -
Whitelock-Jones L., Carswell J., Rassmussen K. C., 1989, “Listeria pneumonia. A case report.”, S Afr Med J 75: 188-189.
Wilkinson V. M., Gould G. W., 1996, “Food irradiation: a reference guide”, Woodhead Publishing, Cambridge (Regno Unito).
Zavizion V. A., Kudryashova V. A., Khurgin Y. I., 1994, “Effect of alpha-amino acids on the interaction of millimeter-wave radiation with water”, Millimetr Volny Biol Med 3: 46-52.
Zhang Q., Monsalve-Gonzalez A., Barbosa-Canovas G. V., Swanson B. G., 1994, “Inactivation of E. coli and S. cerevisiae by pulsed electric fields under controlled temperature conditions”, Trans ASAE 37: 581-587.
Zhang Q., Barbosa-Canovas G. V., Swanson B. G., 1995, “Engineering aspects of pulsed electric field pasteurization”, J Food Eng 25(2): 261-281.
Zientara G. P., Freed J. P., 1979, “Theory of chemically-induced dynamic spin polarization. IV. Low field effects”, J Chem Phys 70(3): 1359-1370.
Zimmermann U., Pilwat G., Riemann F., 1974, “Dielectric breakdown of cell membranes”, Biophys J 14(11): 881-899.



- 153 -
Ringraziamenti
Si ringraziano tutti coloro che a vario titolo, direttamente o indirettamente, con il
loro supporto sia materiale che di conoscenza ed esperienza, hanno contribuito alla
realizzazione del lavoro, e in particolare: il prof. Raffaello Pompei e la dott.ssa
Samuela Laconi del Dipartimento di Scienze e Tecnologie Biomediche (sezione di
Microbiologia Applicata e Tecnologie Biomediche) dell’Università degli Studi di
Cagliari; il prof. Giuseppe Mazzarella, il prof. Giovanni Martines e il dott. Gianluca
Gatto del Dipartimento di Ingegneria Elettrica ed Elettronica dell’Università degli
Studi di Cagliari; il dott. Germano Orrù del Dipartimento di Chirurgia e Scienze
Odontostomatologiche dell’Università degli Studi di Cagliari.
Un ringraziamento doveroso e particolare va infine al prof. Renzo Carta che,
guidando tutta la mia attività di ricerca con sapienza, dedizione e passione, ha
costantemente profuso in me il giusto entusiasmo e mi ha permesso di realizzare, nel
corso degli anni, un’esperienza eccezionale in termini di maturazione scientifica,
professionale e personale; spero sinceramente di avere con lui, in futuro, ulteriori ed
interessanti occasioni di collaborazione.


















