Unique Toxin Profile of a Mediterranean Ostreopsis cf. ovata Strain: HR LC-MS n Characterization of...
10
Unique Toxin Profile of a Mediterranean Ostreopsis cf. ovata Strain: HR LC-MS n Characterization of Ovatoxin-f, a New Palytoxin Congener Patrizia Ciminiello, † Carmela Dell’Aversano,* ,† Emma Dello Iacovo, † Ernesto Fattorusso, † Martino Forino, † Luciana Tartaglione, † Cecilia Battocchi, § Rita Crinelli, ‡ Elisa Carloni, ‡ Mauro Magnani, ‡ and Antonella Penna § † Department of Chemistry of Natural Products, University of Napoli Federico II, via D. Montesano 49, Napoli 80131, Italy ‡ Section of Biochemistry and Molecular Biology and § Section of Environmental Biology, Department of Biomolecular Sciences, University of Urbino, via A. Saffi 2, Urbino (PU) 61029, Italy * S Supporting Information ABSTRACT: Currently, the benthic dinoflagellate Ostreopsis cf. ovata represents a serious concern to human health in the whole Mediterranean basin due to the production of palytoxin congeners, a putative palytoxin and ovatoxins (ovatoxin-a, -b, -c, -d/-e), listed among the most potent marine toxins. High resolution liquid chromatography-mass spectrometry (HR LC- MS) based investigation of a North Western Adriatic strain of Ostreopsis cf. ovata collected at Portonovo (Italy) in 2008 is reported herein. Toxin profile was different from those previously reported for other O. cf. ovata, both qualitatively and quantitatively. For the first time, ovatoxin-a did not dominate the toxin profile, and a new palytoxin congener, here named ovatoxin-f, was detected. Ovatoxin-f and its elemental formula present C 2 H 4 more than ovatoxin-a. HR CID MS n experiments allowed us to restrict structural differences between ovatoxin-a and -f to the region between C-95 and C-102, a region not previously been described to be modified in other palytoxins. Ovatoxin-f represents the major component of the toxin profile of the analyzed strain accounting for 50% of the total toxin content, while ovatoxin-a, the dominant toxin in most of the Mediterranean O. cf. ovata strains we have analyzed so far, is the second major component of the toxin profile (23%). Thus, the presence of ovatoxin-f should be taken into account when monitoring programs for palytoxin-like compounds in microalgae and/ or seawater are carried out. ■ INTRODUCTION Tropical and subtropical benthic dinoflagellates belonging to genus Ostreopsis have been detected in temperate waters such as those of the Mediterranean Sea since the late ‘70s. 1,2 However, only in the past decade have blooms of O. confronta (cf.) ovata and less frequently of O. cf. siamensis been occurring along the Mediterranean coasts of Europe and North Africa and the Atlantic coasts of Portugal 3-5 resulting in benthic biocenosis suffering and associated human health effects. The most extensive sanitary events associated with Ostreopsis blooms occurred in Italy in 2001 6 and 2005, 7 in Spain in 2004 and 2006, 8,9 in France in the period 2006-2009, 10,11 and in Croatia in 2010 (Pfannkuchen M. et al., personal communication). During these events, up to 200 people exposed to marine aerosols during recreational or working activities required medical attention due to symptoms such as fever, cough, dyspnea, sore throat, rhinorrhea, etc. 12,13 High concentration of Ostreopsis spp. (in most cases O. cf. ovata) were recorded in seawater both using traditional and innovative molecular methods, 14 and symptoms in humans disappeared as bloom intensity decreased. 12 To date, resolving the taxonomy of Ostreopsis species based only on morphology has been di fficult due to the morphological variability of both field material and cultured specimens. 15 Further, none of the original isolates from tropical areas from which species of Ostreopsis species were described has yet been sequenced for the genotype assignment. Given that for many dinoflagellate genera there is a discrepancy between morphology-based taxonomy and genetic studies, 15 the species names applied to strains of different genetic lineages must be treated with caution. It is therefore advisable to use the designation O. cf. ovata when referring to isolates from the Mediterranean sea. Liquid chromatography-mass spectrometry (LC-MS) stud- ies on O. cf. ovata demonstrated it to produce several congeners of palytoxin (Figure 1), one of the most potent nonprotein Received: February 27, 2012 Published: April 13, 2012 Article pubs.acs.org/crt © 2012 American Chemical Society 1243 dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243-1252
Transcript of Unique Toxin Profile of a Mediterranean Ostreopsis cf. ovata Strain: HR LC-MS n Characterization of...

Unique Toxin Profile of a Mediterranean Ostreopsis cf. ovata Strain:HR LC-MSn Characterization of Ovatoxin-f, a New Palytoxin CongenerPatrizia Ciminiello,† Carmela Dell’Aversano,*,† Emma Dello Iacovo,† Ernesto Fattorusso,†
Martino Forino,† Luciana Tartaglione,† Cecilia Battocchi,§ Rita Crinelli,‡ Elisa Carloni,‡ Mauro Magnani,‡
and Antonella Penna§
†Department of Chemistry of Natural Products, University of Napoli Federico II, via D. Montesano 49, Napoli 80131, Italy‡Section of Biochemistry and Molecular Biology and §Section of Environmental Biology, Department of Biomolecular Sciences,University of Urbino, via A. Saffi 2, Urbino (PU) 61029, Italy
*S Supporting Information
ABSTRACT: Currently, the benthic dinoflagellate Ostreopsis cf.ovata represents a serious concern to human health in the wholeMediterranean basin due to the production of palytoxincongeners, a putative palytoxin and ovatoxins (ovatoxin-a, -b,-c, -d/-e), listed among the most potent marine toxins. Highresolution liquid chromatography−mass spectrometry (HR LC-MS) based investigation of a North Western Adriatic strain ofOstreopsis cf. ovata collected at Portonovo (Italy) in 2008 isreported herein. Toxin profile was different from those previouslyreported for other O. cf. ovata, both qualitatively andquantitatively. For the first time, ovatoxin-a did not dominatethe toxin profile, and a new palytoxin congener, here namedovatoxin-f, was detected. Ovatoxin-f and its elemental formulapresent C2H4 more than ovatoxin-a. HR CID MSn experimentsallowed us to restrict structural differences between ovatoxin-a and -f to the region between C-95 and C-102, a region notpreviously been described to be modified in other palytoxins. Ovatoxin-f represents the major component of the toxin profile ofthe analyzed strain accounting for 50% of the total toxin content, while ovatoxin-a, the dominant toxin in most of theMediterranean O. cf. ovata strains we have analyzed so far, is the second major component of the toxin profile (23%). Thus, thepresence of ovatoxin-f should be taken into account when monitoring programs for palytoxin-like compounds in microalgae and/or seawater are carried out.
■ INTRODUCTIONTropical and subtropical benthic dinoflagellates belonging togenus Ostreopsis have been detected in temperate waters such asthose of the Mediterranean Sea since the late ‘70s.1,2 However,only in the past decade have blooms of O. confronta (cf.) ovataand less frequently of O. cf. siamensis been occurring along theMediterranean coasts of Europe and North Africa and theAtlantic coasts of Portugal3−5 resulting in benthic biocenosissuffering and associated human health effects. The mostextensive sanitary events associated with Ostreopsis bloomsoccurred in Italy in 20016 and 2005,7 in Spain in 2004 and2006,8,9 in France in the period 2006−2009,10,11 and in Croatiain 2010 (Pfannkuchen M. et al., personal communication).During these events, up to 200 people exposed to marineaerosols during recreational or working activities requiredmedical attention due to symptoms such as fever, cough,dyspnea, sore throat, rhinorrhea, etc.12,13 High concentration ofOstreopsis spp. (in most cases O. cf. ovata) were recorded inseawater both using traditional and innovative molecular
methods,14 and symptoms in humans disappeared as bloomintensity decreased.12
To date, resolving the taxonomy of Ostreopsis species basedonly on morphology has been difficult due to themorphological variability of both field material and culturedspecimens.15 Further, none of the original isolates from tropicalareas from which species of Ostreopsis species were describedhas yet been sequenced for the genotype assignment. Giventhat for many dinoflagellate genera there is a discrepancybetween morphology-based taxonomy and genetic studies,15
the species names applied to strains of different genetic lineagesmust be treated with caution. It is therefore advisable to use thedesignation O. cf. ovata when referring to isolates from theMediterranean sea.Liquid chromatography−mass spectrometry (LC-MS) stud-
ies on O. cf. ovata demonstrated it to produce several congenersof palytoxin (Figure 1), one of the most potent nonprotein
Received: February 27, 2012Published: April 13, 2012
Article
pubs.acs.org/crt
© 2012 American Chemical Society 1243 dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−1252
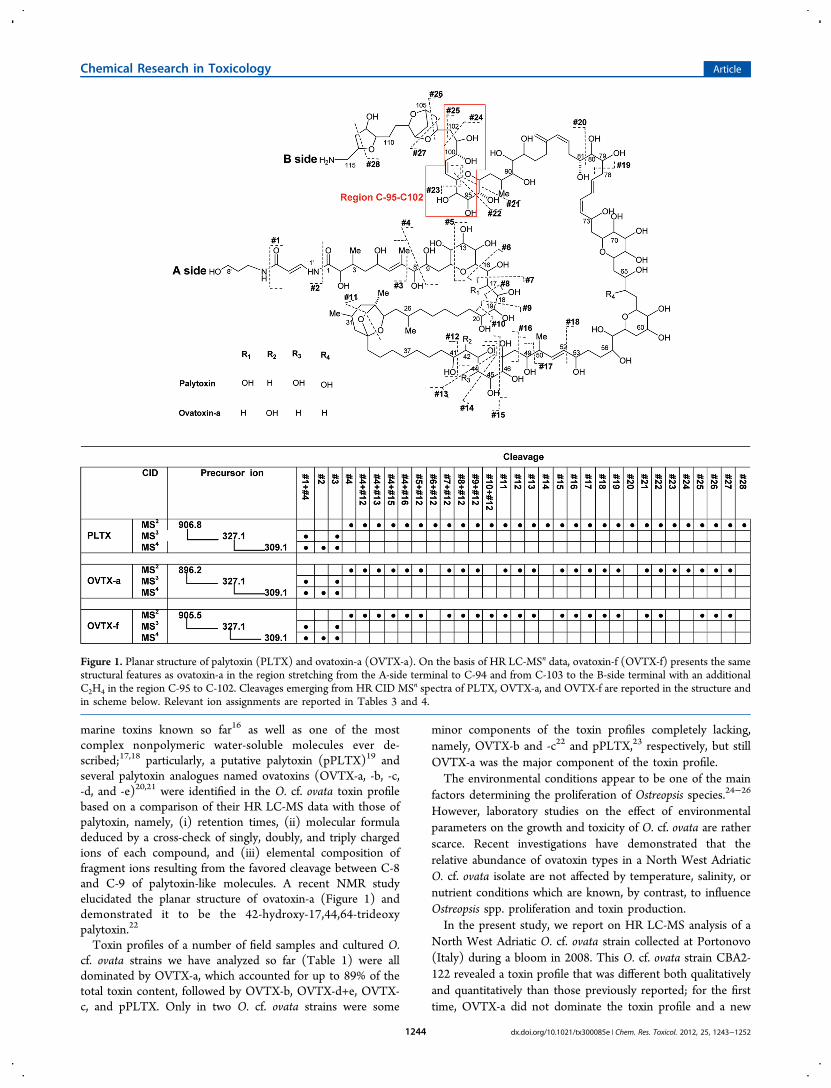
marine toxins known so far16 as well as one of the mostcomplex nonpolymeric water-soluble molecules ever de-scribed;17,18 particularly, a putative palytoxin (pPLTX)19 andseveral palytoxin analogues named ovatoxins (OVTX-a, -b, -c,-d, and -e)20,21 were identified in the O. cf. ovata toxin profilebased on a comparison of their HR LC-MS data with those ofpalytoxin, namely, (i) retention times, (ii) molecular formuladeduced by a cross-check of singly, doubly, and triply chargedions of each compound, and (iii) elemental composition offragment ions resulting from the favored cleavage between C-8and C-9 of palytoxin-like molecules. A recent NMR studyelucidated the planar structure of ovatoxin-a (Figure 1) anddemonstrated it to be the 42-hydroxy-17,44,64-trideoxypalytoxin.22
Toxin profiles of a number of field samples and cultured O.cf. ovata strains we have analyzed so far (Table 1) were alldominated by OVTX-a, which accounted for up to 89% of thetotal toxin content, followed by OVTX-b, OVTX-d+e, OVTX-c, and pPLTX. Only in two O. cf. ovata strains were some
minor components of the toxin profiles completely lacking,namely, OVTX-b and -c22 and pPLTX,23 respectively, but stillOVTX-a was the major component of the toxin profile.The environmental conditions appear to be one of the main
factors determining the proliferation of Ostreopsis species.24−26
However, laboratory studies on the effect of environmentalparameters on the growth and toxicity of O. cf. ovata are ratherscarce. Recent investigations have demonstrated that therelative abundance of ovatoxin types in a North West AdriaticO. cf. ovata isolate are not affected by temperature, salinity, ornutrient conditions which are known, by contrast, to influenceOstreopsis spp. proliferation and toxin production.In the present study, we report on HR LC-MS analysis of a
North West Adriatic O. cf. ovata strain collected at Portonovo(Italy) during a bloom in 2008. This O. cf. ovata strain CBA2-122 revealed a toxin profile that was different both qualitativelyand quantitatively than those previously reported; for the firsttime, OVTX-a did not dominate the toxin profile and a new
Figure 1. Planar structure of palytoxin (PLTX) and ovatoxin-a (OVTX-a). On the basis of HR LC-MSn data, ovatoxin-f (OVTX-f) presents the samestructural features as ovatoxin-a in the region stretching from the A-side terminal to C-94 and from C-103 to the B-side terminal with an additionalC2H4 in the region C-95 to C-102. Cleavages emerging from HR CID MSn spectra of PLTX, OVTX-a, and OVTX-f are reported in the structure andin scheme below. Relevant ion assignments are reported in Tables 3 and 4.
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521244

palytoxin congener, ovatoxin-f, was detected; the HR LC-MSn
based characterization of ovatoxin-f is presented herein.
■ MATERIALS AND METHODSCultures of O. cf. ovata CBA2-122. The O. cf. ovata strain
CBA2-122 was originally isolated from a field sample collected atPortonovo on October 2008 during a bloom of O. cf. ovata. PortonovoBay is a rocky beach with stones and artificial barriers along the beach,located in the Marche Region (Ancona), in the central West Adriaticcoast of Italy. During the bloom of O. cf. ovata local newspapers andbroadcast news reported cases of airborne syndrome in a few peopleattending the beaches, such as swimmers and divers, but not infishermen nor in operators who sampled Ostreopsis. In addition, noofficial source of information about cases of irritation to eyes, cough,and fever were reported, and the information was obtained frompeople visiting the rocky beach. Cultures were grown in 1 L glassbottles containing 0.6 L sterilized f/4 medium27 with an initial cellamount of 3.0 × 104 cells. The temperature was set at 23 ± 1 °C. Lightwas provided by cool white fluorescent bulbs (photon flux of 100 μEm−2 s−1) on a 14−10 h light dark cycle. Culture samples were fixedwith Lugol’s iodine28 and counted using the Utermhol method.29 Atotal volume of 8 L of culture was harvested by centrifugation at 4,000rpm for 15 min at room temperature; a pellet containing 3.0 × 106
cells was stored at −80 °C until chemical analysis.Taxonomic Molecular Identification by a PCR Based Assay.
Genomic DNA was extracted from a 10 mL culture of O. cf. ovataCBA2-122 in logarithmic growth phase using DNeasy Plant Kit(Qiagen, Valencia, CA, USA), according to the manufacturer’sinstructions. PCR amplification using genus- and species-specificprimers was performed as described in Penna et al.30 and Battocchi etal.31
Extraction. The O. cf. ovata cell pellet (3.0 × 106 cells) wasextracted by adding 20 mL of methanol/water (1:1 v/v) and 0.2%acetic acid and sonicating for 30 min in pulse mode, while cooling inan ice bath. The mixture was centrifuged at 3000g for 30 min, thesupernatant was decanted, and the pellet was washed twice more with20 mL of methanol/water (1:1, v/v) and 0.2% acetic acid. The extractswere combined, and the volume was adjusted to 60 mL with extractingsolvent. A 250 μL aliquot of the crude extract was filtered through a0.45 μm filter and directly analyzed by high resolution liquidchromatography−mass spectrometry (HR LC-MS).
HR LC-MS and MSn Experiments. HR LC-MS and MSn analyseswere performed on a hybrid linear ion trap LTQ Orbitrap XL Fouriertransform mass spectrometer (FTMS) equipped with an ESI IONMAX source (Thermo-Fisher, San Jose, CA, USA) and coupled to anAgilent 1100 LC binary system (Palo Alto, CA, USA) including asolvent reservoir, online degasser, binary pump, and thermostatedautosampler. A 3 μm Gemini C18 column (150 × 2.00 mm;Phenomenex, Torrance, CA, USA) was eluted at 0.2 mL/min withwater (eluent A) and 95% acetonitrile/water (eluent B), bothcontaining 30 mM acetic acid (control conditions). A gradient elution(20−50% B over 20 min, 50−80% B over 10 min, 80−100% B in 1min, and held for 5 min) was used. Injection volume was 5 μL.
HR full MS experiments (positive ions) were acquired either in therange m/z 800−1400 or m/z 2000−3000 at resolution setting 30.000to 100.000. The following source settings were used: spray voltage = 4kV, capillary temperature = 290 °C, capillary voltage = 45 V, sheath gas= 35 and auxiliary gas = 1 (arbitrary units), and tube lens voltage = 165V (m/z 800−1400) or 250 V (m/z 2000−3000).
HR collision induced dissociation (CID) MS2 experiments wereacquired on [M + H + Ca]3+ ions of palytoxin and ovatoxins (Table 2)by using 25% collision energy (CE). HR CID MS3 and MS4
experiments were carried out on m/z 905.5 > 327.1 and m/z 905.5> 327.1 > 309.1 of ovatoxin-f at CE = 25% and 20%, respectively. A60000 resolving power, an activation Q of 0.250, and an activationtime of 30 ms were used in all cases.
Calculation of elemental formulas in full MS and CID MSn spectrawas performed by using the monoisotopic ion peak of each ion cluster.T
able
1.Percentage(%
)andCon
tent
onaPer
CellBasis(pg/Cell)of
PutativePalytoxin
(pPLT
X)andOvatoxins
(OVTX-a,-b,-c,-d+-e,and-f)Con
tained
inFieldand
CulturedO.cf.ovataStrainsAnalyzedSo
Fara
pPLT
XOVTX-a
OVTX-b
OVTX-c
OVTX-d+-e
OVTX-f
cultured/
field
samplingsite
(sea,region,
country),d
ate(code)
%pg/cell
%pg/cell
%pg/cell
%pg/cell
%pg/cell
%pg/
cell
cultured
Portonovo(Adriatic,M
arche,Italy),2
008(C
BA2-122)
0.3
0.1
238
176
2.4
0.8
6.7
250
17cultured
Num
ana(Adriatic,M
arche,Italy),2006(O
OAN0601)2
1,22,25,26
0.6
0.2
5418
279
62
124
ndnd
field
Ancona(Adriatic,M
arche,Italy),2
0093
7nd-3
nd-2
55−65
8−49
14−20
2−15
nd-6
nd-4
15−21
3−13
ndnd
field
Trieste(Adriatic,F
riuliVeneziaGiulia,Italy),2009
23nd
nd77−89
45−64
5−14
4−8
nd-1
nd-0.7
4−9
3−5
ndnd
field
Rovinji(Adriatic,C
roatia),2010
(M.P
fannkuchen,p
ersonalcommun.)
0.6−
0.9
0.2−
0.7
58−64
14−46
18−25
5−13
4−5
0.9−
210−12
2−8
ndnd
cultured
Num
ana(Adriatic,M
arche,Italy),2008(O
OAN0816)2
23
77nd
nd20
ndnd
cultured
Num
ana(Adriatic,M
arche,Italy),2007(O
OAN0709)2
2Bari(Adriatic,
Puglia,Italy),2008
(OOAB0801)2
2Latin
a(T
yrrhenian,
Lazio,
Italy),
2007
(OOTL0
707)
22
1−4
53−57
24−26
4−5
12−14
ndnd
and
=notdetected.
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521245

A mass tolerance of 5 ppm was used, and the isotopic pattern of eachion cluster was considered.Extracted ion chromatograms (XIC) were obtained from the HR
full MS spectra by selecting the most abundant and the monoisotopicion peaks of the [M+2H−H2O]
2+ and [M+H+Ca]3+ ion clusters ofeach compound (Table 2). A mass tolerance of 5 ppm was used. Thechromatographic peaks were identified by comparing their retentiontimes and associated HR full MS and CID MS2 spectra to those ofOVTX-a, OVTX-b, OVTX-c, OVTX-d and OVTX-e, and putativePLTX contained in a reference O. cf. ovata extract (OOAN0601)previously characterized21 and analyzed under the same experimentalconditions. In quantitative analyses (triplicate injection), peak areaswere measured and interpolated within the calibration curve ofpalytoxin standard (Wako Chemicals GmbH, Neuss, Germany) at fivelevels of concentrations (50, 25, 12.5, 6.25, and 3.13 ng/mL). Linearityof the calibration curve was indicated by a correlation coefficient (R2)of 0.9980. Because of the lack of standards for ovatoxins, their molarresponses were assumed to be the same as that of palytoxin, on thebasis of structural similarities between palytoxin and ovatoxins. Theobtained results were corrected based on recovery percentage of theextraction procedure (98%).20 Measured limit of quantitation (LOQ)and limit of detection (LOD) for the palytoxin standard under the LC-MS conditions used were 3.13 and 1.9 ng/mL, respectively.
■ RESULTS
Molecular Analysis. PCR amplification based assay of theOstreopsis sp. CBA2-122 strain using genus- and species-specificprimers confirmed the O. cf. ovata genotype identification.Toxin Profile of O. cf. ovata CBA2-122. Crude extract of
the cultured O. cf. ovata strain CBA2-122 from Portonovo(Italy) was analyzed by HR LC-MS in positive full MS and CIDMSn modes versus both a palytoxin standard and a previouslycharacterized culture extract of Adriatic O. cf. ovata(OOAN0601), which was used as reference in containing apPLTX and all the ovatoxins known so far, namely, OVTX-a,-b, -c, d-, and -e.21
Figure 2 shows the total ion chromatogram (TIC) of theCBA2-122 extract and extracted ion chromatograms (XIC) ofthe principal components of the toxin profile obtained selectingthe [M+2H−H2O]
2+ and [M+H+Ca]3+ ions of each com-pound. Recently, we demonstrated that palytoxin and ovatoxinstend to form adducts with various divalent cations and, aboveall, with calcium, which results in triply charged [M+H+Ca]3+
ions dominating their MS spectra.32 Chromatographic separa-tion was not complete; however, the use of a high resolutionMS setting (60.000) and a mass tolerance of 5 ppm in ionassignment, as well as the further confirmation provided by HRCID MSn experiments, avoided ambiguity in the identificationof individual compounds.Toxins eluting under the minor peak of the TIC were
identified as pPLTX, OVTX-b, -c, d-, and -e by comparison oftheir full MS and CID MS2 spectra with those previouslyreported for the reference sample (OOAN0601).21 Analysis offull MS spectrum associated with the most abundant peak of
the TIC (at 11.30 min) highlighted the presence of two sets ofions of different relative intensity, each including singly-,doubly-, and triply charged ions of a palytoxin-like compound:the first set of ions could be assigned to ovatoxin-a (Figure 3a),while the second and more intense set of ions suggested thepresence in the extract of a new palytoxin congener elutingalmost at the same retention time as ovatoxin-a (11.32 min vs11.24 min, respectively); we named it ovatoxin-f (Figure 3b).Because of chromatographic overlapping of OVTX-a and -f, theMS spectrum of each compound unavoidably contains lessintense ions belonging to the other compound.As recently reviewed by Ciminiello et al,33 the high number
of multiply charged ions contained in electrospray (ESI) MSspectra of palytoxin-like compounds, including protonated ions,adducts with monovalent (Na+, K+) and divalent (Ca2+, Mg2+,etc.) cations,32 and ions due to multiple water losses, may leadto ambiguity in assigning true molecular weights to newanalogues. High resolution MS is a prerequisite, but, even using
Table 2. Elemental Formula, Retention Times, and Principal Ions (m/z) of HR Full MS Spectra of Putative Palytoxin (pPLTX)and Ovatoxins (OVTX-) Used in Quantitative Analyses
pPLTX OVTX-a OVTX-b OVTX-c OVTX-d/-e OVTX-f
formula C129H223N3O54 C129H223N3O52 C131H227N3O53 C131H227N3O54 C129H223N3O53 C131H227N3O52
Rt (min) 10.56 11.24 10.98 10.65 10.89 11.32[M+2H−H2O]
2+ most intense 1331.7436 1315.7498 1337.7623 1345.7584 1323.7456 1329.7650monoisotopic 1331.2417 1315.2480 1337.2595 1345.2566 1323.2439 1329.2624
[M+H+Ca]3+ most intense 906.8167 896.1572 910.8318 916.1628 901.4884 905.4976monoisotopic 906.4851 895.8255 910.4976 915.8286 901.1533 905.1632
Figure 2. (a) Total ion chromatogram (TIC) of the cultured O. cf.ovata CBA2-122 and (b) extracted ion chromatograms (XIC) of theprincipal components of the toxin profile.
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521246
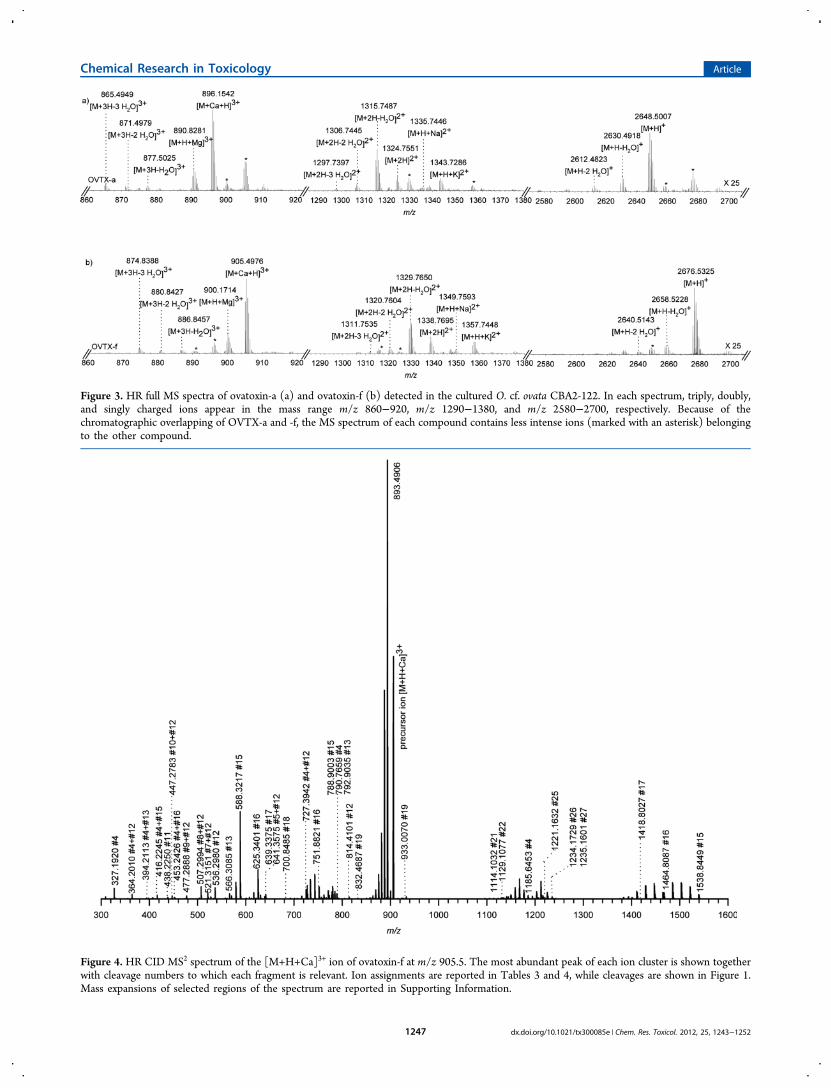
Figure 3. HR full MS spectra of ovatoxin-a (a) and ovatoxin-f (b) detected in the cultured O. cf. ovata CBA2-122. In each spectrum, triply, doubly,and singly charged ions appear in the mass range m/z 860−920, m/z 1290−1380, and m/z 2580−2700, respectively. Because of thechromatographic overlapping of OVTX-a and -f, the MS spectrum of each compound contains less intense ions (marked with an asterisk) belongingto the other compound.
Figure 4. HR CID MS2 spectrum of the [M+H+Ca]3+ ion of ovatoxin-f at m/z 905.5. The most abundant peak of each ion cluster is shown togetherwith cleavage numbers to which each fragment is relevant. Ion assignments are reported in Tables 3 and 4, while cleavages are shown in Figure 1.Mass expansions of selected regions of the spectrum are reported in Supporting Information.
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521247
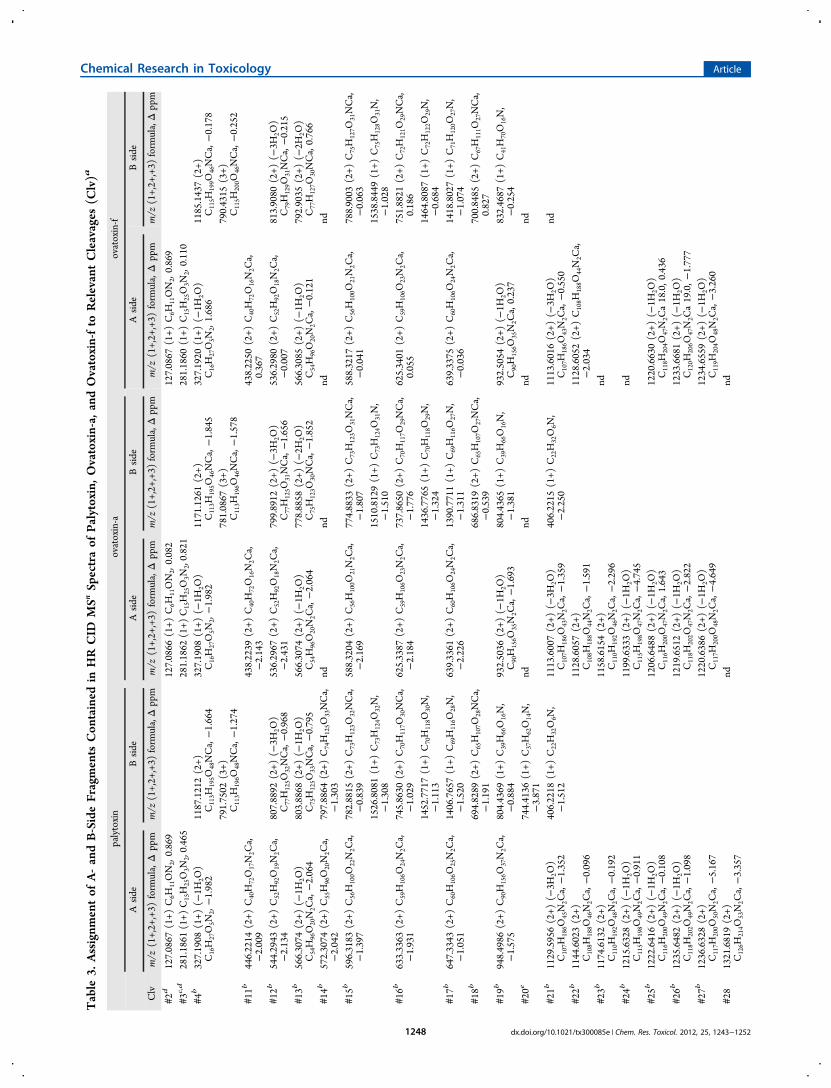
Table
3.Assignm
entof
A-andB-SideFragments
Con
tained
inHRCID
MSn
Spectraof
Palytoxin,Ovatoxin-a,andOvatoxin-fto
RelevantCleavages
(Clv)a
palytoxin
ovatoxin-a
ovatoxin-f
Aside
Bside
Aside
Bside
Aside
Bside
Clv
m/z
(1+,2+
,+3)
form
ula,Δ
ppm
m/z
(1+,2+
,+3)
form
ula,Δppm
m/z
(1+,2+
,+3)
form
ula,Δ
ppm
m/z
(1+,2+
,+3)
form
ula,Δppm
m/z
(1+,2+
,+3)
form
ula,Δ
ppm
m/z
(1+,2+
,+3)
form
ula,Δppm
#2d
127.0867
(1+)
C6H
11ON
2,0.869
127.0866
(1+)
C6H
11ON
2,0.082
127.0867
(1+)
C6H
11ON
2,0.869
#3c,d
281.1861
(1+)
C15H
25O
3N2,0.465
281.1862
(1+)
C15H
25O
3N2,0.821
281.1860
(1+)
C15H
25O
3N2,0.110
#4b
327.1908
(1+)
(−1H
2O)
C16H
27O
5N2,−1.982
1187.1212(2+)
C113H
195O
48NCa,−1.664
327.1908
(1+)
(−1H
2O)
C16H
27O
5N2,−1.982
1171.1261(2+)
C113H
195O
46NCa,−1.845
327.1920
(1+)
(−1H
2O)
C16H
27O
5N2,1.686
1185.1437(2+)
C115H
199O
46NCa,−0.178
791.7502
(3+)
C113H
196O
48NCa,−1.274
781.0867
(3+)
C113H
196O
46NCa,−1.578
790.4315
(3+)
C115H
200O
46NCa,−0.252
#11b
446.2214
(2+)
C40H
72O
17N
2Ca,
−2.009
438.2239
(2+)
C40H
72O
16N
2Ca,
−2.143
438.2250
(2+)
C40H
72O
16N
2Ca,
0.367
#12b
544.2943
(2+)
C52H
92O
19N
2Ca,
−2.134
807.8892
(2+)
(−3H
2O)
C77H
125O
32NCa,−0.968
536.2967
(2+)
C52H
92O
18N
2Ca,
−2.431
799.8912
(2+)
(−3H
2O)
C77H
125O
31NCa,−1.656
536.2980
(2+)
C52H
92O
18N
2Ca,
−0.007
813.9080
(2+)
(−3H
2O)
C79H
129O
31NCa,−0.215
#13b
566.3074
(2+)
(−1H
2O)
C54H
96O
20N
2Ca,−2.064
803.8868
(2+)
(−1H
2O)
C75H
125O
33NCa,−0.795
566.3074
(2+)
(−1H
2O)
C54H
96O
20N
2Ca,−2.064
778.8858
(2+)
(−2H
2O)
C75H
123O
30NCa,−1.852
566.3085
(2+)
(−1H
2O)
C54H
96O
20N
2Ca,−0.121
792.9035
(2+)
(−2H
2O)
C77H
127O
30NCa,0.766
#14b
572.3074
(2+)
C55H
96O
20N
2Ca,
−2.042
797.8864
(2+)
C74H
125O
33NCa,
−1.303
ndnd
ndnd
#15b
596.3183
(2+)
C56H
100O
22N
2Ca,
−1.397
782.8815
(2+)
C73H
123O
32NCa,
−0.839
588.3204
(2+)
C56H
100O
21N
2Ca,
−2.169
774.8833
(2+)
C73H
123O
31NCa,
−1.807
588.3217
(2+)
C56H
100O
21N
2Ca,
−0.041
788.9003
(2+)
C75H
127O
31NCa,
−0.063
1526.8081(1+)
C73H
124O
32N,
−1.308
1510.8129(1+)
C73H
124O
31N,
−1.510
1538.8449(1+)
C75H
128O
31N,
−1.028
#16b
633.3363
(2+)
C59H
106O
24N
2Ca,
−1.931
745.8630
(2+)
C70H
117O
30NCa,
−1.029
625.3387
(2+)
C59H
106O
23N
2Ca,
−2.184
737.8650
(2+)
C70H
117O
29NCa,
−1.776
625.3401
(2+)
C59H
106O
23N
2Ca,
0.055
751.8821
(2+)
C72H
121O
29NCa,
0.186
1452.7717(1+)
C70H
118O
30N,
−1.113
1436.7765(1+)
C70H
118O
29N,
−1.324
1464.8087(1+)
C72H
122O
29N,
−0.684
#17b
647.3343
(2+)
C60H
106O
25N
2Ca,
−1.051
1406.7657(1+)
C69H
116O
28N,
−1.520
639.3361
(2+)
C60H
106O
24N
2Ca,
−2.226
1390.7711(1+)
C69H
116O
27N,
−1.311
639.3375
(2+)
C60H
106O
24N
2Ca,
−0.036
1418.8027(1+)
C71H
120O
27N,
−1.074
#18b
694.8289
(2+)
C65H
107O
28NCa,
−1.191
686.8319
(2+)
C65H
107O
27NCa,
−0.539
700.8485
(2+)
C67H
111O
27NCa,
0.827
#19b
948.4986
(2+)
C90H
156O
37N
2Ca,
−1.575
804.4369
(1+)
C39H
66O
16N,
−0.884
932.5036
(2+)
(−1H
2O)
C90H
156O
35N
2Ca,−1.693
804.4365
(1+)
C39H
66O
16N,
−1.381
932.5054
(2+)
(−1H
2O)
C90H
156O
35N
2Ca,0.237
832.4687
(1+)
C41H
70O
16N,
−0.254
#20e
744.4136
(1+)
C37H
62O
14N,
−3.871
ndnd
ndnd
#21b
1129.5956(2+)
(−3H
2O)
C107H
186O
45N
2Ca,−1.352
406.2218
(1+)
C22H
32O
6N,
−1.512
1113.6007(2+)
(−3H
2O)
C107H
186O
43N
2Ca,−1.359
406.2215
(1+)
C22H
32O
6N,
−2.250
1113.6016(2+)
(−3H
2O)
C107H
186O
43N
2Ca,−0.550
nd
#22b
1144.6023(2+)
C108H
188O
46N
2Ca,−0.096
1128.6057(2+)
C108H
188O
44N
2Ca,−1.591
1128.6052(2+)
C108H
188O
44N
2Ca,
−2.034
#23b
1174.6132(2+)
C110H
192O
48N
2Ca,−0.192
1158.6154(2+)
C110H
192O
46N
2Ca,−2.296
nd
#24b
1215.6328(2+)
(−1H
2O)
C115H
198O
49N
2Ca,−0.911
1199.6333(2+)
(−1H
2O)
C115H
198O
47N
2Ca,−4.745
nd
#25b
1222.6416(2+)
(−1H
2O)
C116H
200O
49N
2Ca,−0.108
1206.6488(2+)
(−1H
2O)
C116H
200O
47N
2Ca,1.643
1220.6630(2+)
(−1H
2O)
C118H
204O
47N
2Ca18.0,0
.436
#26b
1235.6482(2+)
(−1H
2O)
C118H
202O
49N
2Ca,−1.098
1219.6512(2+)
(−1H
2O)
C118H
202O
47N
2Ca,−2.822
1233.6681(2+)
(−1H
2O)
C120H
206O
47N
2Ca19.0,−
1.777
#27b
1236.6328(2+)
C117H
200O
50N
2Ca,−5.167
1220.6386(2+)
(−1H
2O)
C117H
200O
48N
2Ca,−4.649
1234.6559(2+)
(−1H
2O)
C119H
204O
48N
2Ca,−3.260
#28
1321.6819(2+)
C126H
214O
53N
2Ca,−3.357
ndnd
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521248

a mass tolerance of 5 ppm, several plausible elemental formulasfor the monoisotopic ion peaks are possible due to the highvalues of the masses of palytoxin-like compounds associatedwith the high number of C, H, N, and O atoms contained intheir elemental formulas. This drawback can be overcomethrough a combined analysis of all the singly, doubly, and triplycharged ions contained in HR full MS spectra. Therefore,combining assignment of the most abundant singly, doubly, andtriply charged ions of full MS spectra of OVTX-f, namely, [M+H]+ at m/z 2676.5122 (monoisotopic at m/z 2675.5283,C131H228N3O52, Δ = −0.015 ppm), [M+H+K]2+ at m/z1357 . 7 4 78 (mono i s o t op i c a t m / z 1357 . 2 4 22 ,C131H228N3O52K, Δ = −2.615 ppm), [M+2H-1H2O]
2+ at m/z 1329.7640 (monoisotopic at m/z 1329.2624, C131H227N3O51,Δ = −0.095 ppm), [M+H+Ca]3+ at m/z 905.5003 (mono-isotopic at m/z 905.1632, C131H228N3O52Ca, Δ = −0.086ppm), and [M+H+Mg]3+ at m/z 900.1746 (monoisotopic atm/z 899.8364 C131H228N3O52Mg, Δ = −1.143 ppm), theelemental formula C131H227N3O52 was assigned to ovatoxin-f.Compared to OVTX-a (C129H223N3O52), the OVTX-felemental formula contains more C2H4 than that of OVTX-a.In order to individuate the region of the molecule where
structural differences between OVTX-a and -f occur, HR LC-MSn spectra of OVTX-f were acquired and analyzed in parallelto those of OVTX-a whose structure was recently determinedthrough NMR.22 In this regard, we recently demonstrated thatmuch structural information can be obtained from the positiveHR MSn spectra of palytoxin and its congeners, as they undergocharacteristic fragmentation at several sites of their backbone.32
Therefore, a comparative analysis of the fragmentation patternof OVTX-f versus that of OVTX-a was accomplished to gainstructural information on ovatoxin-f. In this comparative study,the [M+H+Ca]3+ ion at m/z 905.5 (ovatoxin-f) was used as theprecursor in MS2 experiments, and some of the resultingproduct ions were fragmented within MS3 and MS4 experi-ments.Figure 4 shows the HR CID MS2 spectrum of OVTX-f.
Tables 3 and 4 contain assignments of all the ions of HR CIDMSn spectra of ovatoxin-f to relevant cleavages shown in Figure1 in comparison to those previously reported for ovatoxin-a andpalytoxin, whose standard is commercially available.32 It has tobe noted that, under the used conditions, fragmentation occursat several sites of each palytoxin-like molecule generating foreach cleavage A-side fragments (containing 2 N atoms) and/orB-side fragments (containing 1 N atom) (Table 3). Each A-sideand B-side fragment can appear in MSn spectra as a singly and/or doubly charged ion and is followed by 1 to 7 ions due tosubsequent water losses (mass expansions are shown in FigureS1, Supporting Information); in some cases ions, correspondingto 1−3 water losses from the A- or the B-side fragments are theonly ones that can be detected; a very limited number of ionsare protonated fragments, whereas most of them are calcium-containing fragments. In addition, some internal fragments dueto a combination of two cleavages also occur, containingneither the A-side nor the B-side terminal of the molecule andthus having no N atom in their elemental formula (Table 4).With the only exception of the internal fragment due tocleavage #1 + #4 (Figure 1) which provides information on thepart of the structure ranging from C-8 to the A-side terminal ofthe molecule, all the other internal fragments originate from theinner portion of the molecule stretching from C-9 to C-48(Figure 1), namely, from the B-side fragment of cleavage #4after further fragmentation at cleavage sites #12, #13, #15, andT
able
3.continued
aElem
entalformulae
ofthemono-isotopicionpeaks(m
/z)arereported
with
ioncharge
state(1+,
2+,or3+
)anderrors(Δ
,ppm
).nd
=notdetected.bIons
intheHRCID
MS2
spectraof
the[M
+H+C
a]3+ionofpalytoxin(m
/z906.8),ovatoxin-a(m
/z896.2),and
ovatoxin-f(m
/z905.5),respectively.c IonsintheHRCID
MS3
spectraofpalytoxin(m
/z906.8>327.1),ovatoxin-a(m
/z896.2>327.1),
andovatoxin-f(m
/z905.5>327.1),respectively.dIons
intheHRCID
MS4
spectraofpalytoxin(m
/z906.8>327.1>309.1),ovatoxin-a(m
/z896.2>327.1>309.1),and
ovatoxin-f(m
/z905.5>327.1>
309.1),respectively.e Ionsin
theHRCID
MS2
spectraof
the[M
+2H−H
2O]2
+ionof
palytoxin(m
/z1331.7).
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521249
#16 and from the A-side fragment of cleavage #12 after furtherfragmentation at cleavage sites #5, #7, #8, #9, and #10.Fragmentation pattern of ovatoxin-f closely paralleled that of
ovatoxin-a, suggesting that they share the same backbone;however, there were some key clues in the spectra thatindicated the region where structural differences between thetwo compounds occur. Such information could be obtained bycomparing elemental formulas of fragments of ovatoxin-a withthose of ovatoxin-f for each cleavage (Tables 3 and 4).In the part of the structure ranging from C-1 to C-94 (Figure
1), cleavages occurred at the same sites in both ovatoxin-a andovatoxin-f and generated A-side and internal fragments havingthe same elemental composition in both compounds (Tables 3and 4) as well as B-side fragments presenting C2H4 more inOVTX-f than the relevant fragments of OVTX-a. Thissuggested that ovatoxin-f and ovatoxin-a share the part of thestructure stretching from the A-side terminal to C-94, and thus,the structural difference between them is located in the regionranging from C-95 to the B-side terminal.A comparative analysis of fragmentation occurring in the part
of the structure close to the B-side terminal of the moleculesallowed us to restrict the region of structural difference betweenOVTX-a and OVTX-f to the region stretching from C-95 to C-102. In fact, the fragmentation pattern of the two molecules wasdifferent in this region: fragments due to cleavages #23 and #24in OVTX-a were lacking in OVTX-f, while the A-side fragmentdue to cleavage #25 (between C-102 and C-103), as well asthose due to cleavages #26, and #27, contained C2H4 more inOVTX-f than in OVTX-a.It has to be noted that within palytoxin analogues,22,34−36
including ovatoxin-a, the region close to the B-side terminal hasnever been described to have structural modifications. Inaddition, considering that, compared to palytoxin, ovatoxin-alacks the hydroxyl groups at C-17 and at C-64 and presents anadditional hydroxyl group at C-42, the whole of the obtainedresults suggest that the same structural features could bepresent in ovatoxin-f.For quantitative analysis, extracted ion chromatograms
(Figure 2b) were obtained from full MS spectrum of the
crude extract by selecting the most abundant bi- and trichargedions of ovatoxins (a−f) and palytoxin (Table 2). Because of thelack of pure standards of ovatoxins, their molar response wasassumed to be the same as palytoxin’s, and the calibration curveof palytoxin at four levels of concentration was used toextrapolate quantitative data. Total toxin content of the O. cf.ovata CBA2-122 strain was thus calculated to be 34 pg/cell.Ovatoxin-f resulted in being the major component of the toxinprofile (17 pg/cell, 50%) followed by ovatoxin-a (8 pg/cell,23%), ovatoxin-b (6 pg/cell, 17%), ovatoxin-c (0.8 pg/cell,2.4%), ovatoxin-d+e (2 pg/cell, 6.7%), and putative palytoxin(0.1 pg/cell, 0.3%). This is a unique toxin profile whencompared to the toxin profiles of most O. cf. ovata strains wehave analyzed so far (Table 1) because for the first time,ovatoxin-a does not represent the major component of thetoxin profile.The Ostreopsis isolate CBA2-122 was genetically identified as
O. cf. ovata, clade Atlantic/Mediterranean.3 Thus, the differentovatoxin profiles found among different isolates of O. cf. ovatacould be related to the different haplotypes within the Atlantic/Mediterranean clade based on the concatenated ITS regions,5.8S and LSU ribosomal genes. Studies are in progress in ourlaboratories to investigate this issue.Since the O. cf. ovata CBA2-122 extract was unique in
containing all the ovatoxins known so far, we believed itopportune to evaluate its bioactivity. The obtained results havebeen recently reported (Crinelli et al., unpublished) andshowed that the toxin extract was able to induce an increase inthe levels of mRNAs encoding inflammation-related proteins inprimary human macrophages via signaling through tran-scription factor-κB (NF−κB) and p38 MAPK (mitogenactivated protein kinases). This is the first report providingthe molecular basis of a direct correlation between ovatoxincellular activity and acute inflammatory reactions recorded inindividuals exposed to Ostreopsis species through inhalation orskin contact. In addition, the fact that the overall toxinscontained in the extract, although at a 10-fold higherconcentration, displayed an activity totally overlapping that ofthe palytoxin standard might suggest that chemically similar but
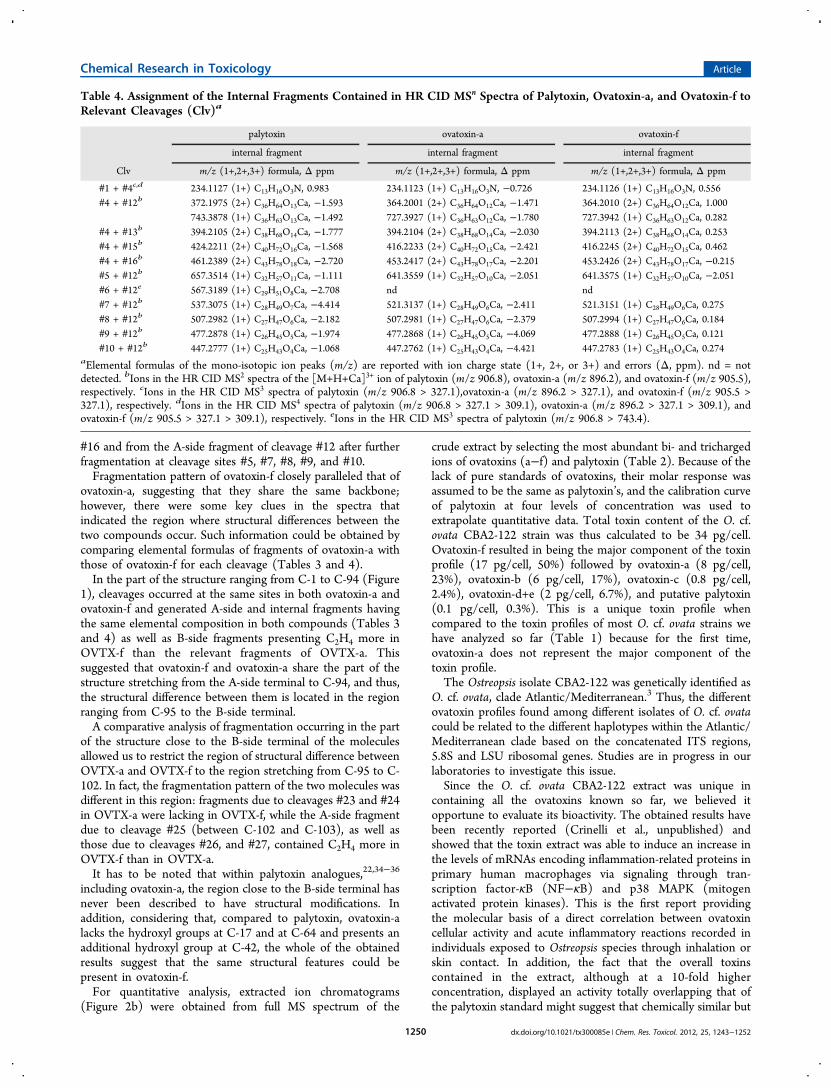
Table 4. Assignment of the Internal Fragments Contained in HR CID MSn Spectra of Palytoxin, Ovatoxin-a, and Ovatoxin-f toRelevant Cleavages (Clv)a
palytoxin ovatoxin-a ovatoxin-f
internal fragment internal fragment internal fragment
Clv m/z (1+,2+,3+) formula, Δ ppm m/z (1+,2+,3+) formula, Δ ppm m/z (1+,2+,3+) formula, Δ ppm
#1 + #4c,d 234.1127 (1+) C13H16O3N, 0.983 234.1123 (1+) C13H16O3N, −0.726 234.1126 (1+) C13H16O3N, 0.556#4 + #12b 372.1975 (2+) C36H64O13Ca, −1.593 364.2001 (2+) C36H64O12Ca, −1.471 364.2010 (2+) C36H64O12Ca, 1.000
743.3878 (1+) C36H63O13Ca, −1.492 727.3927 (1+) C36H63O12Ca, −1.780 727.3942 (1+) C36H63O12Ca, 0.282#4 + #13b 394.2105 (2+) C38H68O14Ca, −1.777 394.2104 (2+) C38H68O14Ca, −2.030 394.2113 (2+) C38H68O14Ca, 0.253#4 + #15b 424.2211 (2+) C40H72O16Ca, −1.568 416.2233 (2+) C40H72O15Ca, −2.421 416.2245 (2+) C40H72O15Ca, 0.462#4 + #16b 461.2389 (2+) C43H78O18Ca, −2.720 453.2417 (2+) C43H78O17Ca, −2.201 453.2426 (2+) C43H78O17Ca, −0.215#5 + #12b 657.3514 (1+) C32H57O11Ca, −1.111 641.3559 (1+) C32H57O10Ca, −2.051 641.3575 (1+) C32H57O10Ca, −2.051#6 + #12e 567.3189 (1+) C29H51O8Ca, −2.708 nd nd#7 + #12b 537.3075 (1+) C28H49O7Ca, −4.414 521.3137 (1+) C28H49O6Ca, −2.411 521.3151 (1+) C28H49O6Ca, 0.275#8 + #12b 507.2982 (1+) C27H47O6Ca, −2.182 507.2981 (1+) C27H47O6Ca, −2.379 507.2994 (1+) C27H47O6Ca, 0.184#9 + #12b 477.2878 (1+) C26H45O5Ca, −1.974 477.2868 (1+) C26H45O5Ca, −4.069 477.2888 (1+) C26H45O5Ca, 0.121#10 + #12b 447.2777 (1+) C25H43O4Ca, −1.068 447.2762 (1+) C25H43O4Ca, −4.421 447.2783 (1+) C25H43O4Ca, 0.274
aElemental formulas of the mono-isotopic ion peaks (m/z) are reported with ion charge state (1+, 2+, or 3+) and errors (Δ, ppm). nd = notdetected. bIons in the HR CID MS2 spectra of the [M+H+Ca]3+ ion of palytoxin (m/z 906.8), ovatoxin-a (m/z 896.2), and ovatoxin-f (m/z 905.5),respectively. cIons in the HR CID MS3 spectra of palytoxin (m/z 906.8 > 327.1),ovatoxin-a (m/z 896.2 > 327.1), and ovatoxin-f (m/z 905.5 >327.1), respectively. dIons in the HR CID MS4 spectra of palytoxin (m/z 906.8 > 327.1 > 309.1), ovatoxin-a (m/z 896.2 > 327.1 > 309.1), andovatoxin-f (m/z 905.5 > 327.1 > 309.1), respectively. eIons in the HR CID MS3 spectra of palytoxin (m/z 906.8 > 743.4).
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521250

not identical compounds could exert their toxic effects via thesame signaling pathways. Thus, in light of these findings, itcould be hypothesized that ovatoxins a−f could indeed differnot in the mechanism of action but rather in their potency.Future experiments will be directed toward testing singleovatoxin types when it is possible to purify them in sufficientamounts to be tested in an in vitro cellular system.
■ CONCLUSIONS
A new palytoxin-like compound, ovatoxin-f, has been detectedin a NW Adriatic O. cf. ovata isolate. Its elemental compositioncompared to that of ovatoxin-a presents additional C2H4 andthe whole of the HR CID MSn data suggests that structuraldifferences between molecules reside in the region between C-95 and C-102.Ovatoxin-f represents the major component of the toxin
profile of the analyzed strain accounting for 50% of the totaltoxin content, while ovatoxin-a, the dominant toxin in most ofthe Mediterranean O. cf. ovata strains we have analyzed sofar,21−23,25,26,37 is the second major component of the toxinprofile (23%). Thus, the presence of ovatoxin-f should be takeninto account when monitoring programs for palytoxin-likecompounds in microalgal cells and/or seawater are carried out.The discovery of a new palytoxin congener in theMediterranean Ostreopsis strain is a major proof that we aredealing with an important emerging and evolving toxicologicalproblem with multiple consequences including the possibility ofMediterranean food chain contamination. Toxic effects exertedby single ovatoxin components should be evaluated both interms of the mechanism of action and potency in order toestablish a possible correlation between toxin profiles and riskassessment for human health.
■ ASSOCIATED CONTENT
*S Supporting InformationMonoisotopic ion peaks of all the fragment ions of the HR CIDMS2 spectra of ovatoxin-a and -f, mass expansions of the HRCID MS2 spectrum of ovatoxin-f. This material is available freeof charge via the Internet at http://pubs.acs.org.
■ AUTHOR INFORMATION
Corresponding Author*Tel: +39-081-678502. Fax: +39-081-678552. E-mail:[email protected].
FundingThis work was supported by PRIN 2009 and by ISPRA Grants.
NotesThe authors declare no competing financial interest.
■ ABBREVIATIONS
HR LC-MS, high resolution liquid chromatography−massspectrometry; CID, collision induced dissociation; PLTX,palytoxin; pPLTX, putative palytoxin; OVTX-a, ovatoxin-a;OVTX-b, ovatoxin-b; OVTX-c, ovatoxin-c; OVTX-d+e,ovatoxin-d+e; OVTX-f, ovatoxin-f; FTMS, Fourier transformmass spectrometry; ESI, electrospray ionization; XIC, extractedion chromatogram; LOQ, limit of quantitation; LOD, limit ofdetection; PCR, polymerase chain reaction; ITS, internaltranscribed spacer; LSU, large subunit; MAPK, mitogenactivated protein kinases; O. cf. ovata, Ostreopsis confronta ovata
■ REFERENCES(1) Taylor, F. J. R. (1979) A Description of the BenthicDinoflagellate Associated with Maitotoxin and Ciguatoxin, IncludingObservations on Hawaiian Material, in Toxic Dinoflagellate Blooms(Taylor, D. L., and Seliger, H. H., Eds) pp 71−77, Elsevier North-Holland, New York.(2) Abboud-Abi Saab, M. (1989) Les dinoflagelles des eaux cotiereslibanaises-especes rares ou nouvelles du phytoplancton marin. Leb. Sci.Bull. 5, 5−16.(3) Penna, A., Fraga, S., Battocchi, C., Casabianca, S., Riobo, P.,Giacobbe, M. G., and Vernesi, C. (2010) A phylogeography study ofthe toxic benthic genus Ostreopsis Schmidt. J. Biogeogr. 37, 830−841.(4) Amorim, A., Veloso, V., and Penna, A. (2010) First detection ofOstreopsis cf. siamensis in Portuguese coastal waters. Harmful AlgaeNews 42, 6−7.(5) Parsons, M. L., Aligizaki, K., Bottein, M.-Y. D., Fraga, S., Morton,S. L., Penna, A., and Rhodes, L. (2012) Gambierdiscus and Ostreopsis:Reassessment of the state of knowledge of their taxonomy, geography,ecophysiology, and toxicology. Harmful Algae 14, 107−129.(6) Casavola, N., Troncone, A., Rizzi, E., Favale, M. G., and Bello, G.(2005). Microalghe marine tossiche nella provincia di Bari:danniambientali, ittiofaunistici, evidenze epidemiologiche, in Diffusione dellefioriture algali tossiche nelle acque italiane: gestione del rischio ed evidenzeepidemiologiche (Mattei D., Melchiorre, S., Messineo, V., and Bruno,M., Eds.) pp. 92−97, Rapporto Istisan 05/29.(7) Brescianini, C., Grillo, C., Melchiorre, N., Bertolotto, R., Ferrari,A., Vivaldi, B., Icardi, G., Gramaccioni, L., Funari, E., and Scardala, S.(2006) Ostreopsis ovata algal blooms affecting human health inGenova, Italy, 2005 and 2006. Euro Surveill. 11, 3040, available onlinehttp://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=3040.(8) Vila, M., Maso, M., Sampedro, N., Illoul, H., Arin, l., Garces, E.,Giacobbe, M. G., and Alvarez, J. (2008) The Genus Ostreopsis inRecreational Waters of the Catalan Coast and Balearic Islands (NWMediterranean Sea): Is This the Origin of Human RespiratoryDifficulties? in Proceedings of the 12th International Conference onHarmful Algae (Moestrup, O. et al., Eds.) pp 334−336, UNESCO,Copenhagen, Denmark.(9) Barroso Garcia, P., de la Puerta, P. R., Carreno, T. P., Martinez, P.M., and Enriquez, J. G. (2008) An epidemic outbreak with respiratorysymptoms in the province of Almeria [Spain] due to toxic microalgaeexposure. Gac. Sanit. 22, 578−584.(10) Tichadou, L., Glaizal, M., Armengaud, A., Grossel, H., Lemee,R., Kantin, R., Lasalle, J. L., Drouet, G., Rambaud, L., Malfait, P., andde Haro, L. (2010) Health impact of unicellular algae of the Ostreopsisgenus blooms in the Mediterranean Sea: experience of the FrenchMediterranean coast surveillance network from 2006 to 2009. Clin.Toxicol. 48 (8), 839−844.(11) Mangialajo, L., Ganzin, N., Accoroni, S., Asnaghi, V., Blanfune,A., Cabrini, M., Cattaneo-Vietti, R., Chavanon, F., Chiantore, M.,Cohu, S., Costa, E., Fornasaro, D., Grossel, H., Marco-Miralles, F.,Maso, M., Rene, A., Rossi, A. M., Sala, M. M., Thibaut, T., Totti, C.,Vila, M., and Lemee, R. (2011) Trends in Ostreopsis proliferation alongthe Northern Mediterranean coasts. Toxicon 57 (3), 408−420.(12) Durando, P., Ansaldi, F., Oreste, P., Moscatelli, P., Gasparini, L.M., and Icardi, G. (2007) Ostreopsis ovata and human health:epidemiological and clinical features of respiratory syndrome out-breaks from a two-year syndromic surveillance, 2005−06, in north-west Italy. Euro Surveill. 12 (23), 3212.(13) Tubaro, A., Durando, P., Del Favero, G., Ansaldi, F., Icardi, G.,Deeds, J. R., and Sosa, S. (2011) Case definitions for humanpoisonings postulated to palytoxins exposure. Toxicon 57, 478−495.(14) Perini, F., Casabianca, A., Battocchi, C., Accoroni, S., Totti, C.,and Penna, A. (2011) New approach using the real-time PCR methodfor estimation of the toxic marine dinoflagellate Ostreopsis cf. ovata inmarine environment. PLoS ONE 6 (3), e17699.(15) Penna, A., Vila, M., Fraga, S., Giacobbe, M. G., Andreoni, F.,Riobo, P., and Vernesi, C. (2005) Characterization of Ostreopsis andCoolia (Dinophyceae) isolates in the western Mediterranean Sea based
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521251

on morphology, toxicity and internal transcribed spacer 5.8S rDNAsequences. J. Phycol. 41, 212−245.(16) Deeds, J. R., and Schwartz, M. D. (2010) Human risk associatedwith palytoxin exposure. Toxicon 56, 150−162.(17) Moore, R. E., and Bartolini, G. (1981) Structure of palytoxin. J.Am. Chem. Soc. 103, 2491−2494.(18) Uemura, D., Ueda, K., Hirata, Y., Naoki, H., and Iwashita, T.(1981) Further studies on palytoxin. II. Structure of palytoxin.Tetrahedron Lett. 22, 2781−2784.(19) Ciminiello, P., Dell’Aversano, C., Fattorusso, E., Forino, M.,Magno, S., Tartaglione, L., Grillo, C., and Melchiorre, N. (2006) TheGenoa 2005 outbreak. Determination of putative palytoxin inMediterranean Ostreopsis ovata by a new liquid chromatographytandem mass spectrometry method. Anal. Chem. 78, 6153−6159.(20) Ciminiello, P., Dell’Aversano, C., Fattorusso, E., Forino, M.,Tartaglione, L., Grillo, C., and Melchiorre, N. (2008) PutativePalytoxin and its new analogue, ovatoxin-a, in Ostreopsis ovatacollected along the ligurian coasts during the 2006 toxic outbreak. J.Am. Soc. Mass Spectrom. 19, 111−120.(21) Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso,E., Forino, M., Grauso, L., Tartaglione, L., Guerrini, F., and Pistocchi,R. (2010) Complex palytoxin-like profile of Ostreopsis ovata.Identification of four new ovatoxins by high-resolution liquidchromatography/mass spectrometry. Rapid Commun. Mass. Spectrom.24, 2735−2744.(22) Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso,E., Forino, M., Grauso, L., Tartaglione, L., Guerrini, F., Pezzolesi, L.,Pistocchi, R., and Vanucci, S. (2012) Isolation and structureelucidation of ovatoxin-a, the major toxin produced by Ostreopsisovata. J. Am. Chem. Soc. 134 (3), 1869−1875.(23) Honsell, G., De Bortoli, M., Boscolo, S., Dell’Aversano, C.,Battocchi, C., Fontanive, G., Penna, A., Berti, F., Sosa, S., Yasumoto,T., Ciminiello, P., Poli, M., and Tubaro, A. (2011) Harmfuldinoflagellate Ostreopsis cf. ovata Fukuyo: detection of ovatoxins infield samples and cell immunolocalization using antipalytoxinantibodies. Environ. Sci. Technol. 45 (16), 7051−7059.(24) Pistocchi, R., Pezzolesi, L., Guerrini, F., Vanucci, S.,Dell’Aversano, C., and Fattorusso, E. (2011) A review on the effectsof environmental conditions on growth and toxin production ofOstreopsis ovata. Toxicon 57, 421−428.(25) Pezzolesi, L., Guerrini, F., Ciminiello, P., Dell’Aversano, C.,Dello Iacovo, E., Fattorusso, E., Forino, M., Tartaglione, L., andPistocchi, R. (2012) Influence of temperature and salinity on Ostreopsiscf. ovata growth and evaluation of toxin content through HR LC-MSand biological assays. Water Res. 46 (1), 82−92.(26) Vanucci, S., Pezzolesi, L., Pistocchi, R., Ciminiello, P.,Dell’Aversano, C., Dello Iacovo, E., Fattorusso, E., Tartaglione, L.,and Guerrini, F. (2012) Nitrogen and phosphorus limitation effects oncell growth, biovolume, and toxin production in Ostreopsis cf. ovata.Harmful Algae 15, 78−90.(27) Guillard, R. R. L. (1975) Culture of Phytoplankton for FeedingMarine Invertebrates, in Culture of Marine Invertebrate Animals (Smith,W. L., and Chanley, M. H., Eds) pp 29−60, Plenum Press, New York.(28) Throndsen, J. (1995) Estimating Cell Numbers, in Manual onHarmful Marine Microalgae, IOC Manual and Guides No. 33(Hallegraeff, G. M., Anderson, D. M., Cembella, A. D., andEnevoldsen, H. O., Eds.) pp 63 and 80, UNESCO, Paris, France.(29) Utermohl, H. (1958) Zur Vervollkommung der quantitativenPhytoplankton-Methodik. Mitt. Int. Verein. Limnol. 9, 1−38.(30) Penna, A., Bertozzini, E., Battocchi, C., Giacobbe, M. G.,Galluzzi, L., Garces, E., Vila, M., Luglie, A., and Magnani, M. (2007)Monitoring of HAB species in the Mediterranean Sea throughmolecular techniques. J. Plank. Res. 29, 19−38.(31) Battocchi, C., Totti, C., Vila, M., Maso, M., Capellacci, S.,Accoroni, S., Rene, A., Scardi, M., and Penna, A. (2010) Monitoringtoxic microalgae Ostreopsis (dinoflagellate) species in coastal waters ofthe Mediterranean Sea using molecular PCR-based assay combinedwith light microscopy. Mar. Pollut. Bull. 60 (7), 1074−1084.
(32) Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso,E., Forino, M., Grauso, L., and Tartaglione, L. (2012) High resolutionLC-MSn fragmentation pattern of palytoxin as template to gain newinsights into ovatoxin-a structure. The key role of calcium in MSbehavior of palytoxins. J. Am. Soc. Mass. Spectrom. 23, 952−963.(33) Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso,E., Forino, M., and Tartaglione, L. (2011) LC-MS of palytoxin and itsanalogues: State of the art and future perspectives. Toxicon 57, 376−389.(34) Ukena, T., Satake, M., Usami, M., Oshima, Y., Naoki, H., Fujita,T., Kan, Y., and Yasumoto, T. (2001) Structure elucidation ofostreocin D, a palytoxin analog isolated from the dinoflagellateOstreopsis siamensis. Biosci. Biotechnol. Biochem. 65, 2585−2588.(35) Uemura, D., Hirata, Y., Iwashita, T., and Naoki, H. (1985)Studies on palytoxins. Tetrahedron 41, 1007−1017.(36) Ciminiello, P., Dell’Aversano, C., Dello Iacovo, E., Fattorusso,E., Forino, M., Grauso, L., Tartaglione, L., Florio, C., Lorenzon, P., DeBortoli, M., Tubaro, A., Poli, M., and Bignami., G. (2009)Stereostructure and biological activity of 42-hydroxy-palytoxin: anew palytoxin analogue from Hawaiian Palythoa subspecies. Chem. Res.Toxicol. 22, 1851−1859.(37) Accoroni, S., Romagnoli, T., Colombo, F., Pennesi, C., DiCamillo, C. G., Marini, M., Battocchi, C., Ciminiello, P., Dell’Aversano,C., Dello Iacovo, E., Fattorusso, E., Tartaglione, L., Penna, A., andTotti, C. (2011) Ostreopsis cf. ovata bloom in the northern Adriatic Seaduring summer 2009: Ecology, molecular characterization and toxinprofile. Mar. Pollut. Bull. 62 (11), 2512−2519.
Chemical Research in Toxicology Article
dx.doi.org/10.1021/tx300085e | Chem. Res. Toxicol. 2012, 25, 1243−12521252