TERMINAZIONE*DELLA* TRASCRIZIONE*IN* … - Montagner.pdf · Terminazione della trascrizione in...
45
TERMINAZIONE DELLA TRASCRIZIONE IN PROCARIOTI ED EUCARIOTI
Transcript of TERMINAZIONE*DELLA* TRASCRIZIONE*IN* … - Montagner.pdf · Terminazione della trascrizione in...

TERMINAZIONE DELLA TRASCRIZIONE IN
PROCARIOTI ED EUCARIOTI

Concetti generali sulla trascrizione batterica
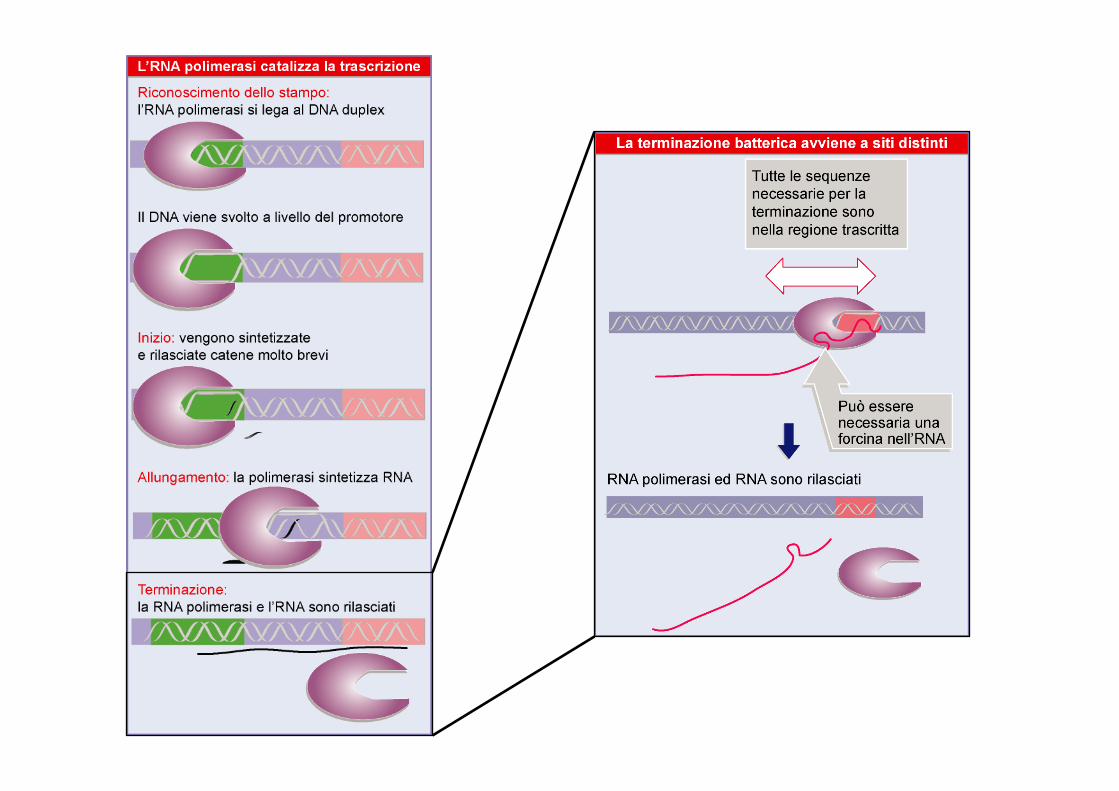
- Allo stesso modo dell’inizio della trascrizione, la terminazione non e’ un fenomeno passivo, ma e’ un modo per controllare la trascrizione
- L’RNA polimerasi sintetizza l’RNA finche’ non trova delle sequenze chiamate TERMINATORI, a quel punto smette di aggiungere nucleotidi, si rompono I legami idrogeni RNA-DNA e l’mRNA si stacca dal template
- Come studiare I siti di terminazione:
- In vivo e’ difficile perche’ non si e’ sicuri se l’mRNA sia stato tagliato o meno
- In vitro e’ difficile perche’ l’attivita’ dell’RNA polimerasi dipende molto dalla forza ionica e dalle altre condizioni chimico-fisiche
Quindi informazioni attendibili sui siti di terminazione arrivano da siti in cui la sequenza al 3’ dell’mRNA e’ uguale in vitro ed in vivo

Concetti generali sulla trascrizione batterica
- Tutte le sequenze richieste per la terminazione risiedono PRIMA dell’ultima base trascritta (quindi al 5’ dell’ultima base), quindi non ci sono sequenze conservate successive all’mRNA che indichino la presenza di terminatori
La responsabilita’ della terminazione risiede nell’mRNA appena trascritto
- La maggior parte delle volte le informazioni per la terminazione risiedono nella STRUTTURA SECONDARIA dell’mRNA, quindi non basta analizzare la sequenza di DNA per capirne la presenza
- Ci sono due classi di terminatori a seconda che l’RNA Polimerasi richieda o no altri cofattori per terminare la trascrizione:
1. Terminatori instrinseci: non hanno bisogno di cofattori, si basano unicamente sulla sequenza dell’mRNA
2. Terminatori Rho-dipendenti: richiedono un fattore proteico chiamato rho
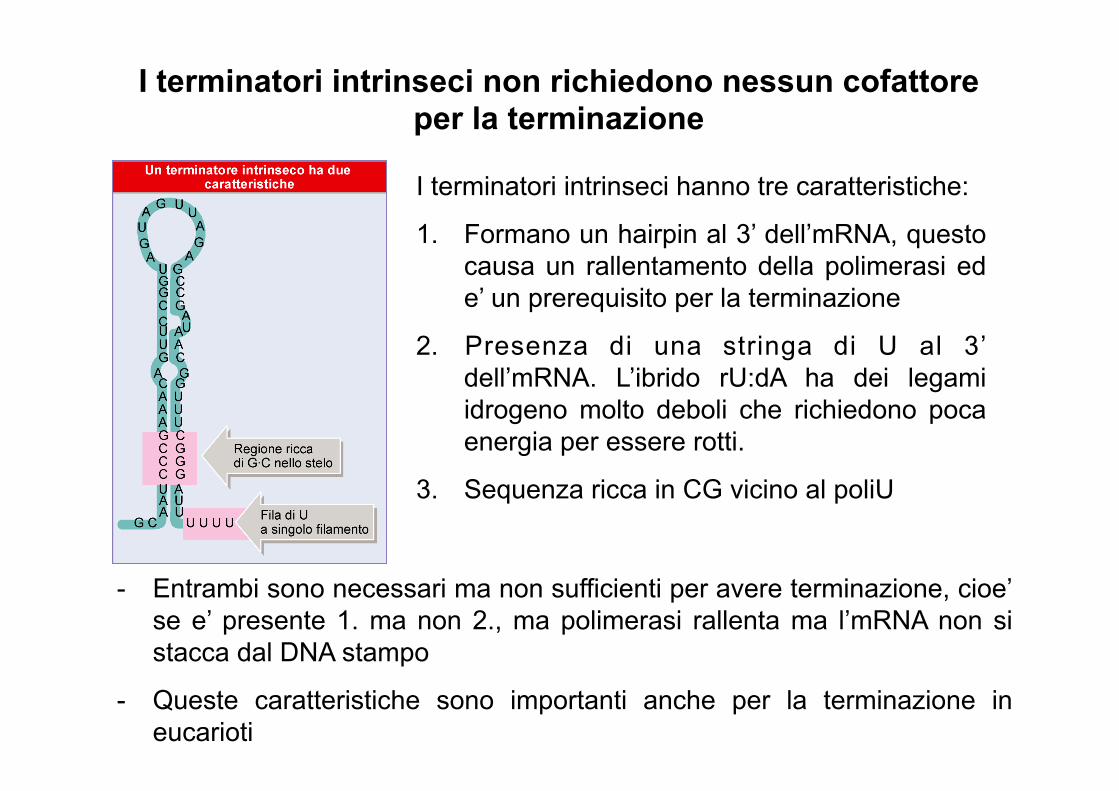
I terminatori intrinseci non richiedono nessun cofattore per la terminazione
I terminatori intrinseci hanno tre caratteristiche:
1. Formano un hairpin al 3’ dell’mRNA, questo causa un rallentamento della polimerasi ed e’ un prerequisito per la terminazione
2. Presenza di una stringa di U al 3’ dell’mRNA. L’ibrido rU:dA ha dei legami idrogeno molto deboli che richiedono poca energia per essere rotti.
3. Sequenza ricca in CG vicino al poliU
- Entrambi sono necessari ma non sufficienti per avere terminazione, cioe’ se e’ presente 1. ma non 2., ma polimerasi rallenta ma l’mRNA non si stacca dal DNA stampo
- Queste caratteristiche sono importanti anche per la terminazione in eucarioti
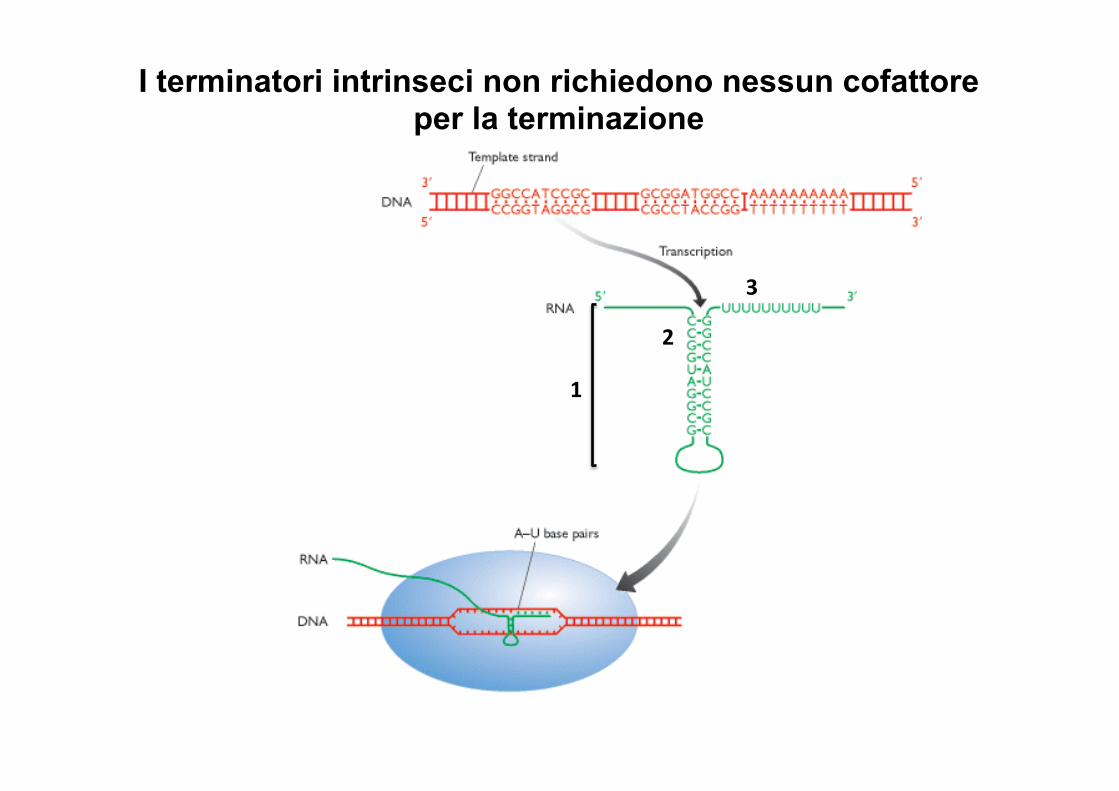
I terminatori intrinseci non richiedono nessun cofattore per la terminazione
1
2
3
Terminatori Rho-dipendenti
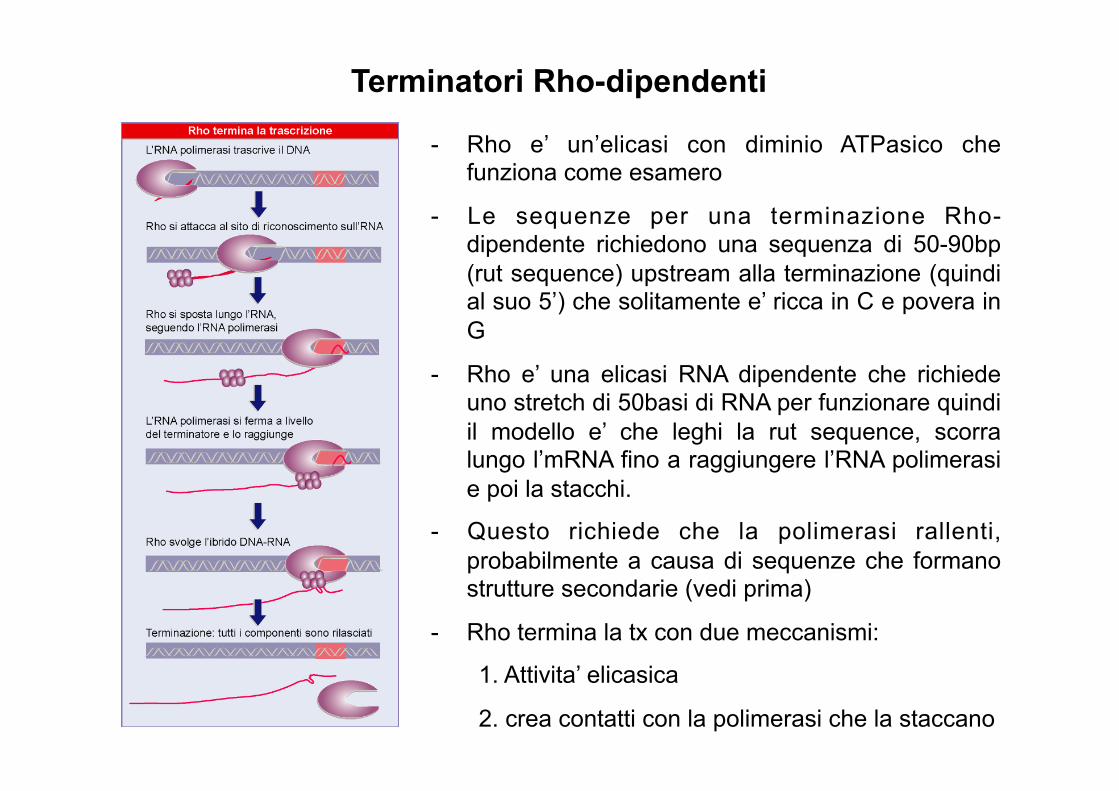
- Rho e’ un’elicasi con diminio ATPasico che funziona come esamero
- Le sequenze per una terminazione Rho-dipendente richiedono una sequenza di 50-90bp (rut sequence) upstream alla terminazione (quindi al suo 5’) che solitamente e’ ricca in C e povera in G
- Rho e’ una elicasi RNA dipendente che richiede uno stretch di 50basi di RNA per funzionare quindi il modello e’ che leghi la rut sequence, scorra lungo l’mRNA fino a raggiungere l’RNA polimerasi e poi la stacchi.
- Questo richiede che la polimerasi rallenti, probabilmente a causa di sequenze che formano strutture secondarie (vedi prima)
- Rho termina la tx con due meccanismi:
1. Attivita’ elicasica
2. crea contatti con la polimerasi che la staccano

Terminatori Rho-dipendenti
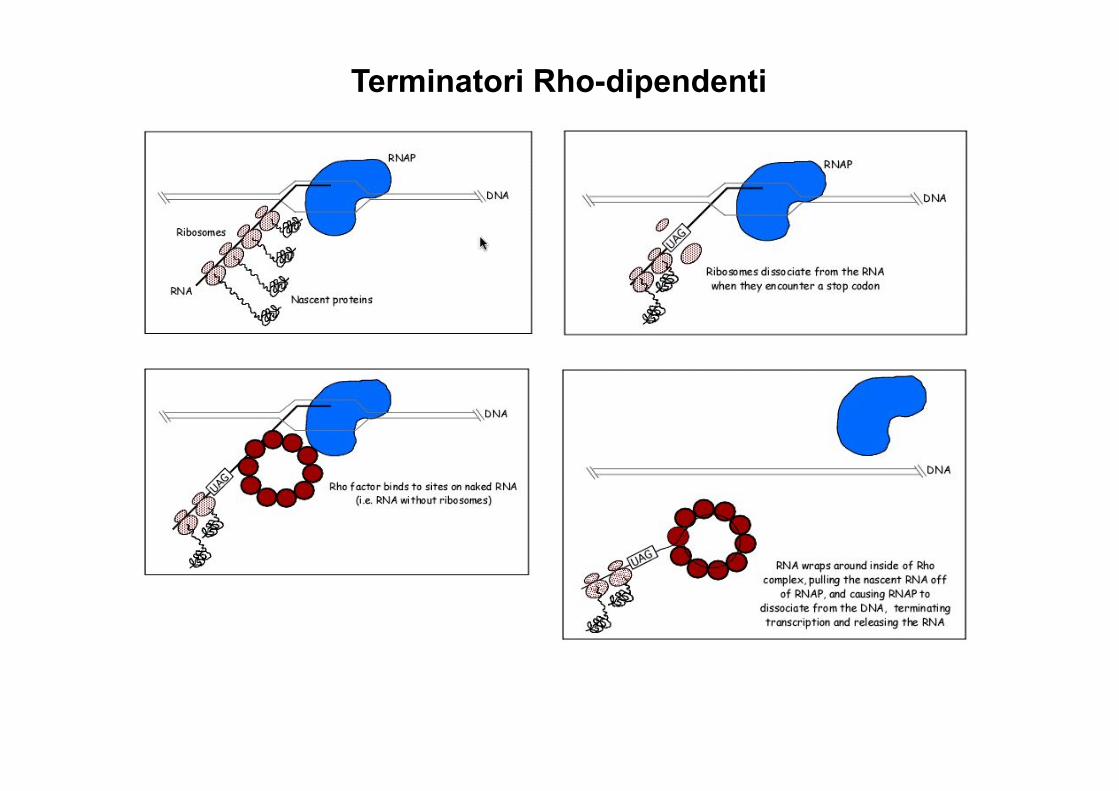
Nei batteri trascrizione e traduzione sono spesso accoppiate, cosa succede alla terminazione Rho-dipendente se l’mRNA e’ gia’ occupato dai ribosomi?
Rho non puo’ funzionare
Spesso molti geni coinvolti nella stessa pathway sono trascritti come unico mRNA nei batteri. Se ci fosse una mutazione non senso in uno di questi geni, avrebbe senso trascrivere gli altri?
Se non ho traduzione di un mRNA trascritto, ha senso continuare a trascriverlo?
Il fattore Rho permette di terminare la tx di sequenze non tradotte o errate grazie alla presenza di sequence terminatrici all’interno degli mRNA, la polimerasi rallenta e, in assenza di ribosomi, Rho raggiunge la polimerasi. Questo fenomeno e’ detto polarita’.
Perche’ ci sono terminatori criptici negli mRNA? O si sono evoluti appositamente o si formando grazie al ripiegamento in 2D di sequenze.
tRNA e rRNA non sono soggetti a polarita’ perche formano strutture secondarie che impediscono il legame di Rho
Terminatori Rho-dipendenti

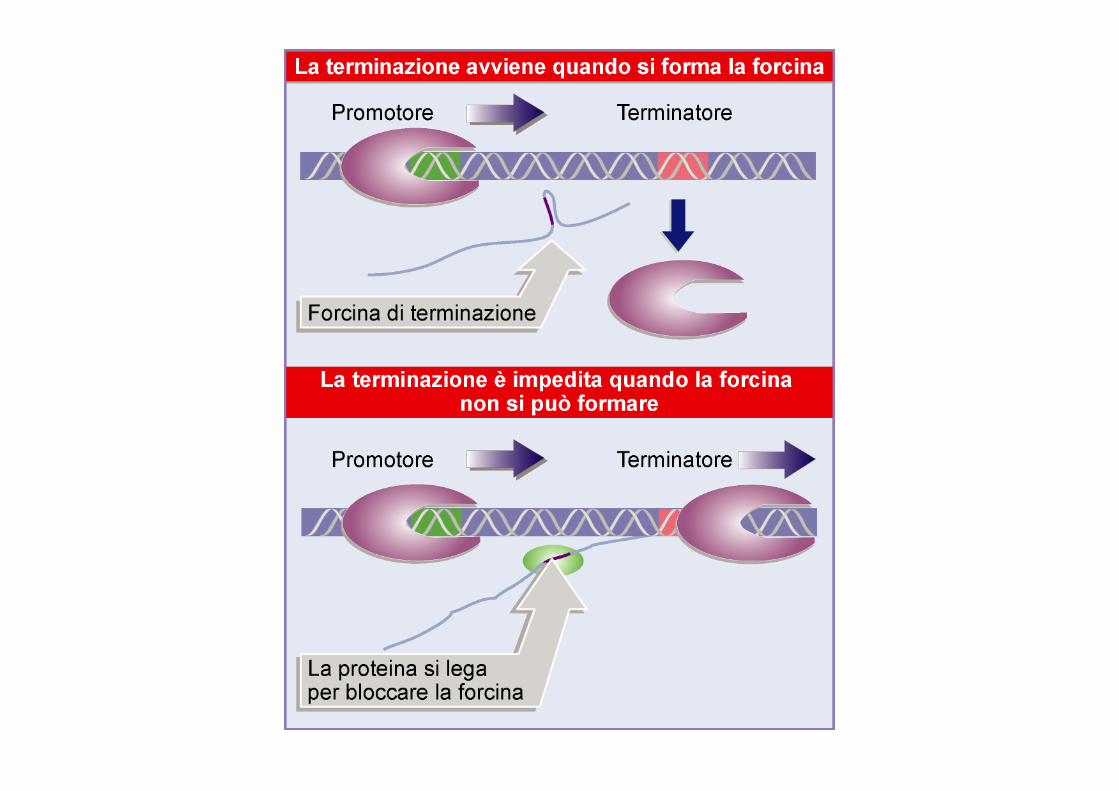
La terminazione batterica puo’ essere modulata
Antiterminatori = Specifiche proteine che consentono alla RNA polimerasi di continuare la trascrizione attraverso il sito di terminazione Attenuatori = terminatori intrinseci localizzati all’inizio di un’unita’ trascrizionale

Antiterminazione: l’esempio del fago λ
Una caratteristica comune della strategia dei fagi e’ che pochi dei geni fagici sono trascritti dall’RNA polimerasi del batterio infettato. Tra questi geni ci sono delle proteine che promuovono la trascrizione dei rimanenti geni fagici con diverse strategie: 1. Sintesi di nuovi fattori sigma che riconoscano I promotori
del fago 2. Sintesi di nuova RNA polimerasi fagica in grado di
riconoscere I promotori fagici senza altri cofattori 3. Sintesi di antiterminatori che consentano di continuare a
trascrivere geni successivi a quelli trascritti per primi. In questo modo tutti I geni saranno controllati dai promotori iniziali
Il caso del fago lambda: Cascata di geni
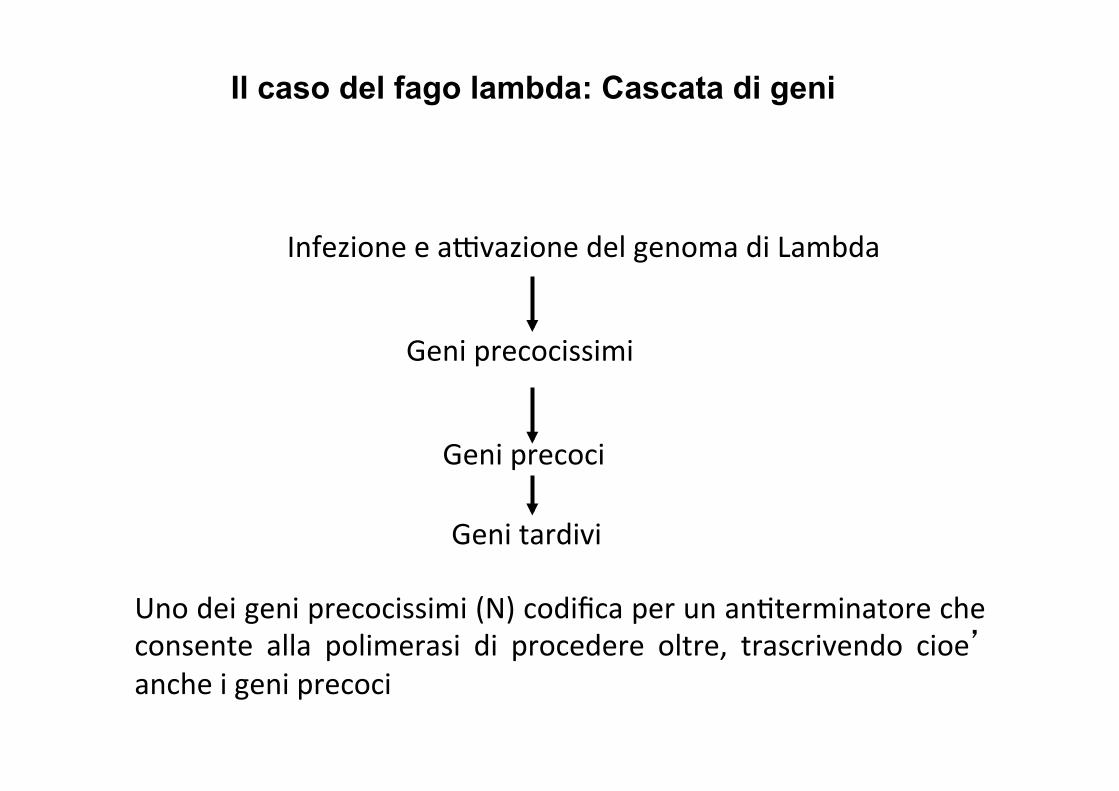
Geni precocissimi
Geni precoci
Geni tardivi
Infezione e aAvazione del genoma di Lambda
Uno dei geni precocissimi (N) codifica per un anIterminatore che consente alla polimerasi di procedere oltre, trascrivendo cioe’ anche i geni precoci
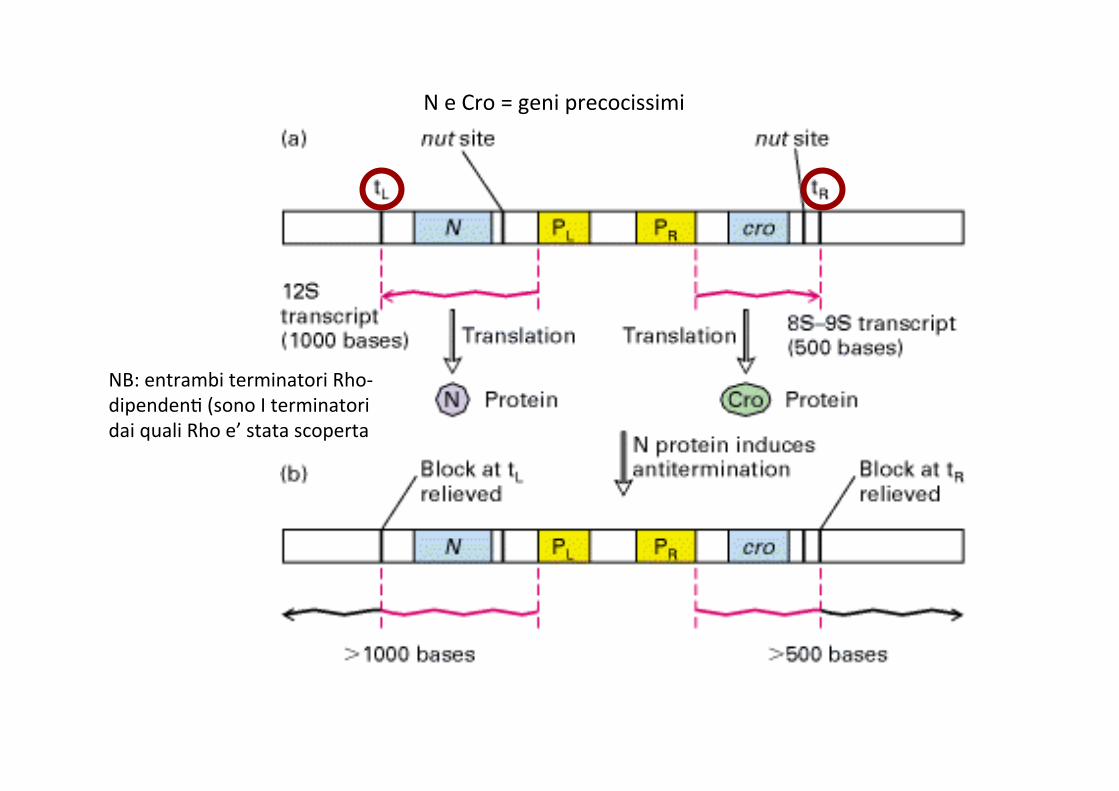
N e Cro = geni precocissimi
NB: entrambi terminatori Rho-‐dipendenI (sono I terminatori dai quali Rho e’ stata scoperta
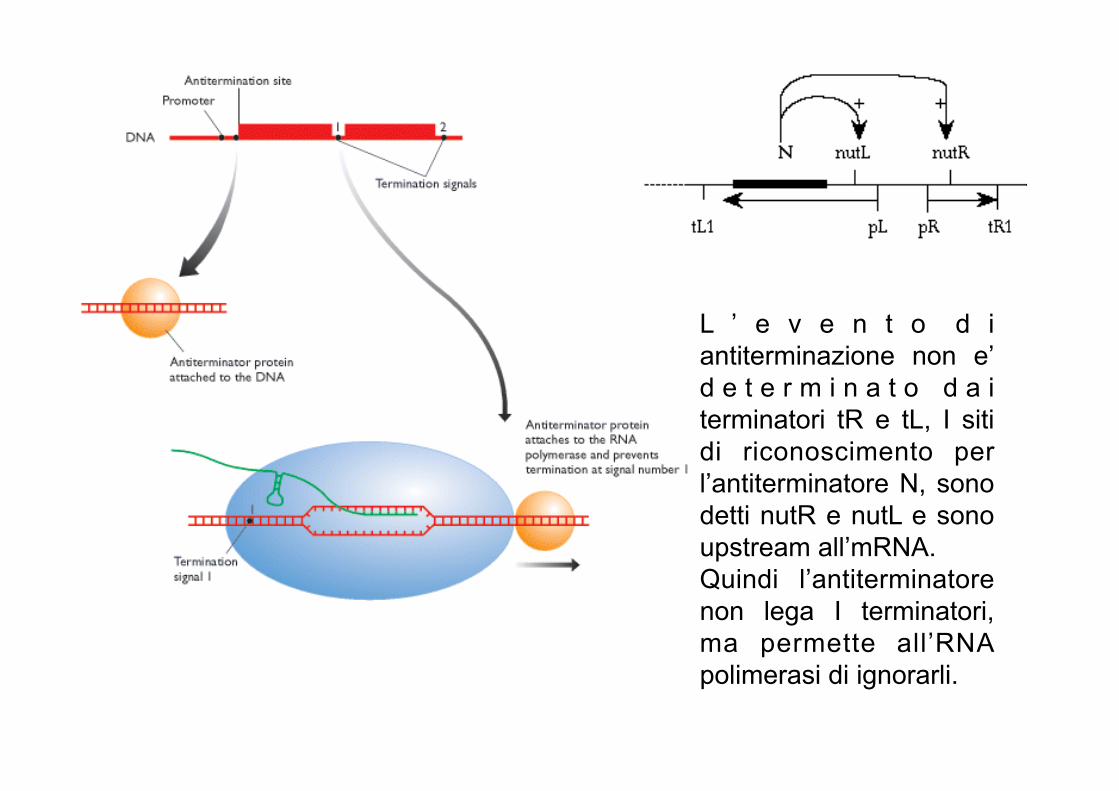
L ’ e v e n t o d i antiterminazione non e’ d e t e r m i n a t o d a i terminatori tR e tL, I siti di riconoscimento per l’antiterminatore N, sono detti nutR e nutL e sono upstream all’mRNA. Quindi l’antiterminatore non lega I terminatori, ma permette all’RNA polimerasi di ignorarli.

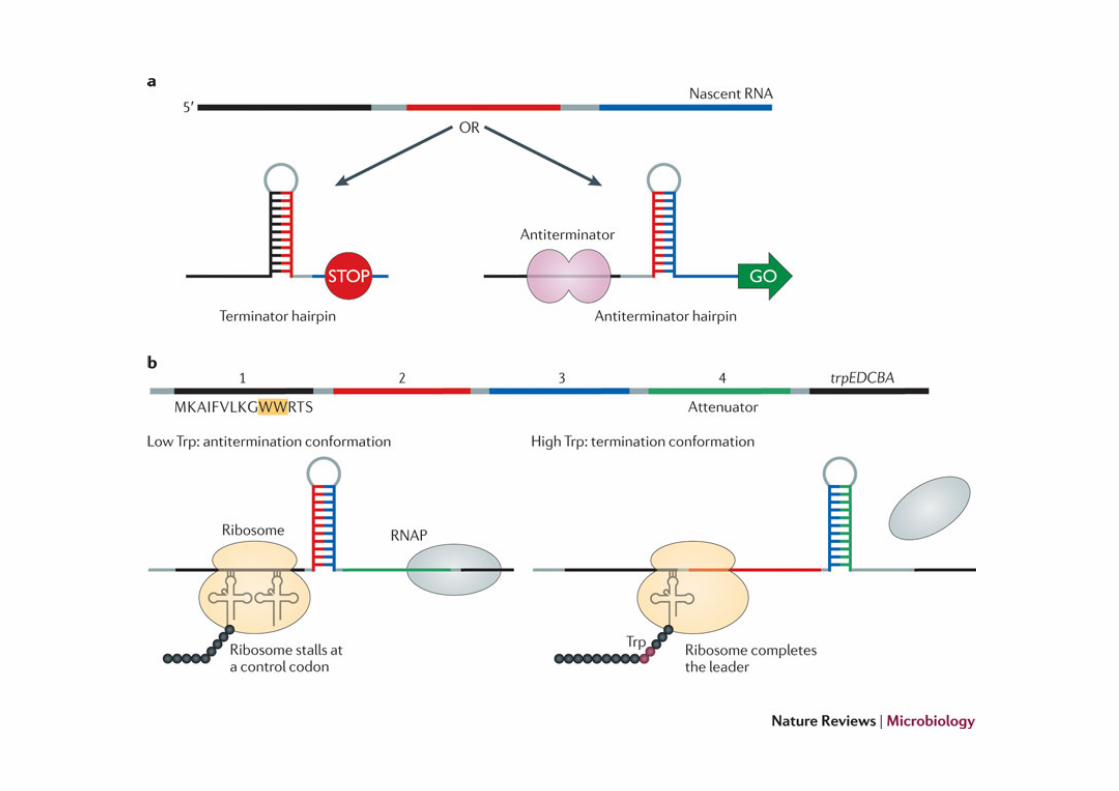
Attenuazione: l’esempio dell’operone triptofano
L’operone triptofano (trp) codifica per cinque geni strutturali disposti l’uno di seguito all’altro, codificanti per tre enzimi che convertono l’acido corismico in triptofano (un amminoacido). L’operone e’ sotto il controllo di due meccanismi che evitano la produzione degli enzimi in caso di abbondanza di triptofano: 1. Repressione dell’espressione attraverso un repressore
che lega la sequenza operatore 2. Attenuazione: indipendente dalla repressione, regola
l’espressione dopo che la polimerasi ha lasciato il promotore.
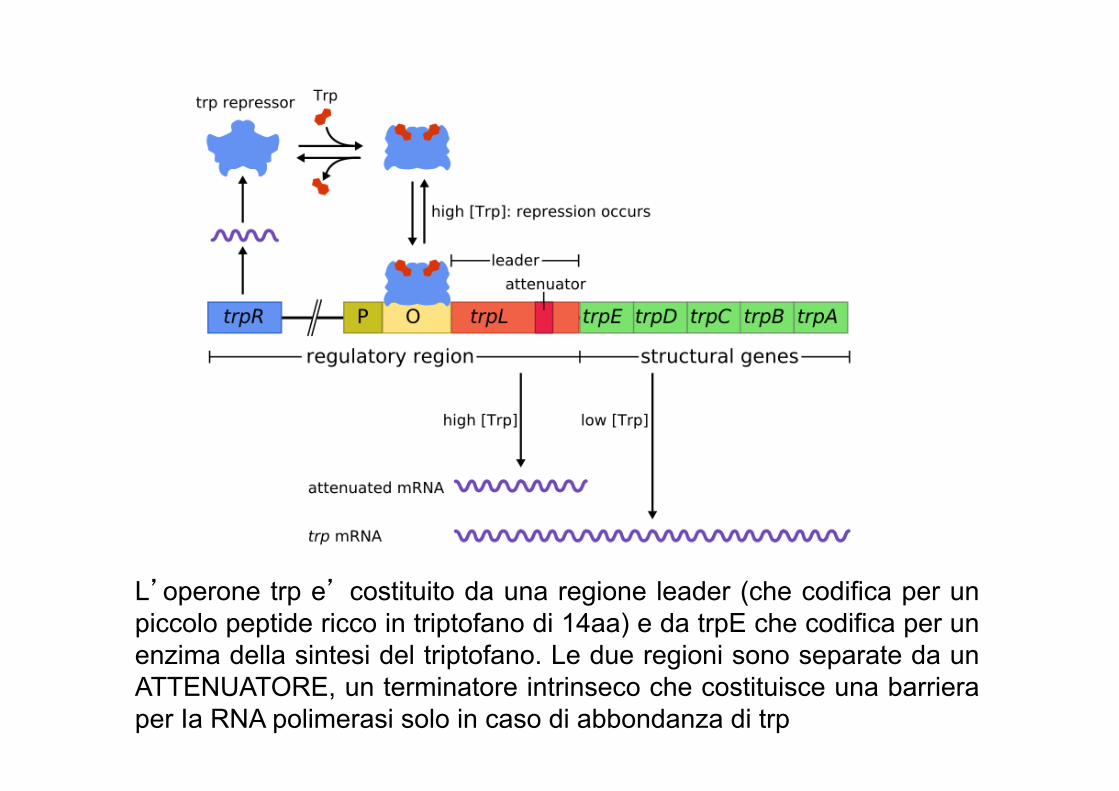
L’operone trp e’ costituito da una regione leader (che codifica per un piccolo peptide ricco in triptofano di 14aa) e da trpE che codifica per un enzima della sintesi del triptofano. Le due regioni sono separate da un ATTENUATORE, un terminatore intrinseco che costituisce una barriera per Ia RNA polimerasi solo in caso di abbondanza di trp
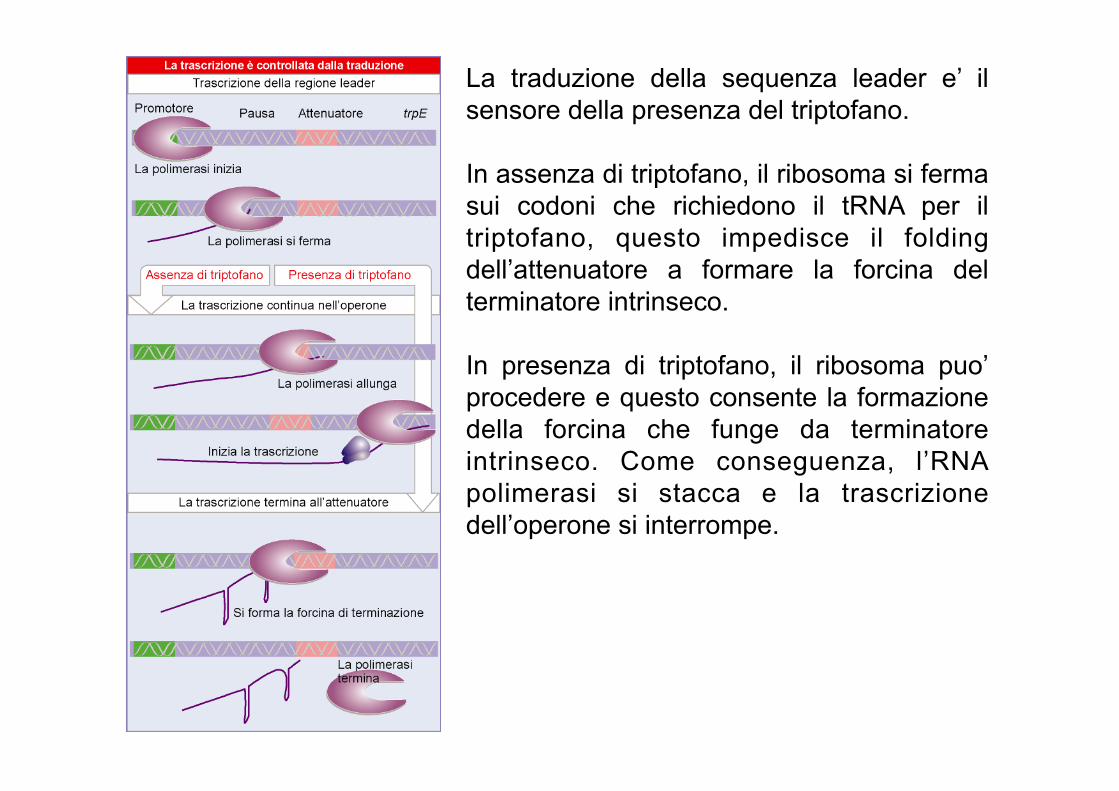
La traduzione della sequenza leader e’ il sensore della presenza del triptofano. In assenza di triptofano, il ribosoma si ferma sui codoni che richiedono il tRNA per il triptofano, questo impedisce il folding dell’attenuatore a formare la forcina del terminatore intrinseco. In presenza di triptofano, il ribosoma puo’ procedere e questo consente la formazione della forcina che funge da terminatore intrinseco. Come conseguenza, l’RNA polimerasi si stacca e la trascrizione dell’operone si interrompe.
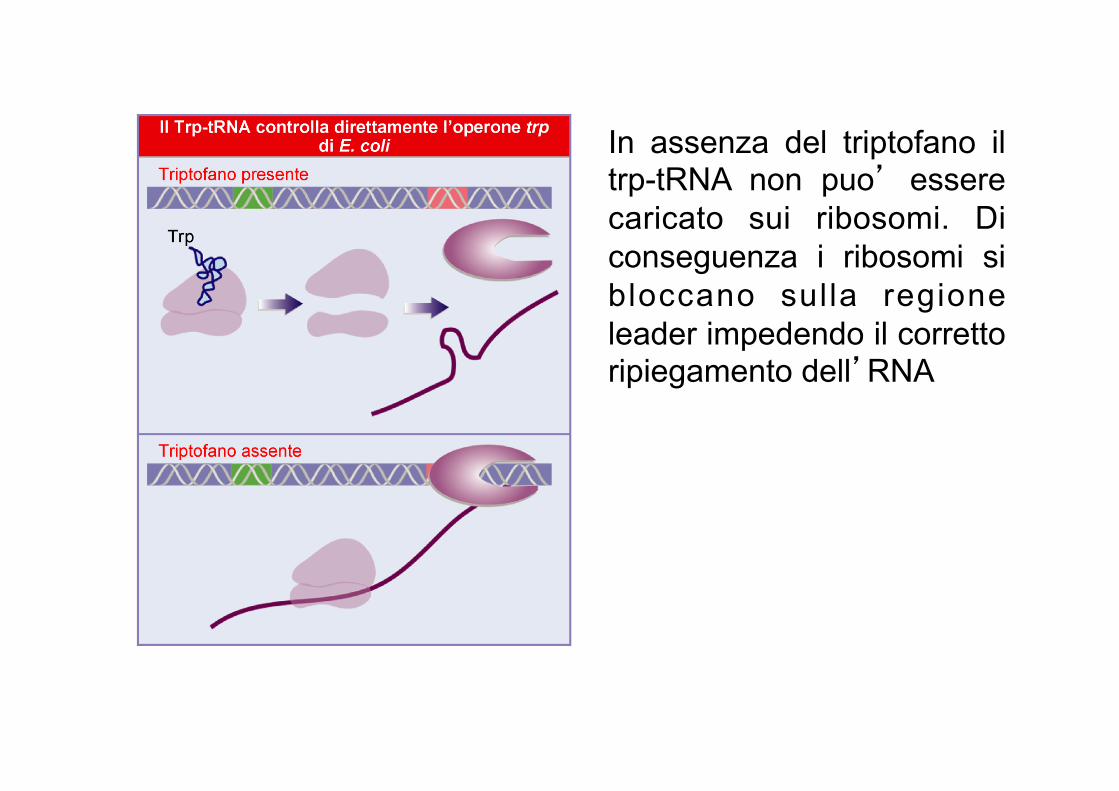
In assenza del triptofano il trp-tRNA non puo’ essere caricato sui ribosomi. Di conseguenza i ribosomi si bloccano sulla regione leader impedendo il corretto ripiegamento dell’RNA
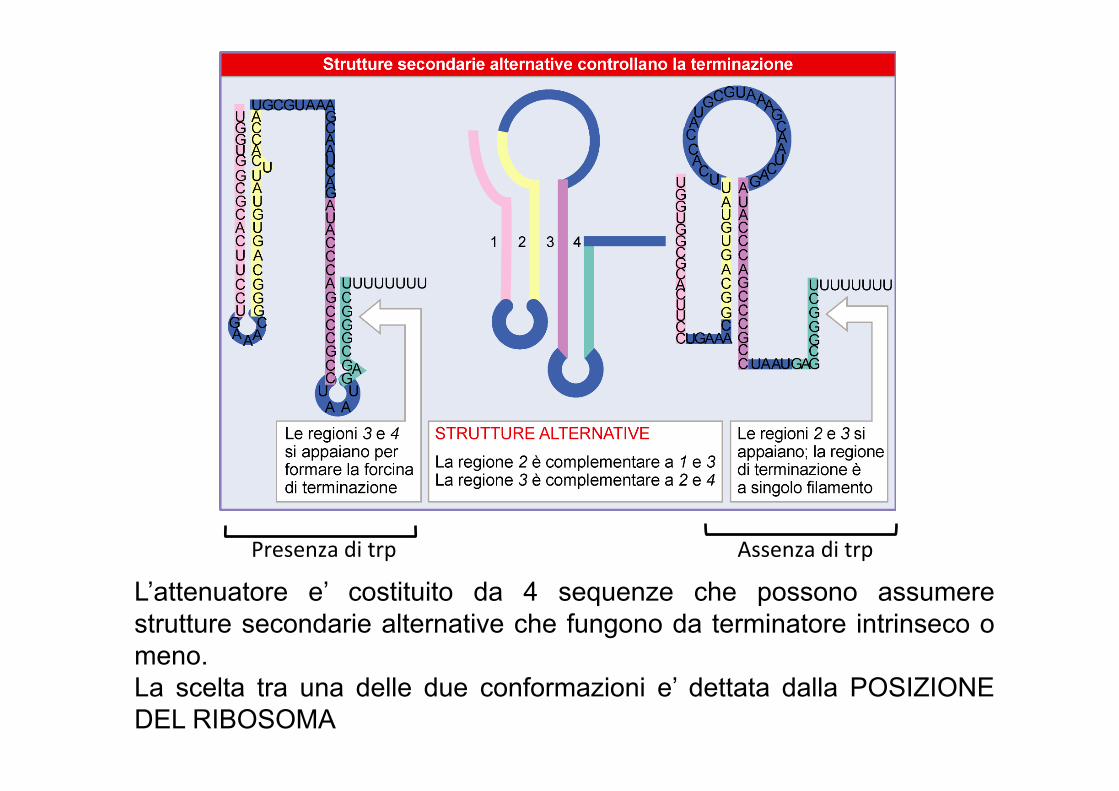
Presenza di trp Assenza di trp
L’attenuatore e’ costituito da 4 sequenze che possono assumere strutture secondarie alternative che fungono da terminatore intrinseco o meno. La scelta tra una delle due conformazioni e’ dettata dalla POSIZIONE DEL RIBOSOMA
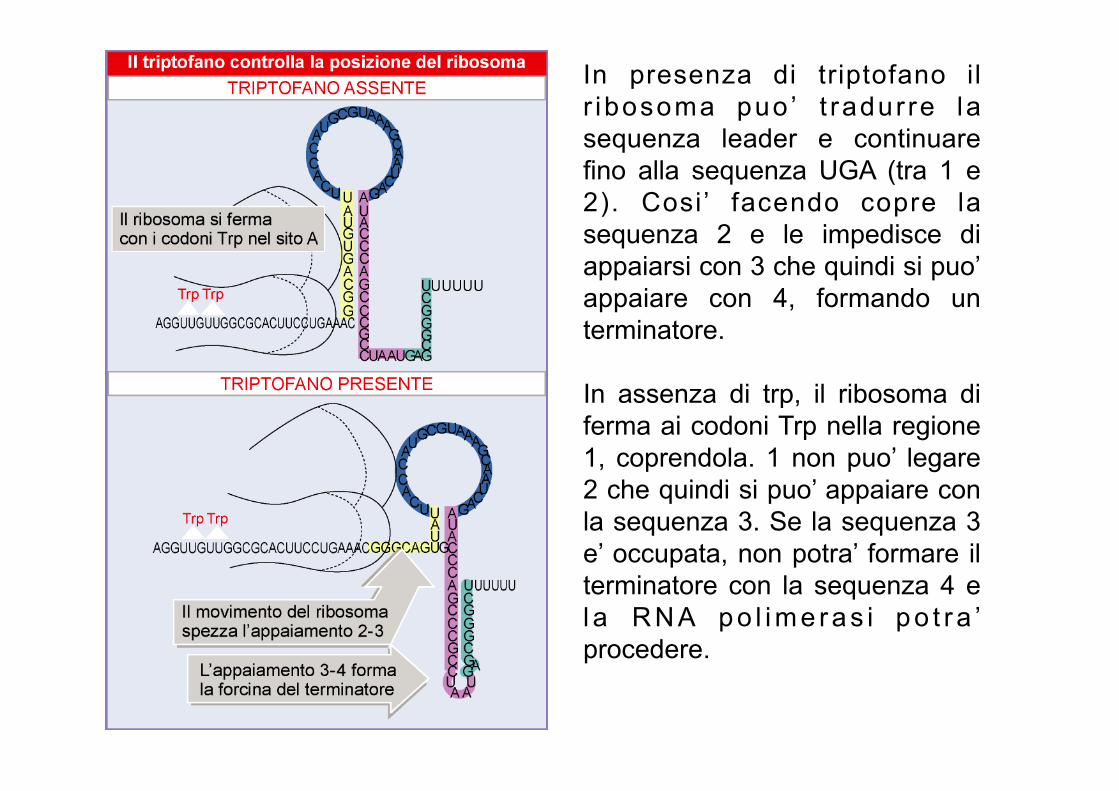
In presenza di triptofano il r ibosoma puo ’ t radur re la sequenza leader e continuare fino alla sequenza UGA (tra 1 e 2). Cosi’ facendo copre la sequenza 2 e le impedisce di appaiarsi con 3 che quindi si puo’ appaiare con 4, formando un terminatore. In assenza di trp, il ribosoma di ferma ai codoni Trp nella regione 1, coprendola. 1 non puo’ legare 2 che quindi si puo’ appaiare con la sequenza 3. Se la sequenza 3 e’ occupata, non potra’ formare il terminatore con la sequenza 4 e l a R N A p o l i m e r a s i p o t r a ’ procedere.
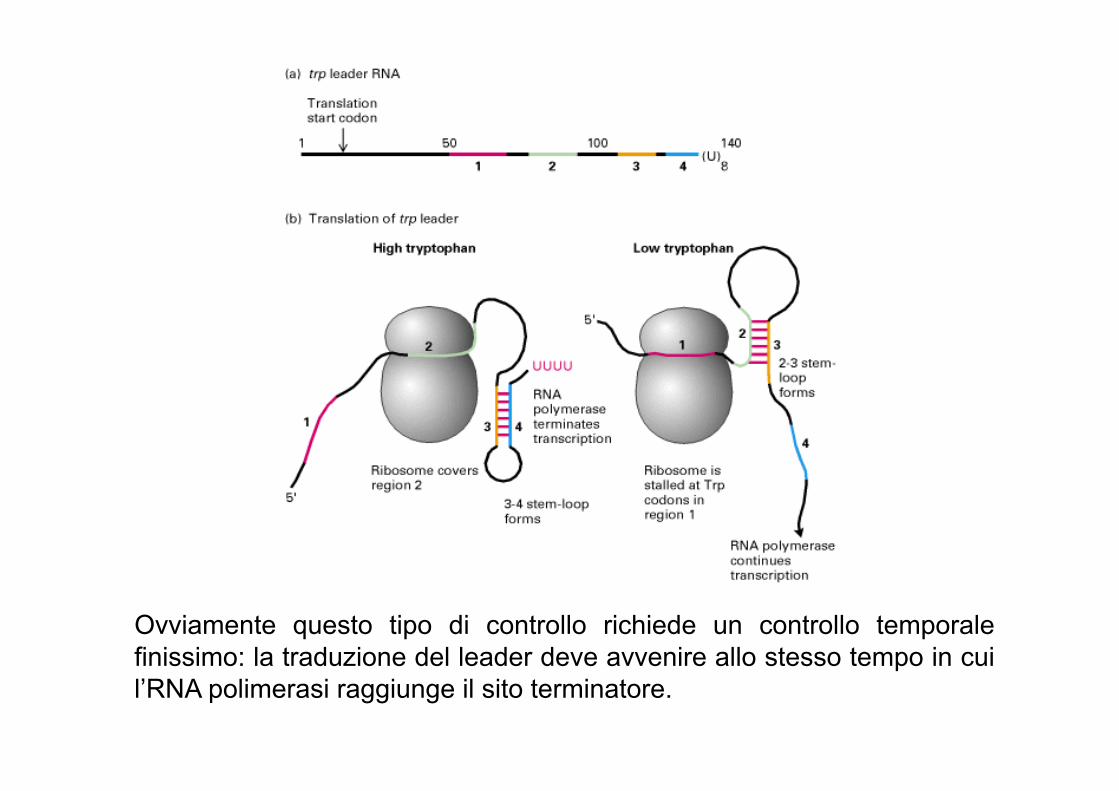
Ovviamente questo tipo di controllo richiede un controllo temporale finissimo: la traduzione del leader deve avvenire allo stesso tempo in cui l’RNA polimerasi raggiunge il sito terminatore.

https://www.youtube.com/watch?v=42RqqAYs8Fk
https://www.youtube.com/watch?v=YAr18UR2dos
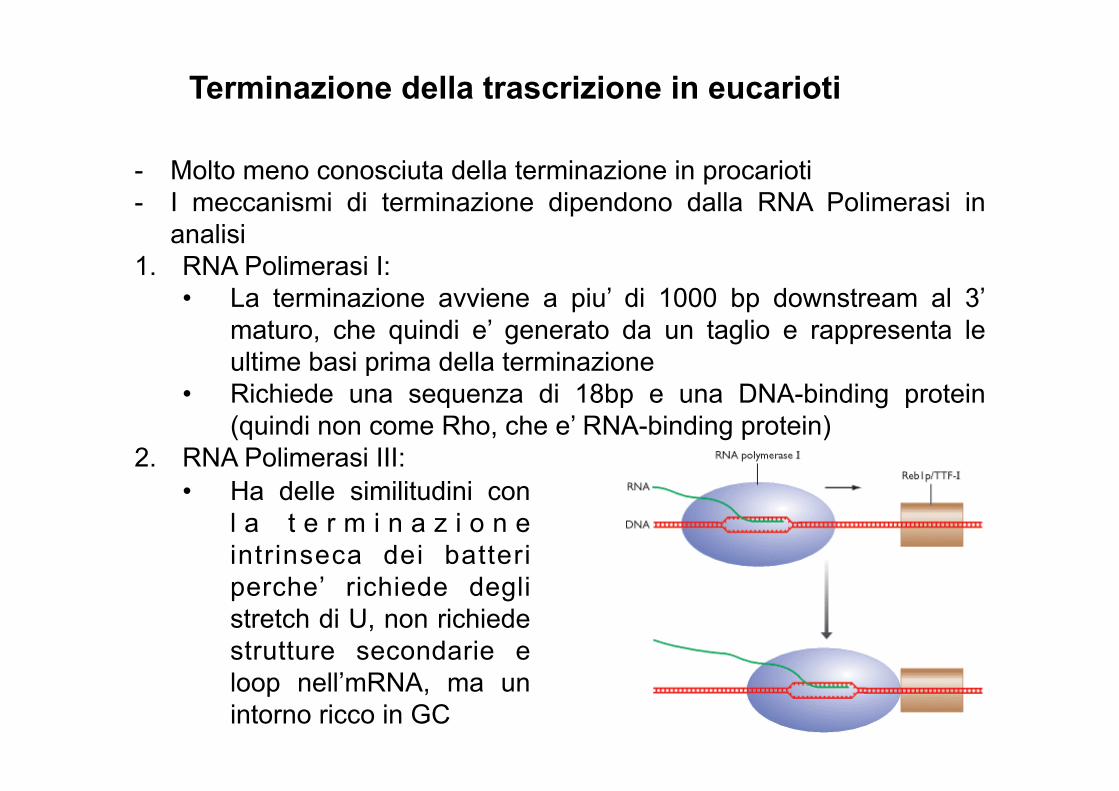
Terminazione della trascrizione in eucarioti
- Molto meno conosciuta della terminazione in procarioti - I meccanismi di terminazione dipendono dalla RNA Polimerasi in
analisi 1. RNA Polimerasi I:
• La terminazione avviene a piu’ di 1000 bp downstream al 3’ maturo, che quindi e’ generato da un taglio e rappresenta le ultime basi prima della terminazione
• Richiede una sequenza di 18bp e una DNA-binding protein (quindi non come Rho, che e’ RNA-binding protein)
2. RNA Polimerasi III:
• Ha delle similitudini con
l a t e r m i n a z i o n e intrinseca dei batteri perche’ richiede degli stretch di U, non richiede strutture secondarie e loop nell’mRNA, ma un intorno ricco in GC

Terminazione della trascrizione in eucarioti
3. RNA Polimerasi II: non ha terminatori, la terminazione e’ strettamente correlata ai processi di maturazione dell’mRNA e puo’ avvenire 0.5-2 Kb a valle del sito di poliadenilazione
La maggior parte degli mRNA eucariotici (tutti tranne quelli degli istoni), sono processati (modificati) nel nucleo. Queste modificazioni determinano: 1. La stabilita’ dell’mRNA 2. La sua capacita’ di essere legato e quindi tradotto dai ribosomi 3. Il distacco dell’RNA Polimerasi II dal DNA e la terminazione della
trascrizione
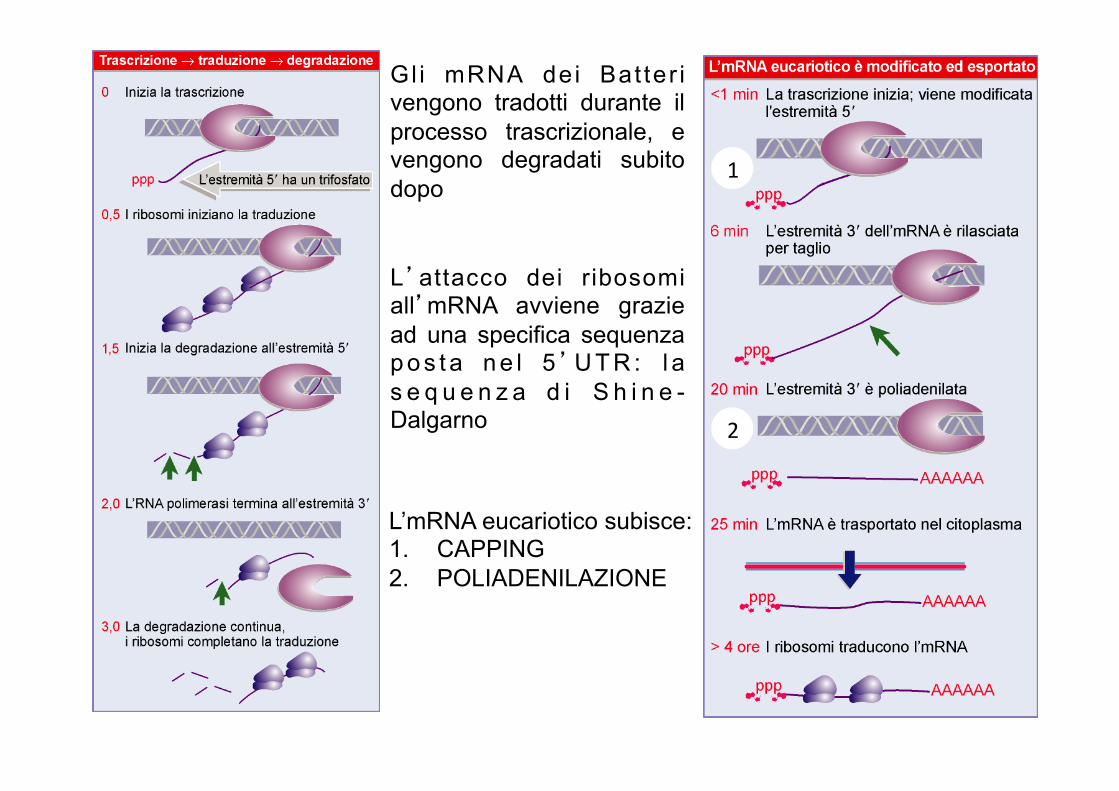
Gl i mRNA dei Bat ter i vengono tradotti durante il processo trascrizionale, e vengono degradati subito dopo
L’attacco dei ribosomi all’mRNA avviene grazie ad una specifica sequenza p o s t a n e l 5’U T R : l a s e q u e n z a d i S h i n e -Dalgarno
L’mRNA eucariotico subisce: 1. CAPPING 2. POLIADENILAZIONE
1
2
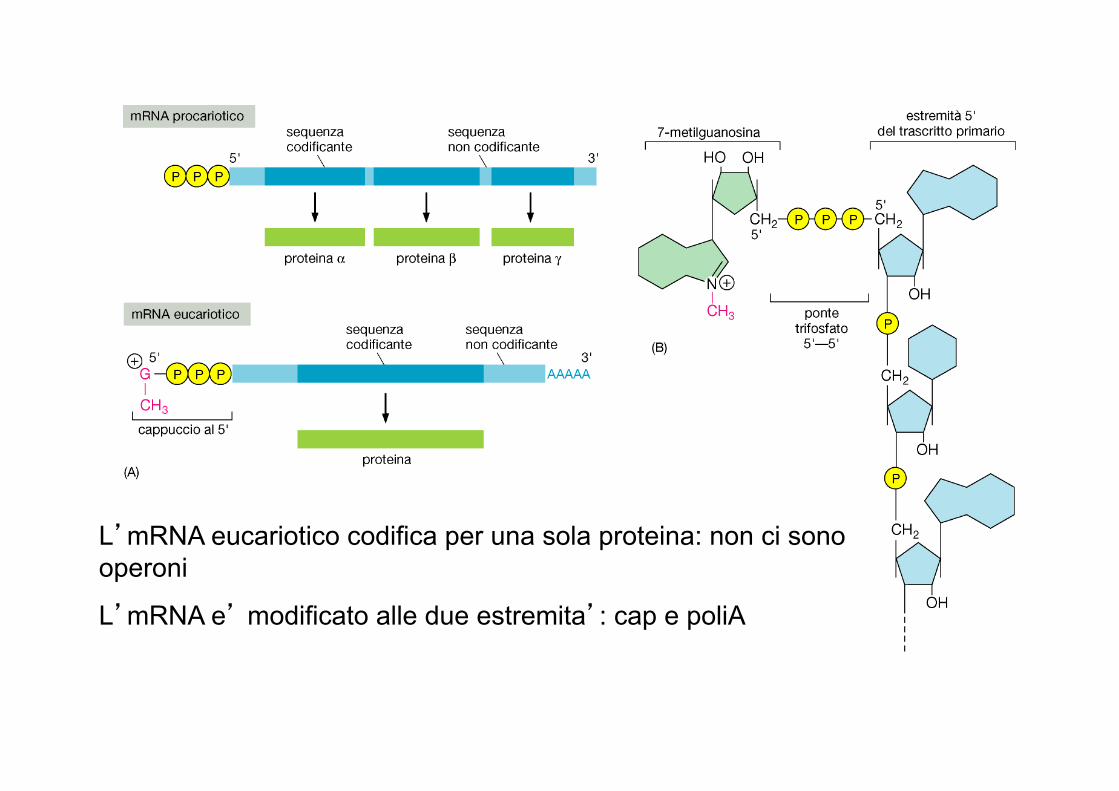
L’mRNA eucariotico codifica per una sola proteina: non ci sono operoni
L’mRNA e’ modificato alle due estremita’: cap e poliA
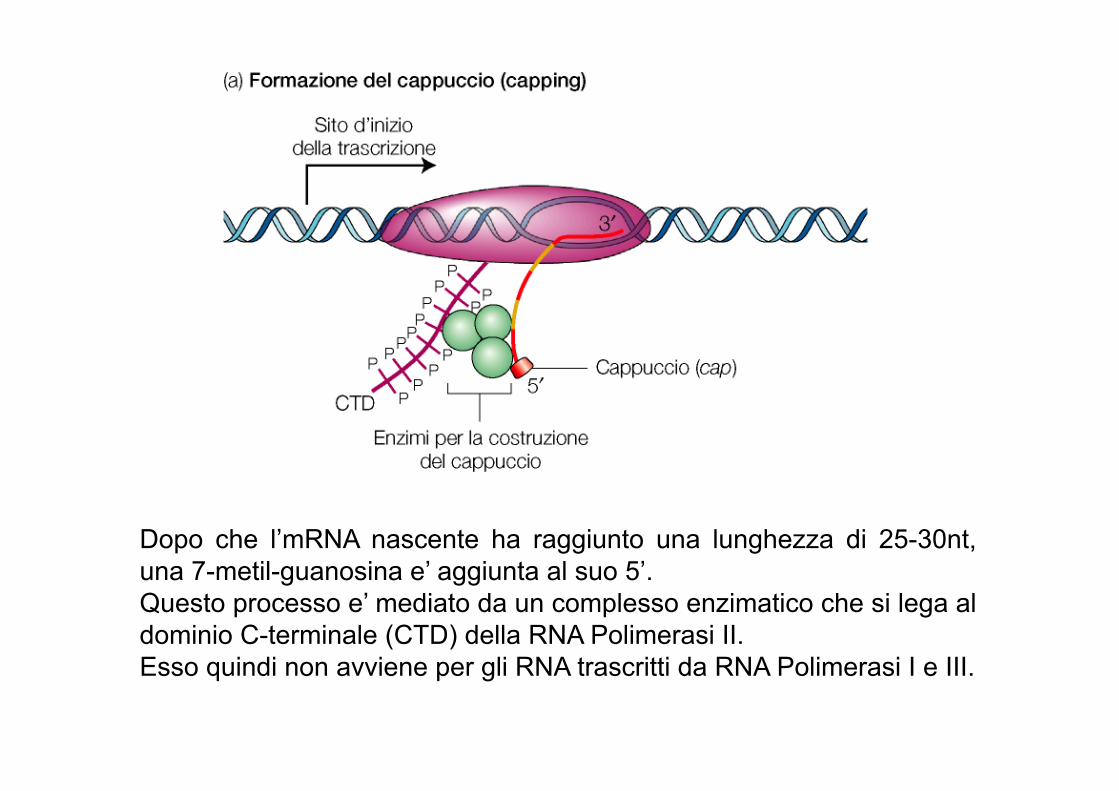
Dopo che l’mRNA nascente ha raggiunto una lunghezza di 25-30nt, una 7-metil-guanosina e’ aggiunta al suo 5’. Questo processo e’ mediato da un complesso enzimatico che si lega al dominio C-terminale (CTD) della RNA Polimerasi II. Esso quindi non avviene per gli RNA trascritti da RNA Polimerasi I e III.
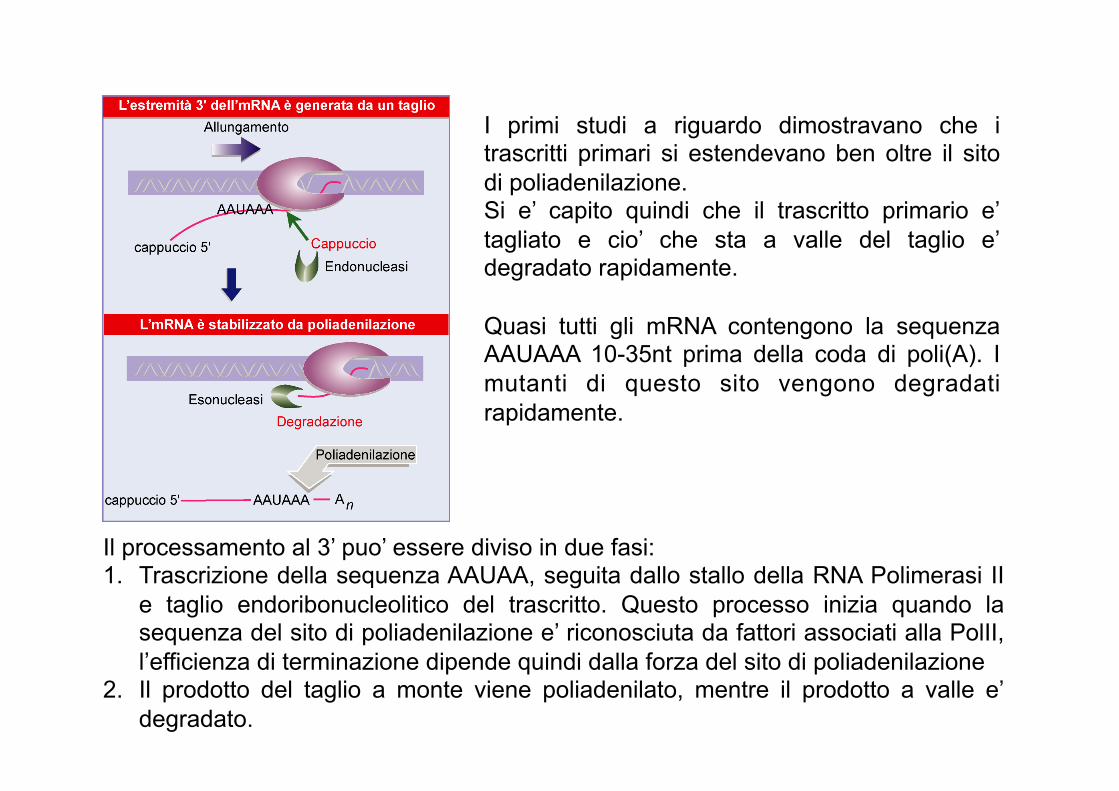
I primi studi a riguardo dimostravano che i trascritti primari si estendevano ben oltre il sito di poliadenilazione. Si e’ capito quindi che il trascritto primario e’ tagliato e cio’ che sta a valle del taglio e’ degradato rapidamente. Quasi tutti gli mRNA contengono la sequenza AAUAAA 10-35nt prima della coda di poli(A). I mutanti di questo sito vengono degradati rapidamente.
Il processamento al 3’ puo’ essere diviso in due fasi: 1. Trascrizione della sequenza AAUAA, seguita dallo stallo della RNA Polimerasi II
e taglio endoribonucleolitico del trascritto. Questo processo inizia quando la sequenza del sito di poliadenilazione e’ riconosciuta da fattori associati alla PolII, l’efficienza di terminazione dipende quindi dalla forza del sito di poliadenilazione
2. Il prodotto del taglio a monte viene poliadenilato, mentre il prodotto a valle e’ degradato.
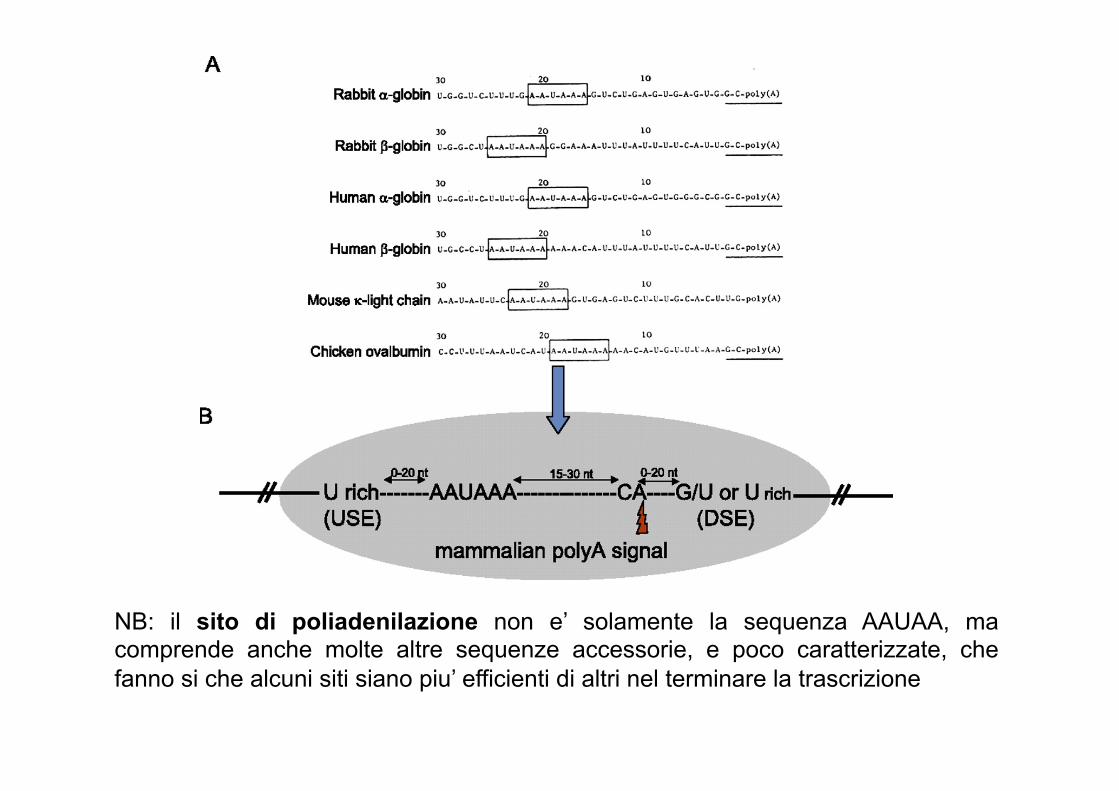
NB: il sito di poliadenilazione non e’ solamente la sequenza AAUAA, ma comprende anche molte altre sequenze accessorie, e poco caratterizzate, che fanno si che alcuni siti siano piu’ efficienti di altri nel terminare la trascrizione
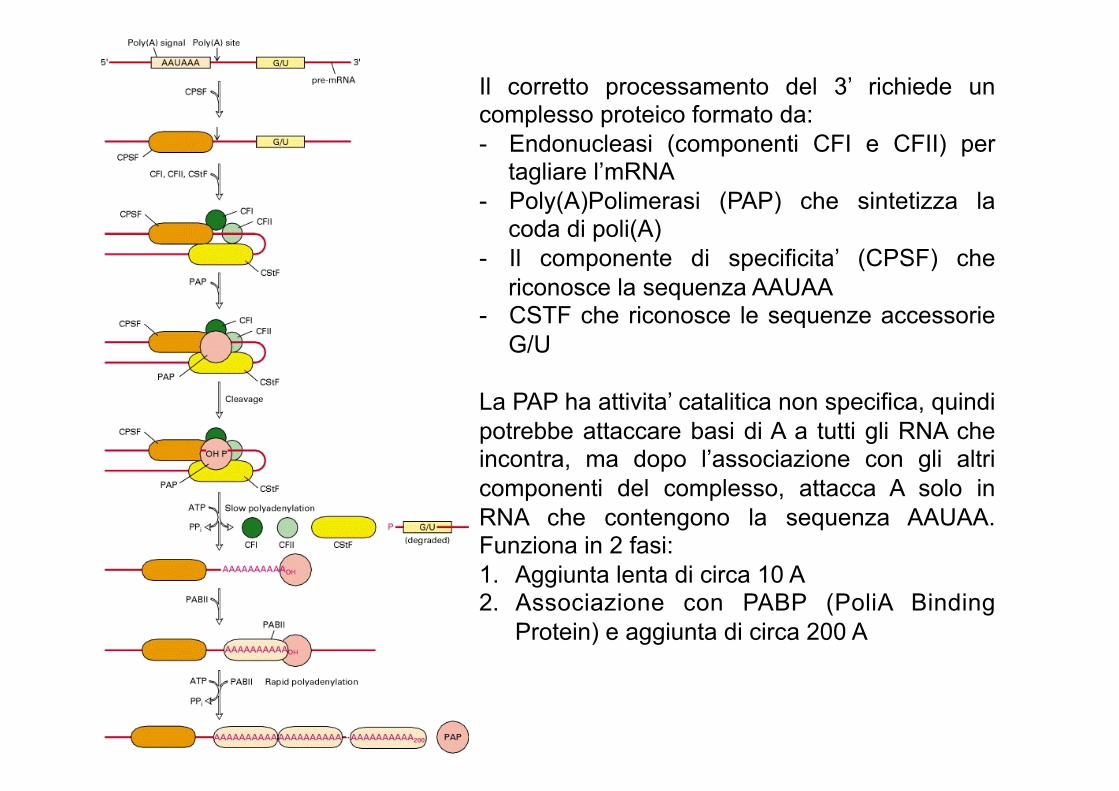
Il corretto processamento del 3’ richiede un complesso proteico formato da: - Endonucleasi (componenti CFI e CFII) per
tagliare l’mRNA - Poly(A)Polimerasi (PAP) che sintetizza la
coda di poli(A) - Il componente di specificita’ (CPSF) che
riconosce la sequenza AAUAA - CSTF che riconosce le sequenze accessorie
G/U
La PAP ha attivita’ catalitica non specifica, quindi potrebbe attaccare basi di A a tutti gli RNA che incontra, ma dopo l’associazione con gli altri componenti del complesso, attacca A solo in RNA che contengono la sequenza AAUAA. Funziona in 2 fasi: 1. Aggiunta lenta di circa 10 A 2. Associazione con PABP (PoliA Binding
Protein) e aggiunta di circa 200 A

https://www.youtube.com/watch?v=DoSRu15VtdM
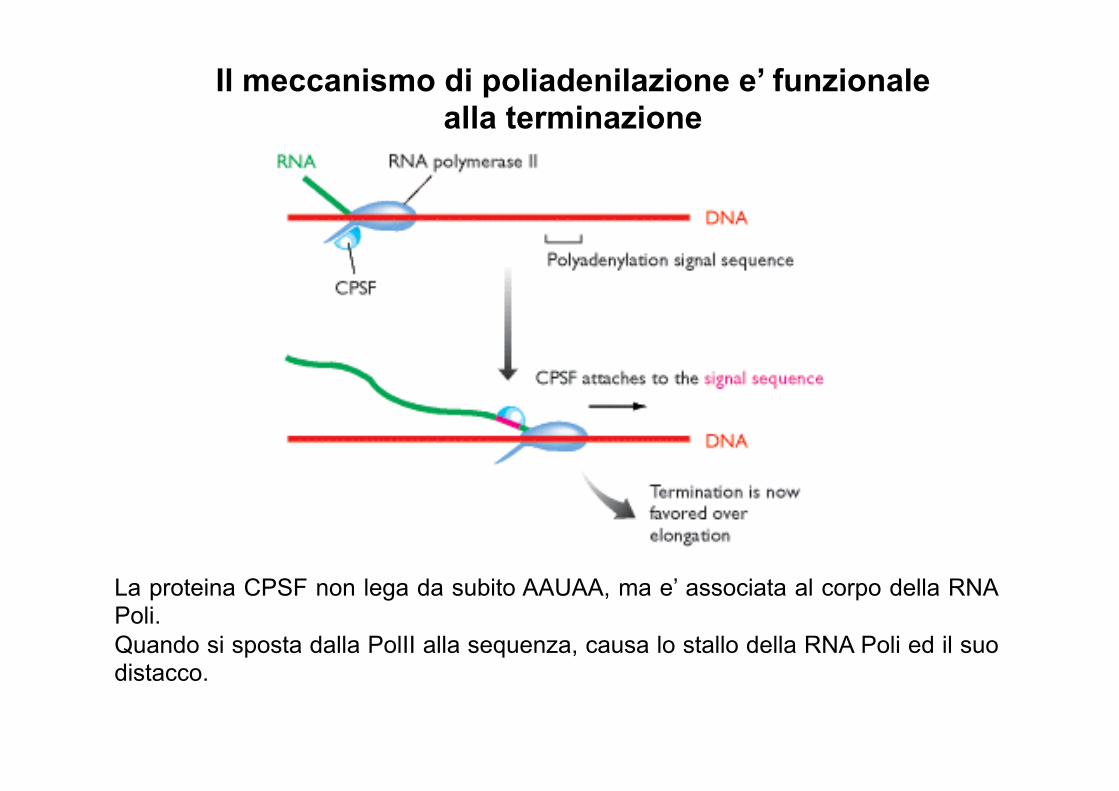
Il meccanismo di poliadenilazione e’ funzionale alla terminazione
La proteina CPSF non lega da subito AAUAA, ma e’ associata al corpo della RNA Poli. Quando si sposta dalla PolII alla sequenza, causa lo stallo della RNA Poli ed il suo distacco.
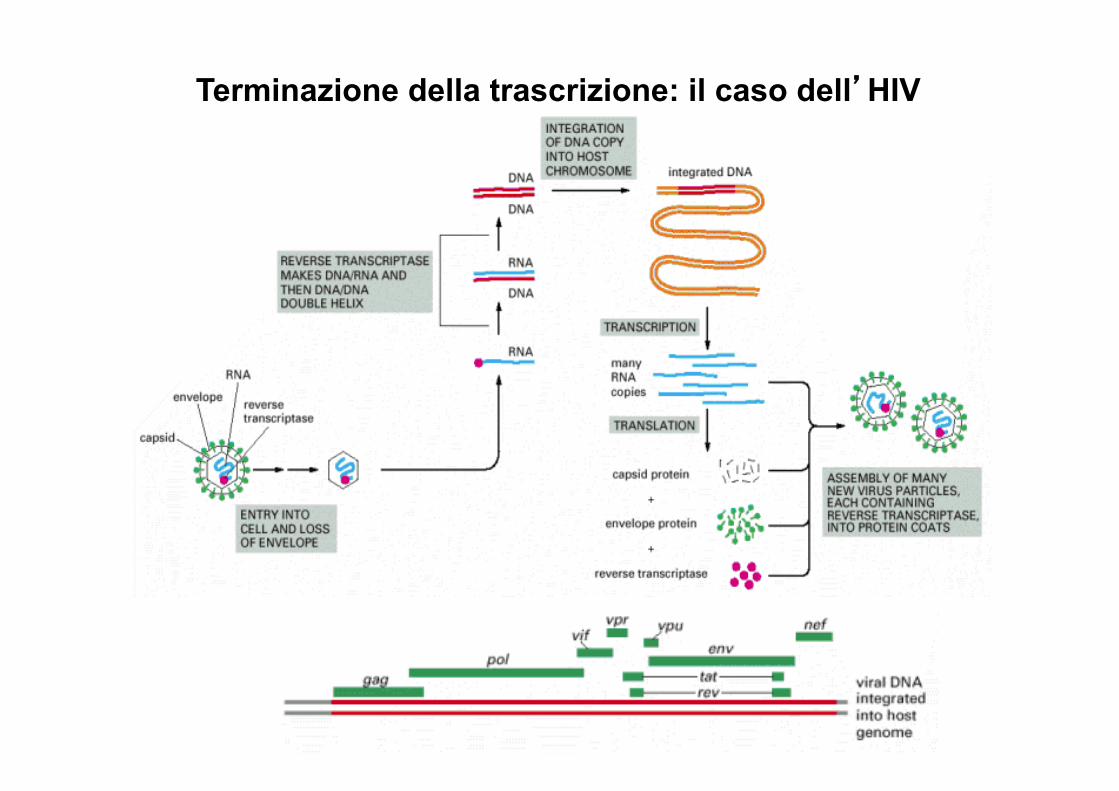
Terminazione della trascrizione: il caso dell’HIV
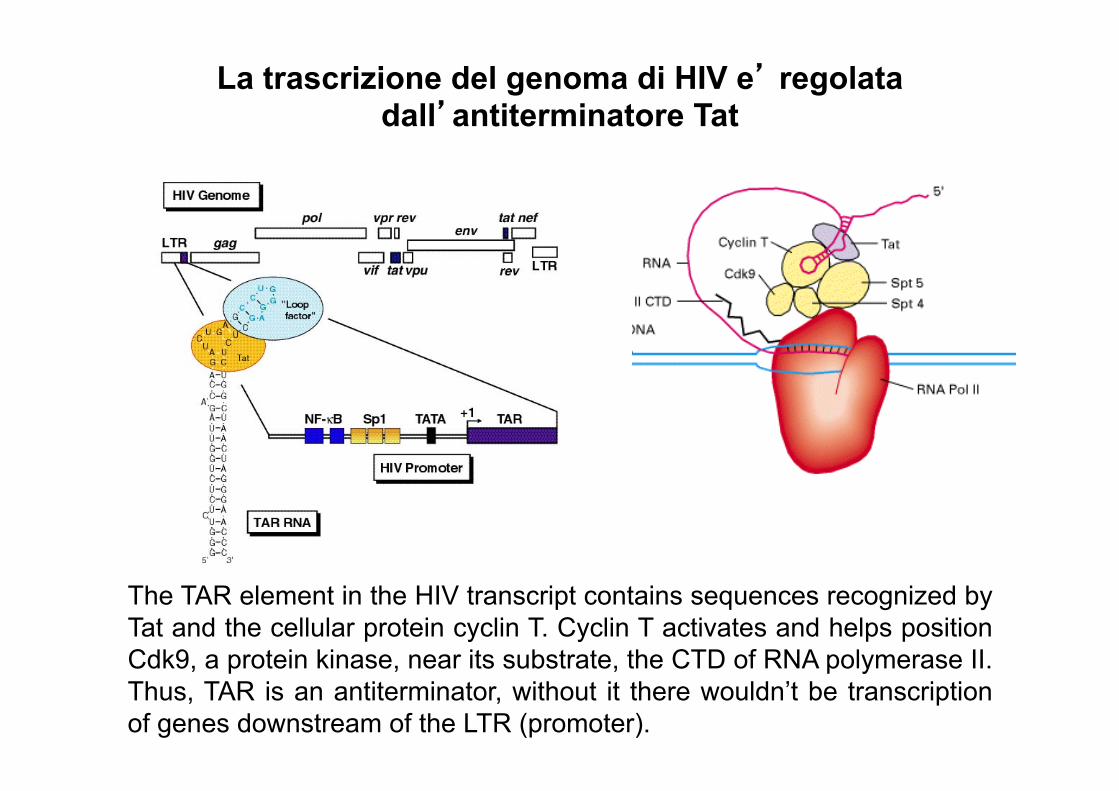
La trascrizione del genoma di HIV e’ regolata dall’antiterminatore Tat
The TAR element in the HIV transcript contains sequences recognized by Tat and the cellular protein cyclin T. Cyclin T activates and helps position Cdk9, a protein kinase, near its substrate, the CTD of RNA polymerase II. Thus, TAR is an antiterminator, without it there wouldn’t be transcription of genes downstream of the LTR (promoter).

Il cap serve per l’attacco ai ribosomi
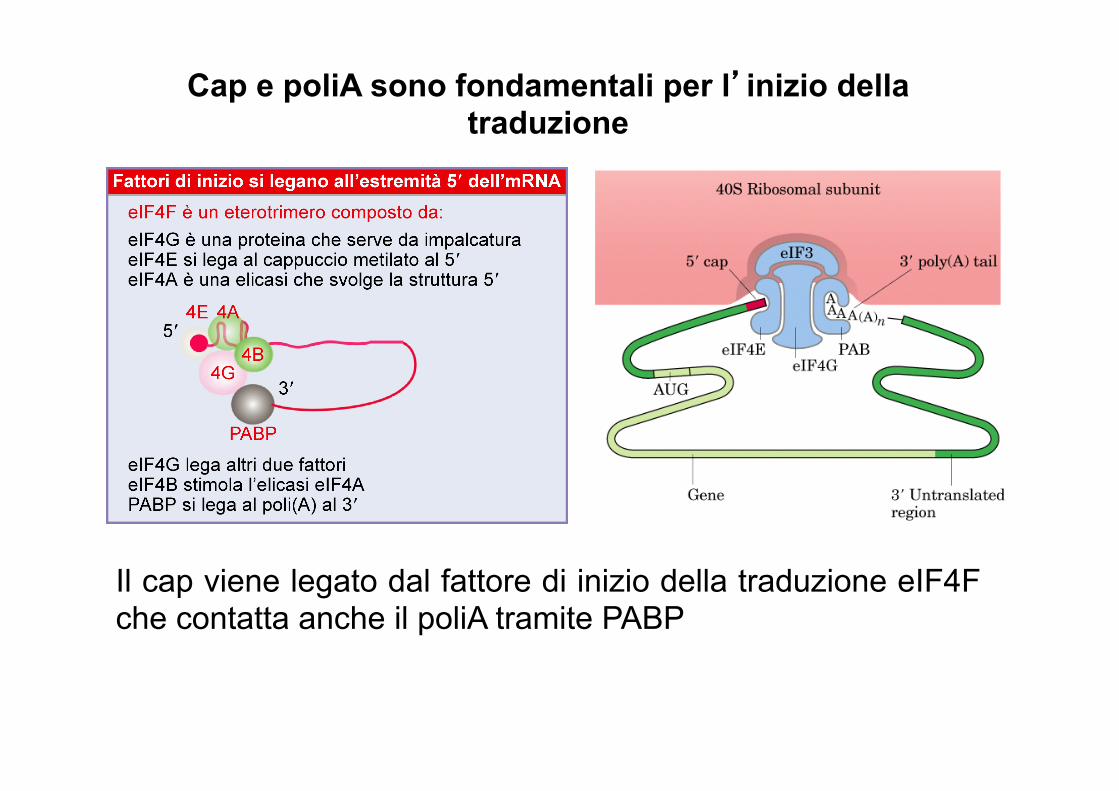
Il cap viene legato dal fattore di inizio della traduzione eIF4F che contatta anche il poliA tramite PABP
Cap e poliA sono fondamentali per l’inizio della traduzione
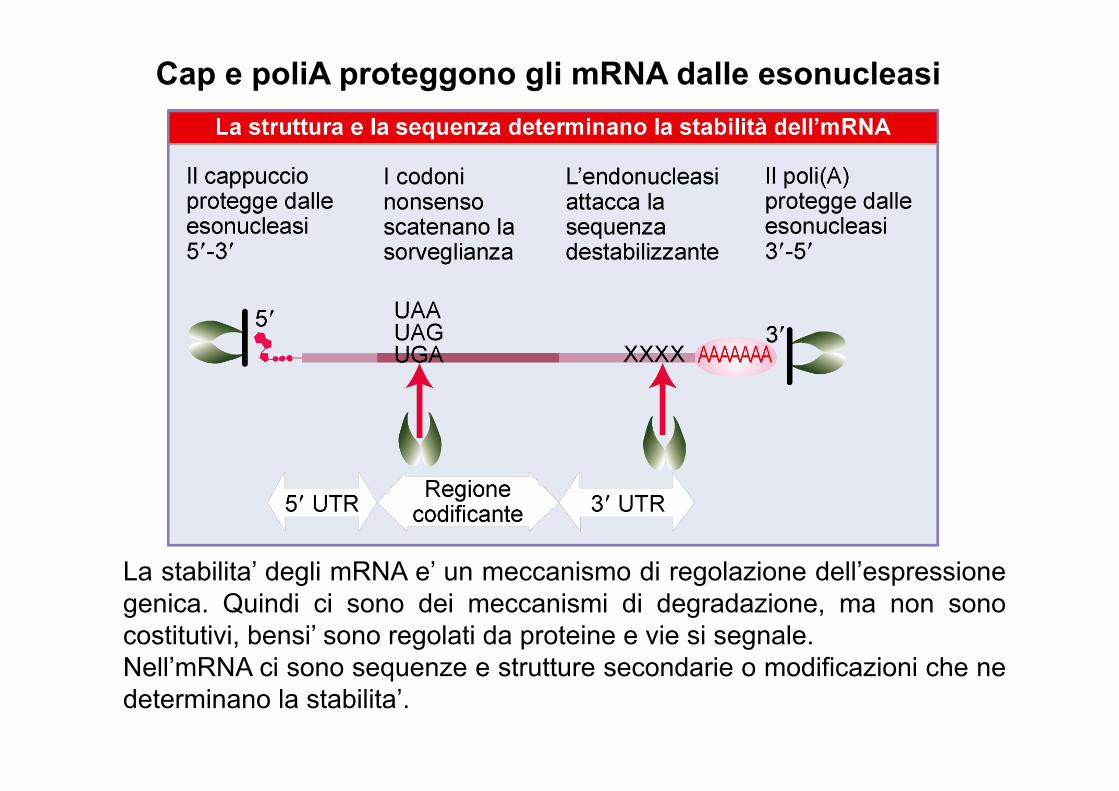
Cap e poliA proteggono gli mRNA dalle esonucleasi
La stabilita’ degli mRNA e’ un meccanismo di regolazione dell’espressione genica. Quindi ci sono dei meccanismi di degradazione, ma non sono costitutivi, bensi’ sono regolati da proteine e vie si segnale. Nell’mRNA ci sono sequenze e strutture secondarie o modificazioni che ne determinano la stabilita’.
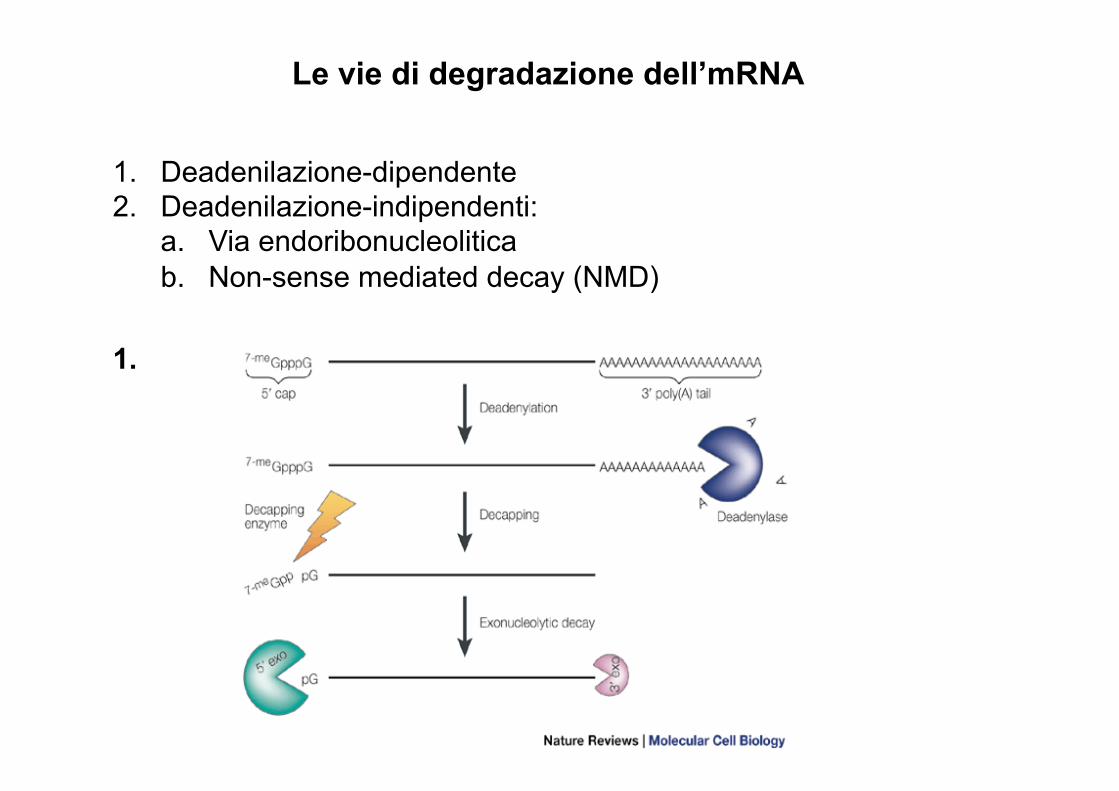
Le vie di degradazione dell’mRNA
1. Deadenilazione-dipendente 2. Deadenilazione-indipendenti:
a. Via endoribonucleolitica b. Non-sense mediated decay (NMD)
1.
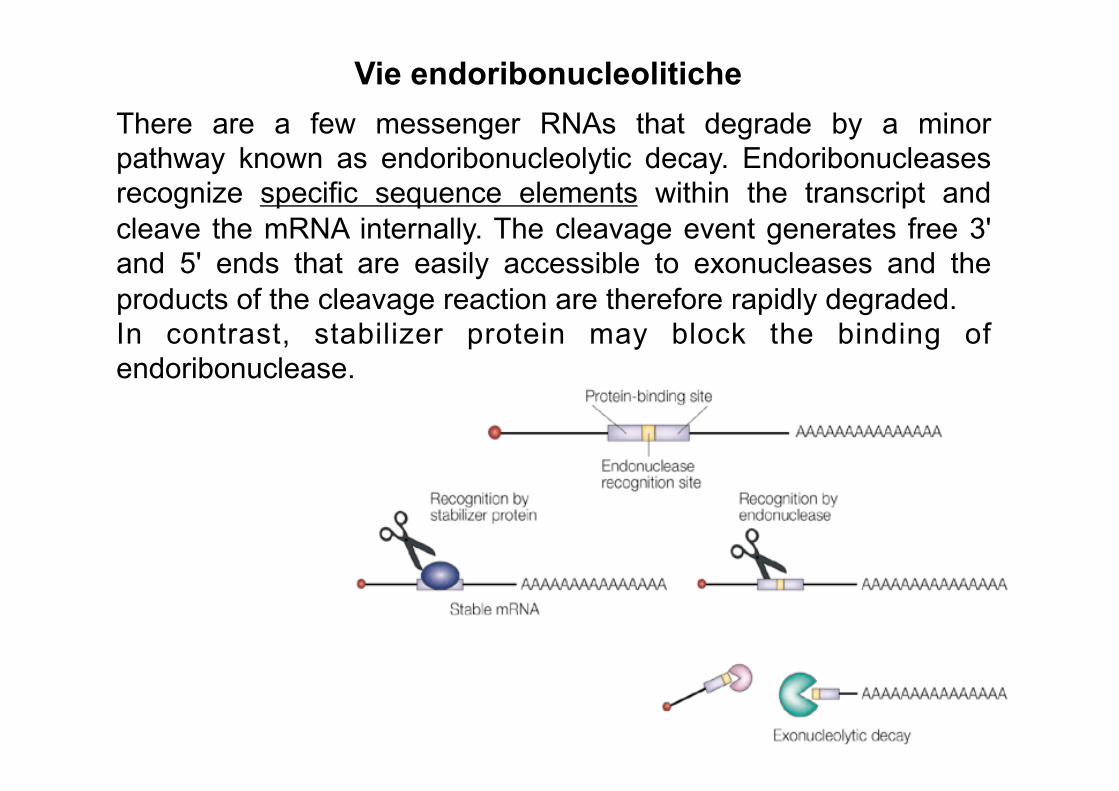
Vie endoribonucleolitiche There are a few messenger RNAs that degrade by a minor pathway known as endoribonucleolytic decay. Endoribonucleases recognize specific sequence elements within the transcript and cleave the mRNA internally. The cleavage event generates free 3' and 5' ends that are easily accessible to exonucleases and the products of the cleavage reaction are therefore rapidly degraded. In contrast, stabilizer protein may block the binding of endoribonuclease.
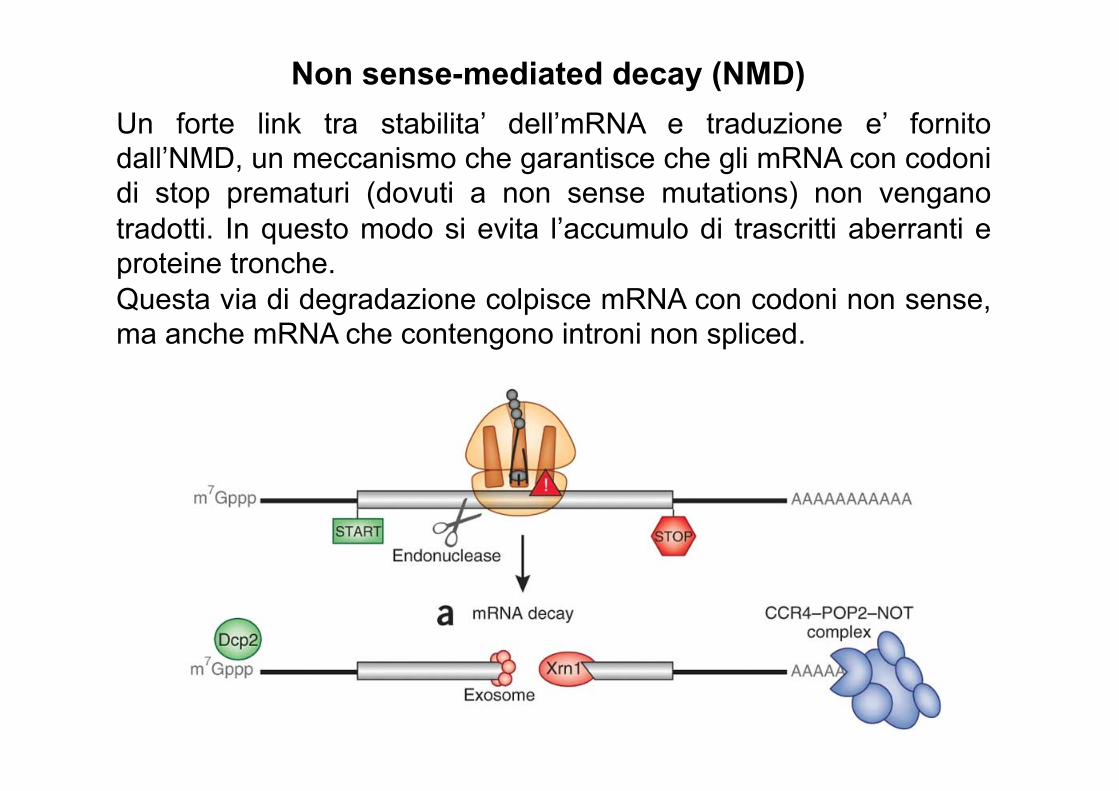
Non sense-mediated decay (NMD) Un forte link tra stabilita’ dell’mRNA e traduzione e’ fornito dall’NMD, un meccanismo che garantisce che gli mRNA con codoni di stop prematuri (dovuti a non sense mutations) non vengano tradotti. In questo modo si evita l’accumulo di trascritti aberranti e proteine tronche. Questa via di degradazione colpisce mRNA con codoni non sense, ma anche mRNA che contengono introni non spliced.
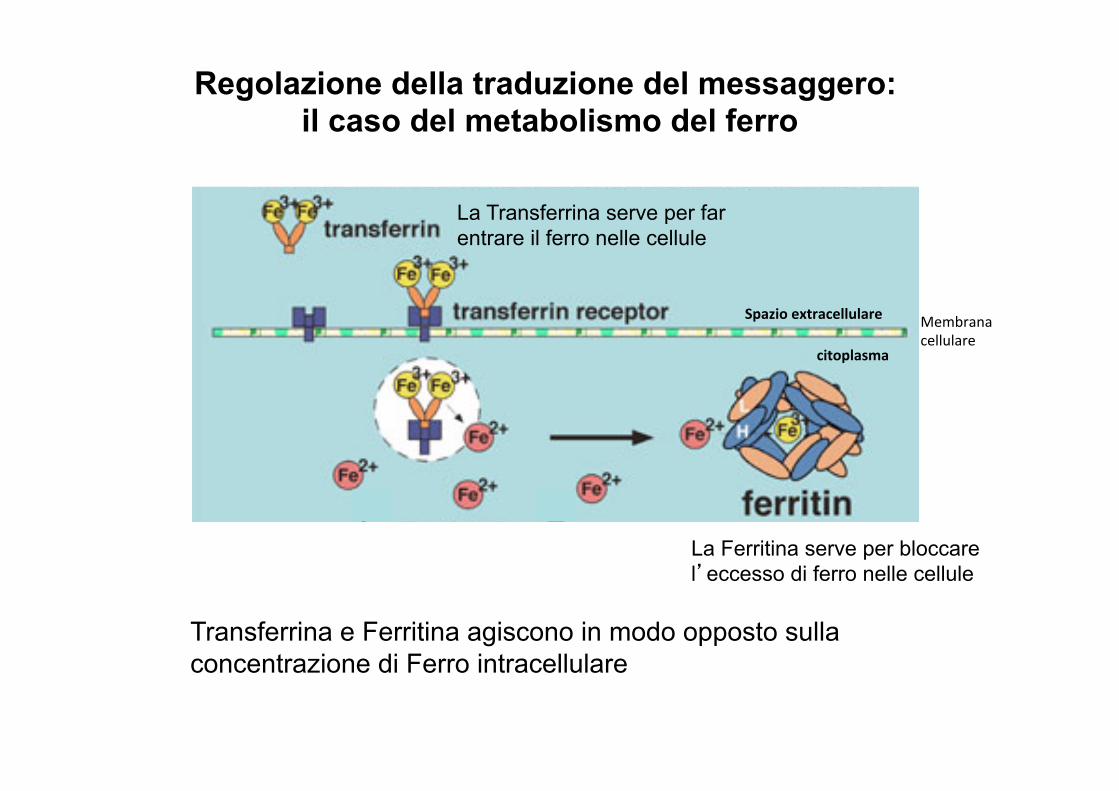
Regolazione della traduzione del messaggero: il caso del metabolismo del ferro
Spazio extracellulare
citoplasma
Membrana cellulare
La Transferrina serve per far entrare il ferro nelle cellule
La Ferritina serve per bloccare l’eccesso di ferro nelle cellule
Transferrina e Ferritina agiscono in modo opposto sulla concentrazione di Ferro intracellulare
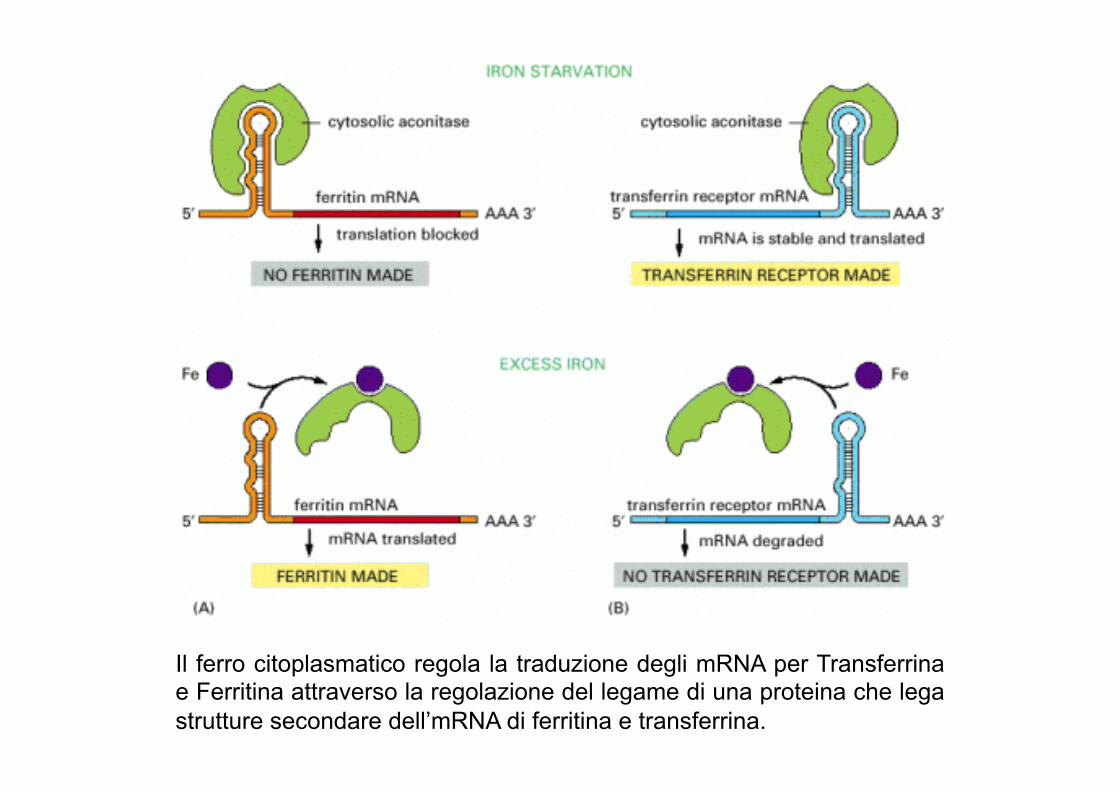
Il ferro citoplasmatico regola la traduzione degli mRNA per Transferrina e Ferritina attraverso la regolazione del legame di una proteina che lega strutture secondare dell’mRNA di ferritina e transferrina.