
Morfogenesi e auto-organizzazione degli aggregati...
49
Morfogenesi e auto-organizzazione degli aggregati cellulari Roberto Natalini Istituto per le Applicazioni del Calcolo – CNR Colloquio Interdisciplinare sulla Biologia – 26 aprile 2017 Universit` a di Roma Tor Vergata Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
Transcript of Morfogenesi e auto-organizzazione degli aggregati...

Morfogenesi e auto-organizzazionedegli aggregati cellulari
Roberto NataliniIstituto per le Applicazioni del Calcolo – CNR
Colloquio Interdisciplinare sulla Biologia – 26 aprile 2017Universita di Roma Tor Vergata
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Outline
1 Short biological background on cell movements2 Continuous models of chemotaxis3 A hybrid model for morphogenesis in zebrafish4 A simple model of collective motion under alignment and chemotaxis5 Cardiac stem cells and the growth of cardiospheres
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
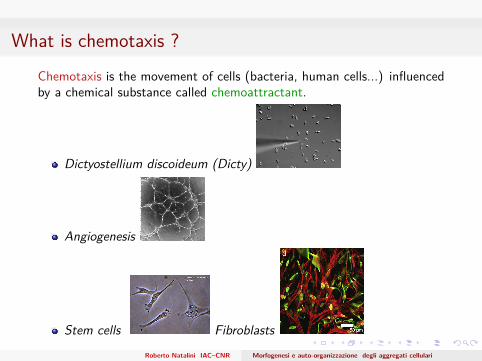
What is chemotaxis ?
Chemotaxis is the movement of cells (bacteria, human cells...) influencedby a chemical substance called chemoattractant.
Dictyostellium discoideum (Dicty)
Angiogenesis
Stem cells Fibroblasts
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Keller-Segel parabolic system
Classical parabolic model [Keller & Segel, 70]:
∂tu = ∇ (Du∇u − χ(u, φ)∇φ) ,
τ∂tφ = Dc ∆φ+ f (u, φ).
• u is the density of bacteria,• φ is the density of chemoattractant.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Keller-Segel parabolic system
Classical parabolic model [Keller & Segel, 70]:
∂tu = ∇ (Du∇u − χ(u, φ)∇φ) ,
τ∂tφ = Dc ∆φ+ f (u, φ).
Numerous theoretical and numerical results [Horstmann, 03 & 04] :• 1D : existence of global solutions• multiD : global existence (small initial mass) vs blow-up of solutions
in finite time (big initial mass).
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

A hyperbolic alternative: the Greenberg-Alt model
Discrete kinetic 1D model∂tu+ + γ ∂x u+ = −µ+(φ, φx ) u+ + µ−(φ, φx ) u−,∂tu− − γ ∂x u− = µ+(φ, φx ) u+ − µ−(φ, φx ) u−,∂tφ = D ∂xxφ+ f (u, φ).
• u± is the density of bacteria with speed ±γ• µ± are the relative turning rates (influenced by chemicals)• φ is the density of chemicals... , it is wave equation with relaxation!
∂tu + ∂x v = 0,∂tv + γ2 ∂x u = αu − βv ,∂tφ = D ∂xxφ+ f (u, φ).
with α = γ(µ− − µ+)and β = µ+ + µ−
• u = u+ + u− total density; v = γ(u+ − u−) total flux
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

A hyperbolic alternative: the Greenberg-Alt model
Discrete kinetic 1D model∂tu+ + γ ∂x u+ = −µ+(φ, φx ) u+ + µ−(φ, φx ) u−,∂tu− − γ ∂x u− = µ+(φ, φx ) u+ − µ−(φ, φx ) u−,∂tφ = D ∂xxφ+ f (u, φ).
• u± is the density of bacteria with speed ±γ• µ± are the relative turning rates (influenced by chemicals)• φ is the density of chemicals... , it is wave equation with relaxation!
∂tu + ∂x v = 0,∂tv + γ2 ∂x u = αu − βv ,∂tφ = D ∂xxφ+ f (u, φ).
with α = γ(µ− − µ+)and β = µ+ + µ−
• u = u+ + u− total density; v = γ(u+ − u−) total flux
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

A continuous model for the movement of brain stem cells after an ischemic event
A Model of Ischemia-Induced Neuroblast Activation in the Adult Subventricular Zone,D. Vergni, F. Castiglione, M. Briani, S. Middei, E. Alberdi, K. G. Reymann, R.Natalini, C. Volonte, C. Matute, F. Cavaliere, Plos One 2009.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Part IIIA hybrid mathematical model for self-organizing cells in the earlydevelopment of the zebrafish lateral lineA joint work with Ezio Di Costanzo and Luigi Preziosi, J. Math. Bio.2015
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Lateral lineA fundamental sensory system present in fish and amphibians.
Large variety of behaviours:detect movement and vibration inthe surrounding water;prey and predator detection;school swimming.
NeuromastsMain sensory organs of the lateral line, embedded in the body surface in arosette-shaped pattern: 1–2 sensory hair cells in the centre, surrounded by othersupport cells (8–12 cells).Neuromasts extend a ciliary bundle into the water, which detect movement in thesurrounding environment.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Attention of biologists in the lateral line to understand:interactions between multiple signaling and the collective migration of cellduring morphogenesis;how an organ responds to injury and replaces damaged components;how the genetic defects cause disorders in the nervous system.
Recent studies have investigated the zebrafish (Danio rerio) lateral line(Gilmour et al, 2008; Nechiporuk et al, 2008).
Zebrafish is an important vertebrate model organism in scientific research formany scientific reasons:
regenerative abilities (major organs are visible in 36 hpf);embryos are robust, transparent, easily observable and testable;genome has been fully sequenced.
Improvements in the fields of oncology, genetics, stem cell research, andregenerative medicine.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Experimental observationsAn initial elongated group (80-100 cells) of mesenchymal cells(primordium), with a trailing region near the head and a leading regiontowards the future tail of the embryo.
Two primary mechanisms in the morphogenesis process:1 a collective cell migration guided by a haptotactic signal, with
constant velocity of about 69 µm h−1;2 a process of differentiation in the trailing region that induces a
mesenchymal–epithelial transition and causes the neuromastsassembly and their detachment.
Movie zebrafish (Gilmour et al, 2006).Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
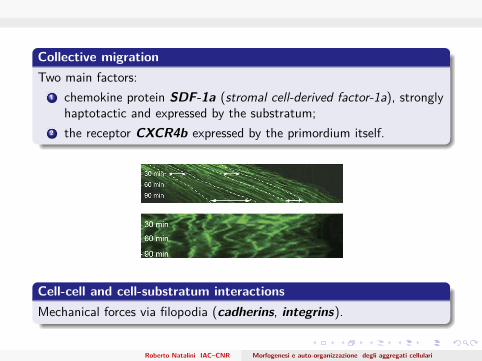
Collective migrationTwo main factors:
1 chemokine protein SDF-1a (stromal cell-derived factor-1a), stronglyhaptotactic and expressed by the substratum;
2 the receptor CXCR4b expressed by the primordium itself.
Cell-cell and cell-substratum interactionsMechanical forces via filopodia (cadherins, integrins).
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Neuromasts assemblyTwo main factors:
1 fibroblast growth factors FGF3–FGF10, strongly chemotactic;2 receptor FGFR.
Experimental observations on the FGF activity1 FGF3 and FGF10 are substantially equivalent (robustness of the
system);2 FGF and FGFR are mutually exclusive.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Leader to follower differentiation
leader mesenchymal cells: produce FGF but the receptor FGFR is not activated;follower epithelial cells: activate FGFR, but do not produce FGF.
Cyclic mechanism
1 at the beginning, all cells are leader;2 leader–follower differentiation (MET
transition) produces rosette-shapedstructures (proto-neuromasts);
3 neuromasts deposition.
Three sufficient conditions for the leader–follower transition
1 a low level of SDF-1a (trailing zone is preferred for transition);2 a high level of FGF;3 a lateral inhibition effect (leader/follower transition favored by a low number of
neighboring cells).
return
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

The mathematical modelOur aim is to obtain a minimal mathematical model which is able to:
1 describe the collective cell migration, the detachment of theneuromasts, in the physical spatial and temporal scale;
2 ensure the existence and stability of the rosette structures of theneuromasts, as stationary solutions.
Request 2) provides a restriction for some parameters.
Hybrid discrete in continuous descriptiondiscrete on cellular scale (but nonlocal sensing area);continuous at molecular scale.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

A second order modelXi (t) : position of the i-th cell;ϕi (t) : switch variable for the i-th cell (ϕi = 0, 1 resp. follower-leader);f (x, t) : concentration of FGF (equivalent FGF3 and FGF10);s(x, t) : concentration of SDF-1a;
acceleration i-th cell︷︸︸︷Xi =
haptotaxis︷ ︸︸ ︷αF1 (∇s) +
chemotaxis︷ ︸︸ ︷γ(1− ϕi )F1 (∇f ) +
alignment︷ ︸︸ ︷F2(X) +
attraction/repulsion︷ ︸︸ ︷F3(X)
−
damping︷ ︸︸ ︷[µF + (µL − µF)ϕi ] Xi ,
leader-follower state︷︸︸︷ϕi =
0, if
SDF conc.︷ ︸︸ ︷δF1(s) −
FGF conc.︷ ︸︸ ︷[kF + (kL − kF)ϕi ] F1(h(f )) +
lateral inhib.︷ ︸︸ ︷λΓ(ni ) ≤ 0,
1, otherwise,
FGF rate in time︷︸︸︷∂t f =
diffusion︷︸︸︷D∆f +
production︷ ︸︸ ︷ξF4(X) −
molecular degradation︷︸︸︷ηf ,
SDF rate in time︷︸︸︷∂t s = −
degradation︷ ︸︸ ︷σsF5(X) ,
Steady statesRoberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Xi = αF1 (∇s) + γ(1− ϕi )F1 (∇f ) + F2(X) + F3(X)− [µF + (µL − µF)ϕi ] Xi .
Alignment effectCucker and Smale–like flocking term:
F2(X) :=1Ni
∑j:Xj∈B(Xi ,R1)\Xi
H(Xj − Xi ),
H(Xj − Xi ) := [βF + (βL − βF)ϕiϕj ]R2
1R2
1 + ||Xj − Xi ||2(Xj − Xi ),
Ni := card j : Xj ∈ B(Xi ,R1) .
Unlike the original Cucker-Smale model:flocking term is coupled with other effects (chemotaxis,attraction/repulsion, damping);it acts on a truncated domain. Alignment effect
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Xi = αF1 (∇s) + γ(1− ϕi )F1 (∇f ) + F2(X) + F3(X)− [µF + (µL − µF)ϕi ] Xi .
Adhesion-repulsion effect
F3(X) :=∑
j:Xj∈B(Xi ,R5)\Xi
K(Xj − Xi ),
K(Xj − Xi ) :=
−ωrep
(1
||Xj − Xi ||−
1R4
)Xj − Xi
||Xj − Xi ||, if ||Xj − Xi || ≤ R4;
ωadh (||Xj − Xi || − R4)Xj − Xi
||Xj − Xi ||, if R4 < ||Xj − Xi || ≤ R5;
Similar terms can be found in D’Orsogna et al (2006), Cristiani et al(2011), Albi and Pareschi (2013).
Three zone models
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Lateral inhibition mechanism
Leader to Follower transition
We count the number ni of cells in a suitable neighborhood of the i-th cell,with radius of influence R2:
Γ(ni ) :=eni
eni + Γ0− 1
1 + Γ0,
ni := card
j : Xj ∈ B(Xi ,R2)\ Xi,
Sharp difference between a cell in the centre or a cell on the boundary;Fast saturation effect as ni increases (possibility to obtain neuromastswith 8-12 cells). Full Model
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Steady states and stability
Particular steady states, biologically relevant, corresponding to theneuromast basic structure.
Stationary model
γ(1− ϕi )
W
∫B(Xi ,R)
∇f (x)wi (x) dx +∑
j:Xj∈B(Xi ,R5)\Xi
K(Xj − Xi ) = 0,
ϕi =
0 if −
kF + (kL − kF)ϕi
W
∫B(Xi ,R)
f (x)
1 + f (x)wi (x) dx + Γ(ni ) ≤ 0,
1 otherwise,
D∗∆f = ηf − ξNtot∑j=1
ϕjχB(Xj ,R3),∂f∂n
= 0, on ∂Ω.
s = 0,
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

DefinitionA N-rosette (N ≥ 2) is a configuration formed by a leader cell surrounded byN follower cells located on the vertices of a regular polygon of N sides centeredin the leader cell.
Hypotheses1 the range of lateral inhibition equal to the range of repulsion between cells;2 the followers are located in the range of the lateral inhibition of the leader;3 there is no repulsion between adjoining followers if N = 2, 3;
there is no repulsion between followers in alternating position if N ≥ 4.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Proposition 1There exist N-rosettes if and only if N ≤ 12.Moreover the distance d1, depending on N,can vary in the following ranges:
12 sin π
N≤ d1
R4≤ 1, if N = 2, 3,
12 sin 2π
N≤ d1
R4≤ 1, if 4 ≤ N ≤ 12.
The maximum number of cells is consistent with the experimental observations(Gilmour et al, 2008).
Proposition 2In a N-rosette there are repulsion and lateral inhibition effects betweenadjoining followers if and only if N ≥ 4. In particular if N ≥ 6 these effects donot depend on d1, and if N = 4, 5 this holds if and only if
12 sin 2π
N≤ d1
R4<
12 sin π
N, N = 4, 5. Parameter range
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Numerical test of the steady solution.Dynamical simulation with initial zero velocity, a FGF concentrationgiven by the stationary equation
D∆f = ηf − ξχB(XL,R3),∂f∂n = 0, on ∂Ω,
and a zero initial concentration of SDF-1a.
Figure : A 8-rosette
0 5 10 15 20 25 30 35 40 45 500
0.5
1
1.5
2
2.5
3x 10
−3
Emax,rel
Time (h)
Figure :maxi ‖Xi (t)− Xi0‖ /R
0 5 10 15 20 25 30 35 40 45 500
1
2
3
4
5
6
7x 10
−4
Time (h)
Vmax(µm
h−1)
Figure : maxi∥∥Xi (t)
∥∥Numerical simulation
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
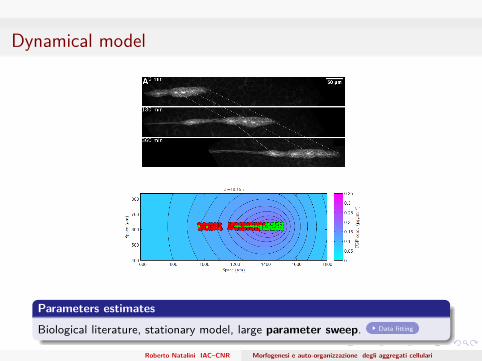
Dynamical model
Parameters estimates
Biological literature, stationary model, large parameter sweep. Data fitting
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
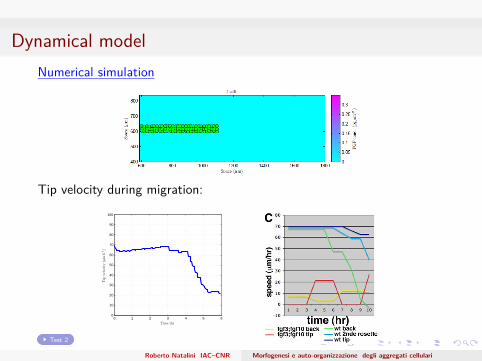
Dynamical modelNumerical simulation
Tip velocity during migration:
0 1 2 3 4 5 60
10
20
30
40
50
60
70
80
90
100
Time (h)
Tip
velocity
(µmh−1)
Test 2
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Part IVA hybrid mathematical model of collective motion under alignment andchemotaxis (joint with Ezio Di Costanzo): a short analytical intermezzo
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Cucker-Smale modelBehaviour of a flock of birds, general phenomena where autonomousagents reach a consensus. Vi =
β
N
N∑j=1
α1
(α2 + ||Xi − Xj ||2)σ(Vj − Vi );
Xi = Vi ,
i = 1, . . . ,N.
σ: rate of decay of the influence between agents.
Cucker and Smale (2007), Ha et al (2009), see also J. A. Carrillo, et al(2010) proved that:
if 0 ≤ σ ≤ 1/2 there is unconditional flocking (Movie with σ = 1/2);if σ > 1/2 there is conditional flocking (Movie with σ = 2).
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Cucker-Smale modelVi =
β
N
N∑j=1
α1(α2 + ‖Xi − Xj‖2)σ (Vj − Vi ),
Xi = Vi ,
Applications: biological field, collective dynamics of different interacting groups(Szabo et al, 2006; Sepulveda et al, 2013; Albi and Pareschi, 2013).Extensions: repulsion, individuals with preferred directions.
Collective motion under alignment and chemotaxis
Vi =β
N
N∑j=1
1(1 +‖Xi−Xj‖2
R2
)σ (Vj − Vi ) + γ∇f (Xi ),
Xi = Vi ,
∂t f = D∆f + ξ
N∑j=1
χB(Xj ,R) − ηf ,
(1)
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Existence and uniqueness
Theorem 1 (Local existence and uniqueness)System (1) has a unique solution on [0,T ], where T is suitably estimated.
Proposition 1 (Continuation of solutions)Let y(t) be a solution of system (1) on a interval [0,T ), if there is aconstant P with ‖y− y0‖ ≤ P on [0,T ), then there is a T > T suchthat y(t) can be continued to [0, T ].
Theorem 2 (Global existence and uniqueness)System (1) has a unique solution in all [0,+∞). go to
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Numerical simulations
Simulation 1. Absence of chemotaxis (γ = 0).Simulation 2. Strong flocking state.Simulation 3. Absence of alignment (β = 0).
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Part VE. Di Costanzo, A. Giacomello, E. Messina, R. Natalini, G. Pontrelli, F.Rossi, R. Smits, M. Twarogowska A discrete in continuous mathematicalmodel of cardiac progenitor cells formation and growth as spheroidclusters (Cardiospheres)
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

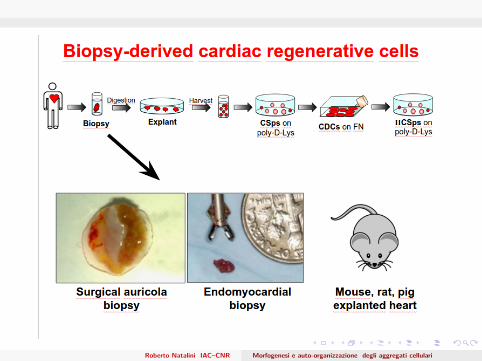
Cardiac stem cells
Cardiac biopsy-derived progenitor cells, growing in vitro as niche-likemicrotissue (Cardiospheres), represent a widely adopted platformtechnology holding great promise to realize a powerful cell therapy systemCardiosphere are spheroid of cellular clusters with a central nucleus ofless differentiated elements surrounded by outer layers of cells morecommitted toward different levels of cardio-vascular differentiation andproliferation. (Messina et al. 2004, Chimenti et al. 2011)
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Cardiac stem cells
Cardiac biopsy-derived progenitor cells, growing in vitro as niche-likemicrotissue (Cardiospheres), represent a widely adopted platformtechnology holding great promise to realize a powerful cell therapy systemCardiosphere are spheroid of cellular clusters with a central nucleus ofless differentiated elements surrounded by outer layers of cells morecommitted toward different levels of cardio-vascular differentiation andproliferation. (Messina et al. 2004, Chimenti et al. 2011)
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

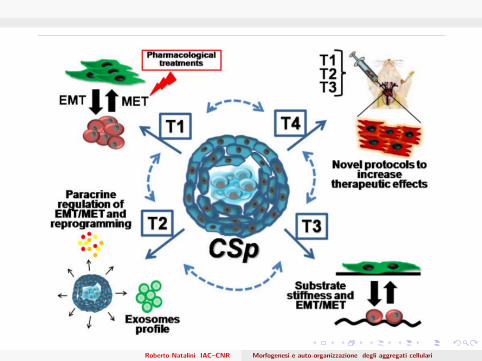
A first model of cardiospheres
Main goalsCell aggregation in spherical layers, with a realistic densityDifferentiation towards the external layersDifferentiation and proliferation are driven by jump processes,submitted to environmental factorsUnderstanding and control the role of the environmental factors(growth factors and nutrients)
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

A first model of cardiospheres
Xi (t) position of the i-eme cell at time tc = c(x, t) concentration of oxigen (nutrients)S = S(x, t) concentation of TGF (chemical signal)
Xi =∑
j:Xj∈B(Xi ,R2)\Xi
K(rij )− µXi + αF(∇S(Xi , t))
∂tc = ∇ · (Dc∇c)−N∑
i=1
λ(ϕi )cγ+1
k(ϕi ) + cγ+1χB(Xi ,R3)+ HB(c0 − c),
∂tS = ∇ · (DS∇S) +N∑
i=1ξ(ϕi )χB(Xi ,R3)
− ηS
with D(x) := Dmax
1+ρA(x), where A(x) is the fraction of occupied volume near
x.
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Differentiation
cells undergoing different states of differentiation:
ϕi (t) = 1, 2, 3, d
(d=dead)
ϕi (t + ∆t) = 1 ϕi (t + ∆t) = 2 ϕ(t + ∆t) = 3ϕi (t) = 1 X Xϕi (t) = 2 X Xϕi (t) = 3 X
Table : Possible levels of differentiation between time t and t + ∆t
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Differentiation II
Differentiation is given by a non homogeneous Poisson processΓi : inhibition factor for the i-eme cell at time t
Γi (t) =
1, if ni ≤ 4,0, if ni > 4, ni := cardj : Xj ∈ B(Xi ,R4), i = 1, . . . ,N.
qi (t) :=σ(ϕi ) · F(S(Xi , t)) · Γi (t)
Smax, i = 1, . . . ,N, diff. treshold
where σ(ϕi ) (ϕi = 1, 2) are constants and Smax is the max of TGF-β
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Differentiation III
qi (t) is the intensity of the Poisson process
P(ϕi (t + ∆t)− ϕi (t) ≥ 1) u qi (t)∆t, (2)
which yields the stochastic differential equation
dϕi = dP(∫ t
0qi (s) ds
), i = 1, . . . ,N, (3)
where P(∫ t
0 qi (s) ds) is the process. A computational approach is
ϕi (t + ∆t) = ϕi (t) +
1, se qi < qi ∆t,0, altrimenti, i = 1, . . . ,N,
where qi is random in [0,1].
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
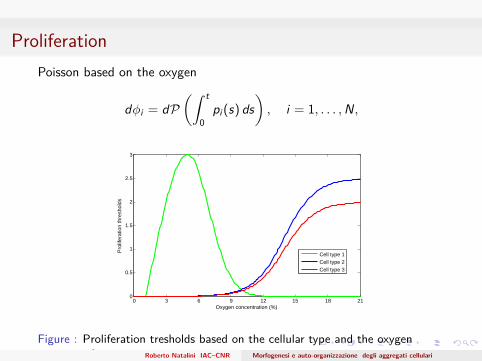
ProliferationPoisson based on the oxygen
dφi = dP(∫ t
0pi (s) ds
), i = 1, . . . ,N,
0 3 6 9 12 15 18 210
0.5
1
1.5
2
2.5
3
Oxygen concentration (%)
Pro
lifer
atio
n th
resh
olds
Cell type 1Cell type 2Cell type 3
Figure : Proliferation tresholds based on the cellular type and the oxygenconcentration Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
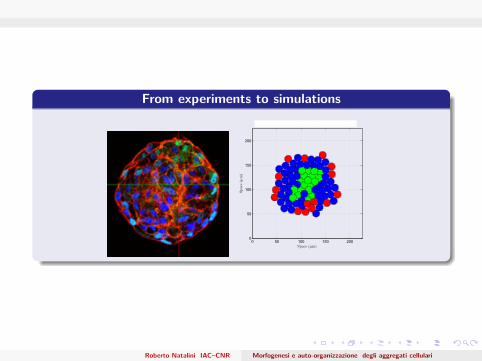
From experiments to simulations
0 50 100 150 2000
50
100
150
200
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
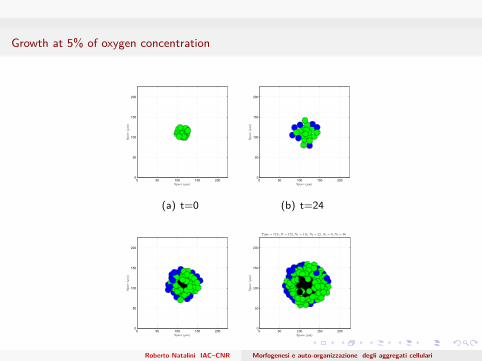
Growth at 5% of oxygen concentration
0 50 100 150 2000
50
100
150
200
(a) t=0
0 50 100 150 2000
50
100
150
200
(b) t=24
0 50 100 150 2000
50
100
150
200
(c) t=48
0 50 100 150 2000
50
100
150
200
Space (µm)
Space(µm)
Time = 72h; N = 172, N1 = 114, N2 = 22, N3 = 0, Nd = 36
(d) t=72Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
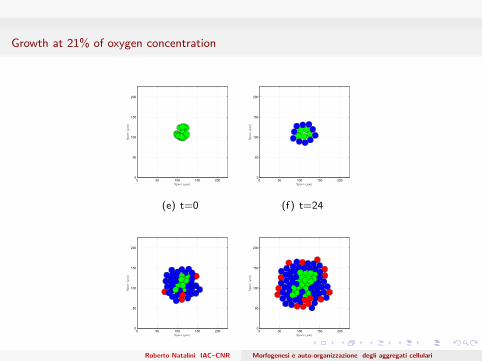
Growth at 21% of oxygen concentration
0 50 100 150 2000
50
100
150
200
(e) t=0
0 50 100 150 2000
50
100
150
200
(f) t=24
0 50 100 150 2000
50
100
150
200
(g) t=48
0 50 100 150 2000
50
100
150
200
(h) t=72Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari
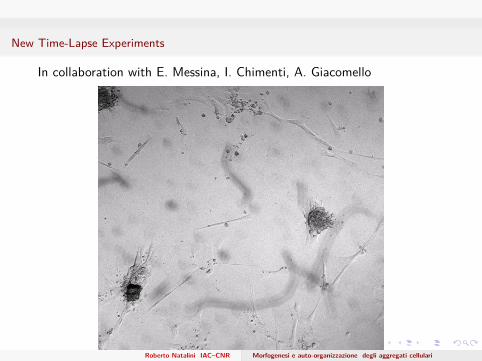
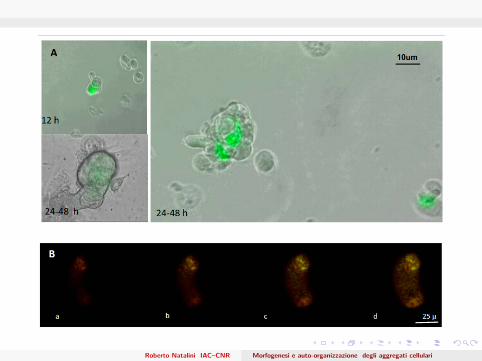
New Time-Lapse Experiments
In collaboration with E. Messina, I. Chimenti, A. Giacomello
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari

Some references1 E. Di Costanzo, R. Natalini, L. Preziosi L (2015) A hybrid mathematical model
for self- organizing tissue migration in the zebrafish lateral line. Journal ofMathematical Biology,
2 E. Di Costanzo, R. Natalini A hybrid mathematical model of collective motionunder alignment and chemotaxis, preprint arXiv:1507.02980.
3 E. Di Costanzo, A. Giacomello, E. Messina, R. Natalini, G. Pontrelli, F. Rossi, R.Smits, M. Twarogowska, (2017) A discrete in continuous mathematical model ofcardiac progenitor cells formation and growth as spheroid clusters(Cardiospheres), Mathematical Medicine & Biology .
Roberto Natalini IAC–CNR Morfogenesi e auto-organizzazione degli aggregati cellulari


















