
L'assemblaggio di un virus -...
4
In questa microfotografia elettronica, scattata da John T. Finch del Medical Research Council Laboratory of Molecular Biology di Cam- bridge (Inghilterra), le particelle del virus del mosaico del tabacco, dalla caratteristica forma a bastoncino, risultano ingrandite di 300 000 volte. Il virus procede in provetta, a un autoassemblaggio spontaneo, a partire dalle molecole di RNA e dalle subunità proteiche. L'assem- blaggio del virus costituisce un modello per sapere qual è l'estensione delle strutture che vengono costruite all'interno delle cellule viventi. / virus sono particelle complesse, costi- tuite da molecole giganti inerti: pro- teine e acidi nucleici (DNA o RNA). Sono inerti nel senso che non hanno alcun metabolismo interno ma, quando entrano in una cellula vivente, diventano vivi essi stessi. Per questo motivo sono «parassiti obbligati», in grado di riprodursi soltanto se utilizzano il meccanismo enzimatico dell'ospite. Grazie alla loro estrema semplicità, essi si sono dimostrati d'incommensurabile valore per gli studiosi di biologia moleco- lare, interessati alla struttura e alla fun- zione dei geni. Inoltre, costituiscono un semplice modello di sviluppo cellulare, in quanto la loro moltiplicazione all'interno della cellula ospite coinvolge l'espressio- ne controllata di un piccolo numero di geni e l'aggregazione di alcune proteine in una struttura estremamente ordinata. L'assemblaggio dei virus è diventato per- tanto paradigmatico per la costruzione di grosse strutture molecolari all'interno delle cellule viventi. Il lavoro del nostro gruppo, presso il Medical Research Council Laboratory of Molecular Biology di Cambridge (Inghil- terra), si è polarizzato sul virus del mosai- co del tabacco, che infetta appunto le fo- glie del tabacco. Uno dei più semplici e vitali virus noti, è formato da un unico filamento di RNA, incluso in una specie di bastoncino cavo di natura proteica. L'RNA virale ha una lunghezza di 6400 nucleotidi e si intercala nelle spire di un'e- lica avvolta in maniera molto compatta e costituita di 2130 subunità proteiche identiche, per cui circa tre n ucleotidi del- l'RNA risultano legati a ciascuna subuni- tà proteica. La proteina circonda e isola l'RNA, proteggendolo da ogni danno fino a quando il virus non ha infettato con successo la cellula ospite. Una volta che l'RNA si trova all'interno di quest'ultima, si ha un processo di liberazione della pro- teina e i geni virali si mettono all'opera per generare grossi quantitativi di nuove particelle virali. Nel 1955, una classica serie di esperi- menti, condotti da Heinz Fraenkel-Con- rat e Robley C. Williams della Università di California a Berkeley, ha dimostrato che il virus del mosaico del tabacco può essere ricostituito in provetta a partire dalla proteina e dall'RNA isolati. Per semplice rimescolamento, si formano particelle virali infettive, strutturalmente indistinguibili dal virus originario. Per- tanto, l'intera informazione necessaria per la costruzione del virus è insita nelle sue parti, che effettuano in soluzione un «autoassemblaggio» spontaneo. L'autoassemblaggio di una struttura a elica, qual è quella di un virus del mosaico del tabacco, può sembrare un fatto non particolarmente impressionante. Si può postulare che le subunità proteiche ab- biano una geometria di superficie ben precisa per cui si possono aggregare sol- tanto in un modo: esse stabiliscono una con l'altra contatti identici, che si ripeto- no continuamente e danno una struttura regolare. In base allo schema più ovvio, l'RNA libero interagisce con le singole subunità proteiche per dare inizio alla formazione dell'elica. Quindi le subunità si aggiungono semplicemente, una o po- che alla volta, al «gradino» che si trova all'estremità dell'elica in via di formazio- ne, proprio come cresce un cristallo con una dislocazione a vite. Solo che, in que- sto caso, le unità intrappolano l'RNA man mano che procedono. Dato che sia la particella virale sia l'RNA hanno estremi- tà distinte, è logico attendersi che la cre- scita cominci a un'estremità dell'RNA e proceda verso l'altra, ma, per quanto plausibili, si sa oggi che queste idee sem- plici sono errate. A posteriori si può capi- re perché il virus abbia adottato una stra- tegia che a prima vista può sembrare la più complessa possibile. primi esperimenti di ricostruzione hanno mostrato di avere molte carat- teristiche in comune con il processo natu- rale di assemblaggio. Il riassemblaggio artificiale del virus del mosaico del tabac- co si svolgeva soltanto a temperatura ambiente e a valori neutri di pH, condi- zioni simili a quelle che si trovano nelle cellule della pianta ospite. Inoltre esso era specifico: avanzava cioè più speditamente con l'RNA proveniente dallo stesso cep- po del virus o da un ceppo molto affine, mentre non procedeva affatto, o soltanto molto poco, con altri RNA naturali o con RNA sintetici. Il solo aspetto del processo di riassemblaggio che lasciava perplessi era la sua bassa velocità: erano necessarie sei ore o più per raggiungere le massime rese di particelle virali aggregate. Questo periodo di tempo sembrava troppo lungo per le condizioni normali dato che l'RNA virale è protetto da eventuali danni solo quando è completamente circondato dal- la proteina. L'assemblaggio di un qualsiasi grosso aggregato di subunità identiche, qual è un cristallo, può essere distinto in due stadi: la nucleazione e la crescita o, nel caso del virus del mosaico del tabacco, l'iniziazio- ne e l'allungamento. La fase che limita la velocità nell'assemblaggio di un virus, come nella maggior parte degli altri casi, è la fase detta «iniziazione» ossia avvio del processo da parte di un iniziatore. Dato l'elevato numero di subunità proteiche per giro d'elica (16 e 1/3), circa 18 subuni- tà separate dovrebbero legarsi alla mole- cola flessibile di RNA prima che la strut- tura in via di aggregazione si chiuda su se stessa e diventi qualcosa di più di un sem- plice aggregato proteico lineare lungo l'RNA. La difficoltà potrebbe essere ag- girata se esistesse una qualche «maschera di montaggio» su cui le prime spire dell'e- lica potrebbero riunirsi, fino a quando essa, raggiunta una sufficiente dimensio- ne, diventa stabile. Si è trovato che la soluzione del pro- blema stava in un'osservazione interes- sante: la proteina del rivestimento, da sola, prima dell'RNA virale, può aggre- garsi in un certo numero di forme distinte anche se correlate, piuttosto che in un'u- nica forma a elica. Donald L. D. Caspar della Brandeis University, di fronte a questa evidenza, ha previsto che alcune di L'assemblaggio di un virus Il virus del mosaico del tabacco consta di un filamento di acido nucleico, incluso in una proteina di forma bastoncellare. I due componenti si aggregano spontaneamente, ma in maniera notevolmente complessa di P. Jonathan G. Butler e Aaron Klug 24 25
-
Upload
trinhkhanh -
Category
Documents
-
view
220 -
download
0
Transcript of L'assemblaggio di un virus -...
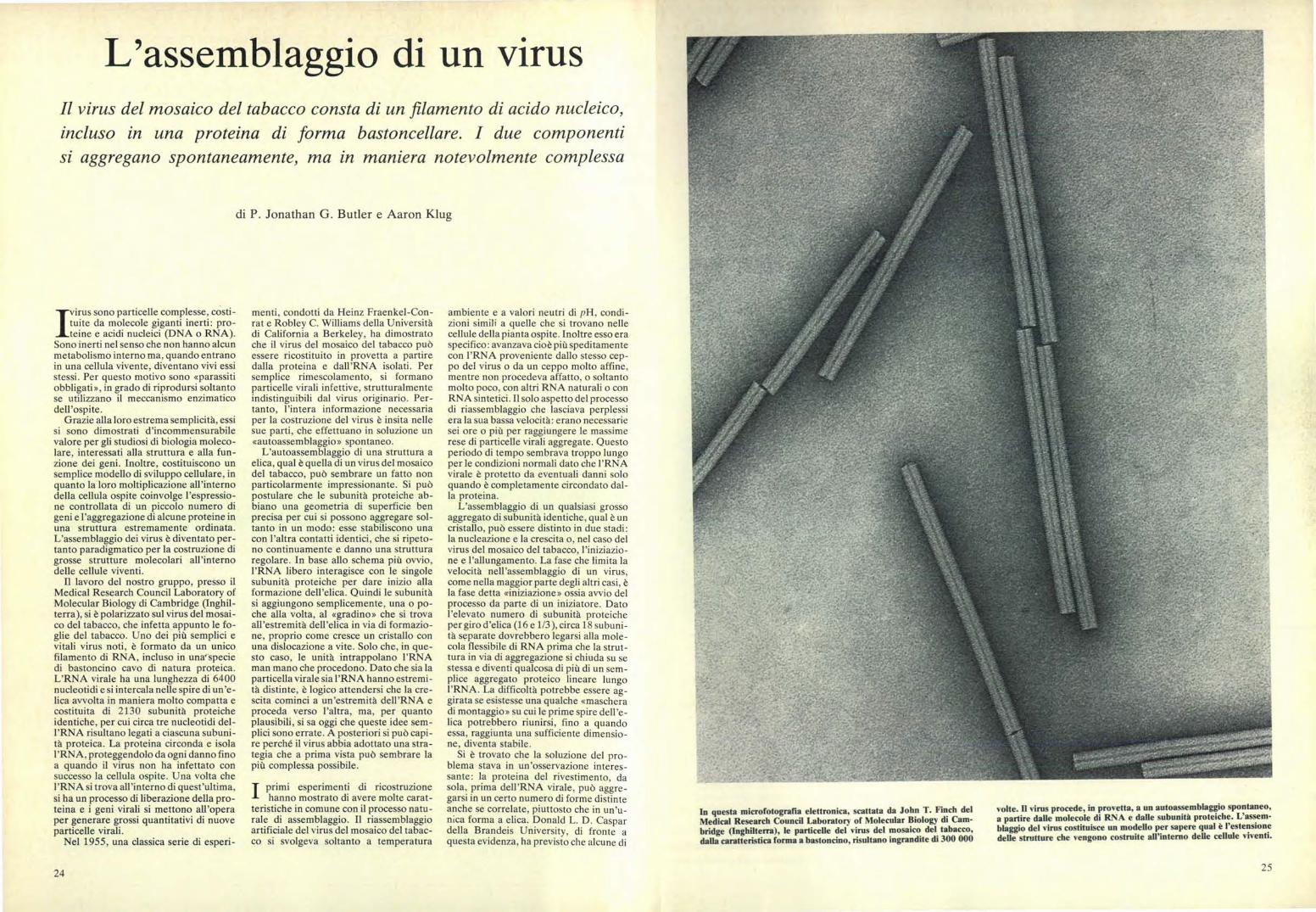
In questa microfotografia elettronica, scattata da John T. Finch delMedical Research Council Laboratory of Molecular Biology di Cam-bridge (Inghilterra), le particelle del virus del mosaico del tabacco,dalla caratteristica forma a bastoncino, risultano ingrandite di 300 000
volte. Il virus procede in provetta, a un autoassemblaggio spontaneo,a partire dalle molecole di RNA e dalle subunità proteiche. L'assem-blaggio del virus costituisce un modello per sapere qual è l'estensionedelle strutture che vengono costruite all'interno delle cellule viventi.
/
virus sono particelle complesse, costi-tuite da molecole giganti inerti: pro-teine e acidi nucleici (DNA o RNA).
Sono inerti nel senso che non hanno alcunmetabolismo interno ma, quando entranoin una cellula vivente, diventano vivi essistessi. Per questo motivo sono «parassitiobbligati», in grado di riprodursi soltantose utilizzano il meccanismo enzimaticodell'ospite.
Grazie alla loro estrema semplicità, essisi sono dimostrati d'incommensurabilevalore per gli studiosi di biologia moleco-lare, interessati alla struttura e alla fun-zione dei geni. Inoltre, costituiscono unsemplice modello di sviluppo cellulare, inquanto la loro moltiplicazione all'internodella cellula ospite coinvolge l'espressio-ne controllata di un piccolo numero digeni e l'aggregazione di alcune proteine inuna struttura estremamente ordinata.L'assemblaggio dei virus è diventato per-tanto paradigmatico per la costruzione digrosse strutture molecolari all'internodelle cellule viventi.
Il lavoro del nostro gruppo, presso ilMedical Research Council Laboratory ofMolecular Biology di Cambridge (Inghil-terra), si è polarizzato sul virus del mosai-co del tabacco, che infetta appunto le fo-glie del tabacco. Uno dei più semplici evitali virus noti, è formato da un unicofilamento di RNA, incluso in una speciedi bastoncino cavo di natura proteica.L'RNA virale ha una lunghezza di 6400nucleotidi e si intercala nelle spire di un'e-lica avvolta in maniera molto compatta ecostituita di 2130 subunità proteicheidentiche, per cui circa tre n ucleotidi del-l'RNA risultano legati a ciascuna subuni-tà proteica. La proteina circonda e isolal'RNA, proteggendolo da ogni danno finoa quando il virus non ha infettato consuccesso la cellula ospite. Una volta chel'RNA si trova all'interno di quest'ultima,si ha un processo di liberazione della pro-teina e i geni virali si mettono all'operaper generare grossi quantitativi di nuoveparticelle virali.
Nel 1955, una classica serie di esperi-
menti, condotti da Heinz Fraenkel-Con-rat e Robley C. Williams della Universitàdi California a Berkeley, ha dimostratoche il virus del mosaico del tabacco puòessere ricostituito in provetta a partiredalla proteina e dall'RNA isolati. Persemplice rimescolamento, si formanoparticelle virali infettive, strutturalmenteindistinguibili dal virus originario. Per-tanto, l'intera informazione necessariaper la costruzione del virus è insita nellesue parti, che effettuano in soluzione un«autoassemblaggio» spontaneo.
L'autoassemblaggio di una struttura aelica, qual è quella di un virus del mosaicodel tabacco, può sembrare un fatto nonparticolarmente impressionante. Si puòpostulare che le subunità proteiche ab-biano una geometria di superficie benprecisa per cui si possono aggregare sol-tanto in un modo: esse stabiliscono unacon l'altra contatti identici, che si ripeto-no continuamente e danno una strutturaregolare. In base allo schema più ovvio,l'RNA libero interagisce con le singolesubunità proteiche per dare inizio allaformazione dell'elica. Quindi le subunitàsi aggiungono semplicemente, una o po-che alla volta, al «gradino» che si trovaall'estremità dell'elica in via di formazio-ne, proprio come cresce un cristallo conuna dislocazione a vite. Solo che, in que-sto caso, le unità intrappolano l'RNAman mano che procedono. Dato che sia laparticella virale sia l'RNA hanno estremi-tà distinte, è logico attendersi che la cre-scita cominci a un'estremità dell'RNA eproceda verso l'altra, ma, per quantoplausibili, si sa oggi che queste idee sem-plici sono errate. A posteriori si può capi-re perché il virus abbia adottato una stra-tegia che a prima vista può sembrare lapiù complessa possibile.
primi esperimenti di ricostruzionehanno mostrato di avere molte carat-
teristiche in comune con il processo natu-rale di assemblaggio. Il riassemblaggioartificiale del virus del mosaico del tabac-co si svolgeva soltanto a temperatura
ambiente e a valori neutri di pH, condi-zioni simili a quelle che si trovano nellecellule della pianta ospite. Inoltre esso eraspecifico: avanzava cioè più speditamentecon l'RNA proveniente dallo stesso cep-po del virus o da un ceppo molto affine,mentre non procedeva affatto, o soltantomolto poco, con altri RNA naturali o conRNA sintetici. Il solo aspetto del processodi riassemblaggio che lasciava perplessiera la sua bassa velocità: erano necessariesei ore o più per raggiungere le massimerese di particelle virali aggregate. Questoperiodo di tempo sembrava troppo lungoper le condizioni normali dato che l'RNAvirale è protetto da eventuali danni soloquando è completamente circondato dal-la proteina.
L'assemblaggio di un qualsiasi grossoaggregato di subunità identiche, qual è uncristallo, può essere distinto in due stadi:la nucleazione e la crescita o, nel caso delvirus del mosaico del tabacco, l'iniziazio-ne e l'allungamento. La fase che limita lavelocità nell'assemblaggio di un virus,come nella maggior parte degli altri casi, èla fase detta «iniziazione» ossia avvio delprocesso da parte di un iniziatore. Datol'elevato numero di subunità proteicheper giro d'elica (16 e 1/3), circa 18 subuni-tà separate dovrebbero legarsi alla mole-cola flessibile di RNA prima che la strut-tura in via di aggregazione si chiuda su sestessa e diventi qualcosa di più di un sem-plice aggregato proteico lineare lungol'RNA. La difficoltà potrebbe essere ag-girata se esistesse una qualche «mascheradi montaggio» su cui le prime spire dell'e-lica potrebbero riunirsi, fino a quandoessa, raggiunta una sufficiente dimensio-ne, diventa stabile.
Si è trovato che la soluzione del pro-blema stava in un'osservazione interes-sante: la proteina del rivestimento, dasola, prima dell'RNA virale, può aggre-garsi in un certo numero di forme distinteanche se correlate, piuttosto che in un'u-nica forma a elica. Donald L. D. Caspardella Brandeis University, di fronte aquesta evidenza, ha previsto che alcune di
L'assemblaggio di un virusIl virus del mosaico del tabacco consta di un filamento di acido nucleico,incluso in una proteina di forma bastoncellare. I due componentisi aggregano spontaneamente, ma in maniera notevolmente complessa
di P. Jonathan G. Butler e Aaron Klug
24 25

pH
Le subunità proteiche si aggregano in forme diverse secondo il pH (concentrazione di protoni oidrogenioni) e la forza ionica (concentrazione salina) del mezzo circostante. Subunità proteichesingole si trovano soltanto in condizioni di forza ionica molto bassa e di forte alcalinità ( p H 10). Incondizioni meno alcaline (pH 8), compaiono ammassi di tre o più subunità. In una soluzioneneutra (pii 7), simile alla condizione che si riscontra nella cellula ospite, le subunità si dispongonoin dischi piatti, consistenti di 34 subunità disposte in due anelli. Se la soluzione viene acidificata(fino a circa pH 6,5), la proteina forma lunghe eliche prive di RNA. A pH neutro, invece, le eli-che si aggregano soltanto quando è presente l'RNA virale. La rondella, che consiste di un unicodisco, costituisce una forma intermedia che compare allorquando i dischi si trasformano in eliche.
La trasformazione del disco in un'elica in assenza di RNA virale si puòeffettuare abbassando ilp H della soluzione per rendere stabile l'aggre-gato elicoidale. È ciò che si vede in queste microfotografie elettronichescattate da Finch. (Nella metà inferiore dell'illustrazione le varie strut-ture sono disegnate schematicamente.) Se l'abbassamento del pH av-viene in modo sufficientemente rapido, i dischi proteici si trasformano
in corte eliche di appena due giri (rondelle), senza dissociazione insubunità (a). Nello spazio di pochi minuti, le rondelle si impilanoverticalmente e in modo casuale e danno delle corte eliche che presen-tano delle tacche (b). Queste si aggregano lentamente in circa 15 mi-nuti e formano una lunga elica, sempre con tacche (c). Tali imperfe-zioni in qualche ora scompaiono e si ottiene l'elica proteica finita (d).
DISCO ELICA CORTA CON TACCHE
RONDELLA
a C
ELICA LUNGA CON TACCHE
quelle forme avrebbero fornito una viad'interpretazione sul modo in cui il virussi aggrega. I vari stati di aggregazionesono stati esaminati in dettaglio per laprima volta dal nostro gruppo (di cui fa-cevano parte anche Anthony C. H. Dur-ham e John T. 7inch) e altri ricercatori, inseguito, hanno contribuito al completa-mento del quadro. Pur essendovi del di-saccordo per quanto riguarda i particola-ri, le linee generali sono ormai chiare: laproteina di rivestimento è progettata inmodo da sapere non solo dove deve anda-re (nell'elica virale), ma anche come.
Il fattore dominante che controlla lostato di aggregazione della proteina dirivestimento è il p H del mezzo circostante(cioè il logaritmo negativo della concen-trazione di protoni liberi, o idrogenioni,del mezzo). In una soluzione lievementealcalina (al disopra di p H 7), la proteina dirivestimento tende a presentarsi come unmiscuglio di piccoli aggregati di parecchiesubunità, miscuglio che viene denomina-to proteina A. Vicino alla neutralità (p H7), compare una struttura diversa e speci-fica: una pila di dischi consistenti di duestrati di subunità. Ogni strato è un anello
di 17 subunità, è formato cioè dallo stessonumero di subunità che si trovano in ungiro dell'elica virale. In condizioni qualiquelle previste per l'interno di una cellulaospite, l'80 per cento circa della proteinadi rivestimento è incorporata nei dischi,mentre il restante 20 per cento è sottoforma di proteine A. Rendendo improv-visamente più acida la soluzione (portan-dola ap H 5), i dischi si trasformano diret-tamente in un corto frammento d'elica disoli due giri, che ricorda come forma unarondella; questi frammenti si impilanoquindi con un registro non esatto e, allafine, si legano per dare origine a eliche dilunghezza indefinita, che hanno una strut-tura molto simile alla struttura della par-ticella virale, anche se non contengonoRNA virale.
',aggregato in forma di disco della pro- teina di rivestimento presenta uncerto numero di proprietà significative. Èla forma dominante nelle condizioni noteper essere ottimali per il riassemblaggiodel virus in provetta e plausibili per l'as-semblaggio naturale nella cellula ospite.Inoltre, dimensione e struttura suggeri-
scono che questo disco potrebbe esserel'ideale maschera di montaggio per l'av-vio dell'assemblaggio. Basandoci sull'ipo-tesi che esso possa servire da centro dinucleazione, ne abbiamo osservato l'ef-fetto su tale reazione. I risultati sono statipiù che evidenti: particelle virali comple-te si formavano nello spazio di dieci minu-ti piuttosto che nelle sei ore necessarienegli esperimenti di ricostituzione, in cuila proteina si trovava in forma disaggrega-ta. Abbiamo pensato allora che, se i dischierano davvero necessari per l'iniziazione,una buona parte di questo tempo dovevatrascorrere nell'attesa che le subunitàseparate si riunissero spontaneamente inessi prima che cominciasse la crescita del-le particelle virali.
L'idea che i dischi fossero coinvolti nelprocesso biologico di iniziazione che av-viene in natura è stata rafforzata da espe-rimenti in cui l'assemblaggio veniva con-dotto con RNA di diverse fonti. Abbiamotrovato che essi interagivano molto piùprontamente con l'RNA del mosaico deltabacco che non con RNA estranei o sin-tetici, garantendo così che solo l'RNAvirale viene utilizzato per essere rivestitocon la proteina virale. La struttura deldisco consente a un giro completo diRNA di legarsi a esso nella prima fase,fornendo così chiaramente una discrimi-nazione molto più forte di quella di trenucleotidi che si legano a una singola sub-unità proteica.
La specificità esige anche che un'unicasequenza di nucleotidi dell'RNA viraleinteragisca energicamente con il discoproteico e questa sequenza deve essere unsegmento significativo di RNA, in mododa rendere ragione dell'elevata selettivitàosservata: in un solo giro del primo discoall'incirca 50 nucleotidi possono interagi-re con le 17 subunità proteiche. Pertantoci siamo accinti a isolare proprio la regio-ne di attacco dell'iniziatore dell'RNA,somministrandogli sufficiente proteina dirivestimento da permettere l'iniziazione,ma non la crescita e digerendo quindi leestremità non rivestite con un enzima.Con il nostro collega David Zimmernsiamo riusciti a isolare una serie di frani-menti di RNA, tutti contenenti una se-quenza comune al centro, ma con lun -ghezze variabili a ogni estremità. Il piùcorto di questi frammenti aveva una lun-ghezza di circa 65 nucleotidi, appena su-periore alla lunghezza necessaria per le-garsi a un giro completo del disco; abbia-mo trovato che esso si legava strettamen-te e specificatamente a esso, il che ci hapermesso di concludere che questo breveframmento contiene tutta l'informazionenecessaria per specificare la normale rea-zione di iniziazione.
La grossa dimensione e la resa relati-vamente bassa della regione di iniziazionedell'RNA del mosaico del tabacco hannoreso tecnicamente difficile la determina-zione della sua sequenza nucleotidica.Mentre stavamo lavorando appunto sul-l'isolamento e sulla determinazione dellasequenza di questa regione, Léon Hirth ecollaboratori dell'Università di Strasbur-go avevano cominciato a determinare la
sequenza nucleotidica di vari frammentidi RNA del virus del mosaico del tabacco,che era stato parzialmente digerito da unenzima. In questo modo hanno potutoottenere rese relativamente buone diquesti frammenti più corti e, in essi, ladeterminazione delle sequenze nucleoti-diche non è stata troppo difficile. Per casouno dei loro frammenti si sovrapponevaalla regione di iniziazione e dai nostri ri-sultati congiunti è stato possibile identifi-care e completare la sequenza.
La sequenza della regione di iniziazio-ne suggerisce per essa una struttura a for-cina: cioè un gambo consistente di unadoppia elica con nucleotidi debolmenteappaiati e, alla sua estremità, un'ansa co-
stituita da nucleotidi non appaiati. L'ansae la parte adiacente di gambo consistonoin una serie insolita di basi nucleotidiche,con un motivo a tripletta che si ripete eche è costituito da guanina (G), adenina(A) e uracile (U), con la guanina che siritrova ogni tre posizioni: AGAAGAA-GUUGUUGAUGA. Essendovi, per su-bunità próteica, tre siti di legame peri nucleotidi, abbiamo avanzato l'ipotesiche, durante il processo di iniziazione,un tipo di tripletta come quella appenariportata potesse condurre al riconosci-mento, da parte del disco proteico, del-l'ansa di RNA esposta.
Con l'identificazione della regione diiniziazione e la conoscenza della sequen-
za dei suoi nucleotidi, è stato possibilelocalizzare questa regione lungo il fila-mento dell'RNA. Ancora una volta l'at-tesa più ovvia - che la sequenza di inizia-zione si trovasse vicino a un'estremità del-l'RNA - è risultata errata. Zimmern e T.Michael A. Wilson hanno mostrato cheessa si trova a circa un sesto della lun-ghezza totale dell'RNA, per cui più di5000 nucleotidi devono essere ricopertiin una direzione, lungo la doppia elicadell'RNA, e circa mille devono esserericoperti nell'altra direzione.
parallelamente agli studi sulla sequenza
dell'RNA, abbiamo indagato insie-me ai nostri collaboratori sulla struttura
26
27

CATENA POLIPEPTIDICA 90 ANGSTROM SUBUNITÀ PROTEICA
ASSE DEL DISCO
Questa sezione trasversale di un disco è stata ricostruita in base airisultati di un'analisi di diffrazione dei raggi X con una risoluzione di 2,8angstrom. I nastri ripiegati indicano le catene polipetidiche che costitui-scono le subunità proteiche. La struttura a doppio strato del disco èevidente: le subunità dei due anelli impilati si toccano in corrisponden-
za di una piccola area vicino al bordo esterno del disco, mentre si al-lontanano ,erso il centro, con un paio di mascelle. Le linee tratteggia-te indicano la porzione flessibile delle catene proteiche, che si estendeverso l'interno dal sito di legame per l'RNA. Tale porzione della cate-na è in costante movimento, perciò non si può risolverne la struttura.
RO CENTRAL
40 ANGSTROM
Le particelle virali durante la crescita hanno a un'estremità due «code»di RNA, come risulta da questa microfotografia elettronica, scattata daGeneviève Lebeurier dell'Università di Strasburgo. Altri segni indica-
no che la coda più corta fuoriesce direttamente dalla particella, mentrela più lunga si ripiega all'interno nella sua cavità centrale. Il virus siallunga essenzialmente all'estremità opposta rispetto alle due code.
del disco proteico, servendoci delle tecni-che di diffrazione dei raggi X. (Nel frat-tempo, Kenneth C. Holmes del MaxPlanck Institut fui- Medizinische For-schung di Heidelberg ha proseguito gli
La struttura «a forcina» della regione dovesi attacca l'iniziatore dell'RNA virale è statadedotta da David Zimmern a partire dal piùprobabile appaiamento di basi nucleotidichenel filamento dell'RNA. Questo ordinamentodà luogo a un gambo a doppia elica con legamideboli, che presenta alla sommità un'ansa conuna speciale sequenza di basi. Si ritiene chequest'ansa si leghi al primo disco proteico percominciare l'assemblaggio del virus. Ha infattiun motivo ben distinto di tre basi nucleotidi-che, con la guanina (G) al centro e l'adenina(A) o l'uracile (U) nelle posizioni esterne.
studi sui virus con i raggi X, avviati da J.D. Bernal fin dal 1936 presso l'Universitàdi Cambridge.) I dischi formano cristallitridimensionali, per cui l'analisi con i rag-gi X è simile alla comune cristallografiaper le proteine, tranne che per la dimen-sione estremamente elevata dell'unitàripetitiva. Il disco, che consiste di 34 su -bunità proteiche e di circa 50 nucleotidi,con un peso molecolare globale di circa600 000 dalton, è stato la prima strutturadi dimensioni tanto grandi a essere esa-minata nei particolari mediante analisicon i raggi X, e ci sono voluti circa dodicianni perché il nostro gruppo di Cambrid-ge (a cui si sono aggiunti di recente AnnC. Bloomer, Gerard Bricogne e John N.Champness) riuscisse a compiere l'inda-gine con un elevato grado di risoluzione. Igrossissimi problemi tecnici sono stati ri-solti solo dopo che, nel nostro laborato-rio, da parte di altri ricercatori è statamessa a punto una speciale apparecchia-tura per affrontare strutture di questedimensioni.
Il disco ha una simmetria di rotazionecon n=17, il che ha dato luogo a informa-zioni ridondanti nei dati utilizzabili del-l'analisi con i raggi X. Tale struttura è ogginota con una risoluzione superiore alle treunità angstrom. (Un angstrom è pari a10- 7 mm, il che corrisponde all'incirca aldiametro di un atomo di idrogeno.) L'a-nalisi con i raggi X dà una mappa delladensità degli elettroni nel disco proteico:noi l'abbiamo interpretata per ottenereun modello atomico dettagliato della pro-teina di rivestimento. Non sono state an-cora dedotte le singole interazioni tra gliamminoacidi nella proteina e i nucleotidinell'RNA virale. Tuttavia, anche allo sta-dio di risoluzione di 5 angstrom, che ab-biamo raggiunto alcuni anni fa, la struttu-ra ha avuto importanti conseguenze per lanostra comprensione del processo di as-semblaggio dei virus.
Nel disco proteico, le subunità dell'a-nello superiore sono praticamente piatte,mentre quelle dell'anello inferiore sonoinclinate verso il basso al centro, per cui idue anelli si toccano soltanto in corri-spondenza della porzione più esterna deldisco e si aprono al centro come un paio dimascelle. Questa geometria è ulterior-mente accentuata dalla flessibilità dellacatena degli amminoacidi nella regionedella proteina che si trova verso l'internorispetto al sito di legame per l'RNA; inquesta parte, la catena ha un aspetto di-sordinato e non risulta ripiegata in manie-ra compatta a costituire una strutturaregolare. Quando il disco si trasforma nel-l'elica virale, le subunità di ambedue glianelli si inclinano verso l'alto al centro,fino a trovarsi tutte con la stessa angolatu-ra e con i giri adiacenti paralleli e in stret-to contatto. Le regioni più interne dellesubunità si addensano in maniera com-patta, assumendo una struttura ben defi-nita e racchiudendo e proteggendo com-pletamente l'RNA virale. Pertanto, latransizione strutturale fa chiudere le ma-scelle del disco proteico, e l'RNA rimaneintrappolato, in questo modo, all'internodella formazione.
Aquesto stadio, gli sforzi per deter-minare la sequenza nucleotidica
nell'RNA virale e quelli per delucidare lastruttura del disco proteico mediante ana-lisi con i raggi X sono proceduti insieme.Abbiamo preso in considerazione la strut-tura del disco e il modo in cui la regione diiniziazione dell'RNA virale, a forma diforcina, poteva interagire con essa perdare inizio alla sua conversione nella for-ma a rondella elicoidale. Con Zimmernabbiamo avanzato, per l'iniziazione, un'i-potesi in cui la forcina dell'RNA si inseri-sce, attraverso il foro centrale del disco,tra le mascelle delimitate dagli anelli dellesubunità. (Le dimensioni del disco e dellaforcina dell'RNA sono quelle giuste perquesto processo.) A questo punto, i nu -cleotidi situati nel gambo a doppia elica sispaiano e il gambo si apre a mano a manoche altro RNA si lega entro le mascelledel disco. Un qualche aspetto ancoraignoto di quest'interazione fa sì che il di-sco si trasformi nella forma a rondella,intrappolando l'RNA entro gli anelli disubunità proteiche allorché le mascelle sichiudono.
L'ipotesi da noi avanzata circa il mec-canismo di iniziazione ci ha portato a unaprevisione che ha potuto essere saggiatasperimentalmente. Se un'ansa di RNAviene inserita al centro del disco proteicoche agisce da iniziatore, ambedue le«code» dell'RNA, cioè le estremità chesporgono, dovrebbero essere associatecon l'estremità della particella virale abastoncino, da cui ha avuto inizio la cre-scita. Ci attendevamo, inoltre, che la codapiù corta sarebbe emersa subito, mentrela più lunga (lungo la quale avviene lamaggior parte dell'allungamento) si sa-rebbe ripiegata all'indietro verso la cavitàcentrale del bastoncino virale. Questaprevisione è stata confermata. Il gruppodi Hirth, a Strasburgo, ha ottenuto alcunemicrofotografie elettroniche di particellevirali in cui si possono vedere le dueestremità dell'RNA sporgere da un'e-stremità di molti bastoncelli (si veda lafigura in basso nella pagina a fronte).
A Cambridge abbiamo usato la micro-scopia elettronica ad alta risoluzione: conessa è stato possibile distinguere le duecode della particella virale per conferma-re che quella più lunga dell'RNA in realtàsi ripiega su se stessa tornando indietrolungo il bastoncino che cresce. Abbiamopreparato, inoltre, particelle virali in cuil'estremità più lunga dell'RNA era libera.E questo rendendo alcalino il pH dellasoluzione e quindi strappando la proteinada una parte delle particelle riunite in-sieme: tale estremità allora si estendevadirettamente fuori dalla particella piut-tosto che ripiegarsi ritornando nella cavi-tà centrale. I bastoncelli in parte denuda-ti crescevano con una velocità inferioreal 10 per cento di quella dei bastoncelliparzialmente montati, dimostrando in talmodo quanto sia importante la geome-tria dell'ansa.
Il nostro modello dell'iniziazione haun'importante conseguenza: la specialeconfigurazione generata dall'inserzionedell'ansa dell'RNA nel foro centrale del
2829
a C
3' 3'
3' 5'
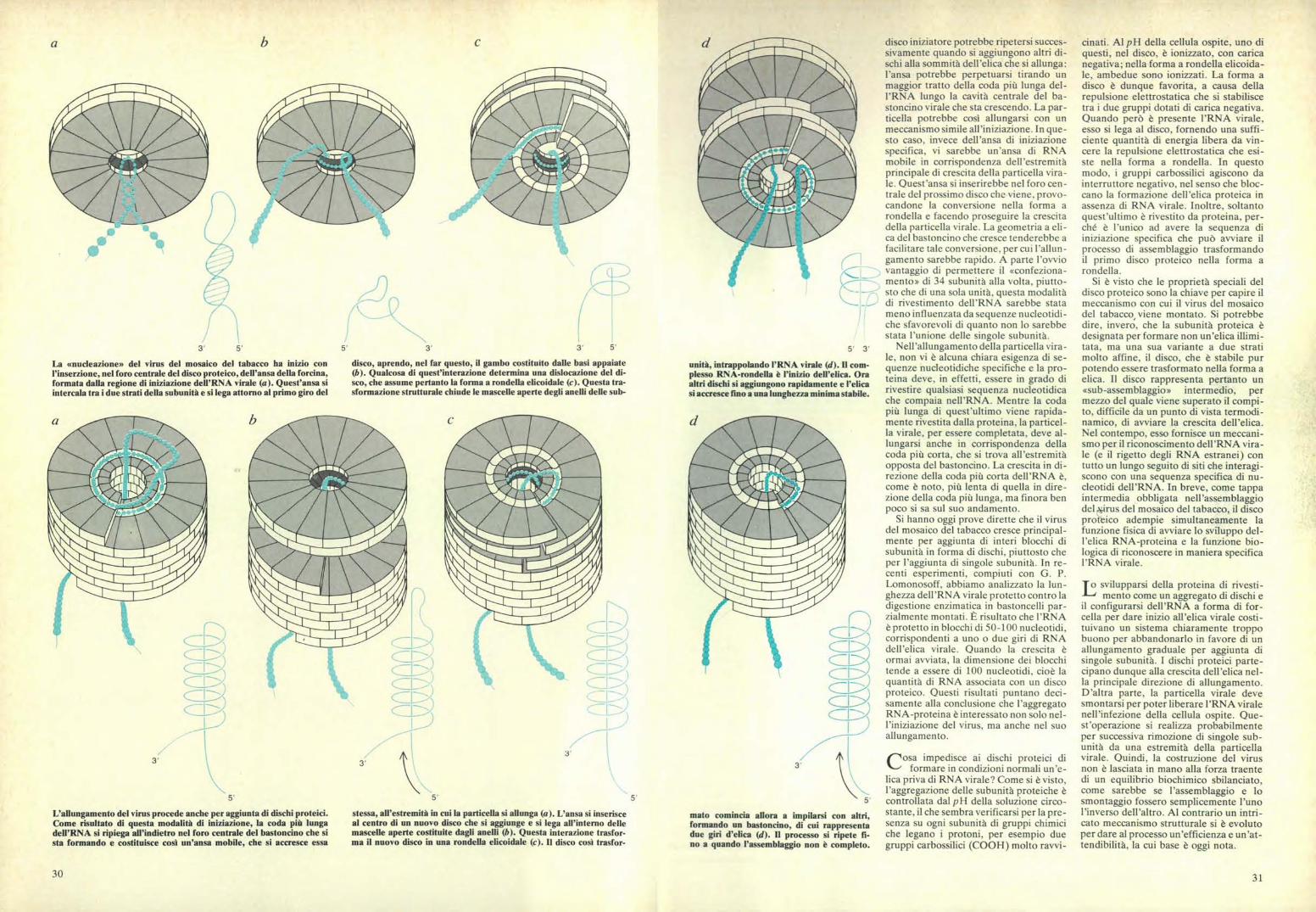
La «nucleazione» del virus del mosaico del tabacco ha inizio conl'inserzione, nel foro centrale del disco proteico, dell'ansa della forcina,formata dalla regione di iniziazione dell'RNA virale (a). Quest'ansa siintercala tra i due strati della subunità e si lega attorno al primo giro del
5' 3' 3' 5'
disco, aprendo, nel far questo, il gambo costituito dalle basi appaiate(b). Qualcosa di quest'interazione determina una dislocazione del di-sco, che assume pertanto la forma a rondella elicoidale (c). Questa tra-sformazione strutturale chiude le mascelle aperte degli anelli delle sub-
5'
L'allungamento del virus procede anche per aggiunta di dischi proteici.Come risultato di questa modalità di iniziazione, la coda più lungadell'RNA si ripiega all'indietro nel foro centrale del bastoncino che sista formando e costituisce così un'ansa mobile, che si accresce essa
5'
stessa, all'estremità in cui la particella si allunga (a). L'ansa si inserisceal centro di un nuovo disco che si aggiunge e si lega all'interno dellemascelle aperte costituite dagli anelli (b). Questa interazione trasfor-ma il nuovo disco in una rondella elicoidale (c). Il disco così trasfor-
5' 3'
unità, intrappolando l'RNA virale (d). Il com-plesso RNA-rondella è l'inizio dell'elica. Oraaltri dischi si aggiungono rapidamente e l'elicasi accresce fino a una lunghezza minima stabile.
3'
5'
mato comincia allora a impilarsi con altri,formando un bastoncino, di cui rappresentadue giri d'elica (d). Il processo si ripete fi-no a quando l'assemblaggio non è completo.
disco iniziatore potrebbe ripetersi succes-sivamente quando si aggiungono altri di-schi alla sommità dell'elica che si allunga:l'ansa potrebbe perpetuarsi tirando unmaggior tratto della coda più lunga del-l'RNA lungo la cavità centrale del ba-stoncino virale che sta crescendo. La par-ticella potrebbe così allungarsi con unmeccanismo simile all'iniziazione. In que-sto caso, invece dell'ansa di iniziazionespecifica, vi sarebbe un'ansa di RNAmobile in corrispondenza dell'estremitàprincipale di crescita della particella vira-le. Quest'ansa si inserirebbe nel foro cen-trale del prossimo disco che viene, provo-candone la conversione nella forma arondella e facendo proseguire la crescitadella particella virale. La geometria a eli -ca del bastoncino che cresce tenderebbe afacilitare tale conversione, per cui l'allun-gamento sarebbe rapido. A parte l'ovviovantaggio di permettere il «confeziona-mento» di 34 subunità alla volta, piutto-sto che di una sola unità, questa modalitàdi rivestimento dell'RNA sarebbe statameno influenzata da sequenze nucleotidi-che sfavorevoli di quanto non lo sarebbestata l'unione delle singole subunità.
Nell'allungamento della particella vira-le, non vi è alcuna chiara esigenza di se-quenze nucleotidiche specifiche e la pro-teina deve, in effetti, essere in grado dirivestire qualsiasi sequenza nucleotidicache compaia nell'RNA. Mentre la codapiù lunga di quest'ultimo viene rapida-mente rivestita dalla proteina, la particel-la virale, per essere completata, deve al-lungarsi anche in corrispondenza dellacoda più corta, che si trova all'estremitàopposta del bastoncino. La crescita in di-rezione della coda più corta dell'RNA è,come è noto, più lenta di quella in dire-zione della coda più lunga, ma finora benpoco si sa sul suo andamento.
Si hanno oggi prove dirette che il virusdel mosaico del tabacco cresce principal-mente per aggiunta di interi blocchi disubunità in forma di dischi, piuttosto cheper l'aggiunta di singole subunità. In re-centi esperimenti, compiuti con G. P.Lomonosoff, abbiamo analizzato la lun-ghezza dell'RNA virale protetto contro ladigestione enzimatica in bastoncelli par-zialmente montati. È risultato che l'RNAè protetto in blocchi di 50-100 nucleotidi,corrispondenti a uno o due giri di RNAdell'elica virale. Quando la crescita èormai avviata, la dimensione dei blocchitende a essere di 100 nucleotidi, cioè laquantità di RNA associata con un discoproteico. Questi risultati puntano deci-samente alla conclusione che l'aggregatoRNA-proteina è interessato non solo nel-l'iniziazione del virus, ma anche nel suoallungamento.
Cosa impedisce ai dischi proteici di
formare in condizioni normali un'e-lica priva di RNA virale? Come si è visto,l'aggregazione delle subunità proteiche ècontrollata dal pH della soluzione circo-stante, il che sembra verificarsi per la pre-senza su ogni subunità di gruppi chimiciche legano i protoni, per esempio duegruppi carbossilici (COOH) molto ravvi-
cinati. Al pH della cellula ospite, uno diquesti, nel disco, è ionizzato, con caricanegativa; nella forma a rondella elicoida-le, ambedue sono ionizzati. La forma adisco è dunque favorita, a causa dellarepulsione elettrostatica che si stabiliscetra i due gruppi dotati di carica negativa.Quando però è presente l'RNA virale,esso si lega al disco, fornendo una suffi-ciente quantità di energia libera da vin-cere la repulsione elettrostatica che esi-ste nella forma a rondella. In questomodo, i gruppi carbossilici agiscono dainterruttore negativo, nel senso che bloc-cano la formazione dell'elica proteica inassenza di RNA virale. Inoltre, soltantoquest'ultimo è rivestito da proteina, per-ché è l'unico ad avere la sequenza diiniziazione specifica che può avviare ilprocesso di assemblaggio trasformandoil primo disco proteico nella forma arondella.
Si è visto che le proprietà speciali deldisco proteico sono la chiave per capire ilmeccanismo con cui il virus del mosaicodel tabacco, viene montato. Si potrebbedire, invero, che la subunità proteica èdesignata per formare non un'elica illimi-tata, ma una sua variante a due stratimolto affine, il disco, che è stabile purpotendo essere trasformato nella forma aelica. Il disco rappresenta pertanto un«sub-assemblaggio» intermedio, permezzo del quale viene superato il compi-to, difficile da un punto di vista termodi-namico, di avviare la crescita dell'elica.Nel contempo, esso fornisce un meccani-smo per il riconoscimento dell'RNA vira-le (e il rigetto degli RNA estranei) contutto un lungo seguito di siti che interagi-scono con una sequenza specifica di nu-cleotidi dell'RNA. In breve, come tappaintermedia obbligata nell'assemblaggiodel akirus del mosaico del tabacco, il discoproitico adempie simultaneamente lafunzione fisica di avviare lo sviluppo del-l'elica RNA-proteina e la funzione bio-logica di riconoscere in maniera specifical'RNA virale.
Io svilupparsi della proteina di rivesti-,
mento come un aggregato di dischi eil configurarsi dell'RNA a forma di for-cella per dare inizio all'elica virale costi-tuivano un sistema chiaramente troppobuono per abbandonarlo in favore di unallungamento graduale per aggiunta disingole subunità. I dischi proteici parte-cipano dunque alla crescita dell'elica nel-la principale direzione di allungamento.D'altra parte, la particella virale devesmontarsi per poter liberare l'RNA viralenell'infezione della cellula ospite. Que-st'operazione si realizza probabilmenteper successiva rimozione di singole sub-unità da una estremità della particellavirale. Quindi, la costruzione del virusnon è lasciata in mano alla forza traentedi un equilibrio biochimico sbilanciato,come sarebbe se l'assemblaggio e losmontaggio fossero semplicemente l'unol'inverso dell'altro. Al contrario un intri-cato meccanismo strutturale si è evolutoper dare al processo un'efficienza e un'at-tendibilità, la cui base è oggi nota.
30 31

















![PARAMYXOVIRUS. Virus parainfluenzali [PIV types 1,2,3,4] Virus della Parotite Morbillivirus: Virus del morbillo Pneumovirus: Virus respiratorio.](https://static.fdocumenti.com/doc/165x107/5542eb76497959361e8e03ed/paramyxovirus-virus-parainfluenzali-piv-types-1234-virus-della-parotite-morbillivirus-virus-del-morbillo-pneumovirus-virus-respiratorio.jpg)