Albini-Crespi-Di SeriCepa 2003 1 Gli effetti delle crisi sui rapporti di lavoro.

Il gatto siamese possiede una variante del gene che provocal'albinismo, sensibile alla temperatura: l'animale quindi è tuttodi colore pallido tranne nelle estremità, più fredde e perciò
nere. Tutti i siamesi presentano anomalie delle vie ottiche. Gliocchi azzurri nei gatti e nelle tigri sono insoliti; gli animalialbini sono privi di pigmento oculare e hanno occhi rosati.
Come vedono gli albini
Le anomalie genetiche dei gatti siamesi, delle tigri e di altrimammiferi albini sono il campo di studio ideale per comprenderecome nel cervello si formi unimmagin.ie ordinata del mondo esterno
di R. W. Guillery
fibre nervose, che di solito restanodalla stessa parte del cervello, si in-crociano e vanno al nucleo genicolatoopposto; ma non risultava chiaro co-me i campi visivi venissero rappresen-tati all'interno del nucleo anormale.Allo scopo di comprendere l'organiz-zazione di tutta la gamma di stimoliche il cervello riceve dalle vie otticheanormali, è necessario determinare sele fibre vengano semplicemente devia-te al loro incrocio nel chiasma otticoo se questo avvenga in maniera piùcomplessa, in modo che l'immagine or-
dinata che si forma sulla retina risultiscomposta nel passaggio all'interno delnucleo genicolato laterale.
Jon H. Kaas e io abbiamo studiatoquesto problema con due metodi. Ilprimo è stato quello di praticare le-sioni piuttosto lievi nella retina deigatti siamesi, distruggendo una chiaz-za di cellule gangliari in modo da pro-vocare una degenerazione nelle fibrecorrispondenti, dalla retina al nucleogenicolato laterale. Poi abbiamo colo-rato le fibre nervose degeneranti inmodo differenziale, mettendo in evi-
denza la parte del nucleo genicolatoinnervata dalla parte danneggiata del-la retina. Questi esperimenti si sonorivelati piuttosto rozzi, in quanto èdifficile praticare lesioni molto limita-te alla retina, ma ci hanno indicatoche l'organizzazione della proiezionedalla retina al nucleo genicolato eranormale. Le lesioni nella parte supe-periore della retina producevano de-generazione nella porzioni anteriori delnucleo, mentre le lesioni nella parteinferiore della retina provocavano de-generazione nella parte posteriore del
ei gatti siamesi, nelle tigri bian-che, nei visoni argentati e neiratti albini esiste un gruppo di
anomalie genetiche per cui la pigmen-tazione ridotta si associa a un'anorma-lità congenita delle vie nervose ottiche:infatti alcune fibre dei nervi ottici sidirigono verso il settore « sbagliato » delcervello. Questa anormalità, che tal-volta è associata allo strabismo, hafornito un metodo nuovo per lo studiodelle vie ottiche, su cui si sono com-piute recentemente indagini da due di-versi punti di vista. Uno di questi pun-ti è stato quello di determinare comeil cervello analizzi i segnali visivi chericeve dagli occhi. L'altro campo diindagine è stato quello dei processi disviluppo che danno origine ai collega-menti ordinati caratteristici delle vieottiche (si veda l'articolo « L'originedella specificità delle cellule nervose»di Marcus Jacobson e R. K. Hunt in«Le Scienze », n. 57, maggio 1973).
L'anormalità delle vie nervose otti-che è stata studiata a fondo nei gattisiamesi. Descriverò prima di tutto lemodificazioni delle vie ottiche di que-sti animali e il modo con cui il cer-vello elabora la recezione anormale.Poi mi occuperò di alcuni problemigenerali di sviluppo che si presentanoquando si considera come un singologene identificabile possa produrre unaserie di collegamenti anomali fra laretina e il cervello.
Alcuni anni fa, durante uno studiosulle vie nervose ottiche dei gatti
compiuto presso la Scuola Medica del-l'Università del Wisconsin, trovai uncervello il cui nucleo genicolato late-rale, ossia il principale gruppo di cel-lule cerebrali di collegamento tra laretina e la rorterria cerebrale, era ti-picamente anomalo. In un nucleo ge-nicolato laterale normale vi sono stra-ti di cellule nervose chiaramente dif-
ferenziati. Nel cervello anormale lecellule, formavano strati smembrati eparzialmente fusi (si veda la figura apagina 39). Questa anormalità era diparticolare interesse, poiché l'animaleda esperimento in questione era ungatto siamese e i gatti siamesi sono disolito strabici. Mi venne l'idea che laparticolare struttura degli strati geni-colati fosse in rapporto coll'anormaleallineamento degli occhi, perché è pro-prio nel nucleo genicolato laterale chele vie nervose provenienti dagli occhivengono appaiate e trasmesse alla cor-teccia e quindi un accoppiamento ar-monico nella corteccia degli stimoliprovenienti dal mondo esterno richiedeun allineamento corretto degli occhi.
In un cervello normale l'organizza-zione degli stimoli visivi viene realiz-zata grazie alla struttura stratificatadei due nuclei genicolati, ciascuno deiquali riceve uno stimolo ordinato dauna metà del campo visivo (si veda lafigura a pagina 40). La metà sinistradel campo viene trasmessa al nucleodestro, che, a sua volta, manda fibre diproiezione alla zona visiva destra dellacorteccia; la metà destra al nucleo si-nistro (e alla parte sinistra della cor-teccia). Ogni nucleo riceve le fibreprovenienti da cellule gangliari di en-trambe le retine, cosicché ciascuna me-tà del campo visivo che può essere vi-sta da entrambi gli occhi viene rappre-sentata due volte: lo stimolo prove-niente da un occhio va nel primo stra-to chiamato strato A, mentre quello pro-veniente dall'altro occhio va nello stra-to sottostante A l . Questo sarebbe il ca-so più semplice, in cui vi sono solodue strati; in realtà quasi tutti i mam-miferi possiedono più di due strati;questi formano due serie ciascuna del-le quali riceve un gruppo ordinato distimoli da un occhio. Ogni strato per-ciò riceve la mappa di una determina-ta metà del campo visivo; le mappe
nei diversi strati devono essere sem-pre corrispondenti l'una con l'altra,per cui si possono disegnare « linee diproiezione » attraverso il nucleo geni-colato, grossolanamente perpendicolariagli strati, che corrispondono ai sin-goli punti del campo visivo.
Il cervello in cui ho potuto osser-vare per la prima volta un nucleo ge-nicolato laterale con strati irregolariproveniva da un animale in cui le fi-bre di un singolo nervo ottico (ossia ilfascio di fibre collegato a un occhio)erano state tagliate. Le fibre tagliateerano degenerate e in seguito trattatecon uno speciale colorante, rivelandocosì quali parti del nucleo genicolatolaterale erano state innervate dal ner-vo ottico tagliato. In un gatto norma-le alcune delle fibre in degenerazionesi sarebbero incrociate per innervarelo strato A sulla parte del cervello op-posta al taglio, e le altre sarebbero ri-maste sul medesimo lato ma avrebbe-ro innervato lo strato A 1 (si veda lafigura a pagina 42). Nel gatto siameseinvece alcune fibre, dopo essersi incro-ciate, proseguivano per innervare partidello strato A / , che normalmente a-vrebbero ricevuto uno stimolo nervo-so non incrociato. Viceversa, sulla par-te del cervello dove il nervo ottico erastato tagliato, solo le parti degli stra-ti che avevano ricevuto uno stimolonon incrociato (e che perciò mostra-vano fibre degenerate) erano in realtàinnervate dal lato tagliato. In seguitoa ulteriori ricerche compiute nel no-stro e in altri laboratori, si è potutodimostrare che tutti i gatti siamesipossiedono questa anomalia, anchequelli che non presentano uno stra-bismo evidente: comunque sembra chei gatti siamesi strabici abbiano un nu-mero maggiore di fibre ottiche devia-te rispetto a quelli con occhi normali.
Dato il tipo di degenerazione subitadalle fibre, abbiamo capito che alcune
36
37
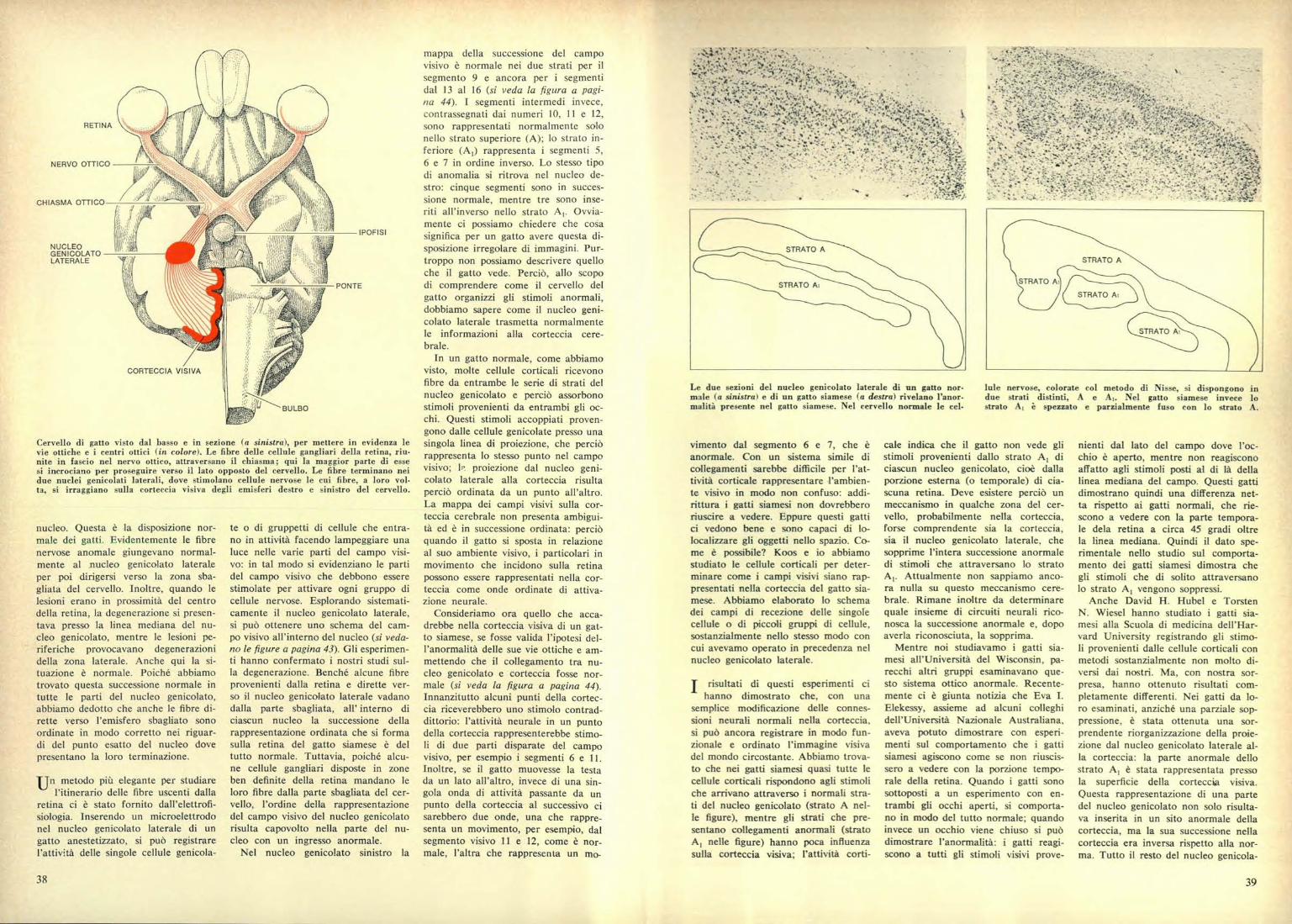
RETINA
NERVO OTTICO
CH IASMA OTTICO
NUCLEOGENI COLATOLATERALE
CORTECCIA VISIVA
Cervello di gatto visto dal basso e in sezione (a sinistra), per mettere in evidenza levie ottiche e i centri ottici (in colore). Le fibre delle cellule gangliari della retina, riu-nite in fascio nel nervo ottico, attraversano il chiasma; qui la maggior parte di essesi incrociano per proseguire verso il lato opposto del cervello. Le fibre terminano neidue nuclei genicolati laterali, dove stimolano cellule nervose le cui fibre, a loro vol-ta, si irraggiano sulla corteccia visiva degli emisferi destro e sinistro del cervello.
Le due sezioni del nucleo genicolato laterale di un gatto nor-male (a sinistra n e di un gatto siamese (a destra) rivelano l'anor-malità presente nel gatto siamese. Nel cervello normale le cel-
lule nervose, colorate col metodo di Nisse, si dispongono indue strati distinti, A e A,. Nel gatto siamese invece lostrato A, è spezzato e parzialmente fuso con lo strato A.
• • ,,. • -.)
5"" • • •:h•,;44.
• s••2 •
; • ;, 2.."Zpr••n:Nii*Iti.••'•••l'' • •• • ; • 1, •4;:e • y e
• ••• • t e • • *>-3'•"""-• ^:•-• :'
STRATO A
STRATO Ai
STRATO Al
nucleo. Questa è la disposizione nor-male dei gatti. Evidentemente le fibrenervose anomale giungevano normal-mente al nucleo genicolato lateraleper poi dirigersi verso la zona sba-gliata del cervello. Inoltre, quando lelesioni erano in prossimità del centrodella retina, la degenerazione si presen-tava presso la linea mediana del nu-cleo genicolato, mentre le lesioni pe-riferiche provocavano degenerazionidella zona laterale. Anche qui la si-tuazione è normale. Poiché abbiamotrovato questa successione normale intutte le parti del nucleo genicolato,abbiamo dedotto che anche le fibre di-rette verso l'emisfero sbagliato sonoordinate in modo corretto nei riguar-di del punto esatto del nucleo dovepresentano la loro terminazione.
Un metodo più elegante per studiarel'itinerario delle fibre uscenti dalla
retina ci è stato fornito dall'elettrofi-siologia. Inserendo un microelettrodonel nucleo genicolato laterale di ungatto anestetizzato, si può registrarel'attività delle singole cellule genicola-
te o di gruppetti di cellule che entra-no in attività facendo lampeggiare unaluce nelle varie parti del campo visi-vo: in tal modo si evidenziano le partidel campo visivo che debbono esserestimolate per attivare ogni gruppo dicellule nervose. Esplorando sistemati-camente il nucleo genicolato laterale,si può ottenere uno schema del cam-po visivo all'interno del nucleo (si veda-no le figure a pagina 43). Gli esperimen-ti hanno confermato i nostri studi sul-la degenerazione. Benché alcune fibreprovenienti dalla retina e dirette ver-so il nucleo genicolato laterale vadanodalla parte sbagliata, all' interno diciascun nucleo la successione dellarappresentazione ordinata che si formasulla retina del gatto siamese è deltutto normale. Tuttavia, poiché alcu-ne cellule gangliari disposte in zoneben definite della retina mandano leloro fibre dalla parte sbagliata del cer-vello, l'ordine della rappresentazionedel campo visivo del nucleo genicolatorisulta capovolto nella parte del nu-cleo con un ingresso anormale.
Nel nucleo genicolato sinistro la
mappa della successione del campovisivo è normale nei due strati per ilsegmento 9 e ancora per i segmentidal 13 al 16 (si veda la figura a pagi-na 44). I segmenti intermedi invece,contrassegnati dai numeri 10, 11 e 12,sono rappresentati normalmente solonello strato superiore (A); lo strato in-feriore (A,) rappresenta i segmenti 5,6 e 7 in ordine inverso. Lo stesso tipodi anomalia si ritrova nel nucleo de-stro: cinque segmenti sono in succes-sione normale, mentre tre sono inse-riti all'inverso nello strato A l . Ovvia-mente ci possiamo chiedere che cosasignifica per un gatto avere questa di-sposizione irregolare di immagini. Pur-troppo non possiamo descrivere quelloche il gatto vede. Perciò, allo scopodi comprendere come il cervello delgatto organizzi gli stimoli anormali,dobbiamo sapere come il nucleo geni-colato laterale trasmetta normalmentele informazioni alla corteccia cere-brale.
In un gatto normale, come abbiamovisto, molte cellule corticali ricevonofibre da entrambe le serie di strati delnucleo genicolato e perciò assorbonostimoli provenienti da entrambi gli oc-chi. Questi stimoli accoppiati proven-gono dalle cellule genicolate presso unasingola linea di proiezione, che perciòrappresenta lo stesso punto nel campovisivo; b‘ proiezione dal nucleo geni-colato laterale alla corteccia risultaperciò ordinata da un punto all'altro.La mappa dei campi visivi sulla cor-teccia cerebrale non presenta ambigui-tà ed è in successione ordinata: perciòquando il gatto si sposta in relazioneal suo ambiente visivo, i particolari inmovimento che incidono sulla retinapossono essere rappresentati nella cor-teccia come onde ordinate di attiva-zione neurale.
Consideriamo ora quello che acca-drebbe nella corteccia visiva di un gat-to siamese, se fosse valida l'ipotesi del-l'anormalità delle sue vie ottiche e am-mettendo che il collegamento tra nu-cleo genicolato e corteccia fosse nor-male (si veda la figura a pagina 44).Innanzitutto alcuni punti della cortec-cia riceverebbero uno stimolo contrad-dittorio: l'attività neurale in un puntodella corteccia rappresenterebbe stimo-li di due parti disparate del campovisivo, per esempio i segmenti 6 e 11.Inoltre, se il gatto muovesse la testada un lato all'altro, invece di una sin-gola onda di attività passante da unpunto della corteccia al successivo cisarebbero due onde, una che rappre-senta un movimento, per esempio, dalsegmento visivo 11 e 12, come è nor-male, l'altra che rappresenta un mo-
vimento dal segmento 6 e 7, che èanormale. Con un sistema simile dicollegamenti sarebbe difficile per l'at-tività corticale rappresentare l'ambien-te visivo in modo non confuso: addi-rittura i gatti siamesi non dovrebberoriuscire a vedere. Eppure questi gattici vedono bene e sono capaci di lo-localizzare gli oggetti nello spazio. Co-me è possibile? Koos e io abbiamostudiato le cellule corticali per deter-minare come i campi visivi siano rap-presentati nella corteccia del gatto sia-mese. Abbiamo elaborato lo schemadei campi di recezione delle singolecellule o di piccoli gruppi di cellule,sostanzialmente nello stesso modo concui avevamo operato in precedenza nelnucleo genicolato laterale.
Irisultati di questi esperimenti ci
hanno dimostrato che, con unasemplice modificazione delle connes-sioni neurali normali nella corteccia,si può ancora registrare in modo fun-zionale e ordinato l'immagine visivadel mondo circostante. Abbiamo trova-to che nei gatti siamesi quasi tutte lecellule corticali rispondono agli stimoliche arrivano attraverso i normali stra-ti del nucleo genicolato (strato A nel-le figure), mentre gli strati che pre-sentano collegamenti anormali (stratoA, nelle figure) hanno poca influenzasulla corteccia visiva; l'attività corti-
cale indica che il gatto non vede glistimoli provenienti dallo strato A, diciascun nucleo genicolato, cioè dallaporzione esterna (o temporale) di cia-scuna retina. Deve esistere perciò unmeccanismo in qualche zona del cer-vello, probabilmente nella corteccia,forse comprendente sia la corteccia,sia il nucleo genicolato laterale, chesopprime l'intera successione anormaledi stimoli che attraversano lo stratoA,. Attualmente non sappiamo anco-ra nulla su questo meccanismo cere-brale. Rimane inoltre da determinarequale insieme di circuiti neurali rico-nosca la successione anormale e, dopoaverla riconosciuta, la sopprima.
Mentre noi studiavamo i gatti sia-mesi all'Università del Wisconsin, pa-recchi altri gruppi esaminavano que-sto sistema ottico anormale. Recente-mente ci è giunta notizia che Eva I.Elekessy, assieme ad alcuni colleghidell'Università Nazionale Australiana,aveva potuto dimostrare con esperi-menti sul comportamento che i gattisiamesi agiscono come se non riuscis-sero a vedere con la porzione tempo-rale della retina. Quando i gatti sonosottoposti a un esperimento con en-trambi gli occhi aperti, si comporta-no in modo del tutto normale; quandoinvece un occhio viene chiuso si puòdimostrare l'anormalità: i gatti reagi-scono a tutti gli stimoli visivi prove-
nienti dal lato del campo dove l'oc-chio è aperto, mentre non reagisconoaffatto agli stimoli posti al di là dellalinea mediana del campo. Questi gattidimostrano quindi una differenza net-ta rispetto ai gatti normali, che rie-scono a vedere con la parte tempora-le dela retina a circa 45 gradi oltrela linea mediana. Quindi il dato spe-rimentale nello studio sul comporta-mento dei gatti siamesi dimostra chegli stimoli che di solito attraversanolo strato A, vengono soppressi.
Anche David H. Hubel e TorstenN. Wiesel hanno studiato i gatti sia-mesi alla Scuola di medicina dell'Har-vard University registrando gli stimo-li provenienti dalle cellule corticali conmetodi sostanzialmente non molto di-versi dai nostri. Ma, con nostra sor-presa, hanno ottenuto risultati com-pletamente differenti. Nei gatti da lo-ro esaminati, anziché una parziale sop-pressione, è stata ottenuta una sor-prendente riorganizzazione della proie-zione dal nucleo genicolato laterale al-la corteccia: la parte anormale dellostrato A, è stata rappresentata pressola superficie della corteccia visiva.Questa rappresentazione di una partedel nucleo genicolato non solo risulta-va inserita in un sito anormale dellacorteccia, ma la sua successione nellacorteccia era inversa rispetto alla nor-ma. Tutto il resto del nucleo genicola-
38
39

41
to era rappresentato in una successio-ne normale, ma su questa rappresen-tazione normale si inseriva una por-zione anormale (si veda la figura apagina 45). Quindi i segmenti delcampo visivo erano rappresentati inordine (eccetto una leggera interru-
zione tra i segmenti normali e anor-mali, cioè, nella figura, tra i segmenti8 e 10). In altre parole, in questo ca-so la successione, spezzata nei nucleigenicolati, veniva riordinata nella cor-teccia. Si è portati a concludere chequesto rimescolamento, che avviene
nel tratto dal nucleo genicolato allacorteccia, si verifica allo scopo di com-pensare la proiezione anormale che av-viene tra la retina e il nucleo. Sembrainvece improbabile che la correzionesia il risultato di una pressione dovu-ta alla selezione naturale, in quanto igatti siamesi vengono allevati per labellezza del loro aspetto e non per laloro capacità visiva.
Il modello di rappresentazione cor-ticale descritto da Hubel e Wiesel eratalmente diverso dal modello da noistudiato che bisognava trovare unaspiegazione soddisfacente. le ,yri pote-vamo certo imputare questa differen-za alle diverse tecniche, poiché i me-todi seguiti dai due autori e da noierano fondamentalmente simili e l'in-terpretazione dei risultati in entrambii casi non presentava dubbi. Tuttavia,per essere del tutto sicuri dei nostririsultati, abbiamo studiato la proiezio-ne dal nucleo genicolato alla cortec-cia con un altro metodo: quello delladegenerazione retrograda. Poiché le cel-lule nervose genicolate mandano fi-bre alla corteccia visiva in modo deltutto ordinato, una piccola lesione cor-ticale danneggia un gruppo ben defi-nito di fibre nervose genicolate: le cel-lule che danno origine a queste fibremostrano una degenerazione retrogra-da. In un gatto con una proiezionenormale dal nucleo genicolato lateralealla corteccia, una lesione corticaleproduce sempre una zona di degene-razione retrograda che attraversa com-pletamente tutti gli strati del nucleoed è limitata dalle linee di proiezione.I collegamenti descritti da Hubel eWiesel dovrebbero portare a un di-verso tipo di degenerazione retrogra-da, con zone di degenerazione noncorrispondenti nei due strati (si vedala figura in alto a pagina 46).
Con questo metodo abbiamo studiato
Il gatti siamesi. I primi 10 dimo-stravano un tipo di degenerazione nor-male. Alla fine, però, l'undicesimo gat-to presentava un tipo di degenerazioneprevedibile sulla base dei risultati diHubel e Wiesel. Abbiamo perciò de-dotto che nei gatti siamesi debbonoesistere due distinti tipi di proiezionedal nucleo genicolato laterale alla cor-teccia. Il primo tipo, quello che noiabbiamo incontrato in quasi tutti i no-stri esperimenti, consiste nella soppres-sione pura e semplice del gruppo distimoli anormali al momento dell'en-trata nella corteccia. Abbiamo chiama-to questo tipo Midwestern. Il secondotipo di proiezione è più complesso,poiché ricrea una rappresentazione or-dinata dell'ambiente visivo circostante
40
La tigre bianca è un mammifero albino che possiede anomalieottiche collegate a una pigmentazione ridotta: una tigre femmi-na albina di nome Rewati si trova presso lo zoo di Washington.Le tigri bianche sono ornate da strisce bianche che sostituisco-
no le normali arancioni, e da strisce marroni. Gli occhi sonoazzurri. L'insolita colorazione e lo strabismo sono collegati aun'anormalità delle vie ottiche, scoperta nei gatti siamesi e inaltri albini e riscontrata anche nel fratello di Rewati, Moni.
CAMPO VISIVO
16
STRATO A
NUCLEOGEN ICOLATO
LATERALE
15
RETINA
NERVOOTTICO
5, 4,
3
STRATO Ai STRATO Ai
CORTECCIAVISIVA
Rappresentazione schematica del campo visivo sulla retina e nel cervello di un gattonormale. Il campo è diviso in 16 segmenti disuguali. Ogni metà del cervello « vede »la metà del campo visivo del lato opposto; una quantità relativamente maggiore ditessuto cerebrale è interessata alla rappresentazione delle parti centrali del campo ri-spetto alle parti laterali. Il cristallino proietta sulla retina un'immagine capovolta delcampo. L'occhio sinistro non vede la parte all'estrema destra del campo complessivo(segmenti 15 e 16), mentre l'occhio destro non vede i segmenti I e 2. Le rappresenta-zioni del campo visto da entrambi gli occhi (segmenti dal 3 al 14) sono localizzate indue serie negli strati del nucleo genicolato (rappresentati qui come strati A e 21 1 ), aseconda che provengano dall'occhio sullo stesso lato o dalla parte opposta; le rappre-sentazioni nei due strati sono corrispondenti, cosicché le « linee di proiezione » (li-nee tratteggiate) attraverso gli strati rappresentano singoli punti nel campo visivo.

\i ,Iliv h
.1.'4,q
, A
I
tri:::.f,(lì::;:1:::;!,\4,
o20 1030
NUCLEO GENICOLATO LATERALEe
GATTO NORMALE
RETINA
NERVO OTTICO TAGLIATO \ss.,SS 5
n
NUCLEOGENICOLATOLATERALE
STRATO A
GATTO SIAMESE
RETINA
NERVO OTTICO TAGLIATO \\•sss,
NUCLEOGENICOLATOLATERALE
STRATO A/ STRATO A
La via seguita dalle fibre tra la retina e i nuclei genicolati può essere messa in evi-denza mediante colorazione selettiva delle fibre nervose degenerate. Tagliando un nervoottico, le sue fibre degenerano. In un gatto normale (sopra) il colorante (disegno a pun-tini) dimostra che alcune fibre si incrociano per innervare lo strato A (superiore) delnucleo genicolato laterale della parte opposta del cervello rispetto al taglio; altrefibre vanno allo strato A, (inferiore) dalla medesima parte del taglio. Nel gatto sia-mese (in basso) vi sono alcune fibre in più che si incrociano per dirigersi verso lostrato A, opposto, e un numero minore di fibre che si mantengono sul medesimo lato,.
La proiezione del campo visivo sul nucleo genicolato lateraleviene determinata registrando gli impulsi nel nucleo genicolato(a destra) mentre si inviano stimoli da varie parti del campo(a sinistra). Qui sono stati registrati i risultati ottenuti con ungatto normale. Ogni linea indicata da una lettera rappresentauna inserzione di microelettrodo, col punto preciso di registra-zione indicato da un trattino orizzontale. Il campo recettivo cor-
rispondente a ciascun sito di registrazione è anch'esso numera-to; i circoletti colorati fanno parte del campo inviato allo stra-to A, mentre quelli neri vengono inviati allo strato A,. Il discoottico corrisponde al punto cieco della retina, dove le fibre ner-vose si riuniscono in un fascio per dare origine al nervo ot-tico; si noti che esiste una piccola zona di discontinuitàpriva di cellule tra le linee b e c nel nucleo genicolato.
Proiezione del campo visivo di un gatto siamese. La rappre-sentazione del campo nello strato A (in colore) è normale; isiti tra la parte esterna e la parte centrale del nucleo genico.lato laterale destro, hanno campi recettivi disposti tra la par.te estrena e la parte centrale del campo visivo sinistro. Invece
nello strato A, la rappresentazione del campo visivo è anorma-le (circoletti neri); subisce cioè un capovolgimento speculare.In questo caso le inserzioni del microelettrodo non riescono acentrare il punto in cui il disco ottico è rappresentato nel nucleogenicolato, cosicché non si nota alcuna discontinuità cellulare.
•) 3060 50 2(GRADI)
"tadhardlal.WInfrar4v
10
NUCLEO GENICOLATO LATERALE
f ed c b aDISCO OTTICO
C f e
10
A,
CAMPO VISIVO
a
CAMPO VISIVOa livello corticale: lo abbiamo battez-zato col nome di Boston. Entrambi imodelli di proiezione permettono allacorteccia di ricevere una rappresenta-zione non ambigua e ordinata dei cam-pi visivi, usando tutto il tessuto corti-cale a disposizione. La sovrapposizio-ne tra la visuale dei due occhi a li-vello corticale è tuttavia scarsa.
Non sappiamo come mai taluni cer-velli di gatto risolvano il loro proble-ma visivo in un certo modo, mentrealtri lo risolvono in un modo diverso.La soluzione dipende probabilmentedalle dimensioni del « segmento media-no normale », ossia la porzione dellostrato A 1 che non è anormale ed è
indicata dai segmenti 8 e 9 (si vedala figura a pagina 44). La frattura trala rappresentazione normale e anor-male sulla corteccia dei gatti Bostonè riferibile alle dimensioni di questosegmento mediano normale. È chiaroche il tipo Boston realizza una rap-presentazione completa del campo vi-sivo solo se il segmento mediano è as-sente o molto piccolo. Il segmento me-diano normale riceve stimoli dalle re-gioni centrali della retina e perciò èprobabilmente molto importante perl'allineamento degli occhi. I gatti incui il segmento mediano normale èmolto piccolo sono per lo più strabici;probabilmente i gatti siamesi netta-
mente strabici presentano il tipo Bo-ston, mentre i gatti con occhi norma-li tendono a presentare il tipo Mid--western.
Tuttora non sappiamo perché i gat-ti siamesi siano strabici; forse in uncervello di gatto siamese pochissimecellule ricevono Stimoli binoculari, percui gli stimoli provenienti dai due oc-chi generalmente non sono mai sovrap-posti. Questa loro anormalità ci ha datola possibilità di comprendere parecchiaspetti delle vie ottiche del gatto. Sap-piamo che le fibre ottiche possonogiungere alla regione sbagliata del cer-vello pur senza modificare le tappeprecedenti del loro percorso. Il nucleogenicolato laterale riceve queste fibrea seconda del luogo d'origine nella re-tina; viene mantenuto l'ordine topogra-fico normale delle fibre che giungonoal nucleo genicolato laterale anche sequesto mantenimento provoca una rap-presentazione spezzata del campo visivoall'interno del nucleo genicolato latera-le. Invece l'ordine topografico dellaproiezione dal nucleo genicolato allacorteccia in alcuni gatti può essere alte-rato in modo nettissimo, per cui ilriordinamento delle fibre ricrea unarappresentazione sequenziale del cam-po visivo. In alcuni gatti questo rior-dinamento non sembra possibile, perciòla rapresentazione, risultando spezzata,viene eliminata. Abbiamo potuto ap-purare che la natura dei collegamentitra la retina e il nucleo genicolatolaterale è del tutto diversa da quelladei collegamenti tra nucleo genicolatoe corteccia; abbiamo così posto pro-blemi nuovi e di difficile soluzione cir-ca i meccanismi di sviluppo delle vienervose centrali.
Il gatto siamese non è un animaleche si presti facilmente allo studio del-lo sviluppo embrionale, ma si è sco-perto che altri mammiferi presentanole stesse anormalità. D. J. Creel, del-l'Università dell'Utah, fu il primo asuggerire che le vie nervose aberranti,presenti nei gatti siamesi, fossero soloun esempio di un'anomalia molto dif-fusa, che colpisce tutti gli animali al-bini; questo ricercatore fece notareche alcuni anni fa R. D. Lund, del-l'University College di Londra, avevatrovato un numero minore di fibre ner-vose non incrociate provenienti dallaretina dei ratti albini, rispetto a quel-le dei ratti a normale pigmentazione.Creel inoltre mise in evidenza il fattoche i gatti siamesi, dal punto di vistagenetico, rappresentano una variantedell'albinismo. Gli animali che hannoereditato il gene albino da entrambi igenitori (e che perciò sono omozigotiper questo carattere) non sono affatto
90 80 70 60 50 40(GRADI)
in grado di fabbricare melanina, poi-ché il gene albino non riesce a orga-nizzare la sintesi della tirosinasi, unenzima essenziale per la formazione diquesto pigmento. Il gene himalaiano,che è una variante del gene albino eche appare nello stesso punto sul cro-mosoma, codifica un enzima che riescea sintetizzare la melanina solo a tem-perature relativamente basse perciò glianimali come i gatti siamesi, i coniglio i topi che sono omozigoti per il ge-ne himalaiano, sono in genere bianchio di colore molto chiaro, ma presen-tano estremità scure
L'ipotesi che vi fosse un'anormalità
ottica comune a tutti gli albini, dicui i gatti siamesi rappresentavano so-lo un esempio, era di particolare in-
teresse. Se si fosse potuto confermarequesta ipotesi, numerose specie avreb-bero costituito un ottimo materiale daesperimento per queste anomalie eavrebbero permesso di superare le dif-ficoltà sperimentali connesse coi gatti.Se tutti gli albini sono anormali, èprobabile vi sia un'unica base geneti-ca per questa anormalità. Sarebbe poiparticolarmente interessante esaminarequanto le anormalità visive riconosciu-te da molto tempo negli albini umanisiano da riferirsi a vie ottiche abnormi.
Per dimostrare che esiste una anor-malità generale legata all'albinismo eranecessario dimostrare che gli strati delnucleo genicolato laterale nel ratto al-bino, per esempio, erano spezzettatipiù o meno come nel gatto siamese, equindi riscontrare simili anomalie in
altre specie. Si scoprì che esistevanogià da diversi anni prove evidenti diquesta anomalia nel nucleo genicolatodei ratti albini, considerate per lo piùsolo come una curiosità. Le linee diproiezione, che rappresentano i singolipunti del campo visivo, sono state de-terminate per il ratto da Vicente M.Montero e collaboratori alla Scuola dimedicina dell'Università del Wisconsin,mediante l'uso di microelettrodi: talilinee percorrono il nucleo con anda-mento leggermente obliquo (si veda lafigura in basso a pagina 46). Studian-do i segmenti di degenerazione retro-grada prodotti nel nucleo genicolatolaterale del ratto in seguito a lesioninella corteccia visiva, Montero e ioabbiamo rilevato che le estremità diquesti segmenti definivano linee di
42
43
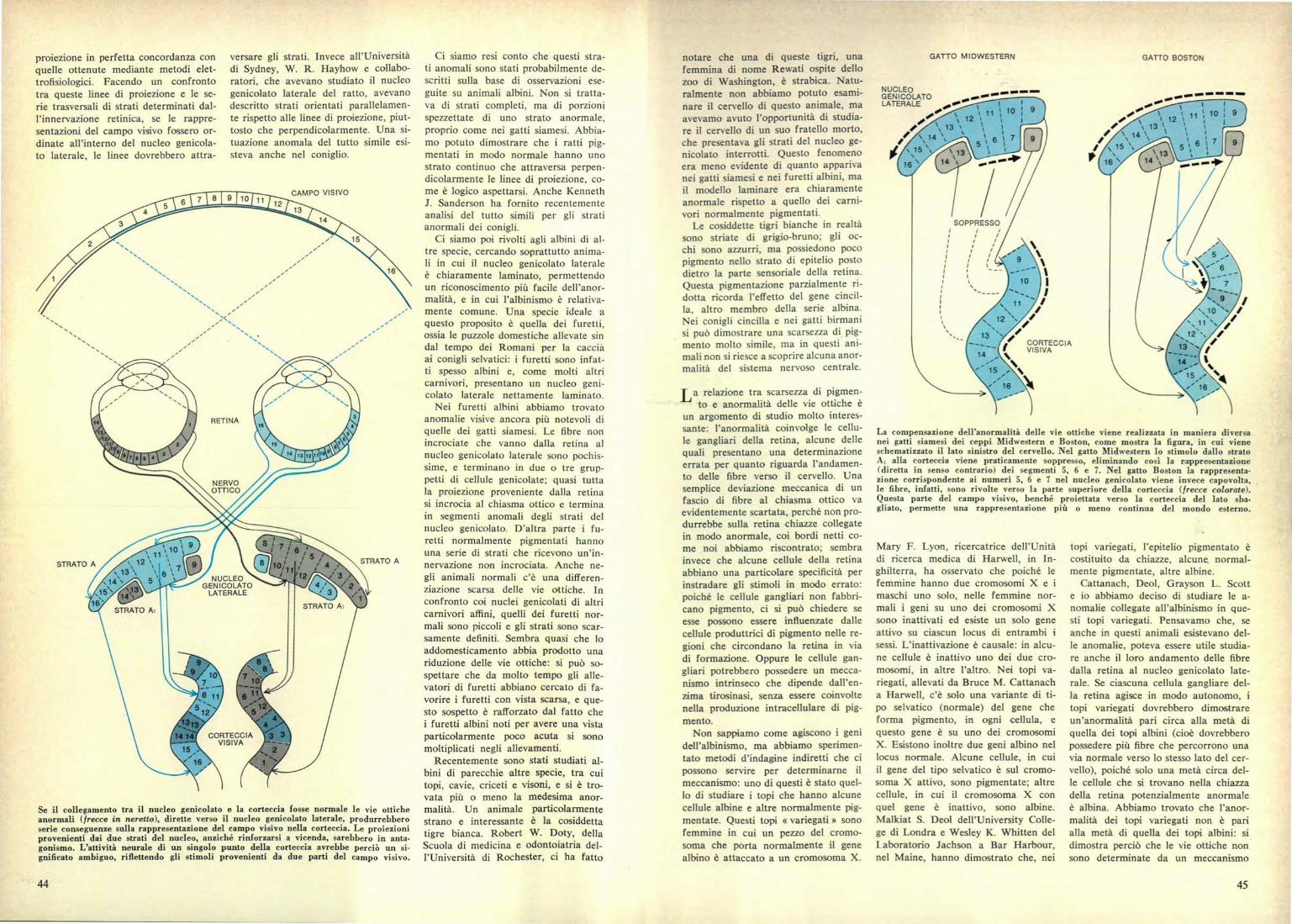
GATTO M IDWESTERN
GATTO BOSTON
La compensazione dell'anormalità delle vie ottiche viene realizzata in maniera diversanei gatti siamesi dei ceppi Midwestern e Boston, come mostra la figura, in cui vieneschematizzato il lato sinistro del cervello. Nel gatto Midwestern lo stimolo dallo stratoA, alla corteccia viene praticamente soppresso, eliminando così la rappresentazione(diretta in senso contrario) dei segmenti 5, 6 e 7. Nel gatto Boston la rappresenta-zione corrispondente ai numeri 5, 6 e 7 nel nucleo genicolato viene invece capovolta,le fibre, infatti, sono rivolte verso la parte superiore della corteccia (frecce colorate).Questa parte del campo visivo, benché proiettata verso la corteccia del lato sba-gliato, permette una rappresentazione più o meno continua del mondo esterno.
10
11
CORTECCIAI VISIVA
NUCLEOGENICOLATOLATERALE
~~~~~ n•n
SOPPRESSOi
I
CAMPO VISIVO
NUCLEOGENICOLATO
LATERALE
CORTECCIAVISIVA
Se il collegamento tra il nucleo genicolato e la corteccia fosse normale le vie otticheanormali (frecce in neretto), dirette verso il nucleo genicolato laterale, produrrebberoserie conseguenze sulla rappresentazione del campo visivo nella corteccia. Le proiezioniprovenienti dai due strati del nucleo, anziché rinforzarsi a vicenda, sarebbero in anta-gonismo. L'attività neurale di un singolo punto della corteccia avrebbe perciò un si-gnificato ambiguo, riflettendo gli stimoli provenienti da due parti del campo visivo.
proiezione in perfetta concordanza conquelle ottenute mediante metodi elet-trofisiologici. Facendo un confrontotra queste linee di proiezione e le se-rie trasversali di strati determinati dal-l'innervazione retinica, se le rappre-sentazioni del campo visivo fossero or-dinate all'interno del nucleo genicola-to laterale, le linee dovrebbero attra-
versare gli strati. Invece all'Universitàdi Sydney, W. R. Hayhow e collabo-ratori, che avevano studiato il nucleogenicolato laterale del ratto, avevanodescritto strati orientati parallelamen-te rispetto alle linee di proiezione, piut-tosto che perpendicolarmente. Una si-tuazione anomala del tutto simile esi-steva anche nel coniglio.
Ci siamo resi conto che questi stra-ti anomali sono stati probabilmente de-scritti sulla base di osservazioni ese-guite su animali albini. Non si tratta-va di strati completi, ma di porzionispezzettate di uno strato anormale,proprio come nei gatti siamesi. Abbia-mo potuto dimostrare che i ratti pig-mentati in modo normale hanno unostrato continuo che attraversa perpen-dicolarmente le linee di proiezione, co-me è logico aspettarsi. Anche KennethJ. Sanderson ha fornito recentementeanalisi del tutto simili per gli stratianormali dei conigli.
Ci siamo poi rivolti agli albini di al-tre specie, cercando soprattutto anima-li in cui il nucleo genicolato lateraleè chiaramente laminato, permettendoun riconoscimento più facile dell'anor-malità, e in cui l'albinismo è relativa-mente comune. Una specie ideale aquesto proposito è quella dei furetti,ossia le puzzole domestiche allevate sindal tempo dei Romani per la cacciaai conigli selvatici: i furetti sono infat-ti spesso albini e, come molti altricarnivori, presentano un nucleo geni-colato laterale nettamente laminato.
Nei furetti albini abbiamo trovatoanomalie visive ancora più notevoli diquelle dei gatti siamesi. Le fibre nonincrociate che vanno dalla retina alnucleo genicolato laterale sono pochis-sime, e terminano in due o tre grup-petti di cellule genicolate; quasi tuttala proiezione proveniente dalla retinasi incrocia al chiasma ottico e terminain segmenti anomali degli strati delnucleo genicolato. D'altra parte i fu-retti normalmente pigmentati hannouna serie di strati che ricevono un'in-nervazione non incrociata. Anche ne-gli animali normali c'è una differen-ziazione scarsa delle vie ottiche. Inconfronto coi nuclei genicolati di altricarnivori affini, quelli dei furetti nor-mali sono piccoli e gli strati sono scar-samente definiti. Sembra quasi che loaddomesticamento abbia prodotto unariduzione delle vie ottiche: si può so-spettare che da molto tempo gli alle-vatori di furetti abbiano cercato di fa-vorire i furetti con vista scarsa, e que-sto sospetto è rafforzato dal fatto chei furetti albini noti per avere una vistaparticolarmente poco acuta si sonomoltiplicati negli allevamenti.
Recentemente sono stati studiati al-bini di parecchie altre specie, tra cuitopi, cavie, criceti e visoni, e si è tro-vata più o meno la medesima anor-malità. Un animale particolarmentestrano e interessante è la cosiddettatigre bianca. Robert W. Doty, dellaScuola di medicina e odontoiatria del-l'Università di Rochester, ci ha fatto
notare che una di queste tigri, unafemmina di nome Rewati ospite dellozoo di Washington, è strabica. Natu-ralmente non abbiamo potuto esami-nare il cervello di questo animale, maavevamo avuto l'opportunità di studia-re il cervello di un suo fratello morto,che presentava gli strati del nucleo ge-nicolato interrotti. Questo fenomenoera meno evidente di quanto apparivanei gatti siamesi e nei furetti albini, mail modello laminare era chiaramenteanormale rispetto a quello dei carni-vori normalmente pigmentati.
Le cosiddette tigri bianche in realtàsono striate di grigio-bruno; gli oc-chi sono azzurri, ma possiedono pocopigmento nello strato di epitelio postodietro la parte sensoriale della retina.Questa pigmentazione parzialmente ri-dotta ricorda l'effetto del gene cincil-la, altro membro della serie albina.Nei conigli cincilla e nei gatti birmanisi può dimostrare una scarsezza di pig-mento molto simile, ma in questi ani-mali non si riesce a scoprire alcuna anor-malità del sistema nervoso centrale.
La relazione tra scarsezza di pigmen-to e anormalità delle vie ottiche è
un argomento di studio molto interes-sante: l'anormalità coinvolge le cellu-le gangliari della retina, alcune dellequali presentano una determinazioneerrata per quanto riguarda l'andamen-to delle fibre verso il cervello. Unasemplice deviazione meccanica di unfascio di fibre al chiasma ottico vaevidentemente scartata, perché non pro-durrebbe sulla retina chiazze collegatein modo anormale, coi bordi netti co-me noi abbiamo riscontrato; sembrainvece che alcune cellule della retinaabbiano una particolare specificità perinstradare gli stimoli in modo errato:poiché le cellule gangliari non fabbri-cano pigmento, ci si può chiedere seesse possono essere influenzate dallecellule produttrici di pigmento nelle re-gioni che circondano la retina in viadi formazione. Oppure le cellule gan-gliari potrebbero possedere un mecca-nismo intrinseco che dipende dall'en-zima tirosinasi, senza essere coinvoltenella produzione intracellulare di pig-mento.
Non sappiamo come agiscono i genidell'albinismo, ma abbiamo sperimen-tato metodi d'indagine indiretti che cipossono servire per determinarne ilmeccanismo: uno di questi è stato quel-lo di studiare i topi che hanno alcunecellule albine e altre normalmente pig-mentate. Questi topi « variegati » sonofemmine in cui un pezzo del cromo-soma che porta normalmente il genealbino è attaccato a un cromosoma X.
Mary F. Lyon, ricercatrice dell'Unitàdi ricerca medica di Harwell, in In-ghilterra, ha osservato che poiché lefemmine hanno due cromosomi X e imaschi uno solo, nelle femmine nor-mali i geni su uno dei cromosomi Xsono inattivati ed esiste un solo geneattivo su ciascun locus di entrambi isessi. L'inattivazione è causale: in alcu-ne cellule è inattivo uno dei due cro-mosomi, in altre l'altro. Nei topi va-riegati, allevati da Bruce M. Cattanacha Harwell, c'è solo una variante di ti-po selvatico (normale) del gene cheforma pigmento, in ogni cellula, equesto gene è su uno dei cromosomiX. Esistono inoltre due geni albino nellocus normale. Alcune cellule, in cuiil gene del tipo selvatico è sul cromo-soma X attivo, sono pigmentate; altrecellule, in cui il cromosoma X conquel gene è inattivo, sono albine.Malkiat S. Deol dell'University Colle-ge di Londra e Wesley K. Whitten delL aboratorio Jachson a Bar Harbour,nel Maine, hanno dimostrato che, nei
topi variegati, l'epitelio pigmentato ècostituito da chiazze, alcune normal-mente pigmentate, altre albine.
Cattanach, Deol, Grayson L. Scotte io abbiamo deciso di studiare le a-nomalie collegate all'albinismo in que-sti topi variegati. Pensavamo che, seanche in questi animali esistevano del-le anomalie, poteva essere utile studia-re anche il loro andamento delle fibredalla retina al nucleo genicolato late-rale. Se ciascuna cellula gangliare del-la retina agisce in modo autonomo, itopi variegati dovrebbero dimostrareun'anormalità pari circa alla metà diquella dei topi albini (cioè dovrebberopossedere più fibre che percorrono unavia normale verso lo stesso lato del cer-vello), poiché solo una metà circa del-le cellule che si trovano nella chiazzadella retina potenzialmente anormaleè albina. Abbiamo trovato che l'anor-malità dei topi variegati non è parialla metà di quella dei topi albini: sidimostra perciò che le vie ottiche nonsono determinate da un meccanismo
44
45

/
STRATO A
STRATO Ai
C
STRATO A
STRATO A
DEGENERAZIONERETROGRADA
16
GATTO BOSTON
DEGENERAZIONERETROGRADA
NUCLEOGENICOLATO
LATERALE MEDICINAFin dai suoi primi numeri, LE SCIENZE, edizione italiana di
SCIENTIFIC AMERICAN, ha dedicato numerosi articoli a pro-
blemi medici di particolare importanza tra cui:
IL COLERA
di N. Hirschhorn e W. Greenough III(n. 39)
Questa malattia può essere facilmentecurata con la sostituzione dei liquidi or-ganici perduti. La conoscenza del mec-canismo d'azione della tossina consenti-rebbe però un trattamento piú semplice.
INSUFFICIENZARESPIRATORIA ACUTA
di P. Winter e E. Lowenstein (n. 19)
Questa « causa mortis » deve essere con-siderata una entità clinica a se stante.Nei centri di terapia respiratoria inten-siva viene fronteggiata da équipe dimedici e tecnici altamente specializzati.
Negli ospedali provvisti di « unità co-ronariche » la mortalità per infarto puòscendere di un terzo. Una larga diffu-sione di queste nuove terapie potrebbesalvare un gran numero di vite umane.
IL PROBLEMA DELLA SCLEROSIMULTIPLA
di G. Dean (n. 26)
La causa di questa malattia del sistemanervoso centrale è sconosciuta. Le note-voli variazioni di frequenza fanno peròsupporre che essa dipenda dall'infezio-ne da parte di un virus a lungo perio-do di latenza.
FATTORI PSICOLOGICINELLO STRESS E NELLE MALATTIE
di J.M. Weiss (n. 49)
Una nuova tecnica permette di __separarenelle situazioni di stress i fattori psico-logici da quelli fisici. In studi condottisui topi i fattori psicologici si sono ri-velati la causa principale dell'ulcera ga-strica e di altri disturbi.
AVVELENAMENTODA PIOMBO
di J. Chisolm jr. (n. 33)
Delle sostanze naturali con le quali l'uo-mo viene a contatto, il piombo è sicu-ramente una delle piú diffuse. Ce neoccupiamo in questa sede per l'effettoche esso ha sui bambini che vivono invecchie abitazioni.
DIAGNOSI PRENATALEDELLE MALATTIE GENETICHE
di T. Friedmann (n. 42)
Nuove tecniche rendono possibile indi-viduare malattie ereditarie nelle fasi pre-coci della gravidanza. In quale misurail controllo di tali nascite è giustificatosul piano biologico e morale?
IL SISTEMA IMMUNITARIO
di N.K. Jerne (n. 63)
Ha il compito di salvaguardare l'identitàdel corpo. I suoi costituenti di base so-no i linfociti e gli anticorpi, molecoleche riconoscono sia le molecole estraneesia quelle appartenenti allo stesso orga-nismo.
MALATTIE E RISPOSTEIMMUNITARIE
di A.L. Notkins e H. Koprowski (n. 56)
Il meccanismo di difesa del corpo nonsempre è utile. In molti casi è lo stes-so processo che dovrebbe combattere unvirus a provocare il danno connesso conla malattia virale.
VIRUS ERPETICI E CANCROdi K.A. Rafferty jr. (n. 65)
É noto da tempo che questi virus ubi-quitari possono provocare il cancro ne-gli animali da esperimento; ora è statodimostrato che virus di questo tipo so-no implicati anche in alcuni tipi di can-cro dell'uomo.
TERAPIA INTENSIVADELL'INFARTO
di B. Lown (n. 5)
GATTO MIDWESTERN
A
CORTECCIA VISIVA
La degenerazione retrograda dei corpi cellulari nel nucleo genicolato laterale (puntini
neri) che si verifica dopo che le terminazioni delle fibre nella corteccia sono statedanneggiate (aree grige), mette in evidenza il percorso delle fibre dal nucleo genico-lato laterale alla corteccia. Queste vie differiscono per i gatti Midwestern e Boston;i collegamenti « normali » dal nucleo genicolato laterale alla corteccia nei gatti Mid-western producono zone di degenerazione che attraversano entrambi gli strati. I col-legamenti anormali (ma correttivi) verso la corteccia nei gatti Boston producono zo-ne di degenerazione non corrispondenti nei due strati del nucleo genicolato laterale.
a i n
Si può dimostrare che nei ratti albini l'anormalità del nucleo genicolato laterale èperfettamente paragonabile a quella che si riscontra nei gatti siamesi. Grazie agli studicompiuti con microelettrodi e con il metodo della degenerazione retrograda si sonopotute tracciare linee di proiezione (linee tratteggiate) nel ratto (a) che non sembranoattraversare strati diversi; al posto degli strati si notano isole (in grigio) orientate pa-rallelamente alle linee anziché perpendicolarmente. I ratti dell'esperimento sono al.bini: se però consideriamo le isole segmenti interrotti anormali, la situazione è lamedesima di quella dei gatti siamesi (b). I ratti normalmente pigmentati hanno strati dicellule continui (c), che si possono considerare analoghi agli strati dei gatti normali (d).
autonomo intracellulare (stranamente,abbiamo trovato che i topi variegatiin realtà hanno più fibre che vannodallo stesso lato rispetto ai topi nor-mali, e stiamo tentando di decifrarele ragioni di questo strano risultato).Per ora la sola conclusione che pos-siamo ricavare dalle nostre osservazio-ni è che il meccanismo che determinale vie nervose presuppone probabil-mente alcune interazioni tra le celluleall'interno della retina. In altre paro-le, sembra che il gene albino agiscamediante un meccanismo intracellula-re per produrre pigmento e median-te un meccanismo intercellulare nellaregolazione delle vie ottiche.
Altri ragguagli sul fondamento ge-netico dell'anormalità delle vie ottichesono stati ottenuti grazie a studi suvisoni d'allevamento. Gli allevatori divisoni apprezzano enormemente i viso-ni bianchi, o argentati, perciò hannoselezionato e mantenuto una gammadi combinazioni di geni che determi-nano una scarsezza di pigmento. Il vi-sone ha una buona vista binoculare epossiede strati ben definiti nel nucleogenicolato. Sanderson, Richard Sha-ckelford e io abbiamo studiato parec-chie mutazioni geneticamente distintenel visone e abbiamo trovato che tut-te le combinazioni di geni che produ-cono mancanza di pigmento nella re-tina sono associate a vie ottiche anor-mali. Questa anormalità è inversamen-te proporzionale alla quantità di pig-mento nella retina: gli animali bianchi,ma con occhi normalmente pigmentatinon dimostrano alcuna anomalia nellevie ottiche centrali.
Da queste osservazioni sul visone sideducono due importanti conclu-
sioni. La prima è che il gene albinonon è l'unico a produrre anormalitànelle vie ottiche. Alcuni loci dei geninettamente distinti dal locus albinopossono provocare anomalie nelle vieottiche, almeno nel visone: perciò nonpossiamo considerare l'assenza di tiro-sinasi o di ogni altro enzima preso sin-golarmente come unica causa dell'a-normalità. I particolari di questa re-lazione non sono noti, ma è da rile-vare che la formazione del pigmentonella retina precede la differenziazionedelle cellule gangliari e che la divisio-ne definitiva che produce queste cellu-le avviene in uno strato adiacente al-l'epitelio pigmentato. Quasi nello stes-so periodo in cui le cellule gangliarisi stanno formando, il calice oculareembrionale subisce una modificazioneimportante: la fessura fetale, attraver-so la quale l'arteria retinica penetranell'occhio, si va chiudendo. Il fatto
curioso è che la regione in cui avvie-ne questa chiusura è completamentepriva di pigmento. Rimane tuttora dadeterminare esattamente quale relazio-ne reciproca vi sia tra la distribuzionedel pigmento, la chiusura della fessu-ra fetale e la specificazione delle cel-lule gangliari.
Infine, ovviamente, ci si può chie-dere che cosa sappiamo sugli albiniumani. Vi sono anche in loro vie ot-tiche anormali? Se così avviene, qualeinfluenza ha questa anormalità sullaloro capacità visiva? 1.,'albinismo nel-l'uomo è generalmente associato a unavista poco acuta e a un movimentoorizzontale ritmico degli occhi, chia-mato nistagmo. Spesso si nota ancheun allineamento errato degli occhi, percui il responso sulla capacità visú/a diun albino è assai spesso difficile. Loesame del cervello dopo la morte po-trebbe rivelare un'anormalità, comenel nostro esame eseguito sul cervellodella tigre: finora però nessuno ha in-trapreso questo tipo di ricerca.
Le ricerche future sull' anormalitàlegata all'albinismo possono condurreverso varie direzioni. La capacità vi-siva e i collegamenti ottici centrali de-gli albini umani sono ovviamente mol-to interessanti, ma difficili da esami-nare. Gli studi sperimentali sullo svi-luppo embrionale negli animali albinici potrebbero mostrare il meccanismograzie al quale i geni agiscono sullamodificazione di sviluppo delle vie ner-vose cerebrali. La comprensione delmodo con cui un gene agisce sullo svi-luppo cerebrale ci potrebbe servire perdeterminare il meccanismo d'azione dimolti altri geni che prendono parteallo sviluppo del sistema nervoso. For-se l'aspetto più interessante della ca-pacità visiva degli albini è che in al-cuni cervelli i collegamenti centralipossono essere modificati per correg-gere una serie di stimoli mescolati di-sordinatamente. Se potessimo scoprirequalche altro aspetto dei processi disviluppo mediante i quali le rappresen-tazioni sensoriali riescono a essere co-sì corrette, comprenderemmo megliocome il cervello acquisisca normalmen-te una rappresentazione ordinata delmondo circostante.
L' anormalità genetica può essereconsiderata come un esperimento de-licato, che non si sarebbe potuto or-ganizzare con alcun mezzo chirurgico.Da un lato ci permette di studiare iprocessi di sviluppo che fanno insor-gere anormalità cerebrali: dall'altro cifornisce un'opportunità per determina-re il grado con cui le vie centrali sipossono adattare per far fronte a mo-delli anormali di attività cerebrale.
4647