Coinvolgimento del sistema olfattivo in corso di scrapie ... · olfattivo e corteccia frontale, con...
12
Coinvolgimento del sistema olfattivo in corso di scrapie naturale RIASSUNTO Recenti esperimenti evidenziano il coinvolgimento del sistema olfattivo (SO) nelle encefalopatie spongiformi: in criceti infet- tati per via intracerebrale la proteina prionica è stata rinvenuta nella mucosa nasale; altri studi hanno dimostrato anche il ruo- lo delle fibre olfattive nella neuroinvasione da parte dei prioni. Questo studio valuta il SO in corso di infezione naturale da scrapie. Campioni di mucosa nasale prelevati a livello di conca na- sale media, conca nasale ventrale e setto nasale da 24 pecore con scrapie naturale sono stati esaminati mediante immunoisto- chimica (IHC), immunofluorescenza (IF), PET blot e Western Blot (WB) per la presenza di proteina prionica (PrP Sc ). La de- posizione di PrP Sc è stata anche valutata mediante IHC sulle aree encefaliche in relazione col SO (bulbo e tratto olfattivo, cor- teccia frontale, lobo piriforme e ippocampo). La distribuzione di PrP Sc è apparsa nel SO sia a livello periferico che centrale. L’esame WB ha riscontrato come positivi 21 cam- pioni di mucosa olfattiva; in 14 di questi anche l’esame IHC ha confermato la positività. La deposizione di PrP Sc è apparsa nel- la conca nasale media, a livello del perinevrio dei nervi olfattivi, ma anche nel tessuto linfatico associato alla mucosa nasale; in nessun caso è stata rinvenuta in neuroni e fibre olfattive. Nel cervello un’intensa deposizione di PrP Sc è stata rilevata in bulbo olfattivo e corteccia frontale, con pattern di tipo submeningeo, subependimale e perivascolare. La presenza di PrP Sc sia nelle aree cerebrali a diretto contatto con il liquor che nel perinevrio dei nervi olfattivi, a ridosso del- l’estensione dello spazio subdurale che ne circonda le radici, supporta la recente scoperta della presenza di PrP Sc nel liquido ce- falo-rachidiano. PAROLE CHIAVE Pecora, scrapie, sistema olfattivo, PrP Sc , perinevrio. C. PORCARIO, F. MARTUCCI, B. IULINI, E. VALLINO COSTASSA, D. CORBELLINI, M.N. CHIEPPA, M.D. PINTORE, M. MAZZA, P.L. ACUTIS, C. CASALONE, C. CORONA Istituto Zooprofilattico Sperimentale del Piemonte, Liguria e Valle d’Aosta - Centro di Referenza Nazionale per le Encefalopatie Animali - Via Bologna, 148 - 10154 Torino C. Porcario et al. Large Animal Review 2010; 16: 21-32 21 l INTRODUZIONE Le encefalopatie spongiformi trasmissibili (EST) sono un gruppo di malattie neurodegenerative ad esito fatale causate dalla conversione conformazionale della proteina prionica cellulare (PrP C ) in un’isoforma patologica, proteasi-resisten- te, denominata proteina prionica patologica (PrP Sc ) 40 . Le EST includono la malattia di Creutzfeldt-Jacob (CJD) nell’uomo, l’encefalopatia spongiforme nel bovino (BSE), la scrapie nel- la pecora e nella capra e la malattia da deperimento cronico del cervo (CWD) nei cervidi selvatici 41 . Da tempo si ritiene che la via di trasmissione orizzontale, benché i suoi meccani- smi ancora non siano stati completamente chiariti 26 , possa giocare un ruolo preponderante nel mantenimento in cam- po dell’infezione da prioni, soprattutto in corso di scrapie e CWD 35 . Si ritiene che la possibilità di contatto con ambienti contaminati e fra animali infetti siano le due principali vie di trasmissione orizzontale delle malattie prioniche. In entram- bi i casi la diffusione dell’infezione è stata ricondotta ai tes- suti ed ai liquidi corporei degli animali infetti 45 . Si conosce da tempo il problema della contaminazione dei pascoli da placente 42 e carcasse 6 in decomposizione di animali affetti da malattie prioniche. Inoltre, la scoperta che i prioni possono aderire ai minerali del suolo mantenendo la capacità infetti- va 25 conferma il ruolo del terreno come serbatoio di infetti- vità. Quest’ultima, recentemente, è stata riscontrata in diver- si liquidi corporei, in particolare nell’urina di topi sperimen- talmente coinfettati da scrapie e da agenti di nefrite 48 e nella saliva e nel sangue di cervidi colpiti da CWD 34 . In corso di scrapie naturale la deposizione di PrP Sc è stata riscontrata nelle ghiandole salivari 57 , ed è stato anche dimostrato che il latte di pecora può rappresentare una via di trasmissione del- la malattia 31 . Nell’insieme questi riscontri indicano un pro- babile ruolo di sangue, saliva, urina e latte nella trasmissione orizzontale delle malattie prioniche, e lasciano supporre che il coinvolgimento degli organi di senso derivi dalla trasmis- sione dell’infettività tramite contatto con questi liquidi cor- porei. Alcune ricerche hanno già accertato il ruolo degli or- gani di senso come via di diffusione dei prioni verso il cer- vello. Il fenomeno della neuroinvasione è stato dimostrato in criceti a seguito di infezione sperimentale dei tessuti lingua- li, e questa via è apparsa addirittura più efficiente della tra- smissione per os nel provocare la malattia. Il sistema olfattivo (SO), oggetto del presente studio, è alta- mente sviluppato negli animali. Il senso dell’olfatto è alla ba- se di molti dei loro comportamenti etologici, come l’approv- vigionamento del cibo, l’esplorazione dell’ambiente, il corteg- giamento ed il riconoscimento della prole. In queste circo- Autore per la corrispondenza: Chiara Porcario ([email protected]).
Transcript of Coinvolgimento del sistema olfattivo in corso di scrapie ... · olfattivo e corteccia frontale, con...

Coinvolgimento del sistema olfattivoin corso di scrapie naturale
RIASSUNTORecenti esperimenti evidenziano il coinvolgimento del sistema olfattivo (SO) nelle encefalopatie spongiformi: in criceti infet-tati per via intracerebrale la proteina prionica è stata rinvenuta nella mucosa nasale; altri studi hanno dimostrato anche il ruo-lo delle fibre olfattive nella neuroinvasione da parte dei prioni.Questo studio valuta il SO in corso di infezione naturale da scrapie. Campioni di mucosa nasale prelevati a livello di conca na-sale media, conca nasale ventrale e setto nasale da 24 pecore con scrapie naturale sono stati esaminati mediante immunoisto-chimica (IHC), immunofluorescenza (IF), PET blot e Western Blot (WB) per la presenza di proteina prionica (PrPSc). La de-posizione di PrPSc è stata anche valutata mediante IHC sulle aree encefaliche in relazione col SO (bulbo e tratto olfattivo, cor-teccia frontale, lobo piriforme e ippocampo).La distribuzione di PrPSc è apparsa nel SO sia a livello periferico che centrale. L’esame WB ha riscontrato come positivi 21 cam-pioni di mucosa olfattiva; in 14 di questi anche l’esame IHC ha confermato la positività. La deposizione di PrPSc è apparsa nel-la conca nasale media, a livello del perinevrio dei nervi olfattivi, ma anche nel tessuto linfatico associato alla mucosa nasale; innessun caso è stata rinvenuta in neuroni e fibre olfattive. Nel cervello un’intensa deposizione di PrPSc è stata rilevata in bulboolfattivo e corteccia frontale, con pattern di tipo submeningeo, subependimale e perivascolare.La presenza di PrPSc sia nelle aree cerebrali a diretto contatto con il liquor che nel perinevrio dei nervi olfattivi, a ridosso del-l’estensione dello spazio subdurale che ne circonda le radici, supporta la recente scoperta della presenza di PrPSc nel liquido ce-falo-rachidiano.
PAROLE CHIAVEPecora, scrapie, sistema olfattivo, PrPSc, perinevrio.
C. PORCARIO, F. MARTUCCI, B. IULINI, E. VALLINO COSTASSA,D. CORBELLINI, M.N. CHIEPPA, M.D. PINTORE, M. MAZZA, P.L. ACUTIS,C. CASALONE, C. CORONA
Istituto Zooprofilattico Sperimentale del Piemonte, Liguria e Valle d’Aosta - Centro di Referenza Nazionaleper le Encefalopatie Animali - Via Bologna, 148 - 10154 Torino
C. Porcario et al. Large Animal Review 2010; 16: 21-32 21
l
INTRODUZIONE
Le encefalopatie spongiformi trasmissibili (EST) sono ungruppo di malattie neurodegenerative ad esito fatale causatedalla conversione conformazionale della proteina prionicacellulare (PrPC) in un’isoforma patologica, proteasi-resisten-te, denominata proteina prionica patologica (PrPSc)40. Le ESTincludono la malattia di Creutzfeldt-Jacob (CJD) nell’uomo,l’encefalopatia spongiforme nel bovino (BSE), la scrapie nel-la pecora e nella capra e la malattia da deperimento cronicodel cervo (CWD) nei cervidi selvatici41. Da tempo si ritieneche la via di trasmissione orizzontale, benché i suoi meccani-smi ancora non siano stati completamente chiariti26, possagiocare un ruolo preponderante nel mantenimento in cam-po dell’infezione da prioni, soprattutto in corso di scrapie eCWD35. Si ritiene che la possibilità di contatto con ambienticontaminati e fra animali infetti siano le due principali vie ditrasmissione orizzontale delle malattie prioniche. In entram-bi i casi la diffusione dell’infezione è stata ricondotta ai tes-suti ed ai liquidi corporei degli animali infetti45. Si conosceda tempo il problema della contaminazione dei pascoli daplacente42 e carcasse6 in decomposizione di animali affetti da
malattie prioniche. Inoltre, la scoperta che i prioni possonoaderire ai minerali del suolo mantenendo la capacità infetti-va25 conferma il ruolo del terreno come serbatoio di infetti-vità. Quest’ultima, recentemente, è stata riscontrata in diver-si liquidi corporei, in particolare nell’urina di topi sperimen-talmente coinfettati da scrapie e da agenti di nefrite48 e nellasaliva e nel sangue di cervidi colpiti da CWD34. In corso discrapie naturale la deposizione di PrPSc è stata riscontratanelle ghiandole salivari57, ed è stato anche dimostrato che illatte di pecora può rappresentare una via di trasmissione del-la malattia31. Nell’insieme questi riscontri indicano un pro-babile ruolo di sangue, saliva, urina e latte nella trasmissioneorizzontale delle malattie prioniche, e lasciano supporre cheil coinvolgimento degli organi di senso derivi dalla trasmis-sione dell’infettività tramite contatto con questi liquidi cor-porei. Alcune ricerche hanno già accertato il ruolo degli or-gani di senso come via di diffusione dei prioni verso il cer-vello. Il fenomeno della neuroinvasione è stato dimostrato incriceti a seguito di infezione sperimentale dei tessuti lingua-li, e questa via è apparsa addirittura più efficiente della tra-smissione per os nel provocare la malattia.Il sistema olfattivo (SO), oggetto del presente studio, è alta-mente sviluppato negli animali. Il senso dell’olfatto è alla ba-se di molti dei loro comportamenti etologici, come l’approv-vigionamento del cibo, l’esplorazione dell’ambiente, il corteg-giamento ed il riconoscimento della prole. In queste circo-
Autore per la corrispondenza:Chiara Porcario ([email protected]).
Porcario_imp:Porcario_imp 26-01-2010 15:37 Pagina 21

22 Coinvolgimento del sistema olfattivo in corso di scrapie naturale
stanze è fattibile che la mucosa nasale venga a contatto conmateriale infetto da prioni, offrendo così la possibilità alla Pr-PSc di penetrare nell’organismo e ivi diffondersi. La diffusio-ne centripeta verso l’encefalo della proteina prionica è statainfatti dimostrata per via sperimentale a seguito sia di infe-zione di criceti con l’hyper strain dell’encefalopatia trasmissi-bile del visone (HY TME)28 che di pecore con la scrapie18.Nelle malattie da prioni dell’uomo un possibile ruolo del-l’olfatto è stato anche ipotizzato per la patogenesi della va-riante di Creutzfeldt-Jacob36. A riguardo si ritiene infatti cheil SO rappresenti una possibile via di risalita verso l’encefaloper l’agente eziologico della BSE, presente nella polvere di fa-rine animali contaminate46,49,51. Sperimentalmente è statopossibile dimostrare non solo che i prioni si propagano pervia centripeta attraverso le vie olfattive ma anche che può av-venire la loro migrazione centrifuga dal cervello alla mucosaolfattiva. Criceti inoculati per via intracerebrale con l’agentedella TME hanno mostrato deposizione di PrPSc nei neuroniolfattivi, nelle cellule di supporto della mucosa nasale e neltessuto linfatico associato alla mucosa nasale (NALT)10. Que-sti recenti studi di patogenesi hanno quindi indicato uncoinvolgimento del SO in corso di infezione sperimentale daprioni, ma pochi hanno indagato questo aspetto in corso diinfezione naturale. La deposizione di PrPSc in mucosa olfat-tiva è stata osservata in pazienti affetti da CJD sporadica(sCJD)63, con maggiore intensità nelle ciglia dei neuroni ol-fattivi e in grado minore nelle cellule basali. Lo studio di unabiopsia olfattiva prelevata da un paziente affetto da sCJD hasvelato la presenza di PrPSc a 45 giorni dall’inizio dei sintomiclinici, dato che suggerisce un precoce coinvolgimento dellamucosa nasale nella diffusione dell’infezione55.Ad oggi non è stato ancora riportato alcuno studio sul ri-scontro di PrPSc nel SO di animali con infezione naturale damalattie da prioni. Ai fini di sopperire a questa mancanza, ilpresente studio si è posto l’obiettivo di determinare se e inche misura il SO centrale e periferico fossero coinvolti nellaspecie ovina in corso di infezione naturale da scrapie.In particolare, l’intenzione di indagare in condizioni natura-li d’infezione la deposizione di PrPSc nelle cellule della mu-cosa nasale, che sono soggette ad un turnover molto rapido,è stato fondamentale per determinare la possibilità del rila-scio dei prioni nell’ambiente attraverso le secrezioni nasali,data per ipotesi la fattibilità di un loro ruolo nella trasmis-sione orizzontale della malattia. In quest’ottica, campioniprelevati dalle cavità nasali e dalle aree del cervello relazio-nate al SO di pecore affette da scrapie naturale sono stati rac-colti ed esaminati con diverse metodiche.
MATERIALI E METODI
Animali e raccolta dei tessuti Ventiquattro pecore di razza Biellese, da 3 ad 8 anni di età eprovenienti da uno stesso gregge piemontese affetto da scra-pie, sono state abbattute nell’ambito di un programma di ab-battimento selettivo per eradicare la malattia. Tutte le pecoreappartenevano a genotipi suscettibili (ARQ/ARQ [n = 20] eARQ/AHQ [n = 4]). Tutti gli animali sono stati sottoposti adun’adeguata visita clinica neurologica ante mortem prima del-l’eutanasia. Quattro delle 24 pecore mostravano sintomi clini-ci (tremori, emaciazione o incapacità di stazione); le rimanen-ti pecore non mostravano sintomi (Tabella 1). Tutte le 24 pe-
core sono risultate positive al test rapido per scrapie (Bio-RadTeSeE rapid assay; Bio-Rad Laboratories Inc., Hercules, CA);la malattia è stata poi confermata per mezzo di analisi di isto-logia, IHC e WB sul tronco encefalico. I controlli erano 10 pe-core regolarmente macellate risultate negative ai test di con-ferma per TSE.Alla necroscopia il cervello è stato rimosso e poi tagliato lon-gitudinalmente in due metà. Una metà è stata congelata a–80°C in vista di analisi biochimiche, l’altra è stata fissata informalina tamponata al 10% per analisi immunoistochimi-che. Il cranio è stato tagliato longitudinalmente in due metàper studiare i tessuti delle cavità nasali. Nella pecora le duecavità nasali, separate dal setto nasale (SN), contengono cia-scuna una conca nasale dorsale, una conca nasale media(CNM) (facente parte del sistema degli endoturbinati) e unaconca nasale ventrale (CNV). I campioni sono stati preleva-ti da ciascun lato di SN, CNM e CNV, poiché la concentra-zione dei neuroni olfattivi è più elevata in queste aree30. Piùin dettaglio, i campioni sono stati prelevati nella porzionepiù aborale della CNM e del SN, nelle immediate vicinanzedella lamina cribrosa dell’etmoide, dove la mucosa olfattiva èfacilmente riconoscibile per via di un naturale colore gialla-stro. I campioni di CNV sono stati ricavati solo dalla porzio-ne più aborale, che è l’unica a presentare l’epitelio olfattivo(EO). I campioni di un lato sono stati fissati in formalinatamponata al 10% per le analisi di IHC, di IF e di PET blot,e i tessuti corrispondenti dell’altro lato sono stati congelatiper l’analisi WB.
WB e quantificazione della PrPSc
(i) Pre-trattamento dei tessuti extraneurali. LaCNM, la mucosa del SN e i campioni di CNV sono stati taglia-ti in piccoli pezzi col bisturi e incubati overnight in PBS all’1%di tripsina, a temperatura ambiente e in costante agitazione.(ii) Analisi WB. Una quantità non inferiore a 200 mg deidiversi tessuti è stata omogeneizzata in sarcosyl al 10% e chia-rificata per mezzo di centrifugazione a 22,000 × g per 20 mi-nuti. Per i campioni di tessuto nervoso extraneurale, il surna-tante è stato raccolto ed incubato con enzima nucleasi benzo-nasi (50 meq/ml) (Novagen, San Diego, CA) per 30 min a37°C. I campioni sono stati chiarificati mediante centrifuga-zione a 22,000 × g per 20 minuti a 10°C, e i surnatanti sonostati incubati con 40 µg/ml di proteinasi K per 1 ora a 37°C inagitazione continua. Dopo centrifugazione a 215,000 × g per1 ora a 10°C (Optima TLX ultracentrifuga, rotore TLA 110;Beckman Coulter, Fullerton, CA), il pellet è stato dissolto inLaemmli buffer e fatto bollire per 10 minuti a 99°C. I cam-pioni sono stati separati in elettroforesi su minigel al 12% dipoliacrilammide e poi trasferiti su di una membrana di PVDF(Immobilion P; Millipore, Billerica, MA) mediante wet blot-ting. La PrPSc è stata marcata mediante impiego dell’anticor-po monoclonale P4 (0,1 µg/ml; R-Biopharm, Darmstadt,Germania)19 e di un anticorpo anti-topo coniugato con fosfa-tasi alcalina (Zymed, Invitrogen, Carlsbad, CA). La reazione èstata rivelata in chemiluminescenza (ImmunoStar; Bio-Rad,Hercules, CA) e visualizzata su lastre radiografiche HyperfilmECL (GE-Healthcare LTD., St. Giles, United Kingdom) o me-diante analizzatore di immagini UVI Prochemi (Uvitec,Cambridge, United Kingdom).(iii) Quantificazione della PrPSc. Al fine di stimarela concentrazione relativa di PrPSc nella CNM, nella mucosadel SN e nella CNV, abbiamo confrontato le intensità dei se-
Porcario_imp:Porcario_imp 26-01-2010 15:37 Pagina 22
C. Porcario et al. Large Animal Review 2010; 16: 21-32 23
gnali che apparivano al WB a seguito della digestione conproteinasi K con curve di calibrazione ottenute diluendo iltronco encefalico della pecora positiva per scrapie esaminatacon un omogenato di mucosa nasale di pecora sana. Questoconfronto è stato eseguito su 3 animali affetti da scrapie fraquelli considerati nello studio, due dei quali avevano genoti-po ARQ/ARQ e uno ARQ/AHQ. Il metodo di estrazione e latecnica WB erano identiche a quelle prima descritte. Le ana-lisi di quantificazione sono state effettuate utilizzando unutilizzatore d’immagini UVI Prochemi (Uvitec, Cambridge,United Kingdom).
IHCA seguito di fissazione, sono state tagliate sezioni coronali dicervello dello spessore di 5 mm. Le aree encefaliche relazio-nate al SO (bulbo olfattivo, tratto olfattorio, corteccia fronta-le a livello dei nuclei della base, lobo piriforme e ippocampo)sono state campionate, processate secondo i metodi di routi-ne e incluse in paraffina7.Ad eccezione della mucosa del SN, gli altri campioni di tes-suto nasale sono stati posti in una soluzione decalcificante(Carlo Erba, Rodano, Italy) a temperatura ambiente per 4
giorni (CNM) o per 2 giorni (CNV). I campioni sono statirisciacquati sotto acqua corrente, tagliati trasversalmente inpezzi dello spessore di 0,5 cm e poi processati come sopra de-scritto per le sezioni di tessuto cerebrale.Per ciascuna pecora sono state tagliate sezioni dello spessoredi 5 µm di ogni area relazionata al SO e di ogni tessuto di ca-vità nasale. I tessuti cerebrali e quelli delle cavità nasali sonostati quindi sottoposti ad IHC per PrPSc. Le sezioni sono sta-te sparaffinate e reidratate secondo protocollo di routine epoi immerse in acido formico al 98% per 25 minuti. Dopolavaggio in acqua distillata le sezioni sono state sottoposte adautoclave per 30 minuti a 121°C in tampone citrato (pH 6,1).Le perossidasi endogene sono state bloccate con perossido diidrogeno al 3% in metanolo per 20 minuti a temperaturaambiente. Per bloccare gli antigeni tissutali non specifici, lesezioni sono state incubate con siero aspecifico di cavallo al5% per 20 min a temperatura ambiente e a seguito incubateper un’ora con l’anticorpo primario monoclonaleF99/97.6.152 (epitopo QYQRES, aminoacidi 220-225 dellaPrP ovina, diluizione 1:1000, VMRD Inc., Pullman, WA).Dopo lavaggio sulle sezioni è stato applicato un anticorpo se-condario biotinilato di capra, anti-topo (diluizione 1:200,
a +, immunoreattività per PrPSc; –, assenza di immunoreattività per PrPSc
b BO: bulbo olfattivo; TO: tratto olfattorio; CF: corteccia frontale; LP: lobo piriforme; Hi: ippocampo; ND: non disponibile; –: assente; – / +: poco evidente;+: lieve; + +: moderato; + + +: marcato; + + + +: molto marcato.
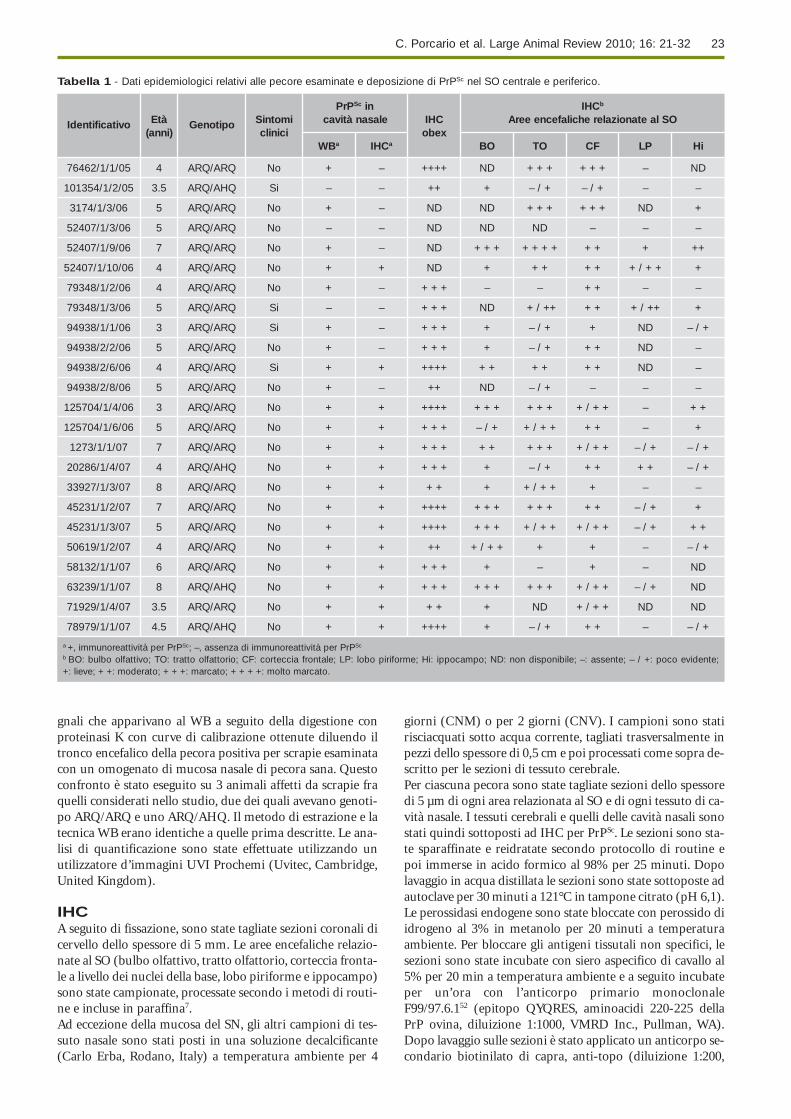
Tabella 1 - Dati epidemiologici relativi alle pecore esaminate e deposizione di PrPSc nel SO centrale e periferico.
PrPSc in IHCb
Identificativo Età Genotipo Sintomi cavità nasale IHC Aree encefaliche relazionate al SO(anni) clinici obex
WBa IHCa BO TO CF LP Hi
76462/1/1/05 4 ARQ/ARQ No + – ++++ ND + + + + + + – ND
101354/1/2/05 3.5 ARQ/AHQ Si – – ++ + – / + – / + – –
3174/1/3/06 5 ARQ/ARQ No + – ND ND + + + + + + ND +
52407/1/3/06 5 ARQ/ARQ No – – ND ND ND – – –
52407/1/9/06 7 ARQ/ARQ No + – ND + + + + + + + + + + ++
52407/1/10/06 4 ARQ/ARQ No + + ND + + + + + + / + + +
79348/1/2/06 4 ARQ/ARQ No + – + + + – – + + – –
79348/1/3/06 5 ARQ/ARQ Si – – + + + ND + / ++ + + + / ++ +
94938/1/1/06 3 ARQ/ARQ Si + – + + + + – / + + ND – / +
94938/2/2/06 5 ARQ/ARQ No + – + + + + – / + + + ND –
94938/2/6/06 4 ARQ/ARQ Si + + ++++ + + + + + + ND –
94938/2/8/06 5 ARQ/ARQ No + – ++ ND – / + – – –
125704/1/4/06 3 ARQ/ARQ No + + ++++ + + + + + + + / + + – + +
125704/1/6/06 5 ARQ/ARQ No + + + + + – / + + / + + + + – +
1273/1/1/07 7 ARQ/ARQ No + + + + + + + + + + + / + + – / + – / +
20286/1/4/07 4 ARQ/AHQ No + + + + + + – / + + + + + – / +
33927/1/3/07 8 ARQ/ARQ No + + + + + + / + + + – –
45231/1/2/07 7 ARQ/ARQ No + + ++++ + + + + + + + + – / + +
45231/1/3/07 5 ARQ/ARQ No + + ++++ + + + + / + + + / + + – / + + +
50619/1/2/07 4 ARQ/ARQ No + + ++ + / + + + + – – / +
58132/1/1/07 6 ARQ/ARQ No + + + + + + – + – ND
63239/1/1/07 8 ARQ/AHQ No + + + + + + + + + + + + / + + – / + ND
71929/1/4/07 3.5 ARQ/ARQ No + + + + + ND + / + + ND ND
78979/1/1/07 4.5 ARQ/AHQ No + + ++++ + – / + + + – – / +
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 23

24 Coinvolgimento del sistema olfattivo in corso di scrapie naturale
Vector Laboratories, Burlingame, CA), per 30 min a tempe-ratura ambiente, seguito dal complesso avidina-biotina-pe-rossidasi (Vectastain ABC kit, Vector Laboratories, Burlinga-me, CA), secondo le istruzioni fornite. La PrPSc è stata visua-lizzata utilizzando la 3, 3’ diaminobenzidina (Dakocytoma-tion, Carpenteria, CA) come cromogeno; le sezioni sono sta-te quindi controcolorate con ematossilina di Meyer. I cam-pioni impiegati come controlli negativi sono stati anch’essitestati.La deposizione di PrPSc nelle aree relazionate al SO è stata va-lutata mediante microscopia ottica ed è stato assegnato ungrado di intensità ai diversi pattern riscontrati: assente (-),poco evidente (-/+), lieve (+), moderato (+ +), marcato (+ ++) o molto marcato (+ + + +)24.
ImmunofluorescenzaSono state tagliate sezioni seriali dei tessuti delle cavità nasaliinizialmente risultati positivi in IHC per la presenza di PrPSc.Le indagini di IF sono state condotte secondo i due diversiprotocolli qui di seguito descritti.(i) Detezione di PrPSc e carnosina. Le sezioni ditessuto sono state sparaffinate, reidratate, trattate con acidoformico, e autoclavate come nel protocollo di IHC sopra de-scritto. Dopo autoclavaggio, le sezioni sono state lavate in ac-qua distillata e processate secondo un protocollo di doppiaIF. Per bloccare gli antigeni tissutali non specifici, le sezionisono state incubate con siero di capra diluito al 5% in PBSper 20 minuti a temperatura ambiente. L’anticorpo primariomonoclonale F99/97.6.1 (VMRD Inc., Pullman, WA), diluito1:200, è stato applicato per 1 ora a temperatura ambiente. Lesezioni di tessuto delle cavità nasali sono state quindi incu-bate a temperatura ambiente con un anticorpo secondario dicapra anti-topo coniugato con Cy3 (Jackson Immunoresear-ch Laboratories Inc., West Grove, PA), diluito 1:100, per 30min. Dopo lavaggio in tampone TRIS, le sezioni di tessutosono state incubate a temperatura ambiente con un anticor-po primario policlonale anti-carnosina (gentilmente fornitodalla Dott.ssa S. De Marchis, Dipartimento di Biologia Ani-male e dell’uomo, Università di Torino) diluito 1:150 in tam-pone TRIS addizionato di Tween 20. Dopo risciacquo intampone TRIS addizionato di Tween 20, è stato applicato per30 min a temperatura ambiente un anticorpo secondario dicapra, anti-coniglio, coniugato con Cy2 (Jackson Immuno-Research Laboratories Inc., West Grove, PA), diluito 1:100. Lesezioni sono state montate in PBS diluito 1:3 in glicerolo edesaminate con un microscopio Nikon Eclipse 80i per fluore-scenza (Nikon Instruments, Florence, Italy), usando filtri perfluoresceina isotiacianato e rodamina tetrametile. La specifi-cità degli anticorpi secondari è stata testata applicando i pre-cedenti antisieri senza gli anticorpi primari. Non è stata ri-scontrata alcuna marcatura né per PrPSc né per carnosinaomettendo l’anticorpo primario.(ii) Detezione di PrPSc e citocheratine. Non èstato possibile applicare un protocollo di doppia IF per PrPSc
e citocheratine poiché i due antigeni necessitano di esseresmascherati mediante due diverse tecniche al fine di massi-mizzarne l’intensità di marcatura. Perciò si è deciso di adot-tare l’impiego di singoli protocolli di IF per PrPSc e citoche-ratine su sezioni seriali di tessuto. La marcatura per PrPSc èavvenuta secondo il protocollo in precedenza descritto. Losmascheramento per le citocheratine è avvenuto a bagnoma-ria (95-99°C) in tampone Tris-EDTA pH 9 per 20 minuti. Le
sezioni sono state in seguito incubate per 1 ora con un anti-corpo primario monoclonale anti-citocheratine umane, clo-ne AE1/AE3 (Dakocytomation, Carpenteria, CA), diluito1:50, in siero di capra diluito al 5% in PBS. Come anticorposecondario è stato applicato un anticorpo anti-topo di capraconiugato con Cy2 (Jackson ImmunoResearch LaboratoriesInc., West Grove, PA), diluito 1:100, per 30 minuti a tempe-ratura ambiente.Al fine di studiare la colocalizzazione di PrPSc e citocheratinesono state scattate immagini di aree strettamente corrispon-denti su sezioni seriali di tessuto, e la sovrapposizione digita-le e la manipolazione delle immagini sono state eseguite uti-lizzando il software CS Photoshop 8.0.
Analisi PET blotIl PET blotting è stato eseguito come altrove descritto43 conlievi modifiche. Sezioni in paraffina dello spessore di 5 µmsono state montate su membrane di nitrocellulosa e lasciateasciugare orizzontalmente overnight a 60°C. Dopo lavaggiole sezioni sono state sottoposte a proteolisi con proteinasi K(25 µg/ml) a 55°C per 2 ore. Le sezioni sono state quindi de-naturate in guanidina isotiocianato 3 M per 10 min, blocca-te con caseina per 30 min, e incubate a 4°C overnight conl’anticorpo monoclonale F99/97.6.1 (diluito 1:2000). Lamarcatura ha previsto l’impiego del sistema di rilevazioneABC-AmP (Vector Laboratories, Burlingame, CA). La mar-catura è stata visualizzata utilizzando 5-bromo-4-cloro-3 in-dolil tetrazolio nitroblu fosfato e osservata allo stereomicro-scopio. Per i controlli negativi, l’anticorpo primario è statoomesso e sostituito con siero aspecifico.
RISULTATI
La PrPSc è stata riscontrata all’analisi WB nei tessuti delle ca-vità nasali di 21 su 24 delle pecore esaminate (Tabella 1); 18delle 21 pecore positive al WB avevano genotipo ARQ/ARQ,e tre erano ARQ/AHQ. Due delle pecore positive all’analisiWB, entrambe di genotipo ARQ/ARQ, mostravano sintomiclinici di malattia. Le 3 pecore negative all’analisi WB aveva-no 2 diversi genotipi: due erano ARQ/ARQ, una delle qualisintomatica, la terza invece, anch’essa sintomatica, aveva ge-notipo ARQ/AHQ.All’analisi WB la deposizione di PrPSc è stata più frequente-mente riscontrata a livello di conca nasale media (n = 19)(Tabella 2 e Figura 1A). In 5 animali la PrPSc è stata identifi-cata esclusivamente a livello di conca nasale media, in 5 ani-mali sia a livello di mucosa di conca nasale media che di set-to nasale, in 2 animali sia nella mucosa della conca nasalemedia che della conca nasale ventrale, e in 7 casi in tutte e 3le aree di cui sopra. Non è stata riscontrata alcuna deposizio-ne di PrPSc all’analisi WB nella conca nasale media di 2 pe-core; in una di queste è stata evidenziata a livello di setto na-sale e di mucosa della conca nasale ventrale e nell’altra sol-tanto nella mucosa della conca nasale ventrale (Tabella 2).L’analisi immunoistochimica per la PrPSc ha confermato lapositività dei risultati all’analisi WB sulle cavità nasali di 14animali e non su quelle dei restanti 7. Tale dato è ascrivibilealla maggiore sensibilità della tecnica WB impiegata in que-sto studio, che, essendo basata sul campionamento di unamaggiore quantità di tessuto rispetto all’immunoistochimicae su di una maggiore concentrazione della PrPSc grazie ad ul-
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 24

C. Porcario et al. Large Animal Review 2010; 16: 21-32 25
tracentrifugazione, aumenta le possibilità di riscontrare que-st’ultima nel caso fosse presente.In 3 animali non è stata riscontrata la presenza di PrPSc innessun tessuto della cavità nasale esaminato né medianteIHC né mediante analisi WB (Tabella 2). Tutti e 10 i control-li negativi sono risultati tali sia all’esame WB che immunoi-stochimico.
Quantificazione della PrPSc
in cavità nasaleL’intensità di segnale delle conche nasali medie positive eraall’incirca simile a quella ottenuta dalla diluizione di 5 mgdel corrispondente omogenato di tronco encefalico in 50 mgdi omogenato di mucosa nasale negativa. Da ciò possiamostimare che i livelli di PrPSc nei 3 casi esaminati fossero piùbassi di quelli riscontrati nei corrispondenti tronchi encefa-lici di un fattore all’incirca pari a 1 x 10–1. Le intensità di se-gnale delle mucose positive del setto nasale e della conca na-sale ventrale erano generalmente più basse di quelle riscon-trate nei corrispondenti tronchi encefalici di un fattore com-preso fra 1 x 10–5 e 1 x 10–6 (Fig. 1B). La rappresentazionegrafica della percentuale di intensità di segnale di PrPSc in
mucosa olfattiva in rapporto al cervello indica che l’am-montare più elevato di PrPSc riscontrata nella conca nasalemedia era approssimativamente pari al 12% di quello delcervello (Fig. 1C).
Localizzazione della PrPSc
in cavità nasaleL’analisi immunoistochimica ha evidenziato la presenza diPrPSc soltanto a livello di perinevrio dei nervi olfattivi (PNO)in 10 animali (Figg. 2A e C), dove è apparsa come fini depo-siti granulari intracellulari. In particolare, la deposizione diPrPSc era localizzata nel PNO dei fila olfactoria nella porzio-ne di cavità nasale adiacente ai fori della lamina cribrosa del-l’etmoide. In due casi, la deposizione di proteina prionica èstata riscontrata sia nel PNO che nel NALT (Fig. 2B). Due ca-si erano positivi soltanto nel NALT. La deposizione di PrPSc
non è mai stata riscontrata né nelle cellule basali e di soste-gno dell’epitelio olfattivo né nel pericario e negli assoni deineuroni olfattivi (Fig. 2F). L’analisi mediante tecnica di PETblot ha confermato la presenza di PrPSc riscontrata dall’im-munoistochimica nel PNO, consentendo così di escludere lapresenza di artefatti (Fig. 3).
a +, immunoreattività per PrPSc; –, assenza di immunoreattività per PrPSc;ND, non disponibile.
Tabella 2 - Localizzazione della deposizione di PrPSc nelle cavitànasali della pecora all’analisi WB e IHC.
WBa IHCa
IdentificativoCNM SN CNV EO PNO NALT
76462/1/1/05 – – + – – –
101354/1/2/05 ND – – – – –
3174/1/3/06 + + – – – –
52407/1/3/06 – – – – – –
52407/1/9/06 + + – – – –
52407/1/10/06 + – – – + –
79348/1/2/06 + – – – – –
79348/1/3/06 – – – – – –
94938/1/1/06 + – – – – –
94938/2/2/06 + + – – – –
94938/2/6/06 + – + – – +
94938/2/8/06 + – + – – –
125704/1/4/06 + + + – + –
125704/1/6/06 + + + – + –
1273/1/1/07 + + – – + +
20286/1/4/07 + + – – + –
33927/1/3/07 + + + – + +
45231/1/2/07 + – – – + –
45231/1/3/07 + + + – + –
50619/1/2/07 + + + – + –
58132/1/1/07 + – – – + –
63239/1/1/07 – + + – – +
71929/1/4/07 + + + – + –
78979/1/1/07 + + + – + –
Figura 1 - (A) Riscontro di PrPSc per mezzo di WB altamente sen-sibile a livello di tronco encefalico e mucosa nasale di 3 pecore po-sitive per scrapie. Bande da 1 a 4: tronco encefalico, CNM, CNV eSN, rispettivamente, di 3 pecore positive per scrapie; banda C-:CNM di un campione di pecora sana. La quantità per ogni campio-ne di equivalenti di tessuto è indicata in milligrammi. Tutti i campio-ni sono stati digeriti con proteinasi K. (B) WB di un campione ditronco encefalico positivo (identificativo 125704/1/4/06) diluito se-rialmente in un campione di mucosa di CNM negativo: 1 × 10–1 (ban-da 1), 1× 10–2 (banda 2), 1 × 10–3 (banda 3), 1 × 10–4 (banda 4), 1 ×10–5 (banda 5), 1 × 10–6 (banda 6). mw, masse molecolari. La quan-tità per ogni campione di equivalenti di tessuto è indicata in milli-grammi. Tutti i campioni sono stati digeriti con proteinasi K. (C)Rappresentazione grafica della percentuale di intensità di segnale diPrPSc a livello di CNM, CNV e SN in rapporto al cervello di 3 pecoreaffette da scrapie.
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 25

26 Coinvolgimento del sistema olfattivo in corso di scrapie naturale
L’innervazione della cavità nasale è fornita da fibre del primonervo cranico, che trasporta le sensazioni olfattive dalla mu-cosa nasale al cervello, e da rami del nervo trigemino, in par-ticolare dalla sua branca oftalmica, che veicola stimoli mec-canici, termici, chimici e nocicettivi47. Utilizzando una tecni-ca di IF per PrPSc e per la carnosina, marker specifico per lefibre olfattive4, abbiamo dimostrato che la deposizione di Pr-PSc circondava i nervi olfattivi (Figg. 4A, B e C). La localizza-
zione della PrPSc soltanto a livello del PNO ha sollevato la do-manda su quale fosse il tipo cellulare coinvolto nella deposi-zione di PrPSc. Secondo quanto descritto in studi preceden-ti12,13,23,60, le cellule del perinevrio a ridosso dei nervi olfattivia livello della lamina cribrosa dell’etmoide sono un’estensio-ne delle meningi, le quali, passando attraverso i fori della la-mina, vengono a circondare i fila olfactoria che la attraversa-no (Fig. 5).
Figura 2 - Distribuzione di PrPSc nelle cavità nasali della pecora. (A e C) PNO con marcatura per PrPSc (× 20 e × 100, rispettivamente);(E) PNO di una pecora utilizzata come controllo negativo (× 100); (B) marcatura per PrPSc nel NALT (× 20). Un’immagine a maggiore in-grandimento (× 40) di NALT con deposizione di PrPSc è rappresentata nell’area rettangolare in alto a destra del pannello B. (D) NALT di unapecora controllo negativo (× 20); (F) assenza di marcatura per PrPSc nella mucosa olfattiva di una pecora positiva per scrapie (× 40).
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 26

C. Porcario et al. Large Animal Review 2010; 16: 21-32 27
Per distinguere le cellule meningee dai fibroblasti ad essestrettamente apposti abbiamo voluto identificare un markerspecifico per le cellule meningee. Al contrario dell’origineembriologica dei fibroblasti16,32, quella delle meningi può es-sere parzialmente ricondotta a cellule di derivazione dell’ec-toderma neurale17,20,29,38, da parte delle quali è nota l’espres-sione di citocheratine15; per questo motivo è stato utilizzatoun anticorpo contro questi markers per evidenziare esclusi-vamente le cellule meningee del PNO. L’immunofluorescen-za per PrPSc e citocheratine su sezioni seriali di tessuto ha di-mostrato che le citocheratine e la PrPSc si colocalizzavano al-l’interno dello strato meningeo del PNO (Figg. 4D, E, F) eche non si evidenziava nessuna deposizione di PrPSc negli al-tri tipi cellulari costituenti il perinevrio.Nelle cavità nasali della pecora il NALT è costituito da singo-li noduli linfatici distribuiti in tutto lo spessore della laminapropria della mucosa olfattiva. La struttura dei noduli linfa-
tici è simile a quella dei follicoli linfatici secondari descrittinella pecora nelle placche del Peyer a livello del digiuno54. Al-l’analisi immunoistochimica l’accumulo di PrPSc nel NALTappariva come depositi granulari localizzati nel citoplasma osulla superficie di cellule che, sulla base della loro morfolo-gia, erano riconducibili a macrofagi e a cellule follicolaridendritiche (Fig. 2B).
Localizzazione della PrPSc
nelle aree encefaliche relazionate al sistema olfattivoNelle aree encefaliche relazionate al SO delle pecore esami-nate la deposizione di PrPSc era più abbondante a livello deibulbi olfattivi dove mostrava generalmente un’intensità mar-cata. Nei bulbi olfattivi sono stati frequentemente evidenzia-ti i pattern submeningeo (Fig. 6A) e subependimale (Fig.6C), in concomitanza con un pattern di tipo finemente pun-tato nei glomeruli olfattivi. Nei campioni dove lo strato deinervi olfattivi era valutabile, la PrPSc veniva evidenziata comedepositi granulari lungo il decorso delle fibre olfattive (datinon pubblicati). Lo strato plessiforme interno, lo strato del-le cellule mitrali e lo strato plessiforme esterno mostravanoprevalentemente un pattern gliale di deposizione della PrPSc.Nel bulbo olfattivo accessorio, laddove esaminabile, si evi-denziava sempre un pattern di tipo finemente puntato neiglomeruli (Fig. 6A).A livello del tratto olfattivo la marcatura per PrPSc apparivada moderata a marcata, e i pattern di deposizione della PrPSc
erano soprattutto di tipo submeningeo e gliale (Fig. 6E). A li-vello di corteccia frontale erano apprezzabili pattern di tiposubmeningeo e gliale di moderata intensità e una deposizio-ne marcata della PrPSc di tipo perivascolare (Fig. 6G).Nelle aree relazionate al SO del sistema limbico (ovvero il lo-bo piriforme e l’ippocampo), l’intensità di marcatura della
Figura 3 - Analisi PET blot di fasci di nervi olfattivi di una pecoraaffetta da scrapie (A) e di un controllo negativo (B). La freccia indi-ca la localizzazione dei depositi di PrPSc.
Figura 4 - Immunofluorescenza su di un nervo olfattivo di una pecora affetta da scrapie. (Da A a C) Doppia IF per PrPSc e carnosina (×60). (A) Anticorpo anti-PrP (Cy3, rosso); (B) anticorpo anti-carnosina (Cy2, verde); (C) sovrapposizione dei pannelli A e B. (Da D a F) IFper PrPSc e citocheratine (× 60). (D) Anticorpo anti-PrP (Cy3, rosso); (E) anticorpo anti-citocheratine (Cy2, verde); (F) sovrapposizione deipannelli D ed E.
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 27

28 Coinvolgimento del sistema olfattivo in corso di scrapie naturale
PrPSc appariva lieve o persino assente, costituita prevalente-mente da depositi di tipo submeningeo (Fig. 6I), subependi-male (Fig. 6K) e perivascolare. Questi erano i principali pat-tern evidenziati in tutte le aree cerebrali relazionate al SO,tuttavia non ne erano esclusive, poiché era possibile riscon-trarli in tutto il cervello.
DISCUSSIONE
Il coinvolgimento del SO nel corso di malattie da prioni ani-mali ad oggi non è stato ancora indagato in maniera ap-profondita. Ad eccezione di tre precedenti studi sperimenta-li su modelli di roditori10,21,28, l’unico studio in merito alla de-posizione di PrPSc nel SO in condizioni d’infezione naturaleè stata una presentazione poster da parte di Vidal e coll. nel200359, che considerava un campione di 19 vacche affette daBSE. In tutte le aree encefaliche relazionate al SO esaminatela presenza di proteina prionica veniva riscontrata in mag-giori quantità nelle vacche a stadi più avanzati della malattia.Tuttavia, nel corso dello studio non venne riscontrata depo-sizione di PrPSc né all’analisi WB né in IHC sia nei neuroniolfattivi che nelle cellule di supporto e basali dell’epitelio ol-fattivo. In base a quanto è ad oggi noto, in letteratura non so-no riportati altri studi che abbiano indagato la deposizionedi PrPSc nel SO di pecore affette da scrapie naturale. I nostridati confermano non solo il coinvolgimento del SO centralein corso di scrapie naturale, come riportato da Vidal e coll.59
in merito alla BSE, ma anche che la deposizione di PrPSc è ri-scontrabile nei tessuti delle cavità nasali. L’analisi WB ha in-fatti riscontrato a questo livello la presenza di PrPSc in 21 del-
le 24 pecore affette da scrapie esaminate, che includevanoanimali sia sintomatici che asintomatici, a dimostrazione delfatto che i tessuti delle cavità nasali sono coinvolti in corso discrapie, persino nelle fasi asintomatiche della malattia. Alcontrario, nell’ambito del nostro studio il SO periferico, co-stituito dai neuroni olfattivi e valutabile solo mediante IHC,è apparso completamente privo di PrPSc.In merito alla distribuzione anatomica della PrPSc a livello dimucosa nasale, sia l’epitelio respiratorio che quello olfattivone apparivano completamente privi. Questa osservazioneconcorda con quanto precedentemente riportato in meritoalla distribuzione della PrPSc in criceti infettati per via extra-nasale con l’agente della TME28. Alla base di questi risultativi può essere il fatto che la PrPC è presente a livello di EOsoltanto in piccole quantità, come è stato riportato nellaspecie umana63 o all’elevato turnover dei neuroni olfattivi,che, nel corso della vita, vanno incontro a continua rigene-razione9. Si ritiene infatti che la breve durata di vita dei neu-roni olfattivi (da 6 a 10 settimane)56 non sia favorevole alladeposizione di PrPSc nell’epitelio olfattivo10. Ciononostante,in letteratura è stato riportato il riscontro di PrPSc nell’EO dicriceti infettati sperimentalmente con l’agente della TME10 edi pazienti affetti da sCJD63; in quest’ultimo caso, tuttavia,va tenuto in considerazione che la concentrazione stimata diPrPSc nell’EO è risultata essere piuttosto bassa (circa il 3%rispetto a quella del bulbo olfattivo). Come è noto i neuritidei neuroni olfattivi, contrariamente a tutte le altre fibrenervose, presentano elevati livelli di PrPC 44. Al contrario diquanto ci si potrebbe aspettare, e tuttavia in accordo conprecedenti riscontri28, l’esame immunoistochimico applica-to nel nostro studio secondo il metodo in precedenza de-scritto non ha evidenziato alcun deposito di PrPSc nei nerviolfattivi in nessuno degli animali esaminati; tuttavia un se-gnale di marcatura, in precedenza mai descritto, è apparso alivello del loro perinevrio.La detezione di PrPSc nel PNO, pressoché esclusivamente a li-vello della lamina cribrosa, può essere spiegata considerandol’organizzazione ultrastrutturale delle guaine dei nervi olfat-tivi in quell’area specifica (Fig. 5). Nella pecora in particola-re mancano ancora studi sull’ultrastruttura del PNO a livel-lo della lamina cribrosa, ma tali descrizioni sono disponibiliin letteratura in merito al coniglio13 e al topo12,60. In base aquesti dati risulta che i nervi olfattivi sono avvolti per tutta laloro lunghezza dalle olfactory ensheathing cells che accom-pagnano nel loro tragitto i nervi olfattivi verso il bulbo12, ve-nendo a costituire lo strato più interno del PNO. La presen-za di una lamina basale separa le olfactory ensheathing cellsda uno strato connettivale più esterno, direttamente appostoal precedente, costituito da fibre collagene e fibroblasti, che siaccolla anch’esso strettamente ai fasci di nervi olfattivi lungoil loro tragitto.Alla zona di transizione fra il sistema nervoso periferico ecentrale, dove i fila olfactoria attraversano la lamina cribrifor-me, al perinevrio si aggiunge un’ulteriore componente di na-tura meningea, costituita dalla pia madre e dall’aracnoide. Ledue meningi si estendono attraverso i fori della lamina cri-brosa apponendosi ai nervi olfattivi, e delimitando in tal mo-do uno spazio perineurale virtuale che è una diretta prosecu-zione dello spazio subaracnoideo encefalico. Questo spazio,compreso fra le membrane della pia madre e dell’aracnoide,contiene liquido cefalo-rachidiano e si estende lungo il de-corso dei nervi olfattivi fino a che le due meningi si fondono
Figura 5 - Diagramma schematico della sezione trasversale di unnervo olfattivo a livello della lamina cribrosa dell’etmoide, che mo-stra gli strati costituenti il suo perinevrio a quel livello. Blu, fila ol-factoria; magenta, olfactory ensheathing cells; verde, endonevrio difibroblasti e fibre collagene; azzurro (pia madre), bianco (spazio pe-rineurale) e rosso (membrana aracnoidea), componenti che formanol’epitelio perineurale; giallo, epinevrio. La natura cellulare è comunea tutti gli strati sopra menzionati; tuttavia, anche una componenteacellulare di fibre collagene prende parte alla costituzione di epine-vrio ed endonevrio.
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 28

C. Porcario et al. Large Animal Review 2010; 16: 21-32 29
Figura 6 - Principali pattern di deposizio-ne della PrPSc riscontrati nelle aree encefali-che relazionate al SO delle pecore affette dascrapie (A, C, E, G, I e K) e dei controllinegativi (B, D, F, H, J e L) esaminati. (A)Marcatura di tipo finemente puntato (+ + +)all’interno dei glomeruli del bulbo olfattivoaccessorio (× 4); (C) recesso olfattorio (RO)del bulbo olfattivo principale che mostra unmarcato pattern subependimale (× 40); (E)tratto olfattivo con moderati depositi di Pr-PSc di tipo submeningeo e gliale; (G) mar-cato pattern di deposizione della PrPSc di ti-po perivascolare nella corteccia frontale a li-vello dei nuclei della base (× 40); (I) mode-rati depositi di PrPSc di tipo submeningeo egliale nel lobo piriforme (× 10); (K) lieve pat-tern di tipo subependimale nell’ippocampo(× 10). (B) Sezioni di tessuto di pecore uti-lizzate come controlli negativi che mostranoassenza di marcatura per PrPSc a livello dibulbo olfattivo accessorio (× 4); (D) recessoolfattivo del bulbo olfattivo principale (× 40);(F) tratto olfattivo (× 10); (H) corteccia fron-tale (× 40); (J) lobo piriforme (× 10); (L) ip-pocampo (× 10).
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 29

30 Coinvolgimento del sistema olfattivo in corso di scrapie naturale
insieme con il perinevrio12,13,60. Pertanto la via olfattiva risul-ta essere, oltre ai villi aracnoidei e alle granulazioni di Pac-chioni nel cervello, un’ulteriore via di drenaggio, a livello disottomucosa nasale, del liquido cefalo-rachidiano, come pre-cedentemente si è già avuto modo di dimostrare in molte spe-cie, fra cui anche quella ovina27,62.Nel corso del presente studio la marcatura per PrPSc a livellodel PNO è apparsa esclusivamente a livello della lamina cri-brosa e nelle adiacenti porzioni della CNM dove essa prendeinserzione. Più in dettaglio, eseguendo uno studio di doppiaIF, abbiamo dimostrato la co-localizzazione della PrPSc con lecitocheratine, accertando così la componente meningea delPNO quale vero sito di deposizione della PrPSc. Questa par-ticolare localizzazione anatomica sembra molto verosimil-mente indicare un probabile ruolo del liquido cefalo-rachi-diano nel trasporto di PrPSc alle cellule meningee che si af-facciano sull’estensione dello spazio subdurale circondantele radici dei nervi. Ad oggi, soltanto la PrPC è stata riscontra-ta nel liquido cefalo-rachidiano della pecora39,58; tuttavia, cisono buone evidenze che anche la PrPSc vi possa essere pre-sente, come è già stato dimostrato mediante cromatografialiquida ad alta pressione (HPLC)2 e PMCA1 nel liquido cefa-lo-rachidiano rispettivamente di topi e criceti sperimental-mente infettati con scrapie.I nostri riscontri sono molto simili a quelli descritti in unostudio con traccianti condotto da Walter e coll.60, che avevaprevisto l’inoculazione di inchiostro nello spazio subaracnoi-deo di topi sperimentali. A livello del PNO l’inchiostro erastato riscontrato tramite microscopia elettronica all’internodelle cellule aracnoidee, ma non nelle cellule della dura ma-dre né nei fibroblasti. Il solo coinvolgimento delle cellulearacnoidee sia nella deposizione di PrPSc che dell’inchiostro,ad esclusione di ogni altro tipo di cellula contigua, potrebbeessere ricondotto al tipo di giunzione intercellulare fra di es-se presente. Le giunzioni fra i fibroblasti del PNO, benché no-tevolmente distanziate fra di loro, sono di tipo tight junc-tion13, e per loro natura limitano i passaggi di sostanze attra-verso gli spazi intercellulari. Inoltre, la presenza di una lami-na basale intorno alle olfactory ensheathing cells a ridosso deinervi olfattivi è un ulteriore aspetto che potrebbe fungere dafattore limitante per la captazione di molecole, fra cui la Pr-PSc, da parte dei neuroni olfattivi sottostanti.Nonostante i nervi olfattivi e la maggior parte delle cellule co-stituenti il loro perinevrio non sembrino essere permissive al-la deposizione di PrPSc, i capillari linfatici della sottomucosanasale e il NALT sono vie ben note per il drenaggio del liqui-do cefalo-rachidiano e le molecole che esso può veicolare. Nel-la pecora è stato infatti calcolato che un ammontare conside-revole del liquido cefalo-rachidiano (dal 40 al 48%) drena neivasi linfatici del NALT e da questi poi nei linfonodi retrofarin-gei e cervicali5. Benché questi linfonodi non siano stati esami-nati nel presente studio, la PrPSc è stata rinvenuta nel NALT in4 delle 24 pecore esaminate. Sembrerebbe lecito ipotizzare,pertanto, un probabile ruolo del liquido cefalo-rachidiano neldeterminare il coinvolgimento del NALT in corso di scrapie.Inoltre, il fatto di aver riscontrato uno stato infiammatorio as-sociato alla presenza di larve di Oestrus ovis nella mucosa na-sale di 2 delle 4 pecore il cui NALT risultava positivo per la pre-senza di PrPSc (dati non pubblicati) è a favore dell’ipotesi cheuno stato infiammatorio concomitante alla scrapie possa por-tare alla diffusione di PrPSc in siti di deposizione inusuali22. Inquest’ottica la presenza di PrPSc nel NALT potrebbe anche es-
sere dovuta ad un’infiammazione a carico della mucosa nasa-le, come appunto si verifica in caso di miasi.In conclusione i dati da noi ottenuti confermano il coinvol-gimento del NALT in corso di scrapie naturale, come prece-dentemente riportato in corso di CWD naturale53 e di TMEsperimentale3,10.L’ipotesi di un probabile ruolo del liquido cefalo-rachidianonel garantire la neuroinvasione da parte della PrPSc in corsodi scrapie viene anche supportata dai risultati dell’analisi im-munoistochimica condotta sui cervelli delle pecore esamina-te in questo studio. Nelle aree encefaliche in relazione al SOla deposizione di PrPSc appariva frequentemente in quellearee a diretto contatto col liquido cefalo-rachidiano, presen-tandosi soprattutto con pattern di tipo subependimale, peri-vascolare e submeningeo. È da notare come questi patternnon fossero specifici per le aree relazionate al SO, in quantoriscontrati a livello dell’intero encefalo, in animali allo stadiosia clinico che preclinico. Questi dati rafforzano ulterior-mente l’ipotesi di un probabile ruolo del liquido cefalo-ra-chidiano nel trasporto della PrPSc all’intero cervello.Altri autori hanno elaborato ipotesi sul possibile ruolo del si-stema acqueduttale encefalico come via alternativa al sistemanervoso autonomo nel fenomeno della neuroinvasione2,33,50.In linea con precedenti segnalazioni14, abbiamo osservato ladeposizione di PrPSc nelle regioni supra e subependimali; ciòindica che gli ependimociti sono suscettibili all’agente dellascrapie e hanno probabilmente un ruolo nello scambio di Pr-PSc fra il parenchima cerebrale e il liquido cefalo-rachidiano.A dimostrazione di una possibile relazione fra il liquido ce-falo-rachidiano e il pattern di deposizione della PrPSc vi è lacostante identificazione di un pattern perivascolare nelle areerelazionate al SO che abbiamo esaminato. La particolare ul-trastruttura delle arterie del cervello è tale per cui lo spazioperivascolare delle arterie intracerebrali è in diretta conti-nuità con quello delle arterie subaracnoidee64. Tale architet-tura consente al liquido interstiziale encefalico di essere dre-nato nello spazio perivascolare o anche molto verosimilmen-te di infiltrarsi, in misura ancora non nota, nel liquor dellospazio subaracnoideo. Come osservato in alcuni animali spe-rimentali37,61,65, la comunicazione esistente fra il liquido in-terstiziale e il liquor potrebbe rappresentare una via di diffu-sione di diverse molecole e traccianti nel tessuto cerebrale,verosimilmente anche della PrPSc.La frequente identificazione di un pattern di deposizionesubmeningeo della PrPSc, senza il coinvolgimento delle sotto-stanti aree della corteccia cerebrale, potrebbe essere attribui-to a un possibile trasporto della PrPSc da parte del liquor al-l’interno del cervello. Questa ipotesi resta ancora da chiarire;tuttavia, precedenti studi hanno dimostrato la permeabilitàdella pia madre ad alcuni soluti, molecole ed anche enzimiquali, ad esempio, la perossidasi64.Riguardo al coinvolgimento delle aree cerebrali relazionate alSO, la deposizione di PrPSc, in tutte le pecore esaminate, ap-pariva principalmente a livello delle strutture più frontali(cioè bulbo olfattivo, tratto olfattorio e corteccia frontale),che costituiscono le zone primarie di afferenza dei neuroni ol-fattivi. Al contrario, le aree di afferenza delle fibre olfattive disecondo e terzo ordine, ovvero, rispettivamente, il lobo pi-riforme e l’ippocampo, hanno mostrato solo una debole mar-catura per PrPSc. Questo dato non esclude la possibilità di untrasporto anterogrado della PrPSc attraverso le fibre olfattive,benché questa non sia mai stata riscontrata né in esse né nel-
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 30

l’epitelio olfattivo. È necessario sottolineare che la mancatadetezione di PrPSc nelle fibre olfattive potrebbe essere dovutaal loro elevato fisiologico turnover, che, tuttavia, potrebbenon precludere il trasporto verso il cervello della PrPSc. Infat-ti, l’immunoreattività per PrPSc osservata nello strato delle fi-bre olfattive sembrerebbe essere a sostegno di questa ipotesi.Nelle pecore esaminate in questo studio l’intensità di marca-tura della PrPSc nelle aree olfattive encefaliche è sempre ap-parsa, sia in animali con sintomi clinici che in fase preclini-ca, da poco a molto più debole di quella evidenziata negliobex corrispondenti a livello del nucleo motore dorsale delvago. Pertanto non può essere esclusa l’ipotesi che la neu-roinvasione possa aver avuto luogo dal tratto gastrointesti-nale attraverso la già ben nota via del nervo vago.Non è ancora noto a quale delle tre vie ipotizzate, ovvero se (i)a quella mediata dal liquor, (ii) a quella mediata dal sistemalinfatico o (iii) a quella associata ai nervi (o, ancora, se adun’interazione fra tutte queste) sia da ascrivere la deposizionedi PrPSc nel SO nel corso delle encefalopatie spongiformi tra-smissibili acquisite per via naturale. Ciononostante, la quan-tità relativamente elevata di PrPSc (1 x 10–1 di quella presente alivello encefalico) quantificata nei tessuti delle cavità nasalidelle pecore esaminate suggerisce un loro coinvolgimento pre-ponderante nella patogenesi della scrapie.La presenza di PrPSc in mucosa nasale è un forte elemento asostegno della sua possibile diffusione nell’ambiente attra-verso il muco nasale e parassiti come le larve di Oestrus ovis8.La fattibilità della trasmissione orizzontale della scrapie nel-la popolazione ovina è anche supportata dal riscontro speri-mentale che il SO rappresenta una valida via di neuroinva-sione in questa specie18. Si potrebbero intraprendere ulterio-ri studi per stabilire possibili differenze dovute al genotipo, alceppo di scrapie e per determinare il momento dell’insor-genza della deposizione della PrPSc nei tessuti della cavità na-sale della pecora. In ragione dei dati ottenuti al momento at-tuale, vi sono forti evidenze che indicano che il SO sia pre-cocemente coinvolto nella patogenesi della scrapie, persinonegli stadi precoci della malattia quando gli animali ancoraappaiono asintomatici. I nostri risultati ben si correlano conquelli riportati in merito ad altre malattie neurodegenerativedell’uomo, in particolare l’Alzheimer ed il Parkinson, nelcorso della cui patogenesi, in molti casi, già è stato accertatocon sicurezza il coinvolgimento del SO11.
RINGRAZIAMENTI
Gli autori ringraziano il Dott. Danilo Muratore e la Dott.ssaPatrizia Davico del Presidio Multizonale Profilassi Polizia Ve-terinaria dell’ASL TO 1 per la gentile assistenza tecnica for-nita durante i campionamenti. Questo studio è stato finan-ziato da fondi per la ricerca erogati dal Ministero della Salu-te (IZSPLV2003RF e IZSPLV20/04) e dalla Regione Piemon-te (Ricerca scientifica applicata 2004 - B44).
❚ Olfactory system involvement in natural scrapie disease
SUMMARYThe olfactory system (OS) involvement in transmissiblespongiform encephalopathies has lately been acknowledged
in experimental studies: prion spread to the nasal mucosa isknown to occur in hamsters after intracerebral challenge,and olfactory neurons have been assessed as a route for prionneuroinvasion too.This study investigated whether the OS is involved in natu-rally occurring prion diseases. Samples of nasal mucosataken at the level of medial nasal concha, ventral nasal con-cha and nasal septum from 24 natural scrapie affected sheepwere examined by immunohistochemistry (IHC), immuno-fluorescence (IF), PET blot and Western Blot (WB) for scra-pie prion protein (PrPSc). OS related brain areas of the selec-ted sheep (olfactory bulb, olfactory tract, frontal cortex, py-riform lobe and hippocampus) were analyzed too.Prion spread was assessed both in peripheral and central OSof the examined sheep. Twenty one samples of olfactory mu-cosa were positive by WB; IHC confirmed WB positive re-sults in 14 cases. PrPSc was mainly localized in the medial na-sal concha at the level of the olfactory nerve perineurium,and it was also disclosed in the nasal associated lymphoid tis-sue. It was never detected in olfactory neurons and fibers. Inthe brain PrPSc staining intensity was higher in olfactory bulband frontal cortex, where it appeared as submeningeal, sube-pendymal and perivascular patterns.The finding of PrPSc both in the brain areas directly in con-tact to the cerebrospinal fluid (CSF) and in the olfactory ner-ve perineurium bounding the subdural space extension thatsurrounds nerve rootlets might be consistent with the recen-tly discovered PrPSc presence in CSF.
KEY WORDSSheep, scrapie, olfactory system, PrPSc, perineurium.
Bibliografia
1. Atarashi, R., R.A. Moore, V.L. Sim, A.G. Hughson, D.W. Dorward, H.A.Onwubiko, S.A. Priola, and B. Caughey. 2007. Ultrasensitive detectionof scrapie prion protein using seeded conversion of recombinant prionprotein. Nat. Methods 4:645-650.
2. Banks, W.A., M.L. Niehoff, C. Adessi, and C. Soto. 2004. Passage of mu-rine scrapie prion protein across the mouse vascular blood-brain bar-rier. Biochem. Biophys. Res. Commun. 318:125-130.
3. Bartz, J.C., A.E. Kincaid, and R.A. Bessen. 2003. Rapid prion neuroin-vasion following tongue infection. J. Virol. 77:583-591.
4. Bonfanti, L., P. Peretto, S. De Marchis, and A. Fasolo. 1999. Carnosi-ne related dipeptides in the mammalian brain. Prog. Neurobiol. 59:333-353.
5. Boulton, M., M. Flessner, D. Armstrong, J. Hay, and M. Johnston. 1998.Determination of volumetric cerebrospinal fluid absorption into extra-cranial lymphatics in sheep. Am. J. Physiol. 274:R88-R96.
6. Brown, P., and D.C. Gajdusek. 1991. Survival of scrapie virus after 3years’ internment. Lancet 337:269-270.
7. Casalone, C., G. Zanusso, P. Acutis, S. Ferrari, L. Capucci, F. Tagliavini,S. Monaco, and M. Caramelli. 2004. Identification of a second bovineamyloidotic spongiform encephalopathy: molecular similarities withsporadic Creutzfeldt-Jacob disease. Proc. Natl. Acad. Sci. USA101:3065-3070.
8. Corona, C., F. Martucci, B. Iulini, M. Mazza, P.L. Acutis, C. Porcario, M.Pezzolato, B. Manea, A. Maroni, S. Barocci, E. Bozzetta, M. Caramelli,and C. Casalone. 2006. Could Oestrus ovis act as vector for scrapie?, ab-str. PA-10, p. 239. Abstr. Prion 2006: strategies, advances and trendstowards protection of society, Turin, Italy.
9. Cowan, C.M., and A.J. Roskams. 2002. Apoptosis in the mature and de-veloping olfactory neuroepithelium. Microsc. Res. Tech. 58:204-215.
10. DeJoia, C., B. Moreaux, K. O’ Connell, and R.A. Bessen. 2006. Prion in-fection of oral and nasal mucosa. J. Virol. 80:4546-4556.
11. Doty, R.L. 2008. The olfactory vector hypothesis of neurodegenerativedisease: is it viable? Ann. Neurol. 63:7-15.
12. Doucette, R. 1991. PNS-CNS transitional zone of the first cranial ner-ve. J. Comp. Neurol. 312:451-466.
C. Porcario et al. Large Animal Review 2010; 16: 21-32 31
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 31

32 Coinvolgimento del sistema olfattivo in corso di scrapie naturale
13. Erlich, S.S., J.G. McComb, S. Hyman, and M.H. Weiss. 1986. Ultra-structural morphology of the olfactory pathway for cerebrospinal fluiddrainage in the rabbit. J. Neurosurg. 64:466-473.
14. Fournier, J.G., K. Adjou, V. Grigoriev, and J.P. Deslys. 2008. Ultrastruc-tural evidence that ependymal cells are infected in experimental scra-pie. Acta Neuropathol. 115:643-650.
15. Frank, E.H., B.W. Burge, B.H. Liwnicz, L.J. Lotspeich, J.C. White, S.L.Wechsler, F.H. Mayfield, and J.T. Keller. 1983. Cytokeratin provides aspecific marker for human arachnoid cells grown in vitro. Exp. CellRes. 146:371-376.
16. Franke, W.W., D. Mayer, E. Schmid, H. Denk, and E. Borenfreund. 1981.Differences of expression of cytoskeletal proteins in cultured rat hepa-tocytes and hepatoma cells. Exp. Cell Res. 134:345-365.
17. Gil, D.R., and G.D. Ratto. 1973. Contribution to the study of the originof leptomeninges in the human embryo. Acta Anat. 85:620-623.
18. Hamir, A.N., R.A. Kunkle, J.A. Richt, J.M. Miller, and J.J. Greenlee. 2008.Experimental transmission of US scrapie agent by nasal, peritoneal, andconjunctival routes to genetically susceptible sheep. Vet. Pathol. 45:7-11.
19. Harmeyer, S., E. Pfaff, and M.H. Groschup. 1998. Synthetic peptidevaccines yield monoclonal antibodies to cellular and pathologicalprion proteins of ruminants. J. Gen. Virol. 79:937-945.
20. Harvey, S.C., and H.S. Burr. 1926. The development of the meninges.Arch. Neurol. Psychiat. 15:545-567.
21. Haybaeck, J., M. Heikenwalder, I. Margalith, N. Zeller, C. Bridel, P.Schwarz, K. Merz, L. Stitz, and A. Aguzzi. 2007. Intranasal and aereoso-lic prion transmission, abstr. P03.103, p. 72. Abstr. Prion 2007, Edin-burgh, Scotland, United Kingdom.
22. Heikenwalder, M., N. Zeller, H. Seeger, M. Prinz, P.C. Klohn, P. Schwarz,N.H. Ruddle, C. Weissman, and A. Aguzzi. 2005. Chronic lymphocytic in-flammation specifies the organ tropism of prions. Science 307:1107-1110.
23. Jackson, R.T., J. Tigges, and W. Arnold. 1979. Subarachnoid space of theCNS, nasal mucosa, and lymphatic system. Arch. Otolaryngol. 105:180-184.
24. Jeffrey, M., S. Martin, L. González, S. J. Ryder, S.J. Bellworthy, and R.Jackman. 2001. Differential diagnosis of infections with the bovinespongiform encephalopathy (BSE) and scrapie agents in sheep. J.Comp. Pathol. 125:271-284.
25. Johnson, C.J., K.E. Phillips, P.T. Schramm, D. McKenzie, J.M. Aiken,and J.A. Pedersen. 2006. Prions adhere to soil minerals and remain in-fectious. PloS Pathog. 2:296-302.
26. Johnson, R.T. 2005. Prion diseases. Lancet Neurol. 4:635-642.27. Johnston, M., A. Zakharov, C. Papaiconomou, G. Salmasi, and D. Arm-
strong. 2004. Evidence of connections between cerebrospinal fluid andnasal lymphatic vessels in humans, non-human primates and othermammalian species. Cerebrospinal Fluid Res. 1:2.
28. Kincaid, A.E., and J.C. Bartz. 2007. The nasal cavity is a route for prioninfection in hamsters. J. Virol. 81:4482-4491.
29. Klika, E. 1968. Ultrastructure of the meninges during human ontogeny.Z. Mikrosk. Anat. Forsch. 79:209-222.
30. Kolb, A. 1979. Lichtmikroskopische Untersuchungen an der Riechsch-leimhaut des Hausschafes (Ovis aries L.). Anat. Anz. 146:444-455.
31. Konold, T., S.J. Moore, S.J. Bellworthy, and H.A. Simmons. 2008. Evi-dence of scrapie transmission via milk. BMC Vet. Res. 4:14.
32. Lazarides, E. 1980. Intermediate filaments as mechanical integrators ofcellular space. Nature 283:249-256.
33. Marsh, R.F., and R.H. Kimberlin. 1975. Comparison of scrapie andtransmissible mink encephalopathy in hamsters. II. Clinical signs,pathology and pathogenesis. J. Infect. Dis. 131:104-110.
34. Mathiason, C.K., J.G. Powers, S.J. Dahmes, D.A. Osborn, K.V. Miller,R.J. Warren, G.L. Mason, S.A. Hays, J. Hayes-Klug, D.M. Seelig, M.A.Wild, L.L. Wolfe, T.R. Spraker, M.W. Miller, C.J. Sigurdson, G.C. Telling,and E.A. Hoover. 2006. Infectious prions in the saliva and blood of deerwith chronic wasting disease. Science 314:133-136.
35. Miller, M.W., and E.S. Williams. 2003. Prion disease: horizontal priontransmission in mule deer. Nature 425:35-36.
36. Mori, I., Y. Nishiyama, T. Yokochi, and Y. Rimura. 2005. Olfactory tran-smission of neurotropic viruses. J. Neurovirol. 11:129-137.
37. Nicholson, C., and E. Syková. 1998. Extracellular space structure revea-led by diffusion analysis. Trends Neurosci. 21:207-215.
38. Pease, D.C., and R.L. Schultz. 1958. Electron microscopy of rat cranialmeninges. Am. J. Anat. 102:301-321.
39. Picard-Hagen, N., V. Gayrard, C. Viguie, M. Moudjou, C. Imbs, and P.L.Toutain. 2006. Prion protein in the cerebrospinal fluid of healthy andnaturally scrapie-affected sheep. J. Gen. Virol. 87:3723-3727.
40. Prusiner, S.B. 1982. Novel proteinaceus infectious particles cause scra-pie. Science 216:136-144.
41. Prusiner, S.B. 1998. The prion diseases. Brain Pathol. 8:499-513.42. Race, R., A. Jenny, and D. Sutton. 1998. Scrapie infectivity and protei-
nase K-resistant prion protein in sheep placenta, brain, spleen andlymph node: implications for transmission and antemortem diagnosis.J. Infect. Dis. 178:949-953.
43. Ritchie, D.L., M.W. Head, and J.W. Ironside. 2004. Advances in the de-tection of prion protein in peripheral tissues of variant Creutzfeldt-Jakob disease patients using paraffin-embedded tissue blotting. Neuro-pathol. Appl. Neurobiol. 30:360-368.
44. Sales̀, N., K. Rodolfo, R. Hässig, B. Faucheux, L. Di Giamberardino, andK.L. Moya. 1998. Cellular prion protein localization in rodent and pri-mate brain. Eur. J. Neurosci. 10:2464-2471.
45. Salman, M.D. 2003. Chronic wasting disease in deer and elk: scientificfacts and findings. J. Vet. Med. Sci. 65:761-768.
46. Sawcer, S.J., G.M. Yuill, T.F.G. Esmonde, P. Estibeiro, J.W. Ironside, J.E.Bell, and R.G. Will. 1993. Creutzfeldt-Jacob disease in an individual oc-cupationally exposed to BSE. Lancet 341:642.
47. Schaefer, M.L., B. Böttger, W.L. Silver, and T.E. Finger. 2002. Trigeminalcollaterals in the nasal epithelium and olfactory bulb: a potential routefor direct modulation of olfactory information by trigeminal stimuli. J.Comp. Neurol. 444:221-226.
48. Seeger, H., M. Heikenwalder, N. Zeller, J. Kranich, P. Schwarz, A. Gaspert,B. Seifert, G. Miele, and A. Aguzzi. 2005. Coincident scrapie infection andnephritis lead to urinary prion excretion. Science 310:324-326.
49. Shaw, I. C. 1995. BSE and farmworkers. Lancet 346:1365.50. Sisó, S., M. Jeffrey, H. Baird, S. Martin, S. Bellworthy, F. Chianini, and
L. Gonzalez. 2006. The topographical distribution of prion protein inthe brains of sheep challenges the current hypothesis of neuroinvasion,abstr. PA-52, p. 281. Abstr. Prion 2006: Strategies, advances and trendstowards protection of society, Turin, Italy.
51. Smith, P.E.M., M. Zeidler, J.W. Ironside, P. Estibeiro, and T.H. Moss.1995. Creutzfeldt-Jacob disease in a dairy farmer. Lancet 346:898.
52. Spraker, T.R., K.I. O’ Rourke, A. Balachandran, R.R. Zink, B.A. Cum-mings, M.W. Miller, and B.E. Powers. 2002. Validation of monoclonalantibody F99/97.6.1 for immunohistochemical staining of brain andtonsil in mule deer (Odocoileus hemionus) with chronic wasting di-sease. J. Vet. Diagn. Investig. 14:3-7.
53. Spraker, T.R., R.R. Zink, B.A. Cummings, M.A. Wild, M.W. Miller, andK.I. O’ Rourke. 2002. Comparison of histological lesions and immu-nohistochemical staining of proteinase-resistant prion protein in a na-turally occurring spongiform encephalopathy of free-ranging muledeer (Odocoileus hemionus) with those of chronic wasting disease ofcaptive mule deer. Vet. Pathol. 39:110-119.
54. Stanley, A.C., J.F. Huntley, M. Jeffrey, and D. Buxton. 2001. Characteri-zation of ovine nasal-associated lymphoid tissue and identification ofM cells in the overlying follicle-associated epithelium. J. Comp. Pathol.125:262-270.
55. Tabaton, M., S. Monaco, M.P. Cordone, M. Colucci, G. Giaccone, F. Ta-gliavini, and G. Zanusso. 2004. Prion deposition in olfactory biopsy ofsporadic Creutzfeldt-Jacob disease. Ann. Neurol. 55:294-296.
56. Treloar, H.B., J.C. Bartolomei, B.W. Lipscomb, and C.A. Greer. 2001.Mechanisms of axonal plasticity: lessons from the olfactory pathway.Neuroscientist 7:55-63.
57. Vascellari, M., R. Nonno, F. Mulinelli, M. Bagolaro, M.A. Di Bari, E.Melchiotti, S. Marcon, C. D’Agostino, G. Vaccai, M. Conte, L. De Gros-si, F. Rosone, F. Giordani, and U. Agrimi. 2007. PrPsc in salivary glandsof scrapie affected sheep. J. Virol. 81:4872-4876.
58. Vella, L.J., D.L.V. Greenwood, R. Cappai, J.P.Y. Scheerlinck, and A.F.Hill. 2008. Enrichment of prion protein in exosomes derived from ovi-ne cerebral spinal fluid. Vet. Immunol. Immunopathol. 124:385-393.
59. Vidal, E., M. Ordonez, J.M. Vela, and M. Pumarola. 2003. Study of theolfactory pathway and related structures of 19 BSE naturally affectedcows, abstr. PG-26, p. 203. Abstr. Prion 2003: prion diseases: from ba-sic research to intervention concepts, Gasteig, Munich, Germany.
60. Walter, B.A., V.A. Valera, S. Takahashi, and T. Ushiki. 2006. The olfac-tory route for cerebrospinal fluid drainage into the peripheral lympha-tic system. Neuropathol. Appl. Neurobiol. 32:388-396.
61. Weller, R.O. 1998. Pathology of cerebrospinal fluid and interstitial fluidof the CNS: significance for Alzheimer disease, prion disorders andmultiple sclerosis. J. Neuropathol. Exp. Neurol. 57:885-894.
62. Zakharov, A., C. Papaiconomou, J. Djenic, R. Midha, and M. Johnston.2003. Lymphatic cerebrospinal fluid absorption pathways in neonatalsheep revealed by subarachnoid injection of Microfil. Neuropathol. Ap-pl. Neurobiol. 29:563-573.
63. Zanusso, G., S. Ferrari, F. Cardone, P. Zampieri, M. Gelati, M. Fiorini,Farinazzo, A., M. Gardiman, T. Cavallaro, M. Bentivoglio, P.G. Rigetti,M. Pocchiari, N. Rizzato, and S. Monaco. 2003. Detection of pathologicprion protein in the olfactory epithelium in sporadic Creutzfeldt-Jacobdisease. N. Engl. J. Med. 348:711-719.
64. Zhang, E.T., C.B.E. Inman, and R.O. Weller. 1990. Interrelationships ofthe pia mater and the perivascular (Virchow-Robin) spaces in the hu-man cerebrum. J. Anat. 170:111-123.
65. Zhang, E.T., H.K. Richards, S. Kida, and R.O. Weller. 1992. Directionaland compartimentalised drainage of interstitial fluid and cerebrospinalfluid from the rat brain. Acta Neuropathol. 83:233-239.
Porcario_imp:Porcario_imp 26-01-2010 15:38 Pagina 32