“Sperimenta il BioLab” Da un albero genealogico al DNA e ... · Tab.1.1 Classificazione delle...
15
“Sperimenta il BioLab” Da un albero genealogico al DNA e ritorno Università degli Studi di Milano
Transcript of “Sperimenta il BioLab” Da un albero genealogico al DNA e ... · Tab.1.1 Classificazione delle...

“Sperimenta il BioLab”
Da un albero genealogico al DNA e ritorno
Università degli Studi di Milano

2
Indice 1. Conoscenze propedeutiche p. 3
• 1.1 Le mutazioni p. 3 • 1.2 Mutazioni geniche • 1.3 Alcuni concetti base e terminologia essenziale riguardanti le malattie
monogeniche e le leggi di Mendel p. 4
2. Malattie genetiche monogeniche e analisi del pedigree p. 7 • 2.1 Che cosa è il pedigree p. 7 • 2.2 Principali modalità di trasmissione di malattie o caratteri monogenici p. 8
2.2.1 Trasmissione autosomica recessiva p. 8 2.2.2 Trasmissione autosomica dominante p. 8 2.2.3 Trasmissione di caratteri recessivi associati al cromosoma X p. 8 2.2.4 Trasmissione di caratteri dominanti associati al cromosoma X p. 9 2.2.5 Trasmissione di caratteri associati al cromosoma Y p. 9
• 2.3 La banca dati OMIM p. 9 • 2.4 Eterogeneità genetica p.10 • 2.5 Pleiotropia p.10
3. I polimorfismi di restrizione (RFLP) e il loro utilizzo nella
diagnostica delle malattie genetiche p.11
• 3.1 Gli enzimi di restrizione p.11 • 3.2 I polimorfismi del DNA p.12
• 3.3 I polimorfismi di sequenza del DNA p.12
• 3.4 I polimorfismi di lunghezza dei frammenti di restrizione (RFLP) p.12 3.4.1 La tecnica della PCR per l’analisi degli RFLP p.13
• 3.5 I polimorfismi di restrizione (RFLP) nella diagnosi delle malattie
genetiche p.14 3.5.1 Utilizzo di un polimorfismo di restrizione (RFLP) nella diagnosi diretta.
Diagnosi di anemia falciforme p.14 3.5.2 Utilizzo di un polimorfismo di restrizione (RFLP) nella diagnosi indiretta.
Ricerca di un RFLP associato a un allele malattia p.15 4. Sano o malato? p.16
• 4.1 Schede informative di 3 malattie monogeniche p.16 (anemia falciforme, ipercolesterolemia familiare, emofilia A) • 4.2 A ciascuna storia il suo pedigree. Scenari e simulazione di consulenza genetica p.20
5. Tecniche utilizzate in laboratorio p.23
• 5.1 PCR p.23 5.1.1 termociclatori p.23 5.2.2 Taq polimerasi p.24 5.2.3 scelta dei primer p.24
• 5.2 Digestione con gli enzimi di restrizione p.24 • 5.3 Elettroforesi su gel di agarosio p.24 • 5.4 Strumentazione e materiale a disposizione p.25 • 5.5 Principali prefissi e unità di misura usate in biologia molecolare e cellulare p.25
6. Dal pedigree al DNA e ritorno p.26
• 6.1 Schema delle PCR e digestione con enzimi di restrizione p.26 • 6.2 Elettroforesi su gel di agarosio. Soluzioni e reagenti p.27 • 6.3 Preparazione del gel di agarosio p.27 • 6.4 Corsa elettroforetica p.28 • 6.5 Analisi dei risultati p.29 • 6.6 Consulenza genetica p.30
7. Norme di sicurezza in laboratorio p.30
8. Test di autovalutazione p.31
9. Glossario p.36
10. Siti web p.38 11. Concorso "Una settimana da ricercatore" p.38

3
1. Conoscenze propedeutiche 1.1 Le mutazioni Nella specie umana esistono 23 coppie di cromosomi che contengono circa 24.000 geni. Le malattie genetiche sono causate da mutazioni, cioè da alterazioni del corredo genetico (genoma) di un individuo. • Se le mutazioni avvengono nelle cellule che formano i gameti, cioè nelle cellule germinali, allora
possono essere trasmesse alla discendenza dove saranno presenti in ogni cellula dell’individuo affetto. Queste mutazioni possono sorgere come mutazioni nuove nei gameti di uno dei genitori oppure possono essere state ereditate dai genitori dalle generazioni precedenti.
• Viceversa le mutazioni che avvengono in altre cellule del corpo, le cellule somatiche, non sono trasmesse alle generazioni successive, ma determinano una differenza genetica tra le cellule dello stesso organismo (mosaicismo). La conseguenza può essere l’insorgenza e lo sviluppo di un cancro.
Le mutazioni possono riguardare il numero dei cromosomi (mutazioni genomiche), la struttura dei cromosomi (mutazioni cromosomiche) o i singoli geni (mutazioni geniche). 1.2 Mutazioni geniche Le mutazioni che coinvolgono singoli geni sono, di solito, il risultato della sostituzione di una base con un'altra, della delezione (perdita) o dell’inserzione di basi (Fig. 1.1). Se insorgono all’interno di un gene che codifica una proteina, queste mutazioni possono alterare la proteina prodotta. Per esempio, una sostituzione può modificare il significato di un codone e portare all’inserimento nella proteina di un amminoacido al posto di un altro (mutazione mis-senso); oppure può cambiare un codone per un amminoacido in un codone di stop provocando la fine prematura della sintesi della proteina e un prodotto proteico incompleto (mutazione non-senso); l’inserzione o la delezione di una o più basi (non multipli di tre!) portano a una sfasatura
nella lettura del codice e quindi all’alterazione della sequenza della proteina a valle della mutazione (mutazione frameshift). La conseguenza di una mutazione è quasi sempre la produzione di una proteina che non è in grado di svolgere la sua funzione biologica (ad esempio, di un enzima che non è in grado di catalizzare una reazione). In anni recenti, si è individuato un nuovo tipo di mutazione che consiste nella presenza di copie multiple (da decine a più di mille, a seconda delle diverse patologie) di una tripletta in un punto del gene (malattie da ripetizione di
triplette); nella corea di Huntington, una malattia genetica neurodegenerativa molto grave a trasmissione autosomica dominante e ad insorgenza tardiva, la tripletta ripetuta è CAG. Questa ripetizione porta ad un incremento del numero di copie dell’amminoacido glutammina nella proteina che diventa tossica per le cellule nervose. Le mutazioni all’interno delle regioni di controllo di un gene possono invece influenzarne il livello di trascrizione. Un esempio è una forma di talassemia (nota anche come anemia mediterranea) in cui vengono prodotte in quantità insufficiente le catene dell’emoglobina. Una mutazione che interessa una sequenza nucleotidica cruciale per il meccanismo di splicing (rimozione degli introni) di un trascritto primario di RNA avrà come conseguenza il blocco dello splicing o la modificazione dello schema di splicing di quel trascritto e quindi la mancata produzione del mRNA corrispondente o la produzione di un mRNA modificato.
Fig. 1.1 Tipi diversi di mutazione. Le mutazioni puntiformi (fatta eccezione per le mutazioni sinonime) e le piccole delezioni causano quasi sempre la produzione di una proteina con funzionalità alterata.

4
1.3. Alcuni concetti base e terminologia essenziale sulle malattie monogeniche e le leggi di Mendel Le malattie ereditarie monogeniche sono determinate da mutazioni in singoli geni che si trasmettono nelle famiglie secondo leggi ben definite scoperte dall’abate Gregorio Mendel nella seconda metà dell’Ottocento. Per questo motivo le malattie monogeniche o monofattoriali vengono anche chiamate “mendeliane”. In particolare bisogna ricordare che: I geni possono avere alleli diversi (varianti alternative di uno stesso gene) ossia essere polimorfici;
Gli organismi diploidi hanno due copie di ogni gene, cioè due alleli. Se i due alleli sono uguali, l’individuo è omozigote (ad es, ha genotipo AA oppure aa). Se i due alleli sono diversi, l’individuo è eterozigote (ad es, il genotipo è Aa);
La I legge di Mendel o legge dell’uniformità della prima generazione ibrida, afferma che l’incrocio tra individui della generazione parentale ciascuno omozigote per due alleli diversi di uno stesso gene (ad es, AA x aa) e che quindi differisce dall’altro genitore per una caratteristica (ad es, pelo nero o marrone), dà una progenie costituita da individui tutti identici tra loro (tutti eterozigoti; ad es, Aa);
Durante la meiosi i due alleli di un gene segregano e si distribuiscono ciascuno in un gamete aploide (II legge di Mendel, legge della segregazione). Un individuo omozigote produce un solo tipo di gamete (A oppure a) relativamente a un dato locus (la posizione occupata da un gene in un cromosoma). Un individuo eterozigote produce due tipi di gameti (A e a) in ugual quantità, cioè in rapporto (50%) ciascuno. Una buona rappresentazione grafica della segregazione si ottiene costruendo il “quadrato di Punnett” (Fig. 1.2 e 1.3);
Alleli appartenenti a geni diversi localizzati su cromosomi diversi segregano in modo indipendente (III legge di Mendel, legge della segregazione indipendente);
Allele dominante: allele che si manifesta allo stato eterozigote;
Allele recessivo: allele mascherato dall’allele dominante, che si manifesta solo allo stato omozigote;
Allele codominante: due alleli entrambi espressi allo stato eterozigote, che agiscono sul fenotipo in modi indipendenti e distinguibili. Un esempio classico si ha nel caso degli alleli IA e IB del gruppo sanguigno ABO.
L’1% circa dei neonati è affetto da una malattia genetica monogenica. La mutazione genica è presente fin dal concepimento, quindi anche alla nascita, anche se non sempre si manifesta fenotipicamente alla nascita. Alcune di queste malattie comportano conseguenze già apprezzabili nel neonato. Altre malattie monogeniche si manifestano, invece, durante le età successive della vita. La corea di Huntington, ad esempio, è una malattia genetica a insorgenza tardiva che si manifesta solo in età adulta.
Fig. 1.3 Il quadrato di Punnett illustra la modalità di trasmissione di un carattere recessivo legato al cromosoma X e i possibili genotipi dei figli di un padre malato (a sinistra) o di una madre portatrice (a destra).
Fig. 1.2 Rappresentazione della legge della segregazione dei caratteri (I legge di Mendel): nel quadrato di Punnett sono riportati i possibili genotipi dei figli di genitori eterozigoti per un carattere recessivo (sopra) e per un carattere dominante (sotto).

5
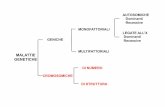
Si definisce malattia autosomica quella in cui l’allele malato è presente nelle coppie di cromosomi non coinvolti nella determinazione del sesso, gli autosomi; si parla invece di malattia legata all’X quella in cui l’allele si trova nel cromosoma sessuale X. In questo caso la trasmissione segue modalità particolari in quanto la maggior parte dei geni rappresentati sul cromosoma X non sono rappresentati sul cromosoma Y. Il cromosoma Y, infatti, è molto più piccolo e contiene quasi esclusivamente i geni per la determinazione del sesso. La Fig. 1.4 rappresenta la mappa del cariotipo umano in cui è indicata la localizzazione cromosomica dei geni responsabili di alcune malattie monogeniche autosomiche e associate al cromosoma X. Per schematizzare (vedere Tab. 1.1): • malattie autosomiche: se l’allele normale è dominante, la malattia si rende manifesta solo nella
condizione omozigote recessiva, ossia negli individui in cui entrambi gli alleli del gene sono alterati (malattia autosomica recessiva); se invece è l’allele alterato a prevalere su quello sano, la malattia si rende sempre evidente, anche nella condizione eterozigote, ossia è sufficiente che uno solo dei due alleli sia alterato per avere la malattia (malattia autosomica dominante) (Fig.1.2).
• malattie legate all’X: se l’allele è recessivo (Fig.1.3), come nella maggior parte dei casi, i maschi ricevono l’X difettoso dalla madre, ma non possono bilanciarlo con un allele sano nell’Y in quanto questo cromosoma non contiene l’allele corrispondente; le femmine invece manifestano la malattia solo nelle condizione omozigote. Di conseguenza, la frequenza delle malattie recessive legate al
cromosoma X è maggiore nei maschi che nelle femmine. Nel caso di malattie dominanti le femmine
manifestano la malattia anche nella condizione eterozigote.
Tab.1.1 Classificazione delle malattie a ereditarietà mendeliana
Dominanti Autosomiche
Recessive Dominanti
Malattie a ereditarietà mendeliana
Legate al cromosoma X Legate al cromosoma Y
Recessive
Fig. 1.4 Mappa del cariotipo umano in cui sono evidenziate alcune delle malattie monogeniche autosomiche dominanti, autosomiche recessive e legate al cromosoma X recessive.

6
Nella Tabella 1.2 sono riportate alcune delle principali malattie monogeniche di cui è indicata la frequenza nella popolazione adulta, il tipo di trasmissione, la localizzazione cromosomica e il numero del catalogo OMIM (Online Mendelian Inheritance in Man).
Tab. 1.2 Alcune malattie monogeniche di cui è indicata la frequenza nella popolazione adulta, il tipo di trasmissione, la localizzazione cromosomica e il numero del catalogo OMIM (Online Mendelian Inheritance in Man).
Malattie
monogeniche
Frequenza in popolazione adulta Tipo di
trasmissione
Localizzazione
cromosomica
Ref. OMIM
Ipercolesterolemia familiare
eterozigoti 1 / 500 Autosomica dominante
Cromosoma 19 143890
Corea di Huntington 1 / 20000 Autosomica dominante
Cromosoma 4 143100
Policistite renale dell’adulto
1 / 1000 nati vivi Autosomica dominante
Cromosoma 16 173900
Fibrosi cistica 1 / 2000 – 4000 popolazione caucasica
Autosomica recessiva
Cromosoma 7 219700
Anemia falciforme (HbS)
Molto rara in Europa, molto diffusa nella popolazione nera in Africa (1 / 400 – 600) e in America (1 / 50)
Autosomica recessiva
Cromosoma 11 141900.0243
Talassemia Molto frequente nel bacino Mediterraneo (1 / 250)
Autosomica recessiva
Cromosoma 11 catene beta; cromosoma 16 catene alfa
Alfa: 141800 Beta: 141900
Fenilchetonuria 1 / 10000 nati vivi Autosomica recessiva
Cromosoma 12 261600
Ipofosfatemia familiare
1 / 20000 X-linked dominante
Cromosoma X 307800
Emofilia A 1 / 5000 – 10000 nei maschi X-linked recessiva
Cromosoma X 306700
Daltonismo (Cecità al verde e al rosso)
8% nei maschi; 0,64% nelle femmine
X-linked recessivo
Cromosoma X 303800
Miopatia di Duchenne
1 / 6000 nei maschi X-linked recessiva
Cromosoma X 310200

7
2. Malattie genetiche monogeniche e analisi dei pedigree
2.1 Che cosa e’ il pedigree?
Uno degli aspetti della genetica è lo studio dei meccanismi con cui i geni sono trasmessi dai genitori ai figli. A tale scopo i genetisti effettuano accoppiamenti tra organismi della stessa specie, per analizzare la trasmissione dei caratteri. Nella genetica umana, gli accoppiamenti sperimentali ovviamente non sono possibili. Molte delle nostre conoscenze sull’ereditarietà dei caratteri umani derivano perciò dalla analisi degli alberi genealogici o pedigree. In pratica, il pedigree è la rappresentazione sistematica della storia familiare attraverso l’uso di simboli standardizzati (Fig.2.1). Il pedigree viene stilato partendo da un’intervista ai componenti di una famiglia, al fine di ricostruirne la storia (anamnesi); in questo modo è possibile seguire la trasmissione di un dato carattere attraverso parecchie generazioni in una data famiglia (Fig.2.2). L’analisi del pedigree permette di determinare se il carattere ha una modalità di trasmissione recessiva o dominante, e se il gene in questione è localizzato su un autosoma o su un cromosoma sessuale, permette
in ultima analisi di informare i membri di una famiglia sulla probabilità di trasmettere queste malattie ai propri figli.
Nell’analisi del pedigree ci si basa sui principi dell’eredità di Mendel per escludere le modalità di trasmissione che sono incompatibili con il pedigree. Ad esempio, la presenza nell’albero genealogico di femmine malate ci permette di escludere la trasmissione legata all’Y.
Fig.2.2. Un pedigree che mostra la trasmissione ereditaria di un carattere (i simboli scuri rappresentano gli individui affetti) attraverso numerose generazioni di una famiglia.
Fig.2.1. I simboli usati nell’analisi del pedigree adottati nel 1995 dalla Società Americana di
??!! Ma tu ce l’hai
il pedigree?

8
2.2 L’analisi del pedigree per la determinazione delle modalità di trasmissione di malattie o di
caratteri monogenici
Dall’analisi di un pedigree si può determinare (talvolta non in modo univoco, in quanto un pedigree può essere compatibile con più di una modalità di trasmissione ereditaria) come viene trasmesso un dato carattere. Analizziamo le caratteristiche dei pedigree relativi a caratteri:
autosomici recessivi (AR) autosomici dominanti (AD) legati al cromosoma X recessivi (XR) legati al cromosoma X dominanti (XD) legati al cromosoma Y
2.2.1 Eredità autosomica recessiva
• Possono essere affetti sia i maschi sia le femmine. • In genere gli individui affetti hanno genitori sani
(portatori asintomatici). • I genitori entrambi portatori sani hanno il 25% di
probabilità di avere figli malati ad ogni gravidanza. • L’incidenza della malattia aumenta in caso di
consanguineità. • Tutti i figli di genitori entrambi affetti (omozigoti)
sono a loro volta affetti.
2.2.2 Eredità autosomica dominante
• Possono essere affetti sia i maschi sia le femmine.
• Gli individui affetti possono essere presenti in tutte le generazioni.
• Gli individui affetti hanno sempre un genitore affetto.
• Poiché i geni responsabili di malattie autosomiche dominanti sono rari, in
genere gli individui affetti sono eterozigoti. Rarissimi sono gli omozigoti, che possono nascere solo da genitori entrambi eterozigoti. Ogni affetto (eterozigote) ha il 50% di probabilità di avere figli malati.
• Individui non affetti del pedigree che sposano individui non affetti non hanno discendenti affetti. • Genitori entrambi affetti (eterozigoti) possono avere figli sani (25%). • I genitori di un bambino malato possono essere sani: in questo caso si può dedurre che la malattia origina
da una nuova mutazione verificatasi durante la formazione di un gamete (mutazione “de novo”).
2.2.3 Eredità recessiva legata al cromosoma X
• La frequenza della malattia è maggiore nei maschi che nelle femmine.
• Le femmine possono essere portatrici sane. • Le donne portatrici hanno un rischio del 50% di
avere figli maschi malati. • I maschi affetti hanno figlie portatrici sane e
figli maschi sani. • La madre di un individuo affetto è portatrice
sana. • Il carattere si trasmette a zig-zag con maschi
affetti in generazioni diverse (eredità diaginica). • La metà delle figlie femmine di madri portatrici
sono anche loro portatrici.

9
2.2.4 Eredità dominante legata al cromosoma X
• Maschi affetti generano solo
femmine affette, mentre i figli maschi sono sempre sani.
• Femmine eterozigoti affette trasmettono il carattere al 50% dei figli siano essi maschi o femmine.
• La frequenza della malattia nelle femmine è solitamente doppia di quella nei maschi.
2.2.5 Eredità legata al cromosoma Y
• Sono stati individuati pochi caratteri legati al cromosoma Y.
• I caratteri si manifestano solo nei maschi
• Sono trasmessi direttamente da padre a figlio (eredità olandrica).
2.3 La banca dati OMIM
OMIM, Online Mendelian Inheritance in Man, è una banca dati che contiene informazioni sui geni umani e sulle malattie genetiche realizzata e mantenuta dall’NCBI, the National Center for Biotechnology Information. La banca contiene la descrizione dei geni e delle malattie ad essi associate, i quadri clinici e i riferimenti bibliografici, oltre a link a sequenze e ad altre risorse web. Si tratta della versione on line del testo “Mendelian Inheritance in Man”, ora alla sua dodicesima edizione, a cura del genetista medico Victor A. McKusick e di un gruppo di colleghi della Johns Hopkins University e di altre istituzioni. La banca dati è aggiornata quotidianamente e a metà 2004 conteneva oltre 15400 voci. Essa riporta solo malattie che sono state associate ad uno o più geni. La pagina di accesso ad OMIM si raggiunge tramite un link sulla homepage dell’NCBI: (http://www.ncbi.nlm.nih.gov/). Per entrare, fare click sulla parola OMIM in bianco sulla striscia blu in cima alla pagina.

10
2.4 Eterogeneità genetica
Per alcune malattie ereditarie monogeniche tutti gli individui affetti portano un’identica mutazione: l’anemia falciforme ne è un esempio. Per molte altre malattie ci sono invece più mutazioni che possono produrre lo stesso quadro patologico. Quando le diverse mutazioni sono a carico dello stesso gene si parla di eterogeneità allelica. Nel caso dell’emofilia A, ad esempio, il catalogo OMIM riporta 270 varianti alleliche. La fibrosi cistica è causata da più di 1000 mutazioni diverse nel gene CFTR. Le diverse
mutazioni possono produrre fenotipi di gravità diversa. Nel caso in cui le mutazioni avvengono in geni diversi che collaborano alla determinazione di un fenotipo, si parla di eterogeneità di locus. Ad esempio, in una stessa via metabolica, mutazioni in geni diversi possono portare alla stessa modificazione del fenotipo (Fig.2.3). E’ il caso delle due forme di ipercolesterolemia familiare A e B (vedere §4.1) dovute rispettivamente a mutazioni del gene del recettore che lega le lipoproteine LDL e del gene della apoproteina B-100 che media il legame della particella LDL al recettore.
2.5 Pleiotropia
Questo concetto sta a indicare che un solo gene può essere responsabile di fenotipi diversi. Un esempio di pleiotropia nell’uomo è quello dell’anemia falciforme, una malattia caratterizzata da diversi sintomi a carico di organi e tessuti diversi. Questi possibili effetti fenotipici derivano tutti dall’azione di un solo allele mutato presente in omozigosi. L’effetto diretto dell’allele dell’anemia falciforme è quello di indurre i globuli rossi a produrre molecole anomale di emoglobina che tendono a unirsi fra loro e cristallizzare. Di conseguenza, i globuli rossi si deformano, assumendo forme a falce con contorni frastagliati, sono più fragili e lisano e tendono ad aggregarsi e a occludere i capillari. Tutto questo produce molteplici danni a carico di organi e tessuti diversi (Fig 2.4).
Fig.2.4. La mutazione che causa l’anemia falciforme determina una cascata di effetti fenotipici a livello cellulare e di interi organi.
Fig.2.3. Esempio di uno dei meccanismi alla base dell’eterogeneità genetica di locus. È rappresentata una catena metabolica costituita da tre passaggi catalizzati da differenti enzimi (x, y, z); mutazioni in uno qualunque dei tre geni, che inattivano uno dei tre enzimi portano tutte, come esito finale, al blocco della via e alla mancata produzione dello stesso metabolita.

11
3. I polimorfismi di restrizione (RFLP) e il loro utilizzo nella diagnostica delle
malattie genetiche
3.1 Gli enzimi di restrizione
Gli enzimi di restrizione (ER) riconoscono una sequenza di nucleotidi nella doppia elica del DNA e tagliano il DNA in frammenti di lunghezza definita (Tab.3.1). Sono enzimi prodotti dai batteri che li utilizzano per difendersi dagli attacchi di un DNA estraneo, per esempio di un virus (Fig.3.1). Sono stati fino ad ora purificati più di 1000 enzimi di restrizione prodotti da altrettanti tipi di batteri diversi.
Il DNA endogeno viene protetto dall’aggiunta di gruppi metile (-CH3) ai residui di adenina e citosina (Fig.3.1). Dal punto di vista biochimico gli ER sono delle endo-desossi-ribonucleasi che scindono un
legame fosfodiesterico. Gli ER si possono suddividere in varie classi a seconda della specificità e della modalità di taglio; ER diversi riconoscono e tagliano, in linea di massima, sequenze diverse; uno stesso enzima taglia qualsiasi tipo di DNA dovunque trovi la propria sequenza di riconoscimento, detta sito di restrizione. I siti di restrizione sono sequenze palindromiche (cioè che possono essere lete in entrambe le direzioni, come le parole ANNA, ETNA GIGANTE …) di poche (in genere 4, 5 o 6) coppie di nucleotidi. Gli ER sono uno strumento fondamentale per l’analisi del DNA. A causa del tipo di sequenze riconosciute (sequenze palindromiche) e della loro modalità di taglio (la maggior parte degli ER fa un taglio asimmetrico), tagliando due DNA diversi con uno stesso enzima si mettono allo scoperto le stesse sequenze nucleotidiche, si generano cioè le stesse estremità (estremità adesive o coesive) nei due tipi di molecole. Questo permette di costruire molecole di DNA ricombinante (Fig.3.2)
Fig. 3.1 I batteri producono enzimi di restrizione per difendersi dagli attacchi di DNA esogeno, per esempio DNA fagico. Altri enzimi (metilasi) proteggono il DNA batterico dall’azione delle proprie endonucleasi di restrizione.
Tab 3.1 Gli enzimi di restrizione si indicano con un sistema di lettere e numeri che si riferisce al ceppo batterico da cui sono stati isolati. Sono mostrati alcuni esempi di enzimi di restrizione, le sequenze di DNA che questi tagliano e i prodotti di scissione. Alcuni enzimi tagliano le sequenze in modo da dare origine ad estremità coesive, altri effettuano un taglio che determina la formazione di estremità piatte.
coesive

12
3.2 Polimorfismi del DNA Il termine polimorfismo significa “esistenza di forme diverse”. In genetica, il polimorfismo può essere analizzato a livello di proteina (polimorfismo proteico) oppure di materiale genetico (polimorfismo
genetico). In questo secondo caso, le forme diverse (ossia le varianti genetiche) possono riguardare un gene, vale a dire un tratto di DNA codificante una proteina (polimorfismo allelico), oppure un tratto di DNA non codificante (polimorfismo di sequenza). polimorfismo proteico Polimorfismo polimorfismo allelico polimorfismo genetico polimorfismo di sequenza
3.3 Polimorfismi di sequenza del DNA E’ stato osservato che il DNA di due individui differisce per circa un nucleotide ogni 500/1000. Queste diversità di sequenza si definiscono polimorfismi e dato che >98% del DNA umano è DNA non codificante, e che quindi la maggior parte di queste differenze è localizzata in sequenze non codificanti, il fenotipo di un polimorfismo di sequenza del DNA non è riconoscibile dall’esterno (come nel caso, ad esempio, dell’albinismo, o individuabile biochimicamente, come per i gruppi sanguigni). Dato l‘elevato numero di loci polimorfici (1 coppia di basi ogni 500/1000), i polimorfismi di sequenza del DNA sono molto più frequenti dei polimorfismi allelici tradizionali (albinismo, gruppi sanguigni, alleli della determinazione del colore degli occhi, etc, etc…) e conseguentemente più utili nella ricerca biologica e medica.
3.4 I polimorfismi di lunghezza dei frammenti di restrizione (RFLP)
Quando un polimorfismo interessa una sequenza riconosciuta da un dato ER, la variazione, creando o distruggendo siti di restrizione, darà luogo a differenze nei siti di taglio di quel dato enzima all’interno della popolazione. Digerendo il DNA di individui diversi con quell’enzima, si osserva quindi un polimorfismo di lunghezza dei frammenti di restrizione (RFLP, che per semplicità si usa leggere RIFLIP), e cioè dal DNA di individui diversi si generano frammenti di restrizione diversi. Come tutti i polimorfismi, i RFLP sono ereditabili come caratteri mendeliani semplici (Fig. 3.3) e sono trasmessi come caratteri codominanti. Il fenotipo di un RFLP è evidenziabile in termini di differenze di numero e/o dimensione dei frammenti di DNA ottenuti dalla digestione con un certo enzima di restrizione. I frammenti sono visibili dopo migrazione elettroforetica su un gel (Fig. 3.3 in basso).
Fig 3.2 Costruzione di una molecola di DNA ricombinante a partire da due molecole di DNA (una bianca e una nera) provenienti da fonti diverse ed entrambe tagliate con lo stesso enzima di restrizione EcoRI.

13
Fig. 3.3 Sono mostrati due frammenti di restrizione ottenuti con digestione con l’enzima EcoRI (E = sito di restrizione di EcoRI) di due alleli (1 e 2) di un gene. L’allele 1 è interrotto da un sito di taglio per l’enzima EcoRI (GAATTC), l’allele 2, a causa di una mutazione di una singola base, non presenta il sito di taglio (GACTTC). Trattando questi due alleli con l’enzima di restrizione EcoRI, si otterranno segmenti di lunghezza diversa: due segmenti (350 e 150 bp) nel caso dell’allele 1 e un solo segmento non tagliato (500 bp, 350 + 150) nel caso dell’allele 2. Negli individui eterozigoti sono presenti entrambi gli alleli e quindi si evidenzieranno tutti e tre i segmenti. Nella parte in basso della figura è mostrato un pedigree di una famiglia e sotto il simbolo dei singoli individui è indicato il genotipo relativamente agli alleli 1 e 2. Dal pedigree mostrato si
capisce come i RFLP abbiano ereditarietà mendeliana semplice e siano codominanti. Nel caso osservato, una coppia eterozigote ha una figlia omozigote per l’allele 1 e un figlio omozigote per l’allele 2. Quando il DNA dei genitori e dei figli è digerito con l’enzima di restrizione EcoRI e i frammenti di DNA vengono separati mediante corsa elettroforetica, il DNA dei genitori eterozigoti mostrano 3 bande corrispondenti a frammenti da 500, 350 e da 150 bp; la figlia omozigote 1/1 mostra 2 bande corrispondenti a frammenti da 350 e 150 bp; il figlio omozigote 2/2 mostra una sola banda da 500 bp.
3.4.1 La tecnica della PCR per l’analisi degli RFLP L’introduzione della PCR, la tecnica che consente di amplificare selettivamente un tratto di DNA, ha rivoluzionato la genetica molecolare. (Per la descrizione della tecnica, vedere più avanti al § 5.1). Le applicazioni della PCR sono praticamente infinite. Uno degli ambiti di utilizzo è la diagnosi di malattie genetiche mediante analisi dei RFLP. L’utilizzo della PCR semplifica molto le cose. Ad esempio, la PCR consente di analizzare uno specifico tratto di DNA, invece di dover lavorare su tutto il DNA nucleare di una cellula, ossia sul DNA genomico. Come è illustrato nella Fig. 3.4, i primer (inneschi) della PCR vengono disegnati a monte e a valle del
sito di restrizione. Il segmento di DNA amplificato viene sottoposto a digestione con l’enzima di restrizione e i prodotti della digestione vengono analizzati dopo separazione elettroforetica.
Fig.3.4 I primer della PCR vengono disegnati a monte e a valle del sito di restrizione (sito di taglio dell’ER, cut site). Dopo l’amplificazione, il DNA viene digerito con l’enzima di restrizione e i frammenti di DNA vengono separati mediante corsa elettroforetica. Gli omozigoti e gli eterozigoti sono distinguibili in base alle diverse bande ottenute dalla digestione con l’ER. Gli omozigoti 1/1 e 2/2 daranno rispettivamente due bande e 1 banda; gli eterozigoti daranno 3 bande.

14
3.5 I polimorfismi di restrizione (RFLP) nella diagnosi delle malattie genetiche
3.5.1 Utilizzo di un RFLP nella diagnosi diretta. Diagnosi di anemia falciforme Gli individui affetti da anemia falciforme sono omozigoti per la mutazione HbS, che consiste nella sostituzione di un singolo amminoacido dei 146 che formano la catena dell’emoglobina. Nell’emoglobina HbS, l’acido glutammico (Glu) nella sesta posizione della catena è sostituito dalla valina (Val). La sostituzione amminoacidica è dovuta alla mutazione A T nella posizione mediana del codone 6 (Fig. 3.5). La sostituzione che converte il codone GAG (acido glutammico) nel codone GTG (valina) modifica la sequenza CCTGAGG (che interessa i codoni 5, 6 e 7) riconosciuta e tagliata dall’enzima di restrizione MstII. Gli individui omozigoti ed eterozigoti sono distinguibili in base al tipo di bande ottenute dal loro DNA dopo digestione con l’enzima MstII (Fig. 3.5, in basso).
In questo caso specifico, la stessa mutazione che causa la malattia altera anche un sito di restrizione. La semplice digestione con l’enzima di restrizione rilevante consente di fare diagnosi diretta di malattia e di riconoscere i portatori (Fig. 3.5 e 3.6).
Fig.3.6. E’ mostrato il pedigree di una famiglia in cui soltanto il figlio 1 è affetto da anemia falciforme. Entrambi i genitori sono eterozigoti e per questo motivo hanno avuto un figlio malato. Le bande elettroforetiche per ogni membro della famiglia (visibili al di sotto di ognuno) consentono di identificare il genotipo di ogni individuo (vedere legenda Fig.3.3 e 3.5).
Fig. 3.5 La figura mostra l’enzima di restrizione Mst II che riconosce e taglia la sequenza CCTGAGG. Il DNA dell’allele A viene tagliato dall’enzima di restrizione. Una mutazione di una singola base fa perdere il sito di taglio dell’enzima MstII e il DNA dell’allele S non viene tagliato dall’enzima. Nella parte bassa della figura si vede il risultato della corsa elettroforetica relativa ai DNA di un individuo normale, di un portatore sano e di un individuo affetto amplificati con PCR (utilizzando i primer indicati dalle frecce orizzontali) e successivamente digeriti con l’enzima MstII. L’individuo normale ( A A) mostra 2 bande, il portatore sano ( A S) mostra 3 bande e l’individuo affetto ( S
S) mostra una sola banda di lunghezza corrispondente al DNA non tagliato.

15
3.5.2 Utilizzo di un RFLP nella diagnosi indiretta. Ricerca di un RFLP associato a un allele-
malattia
A differenza di quello che avviene nel caso dell’anemia falciforme, la maggior parte delle malattie genetiche sono dovute a numerose mutazioni diverse dello stesso gene (eterogeneità genetica, vedere pag. 10). In questo caso la mutazione responsabile della malattia non corrisponde a un unico polimorfismo di sequenza (e quindi a un unico eventuale RFLP). In questi casi non è quindi possibile utilizzare un RFLP per la diagnosi diretta. E’ invece possibile trovare uno o più RFLP associati al gene-malattia (in quanto localizzati all’interno o nelle vicinanze del gene) che possono essere utilizzati per la diagnosi: si tratta in questo caso di diagnosi indiretta per associazione e non di diagnosi diretta. Per poter utilizzare il RFLP come marcatore occorre che il gene responsabile della malattia sia collocato così vicino al RFLP che quest’ultimo possa rivelare la presenza stessa del gene malato. Un particolare RFLP può essere associato in alcuni individui con l’allele–malattia ed in altri con l’allele sano. Pertanto, per poter utilizzare un RFLP nella diagnosi indiretta, è necessario esaminare non soltanto il DNA del malato ma il maggior numero possibile di membri della famiglia per identificare il tipo di associazione presente in quel pedigree (Fig. 3.7). L’importanza dei RFLP come marcatori genetici di associazione sta nel fatto che consentono la diagnosi di malattia anche in casi in cui non è noto il gene responsabile. I marcatori RFLP (nel genoma umano sono stati identificati migliaia di RFLP e diverse centinaia sono stati assegnati a ciascun cromosoma) hanno consentito la mappatura di alcuni importanti geni-malattia (ad esempio, è stato possibile assegnare il gene della fibrosi cistica in posizione distale del braccio lungo del cromosoma 7). A differenza della diagnosi diretta, l’accuratezza della diagnosi per associazione è del 100% solo quando il locus malattia e il RFLP sono strettamente associati (vicini sul cromosoma). In questo caso, infatti la frequenza di crossing-over che può modificare l’associazione è trascurabile. Tanto maggiore è la distanza tra il RFLP e il locus genico di interesse, tanto più bassa è la probabilità di una diagnosi accurata. Un secondo aspetto da prendere in considerazione è che non sempre un dato RFLP può essere utilizzato ai fini della diagnosi indiretta. E’ necessario che nella famiglia siano presenti i due alleli (se tutti gli individui della famiglia sono portatori dello stesso allele del RFLP studiare quel RFLP in quella famiglia non è significativo!).
Fig.3.7 Il pedigree mostra l’ereditarietà di un marcatore RFLP attraverso tre generazioni di una famiglia in cui è presente una malattia recessiva legata al cromosoma X. Nella famiglia sono presenti gli alleli (+) e (-). Tutti gli individui malati sono maschi che presentano l’allele (+). Tutti gli individui portatori dell’allele (-) sono sani; si può dedurre che l’allele malattia sia strettamente associato all’allele (+) di questo RFLP. Sia la madre (II 1) che la nonna I 1 sono eterozigoti portatrici sane in cui l’allele (+) è associato all’allele malattia. L’allele (-) del RFLP è associato all’allele sano del gene malattia come dimostrato da I 2, III 5, III 6 e III 7.
L’accuratezza della diagnosi dipende dal grado di associazione dei due loci. Se il marcatore e il locus malattia non sono strettamente associati, il crossing-over durante la formazione dei gameti potrebbe modificare l’associazione e un individuo portatore dell’allele (+) del RFLP non sarebbe portatore dell’allele malattia.
MW
I
II
III
1
1
1
1
2 3 4
2
1 2 3 4 5 6 7 8