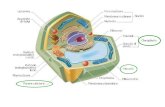
Regolazione dell’espressione genica negli eucarioti · cis-agenti. Prof. Savino; dispense di...
18
Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie Regolazione dell’espressione genica negli eucarioti
-
Upload
trinhkhuong -
Category
Documents
-
view
227 -
download
0
Transcript of Regolazione dell’espressione genica negli eucarioti · cis-agenti. Prof. Savino; dispense di...

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Regolazione dell’espressione genica negli eucarioti

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Il promotore eucariotico
• Il promotore eucariotico è l’insieme di tutti questi siti di legame; la polimerasi si lega solovicino al sito di inizio, non contatta le regioni a monte del promotore. Qunidi sono i fattori di trascrizione che riconoscono il promotore e creano una struttura riconosciuta dall’enzima.
– fattori generici di trascrizione• Le sequenze che costituiscono il promotore sono localizzate vicino al sito di inizio e sono
necessarie per l’inizio stesso. La presenza dei siti intensificatori (enhancer) invece stimolal’inizio. Queste sequenze si possono trovare anche a grande distanza dal sito di inizio, esono il bersaglio per la regolazione dell’espressione tessuto specifica o temporale.
• La regolazione negli eucarioti è positiva.
L’inizio della trascrizione negli eucarioti necessita della RNA polimerasi edei fattori di trascrizione.Qualsiasi proteina sia necessaria per l’inizio della trascrizione, ma nonsia parte della polimerasi, è un fattore di trascrizioneL’inizio della trascrizione necessita di un gran numero di questi fattori,che legano varie sequenze cis-agenti.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Il complesso di inizio • Il primo evento è il legame di TFIID alla TATA.
• Poi si lega TFIIA.• TFIIB contatta il solco
secondario a valle ed il solco principale a monte della TATA box.
• TFIIF (subunità RAP 38) recluta la polimerasi e la porta nel complesso.
• Poi si lega TFIIE.• Dopo TFIIE si legano
TFIIH e TFIIJ.
TFIIE e TFIIH aprono in due filamenti di DNA e permettono alla RNA polimerasi II di iniziare la trascrizione. L’attività chinasica di TFIIH fosforila il CTD, e questo permette alla polimerasi di iniziare la fase di elongazione.
I fattori si staccano.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Il promotore eucariotico
• Il promotore eucariotico è l’insieme di tutti questi siti di legame; la polimerasi si lega solo vicino alsito di inizio, non contatta le regioni a monte del promotore. Qunidi sono i fattori di trascrizione che riconoscono il promotore e creano una struttura riconosciuta dall’enzima.
– fattori generici di trascrizione• Le sequenze che costituiscono il promotore sono localizzate vicino al sito di inizio e sono necessarie
per l’inizio stesso. La presenza dei siti intensificatori (enhancer) invece stimola l’inizio. Queste sequenze si possono trovare anche a grande distanza dal sito di inizio, e sono il bersaglio per laregolazione dell’espressione tessuto specifica o temporale.
• La regolazione negli eucarioti è positiva.
L’inizio della trascrizione negli eucarioti necessita della RNA polimerasi edei fattori di trascrizione.Qualsiasi proteina sia necessaria per l’inizio della trascrizione, ma nonsia parte della polimerasi, è un fattore di trascrizioneL’inizio della trascrizione necessita di un gran numero di questi fattori,che legano varie sequenze cis-agenti.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Brevi sequenze di riconoscimento nel promotore legano gli attivatori
• L’efficienza e la specificità con cui un promotore è riconosciuto dipendono però da altre sequenze, situate a monte, che sono riconosciute da altri fattori, chiamati attivatori
– i siti di legame sono ~100 bp a monte del sito di inizio, ma possono essere anche più distanti.• I motivi di sequenza a monte della TATA influenzano la frequenza di inizio, agendo
direttamente sui fattori generici di trascrizione per aumentare l’efficienza di assemblaggio del complesso di inizio.
QuickTime™ and aGIF decompressor
are needed to see this picture.
Il promotore della RNA polimerasi II è formato da due regioni. Il sito di inizio è identificato dalla regione Inr e dalla TATA box nella sue vicinanze. La RNA polimerasi II, assieme ai fattori generici di trascrizione, forma il complesso di inizio nella regione del sito di inizio.
CAAT boxGC box

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
La struttura del promotore è flessibile
• Sebbene il promotore sia direzionale (la trascrizione avviene solo in una direzione) la “CG box” e la “CAAT box” funzionano in entrambi gli orientamenti
– queste sequenze funzionano solamente come siti di legame al DNA per richiamare i fattori di trascrizione nelle vicinanze del sito di inizio.
• Gli attivatori ubiqitari agiscono su qualsiasi promotore che contenga la loro sequenza di legame; tra queste ci sono la “CAAT box”, la “GC box” e l’ottamero. Tutti i promotori hanno bisogno di uno o più di questi elementi per funzionare efficientemente. Di solito, un attivatore riconosce una sequenza di DNA di <10 paia di basi, ma copre un frammento di DNA di ~20 paia di basi. Considerato il numero degli attivatori e la regione di DNA coperta da ciascuno di essi, le varie proteina coprono interamente la regione di DNA a monte del sito di inizio.
I promotori sono organizzati in maniera combinatoriale.Diversi elementi possono contribuire alla funzionalità del promotore, ma nessuno di essi è essenziale per tutti i promotori. Oltre alla TATA, tre sequenze si trovano in parecchi promotori: la scatola CG (GC box), la scatola CAAT (CAAT box) e l’ottamero (una sequenza di 8 paia di basi). In ogni singolo promotore, sequenze come queste differiscono per numero, posizione e orientamento.Nessuna sequenza è comune a tutti i promotori.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Gli enhancer contengono elementi si sequenza che funzionano in entrambi gli orientamenti e che facilitano l’inizio della trascrizione
• Il promotore è definito dall’insieme delle sequenze di DNA che devono essere in una posizione (relativamente) rigida rispetto al sito di inizio.
• In base a questa definizione, la TATA box (per esempio) fa parte del promotore, le sequenze che formano l’enhancer no.
Non sempre i promotori funzionano da soli. In alcuni casi l’attività di un promotore è aumentata enormemente dalla presenza di un enhancer, che è composto da un altro gruppo di sequenze, site però a distanze variabili da quelle che compongono il promotore stesso.
Non è necessario che la posizione dello enhancer sia fissa rispetto al promotore: essa può essere variata in maniera significativa. Nei genomi, gli enhancers si sono trovati all’interno dei geni o a decine di migliaia di coppie di basi di distanza in entrambe le direzioni.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Gli enhancer contengono gli stessi elementi di sequenzache si trovano nei promotori
• Molte delle sequenze che si trovano negli enhancer sono presenti anche nei promotori; per esempio, l’ottamero.
• La specificità della trascrizione può essere controllata sia dall’enhancer che dal promotore. • Un promotore può essere regolato, e un enhancer nelle “vicinanze” può essere usato per
aumentare l’efficienza di inizio della trascrizione.• Un promotore può non essere regolato, ma diviene veramente efficace quando viene
attivato un enhancer nelle sue “vicinanze”.
Una differenza tra l’enhancer ed il promotore è rappresentata dalla densità dei motivi di sequenza. Quando si effettua un analisi mutazionale, è molto più frequente che una mutazione sia rilevante quando è fatta all’interno di un enhancer piuttosto che all’interno di un promotore.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Ci sono vari tipi di fattori di trascrizione• I fattori generici di trascrizione si legano al sito di
inizio e alla TATA box, assieme alla RNA pol. II– posizionano la polimerasi ed assicurano che l’inizio
della trascrizione avvenga al punto giusto.• Gli attivatori sono dei fattori di trascrizione che
riconoscono specifiche, corte sequenze consenso– si legano a siti nel promotore o nell’enhancer;– aumentano la frequenza di inizio e sono necessari
per un adeguato funzionamento del promotore;– alcuni sono presenti in tutte le cellule (ubiquitari);– altri hanno una funzione regolatoria: sono sintetizzati
o attivati a tempi precisi o in tessuti specifici• questi fattori sono responsabili del controllo della
trascrizione
La diversità di elementi che costituiscono un promotore funzionale e la variabilità della loro posizione suggerisce che gli attivatori possono interagire tra loro in molteplici maniere. E’ stato dimostrato che gli attivatori che si legano sull’enhancer si vengono a trovare vicino al promotore perchè è il DNA che si ripiega.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Come agiscono gli attivatori
• Il dominio di attivazione agisce contattando i fattori generici di trascrizione coinvolti nelle fasi iniziali di assemblaggio del complesso di inizio
– tipicamente TFIID, TFIIB o TFIIA.• La funzione dell’attivatore è quella di facilitare ed
accellerare l’assemblaggio del complesso di inizio.• TFIID è formato da TBP e da 11 TAF: il ruolo
principale dei TAF è di creare un collegamento tra l’apparato basale e gli attivatori
– diversi TAF possono fornire le superfici di contatto per diversi attivatori.
Gli attivatori sono composti da domini indipendenti:• il dominio di legame al DNA (classificazione degli attivatori);• il dominio di connessione (linker);• il dominio di attivazione vero e propio.I motivi di sequenza del promotore possono essere ad una distanza considerevole da sito di inizio, e orientati in entrambe le maniere; gli enhancer possono essere ancora più lontani: la struttura dell’attivatore deve essere flessibile.
Altri attivatori con un dominio di attivazione ad alta carica negativa (acidic activators) aumentano invece l’efficienza con cui TFIIB è reclutato nel complesso di inizio.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Altri tipi di attivatori• A volte l’attivatore
non ha un dominio di attivazione
• In questo caso l’attivatore lega un’altra proteina che ha un dominio di attivazione: un coattivatore, che fornisce il collegamento tra il dominio di legame al DNA e l’apparato di base della trascrizione.
Infine, alcuni regolatori agiscono cambiando la struttura della cromatina e rendendola più accessibile alla trascrizione.In genere la gerarchia é: attivatore→(coattivatore)→ fattori generici di trascrizione→RNA polimerasi II
–questo spiega la flessibilità di costruzione del sistema: i fattori non devono contattare direttamente la polimerasi.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Regolazione degli attivatori• Sintesi.• Modificazione.• Attivazione
indotta da ligando.
• Taglio proteolitico da un precursore inattivo.
• Associazione con altre proteine.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Il legame al DNA:dita con zinco
• Gli attivatori di questa classe hanno da un minimo di 2 “dita” fino a un massimo di 27 “dita”.
• Ci sono mediamente 7-8 amminoacidi tra due “dita”.• La parte C-terminale di ogni “dito” forma una α-elica
che lega il DNA, mentre la parte N-terminale forma un foglietto β.
• Le α-eliche si infilano nel solco maggiore del DNA instaurando delle interazioni sequenza-specifiche.
• Gli amminoacidi non conservati nella regione C-terminale (α-elica) sono responsabili della specificità di riconoscimento.
Le “dita con zinco” (zinc fingers) prendono il nome dalla loro struttura, in cui un piccolo gruppo di amminoacidi conservati legano un atomo di zinco, formando un dominio indipendente nella proteina. Il consenso per un “dito” è:
Cys-X3-Cys-X12-His-X3-His, in complesso 23 amminoacidi e sono chiamate “dita Cys2/His2”.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
I recettori degli steroidi
• I recettori degli steroidi sono attivatori.• Le diverse attività biologiche dei vari
steroidi dipendono da quali geni vengono “accesi” dagli attivatori con cui ogni steroide interagisce.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Il legame dei recettori degli steroidi alla sequenza bersaglio è indotto dall’interazione con il ligando
• Gli steroidi attraversano la membrana per semplice diffusione (sono idrofobici).
• Nella cellula, legano il recettore nella regione C-terminale della proteina– il complesso ormone recettore si ritrova nel nucleo
• Con meccanismi che dipendono dai singoli recettori, il recettore diventa capace di legare la sua sequenza bersaglio sul DNA, che di solito si trova in un enhancer.
• Due copie del recettore/attivatore legano una singola sequenza di riconoscimento– molti attivatori agiscono in realtà come dimeri.
Il dominio di legame al DNA ha 2 “dita di zinco”, di tipo leggermente diverso, con un consenso Cys-X2-Cys-X13-Cys -X2-Cys, chiamato “dito di zinco Cys2/Cys2”.
A questo punto, il promotore associato all’enhancer è attivato e la trascrizione inizia.

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
L’omeodominio
• L’omeodominio forma 3 α-eliche.• L’elica 3 entra nel solco maggiore ed è
responsabile per il maggior numero di contatti.• Altri contatti sono a carico della regione N-
terminale dell’omoedominio, che entra nel solco minore.
• Un numero relativamente piccolo di amminoacidi nell’elica 3 e nella regione amminoterminale sono responsabili della specificità di riconoscimento.
L’omeodominio è una sequenza conservata di 60 amminoacidi presente in alcuni attivatori. Attivatori contenenti un omeodominio sono presenti in tutti gli eucarioti. L’omeodominio è il dominio di legame al DNA

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
Da “I PRINCIPI DI BIOCHIMICA DI LEHNINGER
L’omeodominio• Conseguenze della mutazione della
proteina bithorax.• Conseguenze della mutazione della
proteina antennapedia.
Da “I PRINCIPI DI BIOCHIMICA DI LEHNINGER

Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
La serratura di leucine (leucine zipper)
• Si formano delle α-eliche in cui gli amminocidi idrofobici (tra cui le leucine) sono da un lato e gli amminoacidi polari sono dall’altro.
• Le leucine della α-elica della prima proteina si incastrano nelle leucine della α-elica della seconda proteina, formando il dimero.
• La regione N-terminale delle due proteine è fortemente basica (carica positivamente) e forma il dominio di legame al DNA.
Le proteine a “serratura di leucine” devono il loro nome alla ripetizione di 4 o 5 residui di leucina posti a distanza fissa uno dall’altro. Questa caratteristica fa si che due proteine che contengono queste ripetizioni di leucine interagiscono tra loro a formare dei dimeri.
Si forma una struttura a Y, in cui il gambo della Y è formato dalla serratura di leucine, e le due regioni basiche si biforcano simmetricamente formando le braccia che legano il DNA. La sequenza bersaglio di queste proteine è una palindrome.