
Redalyc.Introdução à morfologia e função nuclearO nucleo das células dos mamiferos é um...
39
REDVET. Revista Electrónica de Veterinaria E-ISSN: 1695-7504 [email protected] Veterinaria Organización España Dias Correia, J. H. R.; Dias Correia, A. A. Introdução à morfologia e função nuclear REDVET. Revista Electrónica de Veterinaria, vol. 12, núm. 8, agosto, 2011, pp. 1-38 Veterinaria Organización Málaga, España Disponível em: http://www.redalyc.org/articulo.oa?id=63621920008 Como citar este artigo Número completo Mais artigos Home da revista no Redalyc Sistema de Informação Científica Rede de Revistas Científicas da América Latina, Caribe , Espanha e Portugal Projeto acadêmico sem fins lucrativos desenvolvido no âmbito da iniciativa Acesso Aberto
Transcript of Redalyc.Introdução à morfologia e função nuclearO nucleo das células dos mamiferos é um...

REDVET. Revista Electrónica de Veterinaria
E-ISSN: 1695-7504
Veterinaria Organización
España
Dias Correia, J. H. R.; Dias Correia, A. A.
Introdução à morfologia e função nuclear
REDVET. Revista Electrónica de Veterinaria, vol. 12, núm. 8, agosto, 2011, pp. 1-38
Veterinaria Organización
Málaga, España
Disponível em: http://www.redalyc.org/articulo.oa?id=63621920008
Como citar este artigo
Número completo
Mais artigos
Home da revista no Redalyc
Sistema de Informação Científica
Rede de Revistas Científicas da América Latina, Caribe , Espanha e Portugal
Projeto acadêmico sem fins lucrativos desenvolvido no âmbito da iniciativa Acesso Aberto

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
1
REDVET - Revista electrónica de Veterinaria - ISSN 1695-7504
Introdução à morfologia e função nuclear (Introduction to nuclear morphology and function)
Dias Correia; J. H. R. : CIISA, Departamento de Morfologia e Função, Faculdade de Medicina Veterinária de Lisboa, Avenida da Universidade Técnica, Pólo Universitário, Alto da Ajuda, 1300-477, Lisboa, Portugal email: [email protected] │ Dias Correia, A. A.: Professor jubilado de Faculdade de Medicina Veterinária de Lisboa, Avenida da Universidade Técnica, Pólo Universitário, Alto da Ajuda , 1300-477, Lisboa, Portugal.
Resumo O núcleo das células dos mamíferos é um organelo compartimentado quanto a espaço e funcionamento, contendo numerosos corpos nucleares proteinaceos assim como domínios do genoma, não colocados ao acaso no seu interior, ao contrário do que se pensava inicialmente de que seria destituído de organização interior. Estes compartimentos são desprovidos de biomembranas e possuem um conjunto de proteínas diferentes consoante o compartimento. O núcleo celular é um organelo altamente dinâmico havendo situações em que se realiza independentemente da utilização de energia ao contrario de outras. A zona periférica do núcleo adjacente ao invólucro nuclear é a zona onde parece ocorrer a activação e repressão dos genes dos cromossomas. No invólucro nuclear é considerada a membrana nuclear externa, a membrana nuclear interna, a lamina nuclear, e os complexos poros. As membranas nucleares e a lamina nuclear interactuam entre si através de diversas proteínas, algumas integrais da membrana nuclear interna, outras associadas com as laminas nucleares e com a proteínas do nucleoplasma dentro da matriz nuclear. A matriz nuclear com grandes quantidades de RNP (ribonucleoproteinas) interactua com determinadas zonas do DNA dos cromossomas. São revistas algumas interações entre estes variadíssimos tipos moleculares, inclusive com o citoesqueleto celular.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
2
Também suborganelos subnucleares contribuem para as movimentações intranucleares, gerando-se provavelmente gradientes de pH intranucleares envolvidos em toda a dinâmica celular. Palavras chave :núcleo, organelos sub-nucleares, nucleoplasma, nucleoesqueleto, compartimentos nucleares, corpos nucleares proteinaceos, invólucro, lamina, matriz, laminopatias Summary The nuclei belonging to cells from mammals have been organells divided in different compartments as far as space and functioning have been concerned. They contain several proteinaceous nuclear bodies as well as genome’s domains that have not been organized randomly inside the nuclei. This view has been in opposition to the initial trought that the nucleus was devoid of internal structure. The compartments in the nucleus are free from membranes and show different array of proteins in separate compartments. The nucleus has been a highly dynamic organell and its dynamics and has in some circumstances been independent of energy consumptiom, but in other circunstances energy expenditure has been implied. The periphery of a nucleus (close to the nuclear envelope) has been the area where it has looked like that the activation and the repression of genes take place. In the nucleus’ envelope one must consider the external nuclear membrane, the internal nuclear membrane, the nuclear lamina and the pores complex. There has been an interaction between the nuclear membranes and the nucleus’ lamina, the interaction being mediated by several proteins. Some of these proteins have been the nucleus’ inner membrane integral proteins, some others have been associated with the nuclear lamina and the proteins of the nucleoplasm inside the nucleus’ matrix. The nuclear matrix, holding large amounts of riboonucleoproteins (RNP) interacts with DNA in cromossomes, In this paper the two authors review these varied molecular types including their interaction with the cell’s cytoskeleton. Subnuclear organells make their contribution towards movement of partcles inside the nucleus probably becoming responsible of the pH gradients inside the nucleus that, in the other hand, have possibly been involved in the whole cellular dynamics. Keywords: nucleus, subnuclear organelles, nucleoplasm, nucleoskeleton nuclear compartments, proteinaceos nuclear bodies,envelope, lamina, matrix, laminopthies

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
3
1-Introdução à morfologia e função nuclear. 1.1-Introdução Sumula global da arquitectura nuclear (Misteli, T. 2005). O nucleo das células dos mamiferos é um organelo compartimentado quanto a espaço e funcionamento, contendo numerosos corpos nucleares proteinaceos assim como domínios do genoma, não colocados ao acaso no seu interior. As mudanças da arquitectura nuclear são fundamentais durante os processos de desenvolvimento e diferenciação celular. O estudo intenso da arquitectura nuclear levou à criação de diversos conceitos gerais através dos quais se tem criado alguma compreensão acerca do funcionamento nuclear.Assim Mistell ,T. (2005) desenvolve para este efeito uma serie de conceitos que se referem seguidamente:
• 1º O conceito dos compartimentos nucleares onde há que considerar os corpos nucleares proteinaceos e os domínios de cromatina.
• 2º O conceito dos territórios ocupados pelos cromosomas. • 3º O conceito da organização nuclear não ao acaso onde se
desenvolvem os genomas em três dimensões e o posicionamento dos corpos nucleares.
• 4º O conceito da dinâmica nuclear. • 5º O conceito de “scaffolds” nucleares. • 6º O conceito da própria organização do núcleo. • 7º O conceito das doenças nucleares.
Segundo Mistelil dadas as dificuldades de definir um “esqueleto” estático para o núcleo, e, por outro lado , dado o elevado dinamismo da arquitectura do núcleo, têm ocorrido sugestões que vão no sentido de que o núcleo é uma entidade que se organiza a si própria. No núcleo das células dos mamíferos sabe-se hoje que existe um número crescente de suborganitos no seu interior, (ao contrário do que se pensava inicialmente de que seria destituído de organização interior) compartimentos ou corpos nucleares ou sejam domínios especializados (Spector, D.L., 2001). A expressão dominio no contexto cientifico pode ter diversas conotações, independentemente da clássica utilização na classificação biológica. Assim por analogia e noutros contextos, conjuntos complexos de variadíssimas moléculas biológicas no núcleo podem constituir

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
4
simplesmente o que se considera serem domínios nucleares, dadas as suas características funcionais. Já quanto aos domínios proteicos estes são considerados como uma parte da sequência da proteína e da sua estrutura que pode transformar-se, funcionar e existir independentemente do resto da cadeia da proteína. Neste caso cada domínio forma uma estrutura tridimensional compacta que pode conter de 25 a 500 ácidos aminados de comprimento. Diversas proteínas podem possuir diversos domínios estruturais. Por outro lado, na organização global dos genomas, os domínios da cromatina ( a cromatina é uma combinação de DNA, histonas e outras proteínas que constituem os cromossomas) podem conter genes vizinhos que podem ser “arrumados” juntos em estruturas cromatínicas específicas que asseguram a sua expressão coordenada ( domínios cromatinicos). Cada dominio cromaíinico pode incluir dezenas de genes por vezes com funções similares. Os domínios cromatinicos são estruturalmente distintos e representam unidades reguladoras para a expressão dos genes e comportamentos dos cromossomas. Os domínios cromatinicos são muitas vezes definidos através de conjuntos distintos de histonas modificadas após a sua tradução. O estabelecimento de distintos domínios é muitas vezes realizado através destas modificações proteicas das histonas, após a sua tradução A organização do núcleo das células eucariotas dos animais superiores constitui um elemento chave para o funcionamento do genoma. Este núcleo encontra-se funcionalmente compartimentado como referimos em suborganitos contendo cada um deles numerosos corpos nucleares proteinaceos, assim como domínios , não posicionados de qualquer maneira mas sim numa determinada organização espacial e temporal. Mudanças na arquitectura nuclear decorrem durante os processos de desenvolvimento e diferenciação celular , sendo conhecido que deficiências nesta arquitectura nuclear podem ser a causa de diversas doenças já conhecidas nos seres humanos e ratos ( Mus musculus). Dentro das doenças nucleares identificadas em seres humanos e em ratinhos experimentais, que são já numerosas, as laminopatias

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
5
causadas por mutações nas proteínas lâminas A e C codificadas pelo gene LMNA originam distrofias musculares, lipodistrofias, neuropatias e outras doenças, em consequência de defeitos mecânicos originados na lâmina nuclear, ou de mutações. Um compartimento nuclear é pois uma região morfologicamente e funcionalmente distinta, dentro do núcleo. Estes compartimentos são desprovidos de biomembranas e possuem um conjunto de proteínas diferentes consoante o compartimento. Assim, como veremos adiante, o nucléolo encerra actividades de transcrição e processamento dos rRNA, os compartimentos com factores de “splicing” encerram componentes spliceossomais, os corpos de Cajal são locais de montagem de snRNP (pequenas ribonucleoproteinas) e os corpos PML tem funções um tanto desconhecidas. Um corpo nuclear é pois um local com actividades particulares. Estes compartimentos nucleares desprovidos de biomembranas parecem resultar mais de interacções transitórias e não específicas entre as proteínas neles residentes,mais do que da presença de um “scaffold” estável. O conceito da existência de “scaffolds” nucleares não tem tido suporte experimental, embora se possa admitir que as proteínas lâmínas (lamins) A, B e C do tipo filamentos intermediários, que são bem conhecidos, possam formar uma rede, a lâmina nuclear, revestindo internamente o invólucro nuclear excepto ao nível dos poros. Nas células dos animais o respectivo núcleo é o habitat natural do genoma onde cada cromossoma parece ocupar um determinado espaço (território) o que parece apontar para o seu desenvolvimento na regulação do funcionamento dos genes e na estabilidade do genoma, quer no estado hÍgido, quer em situações anormais (Meaburn, K.J. e Mistel, 2007). Nas células eucariotas o núcleo desempenha uma série de processos separados no tempo e no espaço de todos os outros componentes e compartimentos celulares (Rzepeck, 2002). Nesses processos ocorridos no núcleo incluem-se a replicação, a transcrição, o processamento do RNA e a montagem das subunidades ribossomais, enquanto a maioria das biossínteses proteicas decorre no citoplasma celular, implicando o transporte posterior de muitas proteínas para o núcleo da célula. Os domínios da cromatina são morfologicamente definidos em regiões genómicas menos condensadas (eucromatina) ou altamente condensadas

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
6
(heterocromatina), as primeiras activas para a transcrição e as segundas ricas em regiões inactivas e silenciadas, o que parece ser uma perspectiva um tanto superficial. Ambas estas cromatinas ( eu ou hetero) têm modificações nas sua histonas que ocorrem após a tradução destas, como veremos adiante, as eucromatinas ricas em histonas H3 e H4 hiperacetiladas e as heterocromatinas com hipoacetilação e trimetilação da histona H3 na lisina 9. Os cromossomas existem ocupando especialmente determinados territórios nos núcleos. A estrutura interna dos cromossomas é ainda um tanto ambígua, admitindo-se em princípio que os genes activos transcriptionalmente se situam à superfície do cromossoma havendo como que canais entre os diversos territórios de cada cromossoma em ordem a facultar a circulação de factores reguladores. Mesmo a cromatina em interfase é dinâmica, com porções dessa cromatina cromossomal desenrolando-se ou enrolando-se (eucromatina ↔ heterocromatina) em ordem a emitir para a sua superfície ansas dessa estruturas aptas a serem transcritas ou silenciadas. Durante a interfase a maioria da heterocromatina da maior parte dos cromossomas situa-se na periferia do núcleo com uma pequena parte à volta do nucleólo ou difundindo-se no nucleoplasma e provavelmente ligada ao invólucro nuclear, enquanto a eucromatina se distribui mais para o interior do núcleo através de múltiplas ansas. Durante a mitose as clássicas bandas claras e escuras dos cromossomas alternam, com a cromatina das bandas escuras (heterocromatina) a replicar-se tardiamente na fase S e com as bandas claras (eucromatina) a replicar-se no início da fase S. No núcleo nos espaços que se situam entre a cromatina encontram-se complexos proteicos e ribonucleoproteícos (snRNP) envolvidos no metabolismo do RNA e noutros metabolismos. A acumulação de diversos factores nucleares nestes compartimentos produz uma elevada concentração desses factores nesses locais favorecendo certo tipo de reacções (Lamond & Sleeman, 2003). De tudo isto ressalta, repete-se, que a organização nuclear não é ao acaso, e os cromossomas ricos em genes parecem posicionar-se sobretudo no centro do núcleo, enquanto os pobres em genes se situariam na periferia deste núcleo. A posição relativa de uns cromossomas em relação aos outros parece ser específica de cada tecido (Boyle et, al.; Parada et, al.;Cremer et, al., citados em Misteli, T. 2005), mas os mecanismos que estão na base deste comportamento são desconhecidos. Os loci dos genes dos diversos cromossomas também não ocupam uma distribuição ao acaso, mas ignora-se quais as razões destes comportamentos.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
7
O núcleo celular é pois um organelo altamente dinâmico e este dinamismo é complexo havendo situações em que se realiza independentemente da utilização da energia ao contrário de outras. As proteínas e os seus complexos exibem “in vivo” movimentos dinâmicos, tal como os corpos nucleares, embora o nucléolo seja, quanto à sua posição, relativamente estável, pelo menos durante curtos períodos de tempo, mas a maioria dos outros componentes nucleares são dinâmicos no seu posicionamento, sendo sensíveis às taxas de ATP, para este efeito. As proteínas podem circular rapidamente no espaço nuclear permutando componentes entre os diversos compartimentos tais como com a cromatina, os cromossomas e os corpos nucleares, bem como com o citoplasma celular e nos dois sentidos (de saída e de entrada de moléculas) através de sinais residentes nessas variadíssimas moléculas e de transportadores e receptores adequados (Carmo-Fonseca, M. 2000). Também os loci dos genes têm movimentações dinâmicas. Repete-se que nos compartimentos do núcleo há que considerar além dos cromossomas, uma série de corpos nucleares no nucleoplasma, onde se incluem os nucléolos, corpos de Cajal, “Gems”, “splicing speckles”, corpos PML (promielocitic leucemia) e outros pior conhecidos (Lamond & Sleeman, 2003), (vide Figura 1 seguinte) É hoje conhecido, como temos referido, que os genes e os cromossomas não se encontram posicionados ao acaso dentro do núcleo, sendo este posicionamento finamente regulado por mecanismos por ora desconhecidos. Esta compartimentação no núcleo das células dos animais superiores contribui em certa medida para o distinto funcionamento do respectivo genoma (Misteli, T., 2004) em função dos diversos graus de organização da cromatina (com graus de condensação muito diferentes, heterocromatina e eucromatina) e dos diversos subcompartimentos proteinaceos nucleares desprovidos de membrana, como é o caso dos nucléolos e de uma diversidade de pequenos corpos no interior desses núcleos. Os próprios cromossomas e a distribuição do material genético em cada cromossoma parecem ocupar territórios próprios dentro do espaço nuclear ocupando posições preferenciais em relação ao centro do núcleo e em relação com os diversos outros cromossomas. As propriedades de cada gene em cada cromossoma estarão dependentes da sua posição relativa em relação com o invólucro nuclear e com os vários domínios cromatínicos bem como com os diversos compartimentos proteináceos contidos no interior do núcleo.
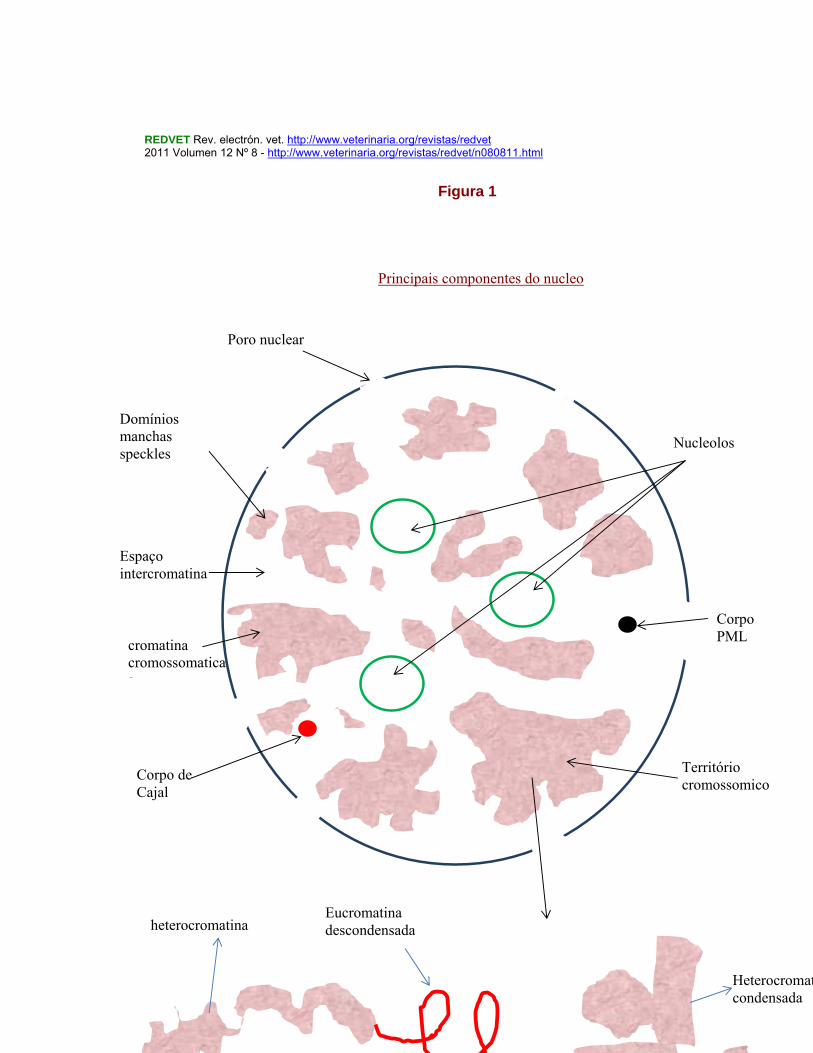
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
8
Nucleolos
Corpo PML
Território cromossomico
Corpo de Cajal
cromatina cromossomaticaa
Espaço intercromatina
Domínios manchas speckles
Poro nuclear
Principais componentes do nucleo
Eucromatina descondensada
Heterocromatina condensada
heterocromatina
Estrutura de cromatina
Figura 1
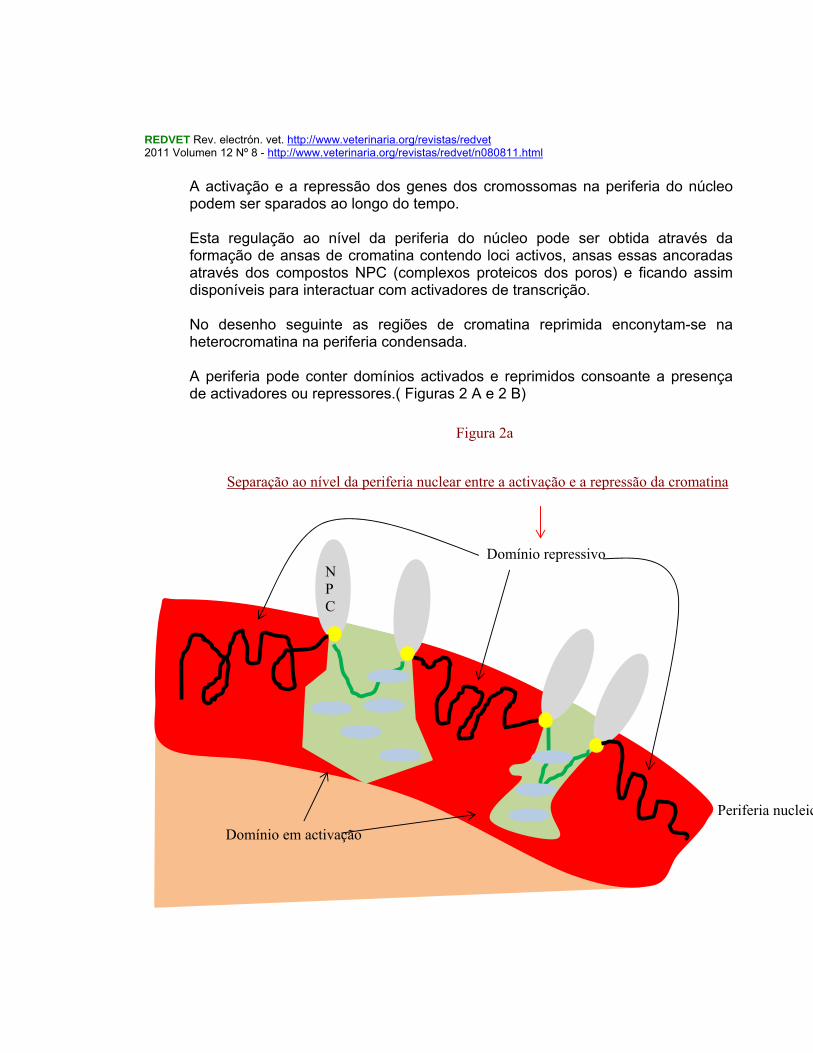
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
9
A activação e a repressão dos genes dos cromossomas na periferia do núcleo podem ser sparados ao longo do tempo. Esta regulação ao nível da periferia do núcleo pode ser obtida através da formação de ansas de cromatina contendo loci activos, ansas essas ancoradas através dos compostos NPC (complexos proteicos dos poros) e ficando assim disponíveis para interactuar com activadores de transcrição. No desenho seguinte as regiões de cromatina reprimida enconytam-se na heterocromatina na periferia condensada. A periferia pode conter domínios activados e reprimidos consoante a presença de activadores ou repressores.( Figuras 2 A e 2 B)
N P C
Domínio em activação
Domínio repressivo
Periferia nucleica
Separação ao nível da periferia nuclear entre a activação e a repressão da cromatina
Figura 2a

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
10
A zona periférica do núcleo adjacente ao invólucro nuclear assume particular relevo dentro deste âmbito pois é a zona onde parece decorrer a repressão da transcrição e onde se encontram grandes blocos de heterocromatina condensada reprimida, parecendo que quando da sua activação esses genes seriam reposicionados mais para o interior do núcleo. Parece pois que a periferia nuclear seria um ambiente favorecedor da repressão da transcrição dos genes cromossomais. 1.2 – Invólucro nuclear Nas células eucariotas as actividades nucleoplásmicas (no interior do núcleo) e citoplásmicas (no citosol) são separadas pelo invólucro nuclear.
NPC
Evolução ao nível da periferia nuclear entre a activação e a repressão Ansas de heterocromatina reprimida
Docagem das ansas aos poros nucleares
Factores de transcrição
activação Ansas de eucromatina activa
Figura 2b

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
11
O invólucro nuclear é uma estrutura com uma dupla membrana, em que a membrana ou folheto externo é contiguo com o retículo endoplásmico rugoso citoplásmico por vezes com ribossomas. Este folheto membranário mais externo (ONM) de membrana nuclear é funcionalmente semelhante ás membranas do retículo endoplásmico (ER) sendo separado do folheto membranário mais interno (INM) ou membrana nuclear interna pelo espaço perinuclear. Estes dois folhetos ou membranas cada um deles com uma dupla camada lipídica são funcionalmente e estruturalmente diferentes. Em certas zonas as duas membranas, do invólucro nuclear, fundem-se para originar os poros nucleares (NPC) compostos por cerca de 30 proteinas diferentes ( nucleoporinas), através das quais se faz o trânsito entre o núcleo e o citoplasma por mecanismos razoavelmente conhecidos. Por dentro do invólucro nuclear liga-se a lâmina nuclear periférica composta por lamínas (lamins) A/C e B que parecem regular a estrutura do invólucro nuclear e servirem de ancoradouro da cromatina à periferia do núcleo durante a interfase. No invólucro nuclear há pois que considerar a membrana nuclear externa, a membrana nuclear interna, a lâmina nuclear fibrosa e os complexos poros nucleares (NPC na figura anterior). 1.3 – Lâmina nuclear. Lamínas. Matriz A lâmína nuclear é uma estrutura fibrosa ( que faz parte da matriz nuclear) composta por cerca de cinco proteínas ( lamíns) A, B, C, D e E relativamente análogas aos filamentos intermediários. Recorda-se que o citoesqueleto das células eucariotas (também envolvido no transporte de vesículas pelas células) é uma estrutura dinâmica formada de três classes de conjuntos filamentosos montados, os microfilamentos, os filamentos intermediários e os microtúbulos. Os filamentos intermediários ( de 7 a 11 nm de diâmetro) contêm conjuntos de 2 ou 3 cadeias helicoidais alfa “enroladas” entre si ( two or three-stranded α helical coiled-coil cores) flanqueadas por diversas sequências de cada lado. Estudos de clonagem e sequenciação do cDNA das laminas ( Goldman, R.D. et al.2002) confirmaram que as lamins têm um domínio com estrutura típica como a dos filamentos intermediários inclusive domínios α-helical coiled-coil flanqueado por domínios não helicoidais.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
12
As lamins estão implicadas numa série de funções inclusive o montagem do invólucro nuclear, a síntese do DNA, a transcrição e a apoptose. Os genomas de diversos mamíferos revelam três genes para as lamins ( LMNA, LMNB 1 e LMNB2) codifricando sete isoformas spliced alternadamente. O tipo-A lamins A, A▲10, C e C2 são todos derivados do gene LMNA. O tipo-B lamins são B1 codificados pelo LMNB1 e B2-B3 codificado pelo LMNB 2. Além da lâmina periférica, há provas evidentes de que as lamins também se formam e existem no interior da estrutura nucleoplásmicas quer originando-se aqui em flocos distintos, ou como que constituindo um véu que preenche o nucleoplasma. Há diversas proteínas que interactuam com as lamins ( Cohen et al. 2001 in Goldman R. D. et al, 2002) e que podem ser distribuídas em dois grupos. O 1º grupo consiste de proteínas integrais da membrana nuclear interna ( Vide adiante) onde se incluem diversas isoformas da proteína 2 associada com lamin ( LAP2), LAP1, emerina, MAN1, otefin, receptor da lamin B (LBR), nurin , nesprin , RING finger-binding protein (RFBP), A-kinase-anchoring protein 149 (AKAP 149), p18 e provavelmente UNC-84. O 2º grupo contem proteínas que não são componentes integrais de membrana, mas que se encontram sobretudo na região da lãmina nuclear como por exemplo GCL, YA (young arrest), PP1 fosfatase e o factor de transcrição Oct 1. Durante a interfase, as lamins nucleares são continuamente sitetizadas e incorporadas na lamina. O envolvimento das lamins na síntese do DNA está assinalado parecendo que a replicação do DNA necessita de uma organização normal das lamins, parecendo haver um papel directo das lamins na síntese do DNA: Há sugestões de que os locais em que regiões da cromatina parecem ancoradas nas lamins estariam envolvidas na regulação da actividade da transcrição, embora se ignore como. As lamins podem interactuar com as sequências MARs e SARs do DNA (vide adiante). A expressão matriz nuclear não é um conceito, mas sim uma estrutura celular observável.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
13
A designação matriz nuclear corresponde a uma estrutura residual do núcleo após este ter sido extraído por solventes variados. Globalmente, na arquitectura do núcleo estão envolvidas estruturas sobrepostas digamos assim, e contendo ácidos nucleicos e que são a cromatina e a matriz nuclear. As estruturas não cromatinicas do núcleo após estudos de microscopia electrónica passaram a ser designados como constituindo a matriz nuclear. Apesar de estarem identificadas diversas proteínas da matriz nuclear, por enquanto não conhecemos como elas interactuam entre si para formar fibras, filamentos de 10 nm e como ocorre a montagem desta estruturas ou seja a arquitectura interior do núcleo(nucleoesqueleto) Estes complexos de fibras são feitos sobre “andaimes” de filamentos ramificados de 10 nm que se ligam com a lâmina nuclear. A ultraestrutura da matriz nuclear consiste da lâmina nuclear e de uma rede nuclear interna de subassociações ligadas entre si por fibras altamente estruturadas. A matriz nuclear possui na sua estrutura e composição, em grande parte, uma rede de ribonucleoproteínas ( RNP). Já há cerca de 30 anos se verificou que esta estrutura RNP constituída por RNA associado com proteínas, estava ligado a uma estrutura não cromatinica intranuclear. Hoje está esclarecido por T Cech e tal. que snRNP (small nucleic ribonucleoproteins) com cerca de 150 nucleótidos de comprimento são complexos de RNA proteínas que se combinam com pré-mRNA não modificado e várias outras proteínas para formar um “ spliceossoma” essencial para a remoção de intrões do pré-mRNA, uma modificação post-transcrição critica no núcleo das células eucariotas, As RNP e os RNA organizados com proteínas, estão ligados a uma estrutura intranuclear não cromatínica, sabendo-se por outro lado que as ligações periódicas e especificas das fibras de cromatina à matriz nuclear criam ou formam as ansas de cromatina ( loop domains). O núcleo é um organito dinâmico, cujas estruturas intranucleares, tais como a cromatina ou subassemblagem da matriz nuclear, se movimentam lentamente enquanto as suas moléculas constituintes se encontram num equilíbrio mais rápido com os “pools” nucleoplásmicos. As estruturas não cromatinicas do núcleo foram primeiramente observadas em secções fixadas dos tecidos e nesta forma, estas estruturas eram ambas estáticas e paradas.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
14
A lâmína nuclear reveste o lado nucleoplásmico do folheto membranário interno, e essa lâmina tem uma composição química que varia consoante o tipo celular e o seu estádio de diferenciação sendo as lâmínas o seu principal componente proteico. Por outro lado, como temos vindo a referir, além deste esqueleto periférico as proteínas lâmínas podem formar outras estruturas mais para o interior do núcleo. Associadas com as proteínas lamínas da lâmina nuclear encontram-se outras proteínas, a maioria das quais são proteínas integrais do folheto interno membranário (INM) onde existem, nos mamíferos, mais de 78 destas proteínas integrais (Margalit et, al. 2005). Estas proteínas membranárias integrais do invólucro nuclear interactuam com as proteínas Lamínas e regulam assim uma enorme variedade de interacções entre as membranas nucleares, a lamina nuclear, a cromatina e o DNA conforme se esquematiza em figura adiante. A lâmina nuclear é pois uma rede fibrosa anastomozada, ligando a superfície do nucleoplasma com a membrana nuclear interna, servindo para ligação da cromatina, ancoragem de filamentos intermediários citoplásmicos ao núcleo e estabilização da membrana nuclear (Georgatos, S.P. et, al., 1988). Resumindo, a membrana interna do invólucro nuclear liga-se portanto do lado do nucleoplasma a uma outra camada de finos filamentos que se distribuem à volta do núcleo excepto nos poros, constituindo a lâmina nuclear, servindo esses finos filamentos como estabilizantes, havendo sugestões acerca da sua semelhança com os filamentos delgados do citoesqueleto citoplásmico das células. Nas células dos mamíferos a lâmina é composta por mais 5 isotipos de lamínas (3 tipos da A e 2 tipos da B) As lâminas do tipo A e do tipo B são pois proteínas helicoidais filamentosas da lâmina nuclear e do nucleoplasma. As lâminas do tipo A encontram-se no interior do nucleoplasma e as laminas do tipo B estão próximas da membrana nuclear interna podendo ligar-se a proteínas integrais que se encontram embutidas nesta membrana nuclear interna. As lâminas estão implicadas na organização e funcionamento do núcleo. Antes e depois da mitose organizam-se e desorganizam-se através de fosfatações que convertem o invólucro em vesículas

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
15
enquanto a desfosfatação reverte este efeito reconstruindo a membrana. Defeitos associados com os genes codificadores das lâminas estão implicados numa série de patologias associadas, como é o caso das laminopatias (Mounkes e Stewart, 2004 citados em Jackson, D, 2005). As lamínas estão reconhecidas como tendo um papel directo na síntese do DNA, não se conhecendo o seu funcionamento específico durante a transcrição (Goldman, R. D. et, al., 2002) mas qualquer um destes processos resulta das interacções das lâmínas e outras proteínas associadas com a cromatina, havendo provas de que sequências MARs do DNA (matrix – attachement regions) e SARs (scaffold – attachment regions) interactuarem com as lâmínas (Goldman, R.D el at..2002). O “Scaffold” nuclear ou matriz nuclear como já referimos são estruturas proteinaceas que ficam após a extracção da membrana nuclear e das histonas e nele parece residir sobretudo uma proteína de 170.000 MW identificada como topoisomerase II, o que é no entanto contestado (Tsutsui, K. et, al. 1993). Nos cromossomas em metáfase também se refere um “Scaffold” do cromossoma como sendo uma estrutura contìnua revelada em cromossomas isolados na metáfase e após extracção severa. Também se refere que as topoisomerases se encontrariam nas ansas (“loop”) do DNA com uma frequência de três cópias de Top.II por cada 70 Kb de DNA. No DNA existiriam sequências dispersas que interactuariam especificamente com o “scaffold” nuclear. A associação do DNA nuclear com a matriz nuclear (scaffold) é pois realizada através de segmentos definidos do DNA, segmentos esses denominados MAR ( região de associação com a matriz ) ou SAR (região de interacção com o scaffold) (Tsutsui,K. et, al. 1993). Estas regiões de ligação da matriz nuclear como DNA (MAR) seriam as responsáveis pela organização da cromatina em domínios em ansas (loop). Os MARs são habitualmente encontrados em regiões não transcritas ricas em A-T (>70%) e têm cerca de 300-1000 bp de comprimento. Segmentos de DNA, MAR ou SAR têm sido identificados em diversas regiões dos genes. A coexistência ou justaposição de MAR/SAR com promotores/”enhancers” da transcrição e com origens de replicação está assinalada (Cockeril e Gamand; Gasser e Laemeli; Amati e Gasser;;DijKwell e Hamlim, citados em Tsutsui, et,al. 1993). Alguns pontos de cisão dos cromossomas também coincidem com a existência de MAR (Sperry et, al. citados em Tsutsui et, al. 1993).

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
16
Parece pois que o MAR está envolvido na organização em ansas do DNA nuclear, uma consequência directa da sua ligação com a matriz nuclear. Noutras proteínas referidas como envolvidas na ancoragem do DNA nuclear ao “scaffold” através dos segmentos MAR estão assinaladas por exemplo a proteína SP120, e a proteína ARBP e ainda a DNA topoisomerase II. Em relação à topoisomerase II há no entanto opiniões contraditórias sobre a sua preponderância. Quadro resumo das características das regiões MARs de interacção com a matriz nuclear ( Adaptado de Microsoft power point- Lecture 10- 2004).
1. São sequências dispersas no DNA do genoma, que interactuam especificamente com o “ scaffold” nuclear.
2. As matrizes de uma espécie são capazes de inter actuar com MARs de outras espécies
3. A homologia entre os MARs não é suficiente para permitir a hibridação cruzada.
4. Estão envolvidas na organização da cromatina em domínios em ansas.
5. Estão envolvidas na replicação do DNA e na expressão dos seus genes
6. O “scaffold” encontra-se por vezes apenas nos cromossomas em metáfase.
7. Os segmentos MAR cingem-se muitas vazes a locais específicos do DNA ricos em A-T (+ 70%) mas esta riqueza em A-T apenas, não é suficiente para a interação com a matriz.
8. Os MARs são mais frequentes em regiões do DNA não transcritas.
9. Os MARs tem habitualmente um comprimento de 300-1000bp.
1.4 – Interacções membrana nuclear interna – Lâmina nuclear – nucleoplasma. A Lâmina nuclear e o folheto interno membranário (INM) do invólucro nuclear constituem como que um esqueleto de filamentos proteicos à volta da periferia do núcleo. Na constituição deste esqueleto encontram-se sobretudo proteínas filamentosas intermediárias do tipo V ou sejam laminas A/C e B que formam uma rede complexa no interior da INM (Foisner, R., 2001). As lâminas do tipo B são essenciais para o desenvolvimento e são expressas ubiquamente, enquanto as do tipo A não são essenciais e

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
17
exprimem-se apenas em células diferenciadas sendo necessárias para manter a integridade de músculos e tecido adiposo. As lamíns são estruturas “coiled-coil) com uma pequena cabeça N-terminal seguida por um domínio em bastão (rod-coiled coil) e uma cauda globular C-terminal . Através destas regiões “coiled coil” das laminas elas podem formar dímeros paralelos e depois a partir destes dímeros originarem tetradas (anti-paralelas) e polímeros anti-paralelos (cabeças com caudas). (Figura 3) O aspecto estrutural mais notório das lamins A, B e C consiste numa região α-helicoidal de heptadas repetidas de ácidos aminados. A lamin A é gerada a partir da prelamin A, em quatro etapas pos tradução: farnesilação, libertação endoproteolitica dos últimos três ácidos aminados da proteína, metilação da farnesil-vcisteína C-terminal e finalmente libertação endoproteolítica dos últimos quinze ácidos aminados da proteína (incluindo o Ester metilfarnesilcisteína. Este esquema de processamento é crucial para a adesão da lamin A ao invólucro nuclear ( Coffinier, e tal. 2010) No Gallus gallus a lamin A apresenta 653 acidos aminados de comprimento contendo oito regiões em toda a sua extensão, ocupando as seguintes posições cada uma delas e com o respectivo comprimento em ácidos aminados Região Posição Comprimento Características 1 2-32 31 acid. aminad. Cabeça molecu. 2 33-382 350 “ Rod 3 33-69 37 “ Coil 1 A 4 70-79 10 “ Linker 1 5 80-217 138 “ Coil 1 B 6 218-241 24 “ Linker 2 7 242-382 141 “ Coil 2 8 383-657 275 “ Cauda A lamina nuclear é dinâmica e pode despolimerizar-se como sucede durante a mitose, para depois se polimerizar novamente quando reentrar na interfase celular, o que parece ser devido a fosfatações e desfosfatações nos resíduos que flaqueiam os domínios “coiled-coil” das laminas (Ref. Wilson et, al. 2001 in NPD nuclear compartments). O reassemble nuclear inicia-se quando as membranas nucleares se estruturam sobre a cromatina.
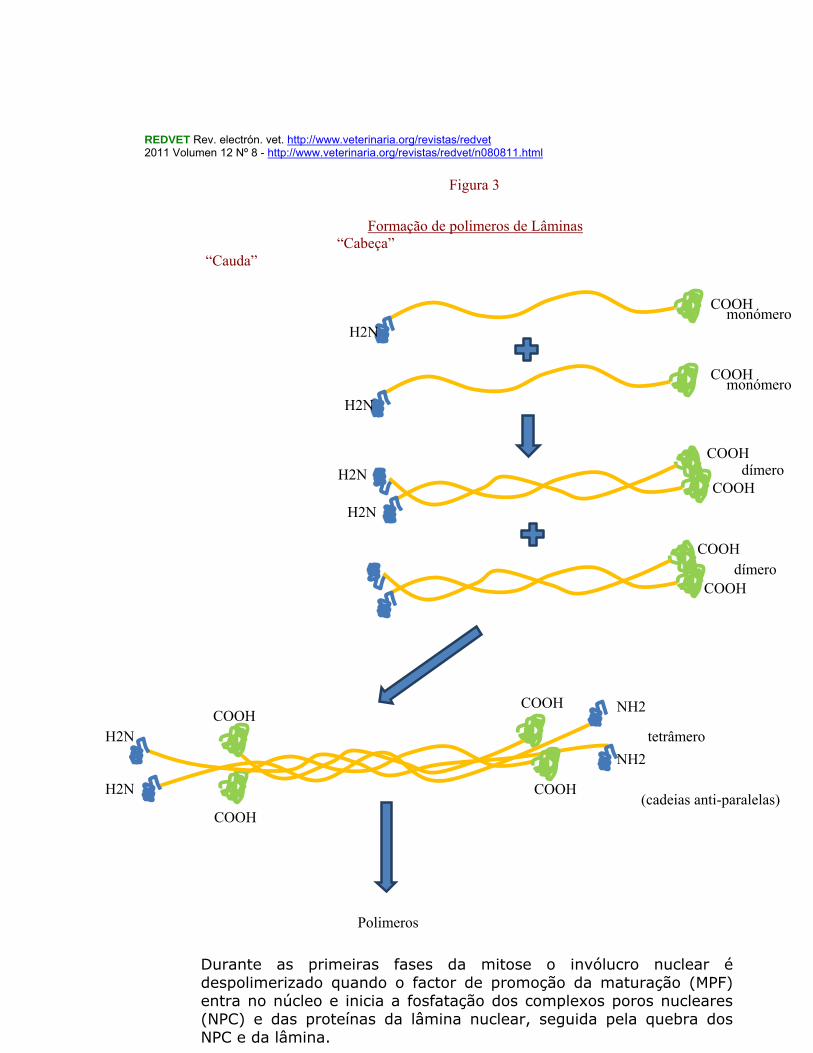
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
18
Durante as primeiras fases da mitose o invólucro nuclear é despolimerizado quando o factor de promoção da maturação (MPF) entra no núcleo e inicia a fosfatação dos complexos poros nucleares (NPC) e das proteínas da lâmina nuclear, seguida pela quebra dos NPC e da lâmina.
H2N
Formação de polimeros de Lâminas “Cabeça” “Cauda”
Polimeros
H2N
H2N
H2N
H2N
H2N
COOH
COOH
COOH
COOH
COOH
COOH
COOH
COOH
COOH
COOH
NH2
NH2
monómero
monómero
dímero
dímero
tetrâmero
(cadeias anti-paralelas)
Figura 3

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
19
O reassemble nuclear inicia-se quando as membranas nucleares se estruturam sobre a cromatina. Nas funções atribuídas à lamina nuclear incluem-se a manutenção da forma do núcleo, a organização dos poros nucleares, a regulação da transcrição, a ancoragem da heterocromatina na interfase, e ainda a replicação do DNA. Está também assinalada a presença de laminas (lamins) no interior do nucleoplasma parecendo que os poros por ela constituídos a este nível estariam implicados na replicação do DNA. Como já referimos, recentemente foram identificadas uma série de doenças (laminopatias) devidas a nutações nos genes quer das proteínas lâminas quer das proteínas que se associam a estas. Durante anos lâminas do tipo A e moléculas com elas associadas foram identificadas como envolvidas numa gama de desordens genéticas (Broers, J.L.V. et, al., 2004). Proteinas do folheto membranário interno (INM) do invólucro nuclear (adaptado de NPD nuclear componente: nuclear lamina)
- Lamins - LAP1 (lamina-associated protein 1) tem três isoformas α, β e ¥ - LAP2 – tem seis isoformas - Emerina - MAN1 - LBR (lamina β receptor) - Otefin - RFBP (ring finger binding protein) - Nurim
Algumas características destas proteÍnas integrais da INM
- RFBP – têm 9 regiões transmembranárias (TMR) - LBR – tem 8 ‘’ ‘’ ‘’ - Nurim – tem 5 ‘’ ‘’ ‘’ - MAN1 – tem 2 ‘’ ‘’ ‘’ - A Emerina, todas as LAP1 isoformas e 4 das 6 LAP2 isoformas (β, §, α e ¥) têm só uma região TMR - As LAP2 α não têm TMR - Todas as isoformas LAP1 e LAP2 α intereactuam preferencialmente com “lamins” do tipo A, enquanto as LBR e LAP2 β interactuam com “lamins” de tipo B e a Emerina interactua com ambos os tipos de “lamins”.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
20
As laminopatias recaiem em três classes, a das doenças que afectam os músculos estriados, as doenças que afectam os tecidos adiposos e as que envolvem as neuropatias periféricas. Estas doenças estão relacionadas com muito raros sÌndromas de envelhecimento precoce (Ref. 52 in Broers, et,. 2004). Referem-se nos dois quadros seguintes algumas proteínas que se encontram no folheto membranário interno (INM) do invólucro nuclear bem como algumas das suas características e nas duas figuras seguintes esquemas das proteínas da membrana interna do núcleo e das suas interacções com a lâmina nuclear e outros componentes nucleoplásmicos. Distribuição de algumas proteínas da membrana nuclear interna e outras proteínas com que interactuam da lâmina nuclear e do nucleoplasma.( Adaptado de Expert Reviews in Molecular Medicine. 2002 - Nurina - RFBP - Emerina (com domínios LEM) Proteínas integrais da - MAN 1 (com domínios LEM) Membrana nuclear interna - LAP-2 - (β, γ, δ, ε) (com domínios LEM) - LAP 1 - LBR Poteinas da lamina nuclear - Lamin B - Lamin A - Rush - RNA polimerase - Complexo 5remodelador da cromatina Proteínas do nucleoplasma - Complexo de “splicing” - BAF -pRb -GCL - DP - E2F - HA95 - HP1

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
21
Algumas interações entre algumas destas proteínas
- Nurim---------------Lamins - RFBP—Rush-----RNA polimerase Complexo remodelador da cromatina Complexo de “splicing” Lamins Cromatina - Emerina—BAF—Cromatina ---Lamins - MAN1—BAF—Cromatina --Lamins - LAP-2—Lamins (β,γ, δ,ε) --HA95 ---BAF --GCL—DP—E2F—pRb--Cromatina Interacções( +) entre diversas proteínas e componentes da membrana interna nuclear com a lâmina nuclear e outros componentes do nucleoplasma ( adaptado de Expert Reviews in molecular medicine, 2002 e de Froisner, R, 2001) Membrana (INM) Nucleoplasma Nurina →►Lamin A+ Lamin B RFBBP →►Rush+Cromatina+RNA polimerase+ Complexo
splicing+ Complexo remodelador da cromatina Emerin →►LEM domínio+BAF+BAF+BAF+LaminsA+B+Cromatina MAN1 →►Lem+ BAF+Cromatina +Lamins A+B LAP2 (β,γ,δ,ε) →►LEM+ BAF+ GCL+ DP+ E2F+ Cromatina+Lamins A+B LAP-1 →► Lamins A+ B LBR →►BAF+ HA95+ Cromatina+ Lamins A+B Cromatina+LAP-2α+ pRb+ BAF+Todos os outros Componentes antes assinalados Lamins A+B +pRb+ todos os outros componentes assinalados
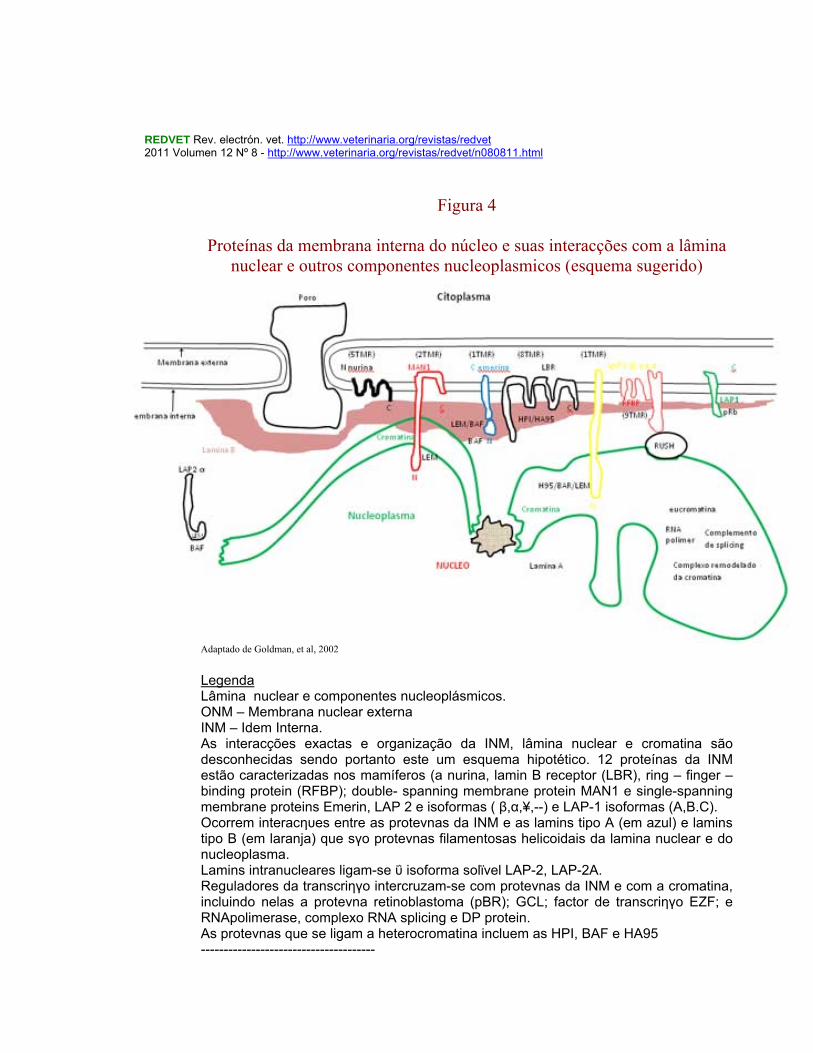
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
22
Figura 4
Proteínas da membrana interna do núcleo e suas interacções com a lâmina
nuclear e outros componentes nucleoplasmicos (esquema sugerido)
Adaptado de Goldman, et al, 2002 Legenda Lâmina nuclear e componentes nucleoplásmicos. ONM – Membrana nuclear externa INM – Idem Interna. As interacções exactas e organização da INM, lâmina nuclear e cromatina são desconhecidas sendo portanto este um esquema hipotético. 12 proteínas da INM estão caracterizadas nos mamíferos (a nurina, lamin B receptor (LBR), ring – finger – binding protein (RFBP); double- spanning membrane protein MAN1 e single-spanning membrane proteins Emerin, LAP 2 e isoformas ( β,α,¥,--) e LAP-1 isoformas (A,B.C). Ocorrem interacηυes entre as proteνnas da INM e as lamins tipo A (em azul) e lamins tipo B (em laranja) que sγo proteνnas filamentosas helicoidais da lamina nuclear e do nucleoplasma. Lamins intranucleares ligam-se ΰ isoforma solϊvel LAP-2, LAP-2A. Reguladores da transcriηγo intercruzam-se com proteνnas da INM e com a cromatina, incluindo nelas a proteνna retinoblastoma (pBR); GCL; factor de transcriηγo EZF; e RNApolimerase, complexo RNA splicing e DP protein. As proteνnas que se ligam a heterocromatina incluem as HPI, BAF e HA95 --------------------------------------
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
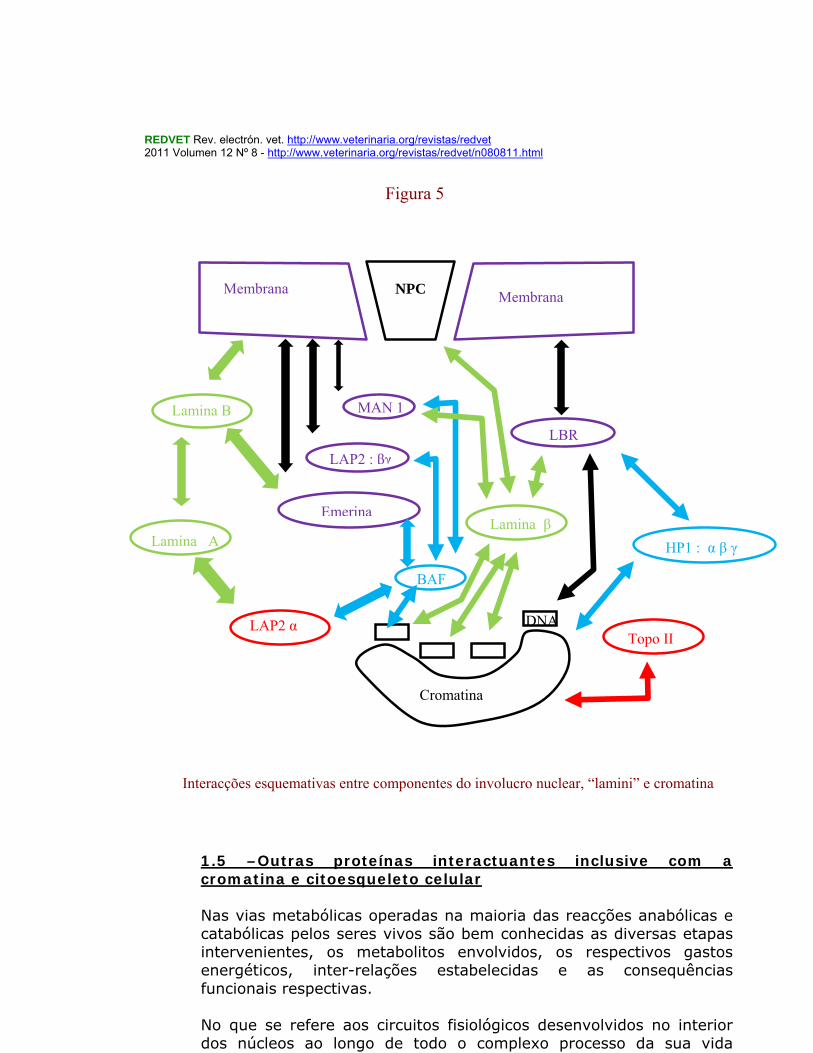
23
1.5 –Outras proteínas interactuantes inclusive com a cromatina e citoesqueleto celular Nas vias metabólicas operadas na maioria das reacções anabólicas e catabólicas pelos seres vivos são bem conhecidas as diversas etapas intervenientes, os metabolitos envolvidos, os respectivos gastos energéticos, inter-relações estabelecidas e as consequências funcionais respectivas. No que se refere aos circuitos fisiológicos desenvolvidos no interior dos núcleos ao longo de todo o complexo processo da sua vida
Lamina β HP1 : α β γ
NPC Membrana Membrana
MAN 1
LAP2 : βγ
Emerina
Lamina B
Lamina A
LAP2 α
Cromatina
Topo II
BAF
LBR
DNA
Figura 5
Interacções esquemativas entre componentes do involucro nuclear, “lamini” e cromatina

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
24
biológica, ainda se ignora muito acerca da sua modelação e regulação. Já se conhecem bastantes moléculas intervenientes, bem como alguns detalhes da sua morfofisiologia, mas ainda se não consegue reproduzir na totalidade todo o decurso operado entre uma determinada causa efectora e o seu consequente efeito. São alguns vislumbres deste complexo campo da biologia que procuramos desenvolver seguidamente. Quadro de interacções entre algumas proteínas LAP1 →► Lamins → ►pRb LAP2α →►pRB BAF Cromatina LBR →►HP1 HA95- Cromatina Lamins Na regulação da cromatina e da transcrição pelas proteínas da INM são conhecidas algumas interacções que são esquematizadas nos quadros seguintes. Regulação da cromatina e transcrição pelas proteínas da INM (NPD – nuclear compartment) -As lamins reprimem a transcrição através de interações da lamin A/C com a proteína retinoblastoma (Rb) e da LBR, a qual pode ligar-se com a HP1 (hetero-Chromatin protein 1) -A LBR e a LAP2β podem interactuar com a proteína cromossomal HÁ 95 -Várias outras proteínas, tais como todas as isoformas da LAP2, Emerina e MAN1 possuem uma região ou domínio LEM (de 43 àcidos aminados) na sua porção N-terminal que pode interactuar com uma proteína BAP (DNA-binding protein) -As lamins podem ainda interactuar com histonas âmago através das suas caudas C-terminais -Também a proteína da INM, RFBP, uma ATPase P-Tipo atípica pode interactuar directamente com a família RUSH dos factores de transcrição SWI/SNF-like. Varias proteínas nucleoplásmicas interactuam com as lamins e/ou proteínas integrais da INM, tais como: -Proteínas da replicação do DNA -RNA polimerase II transcrição -Complexos de splicing -Reguladores específicos da transcrição -Modificadores da cromatina

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
25
A cromatina pode interactuar com as seguintes proteínas, além de outras -Complexo remodelador da cromatina -RNA polimerase -Complexos de splicing -Lamins tipo A - LAP1 Tipo B -LAP1 Rush -BAF--------------Emerinas (dominis LEM) MAN1 (domínios LEM) -pRb - DP---------LAP2 (β,γ,δ,ε) (domínios LEM) -GCL “ -LAP2α -LBR -HP1 -HA95 -RFBP
Vejamos seguidamente algumas características de cada uma destas proteínas. -Rush, é um membro do grupo de proteínas da família Smarca 3 das máquinas remodeladoras de cromatina SWI-SNF, que interactua com uma proteína da membrana nuclear interna e ligando-se também no DNA do promotor proximal de pelo menos o gene uteroglobin. -BAF (Barrier-to-autointegration factor) é uma proteina celular altamente conservada que protége o DNA retroviral contra a autointegração. As interacções do BAF com o DNA são não específicas quanto a sequências do DNA. A interacção dupla do BAF com o DNA e com a LAP 2, uma proteína associada com a lâmina nuclear, sugere que ela tem um papel na organização do cromossoma.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
26
-LAP (Liver activator protein) é um membro da família C/EBP que induz a expressão específica de certos genes ao nível do promotor. -LAP 1 è um polipéptide associado com a lamina dentro do núcleo, localizada na interfase, na membrana nuclear interna, com as formas A e B interactuando com a lâmina A e funcionando na organização da membrana nuclear e na sua biogénese. As proteínas do invólucro nuclear estão envolvidas na regulação da transcrição -A LAP2β é uma proteína integral do invólucro nuclear contem um domínio LEM que se liga à cromatina reprimindo a actividade transcricional desencadeada pelo heterodímero E2F5 – DP3. -EMERIN (EMD –( Emery – Dreifuss muscular dystrophy) é uma proteína da membrana nuclear, rica em serina que medeia a ancoragem da membrana nuclear ao citoesqueleto citosolico podendo uma mutação do seu gene originar uma distrofia muscular. -MAN1 (ou LEMD3) é uma proteína integral da membrana interna nuclear cujas mutações originan osteopoikilosis Buschke – Alendorff Syndrome e melorheostosis. O seu domio C-terminal nucleoplásmico nos humanos liga-se a Smad 2 e Smad 3 e antagoniza os efeitos do factor de crescimento â (TGF-â). -pRb ou Rb (retinoblastona protein) é uma proteína supressora de tumores habitualmente presente como uma fosfoproteína dentro das células sendo uma alvo para fosfatação realizada por diversas cinases. A pRb liga-se e inibe factores de transcrição da família E2F que são dímeros de proteínas E2F e proteínas DP. -DP protein. As duas proteínas DP conhecidas, a TFDP 1, e 2, ligam-se à E2F para formar heterodímeros com alta afinidade para o DNA e com eficiente activação/repressão da transcrição. -GCL protein (Germ-cell-less) nas células de espécies mamiferas está co-localizado com a LAP2β no invólucro nuclear e actua como um componente da matriz nuclear e não como uma proteína integral da membrana. -LBR (Lamin β receptor) – é uma proteína multifuncional da membrana nuclear interna envolvida no assembly nuclear e na ligação à cromatina, ligando a heterocromatina e as lamins à membrana nuclear ,e passível de mutações que desencadeiam anomalias. -HPI –é uma proteína que se encontra associada com a heterocromatina podendo interactuar com outras proteínas o que a implica na regulação dos genes, replicação do DNA e arquitectura nuclear.

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
27
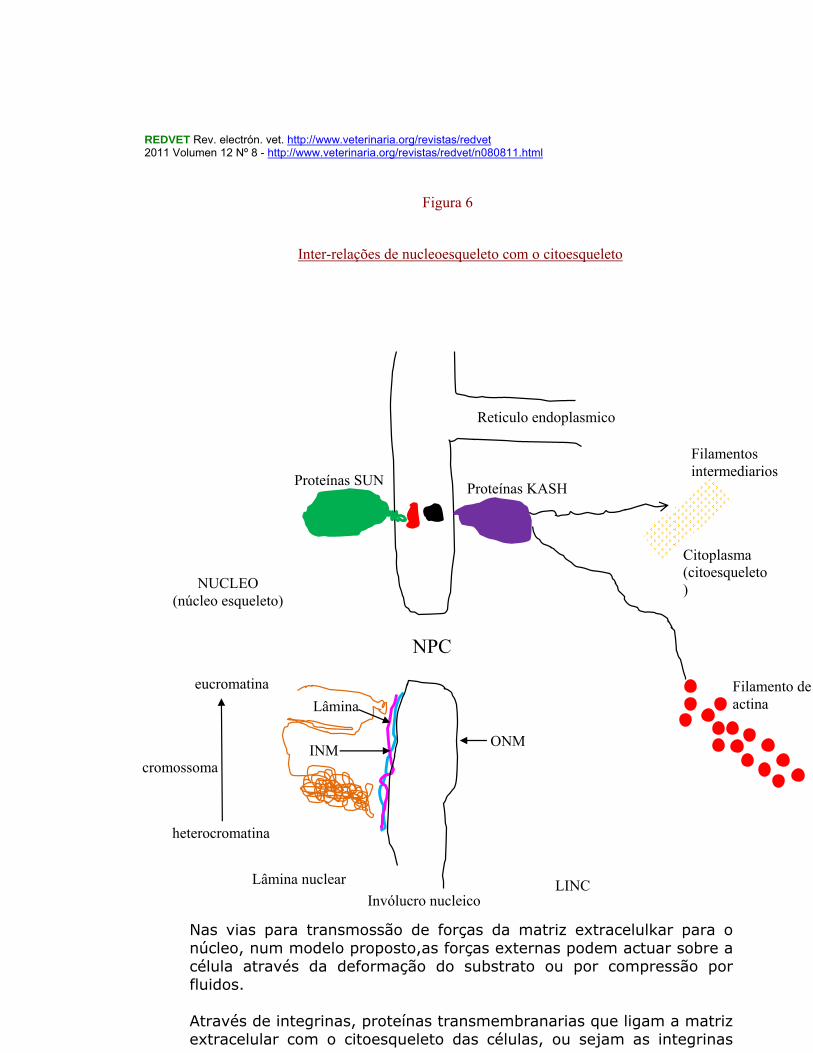
-RFBP (Ring-Finger binding protein) é uma proteina transmembranária da periferia nuclear, localizada na interfase, na membrana nuclear interna que interactua com os factores de transcrição RUSH. A expressão da RPBP é ubíqua e correlacionada com as das proteínas RUSH, sendo essa expressão regulada hormonalmente. As três proteínas integrais da membrana nuclear interna LAP2â e á, Emerina e MAN1 têm cerca de 70% de domínios LEM de 43 ácidos aminados idênticos que parecem servir como um novo domínio para ligação com a cromatina. Nas proteínas que se ligam à heterocromatina incluem-se as HP1, as BAF e as HÁ 95.. -HÁ 95 é uma proteína da cromatina e da matriz nuclear que regula a dinâmica do invólucro nuclear (Martins, S.B.et, al., 2000), Na interfase está firmemente associada com a cromatina e lâmina/matriz nuclear a à cromatina na mitose, formando um complexo com o receptor da lamins β (LBR) e com o LAP2 e Emerina, proteínas integrais da membrana nuclear interna. -E2F são uma família de factores de transcrição uns activadores e outros supressores, todos eles envolvidos na regulação do ciclo celular e na síntese do DNA nas células dos mamíferos. Os E2F ligam-se nas sequências promotoras alvo TTTCGCGC (E2F – Wikipedia, the free encyclopedia). Modelo provável da interação núcleoesqueleto-citoesqueleto( Starr e Fridolfson, 2010) e (Dahl, e tal.,2008) Interações entre o núcleo e o citoesqueleto celular são mediadas por pontes SUN-KASH no invólucro nuclear.As proteínas SUN (Sad1 e UNC-84) e as proteínas KASH (Klarscht, ANC-1 e Syne/Nesperin homology). Algumas combinações CASH_SUN ligam-se com microtúbulos, centrossomas e filamentos de actina a filamentos intermediários à superfície do núcleo. Está descrito um modelo referido como LINC, ou seja, um ligador do nucleoesqueleto com o citoesqueleto, constituído pelas proteínas KASH-SUN, que admite que as proteínas SUN (Sad1 e UNC-84) na membrana INM interactua com proteínas KASH na membrana ONM para abarcar o invólucro nuclear. As forças geradas no citoplasma são assim transferidas para a lamina nuclear. (Figura 6)
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
28
Nas vias para transmossão de forças da matriz extracelulkar para o núcleo, num modelo proposto,as forças externas podem actuar sobre a célula através da deformação do substrato ou por compressão por fluidos. Através de integrinas, proteínas transmembranarias que ligam a matriz extracelular com o citoesqueleto das células, ou sejam as integrinas
Inter-relações de nucleoesqueleto com o citoesqueleto
NPC
Proteínas KASH Proteínas SUN
Reticulo endoplasmico
ONM
Filamentos intermediarios
Filamento de actina
LINC Invólucro nucleico
Lâmina
INM
Lâmina nuclear
heterocromatina
eucromatina
cromossoma
NUCLEO (núcleo esqueleto)
Citoplasma (citoesqueleto)
Figura 6

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
29
ligam-se aos filamentos de actina já no citoesqueleto respondendo desta forma a ligandos extracelulares e a sinais intracelulares. No citoplasma celular os filamentos de actina, filamentos intermediários e microtúbulos podem interactuar com plectin e nesprin-3 nos ONM. Ao nível do invólucro nuclear, as nesprinas (através dos domínios KASH) interactuam com proteinasa SUN que atravessam o espaço perinuclear. Na INM, as proteínas SUN podem ligar-se a lamins e outras proteínas do invólucro nuclear, que por seu turno se podem ligar ao DNA e cromatina, completando-se assim o circuito de transmissão entre o núcleo e o citoesqueleto. 1.6 – Territórios cromossomais e sub organitos subnucleares (Carmo-Fonseca, 2002) O interior dos territórios cromossomais é percorrido por redes de múltiplos canais altamente ramificados e interligados tornando essas sequências genómicas profundamente inseridas nos cromossomas acessíveis a diversíssimos reguladores , quer activadores quer inibidores. Sucede ainda que dentro de cada cromossoma os respectivos braços são mantidos afastados um do outro e as regiões ricas em genes são separadas das regiões pobres em genes. Nos mamíferos as fibras cromatínicas estão dobradas sobre si próprias em domínios com diversos tamanhos (megabases) e ligados uns com os outros, cada um desses domínios com megabases talvez representando uma unidade funcional cuja replicação é coordenada e mantida em ciclos celulares sucessivos (Meaburn e Misteli, 2007). Os territórios cromossomais são dispostos de forma específica dentro dos núcleos, com uns cromossomas tendendo a situar-se no interior do núcleo e outros na periferia como já referimos; contudo a localização de um cromossoma nas células individuais dentro de uma dada população pode variar imenso sendo portanto mais um valor probabilístico do que um valor absoluto. Os arranjos cromossomais podem ainda depender do tipo de células e tipo de tecido podendo variar esses arranjos ao longo da diferenciação celular e desenvolvimento. Na estrutura dos cromossomas os componentes que determinam aspectos fundamentais do seu comportamento biológico são os centrómeros, os telómeros e as origens da replicação (ORI) do DNA. Os princípios básicos da estrutura e funcionamento dos centrómeros e telómeros são relativamente bem conhecidos mas o mesmo não sucede com as ORI. (Jackson, 2005).

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
30
A distribuição destas ORI ao longo do cromossoma tem sido estudada. Estas regiões interactuam com os complexos que as reconhecem (ORC) sendo a síntese de DNA organizada de tal forma que regiões específicas do genoma são replicadas em tempos precisos. Da dinâmica da cromatina sabe-se que os domínios dos genes são muito móveis e apresentam frequentemente movimentos de >500nm as longo de períodos de 10 segundos, enquanto os telómeros e centrómeros são relativamente imóveis (Jackson, 2005). Os telomeros contactam com a membrana nuclear através de um complexo que incorpora reguladores para silenciar a informação (Sir) e Ku. Os rápidos movimentos da cromatina observado nas leveduras está em oposição com a cromatina das células eucariotas que se encontra estavelmente associada com uma estrutura nuclear fixada, o que talvez se justifique por as leveduras não terem filamentos proteicos nucleares como a lamina nuclear que com as lamíns regula a arquitectura genómica nas células eucariotas. Recorda-se novamente que dentro do nucleoplasma os diversos cromossomas encontram-se em territórios próprios, parecendo que os genes residem à superfície destes territórios bem compactados (Cremer et, al., citados em Spector 2001). Na interfase, nos núcleos das células dos mamíferos os cromossomas não aparecem emparelhados. É também conhecido que em alguns tipos celulares a porção da heterocromatina (inactiva portanto) dos cromossomas, se encontra por dentro e associada com a lâmina nuclear, havendo também porções dessa heterocromatina localizadas em zonas mais do interior do núcleo associadas aquelas que se encontram na região pericentromérica, com um grupo de proteínas Polycomb (por ex. RING1, BMII e p PG2) constituindo os corpos PcG. Os corpos PcG nos compartimentos nucleares são locais onde se acumulam proteínas do grupo Polycomb (PcG) e por vezes situam-se próximo dos locais onde se encontra heterocromatina constitutiva (NPD Nuclear Compartments: PcG bodies) embora outras vezes tenham uma distribuição nuclear uniforme. Estes domínios PcG variam em número (desde 2 até centenas), tamanho e composição proteica não se sabendo bem como intervêm no silenciamento dos genes. Proteínas PcG e complexos destas proteínas como por ex: o PRC – 1 e PRC 2 estão envolvidos na modificação das histonas e em mecanismos epigenéticos como por ex: o “imprinting” e a inactivação do

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
31
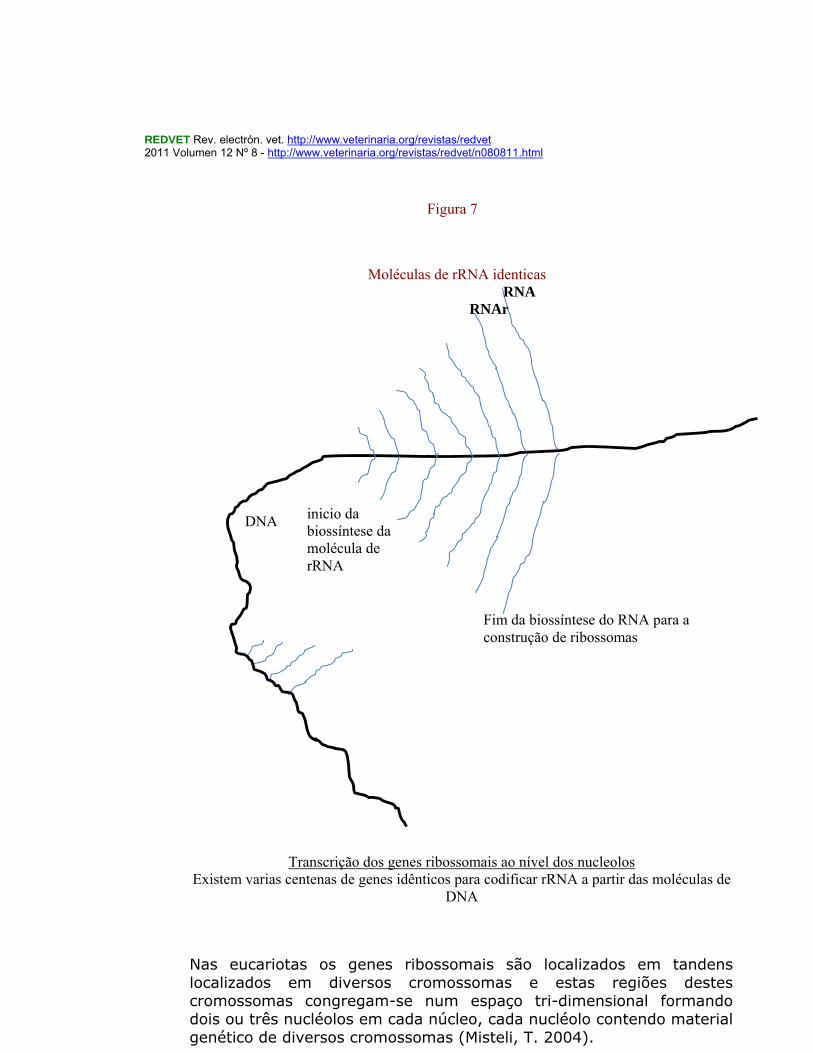
cromossoma X nas fêmeas. Este complexo PRC 2 metila a lisina 27 da histona H3 ( H3– K27). Os PRC 2 e PRC1 são silenciadores da cromatina. Os cromossomas formam portanto subestruturas distintas situadas em posições definidas (O’Brien, T.P. et, al.,2003) durante a progressão do ciclo celular e no núcleo em interfase. Os genes não se encontram distribuídos ao acaso sobre os cromossomas mas antes, como sucede com os genes transcritos em alto grau (como é o caso dos genes “house Keeping”), formando grandes aglomerados separados por outros domínios cromatínicos que possuem genes pouco ou nada expressos. Os complexos envolvidos na biogénese dos ribossomas existem sobretudo nos nucleolos. Os nucléolos onde ocorre a síntese de rRNA, e seu processamento e montagem em subunidades ribossomais encontram-se em número de 1 a 5 nas células dos animais mamíferos existindo neles três regiões bem distintas. Nos três compartimentos que se podem considerar nos nucléolos existe um compartimento fibrilhar central (com os genes r DNA) depois outro compartimento onde ocorre a maturação dos transcritos pré-r RNA e um terceiro compartimento onde ocorre a montagem das partículas dos ribossomas (Carmo-Fonseca, 2000) Nos nucléolos existem um conjunto de genes RNA ribossomais (rRNA) repetidos em tandem tendo por principal função sintetizar e processar rRNA (gerando os 5, 8S, 18S e 28S rRNA) e assemblar as subunidades ribossomais que são depois exportadas para o citoplasma para tomar parte na tradução do mRNA (Lamond & Sleeman, 2003). Os nucléolos durante cada ciclo celular são dinâmicos e ora se auto-organizam ora se desorganizam até estabilizarem. Genes ribossomais activamente transcritos são encontrados ao nível dos nucléolos.(Figura 7)
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
32
Nas eucariotas os genes ribossomais são localizados em tandens localizados em diversos cromossomas e estas regiões destes cromossomas congregam-se num espaço tri-dimensional formando dois ou três nucléolos em cada núcleo, cada nucléolo contendo material genético de diversos cromossomas (Misteli, T. 2004).
Moléculas de rRNA identicas RNA RNAr
DNA inicio da biossíntese da molécula de rRNA
Fim da biossíntese do RNA para a construção de ribossomas
Transcrição dos genes ribossomais ao nível dos nucleolos Existem varias centenas de genes idênticos para codificar rRNA a partir das moléculas de
DNA
Figura 7

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
33
Há um compartimento perinucleolar (PNC) e ainda os SAM 68 “nuclear body” (Huang, S. 2000, citado em Spector, DL, 2001), 1 a 10 por núcleo que se encontram à superfície dos nucléolos e têm um papel no metabolismo do RNA. Outros compartimentos já referidos assinalados no espaço nucleoplásmico entre a cromatina, são os “nuclear Speckles” (contendo os componentes Spliceossomais), os corpos de Cajal (envolvidos na biogénese dos sn RNP), os corpos nucleares PML (ricos em reguladores da transcrição) e uma série de inclusões nucleares (Carmo- Fonseca, M, 2002). Os corpos PML são preferencialmente localizados em regiões do genoma altamente activo transcriptionalmente. 10 a 30 corpos PML (Maul, G et, al. 2000, citados em Spector D.L. 2001) ND10 a POD ou Kr, encontram-se nos núcleos das células dos animais mamíferos, contendo diversas proteínas que parecem intervir na regulação da transcrição. Além dos domínios anteriormente referidos e que se encontram na generalidade dos núcleos das células dos mamíferos,existem outros domínios que são específicos de um dado tipo de células ou de um dado estádio fisiológico como por exemplo os corpos nucleares GATA-1, HSF 1, etc., Os factores responsáveis pelo “splicing” dos pré-mRNA encontram-se localizados em 25-50 manchas (domínios nucleares Speckles) ou dispersos em todo o nucleoplasma (Spector, D. L, 1993 citado em Spector 2001) correspondendo algumas dessas manchas maiores a conjuntos de grânulos intercromatínicos (IGC) parecendo estes intervir na montageme/ou modificação dos factores de “splicing” do pré-mRNA. Os “splicing speckles” são estruturas de forma irregular e de talhes diversos da região intercromatínica, sendo estruturas dinâmicas que contêm múltiplas cinases e fosfatases I parecendo participar na reciclagem dos spliceossomas Os locais de transcrição encontram-se disseminados no nucleoplasma, inclusive na periferia das IGC, constituindo diversos milhares de focos. Contudo diversos factores de transcrição, além de se mostrarem difusamente distribuídos, também se podem concentrar em 1 a 3 compartimentos chamados domínios OPT (OCTI/PTF/transcription). Estes compartimentos contêm transcritos em inicio e outros factores implicados no respectivo processamento (Pombo et, al., 1998, citados em Spector, 2001).

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
34
Estes domínios OPT surgem na fase G1 e por vezes residem próximo do nucléolo e desaparecem durante a fase S. Os domínios OPT são pois locais onde se concentram factores de transcrição necessários para as RNA polimerases. Os corpos de Cajal tendem a co-localizar-se com as histonas e com os genes U2 snRNA. Em diversos tipos de células também aqueles factores de splicing pré-m RNA se localizam em 1 a 10 corpos de Cajal que se pensa intervirem na biogénese de sn RNP e no seu tráfego (as subunidades spliceossomais U1, U2, U4/U6 e U5 e U7 localizam-se nesta estrutura), admitindo-se que possam mover-se através dos corpos de Cajal para as manchas (nuclear speckles) ou nucléolos. Os corpos de Cajal (Lamond & Sleeman, 2003) não contêm rRNA ou genes rRNA mas sim outros factores e por vezes encontram-se associados com loci de genes específicos, como genes das histonas e genes sn RNA, contendo uma pequena parte de “small nuclear” RNP (sn RNP) que são subunidades dos spliceossomas e também “small nucleolar” RNP (sno RNP) que modificam e cindem os pré-rRNA. Os corpos de Cajal, corpos espiralados, ou “coiled bodies” são pois ribonucleoproteínas em forma de bastonete curvado, envolvidos nos spliceossomas. Os corpos “Gems” ou seja Gemins ou gémeos dos corpos de Cajal encontram-se no nucleoplasma próximos dos corpos de Cajal e são do mesmo tamanho que estes e estão envolvidos nos spliceossomas. Diversos factores implicados especificamente na cisão e poliadenilação, no processamento dos pré-m RNA encontram-se dispersos no núcleo e também acantonados em 1 –4 focos nos corpos de quebra (Schal, W. et. Al. 1996, citados em Spector, 2001) que se sobrepõem ou ficam adjacentes aos corpos de Cajal. 1.7 – Movimentações Intranucleares A maioria das estruturas compartimentadas contidas no núcleo possuem uma certa capacidade de movimentação, embora restrita e um tanto ao acaso, como é o caso dos corpos PML e dos corpos de Cajal, que revelam no entanto uma certa direcção nesses movimentos ,que necessitam de energia, para ocorrer. Alguns destes compartimentos nucleares à medida que modificam a sua posição relativa dentro do núcleo podem deslocar-se para locais onde interactuam com outras populações moleculares como sucede

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
35
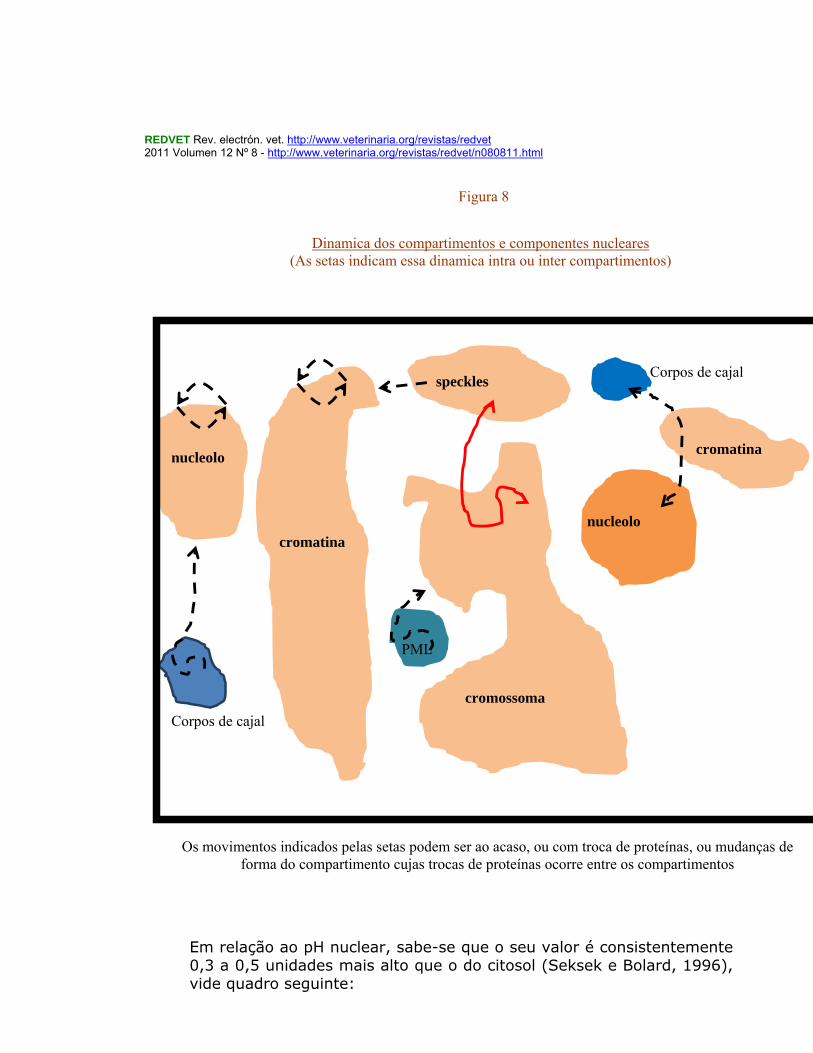
,por exemplo ,com outros domínios “manchas” (speckle domain) quando se dirigem para um território de um cromossoma. Há outras proteínas que se encontram em compartimentos específicos do núcleo e que aqui residem apenas alguns segundos ou minutos, migrando depois através do espaço intercromatínico, havendo no entanto proteínas que se deslocam entre diferentes tipos de compartimentos nucleares (O’Brien et, al., 2003). A localização e as potencialidades de deslocação dos cromossomas, de uns em relação aos outros, durante a interfase refletem-se, na composição dos cromossomas, e nas associações com o invólucro nuclear mediadas por proteínas. Em cada cromossoma existem uma série de conformações estruturais condensadas e não condensadas. Os complexos remodeladores da cromatina condicionam territórios dos cromossomas, e os espaços entre estes. A expressão dos genes ao nível dos cromossomas pode estar associada com a sua vizinhança de um poro do núcleo, ou com a sua localização na zona periférica do núcleo, ou com corpos nucleares específicos como por ex: os PML, ou ainda com o estado de condensação/descondensação da zona cromossomal em questão, e portanto a descondensação levando essa região fibrosa muito grossa de cromatina a tornar-se uma zona fibrosa muito mais fina susceptível de transcrição, inclusive com a formação de ansas de DNA que permitem a activação dos sistemas enzimáticos transcriptionais. Dinâmica dos compartimentos e componentes nucleares (Figura 8)
Os movimentos indicados pelas setas podem ser ao acaso, ou com troca de proteínas, ou mudanças de forma do compartimento, ou de troca de proteínas entre os compartimentos.(adaptado de O’Brien, et, al. 2003)
1.8 – Gradientes de pH intranucleares Admitimos que pequenas flutuações de pH a nível subcelular possam estar envolvidos na dinâmica observada no interior do núcleo. É sabido que o pH intracelular desempenha um papel fundamental no funcionamento celular, no crescimento e desenvolvimento dos animais. Organitos isolados, tais como mitocôndrias ou lisossomas têm gradientes de pH relativamente ao citosol envolvente.
REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
36
Em relação ao pH nuclear, sabe-se que o seu valor é consistentemente 0,3 a 0,5 unidades mais alto que o do citosol (Seksek e Bolard, 1996), vide quadro seguinte:
cromossoma
cromatina
speckles
nucleolo cromatina
nucleolo
Corpos de cajal
Corpos de cajal
Dinamica dos compartimentos e componentes nucleares (As setas indicam essa dinamica intra ou inter compartimentos)
Os movimentos indicados pelas setas podem ser ao acaso, ou com troca de proteínas, ou mudanças de forma do compartimento cujas trocas de proteínas ocorre entre os compartimentos
PML
Figura 8

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
37
Tipo de células Local pH (P<0,05) Nº de células Macrofagos Citoplasma 7.24+-0.05 80 Núcleo 7.55+-0.05 CHO Citoplasma 7.37+-0.18 30 Núcleo 7.63+-0.17 J774g8 Citoplasma 7.20+-0.27 31 Nuclep 7.68+-0.16 3 T 3 Citoplasma 7.49+-0.16 34 Núcleo 7.77+-0.08
Apesar da presença de poros na membrana nuclear, que parecia apontar para a livre difusão de iões e pequenas moléculas para dentro e para fora do núcleo, este trabalho avança que estas células através da membrana nuclear apresentam uma barreira à permeabilidade protonica do H+, indicando portanto restrições ao fluxo de iões monoatómicos. Há portanto um gradiente evidente de pH entre o núcleo e o citoplasma celular. Recorda-se que durante a mitose das células a membrana nuclear desagrega-se voltando a reorganizar-se posteriormente e curiosamente durante a mitose quando a membrana nuclear desaparece os cromossomas condensam-se e certamente nesta fase do ciclo celular o gradiente de pH entre o núcleo e o citoplasma altera-se. Será o pH celular ao nível cromatínico nesta altura do ciclo celular, mais acídico e terá alguma coisa a ver com a condensação cromossómica? Posteriormente, com o decorrer do ciclo celular a membrana nuclear volta a surgir com a descondensação dos cromossomas, ao mesmo tempo que certamente se restabelece o gradiente de pH entre o núcleo e o citosol, com este mais acidico e aquele menos acídico, inclusivamente com uma diferente dinâmica dos gradientes protónicos. Terá este gradiente de pH algo a ver com os domínios cromatínicos, o super enrolamento cromatínico, e os sistemas enzimáticos envolvidos na despolimerização/polimerização da remodelação da cromatina e modificação das histonas?

REDVET Rev. electrón. vet. http://www.veterinaria.org/revistas/redvet 2011 Volumen 12 Nº 8 - http://www.veterinaria.org/revistas/redvet/n080811.html
Introdução à morfologia e função nuclear http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf
38
Bibliografia
• Broers, J. L. N. et al. Laminopathies. Journal of Pathology, 2004, 204, 478-488.
• Carmo-Fonseca ,M. et al. To be or not to be in the nucleolus. Nat. Cell Biol. ,2000, 2. E. 107- E112
• Carmo-Fonseca. M..The contribution of nuclear compartimentalization to gene regulation. Cell ,2002,108. 513-521
• Coffinier, C. et al .Direct synthesis of lamin A by passing prelamin a processing, causes misshapen nuclei in fibroblast but no detectable pathology in mice.Journal of Biological Chemistry, 2010, 285 (27), 20818-26
• E2F- Wikipedia, the free encyclopedia Expert Reviews in Molecular Medicine. 2002 ( httppp://www.expertreviews.org)
• Foisner, R. Inner nuclear membrane proteins and the nuclear lamins.Journal of Cell Science. 2001, 114, 3791-3792.
• Georgatos, S. P. et al. Heterotypic and homotypic associations between the nuclear lamins: site-specificity and control by phosphorylation. PNAS, 1988, 85. 4325-4329.
• Goldman R. D. et al. Nuclear lamins: building blocks of nuclear architecture.Genes 6 Development. 2002, 16, 533-547
• Jackson, D.. Understanding nuclear organization when information become knowledge. Workshop of nuclear organization. Embo Reports, 2005, 6(nº), 213-217.
• Lamond, A. I. & Sleeman, J. E..Nuclear substructure and dynamics. Current biology, 2003, 13 (21):R 825- 828
• Margalit, A. et al. Breaking and making of the nuclear envelope, Journal of Cellular Biochemistry, 2005, 95, 454-465
• Martins, S. B. e tal. HA95 is a protein of the chromatin and nuclear matrix regulating nuclear envelope dynamics. Jounal of Cell Science, 2000,113,3703-3713
• Meaburn, K. J. e Misteli, T..Chromosome territories. Nature 2007,445, 379-381
• Misteli, T. Concepts in nuclear architecture. Bio Essays ,2005, 27, 477-487 • Misteli, T.. Spatial positioning: a new dimension in genome function, Cell,
2004,119,153-156 NPD nuclear compartments • O’Brien,T.P:, et al. Genome function and nuclear architecture: from gene
expression to nanosciences. Genome Research, 2003. 13 (6). 1029-1041 • Rzepecki, R. .The nuclear lamins and the nuclear envelope. Cellular &
Molecular Biology Letters, 2002, 7, 1019-1035. • Seksek, D. & Bolard, J. Nuclear pH gradient in mammalian cells science,
1996, 109, 257-262 • Spector, D. L., Nuclear domains,Journal of Cell Science, 2001, 114, 2891-
2893 • Tsutsui, K. et al, Identification and characterization of a nuclear scaffold
protein that binds the matrix attachement regin of DNA. Journal of Biological Chemistry 1993, 268, 17: 12886-12894.
REDVET: 2011, Vol. 12 Nº 8
Este artículo divulgativo está disponible en http://www.veterinaria.org/revistas/redvet/n080811.html concretamente en
http://www.veterinaria.org/revistas/redvet/n080811/081106.pdf REDVET® Revista Electrónica de Veterinaria está editada por Veterinaria Organización®. Se autoriza la difusión y reenvío siempre que enlace con Veterinaria.org®
http://www.veterinaria.org y con REDVET® - http://www.veterinaria.org/revistas/redvet











![Aplicações Farmacêuticas de Polímeros - SciELOmetacrilato, contendo ligações cruzadas por éteres alílicos do pentaeritritrol ou da alil sacarose[9,13,14]. A forma farmacêutica](https://static.fdocumenti.com/doc/165x107/5eaaa0f244a31b7bf416ca61/aplicaes-farmacuticas-de-polmeros-metacrilato-contendo-ligaes-cruzadas.jpg)






