
MODELLAZIONE DELLE VARIAZIONI ANNUALI DELLA BIOMASSA … ATTI/BMM 13(1) 2006 Trieste/pagine...
10
Biol. Mar. Medit. (2006), 13 (1): 61-70 F. CUMMAUDO, R. PASTRES, D. CURIEL 1 , A. SFRISO 2 Dipartimento di Chimica Fisica, Università Ca’ Foscari di Venezia, Dorsoduro 2137 - 30123 Venezia, Italia. [email protected] 1 SELC p.s.c.a.r.l., Via dell’Elettricità, 5d - 30175 Marghera-Venezia, Italia. 2 Dipartimento di Scienze Ambientali, Università Ca’ Foscari di Venezia, Dorsoduro 2137 - 30123 Venezia, Italia. MODELLAZIONE DELLE VARIAZIONI ANNUALI DELLA BIOMASSA DI FANEROGAME MARINE NELLA LAGUNA DI VENEZIA MODELLING ANNUAL VARIATIONS OF SEAGRASS BIOMASS IN THE LAGOON OF VENICE Abstract The seasonal evolutions of the two most abundant seagrass species in the Lagoon of Venice, Zostera marina L. and Cymodocea nodosa (Ucria) Ascherson, are investigated by means of two simple mathema- tical models, identified on the basis of the specific literature and calibrated using time series of site-specific data. The analysis of input data showed a remarkable spatial and interannual variability of the standing- crop, in particular for C. nodosa. The results obtained indicate that the reduction of irradiance intensity mainly due to an increase of water column’s depth and turbidity, could explain the different growth of Z. marina in the investigated areas. The interpretation of the results is more complex as regards C. nodosa that presents a radical apparatus very stable and a leaves development related to bathymetry. Key-words: seagrass biomass, time series, mathematical models, primary production, Lagoon of Venice. Introduzione Il ruolo ecologico delle praterie di fanerogame nelle aree costiere poco pro- fonde è largamente riconosciuto: esse formano habitat caratterizzati da un’elevata biodiversità (Borum et al., 2004; Leber, 1985) dove trovano rifugio numerosi orga- nismi, quali crostacei, molluschi, pesci ed avannotti. Da un punto di vista mor- fologico, la loro presenza stabilizza il sedimento costituendo così un importante fattore nel contrastare l’erosione del sedimento (Borum et al., 2004). Da un punto di vista funzionale, esse possono rappresentare una frazione rilevante della pro- duzione primaria in corpi idrici costieri (Sfriso e Ghetti, 1998), in quanto la loro densità di biomassa per unità di superficie è superiore rispetto a quella degli altri produttori primari marini (Duarte e Chiscano, 1999). Durante l’ultimo ventennio nella Laguna di Venezia si è assistito ad un notevole incremento della copertura di C. nodosa, ad una marcata ridistribuzione di Z. marina, che ha ricolonizzato alcune aree della Laguna centrale, e ad una notevole riduzione della copertura di Nanozostera noltii (Hornemann) (Sfriso et al., 2003). Contemporaneamente, si è ridotta di circa tre ordini di grandezza la biomassa delle macroalghe (in partico- lare Ulva laetevirens Areschoug), di gran lunga dominanti nella Laguna centrale negli anni ’80 (Sfriso e Marcomini, 1996). Attualmente, le fanerogame marine sono quindi considerate i principali produttori primari dell’ecosistema lagunare veneziano. Per queste ragioni, nell’ultimo decennio si sono intensificati gli studi volti a caratterizzare la loro distribuzione spaziale nella Laguna di Venezia ed a
-
Upload
vuongquynh -
Category
Documents
-
view
215 -
download
0
Transcript of MODELLAZIONE DELLE VARIAZIONI ANNUALI DELLA BIOMASSA … ATTI/BMM 13(1) 2006 Trieste/pagine...

Biol. Mar. Medit. (2006), 13 (1): 61-70
F. Cummaudo, R. PastRes, d. CuRiel1, a. sFRiso2
Dipartimento di Chimica Fisica, Università Ca’ Foscari di Venezia, Dorsoduro 2137 - 30123 Venezia, Italia.
[email protected] p.s.c.a.r.l., Via dell’Elettricità, 5d - 30175 Marghera-Venezia, Italia.
2Dipartimento di Scienze Ambientali, Università Ca’ Foscari di Venezia, Dorsoduro 2137 - 30123 Venezia, Italia.
MODELLAZIONE DELLE VARIAZIONI ANNUALI DELLA BIOMASSA DI FANEROGAME MARINE
NELLA LAGUNA DI VENEZIA
MODELLING ANNUAL VARIATIONS OF SEAGRASS BIOMASS IN THE LAGOON OF VENICE
AbstractThe seasonal evolutions of the two most abundant seagrass species in the Lagoon of Venice, Zostera
marina L. and Cymodocea nodosa (Ucria) Ascherson, are investigated by means of two simple mathema-tical models, identified on the basis of the specific literature and calibrated using time series of site-specific data. The analysis of input data showed a remarkable spatial and interannual variability of the standing-crop, in particular for C. nodosa. The results obtained indicate that the reduction of irradiance intensity mainly due to an increase of water column’s depth and turbidity, could explain the different growth of Z. marina in the investigated areas. The interpretation of the results is more complex as regards C. nodosa that presents a radical apparatus very stable and a leaves development related to bathymetry.
Key-words: seagrass biomass, time series, mathematical models, primary production, Lagoon of Venice.
IntroduzioneIl ruolo ecologico delle praterie di fanerogame nelle aree costiere poco pro-
fonde è largamente riconosciuto: esse formano habitat caratterizzati da un’elevata biodiversità (Borum et al., 2004; Leber, 1985) dove trovano rifugio numerosi orga-nismi, quali crostacei, molluschi, pesci ed avannotti. Da un punto di vista mor-fologico, la loro presenza stabilizza il sedimento costituendo così un importante fattore nel contrastare l’erosione del sedimento (Borum et al., 2004). Da un punto di vista funzionale, esse possono rappresentare una frazione rilevante della pro-duzione primaria in corpi idrici costieri (Sfriso e Ghetti, 1998), in quanto la loro densità di biomassa per unità di superficie è superiore rispetto a quella degli altri produttori primari marini (Duarte e Chiscano, 1999). Durante l’ultimo ventennio nella Laguna di Venezia si è assistito ad un notevole incremento della copertura di C. nodosa, ad una marcata ridistribuzione di Z. marina, che ha ricolonizzato alcune aree della Laguna centrale, e ad una notevole riduzione della copertura di Nanozostera noltii (Hornemann) (Sfriso et al., 2003). Contemporaneamente, si è ridotta di circa tre ordini di grandezza la biomassa delle macroalghe (in partico-lare Ulva laetevirens Areschoug), di gran lunga dominanti nella Laguna centrale negli anni ’80 (Sfriso e Marcomini, 1996). Attualmente, le fanerogame marine sono quindi considerate i principali produttori primari dell’ecosistema lagunare veneziano. Per queste ragioni, nell’ultimo decennio si sono intensificati gli studi volti a caratterizzare la loro distribuzione spaziale nella Laguna di Venezia ed a

F. Cummaudo, R. PastRes, d. CuRiel, a. sFRiso62
valutare in termini quantitativi le variazioni stagionali della loro biomassa (Bel-lato et al., 1995; Curiel et al., 1996a, 1996b; Rismondo et al., 1995, 1997, 2003; Sfriso et al., 1996, 1997, 1998, 1999, 2003, 2004). Le serie storiche di dati raccolti, relativi ad anni ed aree differenti sia per quanto riguarda la tessitura del sedi-mento sia per quanto riguarda le caratteristiche idrodinamiche, sono state par-zialmente interpretate anche mediante modelli matematici (Brigolin et al., 2004; Pastres et al., 2004; Zharova et al., 2001). In questo lavoro vengono presentati i primi risultati ottenuti nel tentativo di interpretare mediante semplici modelli di simulazione le evoluzioni stagionali osservate in Z. marina e C. nodosa. I modelli sono stati elaborati con gli obiettivi di: 1) stimare la produzione primaria ed i flussi di carbonio, azoto e fosforo veicolati dalle fanerogame nei cicli biogeochi-mici, 2) analizzare quantitativamente l’effetto della temperatura e della radiazione solare sui tassi di accrescimento.
Materiali e metodiI modelli qui presentati sono stati elaborati utilizzando serie storiche di dati
rilevate mensilmente nel periodo febbraio 1994 – giugno 2002 (Sfriso, 2000; Sfriso e Ghetti, 1998; Sfriso e Marcomini, 1997, 1999; Sfriso et al., 2004) nelle seguenti stazioni: Petta di Bò, Alberoni e S.Maria del Mare situate in Laguna sud e S.Nicolò situata in Laguna centrale e serie storiche rilevate dal Magistrato alle Acque di Venezia, nell’ambito del progetto di monitoraggio denominato MELa2, coordinato dal Concessionario del Magistrato alle Acque, Consorzio Venezia Nuova-Servizio Ambiente. Queste ultime rilevazioni sono state effettuate nella sta-zione di S.Pietro in Volta nel periodo febbraio 2002 – gennaio 2004 (Magistrato alle Acque, 2003, 2004).
Il campionamento e la determinazione della biomassa delle fanerogame marine è stato effettuato seguendo due metodologie leggermente diverse, riportate in det-taglio rispettivamente in Sfriso e Ghetti (1998) ed in Rismondo et al. (1997). Si ritiene tuttavia che la stima della biomassa ottenuta con questi metodi possa con-siderarsi consistente.
La struttura dei modelli matematici per le due specie oggetto di studio (Z. marina e C. nodosa) si basa su due sole variabili di stato: la biomassa del “com-parto” epigeo, indicata con “S” (shoots), e quella del “comparto” ipogeo, formato da rizomi e radici, indicato con “R” (root-rhizomes). L’evoluzione stagionale delle biomasse è condizionata da due “funzioni forzanti”: la temperatura dell’acqua (T) e la radiazione solare incidente che raggiunge le foglie (I), considerati i fattori chiave nella regolazione della crescita delle fanerogame (Pérez e Romero, 1992). La disponibilità dei nutrienti, azoto e fosforo, non è stata esplicitamente considerata un fattore limitante l’accrescimento, in quanto la loro concentrazione nelle acque interstiziali nella Laguna di Venezia è elevata (Zharova et al., 2001). Le equazioni di stato sono state integrate con passo orario, utilizzando il metodo Runge-Kutta del quarto ordine (Press et al., 1987). Il modello è stato forzato con i dati orari di temperatura dell’acqua ed irradianza, relativi ai periodi in cui sono stati rilevati i dati di biomassa. La temperatura dell’acqua è stata calcolata utilizzando i dati orari di temperatura dell’aria ed il modello di regressione presentato in Pastres et al. (2004). I dati di temperatura dell’aria come pure quelli di irradianza sono disponibili nel sito internet www.ibm.ve.cnr.it. I primi dati di ogni serie temporale di biomassa sono stati utilizzati come condizioni iniziali.

Modellazione delle variazioni annuali della biomassa di fanerogame marine nella laguna di Venezia 63
L’evoluzione temporale delle biomasse epigea ed ipogea di Z. marina è descritta dal seguente sistema di equazioni differenziali, i cui termini sono esplicitati in Appendice:
2
riguarda le caratteristiche idrodinamiche, sono state parzialmente interpretate anche mediante modelli matematici (Brigolin et al., 2004; Pastres et al., 2004; Zharova et al., 2001). In questo lavoro vengono presentati i primi risultati ottenuti nel tentativo di interpretare mediante semplici modelli di simulazione le evoluzioni stagionali osservate in Z. marina e C. nodosa. I modelli sono stati elaborati con gli obiettivi di: 1) stimare la produzione primaria ed i flussi di carbonio, azoto e fosforo veicolati dalle fanerogame nei cicli biogeochimici, 2) analizzare quantitativamente l’effetto della temperatura e della radiazione solare sui tassi di accrescimento.
Materiali e metodi I modelli qui presentati sono stati elaborati utilizzando serie storiche di dati rilevate
mensilmente nel periodo febbraio 1994 – giugno 2002 (Sfriso, 2000; Sfriso e Ghetti, 1998; Sfriso e Marcomini, 1997, 1999; Sfriso et al., 2004) nelle seguenti stazioni: Petta di Bò, Alberoni e S.Maria del Mare situate in Laguna sud e S.Nicolò situata in Laguna centrale e serie storiche rilevate dal Magistrato alle Acque di Venezia, nell’ambito del progetto di monitoraggio denominato MELa2, coordinato dal Concessionario del Magistrato alle Acque, Consorzio Venezia Nuova-Servizio Ambiente. Queste ultime rilevazioni sono state effettuate nella stazione di S.Pietro in Volta nel periodo febbraio 2002 – gennaio 2004 (Magistrato alle Acque, 2003, 2004). Il campionamento e la determinazione della biomassa delle fanerogame marine è stato effettuato seguendo due metodologie leggermente diverse, riportate in dettaglio rispettivamente in Sfriso e Ghetti (1998) ed in Rismondo et al. (1997). Si ritiene tuttavia che la stima della biomassa ottenuta con questi metodi possa considerarsi consistente. La struttura dei modelli matematici per le due specie oggetto di studio (Z. marina e C. nodosa) si basa su due sole variabili di stato: la biomassa del “comparto” epigeo, indicata con “S” (shoots), e quella del “comparto” ipogeo, formato da rizomi e radici, indicato con “R” (root-rhizomes). L’evoluzione stagionale delle biomasse è condizionata da due “funzioni forzanti”: la temperatura dell’acqua (T) e la radiazione solare incidente che raggiunge le foglie (I), considerati i fattori chiave nella regolazione della crescita delle fanerogame (Pérez e Romero, 1992). La disponibilità dei nutrienti, azoto e fosforo, non è stata esplicitamente considerata un fattore limitante l’accrescimento, in quanto la loro concentrazione nelle acque interstiziali nella Laguna di Venezia è elevata (Zharova et al., 2001). Le equazioni di stato sono state integrate con passo orario, utilizzando il metodo Runge-Kutta del quarto ordine (Press et al., 1987). Il modello è stato forzato con i dati orari di temperatura dell’acqua ed irradianza, relativi ai periodi in cui sono stati rilevati i dati di biomassa. La temperatura dell’acqua è stata calcolata utilizzando i dati orari di temperatura dell’aria ed il modello di regressione presentato in Pastres et al. (2004). I dati di temperatura dell’aria come pure quelli di irradianza sono disponibili nel sito internet www.ibm.ve.cnr.it. I primi dati di ogni serie temporale di biomassa sono stati utilizzati come condizioni iniziali.
L’evoluzione temporale delle biomasse epigea ed ipogea di Z. marina è descritta dal seguente sistema di equazioni differenziali, i cui termini sono esplicitati in Appendice:
Il termine μ rappresenta il tasso specifico di accrescimento di biomassa. Esso include il termine di respirazione e può quindi essere paragonato alla Produzione Primaria Netta per unità di biomassa. L’azione delle forzanti sul tasso di accrescimento viene simulata tramite
( ) SkStransdtdS
StransS ⋅Ω−⋅−=⋅Ω−−= μμ )1()(
RSkRStransdtdR
RtransR ⋅Ω−⋅⋅=⋅Ω−⋅= μ
(Eq.1)
(Eq.2)
(Eq. 1)
(Eq. 2)
Il termine µ rappresenta il tasso specifico di accrescimento di biomassa. Esso include il termine di respirazione e può quindi essere paragonato alla Produzione Primaria Netta per unità di biomassa. L’azione delle forzanti sul tasso di accre-scimento viene simulata tramite un modello di tipo moltiplicativo, in cui il tasso massimo (µmax) viene limitato in funzione della disponibilità di luce e temperatura (Bach, 1993) e della “capacità portante” del sistema. La frazione di produzione che viene trasferita dal comparto epigeo a quello ipogeo viene quantificata dal termine indicato con ktrans, mentre i termini di mortalità dei comparti epigeo ed ipogeo sono indicati rispettivamente con ΩS e ΩR.
Sia la letteratura che le osservazioni raccolte nella Laguna di Venezia indicano che l’evoluzione stagionale di C. nodosa si differenzia da quella di Z. marina per le caratteristiche di seguito evidenziate.- La biomassa epigea aumenta più rapidamente nel periodo tardo primaverile in
virtù dell’aumento del numero di germogli piuttosto che della biomassa fogliare di ciascun germoglio (Rismondo et al., 1997);
- La biomassa epigea diminuisce più rapidamente in autunno (Peduzzi e Vukovič, 1990).
Per tener conto di queste differenze, l’evoluzione stagionale delle biomasse è stata descritta mediante le seguenti equazioni di stato:
(Eq. 3)
3
un modello di tipo moltiplicativo, in cui il tasso massimo (μmax) viene limitato in funzione della disponibilità di luce e temperatura (Bach, 1993) e della “capacità portante” del sistema. La frazione di produzione che viene trasferita dal comparto epigeo a quello ipogeo viene quantificata dal termine indicato con ktrans, mentre i termini di mortalità dei comparti epigeo ed ipogeo sono indicati rispettivamente con ΩS e ΩR.
Sia la letteratura che le osservazioni raccolte nella Laguna di Venezia indicano che l’evoluzione stagionale di C. nodosa si differenzia da quella di Z. marina per le caratteristiche di seguito evidenziate. - La biomassa epigea aumenta più rapidamente nel periodo tardo primaverile in virtù dell’aumento del numero di germogli piuttosto che della biomassa fogliare di ciascun germoglio (Rismondo et al., 1997); - La biomassa epigea diminuisce più rapidamente in autunno (Peduzzi e Vukovič, 1990).
Per tener conto di queste differenze, l’evoluzione stagionale delle biomasse è stata descritta mediante le seguenti equazioni di stato:
La fase di reclutamento dei nuovi germogli viene interpretata dal termine indicato con RECL, che viene assunto proporzionale alla biomassa ipogea (R), secondo un coefficiente di trasferimento di biomassa dal comparto radici-rizomi a quello fogliare (KRS). Si è supposto che, così come accade per altre piante terrestri (de Beurs e Henebry, 2004), la fioritura inizi quando la pianta abbia accumulato un certo quantitativo di energia, che viene quantificato mediante i gradi-giorno, definiti come l’accumulo della temperatura media giornaliera tra due soglie termiche (Curry e Feldman, 1987). La fase di rapida diminuzione autunnale della biomassa fogliare invece è stata simulata aumentando il tasso di mortalità dei germogli in tale periodo, sulla base delle indicazioni emerse dall’analisi preliminare dei dati sperimentali.
Risultati Il modello di Z. marina è stato calibrato utilizzando dati mensili rilevati a Petta di Bò
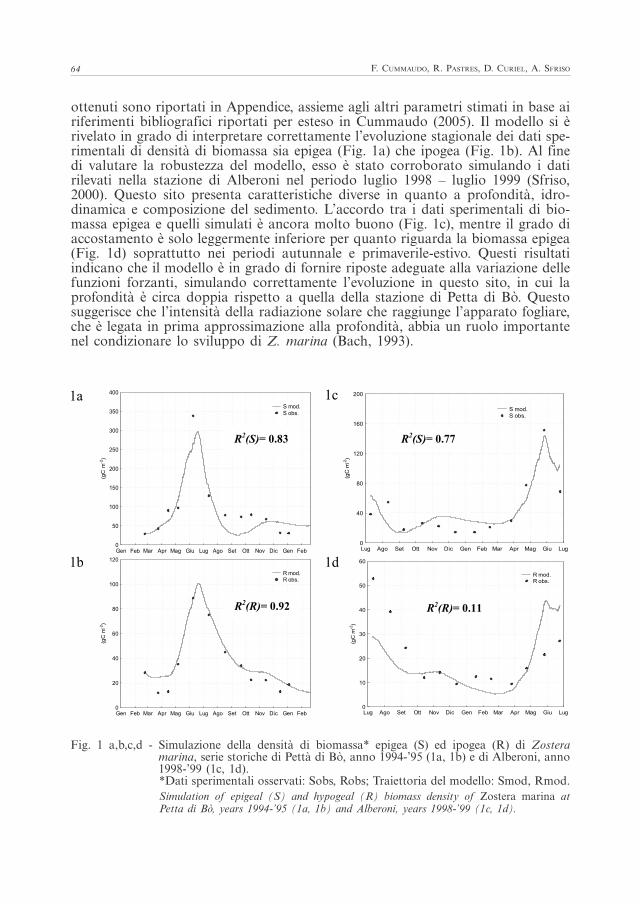
negli anni 1994-95 (Sfriso e Ghetti, 1998). I parametri μmax, Ktrans e σ così ottenuti sono riportati in Appendice, assieme agli altri parametri stimati in base ai riferimenti bibliografici riportati per esteso in Cummaudo (2005). Il modello si è rivelato in grado di interpretare correttamente l’evoluzione stagionale dei dati sperimentali di densità di biomassa sia epigea (Fig. 1a) che ipogea (Fig. 1b). Al fine di valutare la robustezza del modello, esso è stato corroborato simulando i dati rilevati nella stazione di Alberoni nel periodo luglio 1998 – luglio 1999 (Sfriso, 2000). Questo sito presenta caratteristiche diverse in quanto a profondità, idrodinamica e composizione del sedimento. L’accordo tra i dati sperimentali di biomassa epigea e quelli simulati è ancora molto buono (Fig. 1c), mentre il grado di accostamento è solo leggermente inferiore per quanto riguarda la biomassa epigea (Fig. 1d) soprattutto nei periodi autunnale e primaverile-estivo. Questi risultati indicano che il modello è in grado di fornire riposte adeguate alla variazione delle funzioni forzanti, simulando correttamente l’evoluzione in questo sito, in cui la profondità è circa doppia rispetto a quella della stazione di Petta di Bò. Questo suggerisce che l’intensità della radiazione solare che raggiunge l’apparato fogliare, che è legata in prima approssimazione alla profondità, abbia un ruolo importante nel condizionare lo sviluppo di Z. marina (Bach, 1993).
( ) RKSkRECLStransdtdS
RSStransS ⋅+⋅Ω−⋅−=+⋅Ω−−= μμ )1()(
RKRSkRECLRStransdtdR
RSRtransR ⋅−⋅Ω−⋅⋅=−⋅Ω−⋅= μ
(Eq. 3)
(Eq. 4) (Eq. 4)
La fase di reclutamento dei nuovi germogli viene interpretata dal termine indicato con RECL, che viene assunto proporzionale alla biomassa ipogea (R), secondo un coefficiente di trasferimento di biomassa dal comparto radici-rizomi a quello fogliare (KRS). Si è supposto che, così come accade per altre piante terrestri (de Beurs e Henebry, 2004), la fioritura inizi quando la pianta abbia accumulato un certo quantitativo di energia, che viene quantificato mediante i gradi-giorno, definiti come l’accumulo della temperatura media giornaliera tra due soglie ter-miche (Curry e Feldman, 1987). La fase di rapida diminuzione autunnale della biomassa fogliare invece è stata simulata aumentando il tasso di mortalità dei germogli in tale periodo, sulla base delle indicazioni emerse dall’analisi prelimi-nare dei dati sperimentali.
Risultati Il modello di Z. marina è stato calibrato utilizzando dati mensili rilevati a Petta
di Bò negli anni 1994-95 (Sfriso e Ghetti, 1998). I parametri µmax, Ktrans e σ così
F. Cummaudo, R. PastRes, d. CuRiel, a. sFRiso64
ottenuti sono riportati in Appendice, assieme agli altri parametri stimati in base ai riferimenti bibliografici riportati per esteso in Cummaudo (2005). Il modello si è rivelato in grado di interpretare correttamente l’evoluzione stagionale dei dati spe-rimentali di densità di biomassa sia epigea (Fig. 1a) che ipogea (Fig. 1b). Al fine di valutare la robustezza del modello, esso è stato corroborato simulando i dati rilevati nella stazione di Alberoni nel periodo luglio 1998 – luglio 1999 (Sfriso, 2000). Questo sito presenta caratteristiche diverse in quanto a profondità, idro-dinamica e composizione del sedimento. L’accordo tra i dati sperimentali di bio-massa epigea e quelli simulati è ancora molto buono (Fig. 1c), mentre il grado di accostamento è solo leggermente inferiore per quanto riguarda la biomassa epigea (Fig. 1d) soprattutto nei periodi autunnale e primaverile-estivo. Questi risultati indicano che il modello è in grado di fornire riposte adeguate alla variazione delle funzioni forzanti, simulando correttamente l’evoluzione in questo sito, in cui la profondità è circa doppia rispetto a quella della stazione di Petta di Bò. Questo suggerisce che l’intensità della radiazione solare che raggiunge l’apparato fogliare, che è legata in prima approssimazione alla profondità, abbia un ruolo importante nel condizionare lo sviluppo di Z. marina (Bach, 1993).
Fig. 1 a,b,c,d - Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Zostera marina, serie storiche di Pettà di Bò, anno 1994-’95 (1a, 1b) e di Alberoni, anno 1998-’99 (1c, 1d).*Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.Simulation of epigeal (S) and hypogeal (R) biomass density of Zostera marina at Petta di Bò, years 1994-’95 (1a, 1b) and Alberoni, years 1998-’99 (1c, 1d).
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
1
Fig. 1 a,b,c,d - Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Zostera marina, serie
storiche di Pettà di Bò, anno 1994-’95 (1a, 1b) e di Alberoni, anno 1998-’99 (1c, 1d). Simulation of epigeal (S) and hypogeal (R) biomass density of Zostera marina at Petta di Bò,
years 1994-’95 (1a, 1b) and Alberoni, years 1998-’99 (1c, 1d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
120
(gC
m-2
)
R mod. R obs.
R2(R)= 0.92
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
350
400
(gC
m-2
)
S mod. S obs.
R2(S)= 0.83
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
10
20
30
40
50
60
(gC
m-2
)
R mod. R obs.
R2(R)= 0.11
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
40
80
120
160
200
(gC
m-2
)
S mod. S obs.
R2(S)= 0.77
1a
1b
1c
1d
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
1
Fig. 1 a,b,c,d - Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Zostera marina, serie
storiche di Pettà di Bò, anno 1994-’95 (1a, 1b) e di Alberoni, anno 1998-’99 (1c, 1d). Simulation of epigeal (S) and hypogeal (R) biomass density of Zostera marina at Petta di Bò,
years 1994-’95 (1a, 1b) and Alberoni, years 1998-’99 (1c, 1d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
120
(gC
m-2
)
R mod. R obs.
R2(R)= 0.92
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
350
400
(gC
m-2
)
S mod. S obs.
R2(S)= 0.83
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
10
20
30
40
50
60
(gC
m-2
)
R mod. R obs.
R2(R)= 0.11
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
40
80
120
160
200
(gC
m-2
)
S mod. S obs.
R2(S)= 0.77
1a
1b
1c
1d
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
1
Fig. 1 a,b,c,d - Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Zostera marina, serie
storiche di Pettà di Bò, anno 1994-’95 (1a, 1b) e di Alberoni, anno 1998-’99 (1c, 1d). Simulation of epigeal (S) and hypogeal (R) biomass density of Zostera marina at Petta di Bò,
years 1994-’95 (1a, 1b) and Alberoni, years 1998-’99 (1c, 1d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
120
(gC
m-2
)
R mod. R obs.
R2(R)= 0.92
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
350
400
(gC
m-2
)
S mod. S obs.
R2(S)= 0.83
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
10
20
30
40
50
60
(gC
m-2
)
R mod. R obs.
R2(R)= 0.11
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
40
80
120
160
200
(gC
m-2
)
S mod. S obs.
R2(S)= 0.77
1a
1b
1c
1d
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
1
Fig. 1 a,b,c,d - Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Zostera marina, serie
storiche di Pettà di Bò, anno 1994-’95 (1a, 1b) e di Alberoni, anno 1998-’99 (1c, 1d). Simulation of epigeal (S) and hypogeal (R) biomass density of Zostera marina at Petta di Bò,
years 1994-’95 (1a, 1b) and Alberoni, years 1998-’99 (1c, 1d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
120
(gC
m-2
)
R mod. R obs.
R2(R)= 0.92
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
350
400
(gC
m-2
)
S mod. S obs.
R2(S)= 0.83
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
10
20
30
40
50
60
(gC
m-2
)
R mod. R obs.
R2(R)= 0.11
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
40
80
120
160
200
(gC
m-2
)
S mod. S obs.
R2(S)= 0.77
1a
1b
1c
1d
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
1
Fig. 1 a,b,c,d - Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Zostera marina, serie
storiche di Pettà di Bò, anno 1994-’95 (1a, 1b) e di Alberoni, anno 1998-’99 (1c, 1d). Simulation of epigeal (S) and hypogeal (R) biomass density of Zostera marina at Petta di Bò,
years 1994-’95 (1a, 1b) and Alberoni, years 1998-’99 (1c, 1d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
120
(gC
m-2
)
R mod. R obs.
R2(R)= 0.92
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
350
400
(gC
m-2
)
S mod. S obs.
R2(S)= 0.83
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
10
20
30
40
50
60
(gC
m-2
)
R mod. R obs.
R2(R)= 0.11
Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag Giu Lug0
40
80
120
160
200
(gC
m-2
)
S mod. S obs.
R2(S)= 0.77
1a
1b
1c
1d
Modellazione delle variazioni annuali della biomassa di fanerogame marine nella laguna di Venezia 65
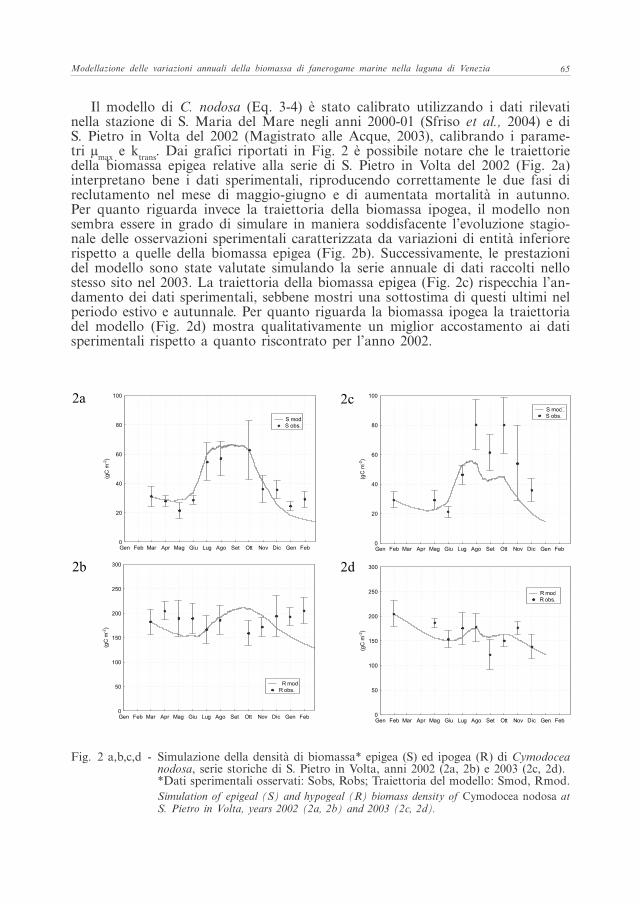
Il modello di C. nodosa (Eq. 3-4) è stato calibrato utilizzando i dati rilevati nella stazione di S. Maria del Mare negli anni 2000-01 (Sfriso et al., 2004) e di S. Pietro in Volta del 2002 (Magistrato alle Acque, 2003), calibrando i parame-tri µmax e ktrans. Dai grafici riportati in Fig. 2 è possibile notare che le traiettorie della biomassa epigea relative alla serie di S. Pietro in Volta del 2002 (Fig. 2a) interpretano bene i dati sperimentali, riproducendo correttamente le due fasi di reclutamento nel mese di maggio-giugno e di aumentata mortalità in autunno. Per quanto riguarda invece la traiettoria della biomassa ipogea, il modello non sembra essere in grado di simulare in maniera soddisfacente l’evoluzione stagio-nale delle osservazioni sperimentali caratterizzata da variazioni di entità inferiore rispetto a quelle della biomassa epigea (Fig. 2b). Successivamente, le prestazioni del modello sono state valutate simulando la serie annuale di dati raccolti nello stesso sito nel 2003. La traiettoria della biomassa epigea (Fig. 2c) rispecchia l’an-damento dei dati sperimentali, sebbene mostri una sottostima di questi ultimi nel periodo estivo e autunnale. Per quanto riguarda la biomassa ipogea la traiettoria del modello (Fig. 2d) mostra qualitativamente un miglior accostamento ai dati sperimentali rispetto a quanto riscontrato per l’anno 2002.
Fig. 2 a,b,c,d - Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Cymodocea nodosa, serie storiche di S. Pietro in Volta, anni 2002 (2a, 2b) e 2003 (2c, 2d).*Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.Simulation of epigeal (S) and hypogeal (R) biomass density of Cymodocea nodosa at S. Pietro in Volta, years 2002 (2a, 2b) and 2003 (2c, 2d).
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
2
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod S obs.
Fig. 2 a,b,c,d – Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Cymodocea nodosa, serie
storiche di S.Pietro in Volta, anni 2002 (2a, 2b) e 2003 (2c, 2d). Simulation of epigeal (S) and hypogeal (R) biomass density of Cymodocea nodosa at S.Pietro
in Volta, years 2002 (2a, 2b) and 2003 (2c, 2d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Tab. 1 – Produzione Primaria Netta di C. nodosa e Z. marina – Confronto con i valori di letteratura Net Primary Production of C. nodosa and Z. marina – Comparison with literature’s values.
Profondità Biomassa epigea Metodo** Sito Fonte
m gC m-2 a-1 1.5 1305 mod Laguna di Venezia Questo lavoro (dati 2002) 1 844 mar Laguna di Urbinu (Corsica) Agostini et al. (2003)* 1 303 mar Laguna di Venezia Rismondo et al. (1997) 4 153 mar Baia di Napoli (Ischia) Cancemi et al. (2002)*
0.8 163 mar Baia Alfaques (E Spagna) Perez e Romero (1994)* 4 176 var Golfo di Trieste Peduzzi e Vucokič (1990)*
C. n
odos
a
0.5 48.3 mar Mar Menor (SE Spagna) Terrados e Ros (1992)* 0.8 1177 mod Laguna di Venezia Questo lavoro (dati 1994-’95) 1 835 mar Laguna di Venezia Sfriso e Ghetti (1998) 413 mar Roscoff (Francia) Jacobs (1979)*
0.75 102 mod Lago Grevelingen (Olanda) Verhagen e Nienhuis (1983)* Z. m
arin
a
0.7 307 var Limfjorden (Danimarca) Olesen e Sand-Jensen (1994)* * Dati in g ps m-2 a-1 convertiti usando il contenuto di carbonio tessutale rilevato nella staz. di S.Maria del Mare. ** Metodo di stima: mod: modello matematico; mar: marcatura; var: variazione di biomassa.
2a
2b
2c
2d
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod2 R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod2 S obs.
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
2
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod S obs.
Fig. 2 a,b,c,d – Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Cymodocea nodosa, serie
storiche di S.Pietro in Volta, anni 2002 (2a, 2b) e 2003 (2c, 2d). Simulation of epigeal (S) and hypogeal (R) biomass density of Cymodocea nodosa at S.Pietro
in Volta, years 2002 (2a, 2b) and 2003 (2c, 2d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Tab. 1 – Produzione Primaria Netta di C. nodosa e Z. marina – Confronto con i valori di letteratura Net Primary Production of C. nodosa and Z. marina – Comparison with literature’s values.
Profondità Biomassa epigea Metodo** Sito Fonte
m gC m-2 a-1 1.5 1305 mod Laguna di Venezia Questo lavoro (dati 2002) 1 844 mar Laguna di Urbinu (Corsica) Agostini et al. (2003)* 1 303 mar Laguna di Venezia Rismondo et al. (1997) 4 153 mar Baia di Napoli (Ischia) Cancemi et al. (2002)*
0.8 163 mar Baia Alfaques (E Spagna) Perez e Romero (1994)* 4 176 var Golfo di Trieste Peduzzi e Vucokič (1990)*
C. n
odos
a
0.5 48.3 mar Mar Menor (SE Spagna) Terrados e Ros (1992)* 0.8 1177 mod Laguna di Venezia Questo lavoro (dati 1994-’95) 1 835 mar Laguna di Venezia Sfriso e Ghetti (1998) 413 mar Roscoff (Francia) Jacobs (1979)*
0.75 102 mod Lago Grevelingen (Olanda) Verhagen e Nienhuis (1983)* Z. m
arin
a
0.7 307 var Limfjorden (Danimarca) Olesen e Sand-Jensen (1994)* * Dati in g ps m-2 a-1 convertiti usando il contenuto di carbonio tessutale rilevato nella staz. di S.Maria del Mare. ** Metodo di stima: mod: modello matematico; mar: marcatura; var: variazione di biomassa.
2a
2b
2c
2d
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod2 R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100(g
C m
-2)
____ S mod2 S obs.
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
2
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod S obs.
Fig. 2 a,b,c,d – Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Cymodocea nodosa, serie
storiche di S.Pietro in Volta, anni 2002 (2a, 2b) e 2003 (2c, 2d). Simulation of epigeal (S) and hypogeal (R) biomass density of Cymodocea nodosa at S.Pietro
in Volta, years 2002 (2a, 2b) and 2003 (2c, 2d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Tab. 1 – Produzione Primaria Netta di C. nodosa e Z. marina – Confronto con i valori di letteratura Net Primary Production of C. nodosa and Z. marina – Comparison with literature’s values.
Profondità Biomassa epigea Metodo** Sito Fonte
m gC m-2 a-1 1.5 1305 mod Laguna di Venezia Questo lavoro (dati 2002) 1 844 mar Laguna di Urbinu (Corsica) Agostini et al. (2003)* 1 303 mar Laguna di Venezia Rismondo et al. (1997) 4 153 mar Baia di Napoli (Ischia) Cancemi et al. (2002)*
0.8 163 mar Baia Alfaques (E Spagna) Perez e Romero (1994)* 4 176 var Golfo di Trieste Peduzzi e Vucokič (1990)*
C. n
odos
a
0.5 48.3 mar Mar Menor (SE Spagna) Terrados e Ros (1992)* 0.8 1177 mod Laguna di Venezia Questo lavoro (dati 1994-’95) 1 835 mar Laguna di Venezia Sfriso e Ghetti (1998) 413 mar Roscoff (Francia) Jacobs (1979)*
0.75 102 mod Lago Grevelingen (Olanda) Verhagen e Nienhuis (1983)* Z. m
arin
a
0.7 307 var Limfjorden (Danimarca) Olesen e Sand-Jensen (1994)* * Dati in g ps m-2 a-1 convertiti usando il contenuto di carbonio tessutale rilevato nella staz. di S.Maria del Mare. ** Metodo di stima: mod: modello matematico; mar: marcatura; var: variazione di biomassa.
2a
2b
2c
2d
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod2 R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod2 S obs.
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
2
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod S obs.
Fig. 2 a,b,c,d – Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Cymodocea nodosa, serie
storiche di S.Pietro in Volta, anni 2002 (2a, 2b) e 2003 (2c, 2d). Simulation of epigeal (S) and hypogeal (R) biomass density of Cymodocea nodosa at S.Pietro
in Volta, years 2002 (2a, 2b) and 2003 (2c, 2d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Tab. 1 – Produzione Primaria Netta di C. nodosa e Z. marina – Confronto con i valori di letteratura Net Primary Production of C. nodosa and Z. marina – Comparison with literature’s values.
Profondità Biomassa epigea Metodo** Sito Fonte
m gC m-2 a-1 1.5 1305 mod Laguna di Venezia Questo lavoro (dati 2002) 1 844 mar Laguna di Urbinu (Corsica) Agostini et al. (2003)* 1 303 mar Laguna di Venezia Rismondo et al. (1997) 4 153 mar Baia di Napoli (Ischia) Cancemi et al. (2002)*
0.8 163 mar Baia Alfaques (E Spagna) Perez e Romero (1994)* 4 176 var Golfo di Trieste Peduzzi e Vucokič (1990)*
C. n
odos
a
0.5 48.3 mar Mar Menor (SE Spagna) Terrados e Ros (1992)* 0.8 1177 mod Laguna di Venezia Questo lavoro (dati 1994-’95) 1 835 mar Laguna di Venezia Sfriso e Ghetti (1998) 413 mar Roscoff (Francia) Jacobs (1979)*
0.75 102 mod Lago Grevelingen (Olanda) Verhagen e Nienhuis (1983)* Z. m
arin
a
0.7 307 var Limfjorden (Danimarca) Olesen e Sand-Jensen (1994)* * Dati in g ps m-2 a-1 convertiti usando il contenuto di carbonio tessutale rilevato nella staz. di S.Maria del Mare. ** Metodo di stima: mod: modello matematico; mar: marcatura; var: variazione di biomassa.
2a
2b
2c
2d
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod2 R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100(g
C m
-2)
____ S mod2 S obs.
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
2
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod S obs.
Fig. 2 a,b,c,d – Simulazione della densità di biomassa* epigea (S) ed ipogea (R) di Cymodocea nodosa, serie
storiche di S.Pietro in Volta, anni 2002 (2a, 2b) e 2003 (2c, 2d). Simulation of epigeal (S) and hypogeal (R) biomass density of Cymodocea nodosa at S.Pietro
in Volta, years 2002 (2a, 2b) and 2003 (2c, 2d). * Dati sperimentali osservati: Sobs, Robs; Traiettoria del modello: Smod, Rmod.
Tab. 1 – Produzione Primaria Netta di C. nodosa e Z. marina – Confronto con i valori di letteratura Net Primary Production of C. nodosa and Z. marina – Comparison with literature’s values.
Profondità Biomassa epigea Metodo** Sito Fonte
m gC m-2 a-1 1.5 1305 mod Laguna di Venezia Questo lavoro (dati 2002) 1 844 mar Laguna di Urbinu (Corsica) Agostini et al. (2003)* 1 303 mar Laguna di Venezia Rismondo et al. (1997) 4 153 mar Baia di Napoli (Ischia) Cancemi et al. (2002)*
0.8 163 mar Baia Alfaques (E Spagna) Perez e Romero (1994)* 4 176 var Golfo di Trieste Peduzzi e Vucokič (1990)*
C. n
odos
a
0.5 48.3 mar Mar Menor (SE Spagna) Terrados e Ros (1992)* 0.8 1177 mod Laguna di Venezia Questo lavoro (dati 1994-’95) 1 835 mar Laguna di Venezia Sfriso e Ghetti (1998) 413 mar Roscoff (Francia) Jacobs (1979)*
0.75 102 mod Lago Grevelingen (Olanda) Verhagen e Nienhuis (1983)* Z. m
arin
a
0.7 307 var Limfjorden (Danimarca) Olesen e Sand-Jensen (1994)* * Dati in g ps m-2 a-1 convertiti usando il contenuto di carbonio tessutale rilevato nella staz. di S.Maria del Mare. ** Metodo di stima: mod: modello matematico; mar: marcatura; var: variazione di biomassa.
2a
2b
2c
2d
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
50
100
150
200
250
300
(gC
m-2
)
____ R mod2 R obs.
Gen Feb Mar Apr Mag Giu Lug Ago Set Ott Nov Dic Gen Feb0
20
40
60
80
100
(gC
m-2
)
____ S mod2 S obs.

F. Cummaudo, R. PastRes, d. CuRiel, a. sFRiso66
I due modelli di Z. marina e C. nodosa sono stati utilizzati per ottenere una stima preliminare della loro Produzione Primaria Netta (PPN) e valutare le quan-tità di carbonio, azoto e fosforo da loro fissate annualmente per m2 di prateria pura nella Laguna di Venezia. La stima della PPN annuale è stata effettuata sulla base degli output del modello integrando i termini di produzione μS in Eq. 1 e 3.
In Tab. 1 viene presentato un confronto tra i valori di PPN reperiti in lettera-tura per le specie C. nodosa e Z. marina in diversi siti ed avvalendosi di metodi di stima differenti. Come si può vedere, i metodi empirici di stima della PPN (marcatura e variazione di biomassa) conducono a risultati in alcuni casi molto diversi tra loro. I valori ottenuti usando il modello sono superiori a quelli ripor-tati in letteratura per entrambe le specie. Il valore ottenuto per Z. marina non si discosta, tuttavia, marcatamente da quello riportato da Sfriso e Ghetti (1998) per la Laguna di Venezia, ottenuto mediante il metodo della marcatura.
Lo scostamento è maggiore per C. nodosa, la cui dinamica stagionale viene riprodotta in maniera meno accurata dal modello proposto. L’estrapolazione delle stime presentate in Tab. 1 all’intero bacino lagunare, effettuata tenendo conto della copertura relativa all’anno 2002 riportata in Rismondo et al. (2003), con-duce a stimare in 4.9⋅104 tC a-1 la quantità di carbonio riciclata annualmente nella Laguna di Venezia da Z. marina, valore superiore a quello riportato in Sfriso e Ghetti (1998) pari a 3.9⋅104 tC a-1, basato sulla precedente mappatura di Caniglia et al. (1992), ma dello stesso ordine di grandezza.
Tab. 1 - Produzione Primaria Netta di C. nodosa e Z. marina – Confronto con i valori di let-teratura.Net Primary Production of C. nodosa and Z. marina – Comparison with literature’s values.
Profondità Biomassa epigea Metodo** Sito Fontem gC m-2 a-1
C.
nodo
sa
1.5 1305 mod Laguna di Venezia Questo lavoro (dati 2002)
1 844 mar Laguna di Urbinu (Corsica) Agostini et al. (2003)*
1 303 mar Laguna di Venezia Rismondo et al. (1997)4 153 mar Baia di Napoli (Ischia) Cancemi et al. (2002)*
0.8 163 mar Baia Alfaques (E Spagna) Perez e Romero (1994)*4 176 var Golfo di Trieste Peduzzi e Vucokič (1990)*
0.5 48.3 mar Mar Menor (SE Spagna) Terrados e Ros (1992)*
Z.
mar
ina 0.8 1177 mod Laguna di Venezia Questo lavoro (dati 1994-’95)
1 835 mar Laguna di Venezia Sfriso e Ghetti (1998)413 mar Roscoff (Francia) Jacobs (1979)*
0.75 102 mod Lago Grevelingen (Olanda) Verhagen e Nienhuis (1983)*0.7 307 var Limfjorden (Danimarca) Olesen e Sand-Jensen (1994)*
* Dati in g ps m-2 a-1 convertiti usando il contenuto di carbonio tessutale rilevato nella staz. di S.Maria del Mare.
** Metodo di stima: mod: modello matematico; mar: marcatura; var: variazione di biomassa.
ConclusioniPer quanto riguarda Z. marina il modello è stato calibrato ottenendo un buon
accordo con la realtà sperimentale, e corroborato applicandolo ad una differente serie sperimentale. I risultati indicano che la struttura del modello è in grado di

Modellazione delle variazioni annuali della biomassa di fanerogame marine nella laguna di Venezia 67
descrivere correttamente l’andamento stagionale delle biomasse epigea ed ipogea. Per quanto riguarda C. nodosa il semplice modello qui proposto non può rite-
nersi del tutto soddisfacente, in quanto non è ancora in grado di simulare in maniera accurata l’evoluzione delle biomasse di entrambi i comparti. Sulla base di questi risultati, si possono quindi indicare alcune direzioni di approfondimento: a) la stima di alcuni parametri del modello tramite misure sperimentali specifiche; b) la quantificazione dell’influenza della profondità nel condizionare lo sviluppo dell’apparato fogliare; c) l’accoppiamento della dinamica di C. nodosa con quella delle epifite, che, sviluppandosi sulle sue foglie, ne riducono il tasso di fotosintesi (Brush e Nixon, 2002) ed agiscono come barriera all’assunzione di carbonio dal-l’acqua (Sand-Jensen, 1977).
Ciononostante, il confronto con i dati di letteratura, vedi Tab. 1, indica che entrambi i modelli possono essere utilizzati per stimare l’ordine di grandezza della Produzione Primaria Netta. A questo proposito è in via di implementazione un modello bidimensionale in grado di simulare l’evoluzione delle biomasse in fun-zione della profondità e della torbidità della colonna d’acqua.
SummaryIn this paper, the seasonal evolutions of the two most abundant seagrass species in the Lagoon of
Venice, Z. marina and C. nodosa, are investigated by means of two simple mathematical models. The models were identified on the basis of the specific literature and calibrated using time series of site-specific data.
The Z. marina model was succesfully calibrated and corroborated. The results of the corroboration indicate that the model enables one to predict the effect of a change in the level of irradiance on the biomass growth.
As regards C. nodosa, the results were not entirely satisfactory, since the simple two-variables model does not allow one to simulate accurately the evolution of the hypogeal biomass. On this basis, further investigation should address: the estimation of the model parameters on the basis of field and laboratory data, the quantification of influence of depth on leaves growth, the coupling of C. nodosa dynamic with that of epiphytes.
The models were used to obtain a preliminary estimation of the Net Primary Production of the pure meadows of the two species. Values of the same order of magnitude of those reported in the literature were obtained. The amounts of carbon, nitrogen and phosphorus annually fixed by Z. marina in the Lagoon of Venice extrapolated from the above estimation are comparable with the values obtained on the basis of field data.
BibliografiaBACH H.K. (1993) – A dynamic model describing the seasonal variations in growth and the
distribution of eelgrass (Zostera marina L.). I. Model theory. Ecological Modelling, 65: 31-50.BELLATO A., RISMONDO A., CURIEL D., MARZOCCHI M. (1995) - Finding of fruits
and germinanting seeds of Cymodocea nodosa (Ucria) Ascherson in the lagoon of Venice. Lavori Soc. Ven. Sc. Nat., 20: 165-166.
BORUM J., DUARTE C.M., KRAUSE-JENSEN D., GREVE T.M. (2004) - European sea-grasses: an introduction to monitoring and management. Managing and Monitoring of Euro-pean Seagrasses Project: 95.
BRIGOLIN D., PASTRES R., SFRISO A., CURIEL D. (2004) - Identificazione di un modello matematico per simulare la dinamica delle praterie di Cymodocea nodosa (Ucria) nella Laguna di Venezia. In: XIV Congresso Nazionale del SItE: 37.
BRUSH M.J., NIXON S.W. (2002) - Direct measurements of light attenuation by epiphytes on eelgrass Zostera marina. Marine Ecology Prog. Ser., 238: 73-79.

F. Cummaudo, R. PastRes, d. CuRiel, a. sFRiso68
CANIGLIA G., BORELLA S., CURIEL D., NASCIMBENI P., PALOSCHI F., RISMONDO A., SCARTON F., TAGLIAPIETRA D., ZANELLA L. (1992) – Distribuzione delle fanrero-game marine Zostera marina L., Zostera noltii Hornem., Cymodocea nodosa (Ucria) Ascher-son in laguna di Venezia. Lavori Soc. Ven. Sc. Nat., 1: 137-150.
CUMMAUDO F. (2005) – Tesi di laurea spec. in Scienze Ambientali: Modellazione dell’evolu-zione stagionale delle fanerogame marine in laguna di Venezia. Relatore Prof. Pastres, Uni-versità Ca’ Foscari di Venezia: 116 pp.
CURIEL D., BELLATO A., MARZOCCHI M., SOLAZZI A., SCATTOLIN M. (1996a) - Aspects of the distributive dynamics of the marine phanerogams in the Venice lagoon. Lavori Soc. Ven. Sc. Nat., 21: 39-51.
CURIEL D., BELLATO A., RISMONDO A., MARZOCCHI M. (1996b) - Sexual reproduc-tion of Zostera noltii Hornemann in the lagoon of Venice (Italy, north Adriatic). Aquat. Bot., 52: 313-318.
CURRY G.L., FELDMAN R.M. Mathematical foundation of population dynamics. The Texas Engineering Experiment Station monograph series, n. 3.
DE BEURS K.M., HENEBRY G.M. (2004) - Land surface phenology, climatic variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sensing of Environm., 89 (4), 497-509.
DUARTE C., CHISCANO C. (1999) - Seagrass biomass and production: a reassessment. Aquat. Bot., 65: 159-174.
LEBER K.M. (1985) - The influence of predatory decapods, refuge, and microhabitat selection on seagrass communities. Ecology, 66: 1951-1964.
MAGISTRATO ALLE ACQUE (2003) - Monitoraggio dell’Ecosistema Lagunare (MELa2) - 2° stralcio triennale. linea A. Rapporto annuale sulle misure periodiche delle caratteristiche fenologiche e dei parametri di crescita delle fanerogame marine (I anno). Consorzio Venezia Nuova. Esecutore SELC.
MAGISTRATO ALLE ACQUE (2004) - Monitoraggio dell’Ecosistema Lagunare (MELa2) - 2° stralcio triennale. linea A. Rapporto annuale sulle misure periodiche delle caratteristiche fenologiche e dei parametri di crescita delle fanerogame marine (II anno). Consorzio Venezia Nuova. Esecutore SELC.
PASTRES R., BRIGOLIN D., PETRIZZO A., ZUCCHETTA M. (2004) - Testing the robustness of primary production models in shallow coastal areas: a case study. Ecological Modelling, 179: 221-233.
PEDUZZI P., VUKOVIC A. (1990) - Primary production of Cymodocea nodosa in the Gulf of Trieste (Northern Adriatic Sea): a comparison of methods. Marine Ecology Prog. Ser., 64: 197-207.
PÉREZ M., ROMERO J. (1992) - Photosynthetic response to light and temperature of the sea-grass Cymodocea nodosa and the prediction of its seasonality. Aquat. Bot., 43: 51-62.
PRESS W.H., FLANNERY B.P., TEUKOLSKY S.A., VETTERLING W.T. (1987) - Numerical recipes, the art of scientific computing. Cambridge University Press: 795 pp.
RISMONDO A., CURIEL. D., MARZOCCHI M., MICHELI C. (1995) - Autoecologia e pro-duzione di Cymodocea nodosa nella Laguna di Venezia. Biol. Mar. Medit., 2 (2): 405-406.
RISMONDO A., CURIEL D., MARZOCCHI M., SCATTOLIN M. (1997) - Seasonal pattern of Cymodocea nodosa biomass and production in the lagoon of Venice. Aquat. Bot., 58: 55-64.
RISMONDO A., CURIEL D., SCARTON F., MION D., CANIGLIA D. (2003) - A new sea-grass map for the Venice lagoon. 6th Conference on the Medit. Coastal Environment, Med-coast 7-11 Oct. 2003, 2: 843-852.
SAND-JENSEN K. (1977) - Effect of epiphytes on eelgrass photosynthesis. Aquat. Bot., 3: 55-63.SFRISO A. (2000) – Eutrofizzazione e inquinamento delle acque e dei sedimenti nella parte
centrale della laguna di Venezia. MAV, CVN, Rapporto Finale, 3 vol.SFRISO A., GHETTI P.F. (1998) - Seasonal variation in biomass, morphometric parameters
and production of seagrasses in the lagoon of Venice. Aquat. Bot., 61: 207-223.SFRISO A., MARCOMINI A. (1996) - Decline of Ulva growth in the lagoon of Venice. Bior.
Tech., 58: 299-307.

Modellazione delle variazioni annuali della biomassa di fanerogame marine nella laguna di Venezia 69
SFRISO A., MARCOMINI A. (1997) - Macrophyte production in a shallow coastal lagoon. Part I: coupling with chemico-physical parameters and nutrient concentrations in waters. Mar. Envir. Res., 44 (4): 351-375.
SFRISO A., MARCOMINI A. (1999) - Macrophyte production in a shallow coastal lagoon. Part II: coupling with sediment, SPM, and tissue carbon, nitrogen and phosphorus concen-trations. Mar. Envir. Res., 47: 285-309.
SFRISO A., FACCA C., CEOLDO S. (2004) - Growth and production of Cymodocea nodosa (Ucria) Ascherson in the Venice lagoon. In: Campostrini, P. (ed), Scientific Research and Safeguarding of Venice. CoRiLa. Research Programme 2001 - 2002 - 2003. Results. Venice: 229-236.
SFRISO A., FACCA C., GHETTI P.F. (2003) - Temporal and spatial changes of macroalgae and phytoplankton in a Mediterranean coastal area: the Venice lagoon as a case study. Mar. Envir. Res., 56: 617-636.
ZHAROVA N., SFRISO A., VOINOV A., PAVONI B. (2001) - A simulation model for the annual fluctuation of Zostera marina biomass in the Venice lagoon. Aquat. Bot., 70: 135-150.

F. Cummaudo, R. PastRes, d. CuRiel, a. sFRiso70
AppendiceForme funzionali
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
..................................................... tasso specifico di accrescimento di biomassa
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
....................... termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
................... termini di mortalità del comparto epigeo ed ipogeo
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
................ funzione che modula l’influenza della temperatura sull’accrescimento
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
......................................funzione che modula l’influenza della temperatura sulla mortalità
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
...........................funzione che modula l’influenza dell’irradianza sull’accrescimento
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
..........................................................................................espressione dell’irradianza al fondo
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
...................................intensità luminosa di compensazione e di saturazione
Modellaz.delle variaz.ann. della biomassa di fan.marine nella Laguna di Venezia.
3
Appendice
FORME FUNZIONALI
)()()(max SfTfIf ⋅⋅⋅= μμ …..................................................tasso specifico di accrescimento di biomassa
μ⋅= transktrans ……………………..termine di trasferimento di biomassa dal comparto epigeo a quello ipogeo
)T(fRR;)T(fSR LOSS20RLOSS20S ⋅=Ω⋅=Ω ………..termini di mortalità del comparto epigeo ed ipogeo
( )⎪⎪⎩
⎪⎪⎨
⎧
>
≤=
⎟⎟⎠
⎞⎜⎜⎝
⎛−
−
⎟⎟⎠
⎞⎜⎜⎝
⎛ −
optm
opt0
TTk
TTkTf
stt
ToptmaxTToptT
stt
ToptTTopt
……….....funzione che modula l’influenza della temperatura sull’accrescimento
( ) 20TlossLOSS Tf −θ= ……………………………...funzione che modula l’influenza della temperatura sulla mortalità
( )
⎪⎪⎪
⎩
⎪⎪⎪
⎨
⎧
≥
<<−
−
≤
=
k
kcck
c
c
II1
IIIII
II
II0
If……………...…funzione che modula l’influenza dell’irradianza sull’accrescimento
zkweII ⋅−⋅= 0 …………………………………………………...………....….espressione dell’irradianza al fondo 20
20−⋅= T
ccc II θ ; 2020
−⋅= Tkkk II θ ……………………...intensità luminosa di compensazione e di saturazione
( )⎪⎪⎩
⎪⎪⎨
⎧
σ>
σ≤⎟⎠⎞
⎜⎝⎛
σ−
=
S0
SS1Sf
2
……………………………………………….funzione limitante della biomassa epigea
PARAMETRI DEI MODELLI DI Z. MARINA E C. NODOSA
Parametro Unità di misura
Valore modello** Descrizione
Cn 0.26 μmax* gg-1 Zm 0.19
Tasso di crescita specifico massimo dei ciuffi a 20°C
Cn 0.018 SR20 gg-1 Zm 0.023 Tasso di mortalità specifico massimo dei germogli a 20°C
Cn 0.009 RR20 gg-1 Zm 0.015 Tasso di mortalità specifico delle radici e dei rizomi a 20°C
Cn 0.64 ktrans* __ Zm 0.21 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello ipogeo
kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli Zm 587.01 σ* g C m-2 Cn # Capacità portante
Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°C Ic20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°C θc __ 1.17 Coefficiente di temperatura per l'intensità luminosa di compensazione θk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazione z m # Profondità kw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acqua k0 gg-1 0.01 Valore della f(T) a T=0°C km gg-1 0.00001 Valore della f(T) a T=Tmax
Cn 30 Topt °C Zm 21 Temperatura ottimale per la fotosintesi
Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibita stt __ 2 Coefficiente di forma della f(T) θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. ** I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.
..................................................................... funzione limitante della biomassa epigea
Parametri dei modelli di Z. marina e C. nodosa
ParametroUnità di misura
Valore modello**
Descrizione
μmax* gg-1 Cn 0.26 Tasso di crescita specifico massimo dei ciuffi a 20°CZm 0.19SR20 gg-1 Cn 0.018 Tasso di mortalità specifico massimo dei germogli a 20°CZm 0.023RR20 gg-1 Cn 0.009 Tasso di mortalità specifico delle radici e dei rizomi a 20°CZm 0.015
ktrans* __Cn 0.64 Coefficiente di trasferimento di biomassa dal comparto epigeo a quello
ipogeoZm 0.21kRS gg-1 Cn 0.003 Coefficiente di reclutamento di nuovi germogli
σ* g C m-2 Zm 587.01 Capacità portanteCn #Ik20 E m-2 gg-1 25.5 Intensità luminosa di saturazione a 20°CIc20 E m-2 gg-1 2.4 Intensità luminosa di compensazione a 20°Cθc __ 1.17 Coefficiente di temperatura per l’intensità luminosa di compensazioneθk __ 1.04 Coefficiente di temperatura per intensità luminosa di saturazionez m # Profonditàkw m-1 0.8 Coefficiente di estinzione della luce da parte dell’acquak0 gg-1 0.01 Valore della f(T) a T=0°Ckm gg-1 0.00001 Valore della f(T) a T=Tmax
Topt °C Cn 30 Temperatura ottimale per la fotosintesiZm 21Tmax °C 34 Temperatura massima oltre la quale la fotosintesi viene inibitastt __ 2 Coefficiente di forma della f(T)θloss __ 1.07 Coefficiente di Arrhenius
* Calibrato in questo lavoro. **I valori relativi alle due specie sono indicati con le iniziali: Cn: C. nodosa; Zm: Z. marina.


















