
METABOLISMO DI OLIGOSACCARIDI PREBIOTICI IN...
198
ALMA MATER STUDIORUM - UNIVERSITA’ DI BOLOGNA DOTTORATO DI RICERCA Biocatalisi Applicata e Microbiologia Industriale Ciclo XX Settore Scientifico Disciplinare di Afferenza: CHIM-11 METABOLISMO DI OLIGOSACCARIDI PREBIOTICI IN BIFIDOBACTERIUM PER IL POTENZIALE SVILUPPO DI NUOVI PRODOTTI ALIMENTARI FUNZIONALI Presentata da: SIMONA ZANONI Coordinatore del Dottorato: Chiar.mo Prof. DIEGO MATTEUZZI Relatore: Chiar.mo Prof. DIEGO MATTEUZZI Esame finale anno 2008
Transcript of METABOLISMO DI OLIGOSACCARIDI PREBIOTICI IN...

ALMA MATER STUDIORUM - UNIVERSITA’ DI BOLOGNA
DOTTORATO DI RICERCA
Biocatalisi Applicata e Microbiologia Industriale
Ciclo XX Settore Scientifico Disciplinare di Afferenza: CHIM-11
METABOLISMO DI OLIGOSACCARIDI PREBIOTICI IN BIFIDOBACTERIUM
PER IL POTENZIALE SVILUPPO DI NUOVI PRODOTTI ALIMENTARI FUNZIONALI
Presentata da: SIMONA ZANONI Coordinatore del Dottorato: Chiar.mo Prof. DIEGO MATTEUZZI
Relatore: Chiar.mo Prof.
DIEGO MATTEUZZI
Esame finale anno 2008


Meravigliarsi di tutto è il primo passo della ragione
verso la scoperta.
LOUIS PASTEUR


I
INDICE
INTRODUZIONE 1
1. L’ECOSISTEMA INTESTINALE 1 1.1. L’apparato gastrointestinale 1 1.2. L’ecosistema microbico 2 1.3. Principali pathway metabolici nel colon 6 2. PROBIOTICI 10 2.1. Effetti benefici sulla salute dell’ospite 10 2.2. Criteri per la scelta di un ceppo probiotico ideale 12 2.3. I batteri lattici 15 2.3.1. Il genere Lactobacillus 15 2.3.2. Streptococcus thermophilus 17 2.4. I bifidobatteri 19 2.4.1. Tassonomia, fisiologia e nutrizione 19 2.4.2. Ecologia 22 2.4.3. Proprietà prebiotiche 24 2.4.4. Catabolismo dei carboidrati in Bifidobacterium 26 2.4.5. Glicosil-idrolasi 27 2.4.6. Shunt dell’esoso fosfato 29 3. CARBOIDRATI PREBIOTICI 34 3.1. Frutto-oligosaccaridi e inulina 36

II
3.2. Oligosaccaridi della soia 38 3.3. Galatto-oligosaccaridi 39 3.4. Isomalto-oligosaccaridi 40 3.5. Xilo-oligosaccaridi e filano 41 3.6. Lattosaccarosio, lattulosio, lattitolo 42 3.7. Amido resistente 43 3.8. Fibra alimentare 44 4. SIMBIOTICI E ALIMENTI FUNZIONALI 45 4.1. I simbiotici 45 4.2. Gli alimenti funzionali 46
SCOPO DELLA TESI 51
MATERIALI E METODI 55 Parte prima 55 1. MICRORGANISMO E CONDIZIONI DI COLTURA 55 1.1. Microrganismo 55 1.2. Terreni di coltura 55 1.3. Reattivi e carboidrati 57 1.4. Bioreattore 59 2. CRESCITA DI B. ADOLESCENTIS MB 239 SU CARBOIDRATI PREBIOTICI 61 2.1. Cinetiche di crescita 61 2.2. Colture in bioreattore 61 3. TECNICHE ANALITICHE 63 3.1. Cromatografia planare HPTLC-AMD 63 3.2. Cromatografia HPLC 66 4. MODELLAZIONE MATEMATICA 66 5. ATTIVITA’ GLICOSIL-IDROLASICHE 71

III
5.1. Preparazione dei campioni 71 5.2. β-galattosidasi 72 5.3. β-xilosidasi 73 5.3.1. Dosaggio enzimatico e quantificazione proteica 73 5.3.2. Purificazione dell’enzima 74 5.3.3. Gel elettroforesi 75 5.3.4. Determinazione del peso molecolare 75 5.3.5. Effetto della temperatura e del pH sull’attività e stabilità dell’enzima 76 5.3.6. Valutazione della specificità di substrato 77 5.3.7. Determinazione dei parametri cinetici: km e Vmax 77 5.4. Xilanasi 78
Parte seconda 81 1. MICRORGANISMI E CONDIZIONI DI COLTURA 81 1.1. Ceppi e condizioni di coltura 81 1.2. Reattivi e carboidrati 82 2. RESISTENZA A SUCCHI GASTRICI E BILE 82 3. CINETICHE DI CRESCITA SU CARBOIDRATI PREBIOTICI 83 4. PROVE DI VITALITA’ DEI CEPPI PROBIOTICI IN
MATRICI ALIMENTARI 84 4.1. Sopravvivenza di Bifidobacterium lactis BAR 30 85 4.2. Vitalità di una miscela dei 3 ceppi probiotici in matrice
di cioccolato 86 5. TRIAL CLINICO 87
5.1. Trial clinico in vivo 88 5.2. Isolamento e conteggio di bifidobatteri e lattobacilli da feci 89

IV
RISULTATI 93 Parte prima 93
1. CRESCITA DI B. adolescentis MB 239 SU CARBOIDRATI PREBIOTICI 93 2. CARATTERIZZAZIONE FERMENTATIVA DI B. adolescentis MB 239 98 2.1. Determinazione del pH ottimale di crescita 98
2.2. Fermentazione di glucosio 101 2.3. Fermentazione di galattosio 103 2.4. Fermentazione di lattosio 105 2.5. Fermentazione di GOS 107
3. MODELLAZIONE MATEMATICA 110 3.1. Modellazione delle fermentazioni batch di B. adolescentis MB 239 110
3.2. Confronto tra i parametri fermentativi su glucosio, galattosio, lattosio e GOS 114
4. FERMENTAZIONE DI XILOSIO E XOS 117
5. PURIFICAZIONE DI UNA β-XILOSIDASI EXTRACELLULARE DI B. adolescentis MB 239 120 5.1. Determinazione del peso molecolare e dei parametri cinetici enzimatici 121 5.2. Attività enzimatica in funzione di pH e temperatura 122 5.3. Specificità di substrato 125
Parte seconda 127
1. SELEZIONE DEI MICRORGANISMI 127 2. DETERMINAZIONE IN VITRO DELLA TOLLERANZA AI SUCCHI GASTRICI E ALLA BILE 128 3. CINETICHE DI FERMENTAZIONE DI L. plantarum, S. thermophilus E B. lactis SU CARBOIDRATI PREBIOTICI 130

V
4. VITALITA’ DEI CEPPI PROBIOTICI IN MATRICI ALIMENTARI 134 4.1. Sopravvivenza di B. lactis BAR 30 134 4.2. Sopravvivenza di L. plantarum, S. thermophilus e B. lactis in matrice cioccolato 142 5. TRIAL CLINICO 146
DISCUSSIONE 151 CONCLUSIONI 163
BIBLIOGRAFIA 167


1
INTRODUZIONE
1. L’ECOSISTEMA INTESTINALE 1.1. L’apparato gastrointestinale
Il tratto gastrointestinale dell’uomo è costituito dalla cavità orale, dall’esofago, dallo stomaco, dall’intestino tenue (suddiviso in duodeno, digiuno e ileo) e dall’intestino crasso (suddiviso in cieco, colon e retto).
Le principali funzioni dell’apparato digerente sono la degradazione e l’assorbimento di componenti alimentari ed acqua. La degradazione avviene soprattutto nella porzione superiore del tratto gastrointestinale, mentre il luogo principale di assorbimento è rappresentato dall’intestino tenue e dall’intestino crasso. Tali processi sono favoriti dalla degradazione operata da un elevato numero di enzimi digestivi quali glicosidasi, lipasi, peptidasi e proteinasi.
Gli alimenti non digeriti e gran parte delle secrezioni intestinali raggiungono il crasso. Il colon è la sede della maggiore proliferazione microbica (si contano circa 1012 cellule per grammo di contenuto fecale) e in questa sede la maggior parte di tali composti viene degradata dalla microflora intestinale.

2
1.2. L’ecosistema microbico
Normalmente il tratto gastrointestinale è colonizzato da un microbiota autoctono che, se ben bilanciato, contribuisce in modo sostanziale allo stato di salute dell’ospite.
La colonizzazione è un processo molto complesso che inizia immediatamente dopo la nascita, quando il neonato viene a contatto con i microrganismi provenienti dal tratto genitourinario della madre e continua con lo sviluppo successivo di molte specie microbiche; conseguentemente, s’instaurano numerose interazioni tra i vari gruppi microbici e fra questi ultimi e l’ospite.
Un microrganismo in grado di colonizzare una superficie epiteliale possiede due importanti proprietà quali capacità di aderenza e di sopravvivenza (Savage et al. 1979). L’adesione viene realizzata attraverso specializzazioni di membrana, produzione di enzimi e tossine e mediante vantaggio metabolico nell’assorbimento e nell’utilizzo dei nutriliti disponibili. L’adesività batterica è inoltre influenzata dal cosiddetto “tropismo” attraverso il quale i batteri si legano a specifi recettori delle cellule ospiti in determinati siti anatomici che presentano le condizioni ambientali più favorevoli per la colonizzazione di alcuni gruppi di microrganismi e non di altri (Chisari et al. 1992). Le caratteristiche fisiologiche dell’ospite quali, la temperatura corporea, la resistenza agli acidi biliari, la composizione della dieta, l’uso di preparati microbiologici, l’uso di farmaci e la quantità di alimenti che ristagnano nel sito di colonizzazione sono fattori discriminanti il tropismo.
Ogni porzione del tratto gastrointestinale è colonizzata da una microflora specifica, la cui composizione è il risultato dell’adattamento alle condizioni ambientali locali e delle interazioni di tipo mutualistico, commensalistico o parassitico che si stabiliscono tanto tra i componenti della comunità microbica stessa quanto tra questa e l’organismo ospite.

3
Il pH, la presenza di enzimi gastrici e sali biliari, la velocità del transito peristaltico, il potenziale redox, la tensione di ossigeno disciolto e la concentrazione dei nutriliti definiscono la composizione e la concentrazione microbica totale della microflora nelle specifiche porzioni del tratto gastrointestinale.
Stomaco Digiuno Ileo Colon
Concentrazione batterica totale
0-3 0-6 3-7 10 - 12
Aerobi stretti o Anaerobi facoltativi
Enterobatteri 0 - 2 0 - 3 2 - 5 4 - 10
Streptococcus 0 - 3 0 - 4 2 - 6 5 - 10 Staphylococcus 0 - 2 0 - 3 2 - 5 4 - 7 Lactobacillus 0 - 3 0 - 4 2 - 5 6 - 10 Anaerobi Bacteroides Rari 0 - 2 3 - 6 9 - 11 Bifidobacterium Rari 0 - 3 3 - 7 8 - 10 Peptococcus Rari 0 - 3 3 - 4 8 - 10 Clostridium Rari Rari 3 - 4 6 - 10 Fusobacterium Rari Rari Rari 9 - 10 Eubacterium Rari Rari 3 - 5 9 - 10 Veillonella Rari 0 - 2 3 - 4 3 - 5 pH 2 - 2.4 7.5 6 5.5 - 7.5
Tab. 1 pH e composizione della flora batterica in stomaco, digiuno, ileo e colon. Le concentrazioni microbiche sono espresse in log10 CFU ml-1.
Come conseguenza di questi fattori ambientali locali la popolazione
dello stomaco e dell’intestino tenue è scarsa e composta prevalentemente da

4
batteri aerobi stretti o anaerobi facoltativi. Il colon, dove si contano fino a 1012 cellule per grammo di contenuto intestinale, è invece sede della maggiore concentrazione microbica. Il microbiota del colon è costituito da oltre 400 specie batteriche, delle quali solo circa 200 sono state caratterizzate, e rappresenta pertanto uno dei più complessi ecosistemi batterici conosciuti (tab. 1). Le informazioni sulla diversità microbica entro questa complessa comunità hanno avuto negli ultimi anni una grande implementazione grazie alle tecniche molecolari che si basano sull’analisi dell’rRNA 16S (Dethlefsen et al. 2006; Flint 2006). Tale analisi ha indicato che nell’ecosistema microbico intestinale più del 75% dei filotipi ritrovati nell’intestino crasso non corrisponde alle specie microbiche note e coltivabili (Flint et al. 2007; Hold et al. 2002).
A causa dell’attività respiratoria dei batteri aerobi e/o anaerobi facoltativi che esaurisce l’ossigeno presente nel lume intestinale, il colon è un ecosistema strettamente anaerobico, con un basso potenziale di ossidoriduzione, caratterizzato da una netta preponderanza dei batteri anaerobici obbligati.
La microflora autoctona dell’intestino crasso è molto stabile ed esercita un’efficace esclusione competitiva che preclude la colonizzazione dell’ecosistema da parte di microrganismi alloctoni. I gruppi batterici predominanti in questo complesso sistema sono i batteri Gram + a basso contenuto in G + C (Firmicutes)e i Gram – Bacteroidetes. Il 72% dei 395 filotipi identificati da Eckburg e colleghi (2005) appartiene al guppo clostridiale dei batteri firmicuti, la maggior parte nel cluster XIVa (anche definito come il gruppodi Clostridium coccoides) e nel cluster IV (Clostridium leptum) e la maggior parte di questi sono microrganismi nuovi e non appartenenti a nessuna collezione microbica. Le specie autoctone numericamente preponderanti appartengono quindi ai generi Bacteroides, Bifidobacterium, Clostridium, Coprococcus, Peptostreptococcus, Eubacterium e Ruminococcus. Fusobacterium, Streptococcus, Lactobacillus, Veillonella, Megasphaera, Propionibacterium ed enterobacteriacee sono invece presenti in

5
concentrazione inferiore. Questo complesso ecosistema può per semplicità essere considerato come un’entità unica; le specie da cui è costituito, che psono avere effetti benefici o patogeni sulla salute dell’ospite (fig. 1), interagiscono sia in modo antagonista sia sinergico. Tali interazioni influenzano lo sviluppo o l’attività metabolica di ogni ceppo presente.
Fig. 1 Effetti dei batteri intestinali predominanti sulla salute dell’ospite.
Una delle più importanti funzioni della microflora intestinale è il cosiddetto “effetto barriera” tale per cui un microrganismo introdotto per via alimentare in un ospite normale viene eliminato in pochi giorni (Ducluzeau et al. 1970) precludendo al microrganismo la possibilità di proliferare; questa esclusione

6
competitiva impedisce a molti microrganismi alloctoni di stabilirsi nel canale digerente, di proliferare e di esercitare eventualmente un’azione patogena.
Questo effetto contribuisce inoltre a mantenere al di sotto di una certa soglia la popolazione di gruppi come Clostridium o Enterococcus che, pur essendo normalmente presenti nell’intestino, sono potenzialmente patogeni. Talvolta però, l’indebolimento dell’effetto barriera può portare a disturbi intestinali più gravi, determinati dalla crescita di agenti opportunistici come Salmonella typhimurium, Sigella flexneri, Vibrio cholerae e Clostridium difficile.
1.3. Principali pathway metabolici nel colon
La microflora intestinale e l’ospite presentano interazioni metaboliche
molto strette. Tali interazioni non vanno intese esclusivamente da un punto di vista nutrizionale, in quanto condizionano anche la salute dell’ospite. Infatti, enzimi di origine microbica possono catalizzare trasformazioni chimiche dei componenti alimentari o di substrati prodotti dall’ospite tali da alterare la biochimica, la fisiologia e le caratteristiche immunologiche dell’intestino.
Le fonti di carbonio e di azoto disponibili nel colon, che la microflora si è adattata ad utilizzare, sono costituite dai carboidrati e dalle proteine che sfuggono all’idrolisi da parte degli enzimi digestivi dell’ospite e raggiungono il colon senza che ne avvenga l’assorbimento durante il transito nell’intestino tenue. Nel colon la degradazione di queste sostanze è dovuta all’insieme delle attività metaboliche attuate dall’intera comunità batterica e ha l’effetto di instaurare interazioni di sinergia o interdipendenza nutrizionale, oltre che di competizione, tra diversi gruppi microbici.
A causa dell’assorbimento nel tenue, il colon è un ambiente povero di zuccheri semplici, quali mono- e disaccaridi. Sono invece presenti quegli

7
oligo- e polisaccaridi che resistono all’idrolisi da parte delle idrolasi dell’ospite: si tratta sia di carboidrati introdotti con l’alimentazione (ad esempio fibre, amidi resistenti, pectine), sia di glicoproteine e glicoconiugati prodotti dall’ospite. Questi carboidrati non-digeribili raggiungono il colon dove sono depolimerizzati e fermentati dalla flora batterica (Salminen et al. 1998).
L’abilità di utilizzare e degradare queste fibre insolubili può essere limitata dalla presenza di gruppi di batteri definiti “primary degraders”, come nel caso della degradazione della cellulosa (Robert and Bernalier-Donadille 2003) e della mucina (Derrien et al. 2004). Recenti studi in vitro hanno dimostrato che l’utilizzo di un determinato substrato è altamente specie-specifico e che i motivi di questa specializzazione potrebbero risiedere nella necessità di un appropriato sistema di legame, degradazione e uptake del substrato (McWilliam Leitch et al. 2007). I consorzi microbici, che includono batteri coinvolti nella degradazione primaria e batteri coinvolti in quella secondaria, svolgono un ruolo fondamentale nell’ecologia microbica dell’intestino. E’ stato dimostrato che le capacità metaboliche potrebbero differenziarsi tra batteri aderenti e batteri non aderenti ai residui alimentari evidenziando quindi il ruolo fondamentale nella capacità di sviluppare biofilm (MacFarlane and MacFarlane 2006)
La fermentazione dei carboidrati è fondamentale per quello che riguarda la microecologia e la fisiologia dell’intestino crasso in quanto genera acido lattico e SCFA (acidi grassi a catena corta: acetico, propionico, butirrico), che saranno assorbiti dall’ospite oppure utilizzati da altri componenti microbici (Cummings and MacFarlane 1991; Hoverstad et al. 1984; Hoverstad and Bjorneklett 1984; Ruppin et al. 1980). Quindi l’idrolisi di carboidrati complessi crea l’opportunità non solo per la competizione ma anche per la cooperazione mediante cross-feeding metabolico (fig. 2). Queste interazioni includono il trasferimento d’idrogeno e l’utilizzo di prodotti di fermentazione quali lattato, succinato e oligosaccaridi residui dall’idrolisi dei

8
polimeri complessi. Alcuni studi hanno dimostrato che nell’ecosistema intestinale è possibile ritrovare la conversione del lattato sia in butirrato sia in propionato (Bourriaud et al. 2005; Morrison et al. 2006).
Fig. 2 Ruolo delle interazioni di cross-feeding nella conversione di carboidrati non digeribili da parte del consorzio dei batteri anaerobi nell’intestino crasso.
Quindi la competizione tra differenti gruppi batterici in grado di
utilizzare il lattato, tra cui anche i batteri solfato riduttori (SRB), potrebbe essere un importante fattore nel determinare l’equilibrio dei prodotti di fermentazione.
Il metabolismo dei carboidrati è quantitativamente più importante nel tratto ascendente del colon, dove la disponibilità di substrato fermentabile è maggiore; la produzione di acidi organici causa l’abbassamento del pH di questo tratto di intestino ostacolando la crescita dei batteri proteolitici putrefattivi, potenzialmente patogeni. Inoltre gli SCFA esercitano un effetto stimolatorio su sull’uptake di acqua ed elettroliti da parte della mucosa del colon ne è stata dimostrata la capacità di inibire la proliferazione dei colonociti neoplastici (Livesey and Elia 1995, Kim et al. 1994).

9
La maggior parte dei gruppi batterici del colon (Bacteroides, Bifidobacterium, Eubacterium, Lactobacillus e Clostridium) è saccarolitica, tuttavia esistono notevoli differenze per quello che riguarda sia le capacità degradative sia le cinetiche di crescita. Infatti, le attività glicosil-idrolasiche responsabili dell’idrolisi dei polisaccaridi sono variamente distribuite tra i diversi gruppi. Mentre i bifidobatteri sviluppano con elevate velocità di crescita in presenza di elevate concentrazioni di carboidrati, i Bacteroides sono competitivi in condizioni di limitazione di fonte di carbonio e sviluppano a basse velocità di crescita (MacFarlane et al. 1995; Salminen et al. 1998).
Nel colon sono presenti proteine e peptidi di origine alimentare, batterica o endogena (Chacko and Cummings 1988; Cummings and MacFarlane 1997). Si tratta di residui alimentari, di enzimi prodotti dall’ospite o dal microbiota intestinale, di glicoproteine del muco e di proteine rilasciate con lo sfaldamento delle cellule epiteliali o con la lisi dei batteri intestinali. Proteine e peptidi sono depolimerizzati ad opera dell’insieme delle endopeptidasi batteriche e pancreatiche presenti nel colon, che difatti costituiscono uno degli ecosistemi conosciuti con la maggiore attività proteolitica. Le proteasi prodotte dai gruppi microbici proteolitici idrolizzano le proteine ad amminoacidi e peptidi semplici, che rappresentano un’importante fonte di azoto per l’intero microbiota, inclusi i saccarolitici obbligati. Aminoacidi e peptidi possono inoltre essere utilizzati come fonti di carbonio ed energia da batteri che dipendono in modo obbligato dal metabolismo degli aminoacidi o che sono in grado di fermentare sia amminoacidi sia carboidrati (MacFarlane et al. 1995, MacFarlane and MacFarlane 1995, Salminen 1998).
Clostridium, Bacteroides, Peptococcus ed Eubacterium sono i principali batteri proteolitici a livello intestinale. La secrezione di proteasi da parte di questi gruppi microbici avviene in risposta alla limitazione di nutrienti e ha luogo principalmente a livello del colon discendente. Lungo il transito lungo l’intestino crasso la concentrazione dei carboidrati fermentabili si riduce

10
gradualmente fino all’esaurimento facendo sì che il metabolismo fermentativo ceda il passo a quello proteolitico putrefattivo, come evidenziato dal progressivo aumento del pH. Gli amminoacidi sono catabolizzati attraverso reazioni di deaminazione, idrossilazione, deidrossilazione, desaturazione e decarbossilazione che rilasciano prodotti dannosi per la salute dell’ospite, quali ammoniaca, ammine e fenoli. L’ammoniaca diffonde attraverso la parete intestinale ed esercita un’azione tossica a livello cerebrale; le ammine, specialmente nei casi in cui un’insufficienza epatica ne compromette la detossificazione, sono nocive in quanto alterano la circolazione e l’attività muscolare e risultano mutagene e cancerogene; i fenoli, principalmente scatolo e indolo, sono fortemente cancerogeni.
2. PROBIOTICI
2.1. Effetti benefici sulla salute dell’ospite
Poiché i prodotti dei pathway metabolici fermentativi hanno effetti positivi sulla salute dell’ospite, al contrario di quelli dei pathway putrefattivi, sono state sviluppate strategie alimentari o farmaceutiche finalizzate a spostare l’equilibrio della microflora intestinale a favore dei gruppi microbici e delle attività metaboliche fermentative a scapito dei gruppi e delle attività proteolitiche.
I probiotici sono supplementi alimentari microbici vivi che influenzano positivamente la salute dell’ospite, migliorando l’equilibrio della microflora intestinale (Salminen et al. 1998, Tannock 1999, Tannock 2002). I batteri probiotici devono essere in grado di sopravvivere ai succhi gastrici e agli acidi biliari, al fine di raggiungere il colon ove eserciteranno la loro azione benefica.

11
La capacità di aderire al tessuto epiteliale dell’intestino è molto importante per permetterne, anche se transitoriamente, la colonizzazione.
I batteri probiotici sono saccarolitici obbligati che contribuiscono ad abbassare il pH intestinale inibendo lo sviluppo di molti microrganismi patogeni, inoltre competono con i patogeni per i siti di adesione alle cellule epiteliali e per i nutriliti. Altre attività benefiche dei probiotici consistono nella riduzione delle attività enzimatiche potenzialmente dannose dovute principalmente a coliformi, Bacteroides e clostridi (azoreduttasi, nitroreduttasi, β-glucuronidasi); nella produzione di vitamine ed aminoacidi; nell’azione antinfiammatoria, immunomodulante e stimolante la produzione di citochine; nella riduzione dell’insorgenza di forme tumorali dei colonociti (Salminen et al. 1998, Tannock 1999, Tannock 2002).
Solo recentemente è stata evidenziata per alcune specie di lattobacilli l’attività antiossidante e l’inibizione dell’accumulo di ROS (Kaizu et al. 1993). Recenti analisi hanno infatti dimostrato come alcuni ceppi di interesse probiotico appartenenti ai generi Lactobacillus e Streptococcus siano in grado di detossificare le specie reattive all’ossigeno (ROS) e di utilizzare i prodotti della detossificazione di questi radicali (H2O2) come antimicrobici di competitor patogeni. Anche i bifidobatteri sono privi di catalasi ed eliminano l’ossigeno per azione di NADH-ossidasi e NADH-perossidasi.
Alti livelli di NADHossidasi e NADH- perossidasi sono stati trovati in diversi ceppi di Bifidobacterium aerotolleranti, dove svolgono quindi un importante ruolo nel ridurre la tossicità dell’ossigeno. La correlazione tra la concentrazione di questi enzimi e i livelli di ossigeno intracellulare suggerisce come quest’ultimo ne induca l’attività. Un effetto simile avviene negli streptococchi (Higuchi, 1984), nei quali l’attività della NADH ossidasi e NADH perossidasi aumenta quando le cellule sono esposte all’ossigeno. In nessun ceppo appartenente ai generi Bifidobacterium, Lactobacillus, Lactococcus e Streptococcus è stata dimostrata la presenza di enzimi in grado

12
di eliminare i radicali idrossilici, funzione che è stata invece associata alla presenza di glutatione, o di altre proteine tioliche con azione di “scavengers”.
I probiotici destinati all’impiego umano si distinguono in probiotici alimentari e probiotici farmaceutici. I primi, oltre ad un effetto positivo sulla microflora intestinale dell’ospite, portano soprattutto ad un miglioramento degli aspetti nutritivi dell’alimento. Infatti è stato osservato che i prodotti caseari fermentati contengono vitamine e sali minerali (MacDonought et al. 1983) in maggiore quantità, proteine e carboidrati in forma più digeribile rispetto a quelli non fermentati. I probiotici farmaceutici, invece, sono impiegati nel trattamento di diarrea, stipsi, flatulenza, acidità gastrica, gastroenterite, ipercolesterolemia e malattie infiammatorie intestinali quali colite ulcerosa, morbo di Crohn e paucite.
2.2. Criteri per la scelta di un ceppo probiotico ideale
Affinchè un microrganismo possa esser impiegato come probiotico in preparati farmaceutici o alimentari sono desiderabili determinate caratteristiche fisiologiche legate non solo all’effetto benefico esercitato sull’ospite ma anche ai fini pratici della produzione su larga scala.
Infatti nel 1983 divenne chiaro che i ceppi di Lactobacillus utilizzati nell’industria casearia (come ad esempio L. bulgaricus, L. casei e L. acidophilus) non possedevano quelle caratteristiche biologiche tali da favorire la loro colonizzazione nell’intestino dell’ospite. Dopo oltre vent’anni di ricerche, è stato stabilito che la capacità di colonizzare l’intestino è la caratteristica fondamentale che un ceppo probiotico deve possere per poter influenzare l’ecosistema intestinale (Gorbach 2002). Venne quindi creato un elenco delle caratteristiche che un ideale ceppo probiotico avrebbe dovuto avere al fine di avere effetti benefici sulla salute dell’ospite e di poter essere usato nell’industria casearia (fig. 3).

13
Fig. 3 Principi teorici per la selezione di microrganismi probiotici
E’ pertanto auspicabile che un microrganismo potenzialmente probiotico abbia le seguenti caratteristiche: a) raggiunga l’intestino in forma vitale e ad alta concentrazione; a tale scopo
esso deve: - prestarsi alla produzione su larga scala, in quanto il preparato deve contenere un elevato numero di cellule vitali per grammo (il trattamento prevede la somministrazione di almeno 107-109 cellule al giorno) - essere stabile durante e dopo il processo di liofilizzazione o altri metodi di essiccamento
- essere acido-resistente, per superare indenne la barriera gastrica essere resistente all’azione degli acidi biliari.
b) permanga nell’intestino per lungo tempo o perlomeno durante tutto il periodo di somministrazione del probiotico. La capacità di adesione all’epitelio favorisce la colonizzazione permettendo di ridurre la dose giornaliera somministrata. Sebbene la colonizzazione

14
permanente abbia il vantaggio di prolungare gli effetti benefici anche dopo il trattamento, è comunque auspicabile che si mantenga solo per la durata della somministrazione in modo da non alterare gli equilibri del microbiota. Inoltre l’eventuale comparsa di effetti indesiderati è revertibile semplicemente sospendendo la cura.
c) non possieda alcuna antibiotico-resistenza. La resistenza agli antibiotici consente la sopravvivenza del ceppo probiotico anche dopo terapia antibiotica, ma espone al rischio di trasmissione orizzontale (es. per coniugazione) dell’antibiotico-resistenza ad eventuali patogeni intestinali.
d) soddisfi i criteri di specificità dell’ospite. Un preparato probiotico ad uso umano contiene in genere ceppi appartenenti ad alcune delle 8-10 specie umane più diffuse, così da favorire la colonizzazione e dunque l’efficacia del prodotto nel maggior numero possibile di pazienti.
e) non avere effetti nocivi: - non deve essere patogeno
- non deve produrre gas quali H2, CH4, H2S - non deve possedere enzimi ad azione tossica.
Per questo motivo i ceppi probiotici maggiormente utilizzati hanno origine intestinale e appartengono ai generi:
Bifidobacterium (B. bifidum, B. adolescentis, B. animalis, B. infantis, B. longum, B. thermophilum) Lactobacillus (L. acidophilus, L. casei, L. delbrueckii subsp. bulgaricus, L. reuteri, L. brevis, L. cellobiosus, L. curvatus, L. fermentum, L. plantarum) Cocchi gram positivi di origine non intestinale (Streptococcus thermophilus, S. diacetilactis, S. intermedius, Lactococcus lactis subsp. cremoris, Enterococcus faecium).

15
2.3. Batteri lattici
I batteri lattici rappresentano un’ampia ed eterogenea famiglia di microrganismi che hanno la caratteristica di convertire i carboidrati fermentabili principalmente in acido lattico (Teuber 1993).
Sono batteri gram positivi, non sporigeni, immobili e microaerofili (Schlegel 1976), cioè sviluppano ottimamente a basse concentrazioni di ossigeno. Sono privi di catalasi, nitrato reduttasi e citocromo ossidasi, infatti non hanno catena respiratoria e il loro metabolismo è di tipo fermentativo. Essendo privi dell’attività di catalasi, questi microrganismi consumano l’ossigeno per azione di flavoproteine (NADH-ossidasi) ed eliminano l’accumulo di H2O2 attraverso l’attività enzimatica della NADH-perossidasi.
Si possono definire quindi microrganismi eterotrofi e chemiorganotrofi, che si sono adattati a vivere su substrati complessi e che necessitano non solo di carboidrati come fonte di energia, ma anche di amminoacidi, nucleotidi e vitamine (Kandler et al. 1986). Riescono a tollerare acidità elevate, dato che alcune specie sviluppano anche a circa pH 3 (Teuber, 1993).
La famiglia dei batteri lattici include diversi generi, tra i quali Lactobacillus e Streptococcus, che occupano nicchie ecologiche abbastanza differenti e sono utilizzati come batteri probiotici. Per distinguere i diversi generi si fa ricorso a caratteristiche fenotipiche quali al crescita a 10 °C e 45 °C , la tolleranza ad alte concentrazioni di sale (6.5% e 18% di NaCl), tolleranza a pH acidi o alcalini, isomeri dell’acido lattico prodotti e tipo di fermentazione del glucosio.
2.3.1. Il genere Lactobacillus
La morfologia cellulare è molto variabile: certe specie appaiono come bastoncelli lunghi e sottili, talvolta ricurvi, mentre in altre le cellule sono corte

16
e tozze (coccobacilli). Tuttavia, all’interno di ciascuna specie, la lunghezza varia anche in funzione dello stadio di sviluppo, della composizione del terreno e della concentrazione di ossigeno (fig. 4).
Fig. 4 Immagine al microscopio elettronico di L. bulgaricus gentilmente concessa dall’Utah
State University
Il genere Lactobacillus è ampiamente diffuso nonostante, come per tutti i batteri lattici, le esigenze nutrizionali siano elevate (Hammes et al. 1991). Dal punto di vista metabolico, il genere Lactobacillus è diviso in tre gruppi (Kandler et al. 1986):
1- lattobacilli omofermentanti obbligati; 2- lattobacilli eterofermentanti facoltativi; 3- lattobacilli eterofermentanti obbligati.
I lattobacilli omofermentanti obbligati fermentano i carboidrati esosi producendo esclusivamente acido lattico, non sono in grado di fermentare i pentosi e non producono gas.
I lattobacilli eterofermentanti facoltativi fermentano gli esosi producendo acido lattico come gli omofermentanti, ma alcune specie, in determinate condizioni, producono anche acido acetico, acido formico e/o etanolo. Sono inoltre in grado di fermentare i pentosi ad acido lattico e acido acetico. Le differenze tra gli omo- e gli etero- fermentanti sta nella presenza o

17
assenza di enzimi chiave come la fruttosio-1,6-difosfatoaldolasi (FDP-aldolasi) per i primi e la fosfochetolasi per i secondi (Kandler et al. 1986). I lattobacilli eterofermentanti obbligati fermentano gli esosi producendo acido lattico, CO2 e acido acetico e/o alcol etilico, sono capaci di fermentare anche i pentosi con produzione di acido lattico e acetico (Kandler et al. 1986).
2.3.2. Streptococcus thermophilus
I microrganismi appartenenti al genere Streptococcus sono batteri gram-positivi, non sporigeni, immobili e anaerobi facoltativi. Le cellule hanno una forma ovoidale e sferoidale con un diametro di 0.5-1 µm, tipicamente organizzate in disposizioni diploidi o a formare catenelle più o meno lunghe come illustrato in fig. 5 (La Placa 1995).
Fig. 5 Immagine al microscopio elettronico di S. thermophilus
Le specie di Streptococcus attualmente conosciute sono circa 40,
distribuite principalmente sulle superfici delle mucose umane ed animali, includendo anche il tratto gastrointestinale; alcune sono state ritrovate anche nel terreno, nelle acque, su superfici di piante ed in svariati alimenti.
Gli streptococchi, oltre che in base alle attività biochimiche, sono raggruppati in rapporto al tipo di emolisi prodotta in piastre di agar-sangue ed in rapporto alle caratteristiche antigeniche di alcuni polisaccaridi della parete cellulare, denominati globalmente “antigene C” ed estraibili mediante idrolisi

18
acida a caldo, da una sospensione cellulare (antigene di Lancefield) (La Placa 1995).
S. thermophilus appartiene alla divisione “viridans” degli streptococchi che include S. salivarius, S. equinus e S. bovis che si ritrovano sulle mucose della bocca e dell’intestino umano ed animale (Schleifer et al. 1987); essi infatti costituiscono la gran parte della popolazione microbica orale e faringea. Gli streptococchi appartenenti a questo gruppo producono particolari polisaccaridi adesivi, i glucani, che favoriscono l’adesività alle superfici lisce come quella dello smalto dentale.
Streptococcus thermophilus è un microrganismo chemiorganotrofo, ossidasi negativo e catalasi negativo; possiede lunghe catene di acidi grassi saturi o monoinsaturi e il suo contenuto in citosina più guanina varia dal 33% al 46% . Viene denominato “thermophilus” data la sua capacità di sviluppare a temperature comprese fra i 37° e i 45°C. A differenza dei lattococchi, gli streptococchi sono in grado di fermentare pochi carboidrati (Hardie 1986; Hutkins 1984); presentano inoltre una marcata preferenza per i disaccaridi, in particolar modo per lattosio e saccarosio. I ceppi di S. thermophilus sono provvisti di una lattosio permeasi, denominata Lac S, che permette l’ingresso del disaccaride nella cellula. Lo zucchero presente a livello intracellulare viene poi scisso in glucosio e galattosio da una β-galattosidasi (Schroeder et al. 1991); quest’ultimo viene poi secreto dalla cellula alla stessa velocità con la quale viene introdotto il lattosio. Si è inoltre osservato che la sintesi di β-galattosidasi è inibita dalla presenza di glucosio.
S. thermophilus viene ampiamente utilizzato per la preparazione di prodotti caseari, in particolare nelle lavorazioni dello yogurt e di formaggi freschi.

19
2.4. I bifidobatteri 2.4.1. Tassonomia, fisiologia e nutrizione
Il genere Bifidobacterium rappresenta uno dei principali gruppi microbici dell’ecosistema intestinale. Sono batteri gram-positivi ad alto contenuto di GC, anaerobi stretti, non motili, asporigeni, che traggono energia fermentando carboidrati in acido acetico e lattico senza produrre gas.
Fig. 6 Immagini al microscopio elettronico a scansione di B. animalis, B. bifidum, B. infantis e B. longum.
Vennero per la prima volta descritti già intorno al 1900 da Tissier che
isolò dalle feci di un bambino allattato al seno un batterio anaerobico dalla morfologia bifida.
I bifidobatteri sono caratterizzati da un elevato pleomorfismo cellulare, la loro morfologia comprende bastoncelli dalle caratteristiche estremità a Y (bifida), bastoncelli curvi, forme maggiormente ramificate e forme irregolari

20
con rigonfiamenti e protuberanze (fig. 6). Le cause di tale pleomorfismo non sono ancora del tutto chiare, ma sono in gran parte correlate alle diverse condizioni di coltura: l’abbondanza di N-acetilglucosammina, alanina, acido aspartico, acido glutammico, serina e ioni Ca2+ nel terreno di crescita influenzano la forma cellulare dei bifidobatteri (Scardovi 1986).
La morfologia a bastoncello si osserva generalmente in condizioni nutrizionali e fisiologiche ottimali, generalmente quelle dell’habitat naturale. Le morfologie più ramificate e irregolari si originano quindi in risposta a una sintesi difettiva della parete cellulare oltre che a pH acidi, temperature elevate, stress nutrizionali, fonti di azoto o composizioni saline inadeguate, ma non rappresentano né forme di resistenza, né forme cellulari morenti o degenerate (Bezkorovainy and Miller-Catchpole 1989).
La classificazione tassonomica odierna, che sfrutta l’rRNA16S, identifica 35 specie diverse all’interno del genere Bifidobacterium (tab. 2). Insieme al genere Gardnerella, il genere Bifidobacterium, è classificato parte della famiglia delle Bifidobacteriaceae, ordine Bifidobacteriales, sottoclasse Actinobacteridae, classe Actinobacteria, linea dei Firmicuti ad alto GC, dominio dei Batteri (Biavati et al. 1986, Biavati and Mattarelli 2001). Il genoma di Bifidobacterium longum NCC2705, recentemente sequenziato (Schell et al. 2002), è costituto da 2.26 Mbp con 1.730 ORF e un contenuto in GC del 60%.
I bifidobatteri sono batteri saccarolitici obbligati che traggono carbonio ed energia dalla fermentazione dei carboidrati disponibili nell’intestino ad acido acetico ed acido lattico. Per quanto riguarda le fonti di azoto, il genere Bifidobacterium è piuttosto eterogeneo. Molte specie (B. bifidum, B. infantis, B. breve, B. adolescentis, B. thermophilum, B. longum, B. pseudolongum) sono in grado di crescere utilizzando sali di ammonio e cisteina come uniche fonti d’azoto mentre altre necessitano di composti azotati organici (Matteuzzi et al. 1978).

21
Specie Origine Specie Origine
1. B. adolescentis adulto 19. B. inopinatum carie dentaria 2. B. angulatum adulto 20. B. lactis latte 3. B. animalis pollo / ratto 21. B. longum infante / adulto 4. B. asteroides ape 22. B. magnum coniglio 5. B. bifidum bambino 23. B. merycicum rumine bovino 6. B. boum rumine bovino 24. B. minimum acque reflue 7. B. breve bambino 25. B. pseudocatenolatum bambino 8. B. catenolatum adulto 26. B. pseudolongum suino 9. B. choerinum lattonzolo 27. B. psychraerophilum suino 10. B. coryneforme ape 28. B. pullorum pollo 11. B. cuniculi coniglio 29. B. ruminantium rumine bovino 12. B. denticolens carie dentaria 30. B. saeculare coniglio 13. B. dentium carie dentaria 31. B. scardovii uomo 14. B. gallicum pollo 32. B. subtile acque reflue 15. B. gallinarum pollo 33. B. suis suino 16. B. globosum rumine bovino 34. B. thermoacidophilum suino 17. B. indicum ape 35. B. thermophilum rumine bovino 18. B. infantis bambino
Tab. 2 Specie del genere Bifidobacterium e loro origine.
La sequenza genomica di Bifidobacterium longum indica, infatti, che
questa specie possiede tutti i geni necessari per la sintesi di almeno 19 amminoacidi a partire da ammonio, utilizzando fosfoenolpiruvato, ossalacetato e fumarato provenienti dal catabolismo dei carboidrati (Schell et al. 2002). In alternativa all’ammonio libero, altre fonti di ammonio possono essere rappresentate da glutamina, asparagina e, nelle specie dotate di ureasi (B. infantis, B. breve, B. magnum e B. subtile), urea (Crociani e Matteuzzi 1982). Dall’analisi del genoma di B. longum si è riscontrata la presenza di tutti gli enzimi necessari alla sintesi di vitamine come acido folico, tiamina e acido nicotinico mentre sono assenti le vie di sintesi per riboflavina, biotina, cobalamina, pantotenato e piridossina.

22
2.4.2. Ecologia L’intestino dei mammiferi, nel quale sono presenti in concentrazioni
che raggiungono 1010 cfu ml-1, rappresenta sicuramente l’habitat più adatto per lo sviluppo dei bifidobatteri. Nell’uomo i bifidobatteri sono presenti nel colon umano a tutte le età. Alla nascita, l’apparato gastrointestinale del neonato è sterile, ma la colonizzazione microbica inizia già nel primo giorno di vita, quando il neonato viene a contatto con i microrganismi del tratto genito-urinario della madre e con quelli presenti nel latte materno o in quello artificiale. I primi colonizzatori sono batteri anaerobi facoltativi come streptococchi ed enterobatteri, incluso E. coli, i quali contribuiscono a generare l’anaerobiosi necessaria alla colonizzazione dei batteri anaerobi.
L’ordine con cui i gruppi microbici si succedono nella colonizzazione del colon varia enormemente da caso a caso e porta all’instaurarsi di una microflora relativamente stabile in poco più di una settimana, per i bambini nati di parto naturale e allattati al seno, o in tempi leggermente superiori. Nel colon dei bambini allattati con latte materno, i bifidobatteri sono i primi anaerobi a subentrare: a partire dai 3 giorni di vita rappresentano il gruppo microbico maggioritario e solo successivamente saranno seguiti nella colonizzazione da Bacteroides e Lactobacillus. La rapidità con cui il colon è colonizzato dai bifidobatteri è dovuta alla composizione del latte materno. Rispetto al latte bovino, infatti, il latte umano contiene una quantità molto superiore di oligosaccaridi che stimolano la crescita di Bifidobacterium.
Questi oligosaccaridi hanno un grado di polimerizzazione (DP) compreso tra 4 e 9 e sono stati classificati in 9 categorie, in funzione del contenuto glucidico, dei legami glicosidici e delle ramificazioni presenti. Tutti i 9 gruppi condividono la presenza di una molecola di lattosio all’estremità riducente, legata ad un’unità di N-acetilglucosamina mediante un legame β-(1→3). Su questo core si innestano catene saccaridiche più o meno ramificate e costituite prevalentemente da unità di galattosio, fucosio, N-

23
acetilglucosamina e acido N-acetilneuraminico (fig. 7) (Bezkorovainy and Miller-Catchpole 1989).
Fig. 7 Oligosaccaridi del latte materno con attività bifidogenica. Glc glucosio, Gal galattosio, Fuc fucosio, GlcNAc N-acetilglucosamina, NANA acido N-acetilneuraminico. Nei riquadri il core oligosaccaride.
Sebbene esistano notevoli differenze legate alle caratteristiche
dell’ospite, quali fisiologia, età, alimentazione, eventuali patologie e terapie, nel lattante la presenza di bifidobatteri è dell’ordine di 1011 cfu g-1, nell’adulto si stabilizza su valori di 109-1010 cfu g-1 e nell’anziano si riduce ulteriormente. I bifidobatteri costituiscono mediamente circa il 12 % dei batteri intestinali totali e non rappresentano il gruppo microbico maggioritario nell’uomo adulto, superati in numero da Bacteroides (25 %) ed Eubacterium (25 %) e seguiti da peptostreptococchi e fusobatteri (entrambi 9 %).
Anche per quello che riguarda le specie presenti esiste una notevole variabilità da individuo a individuo. Nell’intestino adulto le specie mediamente più rappresentate sono, nell’ordine, B. adolescentis, B. longum, B. infantis, B. breve e B. bifidum. Tuttavia, più che l’appartenenza alla specie, è
Gal GlcNAc
Gal GlcNAc Gal Glc
Gal GlcNAc Fuc
β ‐(1‐ 4) β ‐(1‐ 3) β ‐(1‐ 4)
α ‐(1‐ 3)
β ‐(1‐ 4) β ‐(1‐ 3)β ‐(1‐ 3)
β ‐(1‐ 6)
Gal GlcNAc Gal Glc
β ‐(1‐ 4) β ‐(1‐ 3)β ‐(1‐ 3)
NANA NANA
α ‐(2‐ 3) α ‐(2‐ 6)

24
significativo considerare i diversi ecotipi, organismi che, pur avendo una stessa base genetica, hanno sviluppato adattamenti diversi conformi a habitat e fattori diversi (Benno and Mitsuoka 1986).
Oltre all’intestino esistono altri ecosistemi animali colonizzati da bifidobatteri, anche se con concentrazioni drasticamente inferiori. La cavità orale, che ospita infatti B. adolescentis e B. dentium, quest’ultimo forse coinvolto nella formazione della carie, e nella vagina sono presenti B. breve, B. longum e B. adolescentis.
Per quello che riguarda i bifidobatteri di origine non umana, vanno citate le molte specie isolate dalle feci di conigli, polli, suini, bovini, ratti ed insetti, alcune delle quali sembrano essere caratterizzate da una certa specificità d’ospite. Altre specie infine, B. minimum e B. subtile, sono state isolate da acque reflue smentendo l’assunto che i bifidobatteri siano presenti esclusivamente nel tratto gastrointestinale degli animali.
2.4.3. Proprietà probiotiche In virtù dei benefici apportati nell’ospite, i bifidobatteri sono
considerati uno dei più importanti gruppi microbici probiotici. Il loro effetto probiotico, nutritivo e terapeutico, è dovuto a svariati meccanismi (Salminen et al. 1998, Tannock 1999, Tannock 2002).
Uno dei principali effetti benefici dei bifidobatteri consiste nell’abbassamento del pH del tratto ascendente del colon. Infatti, il metabolismo degli carboidrati in Bifidobacterium produce principalmente acidi acetico e acido lattico che determinano inibiscono lo sviluppo di molte specie microbiche acido sensibili potenzialmente patogene favorendone l’eliminazione. Salmonella, Shigella, Clostridium, Bacillus cereus, Staphilococcus aureus e Candida albicans sono tra i patogeni sensibili all’azione inibitoria dei bifidobatteri.

25
E’ stato dimostrato che la somministrazione di preparati probiotici contenenti bifidobatteri riduce l’incidenza di alcuni tipi di tumori, in particolare di quello intestinale. Questa azione antitumorale è correlata alla capacità di questi batteri probiotici di metabolizzare composti potenzialmente cancerogeni e di diminuire la quantità di enzimi microbici coinvolti nella produzione di queste sostanze, quali β-glucuronidasi, nitroreduttasi e azoreduttasi. Infatti, l’acidificazione del pH intestinale inibisce la crescita di molti microrganismi nitrato-riduttori che producono ioni nitrito per riduzione del nitrato, altamente cancerogeno. Analogamente il metabolismo di Bifidobacterium contribuisce a reprimere le specie responsabili della deconiugazione dei sali biliari primari e della loro trasformazione in derivati cancerogeni.
La presenza dei bifidobatteri nel colon ha effetti immunostimolanti, che si traducono in un’aumentata produzione di citochine. La risposta immunitaria nei confronti delle infezioni virali e dell’insorgenza di tumori ne risulta migliorata, così come si verifica un’azione antinfiammatoria particolarmente utile nella prevenzione e nel trattamento delle patologie infiammatorie del colon.
Un ulteriore beneficio apportato da alcune specie i bifidobatteri consiste nella secrezione di aminoacidi e vitamine che saranno disponibili per l’utilizzo da parte del microbiota intestinale o dell’ospite. In particolare la produzione di acido folico da parte di alcuni ceppi di Bifidobacterium risulta di particolare utilità nel prevenire l’insorgenza di forme neoplastiche dei colonociti (Crittenden et al 2003; Pompei et al. 2007a; Pompei et al. 2007b). Infine è stato dimostrato che la somministrazione di yogurt contenente 109 bifidobatteri g-1 causa una riduzione sostanziale del colesterolo plasmatico totale, sebbene il meccanismo alla base di tale azione non sia ancora stato chiarito.

26
2.4.4. Catabolismo dei carboidrati in Bifidobacterium
I bifidobatteri, in quanto saccarolitici fermentanti, utilizzano i carboidrati come fonte di carbonio e di energia e sono in grado di fermentare una grande varietà di zuccheri. I meccanismi di trasporto dei carboidrati e gli enzimi glicosil-idrolasici sono alla base della possibilità di internalizzare ed idrolizzare gli zuccheri e giocano un ruolo fondamentale nel definire le fonti di carbonio utilizzabili dai bifidobatteri. Molti monosaccaridi e disaccaridi, come glucosio, galattosio, fruttosio, saccarosio e lattosio sono utilizzati da tutte le specie di Bifidobacterium. Altri invece, come arabinosio, xilosio, mannosio, sorbitolo, maltosio, cellobiosio, sono utilizzati solo da alcune specie o da alcuni ceppi di bifidobatteri (Degnan e MacFarlane 1991, Hopkins et al. 1998, Mlobeli et al. 1998).
Dal punto di vista ecologico gli zuccheri semplici non rappresentano le fonti di carbonio normalmente utilizzate dai bifidobatteri nel loro ecosistema naturale. Il colon, infatti, è normalmente privo di zuccheri semplici a causa dell’assorbimento nel piccolo intestino, di conseguenza i bifidobatteri, così come gli altri saccarolitici intestinali, si sono evoluti in funzione dell’utilizzo dei carboidrati complessi che sfuggono alla digestione e all’assorbimento da parte dell’ospite. Infatti i bifidobatteri sono in grado di utilizzare come fonti di carbonio molte fibre di origine alimentare e molte glicoproteine, mucine e glicogoniugati di origine umana (Derensy-Dron et al. 1999, Murphy 2001; Hopkins et al. 1998; Crociani et al. 1994; Scardovi 1986). Le principali fonti di carbonio per i bifidobatteri sono costituite dagli oligosaccaridi del fruttosio (FOS), galattosio (TOS e oligosaccaridi della soia), glucosio (IMOS), xilosio (XOS), da fruttani (FOS, inulina) e da polisaccaridi come inulina, amido, emicellulose e pectine. Per il fatto di essere metabolizzati dai bifidobatteri in modo più o meno specifico, molti di questi carboidrati sono utilizzati come prebiotici in preparazioni alimentari e farmaceutiche.

27
La fermentazione di oligo- e polisaccaridi richiede che questi siano preventivamente idrolizzati. L’idrolisi può avvenire a livello intracellulare; in questo caso sono richiesti trasportatori per l’internalizzazione degli oligosaccaridi. In alternativa l’idrolisi può essere extracellulare e sarà seguita dall’uptake delle unità saccaridiche rilasciate. In questo caso le glicosil-idrolasi extracellulari possono essere prodotte tanto dalla cellula stessa quanto da altri membri del microbiota intestinale e rappresentano un importante elemento di interdipendenza nutrizionale tra diversi gruppi microbici.
2.4.5. Glicosil-idrolasi La capacità dei bifidobatteri di crescere utilizzando oligo- e
polisaccaridi come fonti di carbonio è legata alla produzione di glicosil-
idrolasi cellulari o extracellulari. Molte di queste sono state clonate e
caratterizzate a livello biochimico e talune sono utilizzate per la produzione di
oligosaccaridi semisintetici per transglicosilazione.
Le α-galattosidasi (α-Gal) sono enzimi intracellulari in forma
tetramerica che idrolizzano specificamente i legami α-(1→6) galattosidici, ma
è attiva anche su quelli α-(1→3) (van Laere et al. 1999, Leder et al. 1999).
Contribuiscono a idrolizzare importanti prebiotici quali raffinosio e altri
oligosaccaridi della soia come stachiosio e melibiosio.
Le β-galattosidasi (β-Gal) idrolizzano i legami β-(1→4) galattosidici
presenti nel lattosio e nei GOS e possono essere utilizzate per la sintesi galatto-
oligosaccaridi per trans-galattosilazione (Hinz et al. 2004, Moller et al. 2001,
Rossi et al. 2000). Sono state isolate e caratterizzate 3 β-Gal in B. bifidum,
attive rispettivamente in forma esamerica, dimerica e monomerica. Le 3
isoforme differiscono leggermente in quanto a specificità di substrato nei
confronti di lattosio e galatto-oligosaccaridi, inoltre, mentre le prime due
hanno localizzazione intracellulare, la terza si localizza nel periplasma quando

28
espressa in E. coli.
Le β-fruttofuranosidasi (β-Fru), o inulinasi, idrolizzano unità di fruttosio
terminali, in modalità eso-, contribuendo a idrolizzare FOS, inulina e
raffinosio. Nei confronti di saccarosio agiscono come invertasi, mentre non
attaccano legami α-glicosidici, o residui che non siano fruttosio (Ehrmann et
al. 2003, Janer et al. 2004, Warchol et al. 2002). La produzione di tale enzima
è stimolata dalla presenza di fruttosio (Perrin 2001).
Gli enzimi xilolitici includono le eso-β-xilosidasi, che rilasciano unità di xilosio dalle estremità non riducenti degli xilo-oligosaccaridi, e le endo-xilanasi; che idrolizzano in modo random lo xilano a xilo-oligosaccaridi. Sono note β-xilosidasi e xilanasi purificate da numerosi e differenti microrganismi termofili (Tuncer and Ball 2003; Lama et al. 2004; Kambourova et al. 2007; Bachmann et al. 1991; Honda and Kitaoka 2004). Solo due β-xilosidasi (EC 3.2.1.37) sono state caratterizzate in Bifidobacterium adolescentis e Bifidobacterium breve K-110 (Lagaert et al. 2007; Shin et al. 2003), anche se molti enzimi responsabili degli XOS sono stati predetti dall’analisi del genoma di B. longum (Schell et al. 2002; van den Broek et al. 2005).
Le altre attività glicosil-idrolasiche identificate in Bifidobacterium sono:
α-arabinofuranosidasi (Margolles and de los Reyes-Galivàn 2003; van der
Broek et al. 2005), α-fucosidasi (Katayama et al. 2004), α-amilasi (Wang et al.
1999; Lee et al. 1997), α-glucosidasi (Wang et al. 1999; van der Broek et al.
2003), destranasi (Kaster and Brown 1983) e pectinasi (Slovakova et al. 2002).
La sequenza del genoma di B. longum (Schell et al. 2002) suggerisce
che le capacità di fermentare carboidrati non digeribili siano ancora maggiori,
visto che più dell’8.5 % delle ORF predette è classificabile tra le proteine
coinvolte nel metabolismo e nel trasporto dei carboidrati. In particolare sono
state predette 40 proteine responsabili di 25 attività glicosil-idrolasiche diverse
(tab. 3). Tra queste è possibile riconoscere tutte le attività necessarie all’idrolisi
di molti dei carboidrati utilizzati come prebiotici e degli oligosaccaridi di

29
origine materna coinvolti nella colonizzazione dell’intestino nei primi giorni di
vita. Alcuni degli enzimi predetti sono dotati di un leader-peptide putativo, per
cui ne è ipotizzabile la localizzazione extracellulare.
proteine predette
attività enzimatica
proteine predette
attività enzimatica
2 4-α-gluconotrasferasi 1 β-fruttofuranosidasi 1 α-1,4-glucosidasi 3 β-galattosidasi 1 α-1,6-glucosidasi 2 β-glucosidasi 1 α -galattosidasi 2 endo-β-arabinosidasi 1 α-galattosidasi (melibiasi) 2 endo-β-1,4-xilanasi* 2 α-glucosidasi 1 endo-β-N-ac-glucosaminidasi* 3 α-L-arabinofuranosidasi 1 eso-β-1,5-arabinosidasi II 2 α-L-arabinosidasi 1 eso- β-1,3-eso-glucanasi 3 α-mannosidasi 2 isoamilasi 1 amilasi 1 neopullulanasi 1
arabinogalattan- 1 saccarosio fosforilasi endo-β-galattosidasi* 1 sialidasi*
1 arabinosidasi* 1 xilan esterasi 1 α -esoaminidasi 1 xilanasi*
Tab. 3 Glicosil-idrolasi predette dal genoma di B. longum. * sequenze con leader-peptide putativo.
2.4.6. Shunt dell’esoso fosfato
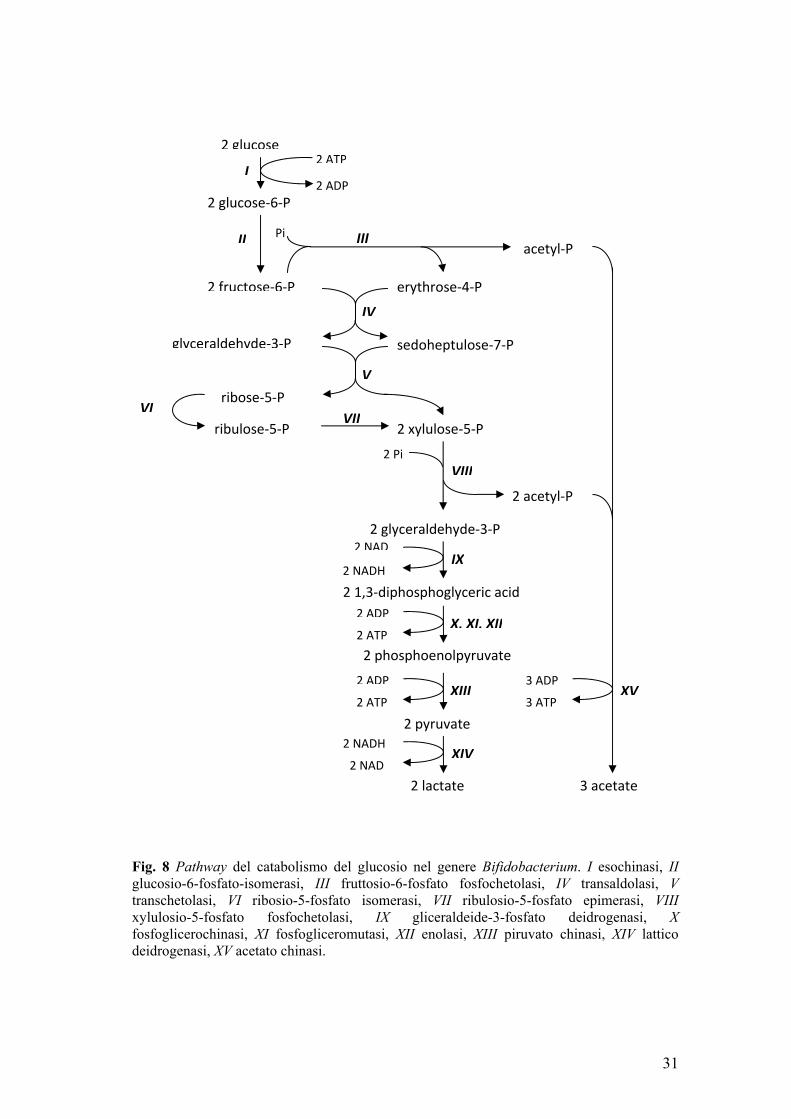
In Bifidobacterium il catabolismo dei monosaccaridi segue una via
caratteristica di questo genere (fig. 8). Tale catabolismo è caratterizzato dalla presenza dell’enzima fruttosio-6-fosfato-fosfochetolasi che scinde il fruttosio-6-fosfato in eritrosio-4-fosfato e acetil-fosfato. La reazione della fruttosio-6-fosfato-fosfochetolasi, presente nei soli bifidobatteri, è utilizzata come marker per l’attribuzione tassonomica al genere Bifidobacterium (Scardovi 1986).
Una volta internalizzato, il glucosio è fosforilato dall’esochinasi

30
generando il glucosio-6-fosfato che è poi isomerizzato a fruttosio-6-fosfato. Il fruttosio-6-fosfato, anziché essere scisso in di-idrossiacetone-fosfato e acetaldeide-3-fosfato come avviene nella normale via glicolitica, è scisso in acetil-fosfato e in eritroso-4-fosfato ad opera della fruttosio-6-fosfato-fosfochetolasi. Eritroso-4-fosfato e fruttosio-6-fosfato sono convertiti in xilulosio-5-fosfato dall’azione successiva della transaldolasi e della transchetolasi. Il pentoso-fosfato è poi scisso in gliceraldeide-3-fosfato ed acetil-fosfato. La gliceraldeide-3-fosfato segue il pathway glicosidico per ottenere piruvato con la formazione di 2ATP e NADH. Il piruvato è infine ridotto a lattato dalla lattico deidrogenasi NADH-dipendente per rigenerare NAD. Dall’acetil-fosfato, invece si ottengono acido acetico e ATP grazie alla transfosforilazione operata dall’acetato chinasi.
Il bilancio complessivo dello shunt prevede la produzione di 1.5 mol di acido acetico e di 1.0 mol di acido lattico secondo la reazione complessiva:
2 Glucosio + 5 ADP + 5Pi → 3 acetato + 2 lattato + 5 ATP In funzione delle richieste di ATP e NADH legate ai processi
metabolici, il flusso di carbonio può essere indirizzato diversamente dal rapporto previsto 1.5:1.0 tra acetato e lattato. Al variare della specie, dello stato fisiologico, del terreno di coltura e della fonte di carbonio la fermentazione dei carboidrati può generare acetato e lattato in rapporto diverso dal previsto così come alcuni sottoprodotti come formiato ed etanolo (De Vries and Stouthamer 1967, Palframan 2003).
31
Fig. 8 Pathway del catabolismo del glucosio nel genere Bifidobacterium. I esochinasi, II glucosio-6-fosfato-isomerasi, III fruttosio-6-fosfato fosfochetolasi, IV transaldolasi, V transchetolasi, VI ribosio-5-fosfato isomerasi, VII ribulosio-5-fosfato epimerasi, VIII xylulosio-5-fosfato fosfochetolasi, IX gliceraldeide-3-fosfato deidrogenasi, X fosfoglicerochinasi, XI fosfogliceromutasi, XII enolasi, XIII piruvato chinasi, XIV lattico deidrogenasi, XV acetato chinasi.
2 glucose
2 glucose‐6‐P
erythrose‐4‐P
acetyl‐P
sedoheptulose‐7‐Pglyceraldehyde‐3‐P
ribose‐5‐P
2 xylulose‐5‐P
2 fructose‐6‐P
ribulose‐5‐P
2 acetyl‐P
2 glyceraldehyde‐3‐P
2 1,3‐diphosphoglyceric acid
2 phosphoenolpyruvate
2 pyruvate
2 ATP
2 ADP
2 Pi
2 NADH
2 ADP
2 ATP
2 lactate
2 NADH
2 NAD
3 acetate
3 ADP
3 ATP
Pi
I
II III
IV
V
VIIVI
VIII
IX
XIII
XIV
2 ADP
2 ATP
XV
X, XI, XII
2 NAD

32
Il piruvato, anziché essere deidrogenato a lattato, può andare incontro ad una reazione fosforoclastica che produce acetil-fosfato e formiato. Dall’acetil-fosfato si possono ottenere acetato o etanolo, producendo 1 mol di ATP nel primo caso o rigenerando 2 mol di NADH nel secondo (fig. 9).
Fig. 9 Reazione fosforoclastica e pathway degli acidi organici in Bifidobacterium. XIV lattico deidrogenasi, XV acetato kinasi, XVI enzima fosforoclastico, XVII aldeide deidrogenasi, XVIII alcool deidrogenasi.
In Bifidobacterium le vie cataboliche di tutti i monosaccaridi sono
convogliate, in modi diversi e a diversi livelli, sullo shunt del fruttosio-6-fosfato (Bezkorovainy and Miller-Catchpole 1989).
pyruvate lactate
NADH NAD
XIV
acetyl‐P
formate
acetate XV
Pi
acetaldehyde
ADP AT
NADH
NAD
ethanol
NADH
NAD
Pi
XVI
XVII
XVIII

33
Il fruttosio richiede la fosforilazione da parte della fruttochinasi ed entra nello shunt come fruttosio-6-fosfato.
Il galattosio, dopo esser fosforilato da una galattochinasi ATP dipendente, può seguire il pathway di Leloir, oppure la via della pirofosforilasi (fig. 10). Entrambe le vie, energeticamente equivalenti, producono glucosio-1-fosfato che, isomerizzato a glucosio-6-fosfato, sarà introdotto nello shunt.
Il saccarosio può essere idrolizzato da due differenti enzimi con la formazione di due prodotti diversi. La β-fruttofuranosidasi idrolizza il saccarosio in fruttosio e glucosio, mentre la saccarosio fosforilasi utilizza un fosfato per generare fruttosio e glucosio-1-fosfato. Questa seconda via è energeticamente molto più favorevole rispetto alla prima in quanto genera un prodotto fosforilato con conseguente risparmio di energia (Sprogoe et al. 2004).
Fig. 10 Pathway di Leloir (a) e via della pirofosforilasi (b). I epimerasi, II esoso-1-fosfato uridil trasferasi, III UDP-galattosio pirofosforilasi.
L’N-acetilglucosamina è deacetilata e fosforilata a glucosamina-6-
fosfato. Quest’ultima è deaminata per mezzo della glucosamina-6-fosfato deaminasi a fruttosio-6-fosfato che entra nello shunt (Bezkorovainy and Miller-Catchpole 1989).
galactose‐1‐P + UDP‐glucose glucose‐1‐P + UDP‐galctose
I
galactose‐1‐P + UTP
II
PPi + UDP‐galactose UDP‐glucose
UTP + glucose‐1‐P
IIII
III
a
b

34
3. CARBOIDRATI PREBIOTICI
I prebiotici sono ingredienti non digeribili o assorbibili da parte dell’ospite che stimolano selettivamente lo sviluppo o l’attività di un numero limitato di batteri benefici a livello intestinale (Gibson and Roberfroid 1995, Salminen et al. 1998, Tannock 2002). La maggior parte dei prebiotici sono carboidrati, ma la definizione non esclude altre molecole che potrebbero essere utilizzate con lo stesso scopo. I prebiotici consentono l’aumento non solo del numero di questi batteri benefici ma anche delle loro attività metaboliche attraverso l’apporto di substrato fermentabile: è proprio questo aumentato metabolismo ad essere la chiave centrale dell’effetto dei prebiotici sulla salute dell’ospite. Infatti il principale obiettivo dell’assunzione di prebiotici è l’aumento del numero e dell’attività dei bifidobatteri e dei batteri lattici già presenti nel colon, con una riduzione dei microrganismi putrefattivi o potenzialmente patogeni quali clostridi e enterobacteriacee (Campbell et al. 1997, Rycroft et al. 2001).
E’ importante però considerare che negli ultimi decenni sono stati definiti alcuni innovativi requisiti che un ingrediente alimentare deve possedere per poter essere considerato prebiotico (Roberfroid 2007; Gibson et al. 2004):
1. resistenza ai succhi gastrici, all’idrolisi da parte di enzimi umani e all’assorbimento intestinale.
Non è necessario che il prebiotico sia completamente non digeribile ma deve garantire che una significativa quantità del prodotto sia biodisponibile nell’intestino crasso come substrato fermentabile.
2. fermentazione da parte della microflora intestinale. Deve essere dimostrata sia in vitro che in vivo la capacità della microflora intestinale di utilizzare e quindi fermentare questo substrato.

35
3. stimolazione selettiva dello sviluppo e/o dell’attività di quei batteri intestinali che contribuiscono positivamente alla salute dell’ospite.
Deve essere dimostrata questa capacità selettiva di indurre l’aumento dei batteri considerati “benefici” per la salute dell’ospite, andando a valutare l’intero equilibrio della microflora intestinale in seguito all’assunzione del prebiotico. Molte tecniche molecolari hanno permesso una più semplice analisi delle variazioni in composizione della microflora intestinale nella sua complessità.
Tutti i prebiotici presenti in commercio sono costituiti da oligo- e
polisaccaridi. I carboidrati prebiotici non sono idrolizzati dagli enzimi idrolitici dell’ospite e fungono da fonti di carbonio fermentabili per la microflora saccarolitica del colon. Il grado di polimerizzazione e i tipi di legami glicosidici tra le unità monosaccaridiche, normalmente galattosio, glucosio e fruttosio, sono responsabili della specificità nell’utilizzo del prebiotico da parte della microflora intestinale. Infatti è desiderabile che il numero e il metabolismo dei batteri benefici presenti nel colon, in particolare bifidobatteri e lattobacilli, siano stimolati quanto più selettivamente possibile (Cummings e MacFarlane 1991, Gibson et al. 2004, Roberfroid 1998; Roberfroid 2007, Tannock 2002).
Molti prebiotici sono ottenuti a partire da materie prime vegetali, mentre altri sono prodotti per via enzimatica mediante l’idrolisi di polisaccaridi complessi o la trans-glicosilazione di mono- o disaccaridi.
Quindi secondo i criteri precedentemente elencati, a tutt’oggi solo 3 ingredienti alimentari possono essere effettivamente considerati prebiotici a pieno titolo (tab. 4), inulina, FOS e galatto-oligosaccaridi. Per la maggior parte degli altri substrati sono disponibili dati scientifici promettenti ma non ancora sufficienti per poterli considerare effettivamente prebiotici.

36
Tab. 4 Sommario e conclusione sull’effetto prebiotico di numerosi carboidrati.
3.1. Frutto-oligosaccaridi e inulina I frutto-oligosaccaridi (FOS) e l’inulina sono catene di unità fruttosio
legate ad un glucosio terminale (formula generale GFn). Le unità di fruttosio sono legate da legami β-(1→2); il glucosio è legato da un legame α-(1→1) analogo a quello presente nel saccarosio. Dal punto di vista chimico FOS e inulina differiscono esclusivamente per il grado di polimerizzazione (DP). Mentre i FOS sono caratterizzati da DP compresi tra 3 e 10, l’inulina è un polimero il cui DP può arrivare a 60 (fig. 11). FOS e inulina sono presenti normalmente nei tessuti di molti vegetali, soprattutto cicoria, carciofo, cipolla, aglio, topinambur. Inoltre i FOS possono essere prodotti a partire dal saccarosio (GF) mediante fruttosiltrasferasi che produce oligomeri di lunghezza crescente, come chestosio (GF2), nistosio (GF3) e fruttosilnistosio (GF4), o mediante l’idrolisi controllata dell’inulina estratta dalla cicoria. Inevitabilmente la miscela di fruttani generata dall’idrolisi dell’inulina conterrà anche oligomeri privi del glucosio terminale.

37
OO
H
OH
OH
HH
OH
OH
OO
CH2H
OH
OH
HH
OH
O
HH
H
OHOH
H OH
H
OH
n
Fig. 11 Struttura generale di FOS (n = 0 - 7) e inulina (n = 0 – 57).
FOS e inulina sono presenti normalmente nei tessuti di molti vegetali,
soprattutto cicoria, carciofo, cipolla, aglio, topinambur. Inoltre i FOS possono essere prodotti a partire dal saccarosio (GF) mediante fruttosiltrasferasi che produce oligomeri di lunghezza crescente, come chestosio (GF2), nistosio (GF3) e fruttosilnistosio (GF4), o mediante l’idrolisi controllata dell’inulina estratta dalla cicoria. Inevitabilmente la miscela di fruttani generata dall’idrolisi dell’inulina conterrà anche oligomeri privi del glucosio terminale.
La fermentazione di FOS e inulina a livello intestinale ne richiede la depolimerizzazione, catalizzata dall’enzima β-fruttofuranosidasi (β-Fru) che idrolizza le unità terminali di fruttosio ed agisce come invertasi nei confronti del saccarosio. L’effetto prebiotico dei FOS e dell’inulina è stato ampiamente dimostrato e numerosi studi riportano che la somministrazione di questi carboidrati causa l’aumento del numero dei bifidobatteri in vivo (Kolida and Gibson 2007; Bornet et al. 2002; Bouhnik et al. 2006, Bouhnik et al. 1999, Gibson et al. 1995,) e in vitro (Bournet et al. 2007; Gibson and Wang 1994,

38
Kaplan and Hutkins 2000, McKellar and Modler 1989, Palframan et al. 2003, Perrin et al. 2001, Wang and Gibson 1993). FOS e inulina costituiscono la categoria di carboidrati non digeribili maggiormente utilizzati come prebiotici.
3.2. Oligosaccaridi della soia Raffinosio e stachiosio sono gli oligosaccaridi principali di questa
categoria di prebiotici (fig. 12). Sono presenti in alte concentrazioni in molti legumi, in particolare nella soia e nei suoi derivati.
Raffinosio e stachiosio contengono entrambi un’unità di saccarosio legata, mediante legame α-(1→6), rispettivamente ad una o due unità di galattosio.
O
O
HH
OH
HOH
H OH
H
OH
CH2
O
O
HH
OH
HOH
H OH
H
O
HH
H
OHOH
H OH
H
CH2
OO
H
OH
OH
HH
OH
OH
sacc
aros
io raff
inos
io mel
ibio
sio
stac
hios
io
Fig. 12 Struttura degli oligosaccaridi presenti nella soia.
L’idrolisi degli oligosaccaridi della soia richiede quindi l’azione di una
α-galattosidasi (α-Gal), per l’idrolisi dei legami α glicosidici, e di una β-Fru

39
per l’inversione del saccarosio. Gli oligosaccaridi della soia hanno una documentata azione prebiotica e bifidogenica (Dinoto et al. 2006, Matteuzzi et al. 2004) e sono utilizzati in molti alimenti funzionali o supplementi alimentari.
3.3. Galatto-oligosaccaridi I galatto-oligosaccaridi sono oligomeri di unità di galattosio legati ad un
glucosio terminale; il più leggero di questa famiglia di oligosaccaridi è il lattosio, disaccaride di glucosio e galattosio (Fig. 13). I legami glicosidici che legano le unità monomeriche sono di tipo β-(1→4).
I galatto-oligosaccaridi sono normalmente presenti nel latte materno, ma possono essere prodotti a partire dal lattosio per l’azione dell’enzima β-galattosidasi (β-Gal). La β-Gal idrolizza i β-(1→4) galattosidi, ma si comporta da galattosil-trasferasi in presenza di elevate concentrazioni di lattosio. La reazione enzimatica porta ad una miscela di oligosaccaridi, definiti anche transgalatto-oligosaccaridi (TOS), che presentano un grado di polimerizzazione variabile tra 3 e 6 con un valore medio di 3-4 unità zuccherine.
O
HH
HOH
HOH
H OH
OH
O
HHH
H
O
OH
H OH
OH
OH
O
HH
HOH
OH
H OH
OH
O
HH
HOH
OH
H OH
OH
nlattosio
Fig. 13 Struttura di galatto-oligosaccaridi (n = 0 - 4) e lattosio.

40
I TOS non sono idrolizzati o assorbiti nell’intestino tenue umano ed è stato osservato che sono rapidamente fermentati nel colon prossimale. Sono utilizzati in particolare dai bifidobatteri ed è stato evidenziato che il loro utilizzo porta alla proliferazione in vivo di questo specifico genere (Vernazza et al. 2006; Bouhnik et al. 1997, Alliet et al. 2007; Gopal et al. 2001, Ito et al. 1990, Rabiu et al. 2001).
3.4. Isomalto-oligosaccaridi
Gli isomalto-oligosaccaridi (IMOS) sono miscele di oligomeri del glucosio di diverso grado di polimerizzazione e variamente ramificati, i cui membri più semplici sono isomaltosio e panosio (fig. 14). Sono caratterizzati da legami α-(1→4) e α-(1→6) glicosidici e sono prodotti attraverso l’idrolisi enzimatica dell’amido o la sintesi mediante destransucrasi a partire dal saccarosio.
OOH
HH
OHOH
H OH
H
CH2
O
O
HH
H
OHOH
H OH
H
OH
O
OHH
HH
OH
H OH
H
OH
maltosio
panosio
isomaltosio
Fig. 14 Struttura fondamentale degli isomalto-oligosaccaridi.

41
Poiché stimolano la proliferazione di bifidobatteri e lattobacilli nel colon stanno trovando crescente applicazione come prebiotici (Vernazza et al. 2006; Chung and Day 2002, Flickinger et al. 2000).
3.5. Xilo-oligosaccaridi e xilano
Gli xilo-oligosaccaridi (XOS) sono formati da 2-6 unità monomeriche
di xilosio legate da legami β-(1→4) (fig. 15). Gli XOS vengono idrolizzati a singole unità di xilosio dalle β-xilosidasi.
O
HH
H
OHOH
H OH
O
O
HH
H
O
OH
H OH
OH
O
HH
HOH
H OHn
Fig. 15 Struttura fondamentale degli xilo-oligosaccaridi (n = 0 - 4).
Gli XOS sono prodotti per idrolisi dello xilano, un polisaccaride di
origine vegetale e stanno trovando crescente applicazione come prebiotici (Vernazza et al. 2006; Rycroft et al. 2001).
Lo xilano appartiene al gruppo delle emicellulose, è un polisaccaride costituito da unità di xilosio legate da legami β-(1→4) con un grado di polimerizzazione compreso tra 70 e 200 (fig. 16). E’ quindi un polisaccaride molto complesso che presenta spesso ramificazioni con catene acetiliche e arabinofuranosidiche. Gli enzimi responsabili dell’idrolisi dello xilano sono le

42
endo-xilanasi, che idrolizzano appunto lo xilano a catene più corte di XOS. Ci sono pochissime evidenze scientifiche sull’utilizzo dello xilano come prebiotico, probabilmente per l’incapacità di esprimere enzimi in grado di idrolizzare tali catene (Salyers et al. 1977).
Fig. 16 Struttura fondamentale dello xilano
3.6. Lattosaccarosio, lattulosio, lattitolo Lattosaccarosio (fig. 17), lattulosio (fig. 18a) e lattitolo (fig. 18b) sono
oligosaccaridi semisintetici prodotti per via enzimatica. Sono resistenti alla digestione nel tratto gastrointestinale superiore, ma sono facilmente idrolizzati da un numero limitato di batteri intestinali. Gli effetti sono un aumento nel numero di Bifidobacterium, Lactobacillus e Streptococcus e una diminuzione della popolazione di Bacteroides, Clostridium, coliformi e Eubacterium (Finney et al. 2007; Okhusa et al. 1995).

43
O
HH
HOH
H OH
H
OH
O
O
O
H
OH
OH
HH
OH
O
HH
HOH
H OH
H
OH
OH
OH
saccarosio
latto
sacc
aros
iolatto
sio
Fig. 17 Lattosaccarosio.
O
O
HH
H
HOH
H OH
OH
OH
O
H
OHHH
OH
OHOH
O
O
HH
H
HOH
H OH
OH
OH
OHOH
HH
H
OH
H OH
OH
a b
Fig. 18 Lattulosio (a) e lattitolo (b).
3.7. Amido resistente Non tutto l’amido assunto con la dieta è idrolizzato e assorbito
nell’intestino tenue. Alcuni amidi sono relativamente resistenti all’idrolisi enzimatica e passano al colon senza essere digeriti costituendo la maggior fonte di carboidrati disponibili per la fermentazione dei batteri (Cummings and MacFarlane 1997). L’entità dell’idrolisi nell’intestino tenue e la sua relativa disponibilità per la fermentazione batterica nel colon dipendono dalla sua

44
origine e struttura. In passato gli amidi resistenti non sono stati considerati come prebiotici in quanto molte specie batteriche, oltre ai bifidobatteri, sono in grado di fermentarli. Tuttavia nonostante la specificità non sia elevata, alcuni studi condotti hanno dimostrato che la somministrazione continua di amidi resistenti, in particolare quelli ricchi di amilosio, favorisce l’aumento della proliferazione di bifidobatteri e lattobacilli (Brown 1997; Kleessen et al. 1997).
3.8. Fibra alimentare Sono definite fibra un’ampia categoria di sostanze che rappresentano la
maggiore componente strutturale dei vegetali. Si tratta di composti ad alto peso molecolare di natura sia polisaccaridica, ramificata o meno (cellulose, emicellulose, pectine, gomme), sia non saccaridica (cutina, lignina) (tab. 5).
Le fibre non sono metabolizzate dagli enzimi digestivi dell’ospite, ma possono fungere da fonti di carbonio fermentabile per la microflora intestinale, qualora il microbiota produca le attività enzimatiche necessarie alla loro idrolisi. Molte pectine ed emicellulose, specialmente se parzialmente idrolizzate, possono essere fermentate dai bifidobatteri in vitro e ne stimolano l’aumento selettivo in vivo, esercitando un’azione prebiotica (Chen et al. 2005, Giannini et al. 2006, Manderson et al. 2005, Olano-Martin et al. 2002 ).
Indipendentemente dai loro effetti sul metabolismo microflora intestinale, le fibre alimentari svolgono un ruolo fondamentale nella regolazione delle funzioni gastrointestinali. Pectine metilate, mucillaggini e gomme sono fibre solubili e tendono a gelificare rallentando lo svuotamento gastrico ed il transito intestinale ed interferendo nell’assorbimento di carboidrati e lipidi. Lignina, pectine non metilate, cellulosa ed emicellulose sono insolubili, aumentano la massa fecale, accelerano il transito intestinale, determinando una minore tensione intestinale e regolarizzando l’evacuazione.

45
fibre alimentari unità monomeriche non saccaridiche
cutina acidi grassi a lunga catena, con più gruppi alcolici
lignina alcol coniferilico saccaridiche cellulosa glucosio (β-1-4) pectine acido galatturonico, ramnosio
(β-1-4) emicellulose: xilano xilosio (β-1-4) arabino-galattano galattosio (β-1-3, ramificazioni
β-1-6), arabinosio (β-1-3) gluco-mannano glucosio e mannosio (β-1-4) galatto-mannano
(gomma guar) mannosio (β-1-4), galattosio (α-1-6)
Tab. 5 Principali fibre alimentari e loro unità monomeriche.
4. SIMBIOTICI E ALIMENTI FUNZIONALI
4.1. I simbiotici
Una possibilità alternativa per modulare o riequilibrare la microflora intestinale consiste nell’uso di simbiotici. I simbiotici sono preparati alimentari o farmaceutici che contengono sia uno o più ceppi probiotici sia un ingrediente prebiotico (Roberfroid 1998). Poiché si allude al sinergismo, questo termine dovrebbe essere utilizzato in senso stretto e riservato a prodotti

46
nei quali il prebiotico favorisce selettivamente lo sviluppo del probiotico addizionato. Tuttavia si può pensare anche che il sinergismo si attui anche in vivo, a livello del colon, mediante l’assunzione di probiotici da un lato e di carboidrati che favoriscono lo sviluppo dei bifidobatteri autoctoni dall’altro.
La somministrazione simultanea di probiotici e di un substrato da loro metabolizzabile offre ai ceppi somministrati maggiori possibilità di colonizzazione e sopravvivenza nel colon dell’ospite, potenziandone o prolungandone gli effetti benefici. Sono presenti in letteratura numerose evidenze scientifiche che supportano il positivo impatto dei simbiotici sulla microflora intestinale (Casiraghi et al. 2007; Rastall et al. 2002; Bielecka et al. 2002; Crittenden et al. 2001; Gmeiner et al. 2000). I simbiotici maggiormente utilizzati e già presenti in commercio riguardano miscele di bifidobatteri e FOS, GOS o amido resistente e miscele di lattobacilli e lattitolo.
4.2. Gli alimenti funzionali La moderna scienza dell’alimentazione è andata oltre i concetti classici,
consistenti nell’evitare carenze di nutrienti e nell’adeguatezza dell’alimentazione di base, passando al concetto di alimentazione "positiva" o "ottimale". La ricerca è oggi incentrata sull’identificazione dei componenti alimentari biologicamente attivi potenzialmente in grado di ottimizzare il benessere fisico e mentale e di ridurre anche il rischio di contrarre malattie. Si è scoperto che molti prodotti alimentari tradizionali, tra cui frutta, verdura, soia, cereali integrali e latte, contengono componenti potenzialmente benefici per la salute. Oltre a questi, si stanno sviluppando nuovi alimenti che rafforzano o incorporano tali benefici come componenti utili per i loro effetti positivi sulla salute o per i favorevoli effetti fisiologici.
Il concetto di functional food o alimenti funzionali ebbe origine in Giappone. Negli anni ‘80, le autorità sanitarie di questo paese riconobbero la

47
necessità di migliorare la qualità della vita parallelamente all’incremento dell’aspettativa di vita di un numero crescente di anziani per poter controllare i costi sanitari. Fu introdotto quindi il concetto di alimenti specificamente sviluppati per favorire la salute o ridurre il rischio di malattie. Inoltre, in seguito alla migliore comprensione delle interazioni esistenti tra geni e nutrizione (Kok 1999), si iniziò a dare maggior rilievo alle necessità biochimiche e nutrizionali individuali.
Secondo Roberfroid (Roberfroid 2000) con il termine “alimenti funzionali” non si può intendere una ben definità e caratterizzata entità ma piuttosto un vero e proprio concetto. Infatti gli alimenti funzionali possono avere molteplici definizioni, tanti quanti sono gli autori che ad essi si sono riferiti:
1. foods that may provide health benefits beyond basic nutrition (IFIC Foundation 1995)
2. foods or food products marketed with the message of the benefit to health (Riemersma 1996)
3. food and drink products derived from naturally occurring substances consumed as part of the daily diet and possessing particular physiological benefits when ingested (Hillian 1995)
4. food derived from naturally occurring substances, which can and should be consumed as part of the daily diet and which serves to regulate or otherwise affect a particular body process when ingested (Smith et al. 1996)
5. food similar in appearance to conventional food, which is consumed as part of the usual diet and has demonstrated physiological benefit and/or reduces the risk of chronic disease beyond basic nutritional functions (Health Canada 1997)

48
6. food that encompasses potentially helpful products, including any modified food or food ingredient that may provide a health benefit beyond that of the traditional nutrient it contains (Food and Nutrition Board 1994).
Qualsiasi definizione si scelga, sono evidenti alcune importanti e comuni caratteristiche. Un alimento funzionale deve:
• essere un alimento convenzionale
• essere consumato come parte usuale della dieta
• essere costituito da ingredienti naturali non sintetici
• avere effetti positivi su una o più funzioni fisiologiche
• contribuire a preservare e/o migliorare lo stato di salute e di benessere e/o ridurre il rischio di insorgenza delle malattie correlate al regime alimentare
• avere evidenze scientifiche a supporto
In seguito al crescente interesse per il concetto di alimenti funzionali e per gli health claims, l’Unione Europea ha realizzato una Azione Concertata della Commissione Europea sulla Functional Food Science in Europe (FUFOSE). Tale programma è stato coordinato con l’obiettivo di stabilire e sviluppare un approccio scientificamente fondato sulle evidenze richieste a sostegno dello sviluppo di prodotti alimentari che possono avere effetti benefici su una specifica funzione biologica, migliorando lo stato di salute e il benessere di una persona e/o riducendo il rischio di malattia.
Questo rapporto sottolinea come gli alimenti funzionali debbano comunque restare “alimenti” e dimostrare la loro efficacia nelle quantità normalmente consumate nella dieta. Funzionale può essere un alimento integrale naturale, un alimento a cui è stato aggiunto un componente, o un alimento da cui è stato eliminato un elemento con mezzi tecnologici o biotecnologici. Può anche trattarsi di un alimento in cui è stata modificata la natura di uno o più componenti, o la biodisponibilità di uno o più elementi, o una qualsiasi combinazione di queste possibilità. Può essere destinato alla

49
popolazione in genere o a gruppi specifici di persone che possono essere definiti, per esempio, in base all’età o alla costituzione genetica.
L’Azione Concertata della UE sostiene lo sviluppo di due tipi di health claims per gli alimenti funzionali, che devono sempre essere validi nell’ambito dell’alimentazione nella sua globalità e devono riferirsi a quantitativi di cibo normalmente consumati in una dieta. Tali health claims sono:
• Tipo A: alimenti che migliorano una specifica funzione fisiologica al di là del loro specifico ruolo nella crescita corporea e nello sviluppo. Questo tipo di alimenti non ha funzioni in relazione a malattie o stati patologici. Esempio può essere il caffè, per il suo contenuto in caffeina che aumenta le capacità cognitive.
• Tipo B: alimenti che riducono il rischio di una malattia. Ad esempio il pomodoro grazie al suo contenuto in licopene può ridurre il rischio di tumori.
Attualmente nell'Unione Europea manca una legislazione specifica su questa categoria di alimenti (Roberfroid 2002), infatti solo alcune nazioni possiedono una precisa legislazione riguardo definizione, etichettatura e commercializzazione degli alimenti funzionali. In Giappone, per esempio, tali alimenti sono riconosciuti e commercializzati con la sigla FOSHU (Food for Specific Health Use) e le proprietà funzionali sono comprovate da indagini scientifiche su popolazione (in vivo).
Negli ultimi anni la ricerca scientifica ha avuto quindi come importante target lo sviluppo di nuovi alimenti funzionali, partendo dalle conoscenze scientifiche di base per trovare nuovi markers da studiare e sviluppare mediante studi sperimentali, secondo la strategia rappresentata in fig. 19. Alimenti contenenti microrganismi probiotici e/o carboidrati prebiotici possono quindi essere considerati alimenti funzionali.

50
Fig. 19 Rappresentazione schematica della strategia per la scoperta e lo sviluppi di nuovi alimenti funzionali.

SCOPO DELLA TESI
I bifidobatteri sono batteri saccarolitici obbligati, appartenenti alla microflora intestinale, che fermentano gli zuccheri presenti nel colon, non assimilati dall’ospite. Poiché la presenza di bifidobatteri è associata ad effetti benefici sulla salute dell’ospite stanno trovando impiego crescente i supplementi alimentari e farmaceutici a base di prebiotici. Si tratta di oligo- e polisaccaridi non digeribili dall’uomo che raggiungono il colon stimolando selettivamente il numero e l’attività dei bifidobatteri. Il metabolismo dei carboidrati in Bifidobacterium è oggetto quindi di notevole interesse, in particolare per quello che riguarda l’idrolisi e la fermentazione dei prebiotici. I carboidrati prebiotici che destano particolare interesse anche dal punto di vista industriale sono principalmente fruttani, quali FOS e inulina, galatto-oligosaccaridi (GOS) e xilo-oligosaccaridi (XOS).
Al fine di valutare le preferenze di Bifidobacterium nei confronti di un ampia gamma di carboidrati prebiotici sono state condotte fermentazioni batch a pH non controllato di B. adolescentis MB 239 su diverse fonti di carbonio prebiotiche e sono state confrontate le affinità e le cinetiche di crescita sui rispettivi componenti mono- e disaccaridici. E’ stato quindi approfondito il comportamento fermentativo di questo ceppo nei confronti dei galatto-oligosaccaridi e xilo-oligosaccaridi, per i quali le informazioni disponibili sono inferiori rispetto ai fruttani. Sono state condotte fermentazioni batch a pH

52
controllato; per garantire la riproducibilità e la diretta confrontabilità delle prove di fermentazione svolte, è stato impiegato uno stesso terreno minimo semisintetico addizionato di volta in volta delle diverse fonti di carbonio in esame. Nel corso delle diverse fermentazioni è stato determinato il consumo dei carboidrati mediante la tecnica cromatografica HPTLC-AMD, la produzione degli acidi grassi a catena corta caratteristici dei bifidobatteri (acido lattico, acido acetico, acido formico ed etanolo) mediante HPLC e la produzione di biomassa cellulare tramite la determinazione del peso secco. È stata inoltre quantificata l’attività della β-galattosidasi in quanto in grado di idrolizzare i legami glicosidici dei GOS e del lattosio, sia a livello citoplasmatico che di superficie.
Al fine di determinare i principali parametri cinetici che descrivono la crescita, il consumo di substrato e l’accumulo di prodotti di B. adolescentis MB 239, è stato utilizzato un algoritmo di calcolo scritto in linguaggio MATLAB in grado di fornire una stima di tali parametri.
È noto che in Bifidobacterium gli oligo- e polisaccaridi possono essere idrolizzati prima o dopo l’uptake da parte del microrganismo. Per approfondire questo importante aspetto della fisiologia dei bifidobatteri, è stata studiata una
β-xilosidasi extracellulare di B. adolescentis MB 239. L’enzima è stato isolato e purificato mediante un protocollo di purificazione multi-step a partire dal surnatante di una coltura sviluppata su xilo-oligosaccaridi durante una fermentazione batch a pH controllato. Il surnatante è stato dializzato, concentrato ed fluito attraverso una colonna a scambio anionico e
successivamente, le frazioni positive al saggio della β-xilosidasi, sono state caricate in una colonna ad esclusione molecolare.
L’attività e la stabilità di questo enzima sono state caratterizzate in funzione della temperatura e del pH, sono stati calcolati i parametri cinetici km e Vmax e ne è stata definita l’affinità di substrato in reazioni di idrolisi monitorate mediante TLC. E’ stato inoltre determinato il peso molecolare della proteina responsabile di tale attività enzimatica.

53
Nell’ottica quindi di evidenziare un risvolto applicativo a queste ricerche sul genere Bifidobacterium, è stata studiata la possibilità di sviluppare prodotti alimentari funzionali contenenti simbiotici, con particolare interesse ai prodotti da forno shelf-stable. Sono stati caratterizzati 3 ceppi probiotici Lactobacillus plantarum BAR 10, Streptococcus thermophilus BAR 20 e Bifidobacterium lactis BAR 30 appartenenti alla collezione dell’azienda Barilla G&R f.lli SpA e scelti a causa della loro elevata attività antiossidante e della capacità di produrre glutatione. In particolare è stata definita la loro tolleranza ai succhi gastrici e alla bile, al fine di mimare il transito nel tratto gastro-intestinale. Successivamente è stata valutata l’affinità di ogni singolo probiotico verso 13 carboidrati prebiotici, al fine di formulare il simbiotico più efficace. Questo è stato ottenuto mediante cinetiche di sviluppo di ogni singolo ceppo, addizionando al terreno di coltura i singoli prebiotici come fonte di carbonio.
Poichè nello sviluppo di un prodotto alimentare funzionale shelf-stable è fondamentale che la popolazione probiotica sia vitale per almeno 6 mesi a temperatura ambiente, è stata studiata la sopravvivenza dei 3 ceppi inoculati in numerose matrici alimentari progettate dall’azienda Barilla.
Infine, allo scopo di valutare l’efficacia del prebiotico e la colonizzazione dei ceppi probiotici addizionati, è stato messo a punto un trial clinico in vivo su 20 soggetti sani che hanno assunto, per 30 giorni, un pre-prototipo simbiotico sviluppato dall’azienda Barilla.

54

55
MATERIALI E METODI
parte I
1. MICRORGANISMO E CONDIZIONI DI COLTURA
1.1. Microrganismo Il ceppo Bifidobacterium adolescentis MB 239 è stato ottenuto dalla
collezione dell’Ex-Istituto di Microbiologia Agraria dell’Università di Bologna (collezione del Prof. V. Scardovi). L’attribuzione tassonomica del ceppo è basata sull’ibridizzazione DNA/DNA con ceppi di collezione ed è stata recentemente confermata a livello di specie mediante la ribotipizzazione automatica utilizzando come enzima di restrizione EcoRI, e come marcatore l’operone ribosomale di E. coli.
1.2. Terreni di coltura
Bifidobacterium adolescentis MB 239 è coltivato in anaerobiosi a 37 °C in terreno Lactobacilli MRS broth addizionato di 0,5 g L-1 di L-cisteina·HCl.

56
Le fermentazioni batch con controllo di pH sono state condotte in terreno semisintetico SM avente la composizione espressa in tab 6.
Il pH del terreno è portato a pH 7.0 con NaOH 0.5 M fino e sterilizzato a 110 °C per 30 min. I carboidrati sono sterilizzati separatamente e aggiunti al terreno alla concentrazione opportuna per il particolare esperimento.
Terreno SM g L -1 Casaminoacids 5.0 acido ascorbico 10 (NH4)2SO4 10 urea 2 KH2PO4 1 MgSO4 · 7H2O 0.7 L-cisteina · HCl 0.5 NaCl 0.2 FeSO4 · 7H2O 0.01 MnSO4 · 7 H2O 0.4e-3 acido borico 0.5e-3 KI 0.1e-3 ZnSO4 · 7 H2O 0.4e-3 Na2MoO4 · 2 H2O 0.2e-3 FeCl3 · 6 H2O 0.2e-3 CuSO4 · 5 H2O 40e-6 nicotinamide 2e-03 acido p-aminobenzoico 1e-03 calcio pantotenato 1e-03 riboflavina 1e-03 tween-80 1.0
Tab. 6 Composizione del terreno SM.

57
Le fermentazioni batch senza controllo di pH di B. adolescentis MB 239 sono state condotte in terreno semisintetico SM a cui sono stati aggiunti 10 g L-1 di acetato di sodio · 3H2O per tamponare il pH durante la crescita fermentativa. Anche in questo caso i carboidrati sono stati sterilizzati separatamente e aggiunti al terreno alla concentrazione opportuna per il particolare esperimento.
La composizione dei terreni SM è tale da non permettere la crescita dei bifidobatteri in assenza di carboidrato. Gli zuccheri oggetto di studio sono aggiunti sempre in quantità tale da garantire che la crescita sia limitata dalla fonte di carbonio e non da altri componenti del terreno.
Nelle prove di fermentazione batch condotte per la determinazione del pH ideale di crescita di B. adolescentis MB 239 è stato utilizzato il terreno SM a cui sono stati aggiunti 10 g L-1 di acetato di sodio · 3H2O e 20 g L-1 di glucosio · 1H2O come fonte di carbonio ed energia.
1.3. Reattivi e carboidrati Tutti i reagenti chimici utilizzati, se non diversamente espresso, sono
stati forniti dalla Sigma-Aldrich (Steinheim, Germany), quelli microbiologici dalla Difco Laboratories (Detroit, USA). Le colonne e le resine sono state fornite dalla Amersham Pharmacia Biotech.
I carboidrati testati nel presente studio sono suddivisibili in: monosaccaridi: glucosio, fruttosio, galattosio e xilosio disaccaridi: saccarosio e lattosio trisaccaridi: raffinosio oligo- e polisaccaridi prebiotici: elencati in tab. 7.
58
PREBIOTICO COMPOSIZIONE SIGLA FORNITORE
Galatto-oligosaccaridi (S)
GOS 59%, lattosio 21%, glucosio 19%, galattosio 1%
GOS Vivinal Borculodomo, Paesi Bassi
Xilo-oligosaccaridi (S) XOS Co. Farmaceutica Milanese SpA, Milano, Italia
Frutto-oligosaccaridi (S)
DP da 2 a 5: f-nistosio 11.3%, nistosio 42.5%, l-ketosio 43.1%, saccarosio 2.4 %
FOS 1 Actilight 950P, Beghin-Meiji, Francia
Frutto-oligosaccaridi (S) DP da 3 a 10 FOS 2 P95, Orafti, Belgio
Inulina (S) DP > 20 INULINA Orafti, Belgio
Isomalto-oligosaccaridi (S) oligosaccaridi 92%, monosaccaridi 4.2% IMOS
Co. Farmaceutica Milanese SpA, Milano, Italia
Destrina resistente (S) contenente legami α-(1-4), (1-6), (1-2), (1-3) DP > 10, 61.5%
RD
Fibersol-2, Matsutani Chemical Industry Co. Ltd., Itami City, Japan
Polidestrosio (S) PD Co. Farmaceutica Milanese SpA, Milano, Italia
Arabino-galattani (S) AG Co. Farmaceutica Milanese SpA, Milano, Italia
Amido resistente (I) RS 1
Hi-Maize, National Starch and Chemical Company, Bridgewater, NJ
Amido resistente (I) RS 2 ActiStar, Cargill, Minneapolis, MN
Destrani (I) DESTRANI Puratos, Groot-Bijgaarden Belgio
Guam guar (I) PHGG Co. Farmaceutica Milanese SpA, Milano, Italia
Tab. 7 Elenco dei carboidrati prebiotici solubili (S) e insolubili (I) utilizzati, composizione se definita, sigla e fornitore.

59
1.4. Bioreattore Tutte le colture batch a pH controllato sono state condotte nel
bioreattore BM-PPS3 (Solaris Biotech, Porto Mantovano, Italia) (fig. 20). Il bioreattore ha una capacità massima di 3 L ed è equipaggiato di 2 turbine Rushton da 58 mm, pari alla metà del diametro interno del vessel, e di uno sparger toroidale da 60 mm a 25 mm dal fondo. Il fermentatore è dotato di un sistema di controllo on-line (BCS, Bioreactor Control System) che provvede al controllo, alla regolazione ed alla registrazione dei seguenti parametri di processo: agitazione, temperatura, ossigeno disciolto, pH, potenziale redox, flusso di gas, pressione, livello di schiuma, feeding e peso.
Fig. 20 Il fermentatore BM-PPS3 3000: 1) Modulo di controllo fluidi; 2) Base del vessel; 3) Vessel in vetro; 4) Piastra di testa; 5) Gas di scarico: condotto, filtro e manometro; 6) Acqua di raffreddamento; 7) Condensatore; 8) Alimentazione aria: condotto, filtro e manometro; 9) Touch-screen; 10) Modulo di misura, controllo e potenza; 11) Pompe peristaltiche per acidi, basi e antischiuma; 12) Cavi delle resistenze riscaldanti; 13) Modulo di supporto; 14) Gruppo di agitazione; 15) Acqua di raffreddamento.

60
Per tutte le prove di fermentazione, il vessel, il piping di aerazione e gli scarichi della condensa sono stati sterilizzati in-situ per 30 min a 121 °C, mentre i terreni di coltura per i bifidobatteri, così come i carboidrati oggetto di studio, sono stati sterilizzati separatamente in autoclave ed introdotti successivamente nel bioreattore sterile. Per tutte le colture batch la temperatura è stata mantenuta a 37 °C, l’agitazione a 300 rpm ed è stato applicato un flusso di N2 sterile pari a 0.05 vvm, alla pressione di 0.05 bar effettivi, per garantire l’anaerobiosi della coltura. Il pH e la concentrazione di ossigeno disciolto sono stati monitorati in continuo mediante le sonde InPro3030 e InPro6800, rispettivamente. Non si è fatto uso di antischiuma.
Le colture batch in controllo di pH sono state condotte con un volume di lavoro di 2 L. Il pH è stato mantenuto costante mediante la titolazione automatica degli acidi prodotti con NaOH 4 M. Lo schema dell’impianto sperimentale, utilizzato per i batch a pH controllato, è riportato in fig. 21.
Fig. 21 Fermentazione batch a pH controllato: schema dell’impianto.
N2
PUMP AIROUT
SAMPLE
pH PROBE
NaOH

61
2. CRESCITA B. adolescentis MB 239 SU CARBOIDRATI PREBIOTICI
2.1. Cinetiche di crescita Il ceppo B. adolescentis MB 239, cresciuto in anaerobiosi a 37 °C per
24 h in MRS addizionato di 0.5 g L-1 di L-cisteina · HCl, è stato inoculato (5% v/v) in terreno semisintetico SM a cui sono stati aggiunti 10 gL-1 di acetato di sodio · 3H2O e contenente 10 g L-1 dei carboidrati in esame come unica fonte di carbonio. Le colture sono state incubate in anaerobiosi a 37°C per 48 h e trapiantate (5 % v/v) per 3 volte nello stesso terreno. Durante l’incubazione anaerobica a 37°C è stata determinata la crescita valutando l’aumento di densità ottica a 600 nm (OD600) per le fibre solubili, mediante conteggio vitale in piastra ed espresso come colony forming unit per ml (cfu/ml) per le fibre insolubili.
. I dati riguardanti lo sviluppo del ceppo sui carboidrati presi in considerazione sono stati confrontati utilizzando il test di Student su campioni accoppiati. Le differenze sono considerate significative per P ≤ 0.05.
2.2. Colture in bioreattore
B. adolescentis MB 239 è stato coltivato in bioreattore in terreno
semisintetico SM contenente singoli carboidrati quali sole fonti di carbonio. Gli zuccheri utilizzati sono stati glucosio, galattosio, xilosio, lattosio, galatto-oligosaccaridi e xilo-oligosaccaridi. Sono state condotte fermentazioni in modalità batch a pH controllato. I carboidrati sono stati utilizzati ad una concentrazione tale che le unità monomeriche totali fossero 100 mM.

62
Le fermentazioni batch a pH controllato sono state svolte inoculando il fermentatore (5 % v/v) con colture in fase di crescita esponenziale su terreno SM contenente il medesimo carboidrato utilizzato in bioreattore. Campioni di brodo colturale delle fermentazioni su glucosio, galattosio, lattosio e galatto-oligosaccaridi sono stati prelevati periodicamente per la determinazione HPTLC-AMD dello zucchero residuo, HPLC degli acidi ed etanolo prodotti e per la misura turbidimetrica della concentrazione di biomassa.
La velocità specifica di crescita µ è stata calcolata nel corso della crescita esponenziale secondo la seguente formula in cui x rappresenta il peso secco cellulare.
ln xt2 = ln xt1 + µ (t2 – t1) La resa di bioconversione del substrato consumato in biomassa è stata
calcolata come:
SXY SX ∆
∆=/
Nelle prime ore di fermentazione, quando la concentrazione del carboidrato è significativamente superiore della costante di saturazione
dell’equazione di Monod, è stato considerato µ = µmax. Per la misura del peso secco (DW) le cellule contenute in 10 ml di
brodo colturale sono state filtrate su filtri di nitrato di cellulosa da 0.22 µm prepesati, lavate con acqua, essiccate per 24 h a 105 °C e pesate. Il rapporto DW/OD600 è stato determinato e corrisponde a 0.682 g L-1 di biomassa per unità di assorbanza.

63
3. TECNICHE ANALITICHE
3.1. Cromatografia planare HPTLC-AMD
I carboidrati presenti nelle colture di B. adolescentis MB 239 sviluppato
su glucosio, galattosio, lattosio e GOS sono stati determinati, in collaborazione con il Prof. G. Vaccari, presso l’Università degli Studi di Ferrara.
L’HPTLC è una tecnica di cromatografia planare su strato sottile ad alte prestazioni che permette la risoluzione, l’identificazione e la quantificazione dei carboidrati presenti in miscela nei brodi di fermentazione.
La metodica AMD, usata per separare miscele complesse, permette variazioni dei parametri sperimentali primari, quali la lunghezza della lastra cromatografica, la composizione della fase mobile in ciascuno sviluppo ed il numero degli sviluppi. Il piatto TLC è sviluppato per una certa distanza e tempi prescelti, quindi lo sviluppo è arrestato ed il solvente allontanato dal piatto per evaporazione. E’ possibile quindi eseguire ripetutamente il processo di eluizione secondo modalità diverse al fine di ottenere la separazione ottimale della miscela in esame. Il processo di eluizione è interamente gestito dall’apparato Camag AMD (Camag, Muttenz, Svizzera). L’apparato sperimentale è schematizzato in fig. 22.
La lastra HPTLC su cui sono stati depositati i campioni in esame è posta nella camera di sviluppo. Le sequenze operative iniziano con uno stadio di essiccazione, ottenuto sotto vuoto nella camera cromatografica chiusa ermeticamente, seguito da uno o più stadi di condizionamento dello strato: tali stadi utilizzano un flusso d’azoto secco oppure umidificato in seguito al passaggio in una bottiglia di lavaggio.
Il gas di condizionamento è immagazzinato in un contenitore di plastica, prima di essere immesso nella camera cromatografica. Possono essere utilizzate atmosfere con diversa umidità relativa o contenenti sostanze volatili,

64
come ammoniaca o acido acetico, allontanabili sotto vuoto. La fase mobile può essere una miscela contenente fino ad otto solventi opportunamente miscelati.
Fig. 22 Camag AMD apparatus. Bottiglia di miscelazione 1, riserva dei solventi 2, valvola ad 8 vie 3, scarico dei solventi 4, pompa a vuoto 5, miscelatore 6, barriera di protezione per la luce 7, camera di sviluppo 8, serbatoio per il gas 9, bottiglia di lavaggio del gas 10, rifornimento di gas 11.
L’eluizione avviene sotto pressione di N2. Lo sviluppo della corsa
cromatografica avviene per il tempo corrispondente alla distanza prestabilita. Al termine del tempo di sviluppo, l’eluente è allontanato dalla camera di sviluppo e la lastra è essiccata sotto vuoto e pronta per un successivo ciclo di eluizione. Al termine della separazione la lastra è opportunamente derivatizzata al fine di permettere la rivelazione densitometrica (Camag Dipping Apparatus e Camag Scanner III).
La separazione dei carboidrati è stata realizzata su piastre 10 x 20 cm in silice Kieselgel 60 F254s (Merck); la deposizione sulle lastre è stata effettuata mediante Camag Linomat V. Per l’eluizione è stato utilizzata una miscela di acqua ultrapura (dal 24 al 30 % v/v) e acetonitrile (Carlo Erba Reagenti) secondo un gradiente multi-step (fig. 23). Ogni stadio è stato seguito da 20 minuti di essiccamento in flusso d’azoto.

65
Per la rilevazione, le lastre sono state trattate con vapori di HCl al 36 % w/v per 30 min, essiccate in stufa a 115 °C e immerse per 2 sec in una soluzione derivatizzante contenente 4 g di acido 4-aminobenzoico, 72 ml di acido acetico glaciale, 80 ml di acqua ultrapura, 4 ml di acido ortofosforico 85% e 240 ml di acetone. Le lastre derivatizzate sono state asciugate in stufa per 12 minuti a 105 °C e successivamente sottoposte a scansione densitometrica in fluorescenza a 366 nm, filtro cut-off 400 nm, utilizzando con uno scanner dotato di lampada al mercurio interfacciato con un computer dotato di software winCATS 1.2.6 (Camag).
STEP
0 2 4 6 8 10 12 14 16
% v
/v H
2O
23
24
25
26
27
28
29
30
Fig. 23 Gradiente multi-step utilizzato nella separazione dei galatto-oligosaccaridi presenti nei brodi di fermentazione.
Per la calibrazione del metodo analitico sono state utilizzate soluzioni standard 10 mg L-1 di glucosio, galattosio e lattosio. Per quanto riguarda i GOS è stata utilizzata una soluzione 20 mg L-1 di Vivinal GOS, le cui componenti oligosaccaridiche sono state precedentemente risolte e quantificate. Le curve di

66
calibrazione sono state costruite depositando 2, 4, 6, 8 e 10 µl delle soluzioni standard sotto forma di bande da 4mm ad una velocità di deposizione di 40
µl/sec in flusso d’azoto. I campioni sono stati diluiti da un minimo di 2 ad un massimo di 100
volte con acqua e depositati su più tracce in volumi tali da fornire risposte all’interno delle rette di calibrazione.
3.2. Cromatografia HPLC
Gli acidi organici presenti nelle colture di B. adolescentis MB 239 sono stati determinati mediante HPLC.
A tal fine è stata utilizzata una colonna Aminex HPX-87H a esclusione di ioni ad una temperatura costante di 42°C.
È stata effettuata un’eluizione isocratica utilizzando come eluente acido solforico 0.01 N, preparato con acqua deionizzata filtrata con filtri da 0.45µm, con velocità di flusso 0.6 ml/min.
L’iniezione dei campione è stata effettuta mediante un sampling loop che permette un volume di iniezione di 40 µl e un’alta riproducibilità dei volumi iniettati, i campioni sono stati iniettati senza alcuna diluizione e per ognuno di essi sono state eseguite iniezioni in triplo. La separazione della sostanze in colonna è stata fatta in base al loro indice di rifrazione.
4. MODELLAZIONE MATEMATICA
Le cinetiche di crescita su glucosio, galattosio, lattosio e GOS di B. adolescentis MB 239 in fermentazione batch sono state elaborate utilizzando un modello matematico, basato sul modello cinetico di Monod “non

67
segregato” (tutte le cellule hanno proprietà identiche) e “non strutturato” (non tiene in considerazione il controllo metabolico e genetico), come descritto dalle equazioni:
Bilancio della biomassa (X):
XKXdtdX
d ⋅−⋅= µ (1)
Bilancio del substrato limitante (glucosio, galattosio, lattosio e
GOS)(S):
XmXYdt
dS
xs
⋅−⋅−=µ (2)
Bilancio dei prodotti legati alla crescita (acido lattico, acido acetico ed
etanolo) (P):
XYdtdP
px ⋅= (3)
dove
SKS
SMAX +
⋅= µµ (4)
I parametri µMAX, KS, YXS e YPX sono stati stimati per mezzo di un algoritmo di calcolo scritto in linguaggio MATLAB (MATLAB software, Versione 6.0, Mathworks). Il programma si serve, per la stima dei parametri: delle equazioni che caratterizzano il modello (eq. (1), (2), (3) e (4)) e della funzione LSQNONLIN (MATLAB Optimization Toolbox) che minimizza l’errore quadratico, tra i dati sperimentali e quelli forniti dal modello secondo l’equazione:
∑ ∑ ∑= = =
−⋅⎟⎟⎠
⎞⎜⎜⎝
⎛+−⋅+−=
n
i
n
i
n
iPXXS PsPm
YSsSmYXsXmF
1 1 1
22
222 )(1)()()( (5)

68
Dove i pedici m ed s stanno rispettivamente per dati del modello e dati sperimentali. Questa funzione è anche chiamata ‘funzione obiettivo’ del modello.
L’algoritmo richiede una stima iniziale di µMAX, KS, YXS, YPX, Kd e m, a partire dalla quale calcola in maniera iterativa il set di parametri che minimizza la somma degli errori quadratici (Larentis et al., 2004; Shene et al., 2004; Boonomee et al., 2002; Ross et al., 2004).
Per uniformare le unità di misura e semplificare i rapporti stechiometrici, tutte le concentrazioni sono fornite come mM di atomi di carbonio; per la biomassa la concentrazione (in mM di atomi di carbonio) si è calcolata a partire dal peso secco secondo la formula C1H1.95O0.46N0.63P0.02S0.01 (PM = 27,8 g mol-1).
Sono di seguiti riportati gli script di MATLAB utilizzati. Inputdata.m richiede l’inserimento delle serie di dati corrispondenti ai tempi di campionamento e alle concentrazioni di biomassa substrato e prodotto; richiede inoltre le stime iniziali dei parametri cinetici. Odes.m dichiara il sistema di equazioni differenziali utilizzate per descrivere il modello. Err.m utilizza la funzione ODE15S per risolvere il sistema di equazioni differenziali nei tempi di campionamento e fornisce il vettore degli scarti tra le concentrazioni sperimentali e quelle calcolate dal modello. Optimizer.m utilizza la funzione LSQNONLIN che chiama iterativamente err.m per minimizzare i quadrati degli scarti, infine fornisce le stime dei parametri richiesti, la radice quadrata della somma degli errori quadratici e traccia in grafico gli andamenti calcolati. inputdata.m
%intput experimental data and first extimation for fermentation %parameters %time, biomass and substrate vectors are required %first extimation for mumax, Ks, Yxs, m, Kd, Ylx and Yax are %required
disp('intput experimental data and first extimation for fermentation parameters')

69
test = 0; while test == 0 disp('enter time, biomass and substrate vectors (same length)') samplet = input('time (hour): '); samplebiomass = input('biomass (mM): '); samplesubstrate = input('substrate (mM): '); samplelactate = input('lactate (mM): '); sampleacetate = input('acetate (mM): '); sampleethanol = input('ethanol (mM): '); test = isequal (length(samplet), length(samplebiomass), length(samplesubstrate), length(samplelactate), length(sampleacetate), length(sampleethanol)); end sample = [samplebiomass' samplesubstrate' samplelactate' sampleacetate' sampleethanol']; y0 = sample(1,:); stop = samplet(length(samplet)); k(1) = input('mumax (h-1): '); k(2) = input('Ks (mM): '); k(3) = input('Yxs (mM/mM): '); k(4) = input('maintenance (h-1) '); k(5) = input('kd (h-1) '); k(6) = input('lactate Ypx (mM/mM): '); k(7) = input('acetate Ypx (mM/mM): '); k(8) = input('ethanol Ypx (mM/mM): ');
odes.m
function dy=odes(t,y,flag,par)
v1=par(1)*y(2)/(par(2)+y(2))*y(1); %Cell growth - Monod equation v2=(v1/par(3)); %Substrate for growth v3=par(4)*y(1); %Substrate for maintanance v4=par(5)*y(1); %Cell death v5=(par(6)+par(7)+par(8))*v1; %Products
dy(1)= v1-v4; %Biomass balance dy(2)=-v2-v3; %substrate balance if y(2) <= 0 %Do not let nutrient drop below 0 dy(2) = 0.; y(2) = 0.; end dy(3)=v5; %Products balance dy = dy'; err.m
%this function err.m integrates odes at experimental times according %to current parameters and calculates a vactor of mismatches between %the data and the model
function e=err(k,y0,samplet,sample) global ym,

70
par=k;
%Odes solver p0 = [y0(3) y0(4) y0(5)]; q0 = [y0(1) y0(2) (y0(3)+y0(4)+y0(5))]; options=odeset('Abstol',1e-8,'Reltol',1e-4); [tm,ym]=ode15s('odes',samplet,q0,options,par);
ratio = [(par(6)/(par(6)+par(7)+par(8))) (par(7)/(par(6)+par(7)+par(8))) (par(8)/(par(6)+par(7)+par(8)))]; Pm = [(ym(:,3)-q0(3))*ratio(1)+p0(1) (ym(:,3)-q0(3))*ratio(2)+p0(2) (ym(:,3)-q0(3))*ratio(3)+p0(3)];
%Error calculation e1 = (ym(:,1) - sample(:,1))'; %Error on biomass e2 = (par(3)*(ym(:,2) - sample(:,2)))'; %Weighed error on substrate e3 = ((pm(:,1) - sample(:,3))/par(6))'; %Weighed error on lactate e4 = ((pm(:,2) - sample(:,4))/par(7))'; %Weighed error on acetate e5 = ((pm(:,3) - sample(:,5))/par(8))'; %Weighed error on ethanol e = [e1 e2 e3 e4 e5]'; %Error vector
optimizer.m
global ym
par0 = k;
%Nonlinear least squares optimization LB = [0 0 0 0 0 0 0 0]; UB = [1000 1000 1000 1000 1000 1000 1000 1000]; options = optimset('Display','iter'); [par,Nm]=lsqnonlin('err',par0,LB,UB,options,y0,samplet,sample);
%Odes solver p0 = [y0(3) y0(4) y0(5)]; q0 = [y0(1) y0(2) (y0(3)+y0(4)+y0(5))]; options=odeset('Abstol',1e-12,'Reltol',1e-5); [t,y]=ode15s('odes',[0:0.2:stop],q0,options,par);
ratio = [(par(6)/(par(6)+par(7)+par(8))) (par(7)/(par(6)+par(7)+par(8))) (par(8)/(par(6)+par(7)+par(8)))]; p = [(y(:,3)-q0(3))*ratio(1)+p0(1) (y(:,3)-q0(3))*ratio(2)+p0(2) (y(:,3)-q0(3))*ratio(3)+p0(3)];
%Plot experimental and modeled data plot(samplet,sample(:,1),'b o', samplet,sample(:,2),'r o',

71
samplet,sample(:,3),'g o', samplet,sample(:,4),'y o', samplet,sample(:,5),'c o', t,y(:,1),'b', t,y(:,2),'r', t,p(:,1),'g', t,p(:,2),'y', t,p(:,3),'c'); xlabel('h'); ylabel('mM'); grid;
%Output SS and parameters SS=(Nm)^0.5 k=par
5. ATTIVITA’ GLICOSIL-IDROLASICHE
5.1. Preparazione dei campioni
Le attività glicosil-idrolasiche sono state determinate a livello intracellulare, di superficie ed extracellulare. A questo fine sono state saggiate le cellule permeabilizzate, le cellule intere e il surnatante della coltura. Le attività presenti sulle cellule intere corrispondono ad attività idrolitiche localizzate sulla superficie cellulare; le attività citoplasmatiche sono determinate per differenza tra cellule permeabilizzate e cellule intere; le attività extracellulari sono misurate direttamente nel surnatante della coltura.
Per ogni singola fermentazione, il campione colturale è centrifugato a 6,000 g per 10 min a 4 °C al fine di separare la biomassa dal brodo esausto.
Il surnatante è filtrato con filtro Millex®-MP da 0.22 µm (Millipore) per eliminare tutte le cellule. Successivamente tutto il surnatante raccolto è dializzato 3 volte (ciascuna per 24 ore) contro tampone fosfato 50mM a pH 7 a 4°C mediante membrana da dialisi con Cut-off da 12-14 KDa. Il surnatante dializzato viene successivamente concentrato 10 volte mediante ultrafiltrazione (membrana NMWL da 3 kDa, Millipore) e raccolto quindi per l’analisi delle attività enzimatiche extracellulari.

72
La biomassa è lavata due volte in tampone Z (0.1 M tampone fosfato pH 7, 10 mM KCl, 1mM MgSO4) e concentrata 10 volte nello stesso tampone. La preparazione del campione per ottenere le cellule lisate è stata effettuata aggiungendo 200 µl di cloroformio (CHCl3) a 1 ml di sospensione cellulare, vortexata per 10 secondi e atteso finchè fase organica e acquosa non si siano separate (1 minuto).
Durante tutte le operazioni di preparazione, i campioni sono stati mantenuti a 4°C.
5.2. β-galattosidasi
Il dosaggio della β-galattosidasi è stato determinato su campioni di cellule permeabilizzate, cellule intere e surnatante delle fermentazioni svolte in bioreattore a pH controllato con glucosio, galattosio, lattosio o GOS come fonte di carbonio. Tale attività enzimatica è valutata mediante il dosaggio spettrofotometrico del nitrofenolo, un cromoforo liberato dall’idrolisi di substrati glicosidici di sintesi in seguito all’azione del proprio enzima specifico (fig. 24). Il substrato glicosidico di sintesi utilizzato per quantificare l’attività della β-galattosidasi è il o-nitrofenil-β-D-galactopiranoside (oNPG)
oNPG galattosio + oNPG
Figura 24 oNPG e prodotti del taglio enzimatico ad opera della β -galattosidasi.

73
Ad 1 ml di campione (cellule permeabilizzate, cellule intere o surnatante) si aggiungono 200 µl di soluzione di oNPG 13 mM (2.2 mM nel volume finale). Dopo l’incubazione a 37 °C per 3 minuti, la reazione è bloccata aggiungendo 0.5 ml di Na2CO3 1M, si centrifuga a 13,000 g per 5 minuti per rimuovere i detriti cellulari (per i dosaggi con le cellule intere e lisate). Infine si legge l’assorbanza a 420 nm.
Noto il coefficiente di estinzione molare del nitrofenolo (1.49 M-1 cm-1), si calcola la concentrazione del substrato idrolizzato. Da questa si risale alla concentrazione delle unità enzimatiche, definendo l’unità come la quantità di enzima in grado di rilasciare 1 µmole di nitrofenolo in 1 minuto. Le attività specifiche sono calcolate rapportando le attività enzimatiche per il valore della biomassa secca.
5.3. β-xilosidasi Per meglio comprendere le capacità fermentative di B. adolescentis MB
239 verso gli xilo-oligosaccaridi, è stata determinata l'attività della β-xilosidasi su cellule permeabilizzate, cellule intere e surnatante delle fermentazioni svolte in bioreattore a pH controllato utilizzando, come fonte di carbonio, xilosio o XOS. La β-xilosidasi (β-Xil) extracellulare presente nel surnatante della coltura sviluppata su XOS è stata successivamente isolata e caratterizzata in collaborazione con il Dipartimento di Chimica dell’Università di Modena e Reggio Emilia.
5.3.1. Dosaggio enzimatico e quantificazione proteica L’attività della β-xilosidasi è stata determinata utilizzando come
substrato il p-nitrofenil-β-D-xylopiranoside (pNPX) alla concentrazione finale di 2.2 mM. Il dosaggio della β-xilosidasi è analogo a quello riportato per la β-

74
galattosidasi, con una variazione sul tempo di incubazione, di 30 minuti per la β-xilosidasi. Una unità di attività β-xilosidasica è definita come la quantità di enzima necessaria per rilasciare 1 µmole di nitrofenolo in 1 minuto nelle condizioni sperimentali. L’attività specifica è stata definita come unità per milligrammo di proteine totali (Amaretti et al. 2007).
La concentrazione proteica del surnatante e delle singole frazioni ottenute durante la purificazione dell’enzima è stata determinata mediante il saggio colorimetrico Lowry (Lowry 1951)
5.3.2. Purificazione dell’enzima L’enzima extracellulare è stato purificato dal surnatante di una coltura
di B. adolescentis MB 239 sviluppato in fermentazione batch a pH controllato utilizzando XOS come sola fonte di carbonio. 500 ml della coltura in tarda fase esponenziale è stata centrifugata a 5000 x g per 20 minuti a 4°C. Il surnatante raccolto è stato dializzato 3 volte (24 ore ciascuna) contro 10 L di Tris 20 mM pH 7 e concentrato a 30 ml mediante ultrafiltrazione utilizzando una membrana con NMWL pari a 3 kDa (Millipore).
Al fine di isolare l’enzima, è stato sviluppato un protocollo di purificazione costituito da 2 steps, in cui sono stati sfruttati 2 differenti tecniche cromatografiche. Il campione di surnatante dializzato e concentrato è stato inizialmente caricato su una colonna Mono-Q Sepharose a scambio anionico ed eluito con un gradiente lineare di NaCl compreso tra 0 e 1 M in tampone Tris 20 mM pH 7. Le singole frazioni da 6 ml sono state raccolte e analizzate per determinarne la concentrazione proteica e l’attività enzimatica della β-xilosidasi mediante il dosaggio colorimetrico precedentemente descritto.
Le frazioni che presentavano attività xilosidasica sono state raccolte in un’unica frazione e concentrate utilizzando il dispositivo Amicon-Ultra con NMWL da 10 kDa (Millipore).

75
Per l’isolamento finale dell’enzima, la β-xilosidasi presente nelle frazioni attive del primo step sono state successivamente sottoposte a gel filtrazione su una colonna Superdex-75 eluita con tampone Tris 20 mM pH 7. Gli step di purificazione della β-xilosidasi sono stati seguiti oltre che mediante il dosaggio dell’attività enzimatica anche mediante SDS-page per poter meglio valutare il fattore di purificazione.
5.3.3. Gel elettroforesi
Gel di poliacrilamide al 12% in condizioni denaturanti sono stati utilizzati al fine di controllare il grado di purificazione della proteina e di determinare il peso molecolare del singolo monomero.
Per confermare l’attività enzimatica della proteina isolata mediante i due step di purificazione, è stato messo a punto uno zimogramma secondo quanto descritto da Mccarthy e Kambourova (Bachman e Mccarthy 1989; Kambourova 2007). Il gel in condizioni native è stato ottenuto mediante uno stacking gel al 5% e un resolving gel al 6% di poliacrilamide. Lo zimogramma è stato poi ottenuto assemblando un sandwich in cui il gel nativo descritto
viene interposto a 2 gel di agarosio al 1% contenenti 50 µg ml-1 4-
methylumbelliferyl-7-β-D-xylopyranoside come substrato per la β-xilosidasi. Questo sandwich è stato incubato a 37°C per 2 h e poi esposto alla luce UV per evidenziare la banda fluorescente sviluppata dal prodotto dell’attività β-xilosidasica.
5.3.4. Determinazione del peso molecolare
Il peso molecolare del monomero di questa proteina è stato determinato mediante SDS-page al 12%. La determinazione del peso molecolare della proteina in forma nativa è stata ottenuta mediante cromatografia ad esclusione molecolare. È stata utilizzata una colonna Sephacryl S200 (1,6 x 70 cm)

76
(Amersham Pharmacia Biotech) attraverso la quale sono state fatte eluire proteine a peso molecolare noto (Cytochrome c 12400 Da, Carbonic Anhydrase 29000 Da, BSA 66000 Da, Alcohol Dehydrogenase 150000 Da, β-Amylase, 200000 Da, Blue dextran 2000000 Da) appartenenti al Kit for Molecular Weights 12000-200000 (Sigma) mediante le quali è stata costruita una retta di taratura. Eluendo la β-xilosidasi nelle stesse condizioni di lavoro, è stato estrapolato dalla retta di taratura il peso molecolare della proteina nativa.
5.3.5. Effetto della temperatura e del pH sull’attività e stabilità dell’enzima E’ stata studiata l’attività e la stabilità dell’enzima in funzione della temperatura e del pH. Tutte le prove sperimentali sono state condotte
incubando 0.16 mg/ml di β‐xilosidasi con una soluzione 2,2 mM di pNPX in
tampone sodio fosfato 50 mM. Inizialmente è stato determinato l’optimum di temperatura incubando le miscele di reazione a pH 7 e a differenti valori di temperatura. Sono state eseguite prove ogni 5°C per un range di temperatura compreso tra 20°C e 70°C. Dopo 30 minuti di incubazione le reazioni sono bloccate e viene letta l’assorbanza a 420 nm, calcolando quindi l’attività enzimatica.
Successivamente è stato calcolato l’optimum di pH svolgendo il dosaggio enzimatico all’optimum di temperatura precedentemente definito e a differenti valori di pH in un range compreso tra 4.5 e 8.5 con intervalli di 0.5 unità.
E’ stata studiata anche la stabilità dell’enzima: all’optimum di reattività (temperatura e pH) l’enzima è stato incubato 1, 2, 4, 6, 12 e 24 h, per poi dosare l’attività enzimatica residua.
La stabilità dell’enzima in funzione del pH è stata studiata ponendo l’enzima in un tampone di sodio fosfato 50 mM a pH 3, 5, 6, 7 e 9 a 4°C e l’attività residua è stata misurata dopo 3, 6, o 24 h riportando il pH al suo

77
optimum e incubando le miscele di reazione all’optimum di temperatura.
5.3.6. Valutazione della specificità di substrato
Per confermare la capacità della β-xilosidasi di idrolizzare gli xilo-oligosaccaridi e valutare la specificità di substrato dell’enzima purificato, è stato messo a punto un esperimento di idrolisi verso XOS, xilano, saccarosio, lattosio, cellobiosio, maltosio e raffinosio.
Le idrolisi sono state condotte incubando le miscele di reazione overnight a pH 6 e 53°C; ogni miscela di reazione, del volume di 20 µl, è composta da:
• 10 µl di β-xilosidasi purificata (596 U/ml)
• 10 µl di carboidrato (4 mg/ml)
L’idrolisi dei carboidrati è stata analizzata mediante cromatografia su strato sottile (TLC), sfruttando un sistema solvente costituito da una miscela di etanolo:n-butanolo:H2O in rapporto 3:5:2. Sulla lastra in silice sono caricati 0.5 µl di ogni miscela di reazione ed è stata condotta una doppia corsa di eluizione per migliorare la separazione dei carboidrati.
I prodotti di idrolisi sono stati visualizzati immergendo la lastra in silice in una soluzione di risoluzione in acetone costituita da:
• difenilammina 2 % (w/v)
• anilina 2 % (v/v)
• acido fosforico 15 % (v/v) e poi riscaldando la lastra a 600°C per pochi minuti.
5.3.7. Determinazione dei parametric cinetici: Vmax e Km
Al fine di completare la caratterizzazione di questo enzima, sono stati
calcolati sperimentalmente i parametric cinetici Vmax e Km. E’ stato messo a

78
punto un dosaggio enzimatico in cui 2,7 x 10-6 M β-xilosidasi sono stati incubati a pH 6 e 53°C per 30 minuti con concentrazioni di pNPX variabili comprese tra 0.2 e 50 mM.
I valori di Km and Vmax sono stati calcolati secondo il diagramma di Lineweaver-Burk (fig. 25).
Fig. 25 Diagramma di Lineweaver-Burk
5.4. Xilanasi La determinazione quantitativa dell’attività della xilanasi sfrutta un
dosaggio enzimatico differente da quello precedentemente descritto. Il campione è messo in contatto con lo xilano e gli zuccheri riducenti liberati dalla xilanasi sono quantificati per via spettrofotometrica. Dalla concentrazione degli zuccheri riducenti si risale alla concentrazione delle unità enzimatiche, definendo l’unità come la quantità di enzima che rilascia dal saccarosio 1 µmole di zucchero riducente in 1 minuto.
La determinazione di un’attività xilanasica è stata svolta sull’enzima purificato dal surnatante della fermentazione condotta su xilo-oligosaccaridi, al

79
fine di confermare la debole affinità dell’enzima anche per questo substrato. Questo dosaggio è stato svolto utilizzando una sospensione di xilano all’1% in tampone acetato 0.2 M pH 4.5. La miscela di reazione comprende:
• 600 µl di campione
• 400 µl di xilano 1%
ed è incubata a 37°C per 25 minuti. Terminata l’incubazione si aggiunge 1 ml di soluzione di Rochelle (Bernfeld et al. 1955 ; Lama et al. 2004 ) contenente:
• acido 3,5-dinitrosalicilico 10 g L-1,
• Na, K tartrato di sodio e potassio · 4H2O 300 g L-1,
• NaOH 0.4M
Si bolle il campione per 5 min e si aggiungono 10 ml di acqua distillata. Infine si legge l’assorbanza a 540 nm.
La curva di calibrazione è costruita utilizzando soluzioni di xilosio in un range di concentrazione compreso tra 0 e 20 mM. Un’unità di attività xilanasica è definita come la quantità di enzima necessario per rilasciare dallo
xilano 1 µmol di zuccheri riducenti al minuto. La relazione tra assorbanza e concentrazione degli zuccheri riducenti è data dalla seguente espressione.
A = (c · 0.0857 M-1) + 0.0163

80

81
MATERIALI E METODI
parte II
1. MICRORGANISMI E CONDIZIONI DI COLTURA 1.1. Ceppi e condizioni di coltura
I 3 microrganismi utilizzati Lactobacillus plantarum BAR 10,
Streptococcus thermophilus BAR 20 e Bifidobacterium lactis BAR 30 sono stati ottenuti dalla collezione dell’azienda Barilla G&R f.lli SpA.
L’attribuzione tassonomica dei 3 ceppi è stata ottenuta mediante amplificazione e sequenziamento del DNA ribosomiale della regione ITS (Internally Transcribed Spacer) compresa tra il 16S e 23S, mediante primers genere-specifici. I primers utilizzati per amplificare il ceppo di Bifidobacterium sono L (5’-GCTGGATCACCTCCTTTCT-3’) e R (5’-CTGGTGCCAAGGCATCCA-3’); i primers per Lactobacillus sono 16-1A (5’-GTCGGAATCGCTAGTAATCG-3’) e 23-1B (5’-GGGTTCCCCCATTCGGA-3’); i primers per Streptococcus thermophilus sono ITS-R (5’-CAAGGCATCCACCGT-3’) e ITS-F (5’-GTCGTAACAAGGTAGCCGTA-3’).

82
I ceppi Lactobacillus plantarum BAR 10 e Bifidobacterium lactis BAR 30 sono stati coltivati in anaerobiosi a 37°C in Lactobacilli MRS Broth addizionato di 0.5 g L-1 di L-cisteina·HCl mentre Streptococcus thermophilus BAR 20 è stato coltivato in aerobiosi a 42°C in M17 Broth.
Il mix di ceppi probiotici utilizzati nel pre-prototipo e quindi nel trial clinico sono Bifidobacterium longum e Lactobacillus acidophilus appartenenti alla collezione dell’azienda Barilla G&R f.lli SpA.
1.2. Reattivi e carboidrati Tutti i reagenti chimici utilizzati, se non diversamente espresso, sono
stati forniti dalla Sigma-Aldrich (Steinheim, Germany), quelli microbiologici dalla Difco Laboratories (Detroit, USA). I carboidrati prebiotici studiati nelle cinetiche di crescita dei 3 ceppi sono stati già precedentemente elencati in tab.7.
2. RESISTENZA A SUCCHI GASTRICI E BILE
Al fine di valutare la tolleranza ai succhi gastrici e alla bile, requisito fondamentale per un microrganismo probiotico, è stato allestito un esperimento in cui è stata determinata la vitalità nel tempo di L. plantarum BAR 10, S. thermophilus BAR 20 e B. lactis BAR 30 dopo essere stati a contatto con:
• succhi gastrici umani (pH 1.8)
• succhi gastrici sintetici (pepsina 3 g L-1, NaCl 0.5 g L-1, pH 2) (Charteris et al. 1998)
• bile suina

83
Ogni coltura sviluppata overnight in terreno MRS o M17 Broth è stata centrifugata a 13000 rpm per 10 minuti a 4°C. Le cellule sono state lavate 2 volte e risospese (10:1 v/v) in terreno sterile Wilkins Chalgren Anaerobe Broth 0.5x contenente 0.5 g L-1 cisteina · HCl. 500 µl di succhi gastrici umani, succhi gastrici sintetici o bile sono stati addizionati a 10 µl di sospensione batterica. Queste miscele sono state incubate per 0, 30, 60 e 90 minuti a 37°C e per ogni intervallo di tempo è stata determinata la vitalità residua mediante conta vitale in piastra utilizzando terreno agarizzato MRS o M17.
Le differenze nel conteggio vitale in piastra sono state valutate usando l’analisi statistica “one-way ANOVA” con misure ripetute, seguite dal confronto “Bonferroni post hoc”. Le differenze sono considerate statisticamente significative per P < 0.05. L’analisi statistica è stata effettuata usando GraphPad Prism 4.0 (Graphpad Software, San Diego, CA).
3. CINETICHE DI CRESCITA SU CARBOIDRATI PREBIOTICI
Al fine di formulare il simbiotico ottimale, sono state determinate
cinetiche di sviluppo di L. plantarum BAR 10, S. thermophilus BAR 20 e B. lactis BAR 30 in terreni di coltura in cui i carboidrati prebiotici selezionati sono stati singolarmente addizionati come sola fonte di carbonio ed è stato valutato lo sviluppo di ogni microrganismo in termini di resa e velocità di crescita e definirne quindi l’affinità per ogni prebiotico.
Le fermentazioni batch sono state condotte a pH non controllato e in terreno MRS o M17 in funzione del probiotico analizzato. I terreni di coltura privi dei carboidrati sono stati autoclavati per 30 minuti a 110°C; i carboidrati sono stati sterilizzati per filtrazione (mediante filtri da 0.22 µm) e addizionati al terreno sterile in modo da ottenere la concentrazione finale di 10 g L-1.

84
Come controllo negativo è stato utilizzato il terreno di coltura privo di fonte di carbonio, mentre come controllo positivo è stato addizionato glucosio.
Le cinetiche di crescita a carico di carboidrati prebiotici solubili sono state condotte in terreno liquido; le colture sono state incubate in anaerobiosi a 37°C per 48 h e trapiantate (5 % v/v) per 3 volte nello stesso terreno. Durante l’incubazione anaerobica a 37°C lo sviluppo è stato monitorato valutando l’aumento di densità ottica a 600 nm (OD600) e calcolando la velocità specifica
di crescita µ nella fase esponenziale della curva di crescita. La fermentazione dei carboidrati prebiotici insolubili è stata svolta
anch’essa in terreno liquido ma lo sviluppo dei 3 ceppi è stato monitorato mediante conteggio vitale in piastra ed espresso come colony forming unit per ml (cfu/ml).
Ogni fermentazione è stata svolta in triplo. I dati riguardanti lo sviluppo su tutti i carboidrati presi in considerazione sono stati confrontati utilizzando il test di Student su campioni accoppiati. Le differenze sono considerate significative per P ≤ 0.05.
4. PROVE DI VITALITA’ DEI CEPPI PROBIOTICI IN MATRICI ALIMENTARI
I ceppi probiotici liofilizzati L. plantarum BAR 10, S. thermophilus
BAR 20 e B. lactis BAR 30 sono stati inoculati (1% w/w) in matrici alimentari con differente composizione e posti in diverse condizioni di conservazione al fine di valutare la variabilità di sopravvivenza in funzione di questi parametri. Ad intervalli regolari di tempo è stata determinata la sopravvivenza residua del microrganismo in esame mediante conta vitale in piastra, utilizzando piastre di terreno agarizzato MRS o M17.

85
4.1. Sopravvivenza di Bifidobacterium lactis BAR 30
Le prime prove di vitalità sono state condotte su un unico ceppo, B. lactis BAR 30, il quale è stato inoculato nelle matrici alimentari descritte in tab. 8.
Matrice Composizione Condizioni
Matrice Barilla 34% grassi, 64 % carboidrati (saccarosio + glucosio)
aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
Matrice Fos 34% grassi + 64% frutto-oligosaccaridi aerobiosi, 25°C
Matrice Inulina 34% grasso + 64% inulina aerobiosi, 25°C
Matrice A matrice Barilla + acido ascorbico 0.05% aerobiosi, 25°C
Matrice B matrice Inulina + acido ascorbico 0.05% aerobiosi, 25°C
PBP 1 matrice Barilla + zuccheri “dry” aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
PBP 3 matrice Barilla + amido 3% aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
PBP 4 matrice Barilla + maltodestrine 3% aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
PBP 7 matrice Barilla + inulina 5% aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
PBP 10 matrice Barilla + proteine del latte 3% aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
PBP 11c matrice Barilla + Raftilose 6% aerobiosi, 25°C anaerobiosi, 25°C

86
aerobiosi, 4°C
PBP 12 impiego processo differente 2 aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
RG impiego processo differente aerobiosi, 25°C anaerobiosi, 25°C aerobiosi, 4°C
Matrice 1 grasso 100% aerobiosi, 25°C
Matrice 2 RG 100% aerobiosi, 25°C
Matrice 3 RG 80 % + inulina 20% aerobiosi, 25°C
Matrice 4 RG 80 % + amido resistente 20% aerobiosi, 25°C
Matrice 5 RG 80 % + saccarosio 20% aerobiosi, 25°C
Matrice 6 RG 80 % + lattosio 20% aerobiosi, 25°C
Matrice 7 RG 80 % + trealosio 20% aerobiosi, 25°C
Matrice 8 RG 80 % + maltosio 20% aerobiosi, 25°C
Matrice 9 RG 80 % + maltodestrine 20% aerobiosi, 25°C
Matrice 10 RG 80 % + FOS 20% aerobiosi, 25°C
Matrice 11 RG 80 % + inulina 10% + maltodestrine 10% aerobiosi, 25°C
Matrice 12 RG 80 % + inulina 10% + trealosio 5% + maltodestrine 5%
aerobiosi, 25°C
Matrice 13 RG 80 % + amido 10% + maltodestrine 10% aerobiosi, 25°C
Tab. 8 Composizione e condizioni di conservazione delle matrici alimentari in cui è stato inoculato il ceppo B. lactis BAR 30 e di cui è stata determinata la vitalità nel tempo.
4.2. Vitalità di una miscela dei 3 ceppi probiotici in matrice di cioccolato
Dopo aver valutato le cinetiche di crescita dei 3 ceppi sui carboidrati
prebiotici e dopo aver studiato la vitalità del probiotico in matrici alimentari a base di grasso e carboidrati, sono state formulate matrici a base di cioccolato

87
fondente. Tali barrette di cioccolato sono state addizionate singolarmente dei 3 prebiotici per i quali i probiotici presentavano una migliore affinità. Queste barrette sono state inoculate all’1% con una miscela dei 3 ceppi liofilizzati Lactobacillus plantarum BAR 10, Streptococcus thermophilus BAR 20 e Bifidobacterium lactis BAR 30 presenti tutti in elevate concentrazioni al momento della preparazione delle barrette. Quindi sono state formulate le seguenti matrici con base cioccolato:
- Cioccolato tq = cioccolato puro - Cioccolato FOS 1 = cioccolato + frutto-oligosaccaridi Actilight - Cioccolato GOS = cioccolato + galatto-oligosaccaridi - Cioccolato FOS 2 = cioccolato + frutto-oligosaccaridi Raftilose P95
Queste matrici sono state conservate ad una temperatura ambiente controllata di 25 °C e sono stati effettuati conteggi di vitalità (mediante conta vitale in piastra) per i microrganismi inoculati agli intervalli di 1, 2, 3, 4, 5 e 6 mesi di conservazione al fine di valutare se i prebiotici addizionati potessero offrire una differenziata protezione per la sopravvivenza dei probiotici in un prodotto alimentare conservato a temperatura ambiente. Il cioccolato è stato addizionato al 20% di fibre prebiotiche e non è stato possibile realizzare un arricchimento in pari quantità di FOS e GOS perché non si dispone di una materia prima fonte di GOS altamente concentrata come per i FOS e che sia anche anidra per l’utilizzo in cioccolato; il dosaggio superiore al 20%, risulta critico per le sensibili difficoltà tecnologiche di produzione delle barrette.
5. TRIAL CLINICO
Dopo la formulazione di un prodotto da forno pre-prototipo da parte della ditta Barilla, è stato messo a punto un trial clinico in vivo su volontari sani per valutare il simbiotico prescelto, osservando quindi l’efficacia del

88
prebiotico e la colonizzazione dei ceppi probiotici addizionati. Il trial clinico è stato svolto dal gruppo del prof. Campieri, presso il dipartimento di Gastroenterologia dell’ospedale S. Orsola.
5.1. Trial clinico in vivo
Per poter svolgere tale prova è stato necessario individuare alcuni volontari sani con una microflora intestinale carente in bifidobatteri e lattobacilli al fine di rendere più evidente la possibile colonizzazione dei probiotici assunti con il prodotto finale e l’effetto prebiotico dei carboidrati addizionati. Questo è stato effettuato analizzando il contenuto fecale di bifidobatteri e lattobacilli di 30 soggetti sani.
Il trial clinico è stato svolto arruolando altri volontari sani in modo random fino a raggiungere un numero totale di 20 soggetti partecipanti.
Prima della sperimentazione, ogni volontario è stato “monitorato” sia dal punto di vista della microflora intestinale che dal punto di vista clinico generale. Dal punto di vista microbiologico, per ciascuno di essi è stata svolta la conta dei lattobacilli e bifidobatteri a livello fecale al tempo 0 della sperimentazione, quindi prima di assumere il prodotto proposto, al fine di valutare la colonizzazione da parte dei ceppi utilizzati nel pre-prototipo sviluppato durante e dopo il trattamento. Dal punto di vista clinico, ogni soggetto sano partecipante alla sperimentazione è stato visitato e sono stati registrati alcuni parametri clinici (età, peso, sesso, pressione sanguigna, tipo di dieta abituale, terapie antibiotiche recenti, frequenza dell’alvo).
Il pre-prototipo sviluppato è un biscotto con cacao: la miscela di 2 ceppi probiotici Bifidobacterium longum e Lactobacillus acidophilus (109 batteri vivi/biscotto) è contenuta nel sottile strato di cioccolato, mentre la fibra prebiotica (frutto-oligosaccaridi) nella parte di biscotto vera e propria (0.5g/biscotto).

89
Il trial clinico è stato svolto per 1 mese e ogni volontario ha assunto 2 biscotti al giorno. Sono state raccolte le feci all’inizio del trattamento, al 30° giorno e dopo 15 giorni di wash-out (quindi dopo 15 giorni dal termine dell’assunzione del biscotto). Questi campioni fecali sono stati analizzati dal punto di vista microbiologico mediante tecniche di microbiologia classica e quindi ne è stato analizzato il contenuto di lattobacilli e bifidobatteri mediante conta vitale in piastra sui terreni selettivi successivamente descritti. Dal punto di vista clinico, al termine del trial è stata consegnata ad ogni volontario una scheda di valutazione (riassunta in modo schematico in fig. 26) in cui venivano valutati i seguenti parametri clinici, durante e dopo il “trattamento”.
Fig. 26 Scheda di valutazione clinica
5.2. Isolamento e conteggio di bifidobatteri e lattobacilli da feci
Le feci raccolte durante il trial clinico sono state analizzate dal punto di
vista microbiologico valutando la concentrazione di bifidobatteri e lattobacilli mediante conta vitale in piastra su terreni selettivi.
Definire i seguenti parametri in un range da 0 (nulla) a 10 (max)
DURANTE e NEL MESE SUCCESSIVO l'assunzione del biscotto:
gonfiore
dolore addominale
diarrea
stipsi
descrivere qualsiasi variazione dell'alvo ("abitudini intestinali")

90
E’ preparata una sospensione batterica partendo da feci fresche omogeneizzate con tampone di Sodio fosfato 100 mM a pH 7, in modo tale da ottenere una concentrazione di materiale fecale del 10 % sul volume totale. La sospensione è filtrata attraverso un setaccio di metallo 100 µm per rimuovere le particelle voluminose.
I campioni sono diluiti serialmente in terreno Wilkins Chalgren Anaerobic Broth (Oxoid, Basingstoke, UK). Per la preparazione del terreno, utilizzato alla concentrazione 0.5x, la polvere è disciolta nel volume doppio rispetto a quello suggerito, la soluzione è addizionata di L-cisteina cloridrato 0.5 g L-1, portata a pH 7 con NaOH 0.5 M, dispensata in tubi da 9 ml e sterilizzata a 110°C per 30 minuti.
Per l’isolamento di ceppi di bifidobatteri sono utilizzate piastre di terreno Raffinose-Bifidobacterium Agar (Hartemink et al., 1996), inoculate con 100 µl di ogni diluizione dei campioni e incubate anaerobicamente a 37°C per 48 h. Il terreno Raffinose-Bifidobacterium Agar ha la seguente composizione (gL-1):
Acetato di sodio 2.5 D(+)Raffinosio 7.5 Caseinato di sodio 5.0 Estratto di lievito 5.0 Cloruro di litio 3.0 Propionato di sodio 15.0 L-Cisteina cloridrato 0.5 Tioglicolato di sodio 0.5 Rosso di bromocresolo 10.0 ml L-1 Soluzione di Sali 40.0 ml L-1
La soluzione di sali RB è così composta (gL-1):
MgSO4 0.2 CaCl2 0.2 K2PO4 1.0

91
KH2PO4 1.0 NaHCO3 10.0 NaCl 2.0
Il pH del terreno è alcalinizzato con NaOH 0.5M fino a pH 6.7-6.8. Dopo l’aggiunta di 18 g/l di agar, il terreno è sterilizzato a 110°C per 30 minuti. Tutte le analisi e le preparazioni dei campioni sono effettuate entro una camera anaerobica (Anaerobic System, Mod. 2028, Forma Scientific Co., Marietta, USA) contenente 10% H2, 10% CO2, 80% N2. Prima dell’utilizzo, il terreno è mantenuto per almeno 24 h in ambiente anaerobico.
Per l’isolamento di ceppi di lattobacilli è utilizzato il terreno LAMVAB Agar (Hartemink et al., 1997).
Per ottenere 1 L di terreno LAMVAB si prepara la soluzione A, sciogliendo in 500 ml di acqua i seguenti componenti:
MRS Broth (DIFCO) 52.2 g/l Cisteina 0.25 g/l Verde Bromocresolo 3 ml/l.
La soluzione A è portata a pH 5 con HCl 0.5M e sterilizzata in autoclave a 110°C per 30’.
18 g di agar sono risospesi in 500 ml di acqua e sterilizzati a 110°C per 30’ (soluzione B). Dopo essere state raffreddate a 55°C, le soluzioni A e B sono unite. Il terreno agarizzato è addizionato di 10 ml di vancomicina (10 mg/ml in acqua) prima di essere versato in piastra.

92

93
RISULTATI
parte I
1. CRESCITA DI B. ADOLESCENTIS MB 239 SU
CARBOIDRATI PREBIOTICI
Bifidobacterium adolescentis MB 239 è stato sottoposto a uno screening per la capacità di utilizzare numerosi carboidrati prebiotici come unica fonte di carbonio. I prebiotici testati sono di diversa natura: glucosidi, galattosidi, fruttosidi e xilosidi. La crescita di B. adolescentis MB 239 su ognuno di essi è stata confrontata con quella dei rispettivi componenti mono- e disaccaridici.
A tale fine sono state condotte fermentazioni batch a pH non controllato su terreno SM contenente 10 g L-1 dei carboidrati in esame monitorando nel tempo l’aumento di torbidità delle colture.
Nelle figure 27, 28, 29 e 30 sono mostrate le curve di crescita del ceppo rispettivamente su fruttosidi (fruttosio, saccarosio, FOS 1, FOS 2 e inulina), galattosidi (galattosio, lattosio, raffinosio, GOS e AG), glucosidi (IMOS, destrina resistente RD, polidestrosio PD) e infine xilosidi (xilosio e XOS). Ogni categoria di carboidrati è stata confrontata con il glucosio come controllo. Le velocità specifiche di crescita delle fermentazioni sui glucosidi,

94
fruttosidi, galattosidi e xilosidi sono invece mostrate nelle tabelle 9, 10, 11 e 12.
B. adolescentis MB 239 fermenta bene tutti i glucosidi testati, la velocità specifica di crescita e le concentrazioni cellulari ottenute in fermentazione sono minori per la destrina resistente RD, maltodestrina resistente altamente ramificata.
t (h)
0 5 10 15 20 25 30
OD
600
0,0
0,5
1,0
1,5
2,0
2,5
3,0
glucosioIMOSRDPD
Fig. 27 Curve di crescita di B. adolescentis MB 239 su diversi glucosidi prebiotici e rispettivi
componenti mono- e disaccaridici.
Carboidrato µ (h-1) Glucosio 0.30
IMOS 0.50 Destrina resistente RD 0.36
Polidestrosio PD 0.54 Tab. 9 Velocità specifiche di crescita (µ) di B. adolescentis MB 239 su glucosidi prebiotici.

95
La fermentazione dei fruttani risulta essere strettamente dipendente dal grado di polimerizzazione (DP) del carboidrato. Infatti il microrganismo cresce bene su frutto-oligosaccaridi a basso DP come FOS 1 (µ = 0.53 h-1) e FOS 2 (µ = 0.48 h-1) mentre non viene fermentata l’inulina (µ = 0.16 h-1), polisaccaride dal DP maggiore di 10.
t (h)
0 5 10 15 20 25 30
OD
600
0,0
0,5
1,0
1,5
2,0
2,5
3,0
glucosiofruttosiosaccarosioFOS 1FOS 2inulina
Fig. 28 Curve di crescita di B. adolescentis MB 239 su diversi fruttosidi prebiotici e rispettivi componenti mono- e dicaccaridici.
Carboidrato µ (h-1)
Glucosio 0.30 Fruttosio 0.19
Saccarosio 0.47 FOS 1 0.53 FOS 2 0.48
Tab. 10 Velocità specifiche di crescita (µ) di B. adolescentis MB 239 su fruttosidi prebiotici.
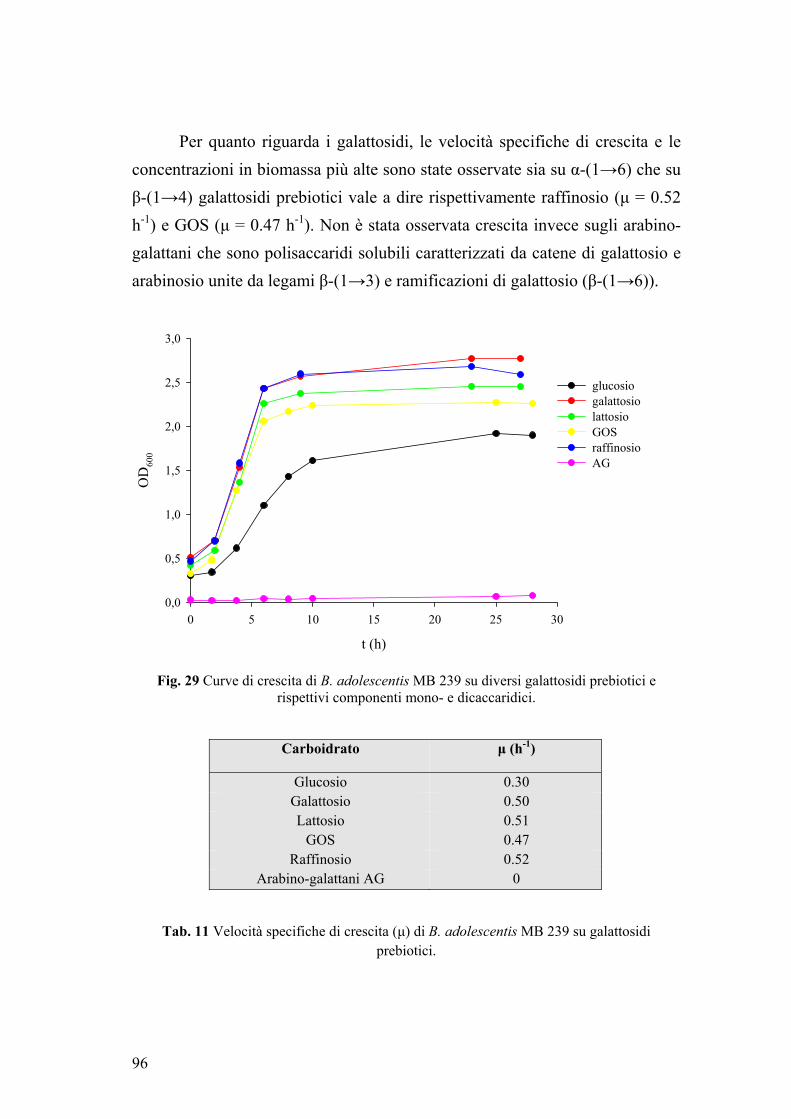
96
Per quanto riguarda i galattosidi, le velocità specifiche di crescita e le concentrazioni in biomassa più alte sono state osservate sia su α-(1→6) che su β-(1→4) galattosidi prebiotici vale a dire rispettivamente raffinosio (µ = 0.52 h-1) e GOS (µ = 0.47 h-1). Non è stata osservata crescita invece sugli arabino-galattani che sono polisaccaridi solubili caratterizzati da catene di galattosio e arabinosio unite da legami β-(1→3) e ramificazioni di galattosio (β-(1→6)).
t (h)
0 5 10 15 20 25 30
OD
600
0,0
0,5
1,0
1,5
2,0
2,5
3,0
glucosiogalattosiolattosioGOSraffinosioAG
Fig. 29 Curve di crescita di B. adolescentis MB 239 su diversi galattosidi prebiotici e
rispettivi componenti mono- e dicaccaridici.
Carboidrato µ (h-1)
Glucosio 0.30 Galattosio 0.50 Lattosio 0.51
GOS 0.47 Raffinosio 0.52
Arabino-galattani AG 0
Tab. 11 Velocità specifiche di crescita (µ) di B. adolescentis MB 239 su galattosidi prebiotici.

97
B. adolescentis MB 239 è inoltre in grado di fermentare xilo-oligosaccaridi con velocità specifica di crescita µ = 0.30 h-1.
t (h)
0 5 10 15 20 25 30
OD
600
0.0
0.5
1.0
1.5
2.0
2.5
3.0
glucosioxilosioXOS
Fig. 30 Curve di crescita di B. adolescentis MB 239 su diversi glucosidi prebiotici e rispettivi
componenti mono- e dicaccaridici.
Carboidrato µ (h-1)
Glucosio 0.30 Xilosio 0.16 XOS 0.30
Tab. 12 Xilosidi: velocità specifiche di crescita (µ) di B. adolescentis MB 239 su xilosidi
prebiotici.
È stato inoltre osservato che B. adolescentis MB 239 è in grado di fermentare fonti di carbonio rappresentate da polisaccaridi insolubili e in particolare amido resistente RS 2 e gomma guar poiché dopo 24 ore il numero

98
di CFU ml-1 aumenta significativamente e rimane circa costante dopo 48 ore (fig. 31).
1,E+06
1,E+07
1,E+08
1,E+09
1,E+10
1 2 3t
CFU
ml-1
Fig. 31 Concentrazione cellulare di B. adolescentis MB 239 sviluppato su carboidrati insolubili. Colori: bianco, controllo negativo; nero, glucosio; rosso, amido resistente RS 2; lilla, amido resistente RS 1; verde, destrani; azzurro, gomma guar PHGG. I tempi 1, 2, 3, indicano rispettivamente 0, 24 e 48 h di incubazione.
2. CARATTERIZZAZIONE FERMENTATIVA DI B. ADOLESCENTIS MB 239
2.1. Determinazione del pH ottimale di crescita
Per determinare il pH ottimale di crescita del ceppo B. adolescentis MB 239 sono state condotte in bioreattore fermentazioni batch a pH controllato in terreno SM contenente 100 mM di glucosio.

99
Le curve di crescita e di consumo del correttore di pH ai pH 4.5, 5.0, 5.5, 6.0 e 6.5 sono riportate in fig. 32.
Come risulta evidente dalla figura la crescita a pH 4.5 risulta essere completamente inibita: non si osserva produzione né di biomassa né di acidi (stimabili in basa alla quantità di NaOH consumata).
Se il pH viene mantenuto a 6.5 e 6.0, B. adolescentis MB 239 mostra una velocità specifica di crescita µ = 0.14 h-1 e 0.15 h-1 rispettivamente, con produzioni massime di biomassa rispettivamente 1.2 e 0.5 g L-1 e rese di bioconversione in biomassa pari a 0.052 C-mol C-mol-1 a pH 6.5 e 0.136 C-mol C-mol-1 a pH 6.0. A pH 5.0 la velocità specifica di crescita risulta essere piuttosto bassa (µ= 0.04 h-1).
Il pH 5.5 è risultato il pH ottimale di crescita con µ = 0.21 h-1 e resa in biomassa 0.148 C-mol C-mol-1. È da notare inoltre che tanto maggiore è la produzione in biomassa minore risulta essere quella degli acidi, infatti a pH 5.5 vengono consumate circa 125 mmoli di NaOH 4M mentre a 5.0, 6.0 e 6.5 vengono consumate rispettivamente circa 140, 220, 230 mmoli di NaOH 4M.
In tabella 13 sono riassunte le velocità specifiche di crescita e le rese di bioconversione di substrato in biomassa.
pH µ (h-1) YX/S (C-mol C-mol-1)
4.5 0.02 0.004 5.0 0.04 0.143 5.5 0.21 0.148 6.0 0.15 0.136 6.5 0.14 0.052
Tab. 13 Confronto delle velocità specifiche di crescita (µ) e delle rese di conversione del
substrato in biomassa (Yx/s) di B. adolescentis MB 239 a pH 4.5, 5.0, 5.5, 6.0 e 6.5

100
a)
t (h)
0 10 20 30 40 50 60 70
Cor
retto
re p
H (m
M)
0
50
100
150
200
250
DW
(mg
L-1)
0
200
400
600
800
1000
1200
1400
1600
pH 4.5pH 5.0pH 5.5pH 6.0pH 6.5
b)
Fig. 32 Fermentazioni batch di B. adolescentis MB 239 a pH 4.5, 5.0, 5.5, 6.0 e 6.5. a) curve di crescita b) consumo di NaOH 4M.

101
2.2. Fermentazione di glucosio
Per la caratterizzazione fermentativa di B. adolescentis MB 239 sono state condotte fermentazioni batch a pH controllato pari a 5.5 precedentemente stimato come il pH ottimale di crescita. La fermentazione del glucosio è stata condotta in terreno SM a cui è stato addizionato glucosio che quindi risulta essere l’unica fonte di carbonio fermentabile.
In figura 33 sono riportati gli andamenti delle curve di crescita della biomassa, del consumo di substrato, della produzione degli acidi organici nonché l’andamento dell’attività specifica della β-galattosidasi.
Dalla curva di crescita della biomassa si può notare che vi è una lag phase della durata di circa 4 ore seguita da un’unica fase di crescita esponenziale, questa fase dura circa 13 ore ed è caratterizzata da una velocità specifica di crescita µ = 0.31 h-1. La coltura successivamente entra in fase stazionaria durante la quale non si osserva più aumento della biomassa ma piuttosto una diminuzione della concentrazione cellulare dovuta alla lisi batterica. Di conseguenza la produzione massima di biomassa risulta essere raggiunta alla 17a ora ed è uguale a 2.75 gl-1 ossia 94.2 mM.
Il glucosio, unico substrato fermentabile, è fornito a una concentrazione a pari a 100 mM equivalente a 600 mM di atomi di carbonio. Come è evidenziato dalla sua cinetica di consumo, il substrato viene velocemente consumato e risulta completamente depleto dal terreno di crescita alla fine della fase esponenziale causando evidentemente l’ingresso della coltura in fase stazionaria di crescita. I prodotti del metabolismo, acido lattico, acido acetico ed etanolo sono tutti prodotti legati alla crescita. Ciò è supportato dal fatto che il loro accumulo, cosi come il consumo del correttore del pH, avviene esclusivamente durante la fase di crescita esponenziale. Le concentrazioni finali dell’acido lattico, dell’acido acetico e dell’etanolo misurate nel terreno al termine della fermentazione sono rispettivamente 74, 90 e 45 mM corrispondenti a 222, 180 e 90 mM di atomi di carbonio. In nessuna delle
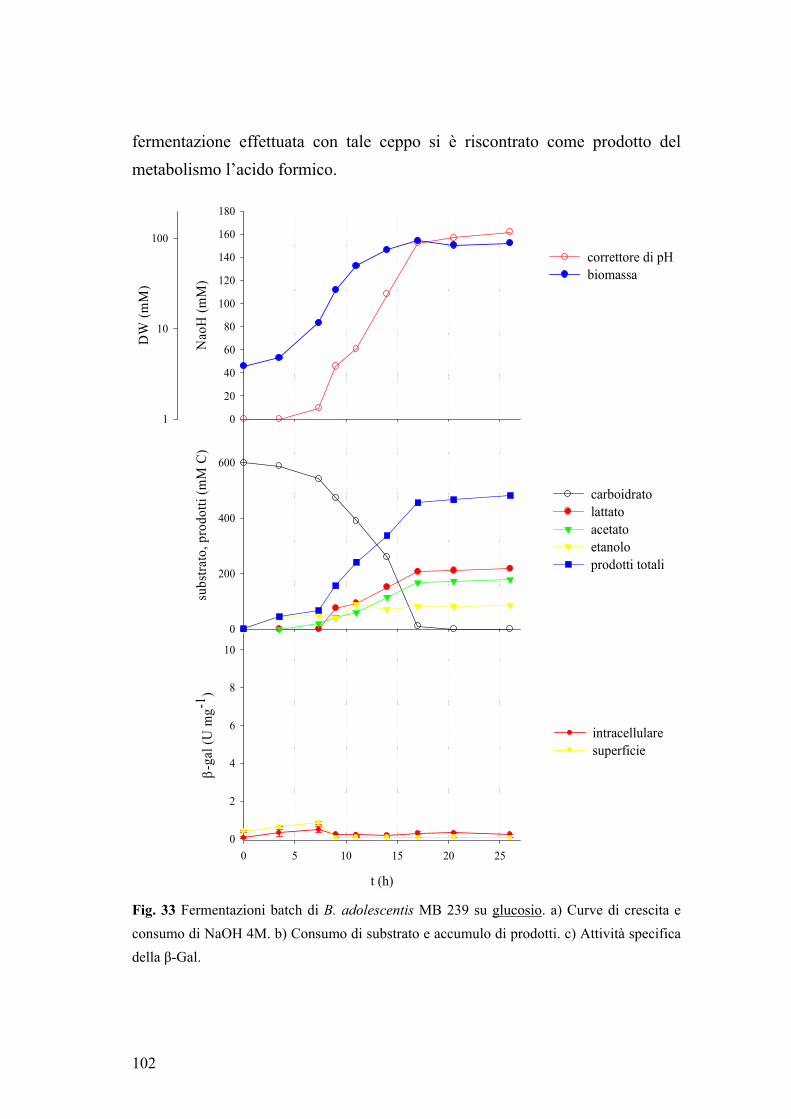
102
fermentazione effettuata con tale ceppo si è riscontrato come prodotto del metabolismo l’acido formico.
t (h)
0 5 10 15 20 25
β-ga
l (U
mg-1
)
0
2
4
6
8
10
intracellularesuperficie
subs
trato
, pro
dotti
(mM
C)
0
200
400
600
carboidratolattatoacetatoetanoloprodotti totali
Nao
H (m
M)
0
20
40
60
80
100
120
140
160
180
DW
(mM
)
1
10
100
correttore di pHbiomassa
Fig. 33 Fermentazioni batch di B. adolescentis MB 239 su glucosio. a) Curve di crescita e consumo di NaOH 4M. b) Consumo di substrato e accumulo di prodotti. c) Attività specifica della β-Gal.

103
Infine è stata determinata l’attività specifica dell’enzima β-galattosidasi sia a livello citoplasmatico che di superficie: è evidente che il microrganismo non produce l’enzima poiché le attività specifiche in entrambi i comparti cellulari sono minori di 0.9 unità per mg di peso secco.
2.3. Fermentazione del galattosio
Anche in questo caso la fermentazione di B. adolescentis MB 239 è
stata condotta in terreno SM a pH 5.5 addizionato questa volta di galattosio come unica fonte di carbonio fermentabile.
In figura 34 sono riportati gli andamenti delle curve di crescita della biomassa, del consumo di substrato, della produzione degli acidi organici e dell’attività specifica del enzima β-galattosidasi. Dalla curva di crescita della biomassa è possibile evidenziare che non vi è una lag phase ma inizia subito la fase esponenziale di crescita caratterizzata da una µ = 0.46 h-1 e da una durata complessiva di 9 ore.
Dopo la fase esponenziale la coltura entra in fase stazionaria durante la quale si ha un lieve calo della concentrazione di biomassa dovuto anche in questo caso a lisi cellulare. La quantità massima di biomassa prodotta è pari a 196 mM e quindi circa 5.4 g L-1. La concentrazione iniziale di galattosio è di 600 mM di atomi di carbonio. Anche in questo caso il substrato viene utililizzato molto rapidamente e risulta a completamente consumato al termine delle 9 ore della fase di crescita esponenziale.
L’analisi dei prodotti rivela che anche in questo caso l’accumulo di acido lattico e acido acetico avviene principalmente durante la crescita esponenziale e rallenta in fase stazionaria. Le concentrazioni finali dei due acidi prodotti sono: 65 mM (195 mM di atomi di C) per l’acido lattico e 109 mM (218 mM di atomi di C) per l’acido acetico, non viene prodotto invece etanolo.

104
subs
trato
, pro
dotti
(mM
C)
0
200
400
600
carboidratolattatoacetato etanoloprodotti totali
NaO
H (m
M)
0
50
100
150
200
250D
W (m
M)
1
10
100
correttore di pHbiomassa
t (h)
0 5 10 15 20 25
β-ga
l (U
mg-1
)
0
2
4
6
8
10
intracellularesuperficie
Fig. 34 Fermentazioni batch di B. adolescentis MB 239 su galattosio. a) Curve di crescita e consumo di NaOH 4M. b) Consumo di substrato e accumulo di prodotti. c) Attività specifica della β-Gal.

105
L’andamento dell’attività specifica della β-Gal nel citoplasma può essere diviso in due fasi: diminuisce durante la fase esponenziale di crescita (da 5.4 a 0.9 U/mgDW) e aumenta durante la fase stazionaria (da 0.9 a 3 U/mgDW). Sulla superficie cellulare invece rimane costante durante le prime 5 ore di fermentazione per poi ridursi in tarda fase esponenziale e all’entrata in fase stazionaria.
2.4. Fermentazione del lattosio
La fermentazione batch è stata effettuata a pH 5.5 in terreno SM
contenente lattosio come sola fonte di carbonio (fig. 35). L’andamento della curva di crescita della biomassa può essere diviso in
fase esponenziale e fase stazionaria dato che non si osserva la lag phase iniziale. La durata della fase esponenziale è di circa 12 ore e la velocità specifica di crescita è µ = 0.39 h-1, durante la fase stazionaria la concentrazione della biomassa rimane costante. La quantità massima di biomassa prodotta è di 3.4 g L-1.
Il lattosio, unico substrato fermentabile, è fornito ad una concentrazione pari a 600 mM di atomi di carbonio.
È riscontrabile dalla sua cinetica di consumo che il substrato viene rapidamente consumato e risulta completamente depleto dal brodo di fermentazione alla fine della fase esponenziale portando la coltura in fase stazionaria di crescita. Quando il lattosio viene utilizzato come sola fonte di carbonio, si osserva l’accumulo di glucosio e galattosio nelle prime 12 ore di fermentazione. I monosaccaridi generati vengono consumati quando il lattosio è completamente depleto.
I principali prodotti della fermentazione del lattosio sono acido lattico e acido acetico e anche in questo caso la produzione di etanolo può essere considerata trascurabile (13 mM-C). La concentrazione finale dell’acido lattico

106
è di 58 mM (174 mM di atomi di carbonio) mentre quella dell’acido acetico è di 101 mM (202 mM di atomi di carbonio).
subs
trato
, pro
dotti
(mM
C)
0
200
400
600
carboidratolattatoacetato etanoloprodotti totali
NaO
H (m
M)
0
50
100
150
200
250
DW
(mM
)
1
10
100
correttore di pHbiomassa
t (h)
0 5 10 15 20 25
β-ga
l (U
mg-1
)
0
2
4
6
8
10
intracellularesuperficie
Fig. 35 Fermentazioni batch di B. adolescentis MB 239 su lattosio. a) Curve di crescita e consumo di NaOH 4M. b) Consumo di substrato e accumulo di prodotti. c) Attività specifica della β-Gal.

107
L’attività specifica dell’enzima β-galattosidasi è stata determinata sia a livello citoplasmatico che di superficie: sulla superficie cellulare l’attività enzimatica rimane pressoché costante mentre a livello citoplasmatico aumenta proporzionalmente alla biomassa durante tutta la fase di crescita. L’attività massima è stata misurata alla 15a ora di fermentazione ed è pari a 11.1 U/mgDW.
Gli andamenti delle curve di crescita della biomassa, del consumo di substrato, della produzione degli acidi organici e dell’attività specifica del enzima β-galattosidasi sono riportati in figura 35.
2.5. Fermentazione dei GOS
La fermentazione di B. adolescentis MB 239 è stata condotta in terreno SM a pH 5.5 addizionato di Vivinal GOS, una miscela di glucosio, galattosio, lattosio e galatto-oligosaccardi con diverso livello di polimerizzazione.
In figura 36 sono riportati gli andamenti delle curve di crescita della biomassa, del consumo di substrato, della produzione degli acidi organici e dell’attività specifica del enzima β-galattosidasi; in figura 37 è presentato l’andamento degli zuccheri residui (in tutte le loro frazioni) analizzati in HPTLC-AMD.
In questa fermentazione il ceppo mostra una fase esponenziale caratterizzata da una velocità specifica di crescita di µ = 0.42 h-1 e della durata di 11.5 ore al termine della quale si ottiene la produzione massima in biomassa che è uguale a 107 mM. Si nota tuttavia una velocità specifica di crescita decrescente rispetto a quello precedentemente descritta; questo comportamento è forse caudato da un differente utilizzo dei componenti della miscela. Infatti la concentrazione iniziale dei GOS nel terreno di coltura è 600 mM di atomi di carbonio e in fig. 39 è descritto l’andamento dei carboidrati monitorati nel tempo.

108
subs
trato
, pro
dotti
(mM
C)
0
200
400
600
800
carboidratolattatoacetato etanoloprodotti totali
NaO
H (m
M)
0
100
200
300
400
DW
(mM
)
1
10
100
correttore di pHbiomassa
t (h)
0 5 10 15 20 25
β-ga
l (U
mg-1
)
0
2
4
6
8
10
intracellularesuperficie
Fig. 36 Fermentazioni batch di B. adolescentis MB 239 su GOS. a) Curve di crescita e consumo di NaOH 4M. b) Consumo di substrato e accumulo di prodotti. c) Attività specifica della β-Gal.

109
t (h)
0 5 10 15 20
Car
boid
rati
(C-m
M)
0
50
100
150
200
250
Bio
mas
sa (m
M)
20
40
60
80
100
120
Fig. 37 Fermentazioni batch di B. adolescentis MB 239 in terreno SM contenente 600 mM-C GOS come sola fonte di carbonio. Andamento della biomassa e dei carboidrati residui: ▲ biomassa, glucosio, ▼ galattosio, lattosio, DP 3, DP 4, DP 5, DP 6, DP 7 - 9.
Inizialmente vengono utilizzati il lattosio e il componente a DP 3, dopo
la quinta ora inizia ad essere fermentato anche il glucosio; glucosio, lattosio e DP 3 sono depleti rispettivamente dopo 12, 14 e 15 h, rispettivamente. Quando il lattosio è completamente esaurito, anche i carboidrati a DP compreso tra 4 e 9 vengono fermentati; in questa fase viene fermentato anche il galattosio che si era precedentemente accumulato dall’idrolisi del lattosio e dei carboidrati a DP 3. Anche con i GOS come sola fonte di carbonio, la produzione di etanolo è quasi trascurabile; al termine della fermentazione si accumulano acido lattico e acido acetico, rispettivamente alle concentrazioni di 246 e 280 mM di atomi di carbonio.
Per quanto riguarda la produzione della β-Gal, l’andamento dell’attività

110
specifica determinata mostra che a livello della superficie cellulare rimane pressoché ma cmq espressa a livelli basali che non superano le 1.7 U/mgDW. Nel citoplasma invece è riscontrabile un continuo aumento nella produzione di tale idrolasi parallelamente all’aumento della concentrazione della biomassa con un’attività specifica massima che raggiunge le 8 U/mgDW .
3. MODELLAZIONE MATEMATICA
3.1. Modellazione delle fermentazioni batch di B. adolescentis MB 239
Fermentazione del glucosio:
Sono riportati graficamente gli andamenti della crescita della biomassa, del consumo del substrato e dell’accumulo di prodotti elaborati dall’algoritmo di calcolo per la fermentazione del glucosio.
Inizialmente sono stati ottimizzati i valori di µMAX, KS e YX/S. I valori ottenuti sono stati forniti come stime iniziali per un secondo ciclo di calcolo al quale sono stati aggiunti i parametri Kd e m. Successivamente sono stati inseriti nel modello i parametri YP/X, che rappresenta la resa del totale dei prodotti di fermentazione (la concentrazione totale dei prodotti è stata calcolata sommando le concentrazioni dei singoli prodotti espresse in molarità di atomi di carbonio), e infine YL/X, YA/X, YE/X che sono rispettivamente le rese dell’acido lattico, dell’acido acetico e dell’etanolo.
Insieme alle stime dei parametri, per ogni processo è fornito il valore della radice degli scarti quadratici tra valori sperimentali e valori calcolati (SS), che rappresenta l’errore del processo di ottimizzazione.
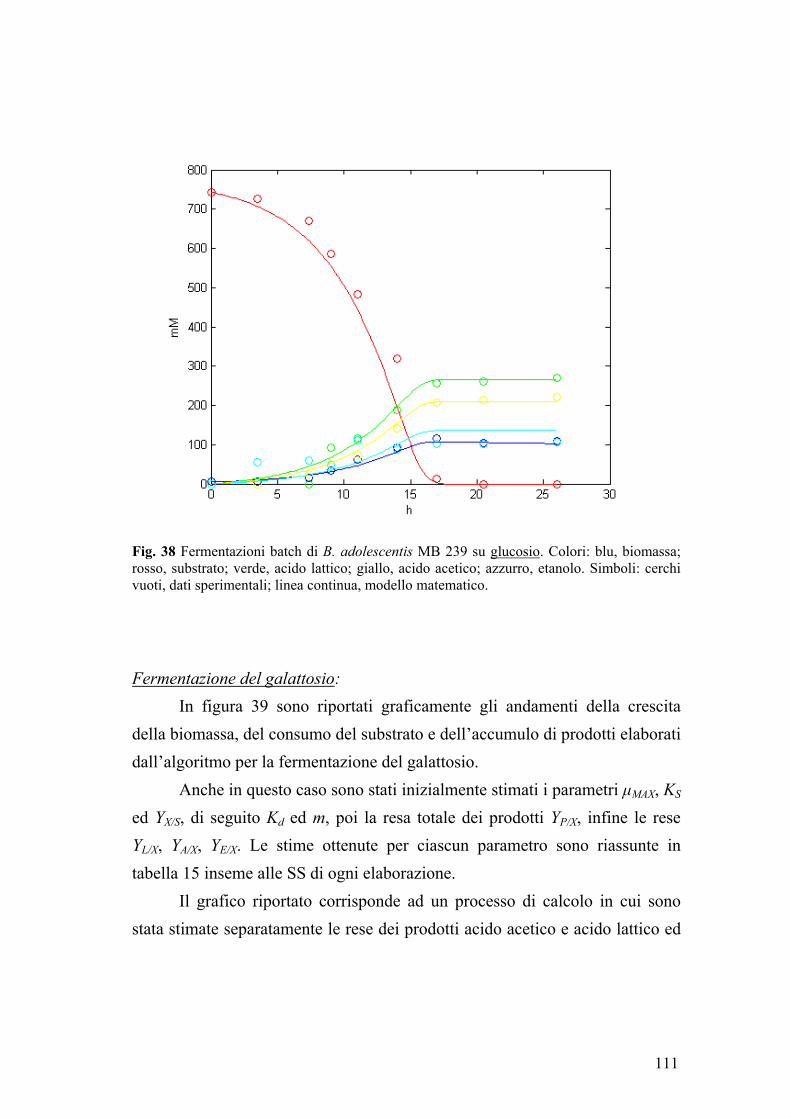
111
Fig. 38 Fermentazioni batch di B. adolescentis MB 239 su glucosio. Colori: blu, biomassa; rosso, substrato; verde, acido lattico; giallo, acido acetico; azzurro, etanolo. Simboli: cerchi vuoti, dati sperimentali; linea continua, modello matematico.
Fermentazione del galattosio:
In figura 39 sono riportati graficamente gli andamenti della crescita della biomassa, del consumo del substrato e dell’accumulo di prodotti elaborati dall’algoritmo per la fermentazione del galattosio.
Anche in questo caso sono stati inizialmente stimati i parametri µMAX, KS ed YX/S, di seguito Kd ed m, poi la resa totale dei prodotti YP/X, infine le rese YL/X, YA/X, YE/X. Le stime ottenute per ciascun parametro sono riassunte in tabella 15 inseme alle SS di ogni elaborazione.
Il grafico riportato corrisponde ad un processo di calcolo in cui sono stata stimate separatamente le rese dei prodotti acido acetico e acido lattico ed
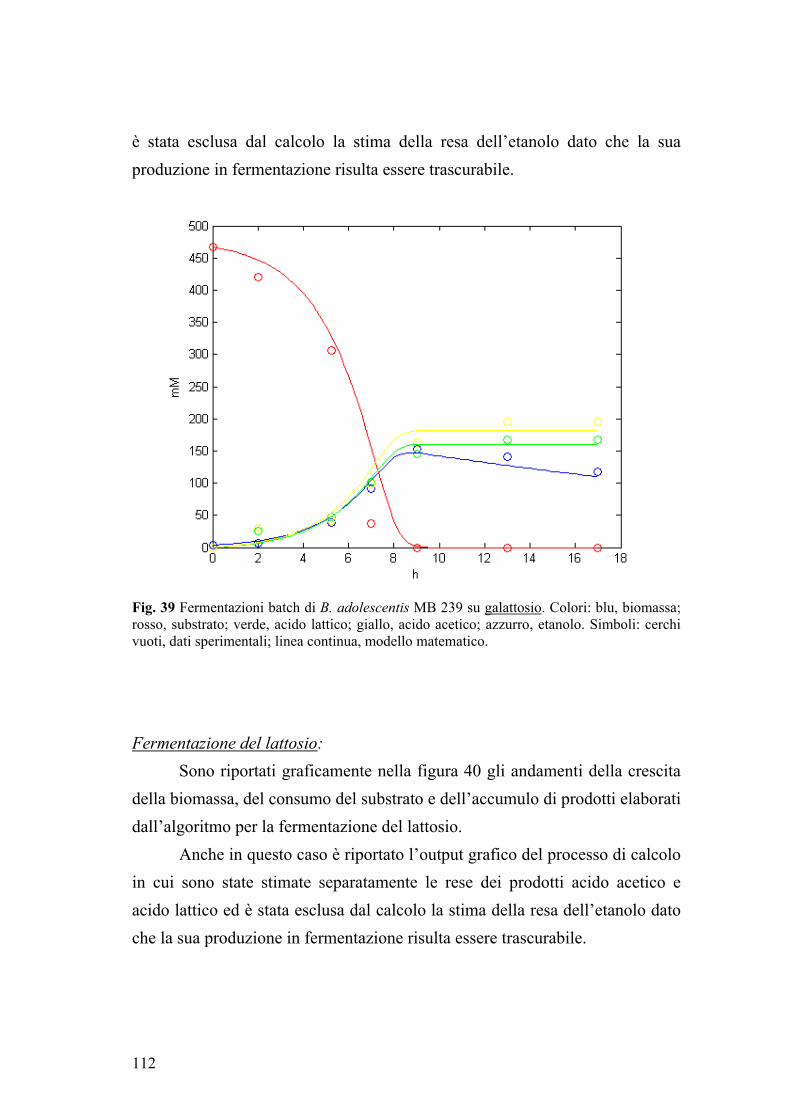
112
è stata esclusa dal calcolo la stima della resa dell’etanolo dato che la sua produzione in fermentazione risulta essere trascurabile.
Fig. 39 Fermentazioni batch di B. adolescentis MB 239 su galattosio. Colori: blu, biomassa; rosso, substrato; verde, acido lattico; giallo, acido acetico; azzurro, etanolo. Simboli: cerchi vuoti, dati sperimentali; linea continua, modello matematico. Fermentazione del lattosio:
Sono riportati graficamente nella figura 40 gli andamenti della crescita della biomassa, del consumo del substrato e dell’accumulo di prodotti elaborati dall’algoritmo per la fermentazione del lattosio.
Anche in questo caso è riportato l’output grafico del processo di calcolo in cui sono state stimate separatamente le rese dei prodotti acido acetico e acido lattico ed è stata esclusa dal calcolo la stima della resa dell’etanolo dato che la sua produzione in fermentazione risulta essere trascurabile.

113
Fig. 40 Fermentazioni batch di B. adolescentis MB 239 su lattosio. Colori: blu, biomassa; rosso, substrato; verde, acido lattico; giallo, acido acetico; azzurro, etanolo. Simboli: cerchi vuoti, dati sperimentali; linea continua, modello matematico.
Fermentazione dei GOS: Sono riportati graficamente nella figura 41 gli andamenti della crescita
della biomassa, del consumo del substrato e dell’accumulo di prodotti elaborati dall’algoritmo per la fermentazione dei GOS.

114
Fig. 41 Fermentazioni batch di B. adolescentis MB 239 su GOS. Colori: blu, biomassa; rosso, substrato; verde, acido lattico; giallo, acido acetico; azzurro, etanolo. Simboli: cerchi vuoti, dati sperimentali; linea continua, modello matematico.
3.2. Confronto tra i parametri fermentativi su glucosio, galattosio, lattosio e GOS.
In tabella 14 sono confrontate le stime dei parametri µMAX, KS ed YX/S ottenuti per tutte le fermentazioni. La µMAX e la YX/S maggiori si ottengono nella fermentazione del galattosio, su glucosio, al contrario si ottengono quelle minori. Lattosio e GOS presentano valori intermedi ai due costituenti monosaccaridici. I valori delle KS sui vari substrati sono tutti compresi tra 32.0 e 112.2 mM. Nei primi processi di elaborazione delle serie di dati del galattosio la stima fornita per la KS convergeva su un valore più elevato, che tuttavia non ottimizza gli andamenti dei singoli prodotti di fermentazione.

115
µMAX KS YX/S h-1 mM (C)
Glucosio 0.22 52.4 0.16 Galattosio 0.51 32.0 0.32 Lattosio 0.32 38.4 0.22 GOS 0.41 112.2 0.18
Tab. 14 Confronto tra le µMAX, KS, YX/S, calcolate dall’algoritmo per ogni substrato.
Sono riportate in grafico (fig. 42, 43, 44, 45) le porzioni del flusso di
carbonio, rappresentato dal substrato di crescita, che vengono trasformate dal ceppo MB 239 di B. adolescentis nei prodotti del suo metabolismo (acido lattico, acido acetico ed etanolo). Su tutte le fonti di carbonio si può notare che la percentuale di flusso di C che convoglia sull’acido lattico oscilla sempre il 43 e il 47% la mentre il resto è rappresentato dalla somma dei prodotti acido acetico ed etanolo. Si nota che la resa della produzione di etanolo è molto variabile a seconda della fonte di carbonio: risulta nulla su galattosio e lattosio, solo del 5.6% su i GOS e del 22.3% su glucosio.
Glucosio
Etanolo22.3%
Acido acetico34.3%
Acido lattico43.4%
Fig. 42 Percentuali relative alle porzioni di flusso di carbonio nei prodotti acido lattico, acido
acetico, etanolo su glucosio come unica fonte di carbonio.

116
Galattosio
Etanolo0%
Acido acetico53.1%
Acido lattico46.9%
Fig. 43 Percentuali relative alle porzioni di flusso di carbonio nei prodotti acido lattico, acido acetico, etanolo su galattosio come unica fonte di carbonio.
Lattosio
Acido lattico45.3%
Acido acetico54.7%
Etanolo0%
Fig. 44 Percentuali relative alle porzioni di flusso di carbonio nei prodotti acido lattico, acido acetico, etanolo su lattosio come unica fonte di carbonio.

117
GOS
Acido lattico44.7%
Acido acetico49.8%
Etanolo5.6%
Fig. 45 Percentuali relative alle porzioni di flusso di carbonio nei prodotti acido lattico, acido acetico, etanolo su GOS come unica fonte di carbonio.
4. FERMENTAZIONE DI XILOSIO E XOS
Le fermentazioni di B. adolescentis MB 239 sono state condotte in terreno SM a pH 5.5 addizionato di xilosio o xilo-oligosaccaridi come sola fonte di carbonio. In figura 46 è riportato l’andamento della fermentazione su xilosio, mentre in figura 50 quello della fermentazione condotta su XOS.
Nella fermentazione dello xilosio il ceppo mostra una fase esponenziale caratterizzata da una velocità specifica di crescita di µ = 0.26 h-1 e della durata di 15 ore circa al termine della quale si ottiene la produzione massima in biomassa che è uguale a 3.1 g L-1.
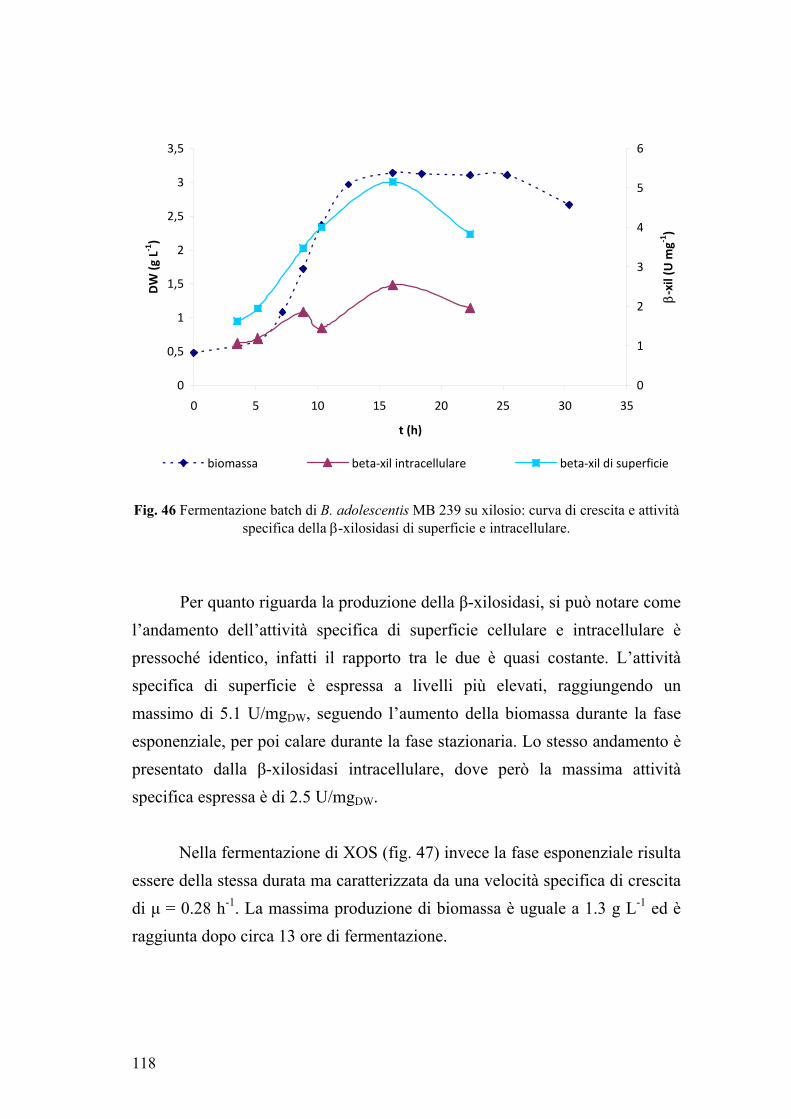
118
0
0,5
1
1,5
2
2,5
3
3,5
0 5 10 15 20 25 30 35
t (h)
DW (g
L‐1)
0
1
2
3
4
5
6
β‐xil (U m
g‐1 )
biomassa beta‐xil intracellulare beta‐xil di superficie
Fig. 46 Fermentazione batch di B. adolescentis MB 239 su xilosio: curva di crescita e attività specifica della β-xilosidasi di superficie e intracellulare.
Per quanto riguarda la produzione della β-xilosidasi, si può notare come
l’andamento dell’attività specifica di superficie cellulare e intracellulare è pressoché identico, infatti il rapporto tra le due è quasi costante. L’attività specifica di superficie è espressa a livelli più elevati, raggiungendo un massimo di 5.1 U/mgDW, seguendo l’aumento della biomassa durante la fase esponenziale, per poi calare durante la fase stazionaria. Lo stesso andamento è presentato dalla β-xilosidasi intracellulare, dove però la massima attività specifica espressa è di 2.5 U/mgDW.
Nella fermentazione di XOS (fig. 47) invece la fase esponenziale risulta
essere della stessa durata ma caratterizzata da una velocità specifica di crescita di µ = 0.28 h-1. La massima produzione di biomassa è uguale a 1.3 g L-1 ed è raggiunta dopo circa 13 ore di fermentazione.

119
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0,0 5,0 10,0 15,0 20,0 25,0
t (h)
DW (g
L‐1)
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
β‐xil (U m
g‐1 )
biomassa beta‐xil di superficie beta‐xil intracellulare
Fig. 47 Fermentazione batch di B. adolescentis MB 239 su XOS: curva di crescita e attività
specifica della β-xilosidasi di superficie e intracellulare.
L’andamento della β-xilosidasi risulta essere quasi speculare a quello della fermentazione su xilosio, infatti anche in questo caso gli andamenti dell’attività specifica di superficie e intracellulare sono analoghi; la β-xilosidasi non viene prodotta durante la fase esponenziale e quindi diminuisce all’aumentare della biomassa, mentre aumenta durante la fase stazionaria.

120
5. PURIFICAZIONE DI UNA β-XILOSIDASI
EXTRACELLULARE DI B. adolescentis MB 239
Il surnatante della coltura di B. adolescentis MB 239 sviluppato in
bioreattore su xilo-oligosaccaridi è stato raccolto in tarda fase esponenziale e ne è stata determinata l’attività β-xilosidasica mediante dosaggio spettrofotometrico. In tab. 15 sono presentati tutti gli step necessari per isolare e purificare la proteina.
Step di
purificazione
Attività specifica
(U/mgprot)
Fattore di
purificazione
Dialisi 5 1
Concentrazione per ultrafiltrazione (cut off 3 kDa) 9 1.8
Interazione ionica Mono Q sepharose (A) 147 29.4
Gel filtrazione (B) 596 119
Tab. 15 Protocollo di purificazione utilizzato per isolare e purificare l’enzima extracellulare β-xilosidasi di B. adolescentis MB 239.
Inizialmente il surnatante è stato filtrato per eliminare residui cellulari,
dializzato e concentrato mediante ultrafiltrazione. Dopo questo primo stadio, il fattore di purificazione della proteina è pari a 1.8, con un’attività specifica che passa da 5 a 9 U/mgprot. (v. Tab. 19)
Lo step successivo, consiste un una cromatografia a scambio ionico che ha permesso di isolare proteine con simili pI; l’attività β-xilosidasica (e quindi l’enzima) è stata individuata mediante dosaggio spettrofotometrico. L’andamento della purificazione è stato seguito anche mediante SDS-page

121
controllando qualitativamente il fattore di purificazione (fig. 48). È stata riscontrata alla fine di questo step un’attività specifica di 147 U/mgprot, con un fattore di purificazione di 29,4 rispetto al surnatante raccolto in fermentazione (v. Tab. 19). Tale fattore di purificazione è aumentato fino a 119 mediante un secondo step cromatografico. In questa fase la proteina purificata presenta un’attività specifica di 596 U/mgprot. Osservando l’SDS-page presentato in fig. 51 si nota come, dopo il secondo step di purificazione, è stato possibile isolare la proteina costituita da un monomero di circa 40 kDa.
Fig. 48 SDS-page del campione eluito in colonna cromatografia a scambio ionico (A) e ad
esclusione molecolare (B).
5.1. Determinazione del peso molecolare e dei parametri cinetici enzimatici Dopo aver isolato e purificato la proteina responsabile dell’attività β-
xilosidasica e aver identificato una singola unità monometrica di 40 kDa, è stata studiata la proteina nella sua forma nativa. Il peso molecolare della proteina nativa è stato determinato mediante gel filtrazione previa taratura con
A B

122
proteine standard. E’ stato definito che il peso molecolare della β-xilosidasi purificata è circa 160 kDa, quindi la proteina è un omotetramero.
La conferma dell’attività enzimatica posseduta dalla proteina isolata è stata ottenuta mediante zimogramma, con la formazione di una banda fluorescente (dovuta al 4-methylumbelliferone, ottenuto per idrolisi dal 4-
methylumbelliferyl-7-β-D-xylopyranoside) in corrispondenza del campione caricato.
I parametri cinetici enzimatici (km e Vmax), determinati mediante dosaggio spettrofotometrico di miscele di reazione in presenza di concentrazioni crescenti di substrato sintetico, sono stati calcolati secondo il diagramma di Lineweaver-Burk e hanno fornito i seguenti valori:
km = 4.36 mM Vmax = 0.93 mM/min.
5.2. Attività enzimatica in funzione di pH e temperatura
La β-xilosidasi isolata è stata caratterizzata dal punto di vista dell’attività e della stabilità in funzione del pH e della temperatura. Sono state condotte reazioni enzimatiche a differenti temperature e a differenti valori di pH ed è stata calcolata l’attività residua, mostrata in fig. 49 e fig. 50. Nelle condizioni di reazione la massima attività è presente per una temperatura di 53°C e mentre l’attività residua diminuisce fino a valori inferiori al 20 % per T > 60°C o < 35°C.
Posta nelle ottimali condizioni di temperatura, la massima attività della
β-xilosidasi è presente per un pH di 6.0, per poi diminuire leggermente a pH 6.5 e 5.5. L’attività residua cala drasticamente per valori di pH inferiori a 5 e superiori a 7.

123
Fig. 49 Attività residua della β-xilosidasi extracellulare di B. adolescentis MB 239 a differenti temperature di reazione.
Fig. 50 Attività residua della β-xilosidasi extracellulare di B. adolescentis MB 239 a
differenti pH di reazione.

124
L’enzima purificato può essere considerato termostabile in quanto mantiene circa il 100% di attività residua dopo essere stato incubato a 53°C per 2 h, per poi calare all’82 % dopo 6 ore e mantenere comunque un’attività residua del 60% circa dopo oltre 24 ore di incubazione (fig. 51).
Fig. 51 Attività residua della β-xilosidasi extracellulare di B. adolescentis MB 239 mantenuta a 53°C per 24 h.
Per quanto riguarda la stabilità in funzione del pH, l’enzima è stato mantenuto a 4°C in un sistema tampone con differenti valori di pH, compresi tra 3 e 9, quindi è stata determinata l’attività residua relativa al pH ottimale (pH 6). In fig. 52 è possibile osservare come per la permanenza a pH 6 l’attività sia stabile per più di 24 h, a pH 5 l’enzima mantenga circa il 90% di attività relativa mentre già dopo 3 h di mantenimento a pH 3 l’attività residua diminuisce drasticamente. L’enzima conservato a pH 7 e 9 subisce un drastico
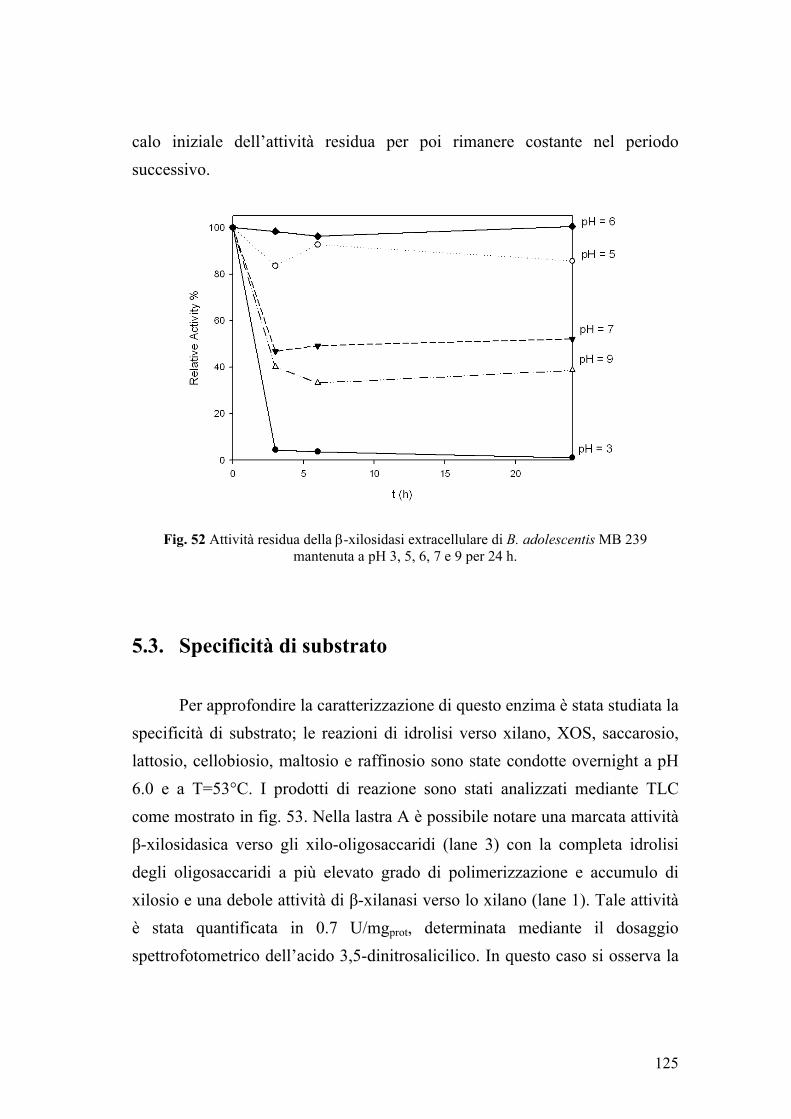
125
calo iniziale dell’attività residua per poi rimanere costante nel periodo successivo.
Fig. 52 Attività residua della β-xilosidasi extracellulare di B. adolescentis MB 239 mantenuta a pH 3, 5, 6, 7 e 9 per 24 h.
5.3. Specificità di substrato Per approfondire la caratterizzazione di questo enzima è stata studiata la specificità di substrato; le reazioni di idrolisi verso xilano, XOS, saccarosio, lattosio, cellobiosio, maltosio e raffinosio sono state condotte overnight a pH 6.0 e a T=53°C. I prodotti di reazione sono stati analizzati mediante TLC come mostrato in fig. 53. Nella lastra A è possibile notare una marcata attività β-xilosidasica verso gli xilo-oligosaccaridi (lane 3) con la completa idrolisi degli oligosaccaridi a più elevato grado di polimerizzazione e accumulo di xilosio e una debole attività di β-xilanasi verso lo xilano (lane 1). Tale attività è stata quantificata in 0.7 U/mgprot, determinata mediante il dosaggio spettrofotometrico dell’acido 3,5-dinitrosalicilico. In questo caso si osserva la
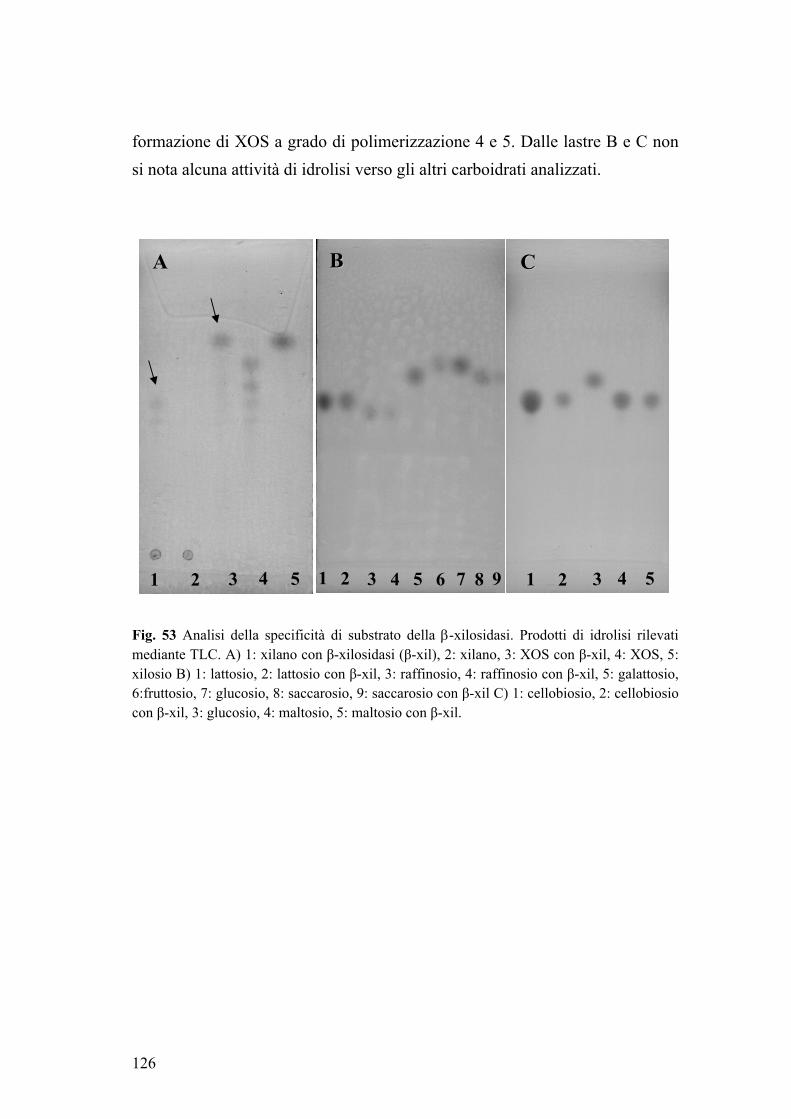
126
formazione di XOS a grado di polimerizzazione 4 e 5. Dalle lastre B e C non si nota alcuna attività di idrolisi verso gli altri carboidrati analizzati.
Fig. 53 Analisi della specificità di substrato della β-xilosidasi. Prodotti di idrolisi rilevati mediante TLC. A) 1: xilano con β-xilosidasi (β-xil), 2: xilano, 3: XOS con β-xil, 4: XOS, 5: xilosio B) 1: lattosio, 2: lattosio con β-xil, 3: raffinosio, 4: raffinosio con β-xil, 5: galattosio, 6:fruttosio, 7: glucosio, 8: saccarosio, 9: saccarosio con β-xil C) 1: cellobiosio, 2: cellobiosio con β-xil, 3: glucosio, 4: maltosio, 5: maltosio con β-xil.
BB
2 4 6 8 3 5 7 9 1
AA
1 2 3 4 5 1 2 3 4 5
CC

127
RISULTATI
Parte II
1. SELEZIONE DEI MICRORGANISMI
Obiettivo di questo progetto di tesi è stato lo sviluppo di una miscela di ceppi probiotici innovativi da veicolare in prodotti da forno shelf-stable. La selezione di ceppi inediti, appartenenti ai generi Bifidobacterium, Lactobacillus e alla specie Streptococcus thermophilus, è stata svolta in uno screening pregresso, nel quale sono state studiate proprietà peculiari di alcuni microrganismi quali l’attività antiossidante e la produzione di glutatione; tali caratteristiche biochimiche e fisiologiche innovative conferiscono un ulteriore vantaggio per la salute dell’ospite, oltre a quelle intrinseche derivanti dall’appartenenza al genere microbico probiotico.
Lactobacillus plantarum BAR 10, Streptococcus thermophilus BAR 20 e Bifidobacterium lactis BAR 30, appartenenti alla collezione dell’azienda Barilla G&R f.lli SpA, sono stati selezionati in quanto ceppi con elevato potere antiossidante e in grado di produrre elevate quantità di glutatione.

128
2. DETERMINAZIONE IN VITRO DELLA TOLLERANZA AI SUCCHI GASTRICI E ALLA BILE
La resistenza agli acidi gastrici e biliari è un pre-requisito fondamentale
che un ceppo probiotico deve possedere, poiché assicura che il microrganismo raggiunga vitale l’intestino, dove svolgerà l’azione benefica. Questa caratteristica biochimica è stata la prima ad essere indagata come marker fondamentale per poter considerare, i microrganismi selezionati, potenziali ceppi probiotici da veicolare in prodotti da forno.
I 3 microrganismi sono stati singolarmente messi a contatto con succo gastrico umano, succo gastrico simulato o bile suina e incubati a 37°C. Nel corso dell’incubazione è stata valutata la cinetica di decadimento della vitalità (facendo dei prelievi rispettivamente dopo 0, 30, 60 e 90 minuti): questa è stata osservata mediante conta vitale su piastra in terreno MRS o M17 in funzione del ceppo (tab. 16).
Tutti i ceppi presentano una completa perdita di vitalità dopo 90 minuti in succo gastrico umano. E’ stata osservata una significativa perdita di vitalità (P < 0.05) nel modello gastrico simulato, nonostante le conte vitali siano sempre più elevate di quelle in succo gastrico umano.
La sopravvivenza di Streptococcus thermophilus BAR 20 in succo gastrico umano diminuisce di ben 6 ordini di grandezza già nei primi 30 minuti con valori inferiori a 2 log10 CFU ml-1. Lo stesso ceppo presenta una vitalità più elevata in succo gastrico simulato dopo 30 minuti, vitalità che però diminuisce fino a 2.51 log10 CFU ml-1 dopo 90 minuti.
Migliore il comportamento di Lactobacillus plantarum BAR 10, le cui conte vitali in succo gastrico umano diminuscono fino al valore di 4.48 log10 CFU ml-1 nei primi 30 minuti. In succo gastrico simulato, la diminuzione di

129
vitalità è sensibilmente inferiore dal momento che L. plantarum presenta valori di 8.26 log10 CFU ml-1 dopo 30 minuti.
In succo gastrico umano B. lactis BAR 30 perde vitalità nei primi 30 minuti passando da valori di 7.78 a valori di 3.70 log10 CFU ml-1 nei primi 30 minuti, per poi dimnuire progressivamente fino a valori inferiori a 2.00 log10 CFU ml-1 dopo 90 minuti di incubazione.
Ceppo
Tempo di
incubazione
(min)
Succo gastrico
umano*
(log10cfu/ml)
Succo gastrico
simulato*
(log10cfu/ml)
Bile *
(log10cfu/ml)
L. plantarum BAR10 0 7.95a 8.62a 8.72a
30 4.48b 8.26a 8.72a
60 3.00c 4.70 b 8.60a
90 < 2d 3.11c 8.60a
S. thermophilus BAR 20 0 6.48a 6.95a 6.95a
30 < 2b 6.60a 6.95a
60 < 2b 3.70b 6.95a
90 < 2b 2.51c 6.52a
B. lactis BAR 30 0 7.78a 8.04a 8.26a
30 3.70b 6.78b 8.26a
60 2.00c 6.68b 8.18a
90 < 2d 6.65b 8.00a
Tab. 16 Conta vitale in piastra di L. plantarum BAR 10, S. thermophilus
BAR20 e B. lactis BAR 30 in succo gastrico umano, succo gastrico simulato o bile suina (*Valore medio, n = 3, SD < 0.4. All’interno dello stesso gruppo microbico e della stessa colonna, i valori che differiscono tra loro possiedono lettere diverse, P < 0.05).

130
Al contrario di ciò che succede con i succhi gastrici, la vitalità di L. plantarum, S. thermophilus e B. lactis non è influenzata dal contatto con la bile suina in questo modello in vitro, dato che le conte vitali in piastra dei 3 ceppi risultano essere pressochè identiche, dopo 90 minuti di incubazione, a quelle del tempo 0 (P < 0.05).
3. CINETICHE DI FERMENTAZIONE DI L. plantarum, S. thermophilus E B. lactis SU CARBOIDRATI PREBIOTICI
Al fine di sviluppare il simbiotico migliore, sono stati selezionati 13
carboidrati, suddivisibili in fibre solubili e fibre insolubili, che sono stati testati come fonte di carbonio per i 3 ceppi in fermentazioni batch a pH non controllato. Sono stati messi a punto terreni di coltura in cui ogni singolo carboidrato è stato addizionato come sola fonte di carbonio, al fine di valutare lo sviluppo di ogni microrganismo e definirne quindi l’affinità per ogni prebiotico.
Lo sviluppo a carico delle fibre solubili è stato monitorato seguendo l’aumento di torbidità (OD600), mentre per quelle insolubili la crescità è stata valutata mediante conta vitale in piastra. Le cinetiche di crescita sui singoli carboidrati sono state confrontate con lo sviluppo in terreno non addizionato di alcuna fonte di carbonio (negativo), in modo da poter essere certi che lo sviluppo del ceppo fosse dovuto solo a carico della frazione glucidica aggiunta.
Analizzando le cinetiche di crescita quasi tutti i prodotti esaminati hanno permesso il raggiungimento del massimo sviluppo dopo 5-8 ore di incubazione per poi mantenersi su valori costanti.
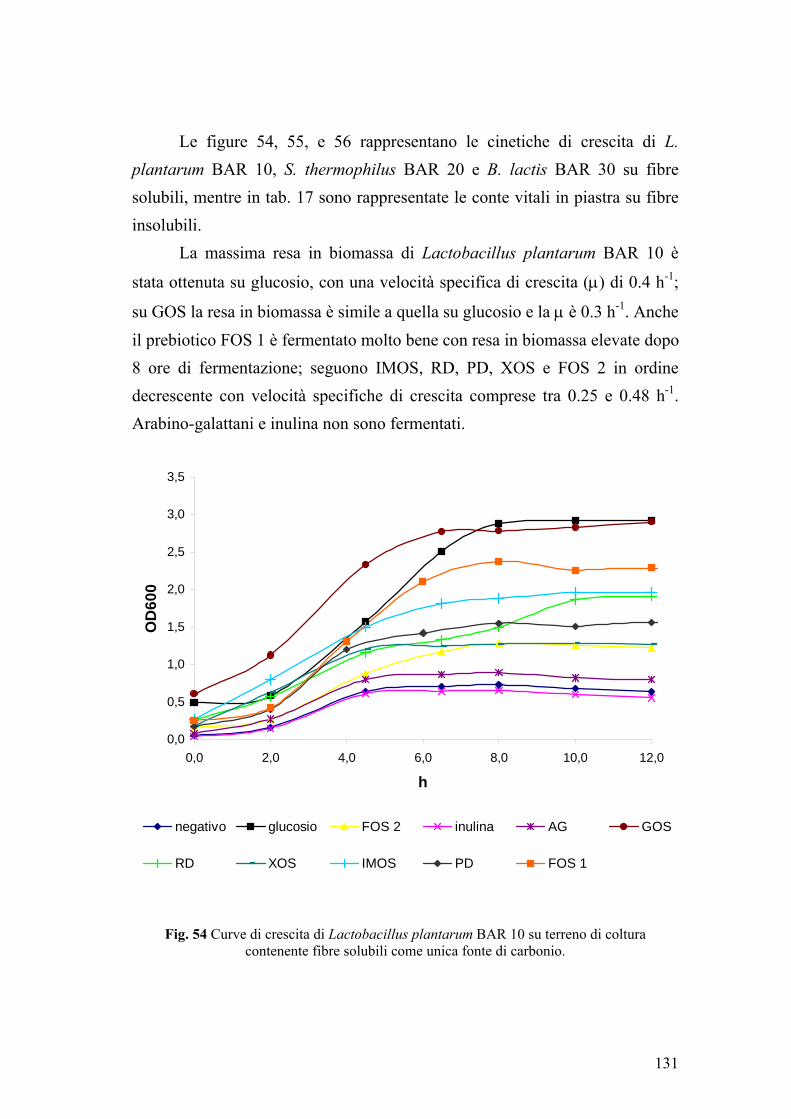
131
Le figure 54, 55, e 56 rappresentano le cinetiche di crescita di L. plantarum BAR 10, S. thermophilus BAR 20 e B. lactis BAR 30 su fibre solubili, mentre in tab. 17 sono rappresentate le conte vitali in piastra su fibre insolubili.
La massima resa in biomassa di Lactobacillus plantarum BAR 10 è
stata ottenuta su glucosio, con una velocità specifica di crescita (µ) di 0.4 h-1;
su GOS la resa in biomassa è simile a quella su glucosio e la µ è 0.3 h-1. Anche il prebiotico FOS 1 è fermentato molto bene con resa in biomassa elevate dopo 8 ore di fermentazione; seguono IMOS, RD, PD, XOS e FOS 2 in ordine decrescente con velocità specifiche di crescita comprese tra 0.25 e 0.48 h-1. Arabino-galattani e inulina non sono fermentati.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
0,0 2,0 4,0 6,0 8,0 10,0 12,0
h
OD
600
negativo glucosio FOS 2 inulina AG GOS
RD XOS IMOS PD FOS 1
Fig. 54 Curve di crescita di Lactobacillus plantarum BAR 10 su terreno di coltura contenente fibre solubili come unica fonte di carbonio.

132
Tra i prebiotici insolubili L. plantarum è in grado di utilizzare solo l’amido resistente RS 2, con rese finali simili a quelle del glucosio (9.04 vs 9.30 log10 CFU ml-1).
Il ceppo Streptococcus thermophilus BAR 20 ha dimostrato una differenziazione molto evidente nell’utilizzo dei vari prebiotici solubili, come è evidente dalla fig. 55. Questo ceppo raggiunge la più elevata resa in biomassa su glucosio (P < 0.05) anche se cresce molto bene su GOS, FOS 1 e FOS 2. Le velocità specifiche di crescita su glucosio, GOS, FOS 1 e FOS 2 sono 0.33, 0.26, 0.35 e 0.43 h-1, rispettivamente.
Infine tra le fibre insolubili Streptococcus thermophilus BAR 20 non metabolizza alcun carboidrato, come è possibile vedere in tab. 17.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
0,0 2,0 4,0 6,0 8,0 10,0 12,0
h
OD
600
negativo glucosio FOS 2 inulina AG GOS
RD XOS IMOS PD FOS 1
Fig. 55 Curve di crescita di Streptococcus thermophilus BAR 20 su terreno di coltura contenente fibre solubili come unica fonte di carbonio.

133
In fig. 56 sono descritte le curve di crescita di Bifidobacterium lactis BAR 30 su fibre solubili. Il ceppo cresce su glucosio, FOS 1 e GOS con simili velocità specifiche di crescita (0.34 h-1) e rese in biomassa. Concentrazioni di biomassa più basse sono state ottenute su FOS 2, IMOS e infine su XOS con
OD di circa 1.5 e µ di 0.49, 0.45, e 0.21 h-1, rispettivamente. Inulina, poli-destrosio, arabino-galattani e destrina resistente non vengono fermentati.
Tra le fibre insolubili, l’amido resistente RS 2 supporta la crescita di B. lactis BAR 30 fino a 9.28 log10 CFU ml-1 dopo 24 h. Il ceppo utilizza debolmente anche destrano e guar gum PHGG che portano ad un aumento di 0.9 e 0.6 ordini di grandezza nella conta in piastra, rispettivamente.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
0,0 2,0 4,0 6,0 8,0 10,0 12,0
h
OD
600
negativo glucosio FOS 2 inulina AG GOS
RD XOS IMOS PD FOS 1
Fig. 56 Curve di crescita di Bifidobacterium lactis BAR 30 su terreno di coltura contenente fibre solubili come unica fonte di carbonio.

134
S. thermophilus BAR 20 e B. lactis BAR 30 non sono in grado di crescere nel terreno senza fonte di carbonio, mentre si può osservare un debole sviluppo per L. plantarum BAR 10.
Ceppo Tempo (h)
Controllo negativo Glucosio Destrano RS 1 RS 2 PHGG
L. plantarum BAR 10 0 7.30 7.30 7.30 7.30 7.30 7.30
8 7.23 8.30 7.76 7.53 8.15 7.60 24 7.45 9.30 7.80 7.57 9.00 7.70 34 7.60 9.30 7.54 7.51 9.04 7.57
S. thermophilus BAR 20 0 6.60 6.60 6.60 6.60 6.60 6.60
8 6.63 8.13 6.68 6.81 6.78 6.48 24 6.45 8.57 6.08 6.85 6.60 6.00 34 6.45 8.50 6.00 6.75 6.60 6.00
B. lactis BAR 30 0 6.68 6.68 6.68 6.68 6.68 6.68
8 6.56 7.45 7.58 6.85 7.41 7.30 24 6.48 8.30 7.08 6.18 9.28 7.20 34 6.48 8.30 7.00 6.10 9.25 7.20
Tab. 17 Conta vitale in piastra (log10cfu/ml) di L. plantarum BAR 10, S. thermophilus BAR 20, e B. lactis BAR 30 su fibre insolubili. Valori mediati, n = 3. SD sempre minore di 0.40.
4. VITALITA’ DEI CEPPI PROBIOTICI IN MATRICI ALIMENTARI
4.1. Sopravvivenza di B. lactis BAR 30
La prima sperimentazione è stata svolta ponendo a confronto la sopravvivenza del ceppo Bifidobacterium lactis BAR 30 inoculato in una

135
crema bianca anidra (“matrice Barilla”), fornita dalla ditta Barilla, e conservata in due differenti condizioni di temperatura (4°C e 25°C).
Tale matrice non subisce un trattamento termico ma solo meccanico.
Fig. 57 andamento di B. lactis BAR 30 in matrice Barilla conservata a 4°C e 25°C
Questa prova preliminare è stata svolta effettuando conteggi in tempi molto brevi per valutare il trend di sopravvivenza del ceppo inoculato nella matrice Barilla. Come si può notare dalla figura 57, il ceppo presenta un andamento di sopravvivenza molto differente in funzione della temperatura di conservazione; infatti B. lactis BAR 30 passa da 3 x 108 a 5 x 102 CFU g-1 in soli 40 giorni se conservato a temperatura ambiente.
Quindi sono stati scelti due carboidrati prebiotici, frutto-oligosaccaridi
e inulina a lunga catena, come potenziali carboidrati sostituivi della frazione glucidica nella matrice Barilla. Queste nuove matrici sono state formulate al fine di valutare se l’inulina o i frutto-oligosaccaridi possano svolgere un’azione di protezione verso la vitalità del probiotico liofilizzato, sostituendo la componente glucidica interamente con l’uno o l’altro dei prebiotici partendo
010
2030
4050
6070
80Ba 25°C
Ba 4°C
0123456789
log1
0 C
FU/g
t (giorni)

136
030
6090
120150
2
3
4
5
6
7
8
9
Barilla FOS inulina
t (giorni)
log 1
0 CFU
/g
dalla comune matrice Barilla. Le matrici “FOS” e “inulina” sono state inoculate con il ceppo liofilizzato B. lactis BAR 30 (1% w/w).
I campioni sono stati conservati in aerobiosi a temperatura ambiente e i conteggi sono stati effettuati ogni mese fino a raggiungere una durata complessiva di conservazione di 5 mesi.
Fig. 58 andamento di B. lactis BAR 30 in matrice Barilla, matrice FOS e matrice inulina conservate a 25°C.
Nella fig. 58 sono presentati i dati relativi ai conteggi effettuati nel
tempo per valutare la vitalità del liofilizzato B. lactis nelle 3 matrici a confronto. Il probiotico, inoculato all’1%, presenta un titolo iniziale di 5.5 x 108 CFU g-1, ma presenta un comportamento sostanzialmente diverso nelle tre matrici utilizzate. Nella matrice Barilla, dopo soli 20 giorni, B. lactis BAR 30 perde più di 5 unità logaritmiche. Nella matrice FOS invece la vitalità del ceppo inoculato è buona, passando da 5.5 x 108 a 1.5 x 106 CFU g-1 nei primi 2 mesi di conservazione e quindi perdendo solo 2 unità logaritmiche. Infine la matrice inulina sembra assicurare la migliore sopravvivenza del ceppo probiotico, il quale, dopo 150 giorni di conservazione a temperatura ambiente

137
0 2040
80120
180
2,0
3,0
4,0
5,0
6,0
7,0
8,0
9,0
Matrice Barilla + ac. ascorbico 0.05% Matrice InulinaMatrice Inulina + ac. ascorbico 0.05 %Matrice BarillaMatrice Barilla ANAEROBIOSI Matrice Barilla 4°C
log 1
0CFU
/g
t (giorni)
e in aerobiosi, rimane vitale con valori di 1.0 x 105 CFU g-1, con una caduta di soli 3 log nella conta vitale in piastra.
Ulteriori prove di sopravvivenza del ceppo Bifidobacterium lactis BAR 30 sono state svolte al fine di valutare la vitalità del liofilizzato in funzione della composizione della matrice e delle condizioni di conservazione dei campioni inoculati (temperatura e atmosfera).
Fig. 59 Conta vitale in piastra di B. lactis BAR 30 inoculato in 4 diverse matrici e conservate in diverse condizioni di temperatura e di atmosfera.

138
Nonostante il ceppo BAR 30 sia particolarmente aerotollerante, sono state progettate matrici contenenti acido ascorbico (0.05% w/w) come agente riducente, al fine di determinare la sopravvivenza del probiotico in matrici più riducenti. In questo stesso esperimento alcuni campioni sono stati preparati in condizioni anaerobiche, chiudendo i falcon entro una camera anaerobica. Il controllo è stato conservato 4°C. Il trend di sopravvivenza del ceppo inoculato in matrice è presentato in fig. 59.
Le due matrici Barilla e Inulina si comportano analogamente all’esperimento precedente, infatti nella matrice Barilla B. lactis perde vitalità arrivando a valori minori di 102 CFU g-1 dopo 120 giorni, mentre la matrice Inulina preserva il ceppo vitale per 180 giorni anche se con una perdità di 5 unità logaritmiche (passando da 1.5 x 109 CFU g-1del T0 a 2.0 x 104 CFU g-1
del T180). L’aggiunta di acido ascorbico ad entrambe le matrici non porta ad una variazione significativa della sopravvivenza del ceppo inoculato, ancora più evidente per la matrice Inulina in cui i conteggi in piastra sono del tutto analoghi alla matrice Inulina addizionata di acido ascorbico.
La matrice Barilla conservata in atmosfera anaerobica e a temperatura ambiente comporta un calo drastico della vitalità di B. lactis che, dopo 120 giorni, risulta essere inferiore a 102 CFU g-1. Anche in questa prova sperimentale, la matrice Barilla conservata a 4°C (controllo) permette un’ottima sopravvivenza del ceppo inoculato che perde all’incirca un’unità logaritmica, passando da 9.2 a 8.1 log10 CFU g-1 in 6 mesi.
Dato che l’addizione di inulina alla crema bianca anidra ha migliorato la sopravvivenza di Bifidobacterium lactis, sono state progettate 8 nuove matrici su base di crema bianca anidra contenenti, come ingredienti addizionali, alcuni carboidrati prebiotici (precedentemente descritti in tab. 7). Le 10 matrici (incluse anche la matrice Barilla e la matrice Inulina) sono state conservate a 4°C (come controllo), a temperatura ambiente in atmosfera aerobica e in atmosfera anaerobica.

139
In tabella 18 e in figura 60 sono presentati i dati relativi ai conteggi (tempi 0, 30, 70, 110, 150 e 180 giorni) in piastra del ceppo B. lactis BAR 30.
Fig 60 Andamento della vitalità di B. lactis BAR 30 nelle 10 matrici conservate a 25°C e in atmosfera aerobica.
Al T0gg i conteggi delle 10 matrici sono tutti confrontabili tra loro, dato
che il titolo del liofilizzato è circa 1.5 x 1011 CFU g-1 ed è stato inoculato all’1% w/w. Le matrici Barilla e Inulina sono state utilizzate come controllo rispetto alle prove precedenti.
Al conteggio T30gg si riscontra che le matrici Inulina, RG, PBP7 e Barilla hanno dato un ottimo risultato, infatti la concentrazione di BAR 30 nelle matrici è calata meno di un ordine di grandezza. Le matrici PBP3, PBP10 e PBP 11c hanno presentato, al contrario, un sensibile calo di concentrazione del liofilizzato inoculato. Al conteggio T70gg si riscontra una diminuzione consistente della concentrazione del liofilizzato in quasi tutte le matrici ad esclusione di PBP7 e RG.
2
3
4
5
6
7
8
9
10
0 30 60 90 120 150 180
t (giorni)
log1
0 C
FU/g
Barilla PBP1 PBP3 PBP4 PBP7
PBP10 PBP12 Inulina PBP11c RG
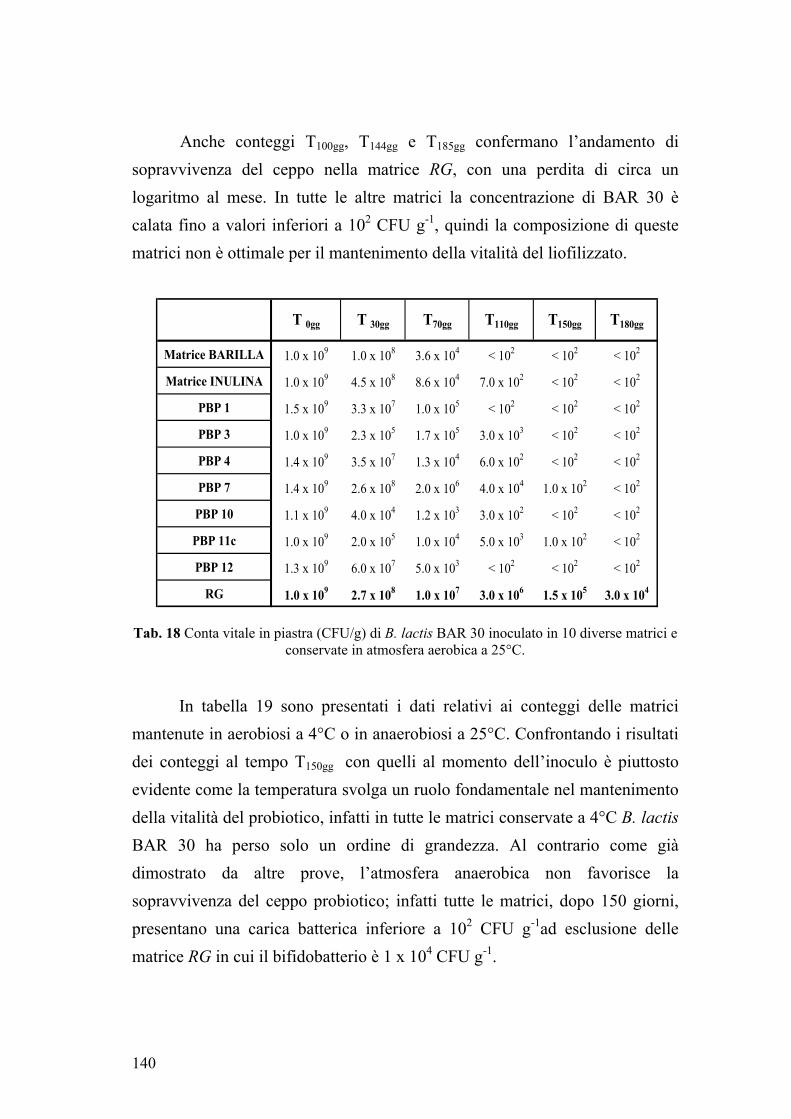
140
Anche conteggi T100gg, T144gg e T185gg confermano l’andamento di sopravvivenza del ceppo nella matrice RG, con una perdita di circa un logaritmo al mese. In tutte le altre matrici la concentrazione di BAR 30 è calata fino a valori inferiori a 102 CFU g-1, quindi la composizione di queste matrici non è ottimale per il mantenimento della vitalità del liofilizzato.
T 0gg T 30gg T70gg T110gg T150gg T180gg
Matrice BARILLA 1.0 x 109 1.0 x 108 3.6 x 104 < 102 < 102 < 102
Matrice INULINA 1.0 x 109 4.5 x 108 8.6 x 104 7.0 x 102 < 102 < 102
PBP 1 1.5 x 109 3.3 x 107 1.0 x 105 < 102 < 102 < 102
PBP 3 1.0 x 109 2.3 x 105 1.7 x 105 3.0 x 103 < 102 < 102
PBP 4 1.4 x 109 3.5 x 107 1.3 x 104 6.0 x 102 < 102 < 102
PBP 7 1.4 x 109 2.6 x 108 2.0 x 106 4.0 x 104 1.0 x 102 < 102
PBP 10 1.1 x 109 4.0 x 104 1.2 x 103 3.0 x 102 < 102 < 102
PBP 11c 1.0 x 109 2.0 x 105 1.0 x 104 5.0 x 103 1.0 x 102 < 102
PBP 12 1.3 x 109 6.0 x 107 5.0 x 103 < 102 < 102 < 102
RG 1.0 x 109 2.7 x 108 1.0 x 107 3.0 x 106 1.5 x 105 3.0 x 104
Tab. 18 Conta vitale in piastra (CFU/g) di B. lactis BAR 30 inoculato in 10 diverse matrici e conservate in atmosfera aerobica a 25°C.
In tabella 19 sono presentati i dati relativi ai conteggi delle matrici mantenute in aerobiosi a 4°C o in anaerobiosi a 25°C. Confrontando i risultati dei conteggi al tempo T150gg con quelli al momento dell’inoculo è piuttosto evidente come la temperatura svolga un ruolo fondamentale nel mantenimento della vitalità del probiotico, infatti in tutte le matrici conservate a 4°C B. lactis BAR 30 ha perso solo un ordine di grandezza. Al contrario come già dimostrato da altre prove, l’atmosfera anaerobica non favorisce la sopravvivenza del ceppo probiotico; infatti tutte le matrici, dopo 150 giorni, presentano una carica batterica inferiore a 102 CFU g-1ad esclusione delle matrice RG in cui il bifidobatterio è 1 x 104 CFU g-1.

141
T 0gg T 150gg
aerobiosi, 4°CT 150gg
anaerobiosi, 25°C
Matrice BARILLA 1.0 x 109 3.0 x 108 < 102
Matrice INULINA 1.0 x 109 3.0 x 108 < 102
PBP 1 1.5 x 109 7.0 x 108 < 102
PBP 3 1.0 x 109 6.0 x 108 < 102
PBP 4 1.4 x 109 8.5 x 108 < 102
PBP 7 1.4 x 109 5.5 x 108 < 102
PBP 10 1.1 x 109 6.0 x 108 < 102
PBP 11c 1.0 x 109 6.0 x 108 < 102
PBP 12 1.3 x 109 2.0 x 108 < 102
RG 1.0 x 109 6.5 x 108 1.0 x 104
Tab. 19 Conta vitale in piastra (CFU/g) di B. lactis BAR 30 inoculato in 10 diverse matrici e conservate in atmosfera anaerobica a 25°C o in atmosfera aerobica a 4°C.
Poiché la vitalità di B. lactis BAR 30 nella matrice RG conservata a 25°C è risultata promettente, al fine di migliorare ulteriormente la vitalità del ceppo sono state progettate 13 differenti matrici composte, all’80% da RG e al 20% da altri componenti quali inulina, FOS, amido, saccarosio, lattosio, trealosio, maltosio e maltodestrine. B. lactis BAR30 è stato inoculato anche in grasso alimentare 100% (matrice 1) al fine di valutare se la migliore sopravvivenza del ceppo possa ascriversi alla frazione lipidica e in matrice costituita al 100% dalla matrice RG (matrice 2).
In figura 61 è presentato l’andamento della vitalità di Bifidobacterium lactis monitorato dopo 45, 90 e 150 giorni. I risultati, dopo 150 giorni di conservazione a 25°C, hanno dimostrato che la sola frazione lipidica non fornisce un effetto di protezione nei confronti del probiotico presente in matrice, infatti in questa matrice il titolo del liofilizzato perde più di 7 unità logaritmiche nei primi 90 giorni, passando da 9.04 a 2.90 log10 CFU g-1. Al contrario, per quanto riguarda le restanti 12 matrici, non è possibile notare alcuna differenza significativa ascrivibile alle addizioni effettuate, se non

142
sottolineare un titolo del liofilizzato più elevato di circa 1 unità logaritmica nelle matrici RG addizionate di inulina, FOS, saccarosio e lattosio (valori nell’intorno di 5 log10 CFU g-1).
2
3
4
5
6
7
8
9
10
0 30 60 90 120 150
t (giorni)
log1
0 C
FU/g
Matrice 1 Matrice 2 Matrice 3 Matrice 4Matrice 5 Matrice 6 Matrice 7 Matrice 8Matrice 9 Matrice 9 Matrice 10 Matrice 11Matrice 12 Matrice 13
Fig. 61 Andamento della vitalità di B. lactis BAR 30 nelle 10 matrici conservate a 25°C e in
atmosfera aerobica.
4.2. Sopravvivenza di L. plantarum, S. thermophilus e B. lactis in matrice cioccolato
E’ stata inoltre studiata la vitalità dei ceppi probiotici in matrici di cioccolato che si presentano nella forma di barrette di cioccolato fondente senza altri ingredienti o addizionate dei carboidrati prebiotici verso i quali i microrganismi avevano presentano una migliore affinità, cioè galatto-

143
oligosaccaridi (GOS), frutto-oligosaccaridi Actilight (FOS 1) e frutto-oligosaccaridi a media catena FOS-Raftilose P95 (FOS 2). E’ stata quindi valutato se, anche nelle matrici cioccolato, la sopravvivenza dei liofilizzati variasse in funzione del carboidrato prebiotico addizionato. Il cioccolato è stato individuato come la matrice per la quale i probiotici presentano affinità ed un'ottima sopravvivenza, probabilmente per un effetto protettivo dovuta all'elevata concentrazione di grassi.
Tali prebiotici sono quindi stati singolarmente inseriti nella formulazione di barrette di cioccolato inoculate all’ 1% con una miscela dei 3 ceppi liofilizzati Lactobacillus plantarum BAR 10, Streptococcus thermophilus BAR 20 e Bifidobacterium lactis BAR 30. La sopravvivenza è risultata ottima fino a 6 mesi con conservazione dei campioni a temperatura ambiente. In particolare è possibile osservare l’andamento della vitalità di Lactobacillus plantarum BAR 10, Bifidobacterium lactis BAR 30 e Streptococccus thermophilus BAR 20 rispettivamente nelle figure 62, 63 e 64. L. plantarum BAR 10 è in grado di sopravvivere nella matrice cioccolato conservando praticamente inalterato il titolo del liofilizzato del conteggio al tempo zero e quindi con valori intorno a 108 CFU g-1; tuttavia non si nota nessuna differenza significativa tra le varie matrici, indice quindi che i carboidrati addizionati non giocano un ruolo fondamentale nella vitalità dei microrganismi in questa matrice. Come si può notare dalla fig. 63 il ceppo Bifidobacterium lactis BAR 30 presenta una buona sopravvivenza anche dopo 180 giorni di conservazione a temperatura ambiente, perdendo solo circa 2 ordini di grandezza rispetto al tempo zero ma anche in questo caso non si osservano significative differenze tra le varie barrette di cioccolato.

144
Fig. 62 Andamento della vitalità di L. plantarum BAR 10 nelle barrette di cioccolato puro (tq), o addizionato di FOS 1, FOS 2 o GOS.
Fig. 63 Andamento della vitalità di B. lactis BAR 30 nelle barrette di cioccolato puro (tq), o
addizionato di FOS 1, FOS 2 o GOS.
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
0 30 60 90 120 150 180
t (giorni)
CFU
/g
cioccolato tq cioccolato FOS 1 cioccolato GOS cioccolato FOS 2
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
0 30 60 90 120 150 180
t (giorni)
CFU
/g
cioccolata tq cioccolata FOS 1 cioccolata GOS cioccolata FOS 2

145
Streptococcus thermophilus BAR 20 è il ceppo che ha subito una maggiore perdita di vitalità passando da valori di circa 3 x 108 CFU g-1 a circa 5 x 105 CFU g-1 dopo 6 mesi, ma mantenendosi comunque su elevate concentrazioni di probiotico nella matrice cioccolato. Anche in questo caso le cinetiche di decadimento del microrganismo, presentate in fig. 64, non variano in modo significativo in funzione del diverso carboidrato prebiotico addizionato.
Fig. 64 Andamento della vitalità di S. thermophilus BAR 20 nelle barrette di cioccolato puro (tq), o addizionato di FOS 1, FOS 2 o GOS.
1,00E+05
1,00E+06
1,00E+07
1,00E+08
1,00E+09
0 30 60 90 120 150 180t (giorni)
CFU
/g
cioccolato tq cioccolato FOS 1 cioccolato GOS cioccolato FOS 2

146
5. TRIAL CLINICO
Dopo aver quantificato il contenuto fecale in bifidobatteri e lattobacilli dei campioni fecali raccolti dai 20 volontari sani, i soggetti partecipanti al trial clinico sono stati suddivisi rispettivamente in 3 gruppi in funzione del contenuto fecale di bifidobatteri o lattobacilli al T0 per meglio analizzare i risultati ottenuti.
BIFIDOBATTERI Gruppo bif-A 108-1010 CFU g-1 (14 soggetti) Gruppo bif-B 106-107 CFU g-1 (4 soggetti) Gruppo bif-C < 106 CFU g-1 (2 soggetti)
LATTOBACILLI Gruppo lat-A 106-107 CFU g-1 (6 soggetti) Gruppo lat-B 104-105 CFU g-1 (8 soggetti) Gruppo lat-C < 104 CFU g-1 (6 soggetti)
Come si può notare in tab. 20, nel gruppo bif-A non si notano significativi aumenti nel contenuto fecale di bifidobatteri durante il trial clinico per quasi tutti i 14 soggetti ad esclusione del soggetto 15, che ha presentato un aumento di 1 unità log al T30g. Nel gruppo bif-B, al contrario, si nota un comune aumento statisticamente significativo (P < 0.05) di circa 1 unità logaritmica nella concentrazione dei bifidobatteri e in particolare il soggetto 9 ha presentato un aumento di 2 unità log, aumento che perdura anche dopo 30 giorni di wash-out. Infine in entrambi i soggetti del gruppo bif-C si può notare un consistente aumento della concentrazione dei bifidobatteri al T30g rispetto al T0g, rispettivamente di quasi 4 (soggetto 12) o 3 (soggetto 18) unità logaritmiche (aumento statisticamente significativo, p< 0.05).
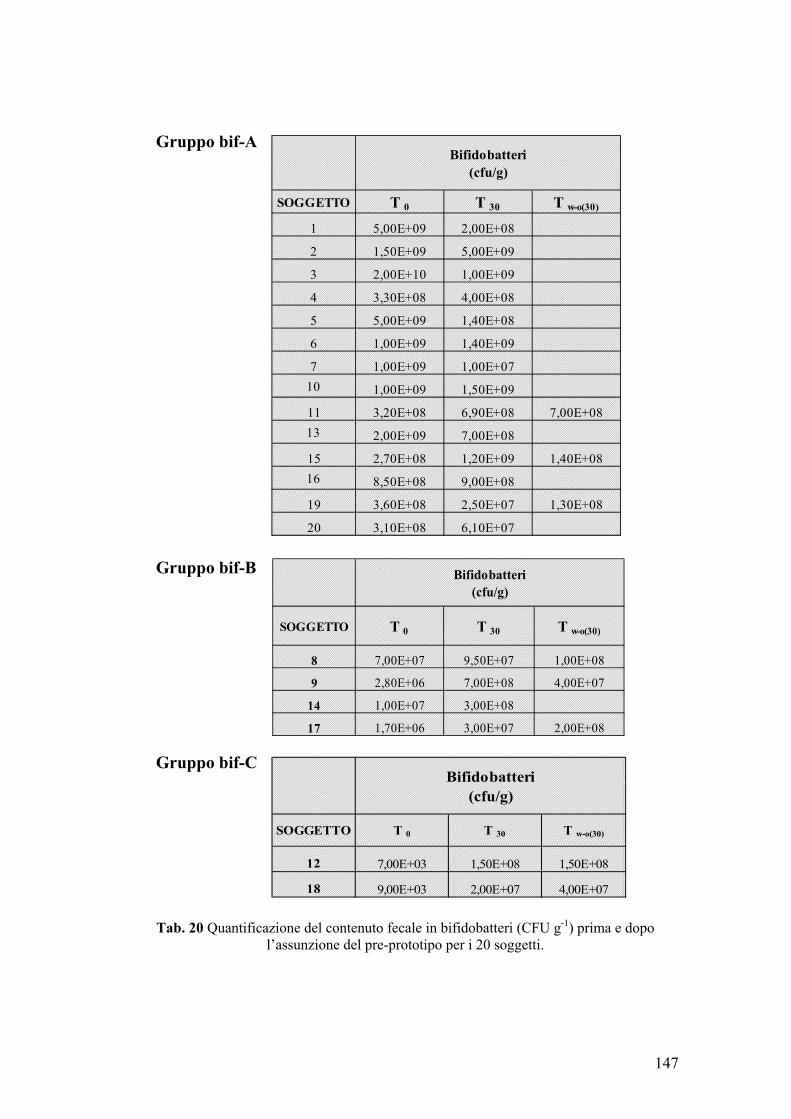
147
Gruppo bif-A
Gruppo bif-B
Gruppo bif-C
Tab. 20 Quantificazione del contenuto fecale in bifidobatteri (CFU g-1) prima e dopo l’assunzione del pre-prototipo per i 20 soggetti.
SOGGETTO T 0 T 30 T w-o(30)
1 5,00E+09 2,00E+08
2 1,50E+09 5,00E+09
3 2,00E+10 1,00E+09
4 3,30E+08 4,00E+08
5 5,00E+09 1,40E+08
6 1,00E+09 1,40E+09
7 1,00E+09 1,00E+0710 1,00E+09 1,50E+09
11 3,20E+08 6,90E+08 7,00E+0813 2,00E+09 7,00E+08
15 2,70E+08 1,20E+09 1,40E+0816 8,50E+08 9,00E+08
19 3,60E+08 2,50E+07 1,30E+08
20 3,10E+08 6,10E+07
Bifidobatteri (cfu/g)
SOGGETTO T 0 T 30 T w-o(30)
8 7,00E+07 9,50E+07 1,00E+08
9 2,80E+06 7,00E+08 4,00E+07
14 1,00E+07 3,00E+08
17 1,70E+06 3,00E+07 2,00E+08
Bifidobatteri (cfu/g)
SOGGETTO T 0 T 30 T w-o(30)
12 7,00E+03 1,50E+08 1,50E+08
18 9,00E+03 2,00E+07 4,00E+07
Bifidobatteri (cfu/g)

148
Gruppo lat-A Gruppo lat-B Gruppo lat-C
Tab. 21 Quantificazione del contenuto fecale in lattobacilli (CFU g-1) prima e dopo l’assunzione del pre-prototipo per i 20 soggetti.
SOGGETTO T 0 T 30 T w-o(30)
1 1,00E+07 5,00E+07
2 2,50E+07 2,00E+05
4 4,80E+07 2,00E+05
6 2,50E+06 9,00E+05
7 3,00E+06 1,00E+05
18 9,00E+07 1,20E+07 1,40E+07
Lattobacilli (cfu/g)
SOGGETTO T 0 T 30 T w-o(30)
3 1,00E+04 1,00E+04
5 4,00E+05 1,00E+05
9 3,00E+04 4,00E+04 7,00E+05
11 7,50E+05 1,00E+07 1,00E+06
14 2,00E+05 2,00E+06
15 6,90E+05 6,90E+03 5,00E+05
16 1,00E+05 2,00E+04
19 6,00E+04 1,40E+04 6,00E+06
Lattobacilli (cfu/g)
SOGGETTO T 0 T 30 T w-o(30)
8 5,00E+03 4,00E+06 1,00E+03
10 6,20E+03 6,00E+05
12 8,50E+03 6,00E+06 <103
13 9,00E+03 4,00E+04
17 4,00E+03 1,50E+04 <103
20 8,70E+03 2,20E+04
Lattobacilli (cfu/g)

149
Quindi il 35% dei soggetti esaminati ha presentato un aumento dei bifidobatteri statisticamente significativo; l’effetto prebiotico e la colonizzazione del ceppo somministrato Bifidobacterium longum sono confermate dall’elevata concentrazione di bifidobatteri presenti anche al wash-out.
In tab. 21 sono presentati i risultati riguardanti il contenuto fecale in lattobacilli. Nei gruppi lat-A e lat-B non si notano aumenti significativi nel contenuto fecale di lattobacilli ad esclusione del soggetto 19 che mostra un aumento di 2 unità log al tempo wash-out rispetto al T0g e T30g e dei soggetti 11 e 14 che presentano un comune aumento di circa 1 unità logaritmica nella concentrazione dei lattobacilli al T30g rispetto al T0g. Nel gruppo lat-C, tutti i 6 soggetti presentano un aumento comune e statisticamente significativo (P<0.05) della concentrazione dei lattobacilli al T30g rispetto al T0g, in particolare i soggetti 8, 10 e 12 hanno mostrato rispettivamente un aumento di 2 o 3 unità logaritmiche.
Quindi il 40% dei soggetti partecipanti al trial clinico ha presentato un aumento dei lattobacilli statisticamente significativo.

150

151
DISCUSSIONE
Questo studio ha analizzato il comportamento fermentativo del ceppo Bifidobacterium adolescentis MB 239 verso oligosaccaridi prebiotici e ha valutato, in sinergia con l’azienda Barilla G&R f.lli SpA, la possibilità di sviluppare nuovi prodotti alimentari funzionali contenenti simbiotici.
Per approfondire le preferenze del genere Bifidobacterium nei confronti di un ampia gamma di carboidrati prebiotici sono state condotte fermentazioni batch a pH non controllato di B. adolescentis MB 239 su diverse fonti di carbonio prebiotiche e sono state confrontate le affinità e le cinetiche di crescita sui rispettivi componenti mono- e disaccaridici.
In particolare è stato osservato che il ceppo presenta affinità soprattutto nei confronti di frutto-oligosaccaridi e queste affinità sono inversamente proporzionali al grado di polimerizzazione del carboidrato. Le fermentazioni condotte su carboidrati glucosidici indicano anche in questo caso la preferenza di B. adolescentis MB 239 per oligosaccaridi a medio grado di polimerizzazione (IMOS) piuttosto che per polisaccaridi o per il glucosio. La crescita su raffinosio e GOS, rispettivamente α-(1→6) e β-(1→4) galattosidi, evidenzia che l’affinità per entrambi è minore di quella osservata per il galattosio mentre è comunque maggiore di quella per il glucosio. È stato inoltre osservato che B. adolescentis MB 239 è in grado di sviluppare su

152
polisaccaridi insolubili forniti come unica fonte di carbonio, in particolare su amido resistente RS 2 e gomma guar PHGG.
La velocità specifica di crescita (µ) e la resa in biomassa non sono parametri sufficienti per determinare l’affinità di un ceppo per una fonte di carbonio, nonostante ciò dai dati ottenuti è possibile affermare che il microrganismo ha maggiore preferenza per gli oligosaccaridi piuttosto che per le loro componenti monosaccaridiche. Questa conclusione risulta essere in accordo con un precedente studio in cui è stato dimostrato che B. adolescentis MB 239 predilige gli oligosaccaridi piuttosto che i monosaccaridi (Amaretti et al. 2006) e con numerosi altri studi in cui questo concetto è stato ampiamente dimostrato per il genere Bifidobacterium (Palframan et al. 2003; Kim et al. 2003; Rada et al. 2002; Hopkins et al. 1998).
Successivamente lo studio è stato rivolto alla caratterizzazione fermentativa di B. adolescentis MB 239 su GOS, oligosaccaridi di notevole interesse poiché hanno uno spiccato potere bifidogenico e come tali stanno trovando di recente numerose applicazioni commerciali, in particolare nelle formulazioni di latte materno artificiale. Inoltre le informazioni da un punto di vista fermentativo su tale categoria di carboidrati sono ancora limitate, soprattutto rispetto a quelle disponibili per i fruttani.
Come primo passo si è determinato il pH ideale di crescita del ceppo in esame confrontandone lo sviluppo in fermentazioni batch a pH controllato in terreno semisintetico ai pH 4.5, 5.0, 5.5, 6.0 e 6.5, e utilizzando come fonte di carbonio fermentabile glucosio. Dal confronto si nota che a pH 5.5 si ottengono le migliori velocità specifiche di crescita ottenute che di resa di conversione di substrato in biomassa; è inoltre emerso che all’aumentare della biomassa prodotta diminuisce la produzione di acidi (stimati in base al consumo di NaOH utilizzata come correttore di pH).
Le fermentazioni batch di B. adolescentis MB 239 sono state condotte al pH ottimale su GOS e i suoi componenti mono- e disaccaridici glucosio, galattosio e lattosio; la velocità specifica di crescita massima (µMAX), il

153
coefficiente di saturazione (KS) e il coefficiente di resa cellulare (YX/S), normalmente determinabili solo mediante fermentazioni in chemostato, sono stati stimati mediante un algoritmo di calcolo elaborato in linguaggio MATLAB.
Questo algoritmo è in grado di fornire una stima di tali parametri a partire dai dati ricavati dalle fermentazioni batch. L’algoritmo si basa su un modello cinetico costituito da equazioni differenziali che descrivono la crescita della biomassa, il consumo di substrato e l’accumulo di prodotti legati alla crescita. Il modello cinetico si applica bene ai casi esaminati nella stima dei parametri che descrivono le cinetiche di crescita e del consumo del substrato producendo un basso SS. Tale valore rappresenta la radice degli scarti quadratici tra valori sperimentali e valori calcolati e stima l’errore nel processo di ottimizzazione.
Dal confronto della velocità specifica di crescita massima, del coefficiente di saturazione e del coefficiente di resa cellulare, si evince che il substrato che meglio viene fermentato è il galattosio sul quale si sono osservate le µMAX e YX/S più alte e la KS più bassa, indice di una migliore affinità per il substrato da parte del microrganismo. A conferma dei risultati ottenuti nelle fermentazioni batch a pH non controllato, il glucosio si è dimostrato il substrato peggiore con µMAX e YX/S più basse. Il valore di KS su GOS è decisamente più elevato rispetto alle altre fonti di carbonio: questo è dovuto al fatto che i componenti della miscela sono consumati con cinetiche differenti e i carboidrati con minore affinità, utilizzati al termine della fase di crescita, influiscono negativamente sul valore della KS.
La miscela di GOS utilizzata per le fermentazioni ha una composizione di oligosaccardi con DP compreso tra 3 e 9 al 58.3%, lattosio al 19.4%, glucosio al 19% e galattosio 3.5%. B. adolescentis MB 239 presenta una stringente selettività basata sul grado di polimerizzazione dal momento che gli oligosaccaridi con DP 2 (lattosio) e 3 vengono consumati per primi, mentre gli oligosaccaridi con grado di polimerizzazione maggiore vengono fermentati

154
solo dopo la completa deplezione del lattosio. Le differenti cinetiche di consumo dei componenti di questa miscela sono in accordo con differenti e decrescenti velocità di crescita. Questa sorta di poliauxia potrebbe essere spiegata dall’attività di diversi enzimi o trasportatori o dalla differente affinità tra enzima/trasportatore e substrato.
Durante le fermentazioni con glucosio, galattosio, lattosio e GOS è stato monitorato l’andamento dell’attività specifica della β-galattosidasi, enzima preposto all’idrolisi dei legami β-(1→4)-galattosidici, sia a livello intracellulare che di superficie. Si evidenzia che su glucosio tale attività è espressa in maniera basale, in presenza di galattosio invece, l’attività specifica diminuisce all’aumentare della concentrazione della biomassa durante la fase esponenziale di crescita per poi ristabilirsi a valori di 3 U/mgDW. In terreno contenente lattosio, come ci si aspettava, l’attività specifica dell’enzima aumenta proporzionalmente all’incremento della biomassa cellulare, come in presenza di GOS. Da ciò si deduce che la presenza di impurezze di glucosio nei GOS non reprime la produzione dell’enzima. L’accumulo di galattosio durante la fermentazione fa presupporre inoltre che la velocità di produzione di questo monomero sia maggiore di quella del suo uptake.
L’analisi effettuata mediante HPLC dei brodi di fermentazione mette in evidenza che i prodotti del metabolismo di B. adolescentis MB 239 su glucosio, galattosio, lattosio e GOS sono i prodotti caratteristici dello shunt dell’esoso fosfato, acidi lattico ed acetico, e quelli derivanti dalla reazione fosforoclastica cioè etanolo, ma in nessuno dei casi si è osservata la produzione di acido formico. A partire dai coefficienti di resa calcolati dall’algoritmo per i prodotti sono state stimate le porzioni del flusso di carbonio, rappresentato dal substrato di crescita, che vengono trasformate dal ceppo di B. adolescentis nei prodotti del suo metabolismo: si può notare che su tutti gli zuccheri la percentuale di flusso di carbonio convogliata sull’acido lattico oscilla sempre tra il 43 e il 47%, mentre il resto è rappresentato dalla somma di acido acetico ed etanolo. Inoltre sono notevoli le differenze

155
riscontrate nella resa di produzione dell’etanolo al variare della fonte di carbonio in particolare tra glucosio e galattosio. Su galattosio l’acetil-P prodotto dallo shunt viene trasformato interamente in acido acetico, mentre su glucosio una percentuale rilevante viene trasformata in etanolo e questo a scapito della resa in ATP; questa constatazione può spiegare la minore produzione di biomassa cellulare della fermentazione del glucosio rispetto a quella del galattosio.
Al fine di approfondire ulteriormente la conoscenza del metabolismo dei carboidrati in Bifidobacterium verso una classe di prebiotici ancora poco studiata come gli xilo-oligosaccaridi (XOS), è stata purificato e caratterizzato un enzima extracellulare di B. adolescentis MB 239 appartenente alla famiglia delle glicosil-idrolasi. Inizialmente sono state svolte fermentazioni batch a pH controllato con xilosio o XOS come sola fonte di carbonio. In questo caso, la
differenza tra la µ su xilosio e XOS è minima (0.26 vs 0.28 h-1); le velocità specifiche di crescita quindi si discostano leggermente dal trend ottenuto per gli altri carboidrati analizzati; in questo caso quindi non si riscontra una forte preferenza del ceppo verso gli oligosaccaridi dello xilosio piuttosto che verso il monosaccaride. Questo concetto è confermato dai valori delle rese in biomassa finali: lo xilosio permette al ceppo di ottenere una più elevata resa in biomassa (3.1 g L-1) rispetto a quella raggiunta su XOS (1.3 g L-1). Questi risultati sono in accordo con Palframan et al. 2003 in cui viene dimostrato che B. adolescentis cresce meglio su xilosio piuttosto che su XOS, e che questa caratteristica è una peculiarità di un gruppo di bifido batteri a cui appartengono anche le specie B. catenulatum e B. gallicum.
Durante le due fermentazioni, è stata monitorata l’attività enzimatica responsabile dell’idrolisi degli xilo-oligosaccaridi espressa sia a livello
intracellulare che a livello superficiale [β-xilosidasi (β-Xil)]. Si evidenzia che su xilosio l’attività specifica della β-Xil aumenta proporzionalmente all’incremento della biomassa cellulare, arrivando ad un massimo di 5.1 U mgDW
-1 all’inizio della fase stazionaria. Al contrario, su XOS, l’attività

156
specifica diminuisce all’aumentare della concentrazione della biomassa durante la fase esponenziale di crescita per poi ristabilirsi a valori di 4.2 U
mgDW-1. Come espresso in altri studi per la β-fruttofuranosidasi (Rossi et al.,
2005; Amaretti et al. 2006) anche in questo caso si può affermare che il
monosaccaride non reprime la produzione della β-xilosidasi, anzi, ne promuove un accumulo. Se il meccanismo di trasporto a livello di superficie per lo xilosio è più efficiente di quello per gli XOS, allora l’affinità per il monosaccaride sarà migliore di quella per i suoi oligosaccaridi, come
dimostrato in questo studio. Dato che l’attività specifica della β-xilosidasi di superficie, durante la fermentazione su XOS, è maggiore che in quella su xilosio, si potrebbe presupporre che in questo ceppo gli oligosaccaridi vengano idrolizzati prima di essere internalizzati e solo dopo fermentati.
Con questo presupposto, si è voluto approfondire il meccanismo di idrolisi extracellulare degli XOS da parte di B. adolescentis MB 239 ed è stata
quindi purificata una β-xilosidasi extracellulare partendo dal surnatante della fermentazione condotta su XOS. L’enzima è stato isolato mediante uno step di cromatografia a scambio anionico e uno successivo di cromatografia ad esclusione molecolare, ottenendo un’attività specifica di 596 U mgprot
-1. Durante gli step di purificazione è stato possibile evidenziare la presenza di un unico monomero per la proteina denaturata, proteina che, nella forma nativa, si presenta come un omotetramero del peso molecolare di circa 160 kDa.
L’attività enzimatica è stata studiata in funzione della temperatura e del pH ed è stato calcolato che i valori ottimali ai quali corrisponde la massima attività sono rispettivamente per T = 53°C e pH 6. Valutata anche la stabilità
dell’enzima nel tempo, si può affermare che questa β-xilosidasi è termostabile in quanto conserva più del 60% di attività residua rispetto al suo massimo se mantenuta a 53°C per più di 24 ore. Queste caratteristiche sono in accordo con
i dati riportati per β-xilosidasi espresse da 3 ceppi di Bifidobacterium (Zeng et
al. 2007) e per β-xilosidasi purificate da altri microrganismi quali ad esempio

157
Termomonospora fusca, Termobifida fusca e Bacillus thermantarcticus (Lama et al. 2004; Bachmann e McCarthy 1991, Tuncer 2000), in cui sono riportati optitmum di pH e di temperatura compresi tra 6-9 e 50-70°C, rispettivamente.
La cinetica enzimatica di questo enzima isolato da B. adolescentis MB 239 è stata caratterizzata calcolando i valori di km = 4.36 mM e di Vmax = 0.93 mM min-1, valori che presentano una minore affinità per questo substrato
sintetico rispetto a quella di altre β-xilosidasi (Bachmann e McCarthy 1989; Deshpande et al. 1986).
Per approfondire l’affinità di substrato di questo enzima e valutare eventuali altre attività enzimatiche secondarie, sono stati condotti esperimenti di idrolisi con numerosi di-, oligo- e polisaccaridi. I prodotti di reazione, visualizzati mediante TLC, hanno permesso di individuare una preponderante attività beta-xilosidasica nei confronti degli XOS, i quali sono stati quasi completamente degradati a xilosio e una debolissima attività xilanasica nei confronti dello xilano. In questo secondo caso è stato possibile notare un’attività di endo-xilanasi dato che lo xilano è stato degradato in parte con la formazione di catene di xilo-oligosaccaridi con grado di polimerizzazione pari a 4 e 5 senza accumulo di xilosio.
Secondo Zeng (Zeng et al. 2007) nei 3 ceppi analizzati di
Bifidobacterium sono espresse solamente β-xilosidasi intracellulari e si sostiene quindi che in questi casi gli xilo-oligosaccaridi dovrebbero essere internalizzati mediante trasportatori di oligosaccaridiad alta affinità MalEFG-type già predetti dalla’analisi del genoma di B. longum (Schell et al. 2002)
La caratterizzazione di questo enzima extracellulare fornisce informazioni importanti sul metabolismo dei bifidobatteri verso una nuova classe di carboidrati prebiotici, anche dal momento che sono presenti
pochissimi studi in letteratura che abbiano caratterizzato β-xilosidasi da Bifidobacterium (Lagaert et al. 2007; Shin et al. 2003; Zeng et al. 2007) e nessuno che si sia occupato di questa classe di enzimi extracellulari.

158
Lo studio del metabolismo di oligosaccaridi prebiotici in Bifidobacterium ha permesso poi di svolgere, insieme all’azienda Barilla G&R f.lli SpA, uno studio per il potenziale sviluppo di prodotti alimentari innovativi e funzionali, ossia contenenti simbiotici. Questo progetto si prefiggeva di - caratterizzare i ceppi probiotici scelti; - garantire la sopravvivenza dei probiotici in prodotti alimentari conservati a temperatura ambiente e con una shelf life di circa 180 giorni; - ottimizzare la formulazione della matrice alimentare al fine di migliorare la sopravvivenza dei ceppi probiotici inoculati; - verificare la colonizzazione e gli effetti benefici dei ceppi prescelti in individui sani.
Come frazione probiotica sono stati scelti 3 ceppi appartenenti alla collezione Barilla, Lactobacillus plantarum BAR 10, Streptococcus thermophilus BAR 20 e Bifidobacterium lactis BAR 30, in quanto ceppi con elevato potere antiossidante e in grado di produrre elevate quantità di glutatione. Infatti questi ceppi erano emersi, durante uno screening svolto precedentemente, come ceppi caratterizzati da peculiarità importanti quali appunto la produzione di glutatione (fino a 30 nmoli/mgprot per B.lactis BAR 30) e la capacità di inibire l’autoossidazione dell’acido ascorbico e l’ossidazione dell’acido linolenico. Parametro fondamentale per un probiotico è l’acido resistenza e quindi la capacità di superare la barriera gastrica per poter raggiungere, ancora vitale, il colon. Il normale tempo di svuotamento gastrico varia da 20 minuti a 3-4 ore in funzione di ciò che si è ingerito, dalla composizione del cibo e dalla dimensione delle particelle alimentari. Sebbene un modello dinamico simuli in modo realistico il transito di un probiotico attraverso il tratto gastro-intestinale, è stato utilizzato un modello statico per la valutazione della tolleranza ai succhi gastrici in modo tale da standardizzare le condizioni di assunzione, come proposto in numerosi lavori (Charteris et al. 1998, Crittenden et al. 2001, Vernazza et al. 2006).

159
E’ stato quindi necessario valutare la resistenza ai succhi gastrici e alla bile di questi 3 ceppi probiotici, per un tempo di contatto rispettivamente di 30, 60 o 90 minuti; l’azione del succo gastrico umano e del succo gastrico simulato è stata confrontata. In accordo con i nostri risultati, è noto che la resistenza di molti ceppi di S. thermophilus è molto bassa (Conway et al. 1987; Marteau et al. 1997), mentre ceppi di B. lactis sopravvivono molto bene al contatto con i succhi gastrici. La conta vitale in succo gastrico umano conferma che B. lactis BAR 30 e L. plantarum BAR 10 sono più acido tolleranti che S. thermophilus BAR 20, anche se in generale, la perdità di vitalità in succo gastrico umano è più drastica di quella in succo gastrico simulato. Quindi, nonostante il contatto con una soluzione di pepsina a pH 2 sia utilizzata da molti ricercatori per simulare il transito gastrico (Masco et al. 2007, Marteau et al. 1997, Charteris et al. 1998 ), probabilmente non rappresenta il modello di simulazione più appropriato, considerato quanto si discostano tra loro i risultati per i 3 ceppi analizzati. La bile suina sembra invece non incidere negativamente sulla vitalità dei ceppi probiotici.
Dato che la somministrazione simultanea di probiotici e di un substrato da loro fermentabile offre ai ceppi somministrati una maggiore probabilità di colonizzazione e di sopravvivenza nel colon dell’ospite, è stata studiata l’affinità dei 3 ceppi probiotici per 13 carboidrati prebiotici, suddivisibili in fibre solubili e fibre insolubili. Nessun ceppo è in grado di fermentare inulina, arabino-galattani (AG), destrano, gomma guar (PHGG) e amido resistente (RS 1), mentre solo L. plantarum BAR 10 and B. lactis BAR 30 riescono ad utilizzare gli xilo-oligosaccaridi (XOS) e alcuni oligosaccaridi del glucosio quali isomalto-oligosaccaridi (IMOS), polidestrosio (PD) e amido resistente (RS 2). I galatto-oligosaccaridi (GOS) e i frutto-oligosaccaridi (FOS 1 e FOS 2) vengono fermentati molto bene dai ceppi analizzati e inducono elevate rese in biomassa; in accordo con la letteratura (Saulnier et al. 2007, Rossi et al. 2005, Gopal et al. 2001).

160
Al fine di valutare quali fossero i parametri che incidono maggiormente sulla sopravvivenza la vitalità dei 3 ceppi probiotici è stata studiata in matrici alimentari e conservate in differenti condizioni di temperatura e di concentrazione di ossigeno. Inizialmente il ceppo B. lactis BAR 30 è stato inoculato in matrici di crema anidra proposte dalla ditta Barilla; si evince che la temperatura di conservazione gioca un ruolo fondamentale nel mantenimento della vitalità dei probiotici in matrice, dato che a temperatura ambiente la vitalità cala drasticamente rispetto a quella dei campioni conservati a 4°C. Lo scopo di ulteriori prove è quello di migliorare la matrice Barilla, al fine di prolungare la sopravvivenza del ceppo probiotico inoculato, variando la componente glucidica con carboidrati semplici o prebiotici introducendo o mantenendo caratteristiche tecnologiche, organolettiche e nutrizionali ottimali. La matrice RG è risultata essere tra quelle analizzate estremamente promettente per la sopravvivenza a temperatura ambiente del ceppo B. lactis BAR 30, esplicando anche un forte effetto di protezione verso il probiotico se conservato a temperatura ambiente. Il titolo del liofilizzato inoculato in matrice Infatti è rimasto abbastanza elevato dopo 6 mesi di conservazione, mentre in tutte le altre matrici la concentrazione di MB 238 è diminuita fino a valori inferiori a 102 CFU g-1.
Essendo noto che i microrganismi probiotici presentano una buona sopravvivenza in cioccolato (Saarela et al. 2006) e al fine di migliorare ulteriormente i risultati ottenuti con le matrici precedenti, L. plantarum BAR 10, S. thermophilus BAR 20 e B. lactis BAR 30 sono stati inoculati in barrette di cioccolato tal quale o addizionato dei 3 carboidrati prebiotici verso i quali i ceppi avevano presentato una migliore affinità: galatto-oligosaccaridi GOS, frutto-oligosaccaridi FOS 1 e frutto-oligosaccaridi FOS 2. Sono stati effettuati conteggi di vitalità al fine di valutare se i prebiotici addizionati potessero offrire una ulteriore e differenziata protezione per la sopravvivenza dei probiotici in un prodotto alimentare conservato a temperatura ambiente.

161
In generale è stato riscontrato che non si notano differenze significative tra le varie matrici, indice quindi che i carboidrati addizionati non giocano un ruolo fondamentale nella vitalità dei microrganismi. I 3 ceppi probiotici conservano una buona vitalità fino a 180 giorni di conservazione a temperatura ambiente, inducendo a considerare il cioccolato come matrice ideale per il potenziale sviluppo di un prodotto da forno funzionale.
E’ stato svolto un trial clinico su volontari sani con lo scopo di valutare in vivo il pre-prototipo sviluppato dall’azienda Barilla e quindi determinare l’efficacia del prebiotico e la colonizzazione dei ceppi probiotici addizionati, è stato svolto un trial clinico su volontari sani. I soggetti, monitorati dal punto di vista clinico e microbiologico, hanno dato risposta positiva all’assunzione del biscotto simbiotico: dal conteggio dei lattobacilli e dei bifidobatteri a livello fecale si è osservato che il 35% dei soggetti esaminati ha presentato un aumento dei bifidobatteri statisticamente significativo, mentre il 40% dei volontari sani ha mostrato un aumento dei lattobacilli statisticamente significativo. Attraverso questa sperimentazione è stata determinata l’efficiacia del pre-prototipo formulato da Barilla, ponendo in risalto sia la colonizzazione dei ceppi probiotici assunti che l’effetto prebiotico delle fibre addizionate al prodotto.

162

163
CONCLUSIONI
Il genere Bifidobacterium è costituito da batteri saccarolitici anaerobi e costituisce uno dei più importanti gruppi batterici della microflora intestinale dell’uomo e degli animali. La presenza e l’attività dei bifidobatteri nel colon è generalmente associata ad effetti benefici sulla salute e per questo motivo i bifidobatteri sono comunemente considerati probiotici e trovano un vasto impiego come supplementi di prodotti alimentari e farmaceutici. Gli oligo- e i polisaccaridi presenti nel colon rappresentano la principale fonte di carbonio fermentabile per i microrganismi appartenenti a tale genere: molti carboidrati non digeribili dall’uomo, in particolare frutto-oligosaccaridi (FOS), inulina, galatto-oligosaccaridi (GOS) e oligosaccaridi della soia, stimolano selettivamente i bifidobatteri a livello intestinale e trovano applicazione come prebiotici. Il metabolismo dei carboidrati in Bifidobacterium è stato oggetto di numerosi studi focalizzati soprattutto sugli effetti bifidogenici dei carboidrati prebiotici, mentre la conoscenza dell’uptake, dell’idrolisi e degli enzimi deputati ad essa e della fermentazione degli zuccheri è ancora incompleta e di notevole interesse. Questo studio è stato svolto al fine di approfondire le caratteristiche fermentative di un ceppo di Bifidobacterium (B. adolescentis MB 239), verso un elevato numero di carboidrati prebiotici attraverso l’analisi delle velocità specifiche di crescita e le rese finali in biomassa. In particolare l’attenzione è

164
stata posta su 2 classi di oligosaccaridi prebiotici, ossia i galatto-oligosaccaridi e gli xilo-oligosaccaridi. Per quanto riguarda i GOS, questo studio ha valutato come le differenze di grado di polimerizzazione di carboidrati prebiotici strutturalmente analoghi, incidano sulle capacità fermentative dei bifidobatteri. Il modello matematico, sviluppato mediante un algoritmo scritto in linguaggio Matlab, ha descritto in modo accurato le cinetiche delle colture batch cresciute su glucosio, galattosio, lattosio e GOS. La preferenza per la fonte di carbonio è stata determinata mediante il confronto delle velocità specifiche, delle rese in biomassa e delle costanti di saturazione. In accordo con studi già presenti in letteratura, B. adolescentis MB 239 è in grado di fermentare i galatto-oligosaccaridi a differente grado di polimerizzazione con una netta preferenza per gli oligosaccaridi piuttosto che per il glucosio.
Nei bifidobatteri, le glicosil-idrolasi sono responsabili dell’idrolisi degli oligo- e polisaccaridi, i cui componenti monosaccaridi vengono successivamente incorporati nello shunt del fruttosio-6-fosfato, fornendo acido lattico e acido acetico come prodotti principali e etanolo e acido formico come prodotti secondari. In risposta a differenti richieste cellulari di ATP o NAD, il piruvato può essere deviato dalla formazione di lattato alla formazione di formiato attraverso la reazione fosforoclastica, l’acetil-P può essere ridotto a etanolo a spese della produzione di acetato. Inoltre, parte del flusso di carbonio deve essere impiegato per la produzione di biomassa; di conseguenza le quantità relative dei prodotti di reazione sono tutt’altro che invariabili e quindi non ci si deve attendere sempre il classico rapporto lattato: acetato uguale a 1:1.5 descritto in letteratura.
L’approfondimento dell’affinità di B. adolescentis MB 239 verso gli xilo-oligosaccardi ha portato all’isolamento e alla caratterizzazione di una glicosil-idrolasi extracellulare, enzima preposto all’idrolisi dei legami β-(1→4) degli XOS, portando alla formazione di xilosio libero. La β-xilosidasi rilascia residui di xilosio dagli XOS che presentano un grado di polimerizzazione compreso tra 2 e 5. La crescente importanza industriale di

165
questi carboidrati deriva dal loro effetto benefico per la salute dell’uomo in quanto prebiotici. Così come i più noti prebiotici (frutto-oligosaccaridi e galatto-oligosaccaridi) anche gli XOS favoriscono lo sviluppo selettivo dei bifidobatteri ma possiedono anche interessanti caratteristiche fisico-chimiche e organolettiche e sono quindi adatti ad essere incorporati in alimenti funzionali.
Questo enzima è stato purificato partendo dal surnatante di una coltura batch a pH costante sviluppata su XOS come sola fonte di carbonio. La purificazione e caratterizzazione di questo enzima è riassunta nella tabella sottostante.
La presenza di questa β-xilosidasi nel surnatante può far presupporre quindi che B. adolescentis MB 239 sia in grado di idrolizzare di xilo-oligosaccaridi a livello extracellulare, per poi internalizzare le unità più semplici di xilosio. Infine lo studio del metabolismo degli oligosaccaridi in Bifidobacterium ha contribuito alla ricerca per il potenziale sviluppo di un prodotto alimentare funzionale shelf-stable contenente un simbiotico. Nello sviluppo di un prodotto da forno funzionale è fondamentale che venga mantenuta elevata nel tempo la popolazione probiotica a temperatura ambiente, quindi la vitalità dei microrganismi probiotici da veicolare. Oltre a promuovere e migliorare le funzioni di equilibrio fisiologico dell’organismo, i probiotici hanno attività
ββ--xxiilloossiiddaassii eexxttrraacceelllluullaarree MMWW ddeellllaa ssuubbuunniittàà 3388 kkDDaa ddaa SSDDSS--PPAAGGEE
MMWW ddeellllaa pprrootteeiinnaa nnaattiivvaa 116600 kkDDaa,, tteettrraammeerroo
AAttttiivviittàà vvss tteemmppeerraattuurraa MMaassssiimmaa aattttiivviittàà aa 5533°°CC AAttttiivviittàà vvss ppHH MMaassssiimmaa aattttiivviittàà aa ppHH==66 SSttaabbiilliittàà aallll’’ooppttiimmuumm ddii tteemmppeerraattuurraa AAttttiivviittàà mmaaggggiioorree ddeell 6600%% ddooppoo 2244 hh
SSttaabbiilliittàà iinn ffuunnzziioonnee ddeell ppHH SSttaabbiillee aa ppHH==55--66 ppeerr ppiiùù ddii 2244 hh
SSppeecciiffiicciittàà ddii ssuubbssttrraattoo IIddrroolliissii ccoommpplleettaa ddeeggllii XXOOSS ((ββ--xxiilloossiiddaassii)),, ddeebboollee iiddrroolliissii ddeelllloo xxiillaannoo ((xxiillaannaassii))
PPaarraammeettrrii cciinneettiiccii kkmm == 44..3366 mmMM ;; VVmmaaxx == 00..9933 mmMM//mmiinn

166
immunostimolante potenziando la risposta immunitaria, attività antitumorale e antinfiammatoria e contribuiscono ad abbassare lo stress ossidativo del colon.
La prima fase della ricerca è stata finalizzata ad un’approfondimento delle caratteristiche metaboliche e fisiologiche della miscela probiotica scelta. Una delle più importanti proprietà che un ceppo probiotico deve possedere è la resistenza ai succhi gastrici, per poter raggiungere ancora vitale e in elevate concentrazioni l’intestino, dove esplicherà le proprie proprietà benefiche. Un secondo aspetto indagato è stato l’affinità dei ceppi probiotici selezionati nei confronti di numerosi carboidrati prebiotici. I ceppi L. plantarum BAR 10, S. thermophilus BAR 20 e B. lactis BAR 30 sono stati coltivati su terreni contenenti tali carboidrati come sole fonti di carbonio e sono state confrontate le rese in biomassa e le velocità specifiche.
Attraverso questi esperimenti in vitro è stato possibile selezionare 3 carboidrati prebiotici verso i quali tutti i ceppi da noi selezionati presentano una maggiore affinità cioè galatto-oligosaccaridi e frutto-oligosaccaridi a corta e media catena. Si è pensato quindi che questi prebiotici potessero essere i candidati ideali nella formulazione del pre-prototipo del prodotto simbiotico da forno. Tali prebiotici sono quindi stati singolarmente inseriti nella formulazione di barrette di cioccolato inoculate con una miscela dei 3 ceppi liofilizzati Lactobacillus plantarum, Streptococcus thermophilus e Bifidobacterium lactis presenti tutti in elevate concentrazioni al momento della preparazione delle barrette. Data l’ottima sopravvivenza dei 3 ceppi a temperatura ambiente per 180 giorni, la ditta Barilla ha sviluppato un prodotto pre-prototipo da utilizzare in un trial clinico in vivo. Dal punto di vista microbiologico, questo trial ha dimostrato l’effettiva colonizzazione dei probiotici assunti con il prodotti e un effettivo aumento dei bifidobatteri e lattobacilli totali a livello fecale, lasciano aperta la possibilità per lo sviluppo di un prodotto alimentare con caratteristiche analoghe.

BIBLIOGRAFIA
1. Alliet P, Scholtens P, Raes M, Hensen K, Jongen H, Rummens JL,
Boehm G, Vandenplas Y. 2007. Effect of prebiotic galacto-oligosaccharide, long-chain fructo-oligosaccharide infant formula on serum cholesterol and triacylglycerol levels. Nutr. 23:719-723.
2. Amaretti A, Tamburini E, Bernardi T, Pompei A, Zanoni S, Vaccari
G, Matteuzzi D, Rossi M. 2006. Substrate preference of Bifidobacterium adolescentis MB 239: compared growth on single and mixed carbohydrates. Appl. Microbiol. Biotechnol. 73:654-662.
3. Amaretti A, Bernardi T, Tamburini E, Zanoni S, Lomma M,
Matteuzzi D, Rossi M. 2007. Kinetics and Metabolism of Bifidobacterium adolescentis MB 239 Growing on Glucose, Galactose, Lactose, and Galactooligosaccharides. Appl. Environ. Microbiol. 73:3637-44.
4. Bachmann SL, McCarthy AJ. 1989. Purification and Characterization
of a Termostable β-Xylosidase from Thermomonospora fusca. J. Gen. Microbiol. 135:293-299.

168
5. Bachmann SL, McCarthy AJ. 1991. Purification and Cooperative Activity of Enzymes Constituting the Xylan-Degrading System of Thermomonospora fusca. Appl. Environ. Microbiol. 57:2121-2130.
6. Benno Y, Mitsuoka T. 1986. Development of intestinal microflora in
human and animals. Bifidobacteria Microflora 5:13-25.
7. Bernfeld P. 1955. S.I. Colowick, N.O. Kaplan (Eds.), Methods in Enzymology, vol. 1, Academic Press, New York, , pp. 149–158.
8. Bezkorovainy A, Miller-Catchpole R. 1989. Biochemestry and
physiology of bifidobacteria. CRC Press, London, United Kingdom.
9. Biaviati B, Castagnoli P, Trovatelli LD. 1986. Species of the genus Bifidobacterium in the feces of the human adults. Microbiologica 9:39-45.
10. Biavati B, Mattarelli P. 2001. The family Bifidobacteriaceae. In M.
Dworkin et al. (ed.) The Prokaryotes: An Evolving Electronic Resource for the Microbiological Community, 3rd edition, release 3.7. Springer-Verlag. New York.
11. Bielecka M, Biedrzycka E, Majkowska A: 2002. Selection of
probiotics and prebiotics for synbiotics and confirmation of their in vivo effectiveness. Food Res. Int. 35:125-131.
12. Boonomee M, Lekswasdi N, Bridge W, Rogers PL. 2002. Batch and
continuous culture of Lactococcus lactis NZ133: experimental data and model development. Biochemical Engineering Journal 14:127-135.

169
13. Bouhnik Y, Flouriè B, D’agay-Abensour L, Pochert P, Gramet G, Durand M, Rambaud JC. 1997. Administration of transgalacto-oligosaccharides increases faecal bifidobacteria and modifies colonic fermentation metabolism in healthy humans. J. Nutr. 127:444-448.
14. Bouhnik Y, Vahedi K, Achour L, Attar A, Salfati J, Pochart P,
Marteau P, Flourie B, Bornet F, Rambaud JC. 1999. Short-chain Fructooligosaccharides administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 429:113-116.
15. Bouhnik Y, Raskine L, Simoneau G, Paineau D, Bornet F. 2006. The
capacity of short-chain fructo-oligosaccharides to stimulate faecal bifidobacteria: a dose-response relationship study in healthy humans. Nutr. J. 28:5-8
16. Brown I, Warhurst M, Arcot J, Playne M, Illman RJ, Topping DL.
1997. Faecal numbers of bifidobacteria are higher in pigs fed Bifidobacterium longum with a high amylose cornstarch than with a low amylose cornstarch. J. Nutr. 127:1822-1827.
17. Bourriaud C, Robins RJ, Martin L, Kozlowski F, Tenaileau E,
Cherbut C, Michel C. 2005. Lactate is mainly fermented to butyrate by human intestinal microfloras but inter-individual variation is evident. J. Appl. Microbiol. 99:201–212.
18. Bornet FR, Brouns F, Tashiro Y, Duvillier V. 2002. Nutritional
aspects of short-chain fructooligosaccharides: natural occurrence, chemistry, physiology and health implications. Dig. Liver. Dis. 34:S111-S120.

170
19. Campbell JM, Fahey GC, Wolf BW. 1997. Selected indigestible oligosaccharides affect large bowel mass, cecal and faecal short-chain fatty acids, pH and microflora in rats. J. Nutr. 127:130-136.
20. Casiraghi MC, Canzi E, Zanchi R, Donati E, Villa L. 2007. Effects of
a synbiotic milk product on human intestinal ecosystem. J. Appl. Microbiol. 103 :499-506.
21. Chacko A, Cummings JH. 1988. Nitrogen losses from the human small
bowel: obligatory losses and the effect of physical form of food. Gut 29:809-815.
22. Charteris WP, Kelly PM, Morelli L, Collins JK. (1998) Development
and application of an in vitro methodology to determine the transit tolerance of potentially probiotic Lactobacillus and Bifidobacterium species in the upper human gastrointestinal tract. J. Appl. Microbiol. 84:759-768.
23. Chen HS, Fan YS, Chen ME, Chan Y. 2005. Unhydrolyzed and
hydrolyzed konjac glucomannans modulated cecal and fecal microflora in Balb/c mice. Nutr. 21:1059-1064.
24. Chisari G, Lo Bue AM, Drago L, Abbiati R, Gismondo MR. 1992.
Adhesion capacity of Enterococcus faecium (SF 68) and Enterococcus faecalis to various substrates. G Ital Chemioter. 39:11-15.
25. Chung CH, Day DF. 2002. Glucooligosaccharides from Leuconostoc
mesenteroides B-742 (ATCC 13146): A potential prebiotic. J. Indust. Microbiol. Biotechnol. 29:196-199.

171
26. Conway PL, Gorbach SL, Goldin BR. 1987. Survival of lactic acid bacteria in the human stomach and adhesion to intestinal cells. J. Dairy Sci. 70:1-12.
27. Crittenden RG, Martinez NR, Playne MJ. 2003. Synthesis and
utilisation of folate by yoghurt starter cultures and probiotic bacteria. Int. J. Food Microbiol. 80:217-222.
28. Crittenden RG, Morris LF, Harvey ML, Tran LT, Mitchell HL,
Playne MJ. 2001. Selection of a Bifidobacterium strain to complement resistant starch in a synbiotic yoghurt. J. Appl. Microbiol. 90:268-278.
29. Crociani F, Alessandrini A, Mucci MM, Biavati B. 1994. Degradation
of complex carbohydrates by Bifidobacterium spp. Int. J. Food. Microbiol. 24:199-210.
30. Crociani F, Matteuzzi D. 1982. Urease activity in the genus
Bifidobacterium. Ann. Microbiol. 133:417-423.
31. Cummings JH, MacFarlane GT. 1991. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 70:443–459.
32. Cummings JH, MacFarlane GT. 1997. Role of intestinal bacteria in
nutrient metabolism. Clin. Nutr. 16:3-11.
33. Degnan BA, MacFarlane GT. 1991. Comparison of carbohydrate substrate preferences in eight species of bifidobacteria. FEMS Microbiol. Lett. 68:151-156.

172
34. Derensy-Dron D, Krzewinski F, Brassart C, Bouquelet S. 1999. Beta-1,3-galactosyl-N-acetylhexosamine phosphorylase from Bifidobacterium bifidum DSM 20082: characterization, partial purification and relation to mucin degradation. Biotechnol. Appl. Biochem. 29:3-10.
35. Derrien M, Vaughan EE, Plugge CM, de Vos WM. 2004.
Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin degrading bacterium. Int. J. Syst. Evol. Microbiol. 54:1469–1476.
36. Deshpande V, Lachke A, Misra C, Keskar S, Rao M. 1986. Mode of
action and properties of xylanase and b-xylosidase fro Neurospora crassa. Biotechnol. Bioeng. 28:1832-1837.
37. Dethlefsen L, Eckburg PB, Bik EM, Relman DA. 2006. Assembly of
the human intestinal microbiota. Trends Ecol. Evol. 21:517-523.
38. de Vries W, Stouthamer AH. 1967. Pathway of glucose fermentation in relation to the taxonomy of bifidobacteria. J. Bacteriol. 93:574-576.
39. Dinoto A, Suksomcheep A, Ishizuka S, Kimura H, Hanada S,
Kamagata Y, Asano K, Tomita F, Yokota A. 2006. Modulation of rat cecal microbiota by administration of raffinose and encapsulated Bifidobacterium breve. Appl. Environ. Microbiol. 72:784-792.
40. Ducluzeau R, Bellier M, Raibaud P. 1970. Transit through the
digestive tract of the inocula of several bacterial strains introduced "per Os" into axenic and "Holoxenic" mice. The antagonistic effect of the microflora of the gastrointestinal tract. Zentralbl. Bakteriol. 213:533-548.

173
41. Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. 2005. Diversity of the human intestinal microbial flora. Science. 308:1635-1638.
42. Ehrmann MA, Korakli M, Vogel RF. 2003. Identification of the gene
for beta-fructofuranosidase of Bifidobacterium lactis DSM10140(T) and characterization of the enzyme expressed in Escherichia coli. Curr. Microbiol. 46:391-397.
43. Finney M, Smullen J, Foster HA, Brokx S, Storey DM. 2007. Effects
of low doses of lactitol on faecal microflora, pH, short chain fatty acids and gastrointestinal symptomology. Eur. J. Nutr. 46:307-314.
44. Flickinger EA, Wolf BW, Garleb KA. 2000. Glucose-based
oligosaccharides exhibit different in vitro fermentation patterns and affect in vivo apparent nutrient digestibility and microbial populations in dogs. J. Nutr. 130:1267-1273.
45. Flint HJ. 2006. The significance of prokaryote diversity in the human
gastrointestinal tract. In Prokaryotic Diversity: Mechanism and Significance. Logan, N.A. Lappin-Scott, H.M. and Oyston, P.C.F. (eds). Cambridge, UK: Cambridge University Press, pp 65-90. SGM Symposium vol. 66.
46. Flint HJ, Duncan SH, Scott KP, Louis P. 2007. Interactions and
competition within the microbial community of the human colon: links between diet and health. Environ Microbiol. 9:1101-1111.

174
47. Food and Nutrition Board, Institute of Medicine, National Academy of Sciences. 1994. Opportunities in the Nutrition and Food Sciences. PR Thomas and R Earl, editors. Washington, DC: National Academy Press.
48. Giannini EG, Mansi C, Dulbecco P, Savarino V. 2006. Role of
partially hydrolyzed guar gum in the treatment of irritable bowel syndrome. Nutr. 22:334-342.
49. Gibson GR, Beatty ER, Wang X, Cummings JH. 1995. Selective
stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterol. 108:975-982.
50. Gibson GR, Roberfroid MB. 1995. Dietary modulation of the human
colonic microbiota: introducing the concept of prebiotic. J. Nutr. 25:1401-1412.
51. Gibson GR, Wang X. 1994. Bifidogenic properties of different types of
fructo-oligosaccharides. Food Microbiol. 11:491-498.
52. Gibson GR, Probert HM, Van Loo JAE, Roberfroid MB. 2004. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 17:257–259.
53. Gmeiner M, Kneifel W, Kulbe KD, Wouters R, De Boever P, Nollet
L, Verstraete W. 2000. Influence of a synbiotic mixture consisting of Lactobacillus acidophilus 74-2 and a fructooligosaccharide preparation on the microbial ecology sustained in a simulation of the human intestinal microbial ecosystem (SHIME reactor). Appl. Microbiol. Biotechnol. 53:219-223.

175
54. Gopal PK, Sullivan PA, Smart JB. 2001. Utilisation of galacto-oligosaccharides as selective substrates of growth by lactic acid bacteria including Bifidobacterium lactis DR10 and Lactobacillus rhamnosus DR20. Int. Dairy J. 11:19-25.
55. Gorbach SL. 2002. Probiotics in the third millennium. Dig. Liver Dis.
34 Suppl 2:S2-7.
56. Hammes WP, Weiss N, Holzapfel W. 1991. The genera Lactobacillus and Carnobacterium, in: The Prokaryotes (Balows, A., Trüper, H., G., Dworkin, M., Harder, W., Schleifer, K., H., Eds.) 2nd Ed., vol 2. pp. 1535-1594 New York: Springer-Verlag.
57. Hardie JM. 1986. Genus Streptococcus. In Bergey’s Manual of
Systematic Bacteriology, 9th edn., Vol. 2 (Sneath, P.H.A., Mair, N.S., Sharpe, M.E., and Holt, J.G., eds.) pp 1043-1071, Williams and Wilkins, Baltimore, MD.
58. Hartemink R, Kok BJ, Weenk GH, Rombouts FM. 1996. Raffinose-
Bifidobacterium (RB) agar, a new selective medium for bifidobacteria. J Microbiol. Meth. 27:33-43.
59. Health Canada. 1997. Policy Options Analysis: Nutraceuticals/
Functional Foods. Ottawa, Ont., Canada: Health Canada, Health Protection Branch, Therapeutic Products Programme and Food Directorate.
60. Higuchi M. 1984. The effect of oxygen on the growth and mannitol
fermentation of Streptococcus mutants. J. Gen. Microbiol. 130:1819-1826.

176
61. Hillian M. 1995. Functional foods: current and future market developments. Food Technology International Europe 25–31.
62. Hinz SW, van den Broek LA, Beldman G, Vincken JP, Voragen AG.
2004 beta-Galactosidase from Bifidobacterium adolescentis DSM20083 prefers beta(1,4)-galactosides over lactose. Appl. Microbiol. Biotechnol. 66:276-284.
63. Hold GL, Pryde SE, Russell VJ, Furrie E, Flint HJ. 2002. Assessment
of microbial diversity in human colonic samples by 16S rDNA sequence analysis. FEMS Microbiol. Ecol. 39:33-39.
64. Honda Y, Kitaoka M. 2004. A family 8 glycoside hydrolase from
Bacillus halodurans C-125 (BH2105) is a reducing end xylose-releasing exooligoxylanase. J. Biol. Chem. 279:55097–55103.
65. Hopkins MJ, Cummings JH, MacFarlane GT. 1998. Inter-species
differences in maximum specific growth rates and cell yields of bifidobacteria cultured on oligosaccharides and other simple carbohydrate sources. J. Appl. Microbiol. 85:381-386.
66. Hoverstad T, Bjorneklett A. 1984. Short-chain fatty acids and bowel
functions in man. Scand. J. Gastroenterol. 19:1059-1065.
67. Hoverstad T, Fausa O, Bjorneklett A, Bohmer T. 1984. Short-chain fatty acids in the normal human feces. Scand. J. Gastroenterol. 19:375-381.
68. Hutkins R. 1984. Carbohydrate metabolism in Streptococcus
thermophilus. Ph.D. thesis, University of Minnesota, St. Paul, MN.

177
69. IFIC Foundation. 1995. Functional foods: opening the door to better health. Food Insight November/December.
70. Ito M, Deguchi Y, Miyamori A, Matsumoto K, Kikuchi H,
Kobayashi Y, Yajiama T, Kan T. 1990. Effects of administration of galacto-oligosaccharides on the human faecal microflora, stool weight and abdominal sensation. Microb. Ecol. Health Dis. 3:285-292.
71. Janer C, Rohr LM, Pelaez C, Laloi M, Cleusix V, Requena T, Meile
L. 2004. Hydrolysis of oligofructoses by the recombinant β-fructofuranosidase from Bifidobacterium lactis. System. Appl. Microbiol. 27:279-285.
72. Kaizu H, Sasaki M, Nakajima H, Suzuki Y. 1993. Effect of
antioxidative lactic acid bacteria on rats fed a diet deficient in vitamin E. J. Dairy Sci. 76:2493-2499.
73. Kambourova M, Mandeva R, Fiume I, Maurelli L, Rossi M, Morana
A. 2007. Hydrolysis of xylan at high temperature by co-action of the xylanase from Anoxybacillus flavithermus BC and the beta-xylosidase/alpha-arabinosidase from Sulfolobus solfataricus Oalpha. J. Appl. Microbiol. 102:1586-1593.
74. Kandler O, Weiss N. Genus Lactobacillus. 1986. In, P. H. A. Sneath, N.
S. Mair, M. E. Sharpe, and J. G. Holt (eds.), Bergey's Manual of Systematic Bacteriology, vol 2, 9th ed. Williams and Wilkins, Baltimore. pp. 1063-1065.

178
75. Kaplan H, Hutkins RW. 2000. Fermentation of fructooligosaccharides by lactic acid bacteria and bifidobacteria. Appl. Environ. Microbiol. 66:2682-2684.
76. Kaster AG, Brown LR. 1983. Extracellular dextranase activity
produced by human oral strains of the genus Bifidobacterium. Infect. Immun. 42:716-720.
77. Katayama T, Sakuma A, Kimura T, Makimura Y, Hiratake J,
Sakata K, Yamanoi T, Kumagai H, Yamamoto K. 2004. Molecular cloning and characterization of Bifidobacterium bifidum 1,2-alpha-L-fucosidase (AfcA), a novel inverting glycosidase (glycoside hydrolase family 95). J Bacteriol. 186:4885-4893.
78. Kim YS, Gum JR, Ho SB. 1994. Colonocyte differentiation and
proliferation. In Short Chain Fatty Acids, Kluwer Academic Press, Lancaster, pp119-134.
79. Kim TB, Song SH, Kang SC, Oh DK. 2003. Quantitative comparison
of lactose and glucose utilization in Bifidobacterium longum cultures. Biotechnol. Prog. 19:672-675.
80. Kleessen B, Stoof G, Proll J, Schmiedel D, Noak J, Blaut M. 1997.
Feeding resistant starch affects faecal and cecal microflora and short-chain fatty acids in rats. J. Anim. Sci. 75:2453-2462.
81. Kok FJ. 1999. Functional foods: relevance of genetic susceptibility. In
Proceedings of Forum on Functional Food, pp. 217–229. Strasbourg: Council of Europe Publishers.

179
82. Kolida S, Gibson GR. 2007. Prebiotic capacity of inulin-type fructans. J. Nutr. 137:2503S-2506S.
83. Lagaert S, Van Campenhout S, Pollet A, Bourgois TM, Delcour JA,
Courtin CM, Volckaert G. 2007. Recombinant expression and characterization of a reducing-end xylose-releasing exo-oligoxylanase from Bifidobacterium adolescentis. Appl. Environ. Microbiol. 73:5374-5377.
84. Lama L, Calandrelli V, Gambacorta A, Nicolaus B. 2004. Purification
and characterization of thermostable xylanase and β-xylosidase by the thermophilic bacterium Bacillus thermantarcticus. Res. Microbiol. 155:283–289
85. La Placa, M. 1995. Principi di Microbiologia Medica, Editrice
Esculapio pp233-242.
86. Larentis AL, Almeida MS, Cabral KMS, Medeiros LN, Kurtenbach E, Coelho MAZ. 2004. Expression of Pisum sativum defensin 1 (Psd1) in shaking flasks and bioreactor cultivations of recombinant Pichia pastoris at different pHs Brasilian Journal of chemichal Engineering. Vol 21, n°2 pagg: 1-16.
87. Leder S, Hartmeier W, Marx SP. 1999. α-galactosidase of
Bifidobacterium adolescentis DSM 20083. Curr. Microbiol. 38:101-106.
88. Lee SK, Kim YB, Ji GE. 1997. Note: purification of amylase secreted from Bifidobacterium adolescentis. J. Appl. Microbiol. 83:267-272.

180
89. Livesey G, Elia M. 1995. Short-chain fatty acids as energy source in the colon: metabolism and clinical implications, p. 427. In Cummings JH, Rombeau JL, and Sakata S (ed.), Physiological and clinical aspects of shortchain fatty acids. Cambridge University Press, Cambridge, United Kingdom.
90. Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ. 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem. 193:265–275.
91. MacDonought FE, Wells P, Wong NP. 1983. Mechanism of lactose
utilisation by lactic acid streptococci: enzymatic and genetic analysis. J. Bacteriol. 102:804-809.
92. MacFarlane GT, Gibson GR, Drasar BS, Cummings JH. 1995
Metabolic significance of the gut microflora. In Whitehead R (ed.), Gastrointestinal and Oesophageal Pathology, 2nd ed., pp. 249–274. Churchill Livingstone, Edinburgh, United Kingdom.
93. MacFarlane GT, MacFarlane S. 1995 Proteolysis and amino acid
fermentation. In Gibson GR and MacFarlane GT (eds), Human Colonic Bacteria: Role in Nutrition, Physiology, and Pathology, pp. 75–100. CRC Press, London, United Kingdom.
94. Macfarlane S and Macfarlane GT. 2006. Composition and metabolic
activities of bacterial biofilms colonizing food residues in the human gut. Appl. Environ. Microbiol. 72:6204–6211.
95. Manderson K, Pinart M, Tuohy KM, Grace WE, Hotchkiss AT,
Widmer W, Yadhav MP, Gibson GR, Rastall RA. 2005. In vitro determination of prebiotic properties of oligosaccharides derived from an

181
orange juice manufacturing by-product stream. Appl. Environ. Microbiol. 71:8383-8389.
96. Margolles A, de los Reyes-Galivàn CG. 2003. Purification and
functional characterization of a novel α-L-Arabinofuranosidase from Bifidobacterium longum B667. Appl. Environ. Microbiol. 69:5096-5103.
97. Marteau P, Minekus M, Havenaar R, Huis in't Veld JH. 1997.
Survival of lactic acid bacteria in a dynamic model of the stomach and small intestine: validation and the effects of bile. J. Dairy Sci. 80:1031-1037.
98. Masco L, Crockaert C, Van Hoorde K, Swings J, Huys G. 2007. In
vitro assessment of the gastrointestinal transit tolerance of taxonomic reference strains from human origin and probiotic product isolates of Bifidobacterium. J. Dairy Sci. 90:3572-3578.
99. Matteuzzi D, Crociani F, Emaldi O. 1978. Aminoacids produced by
bifidobacteria and some clostridia. Ann. Microbiol. Inst. Pasteur 129:175-181.
100. Matteuzzi D, Swennen E, Rossi M, Hartman T, Lebet V. 2004.
Prebiotic effects of a wheat germ preparation in human healthy subjects. Food. Microbiol. 21:119-124.
101. McKellar RC, Modler HW. 1989. Metabolism of
fructooligasaccharides by Bifidobacterium spp. Appl. Microbiol. Biotechnol. 31:537-541.

182
102. McWilliam Leitch EC, Walker AW, Duncan SH, Holtrop G, Flint HJ. 2007. Selective colonization of insoluble substrates by human faecal bacteria. Environ. Microbiol. 9:667–679.
103. Mlobeli NT, Gutierrez NA, Maddox IS. 1998. Physiology and kinetics
of Bifidobacterium bifidum during growth on different sugars. Appl. Microbiol. Biotechnol. 50:125-128.
104. Moller PL, Jorgensen F, Hansen OC, Madsen SM, Stougaard P.
2001. Intra- and extracellular beta-galactosidases from Bifidobacterium bifidum and B. infantis: molecular cloning, heterologous expression, and comparative characterization. Appl. Environ. Microbiol. 67:2276-2283.
105. Morrison DJ, Mackay WG, Edwards CA, Preston T, Dodson B,
Weaver LT. 2006. Butyrate production from oligofructose fermentation by the human faecal flora: what is the contribution of extracellular acetate and lactate? Br. J. Nutr. 96:570–577.
106. Murphy O. 2001 Non-polyol low-digestible carbohydrates: food
applications and functional benefits. Br. J. Nutr. 85:47-53. 107. Olano-Martin E, Gibson GR, Rastall RA. 2002. Comparison of the in
vitro bifidogenic properties of pectins and pectic-oligosaccharides. J. Appl. Microbiol. 93:505-511.
108. Okhusa T, Ozaki Y, Sato C. 1995. Long-term ingestion of lactosucrose
increases Bifidobacterium sp. In human vecal flora. Digest. 56:415-420.

183
109. Palframan RJ, Gibson GR, Rastall RA. 2003. Carbohydrate preferences of Bifidobacterium species isolated from the human gut. Curr. Iss. Intest. Microbiol. 4:71-75.
110. Perrin S, Warchol M, Grill JP, Schneider F. 2001. Fermentation of
fructooligosaccharides and their components by Bifidobacterium infantis ATCC 15697 on batch culture in semi-synthetic medium. J. Appl. Microbiol. 90:859-865.
111. Pompei A, Cordisco L, Amaretti A, Zanoni S, Matteuzzi D, Rossi M.
2007. Folate production by bifidobacteria as a potential probiotic property. Appl. Environ. Microbiol. 73:179-185.
112. Pompei A, Cordisco L, Amaretti A, Zanoni S, Raimondi S, Matteuzzi
D, Rossi M. 2007. Administration of folate-producing bifidobacteria enhances folate status in Wistar rats. J. Nutr. 137:2742-2746.
113. Rabiu BA, Jay AJ, Gibson GR, Rastall RA. 2001 Synthesis and
fermentation properties of novel galacto-oligosaccharides by β-galactosidases from Bifidobacterium species. Appl. Environ. Microbiol. 67:2526-2530.
114. Rada V, Bartonova J, Vlkova E. 2002. Specific growth rate of
bifidobacteria cultured on different sugars. Folia Microbiol. 47:477-480.
115. Rastall RA, Maitin V. 2002. Prebiotics and synbiotics: towards the next generation. Curr. Opin. Biotechnol. 13:490-496.
116. Riemersma RA. 1996. A fat little earner. Lancet 347:775–776.

184
117. Roberfroid MB. 1998. Prebiotics and synbiotics: concepts and nutritional properties. Br. J. Nutr. 80:197-202.
118. Roberfroid MB. 2000. Defining functional foods. In Functional Foods,
Concept to Product, pp. 9–28. GR Gibson and CM Williams, editors. Cambridge, UK: Woodhead Publishing Ltd.
119. Roberfroid MB. 2002. Global view on functional foods: European
perspectives. Br. J. Nutr. 88:S133-138.
120. Roberfroid M. 2007. Prebiotics: the concept revisited. J. Nutr. 137:830S-837S.
121. Robert C and Bernalier-Donadille A. 2003. The cellulolytic microflora
of the human colon: evidence of microcrystalline cellulose-degrading bacteria in methane excreting subjects. FEMS Microbiol. Ecol. 46:81–89.
122. Ross EW, Taub IA, Doona CJ, Feeherry FE, Kustin K. 2004. The
mathematical properties of the quasi-chemical model for microorganism growth- death kinetics in foods . Int. J. of food Microbiol. 99:157-171.
123. Rossi M, Corradini C, Amaretti A, Nicolini M, Pompei A, Zanoni S,
Matteuzzi D. 2005. Fermentation of fructooligosaccharides and inulin by bifidobacteria: a comparative study of pure and fecal cultures. Appl. Environ. Microbiol. 71:6150-6158.
124. Ruppin H, Bar-Meir S, Soergel HK, Wood CM, Schmitt MG. 1980.
Absorption of short-chain fatty acids by the colon. Gastroenterol. 78:1500-1507.

185
125. Rycroft CE, Jones MR, Gibson GR, Rastall RA. 2001 A comparative in vitro evaluation of the fermentation properties of prebiotic oligosaccharides. J. Appl. Microbiol. 91:878-887.
126. Saarela M, Virkajärvi I, Nohynek L, Vaari A, Mättö J. 2006. Fibres
as carriers for Lactobacillus rhamnosus during freeze-drying and storage in apple juice and chocolate-coated breakfast cereals. Int. J. Food Microbiol. 112:171-178.
127. Salyers AA, West SE, Vercellotti JR, Wilkins TD. 1977. Fermentation
of mucins and plant polysaccharides by anaerobic bacteria from the human colon. Appl. Environ. Microbiol. 34:529-533.
128. Salminen S, Bouley C, Boutron-Ruault MC, Cummings JH, Franck
A, Gibson GR, Isolauri E, Moreau MC, Roberfroid M, Rowland I. 1998 Functional food science and gastrointestinal physiology and function. Br. J. Nutr. 80:S147-S171.
129. Saulnier DM, Molenaar D, de Vos WM, Gibson GR, Kolida S. 2007.
Identification of prebiotic fructooligosaccharide metabolism in Lactobacillus plantarum WCFS1 through microarrays. Appl. Environ. Microbiol. 73:1753-1765.
130. Savage DC. 1979. Introduction to mechanism of indigenous microbes.
Am. J. Clin. Nutr. 32:113-118.
131. Scardovi V. 1986. Genus Bifidobacterium. In Sneath H, Mair N, Sharpe M, Holt J (eds), Bergey’s Manual of Systematyc Bacteriology, vol 2, 9th edn. pp. 1418-1434. Williams & Wilkins, Baltimore.

186
132. Schell MA, Karmirantzou M, Snel B, Vilanova D, Berger B, Pessi G, Zwahlen MC, Desiere F, Bork P, Delley M, Pridmore RD, Arigoni F. 2002. The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Microbiol. 99:14422-14427.
133. Schlegel HG. 1976. Microbiologia. Ed. Zanichelli, Bologna.
134. Schleifer KH, Kilpper-Bälz R. 1987. Molecular and chemotaxonomic
approaches to the classification of Streptococci, Enterococci and Lactococci: a review. Syst. Appl. Microbiol. 10:1-19.
135. Schroeder CJ, Robert C, Lenzen G, McKay LL, Mercenier A. 1991.
Analysis of the lacZ gene sequences from two Streptococcus thermophilus strains: comparison to Escherichia coli and Lactobacillus bulgaricus beta galactosidase sequences. J. Gen. Microbiol. 137:369-380.
136. Shene C, Mardones M, Zamora P, Bravo S. 2005. Kinetics of
Bifidobacterium longum ATCC 15707 fermetation: effect of the diluition rate and carbon sources. Appl. Microbiol. Biotechnol. 67:623-630.
137. Shin HY, Lee JH, Lee JY, Han YO, Han MJ, Kim DH. 2003.
Purification and characterization of ginsenoside Ra-hydrolyzing β-D-xylosidase from Bifidobacterium breve K-110, a human intestinal anaerobic bacterium. Biol. Pharm. Bull. 26:1170–1173.
138. Slovakova L, Duskova D, Marounek M. 2002. Fermentation of pectin
and glucose, and activity of pectin-degrading enzymes in the rabbit

187
caecal bacterium Bifidobacterium pseudolongum. Lett. Appl. Microbiol. 35:126-130.
139. Smith BL, Marcotte M, Harman G. 1996. A Comparative Analysis of
the Regulatory Framework Affecting Functional Food Development and Commercialization in Canada, Japan, the European Union and the United States of America. Ottawa, Ont., Canada: Intersector Alliance Inc.
140. Sprogoe D, Van Den Broek L, Mirza Osman, Kastrup JS, Voragen
AGJ, Gajhede M, Skov LK. 2004. Crystal structure of sucrose phosphorylase from Bifidobacterium adolescentis. Biochem. 43:1156-1162.
141. Tannock GV. 1999. Probiotics. A critical review. Horizon Scientific
Press, Norfolk, UK.
142. Tannock GV. 2002. Probiotics and Prebiotics. Where are we going? Caister Academic Press, Norfolk, UK.
143. Teuber M. 1993. Lactic Acid Bacteria. In Sahm H. (ed), Biological
Fundamentals, vol. 1, pp. 325-366
144. Tuncer M, Ball AS. 2003. Co-operative actions and degradation analysis of purified xylan-degrading enzymes from Thermomonospora fusca BD25 on oat-spelt xylan. J. Appl. Microbiol. 94:1030-1035.
145. van den Broek L, Struijs K, Verdoes J, Beldman G, Voragen A.
2003. Cloning and characterization of two α-glucosidases from Bifidobacterium adolescentis DSM20083. Appl. Microbiol. Biotechnol. 61:55-60.

188
146. van den Broek LAM, Lloyd RM, Beldman G, Verdoes JC, McCleary BV, Voragen AGJ. 2005. Cloning and characterization of arabinoxylan arabinofuranohydrolase-D3 (AXHd3) from Bifidobacterium adolescentis DSM20083. Appl. Microbiol. Biotechnol. 67:641–647.
147. van Laere KMJ, Hartemink R, Beldman G, Pitson S, Dijema C,
Schols HA, Voragen AGJ. 1999. Transglycosidase activity of Bifidobacterium adolescentis DSM 20083 α-galactosidase. Appl. Microbiol. Biotechnol. 52:681-688.
148. Vernazza CL, Gibson GR, Rastall RA. 2006. Carbohydrate preference,
acid tolerance and bile tolerance in five strains of Bifidobacterium. J. Appl. Microbiol. 100:846-853.
149. Wang X, Conway PL, Brown IL, Evans AJ. 1999. In vitro utilization
of amylopectin and high-amylose maize (Amylomaize) starch granules by human colonic bacteria. Appl. Environ. Microbiol. 65:4848-4854.
150. Wang X, Gibson GR. 1993. Effects of the in vitro fermentation of
oligofructose and inulin by bacteria growing in the human large intestine. J Appl. Bacteriol. 75:373-380.
151. Warchol M, Perrin S, Grill JP, Schneider F. 2002. Characterization of
a purified β-fructofuranosidase from Bifidobacterium infantis ATCC 15697. Lett. Appl. Microbiol. 35:462-467.
152. Zeng H, Xue Y, Peng T, Shao W. 2007. Properties of xylanolytic
enzyme system in bifidobacteria and their effects on the utilization of xylooligosaccharides. Food Chemistry. 101:1172-1177


















