La FOTOSINTESI rappresenta la - uniroma2.it · I cromofori assorbono la luce di una determinata...
57
La FOTOSINTESI rappresenta la • Fonte principale di carboidrati • Via principale di fissazione del carbonio • Fonte di ossigeno dell’atmosfera Organismi fotosintetici (3.5 miliardi di anni fa): piante, alghe, microrganismi.
-
Upload
truongkhue -
Category
Documents
-
view
214 -
download
1
Transcript of La FOTOSINTESI rappresenta la - uniroma2.it · I cromofori assorbono la luce di una determinata...

La FOTOSINTESI rappresenta la
• Fonte principale di carboidrati
• Via principale di fissazione del carbonio
• Fonte di ossigeno dell’atmosfera
Organismi fotosintetici (3.5 miliardi di anni fa):
piante, alghe, microrganismi.

La CO2 viene ridotta
dagli autotrofi, mentre i
carboidrati (glucosio)
vengono ossidati dagli
eterotrofi.
Nessuno degli atomi di
ossigeno usato per
formare i carboidrati
proviene dalla CO2
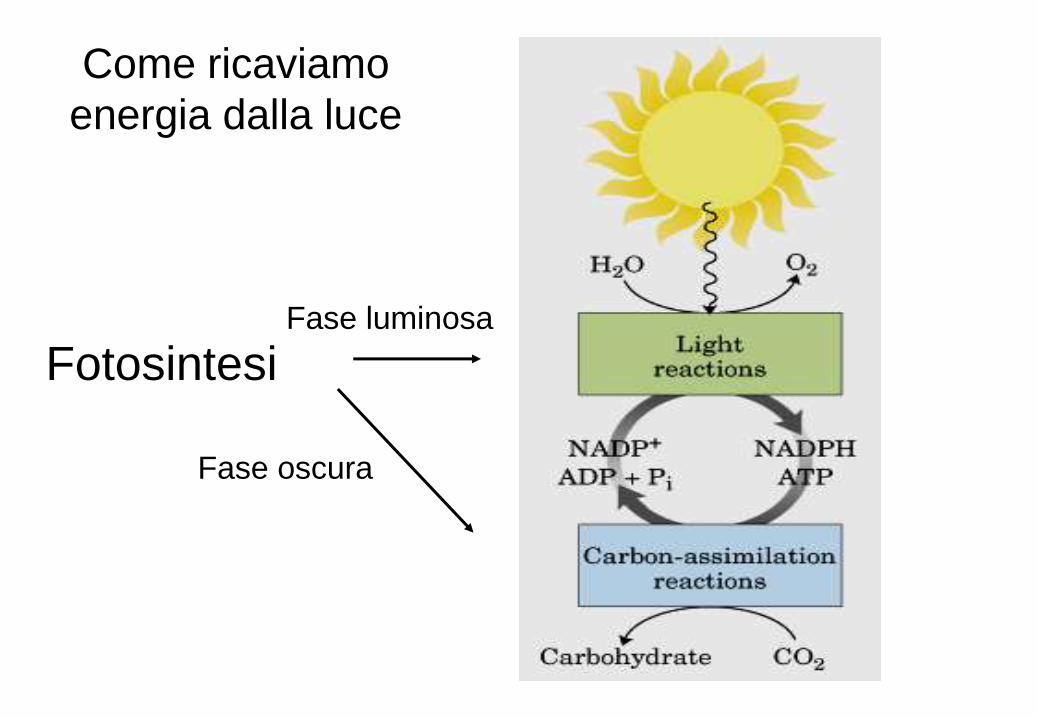
Fotosintesi Fase luminosa
Fase oscura
Come ricaviamo
energia dalla luce

La fase luminosa avviene
sulla membrana e all’interno
del tilacoide, la fase oscura
nello stroma. La sintesi di
ATP avviene analogamente
alla fosforilazione ossidativa
mitocondriale, secondo un
processo chemiosmotico.

NATURA CORPUSCOLARE
fotone
quanto (unità di energia)
E = h E = h c/
(equazione di Planck)
1 mole di fotoni = 1 Einstein
COME SI OTTIENE ENERGIA DALLA LUCE?
NATURA ONDULATORIA
lunghezza d’onda ()
frequenza ()
= c /

La fotosintesi dipende principalmente dalla luce delle regioni dello
spettro del visibile e del vicino infrarosso. Le microonde (lontano
IR) producono solo vibrazioni molecolari, hanno bassa energia.
Le radiazioni UV hanno alta energia, possono rompere legami
covalenti (ma le blocca l’ozono).

Clorofilla a e b sono simili alle protoporfirine (con Mg). Assorbono la luce nel visibile perché hanno sistemi di doppi legami coniugati. Tutti i cromofori si trovano all’interno delle membrane dei tilacoidi. La ficocianina è legata ad una proteina; le clorofille prendono contatto con i lipidi di membrana mediante il fitolo idrofobico. Le ficocianine sono presenti nelle alghe. Sono tetrapirroli lineari. L’insieme dei pigmenti e le proteine a loro associate prendono il nome di FOTOSISTEMI.
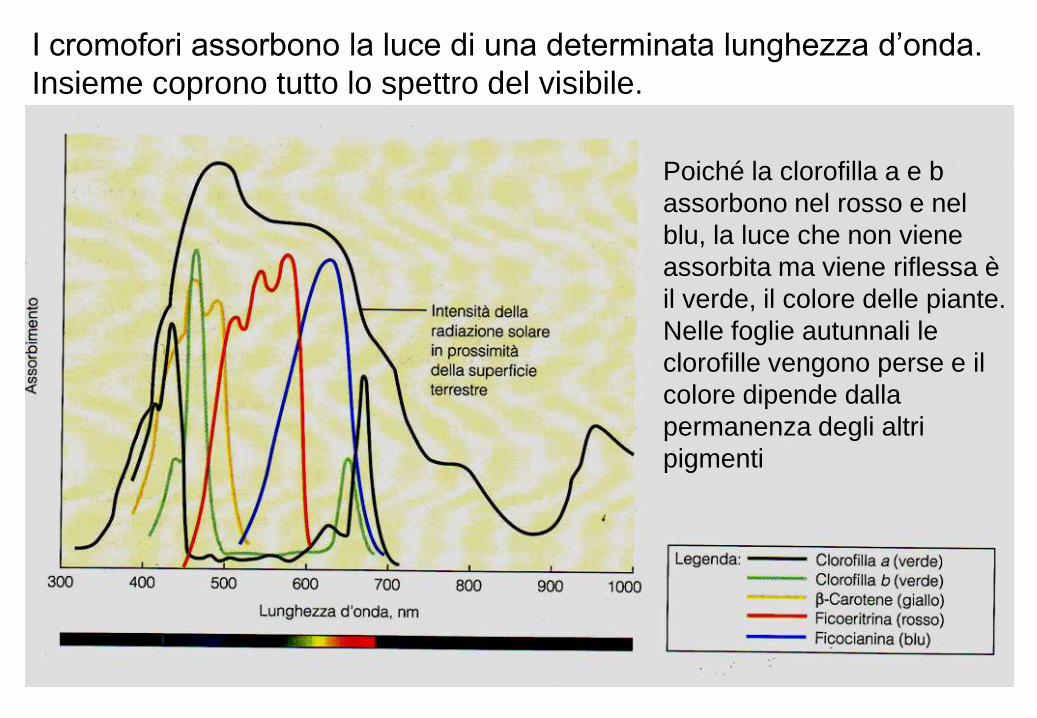
I cromofori assorbono la luce di una determinata lunghezza d’onda.
Insieme coprono tutto lo spettro del visibile.
Poiché la clorofilla a e b
assorbono nel rosso e nel
blu, la luce che non viene
assorbita ma viene riflessa è
il verde, il colore delle piante.
Nelle foglie autunnali le
clorofille vengono perse e il
colore dipende dalla
permanenza degli altri
pigmenti

STATI ELETTRONICI DELLA CLOROFILLA
Una molecola può assorbire fotoni solo di una certa lunghezza d’onda, che abbiano energia sufficiente a far salire l’elettrone da un livello energetico all’altro. L’elettrone che viene eccitato occupa un orbitale π nel sistema dei doppi legami coniugati.
Una volta eccitata la molecola può rilasciare la sua energia in vari modi: calore (conversione interna), fluorescenza (emettendo luce di lunghezza d’onda superiore, quindi di energia minore).

Nei fotosistemi ci sono
COMPLESSI ANTENNA e
CENTRI DI REAZIONE.
L’energia luminosa viene
trasferita tra i pigmenti antenna
mediante trasferimento di
eccitoni per risonanza, fino ad
incontrare il centro di reazione
dove una clorofilla perde un
elettrone perchè lo cede ad una
accettrice che ha uno stato di
eccitazione più basso. Mediante
trasferimento elettronico, una
si ossida, l’altra si riduce e
l’energia viene intrappolata e
trasformata in energia chimica.
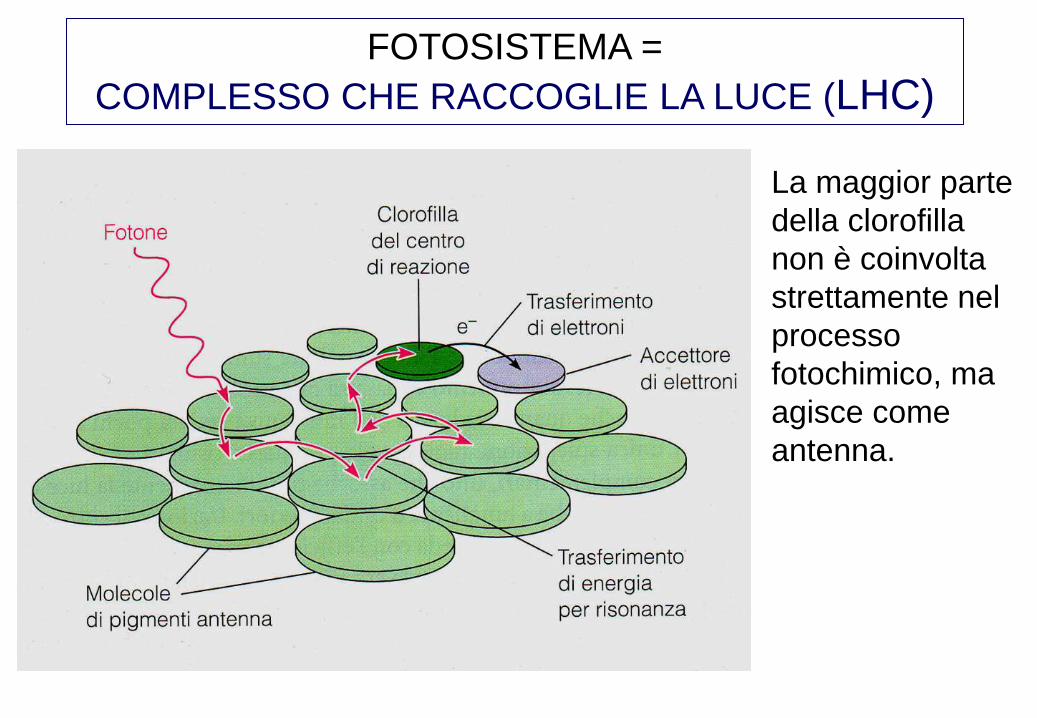
FOTOSISTEMA =
COMPLESSO CHE RACCOGLIE LA LUCE (LHC)
La maggior parte
della clorofilla
non è coinvolta
strettamente nel
processo
fotochimico, ma
agisce come
antenna.
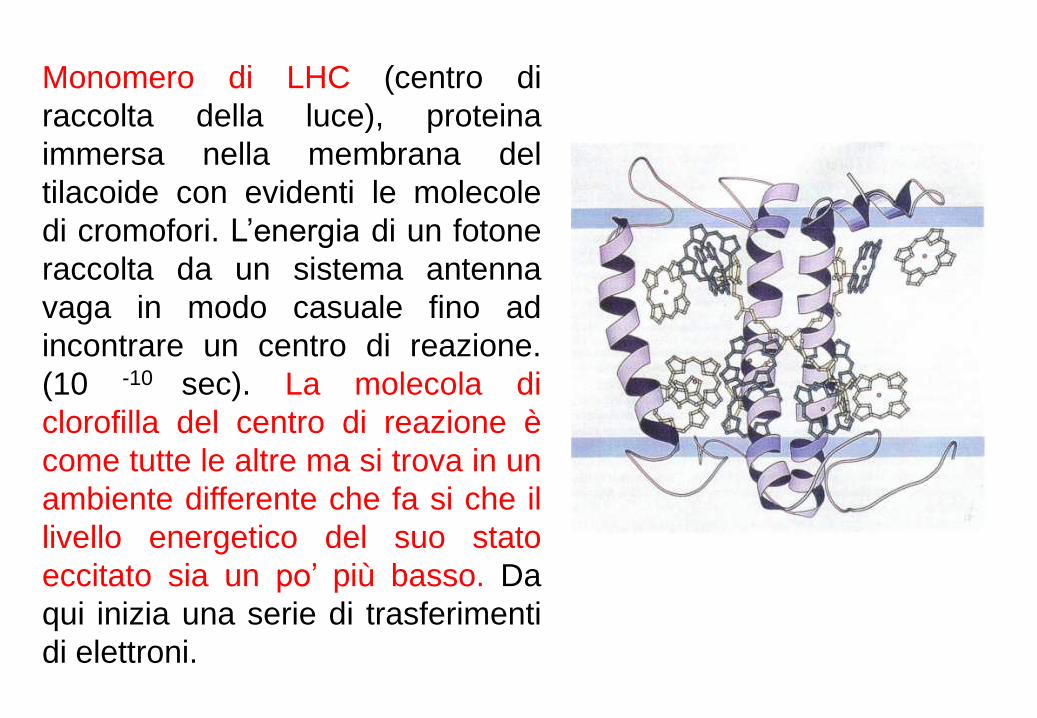
Monomero di LHC (centro di
raccolta della luce), proteina
immersa nella membrana del
tilacoide con evidenti le molecole
di cromofori. L’energia di un fotone
raccolta da un sistema antenna
vaga in modo casuale fino ad
incontrare un centro di reazione.
(10 -10 sec). La molecola di
clorofilla del centro di reazione è
come tutte le altre ma si trova in un
ambiente differente che fa si che il
livello energetico del suo stato
eccitato sia un po’ più basso. Da
qui inizia una serie di trasferimenti
di elettroni.
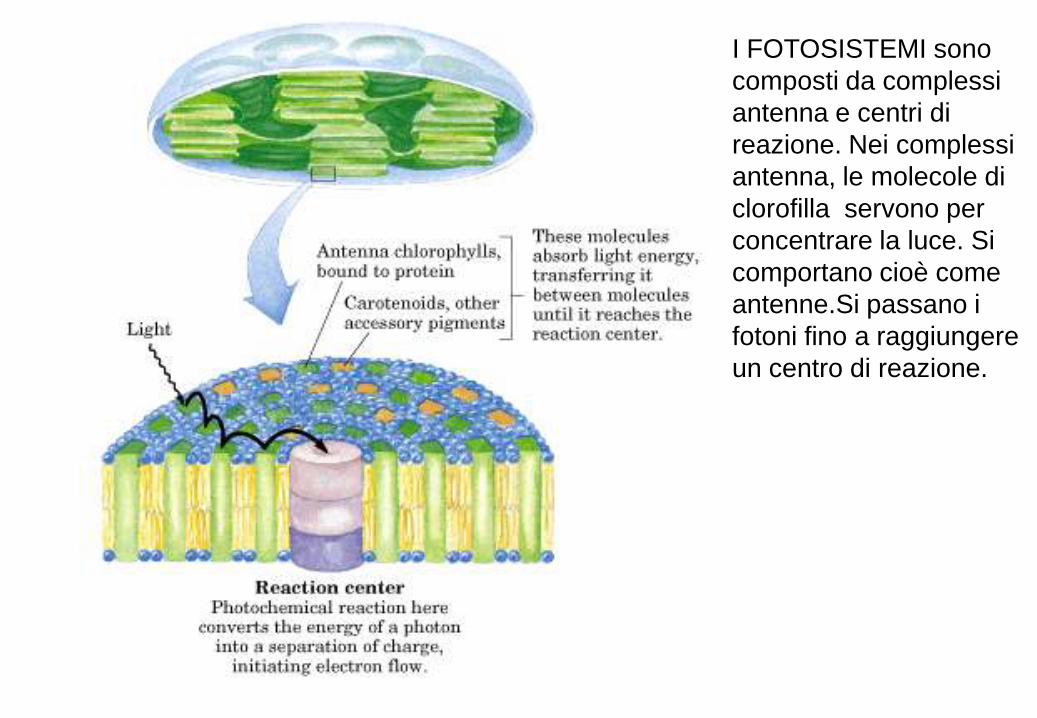
I FOTOSISTEMI sono
composti da complessi
antenna e centri di
reazione. Nei complessi
antenna, le molecole di
clorofilla servono per
concentrare la luce. Si
comportano cioè come
antenne.Si passano i
fotoni fino a raggiungere
un centro di reazione.

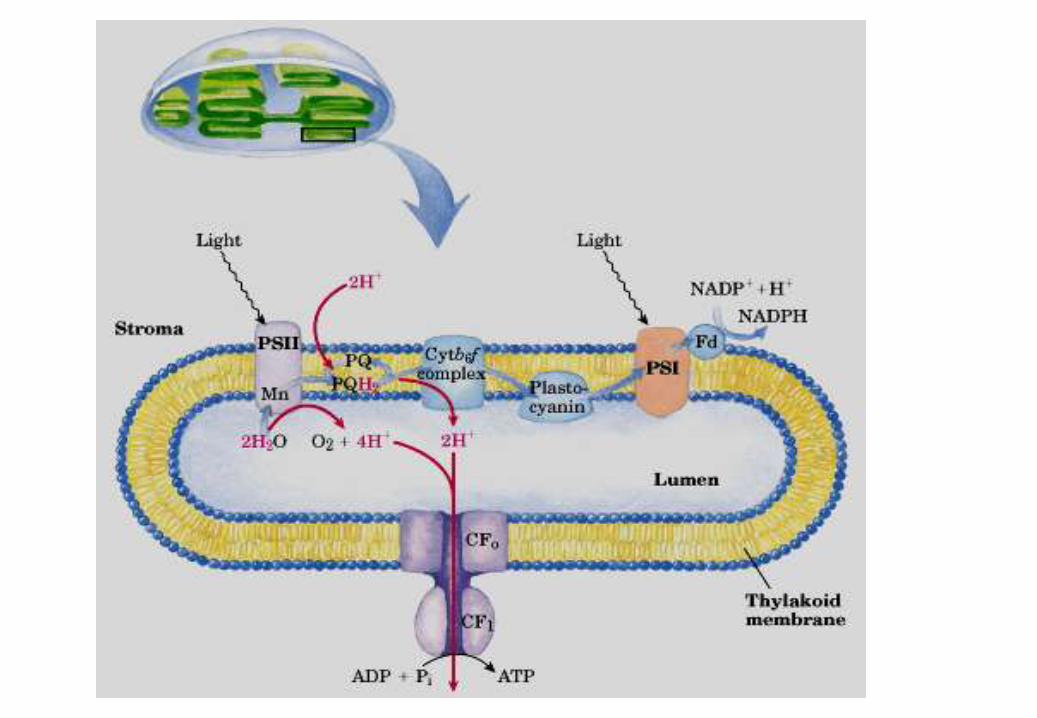
FOTOSISTEMA II
Fotolisi (ossidazione) dell’H2O
evoluzione di O2
formazione del gradiente protonico
FOTOSISTEMA I
produzione di NADPH
PS II e il PS I sono due centri di reazione.
Agiscono in serie
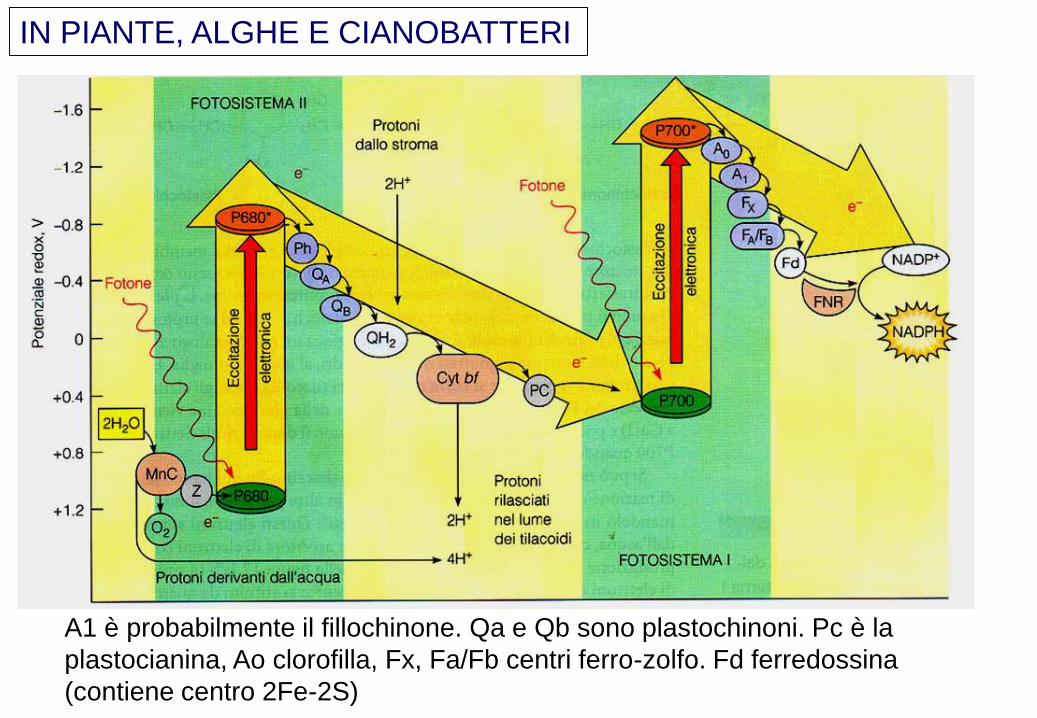
IN PIANTE, ALGHE E CIANOBATTERI
A1 è probabilmente il fillochinone. Qa e Qb sono plastochinoni. Pc è la
plastocianina, Ao clorofilla, Fx, Fa/Fb centri ferro-zolfo. Fd ferredossina
(contiene centro 2Fe-2S)

CENTRI REDOX IMPLICATI NELLA FASE LUMINOSA
Clorofille: P680, P700, feofitina, A0
Chinoni: plastochinone, fillochinone
Citocromi e centri ferro-zolfo
Plastocianina
Ferredossina

Nel PS I
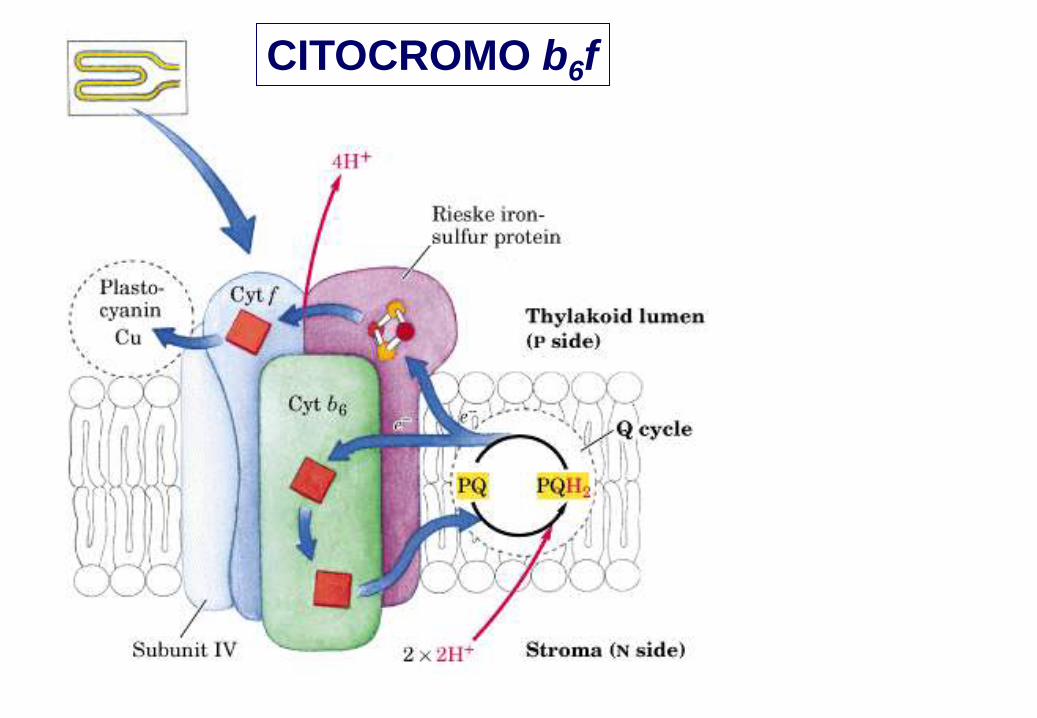
CITOCROMO b6f
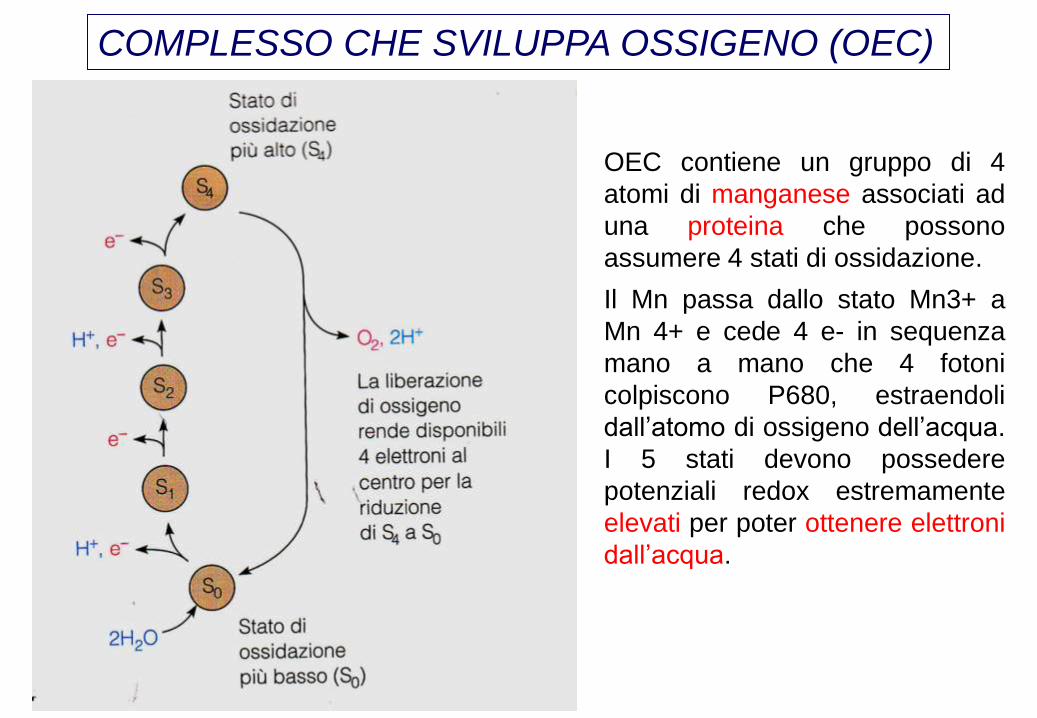
COMPLESSO CHE SVILUPPA OSSIGENO (OEC)
OEC contiene un gruppo di 4
atomi di manganese associati ad
una proteina che possono
assumere 4 stati di ossidazione.
Il Mn passa dallo stato Mn3+ a
Mn 4+ e cede 4 e- in sequenza
mano a mano che 4 fotoni
colpiscono P680, estraendoli
dall’atomo di ossigeno dell’acqua.
I 5 stati devono possedere
potenziali redox estremamente
elevati per poter ottenere elettroni
dall’acqua.

COMPLESSO CHE SVILUPPA OSSIGENO (OEC)
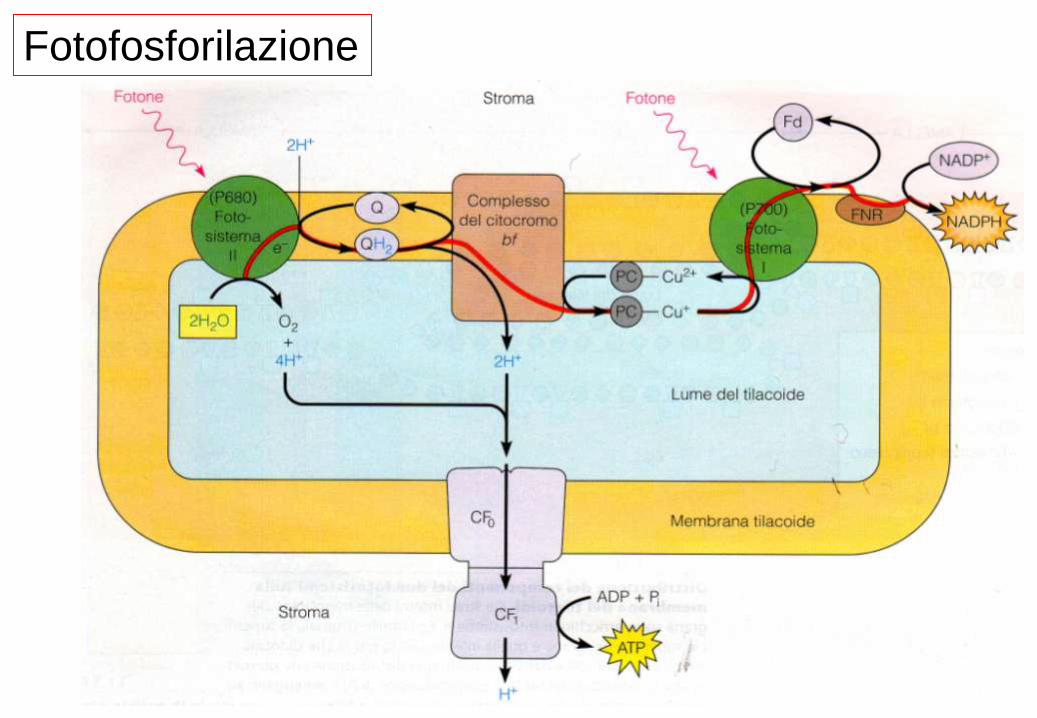
Fotofosforilazione

• I due centri di reazione PS II e il PS I agiscono in serie
in modo da accoppiare l’ossidazione di acqua alla
riduzione di NADPH.
• Il PS II estrae elettroni dall’acqua e li fornisce al PS I.
• Gli elettroni viaggiano da potenziali redox minori verso
potenziali redox maggiori.
• Il P680 ossidato formato in seguito all’eccitazione
luminosa è tra i più potenti ossidanti conosciuti, in grado
di estrarre elettroni dall’H2O tramite il sistema OEC .
• Il P700 eccitato è un debole ossidante e accetta
elettroni dalla plastocianina.

Per ogni elettrone che passa nel PSII vengono traslocati 2 H+. Poiché dall’ H2O provengono 4 elettroni, per ogni molecola di acqua ossidata si
producono 8 H+ che vanno sommati ai 4 H+ provenienti dall’H2O stessa. In totale 12 H+.
Nel trasporto non ciclico si generano 1 ATP per
ogni 3 H+ che passano dal lume del tilacoidi allo
stroma. Quindi per ogni molecola di ossigeno prodotta si generano 4 ATP.

Nei mitocondri i protoni vengono traslocati dalla pompa dallo spazio intermembrana
verso la matrice. Nei tilacoidi i protoni vengono traslocati dal lume del tilacoide verso
lo stroma

TRASPORTO CICLICO DEGLI ELETTRONI
Utilizza il PSI, il complesso b6f e la plastocianina.
Funziona quando il livello di NADP+ è basso, cioè quando NADPH è sufficiente.
La ferredossina passa gli e- al b6f e così via. In questo modo si produce più ATP, perché si liberano più protoni.
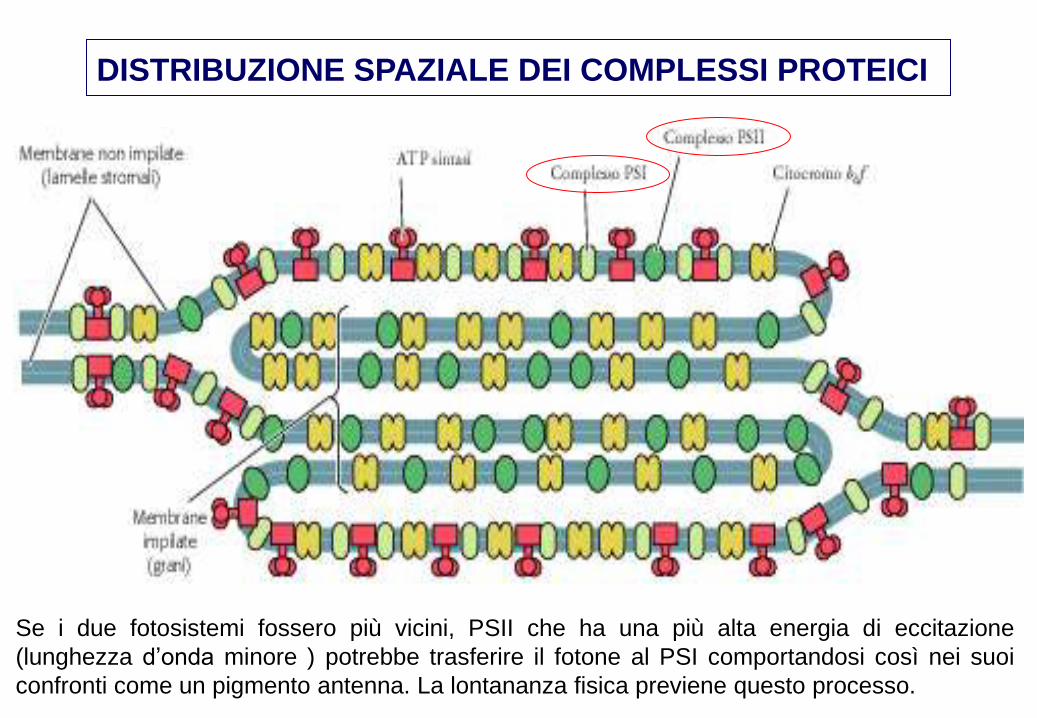
DISTRIBUZIONE SPAZIALE DEI COMPLESSI PROTEICI
Se i due fotosistemi fossero più vicini, PSII che ha una più alta energia di eccitazione
(lunghezza d’onda minore ) potrebbe trasferire il fotone al PSI comportandosi così nei suoi
confronti come un pigmento antenna. La lontananza fisica previene questo processo.

• Due fotosistemi in serie sono operanti nelle alghe fotosintetiche e nelle piante.
• Localizzati nella membrana dei tilacoidi.
• Complesso proteico transmembrana, costituito da pigmenti antenna, centro di reazione
e i trasportatori di elettroni.
• L’evento fondamentale consiste nel trasferimento di un elettrone eccitato.
• Tre complessi proteici PSII, citocromo b6f e PSI, collegati da plastochinone e
plastocianina.
• Ciascuno dei fotosistemi è una catena di trasporto di elettroni, in cui avvengono una
serie di ossidoriduzioni.
• La fonte ultima di elettroni è la molecola d’acqua, l’accettore terminale il NADP+.
• Protoni vengono rilasciati nel lume del tilacoide in due punti.
• Si forma quindi un gradiente protonico, che produrrà ATP.
• ATP e NADPH serviranno a produrre carboidrati nella fase “oscura”.
La fase “luminosa” della fotosisntesi
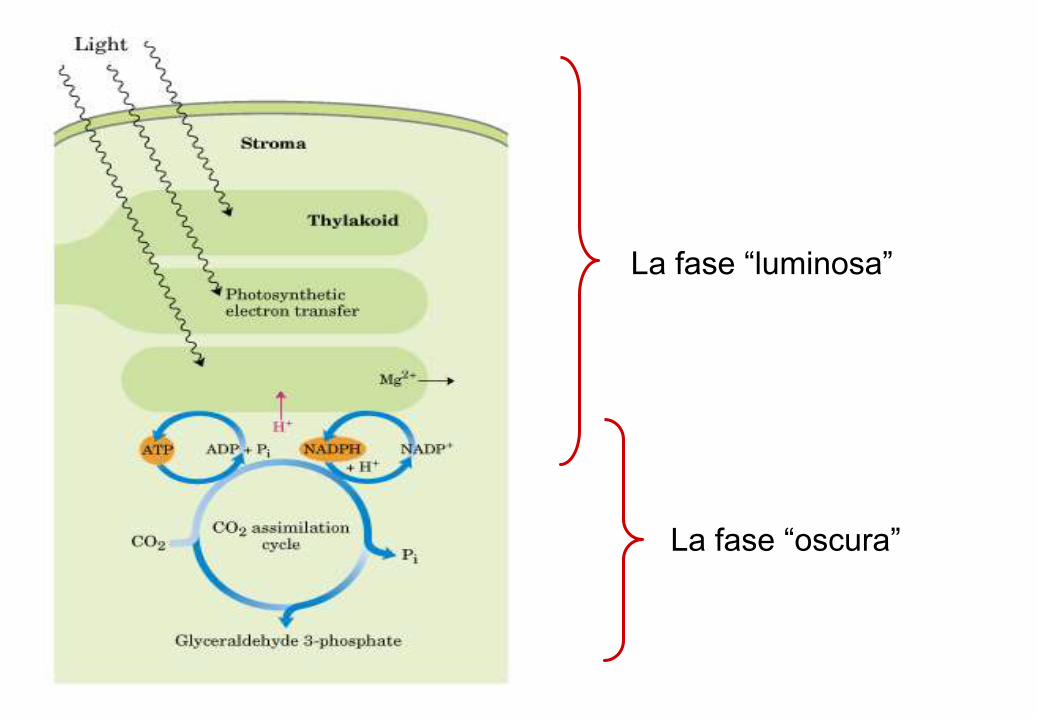
La fase “oscura”
La fase “luminosa”

CICLO DI CALVIN M.Calvin, J.Bassham, A.Besson 1953
14CO2
14CO2
14CO2
14CO2 14CO2
Alga fotosintetica
14CO2
14CO2
14CO2
dopo 1 hr dopo 5 sec
Miscela di metaboliti complessi :
Aminoacidi, zuccheri
3-fosfoglicerato

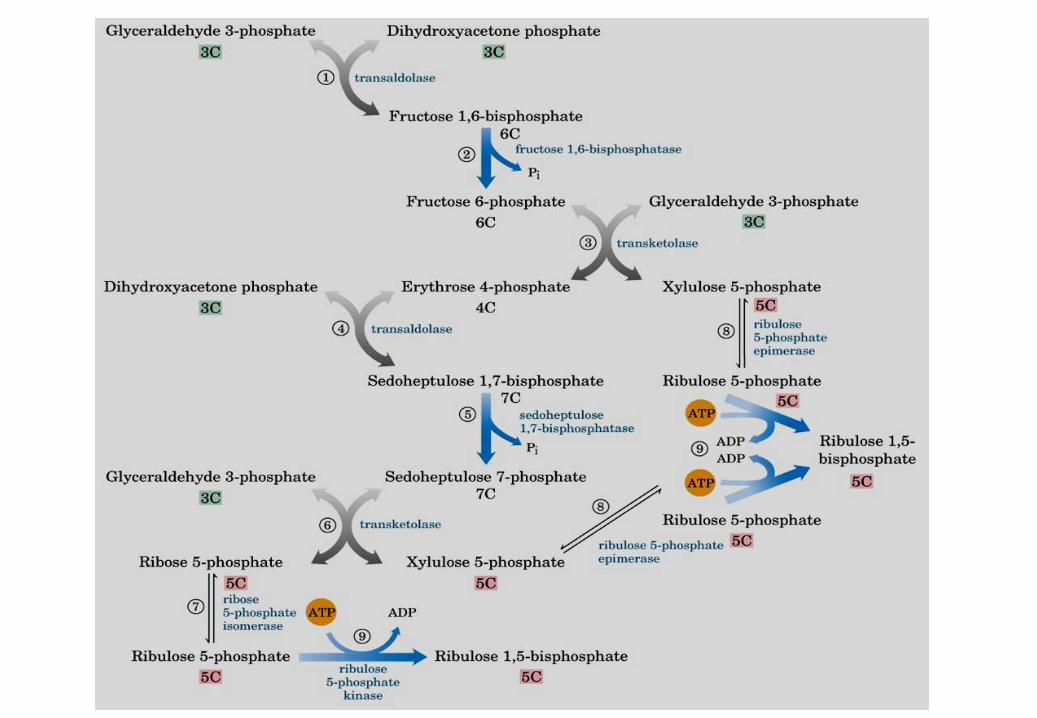
CICLO DI CALVIN
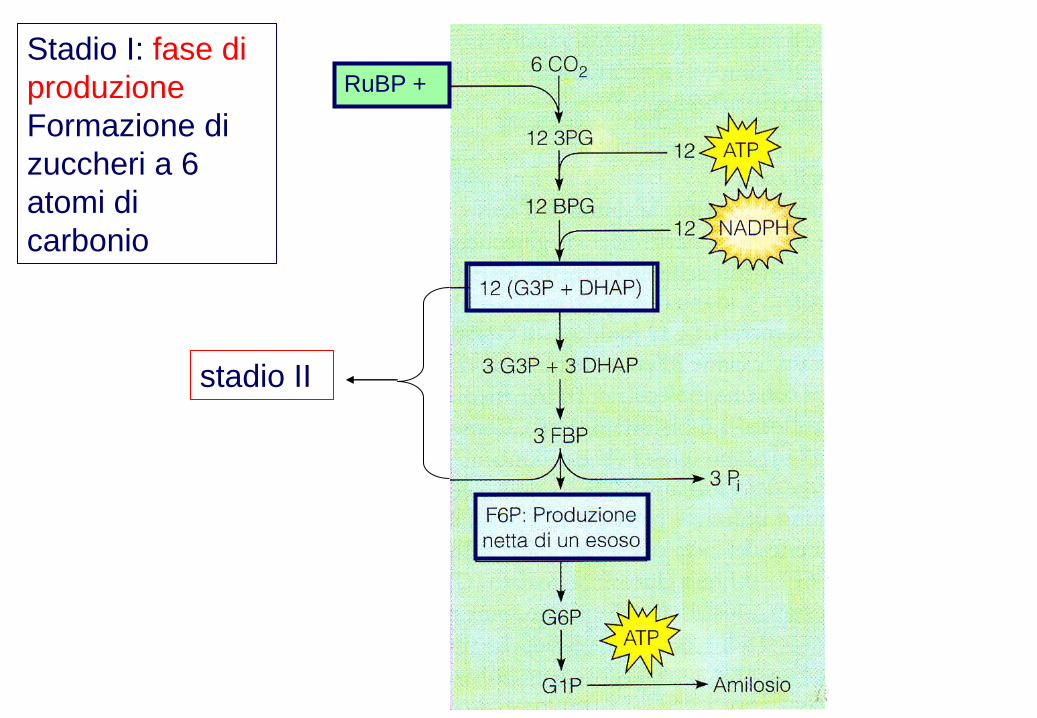
Stadio I: fase di
produzione
Formazione di
zuccheri a 6
atomi di
carbonio
RuBP +
stadio II
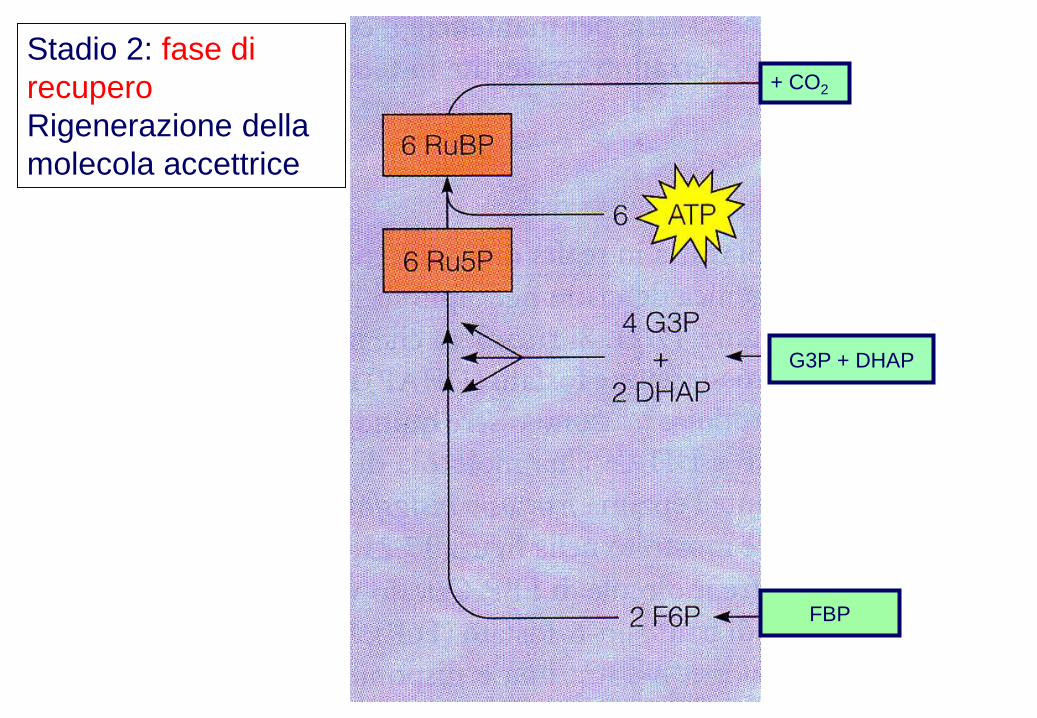
Stadio 2: fase di
recupero
Rigenerazione della
molecola accettrice
+ CO2
G3P + DHAP
FBP
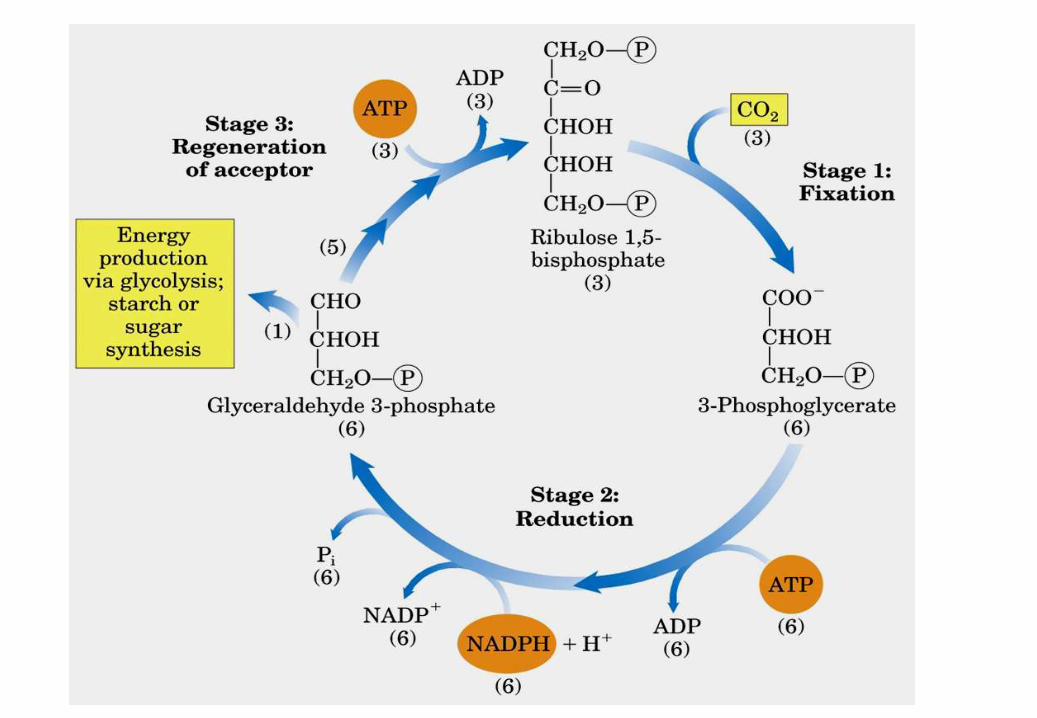
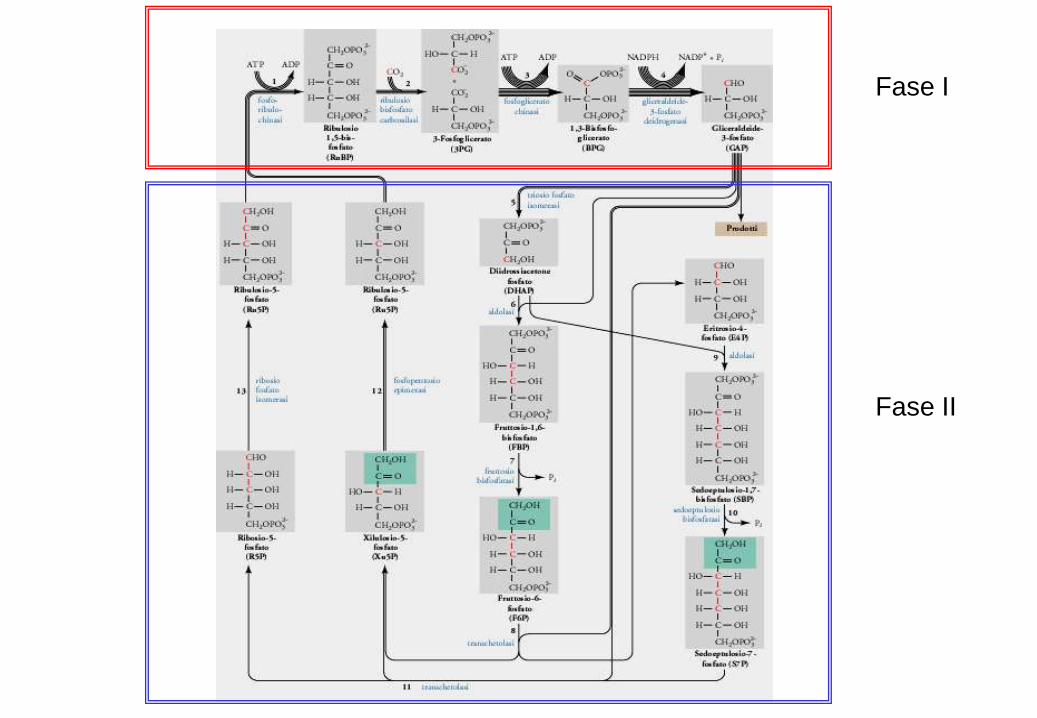
Fase I
Fase II

Ru5P
DHAP Altri prodotti
chinasi carbossilasi chinasi deidrogenasi
isomerasi

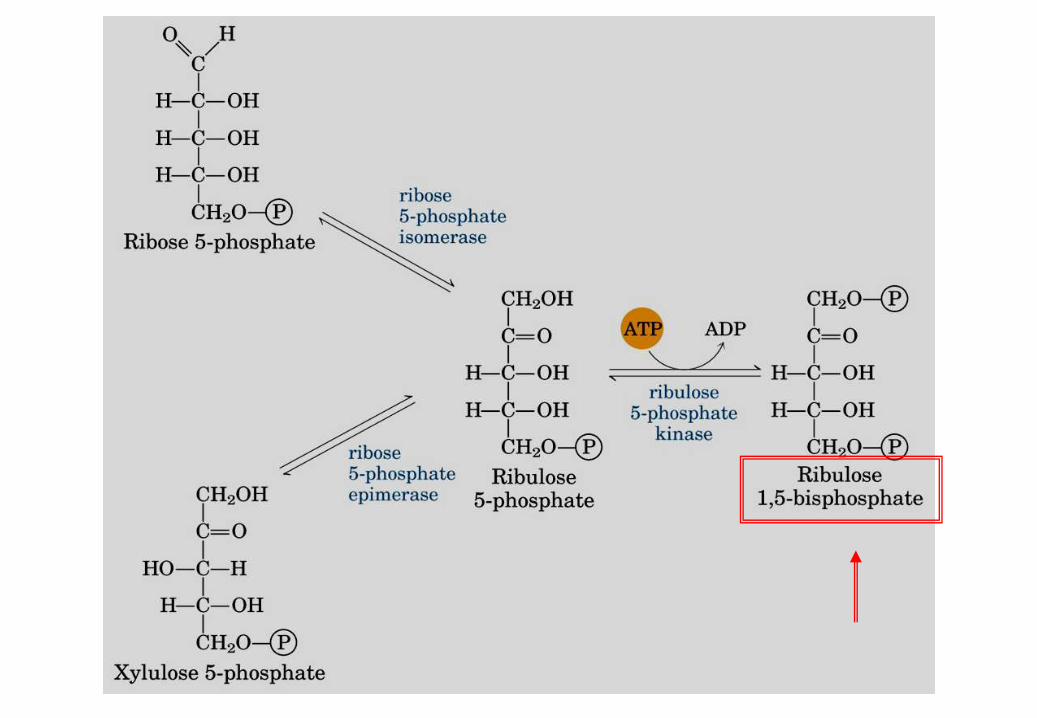
1. Reazione: Condensazione del ribulosio 1,5-bifosfato + CO2

La Ribulosio bisfosfato carbossilasi
(RUBISCO) è un complesso enzimatico.
La reazione dipende da Mg2+ nel sito attivo ed è
favorita da pH alcalino.
La concentrazione di Mg2+ e il pH sono
aumentati dalla luce


Il Ribulosio bisfosfato si lega alle catene L


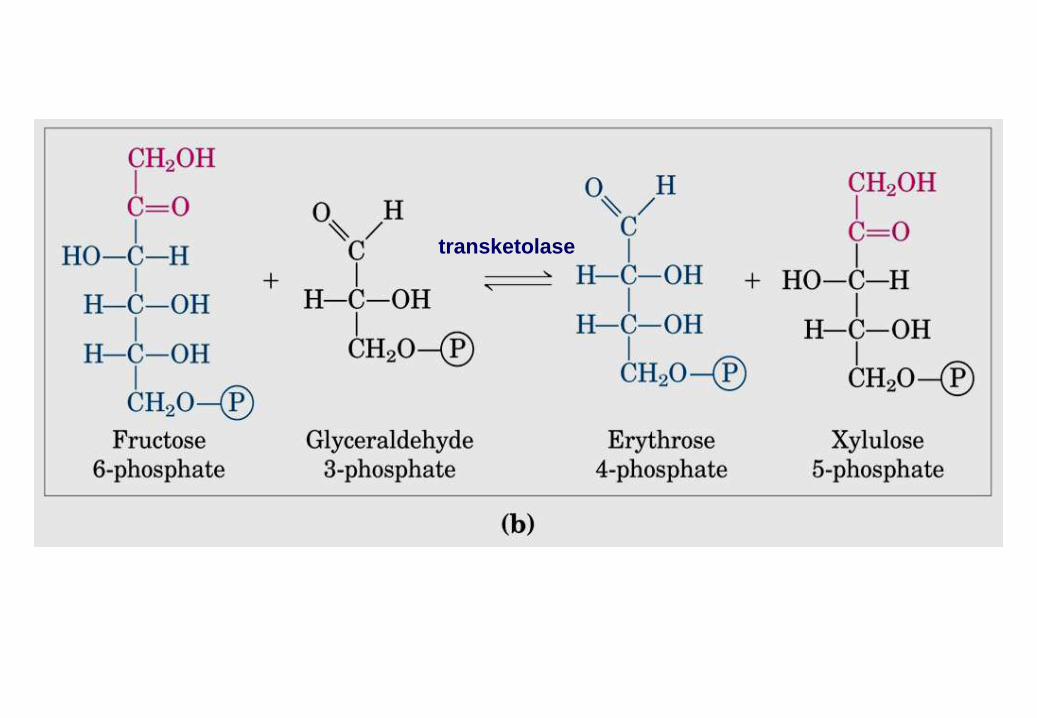
transketolase
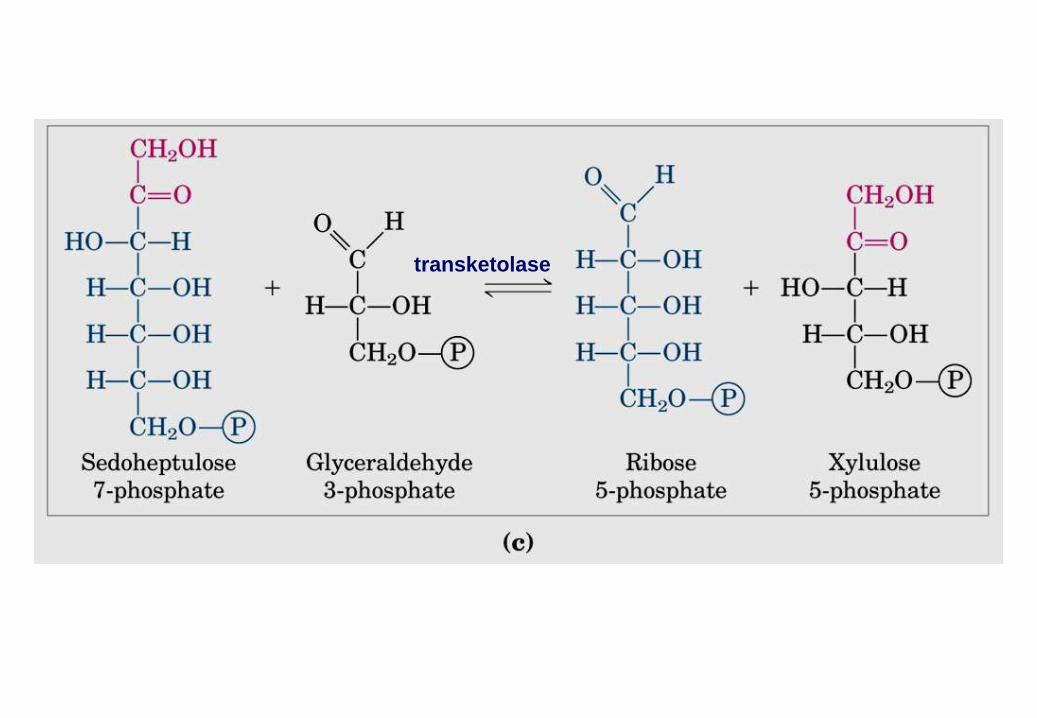
transketolase

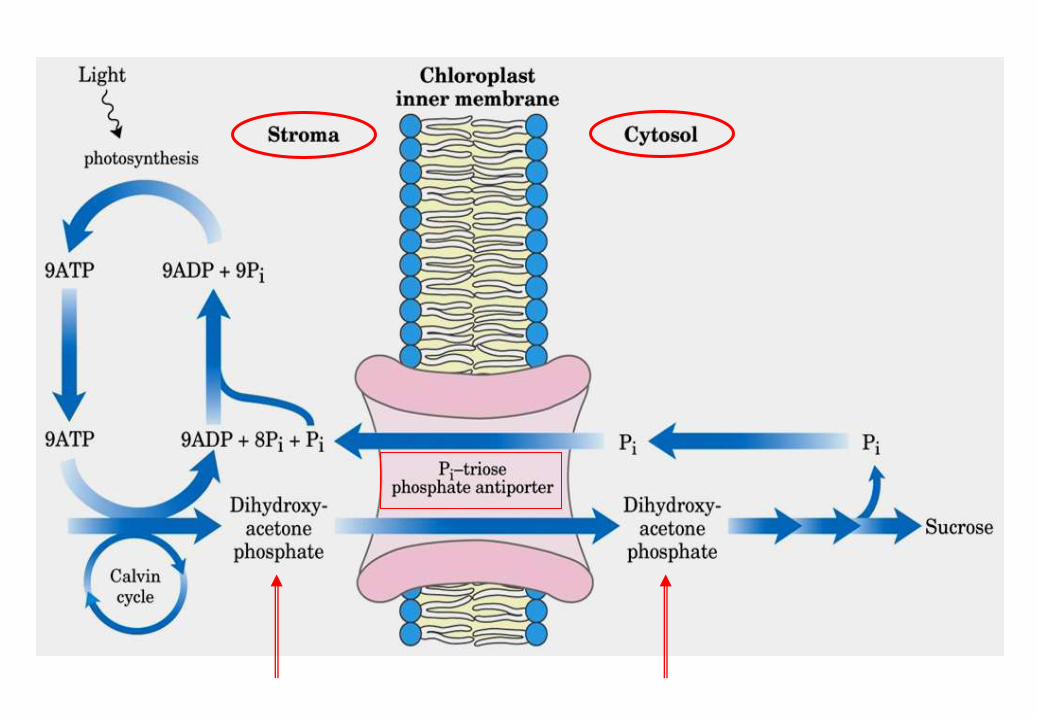
Ogni trioso fosfato sintetizzato
a partire da CO2 costa 9 ATP e
6 NADPH

glicolisi Ciclo di
Calvin

Fotorespirazione
Nelle piante è un processo analogo alla respirazione mitocondriale, ma che NON produce ATP. E’ dovuta alla attività ossigenasica della Rubisco, che non discrimina tra O2 e CO2. In una giornata di sole la velocità della fotorespirazione può essere vicina a quella della fotosintesi e costituire un notevole spreco energetico
Ciclo di Calvin
Riciclato a glicolato, che entra nei perossisomi e diventa glicina
O2

Nelle piante tropicali (ma anche
in alcune piante di interesse
agricolo) la fotorespirazione
viene evitata mediante il ciclo
C4, che usa un intermedio a 4
atomi del carbonio, ma
consuma più energia per
assimilare la CO2

Regolazione del
ciclo di Calvin. 1.

Regolazione del
ciclo di Calvin. 2.
luce
luce
L’attività della fruttosio 1,6-bisfosfatasi
è stimolata da alto pH e alta [Mg2+], che a loro
volta risento della presenza di luce

Regolazione del
ciclo di Calvin. 3.
FBP e SBP sono attivate da
un meccanismo redox
dipendente dalla luce e dalla
azione della FTR
(Ferredossina Tioredossina
Reduttasi) che riduce la
tioredossina.
FTR
FBP,
SBP

FBP