
Influenza dell’eterogeneità ambientale nella valutazione ... · trascorso dall’istituzione)...
105
Università di Pisa Facoltà di Scienze Matematiche Fisiche e Naturali Corso di Laurea Specialistica in Biologia Marina Influenza dell’eterogeneità ambientale nella valutazione dell’effetto riserva: il caso dei popolamenti ittici dell’Arcipelago Toscano Relatore: Lisandro Benedetti-Cecchi Candidato: Martina Dal Bello Anno Accademico 2009-2010
Transcript of Influenza dell’eterogeneità ambientale nella valutazione ... · trascorso dall’istituzione)...

Università di Pisa
Facoltà di Scienze Matematiche Fisiche e Naturali
Corso di Laurea Specialistica in Biologia Marina
Influenza dell’eterogeneità ambientale nella
valutazione dell’effetto riserva:
il caso dei popolamenti ittici dell’Arcipelago Toscano
Relatore: Lisandro Benedetti-Cecchi Candidato: Martina Dal Bello
Anno Accademico 2009-2010

1
INDICE
SUMMARY ...................................................................................................................... 3
RIASSUNTO .................................................................................................................... 4
1. INTRODUZIONE ........................................................................................................ 6
1.1 Ecosistemi marini e attività umane: una visione di insieme ................................ 6
1.2. Le Aree Marine Protette come strumenti per la gestione delle risorse e degli ambienti marini .................................................................................................................... 9
1.2.1. AMP e altri metodi gestionali di conservazione ............................................. 9
1.2.2. AMP: definizioni, categorie e obiettivi ......................................................... 10
1.2.3 AMP: vantaggi, efficacia e limiti ................................................................... 11
1.2.4 Fattori che possono influenzare l’efficacia di una AMP ............................... 15
1.2.4.1 Taglia ed età delle riserve ....................................................................... 15
1.2.4.2 Reti di riserve .......................................................................................... 17
1.2.4.3 Caratteristiche del ciclo vitale ed ecologia delle specie protette ............ 17
1.2.4.5 Eterogeneità spaziale .............................................................................. 19
1.2.5 Aspetti sociali ed economici della conservazione .......................................... 22
1.3 Caratteristiche dei popolamenti ittici protetti del Mediterraneo ..................... 25
1.4 Obiettivi ......................................................................................................................... 27
2. MATERIALI E METODI ......................................................................................... 29
2.1. L’Arcipelago Toscano ............................................................................................... 29
2.2 Le isole oggetto di studio ............................................................................................ 31
2.3 Disegno di campionamento ....................................................................................... 34
2.4 Metodo di campionamento ........................................................................................ 37
2.5 Analisi statistica dei dati ............................................................................................ 39
2.5.1 Cenni teorici .................................................................................................. 39
2.5.1.1 Generalized Additive Models (GAM) .................................................... 39

2
2.5.1.2 Mixed Effect Models (M&M) ................................................................ 40
3.5.1.3 Generalized Additive Mixed Models (GAMM) ..................................... 40
2.5.2 Variabili di risposta e modelli statistici ......................................................... 41
3. RISULTATI................................................................................................................ 45
3.1 Esplorazione dei dati .................................................................................................. 48
3.2 Fit del modello e sua validazione ............................................................................. 57
3.2.1 Influenza dei processi spaziali nella valutazione dell’effetto riserva ............ 57
3.2.1.1 Abbondanza totale .................................................................................. 57
3.2.1.2 Abbondanza delle taglie piccole ............................................................. 60
3.2.1.3 Abbondanza delle taglie medie ............................................................... 63
3.2.1.4 Abbondanza delle taglie grandi .............................................................. 64
3.2.1.5 Abbondanza delle specie predatrici di ricci ............................................ 67
3.2.1.6 Abbondanza delle specie target della pesca ............................................ 69
3.2.1.6 Abbondanza delle specie target della pesca ............................................ 70
3.2.1.7 Ricchezza specifica ................................................................................. 72
3.2.2 Valutazione degli effetti legati ai diversi regimi di protezione applicati ...... 74
4. DISCUSSIONE E CONCLUSIONI ......................................................................... 76
5. BIBLIOGRAFIA ....................................................................................................... 83
APPENDICE A .............................................................................................................. 98

3
SUMMARY
In a world where population and economic growth-rate are incessantly increasing, the
recognition of the profound influence of humans in marine systems and the raising
interest in ocean’s biodiversity and ecological processes lead to strong marine
conservation advocacy. Environmental conservation and sustainable development are
considered the goals to achieve in the imminent future.
Marine Protected Areas (MPA) are globally advocated as efficient and inexpensive
tools to maintain and manage fisheries, while simultaneously preserving biodiversity
and ecosystems’ functioning. Marine reserves lead to increase in total abundance,
biomass and size of fish within their boundaries, especially for species target by
fisheries, as well as enhance local reproductive output. This often results in increasing
diversity of the protected area: the restoration of large fish populations and lost
predatory interactions can cause community-wide effects, such as trophic cascade, that
may trigger the recovery of rocky reef food webs. However, effects of MPA vary both
in direction and magnitude and the reasons are still unknown. This heterogeneity in
response to protection may stem from reserve’s size and age, inclusion in networks of
marine reserves, species life history and ecological traits, as well as habitat patchiness.
The present study investigated the influence of the habitat structure (here represented by
bottom and shore heterogeneity and geographical coordinates), which can be source of
confounding when measuring the effect of protection in four islands of the Tuscany
Archipelago (Capraia, Pianosa, Montecristo and Giannutri). The data were collected
using standard visual census techniques.
Results showed that fish population distribution patterns were inconsistent with effects
of protection; in contrast, environmental processes, especially the effects of shore
heterogeneity and geographical coordinates, are the factors likely to explain a
substantial part of the observed variation. Abundances of fish species increase following
a latitudinal gradient, with direction North to South, and were bigger where shore
heterogeneity showed intermediate values.
Along with considerations of ecological traits of fish species, connectivity, size, age and
enforcement of reserves, our results ascribe a value to spatial heterogeneity as a
fundamental informative criterion to use when measuring the effects of protection.

4
RIASSUNTO In un pianeta in rapida crescita demografica ed economica la presa di coscienza del
progressivo deterioramento degli ecosistemi, da un lato, e il crescente interesse per la
biodiversità degli ambienti marini ed i processi ecologici ad essi associati, dall’altro,
hanno messo in luce l’incapacità dell’uomo di gestire l’immenso patrimonio custodito
negli oceani e la totale mancanza di un approccio lungimirante, teso a quantificare
l’impatto provocato dall’uso indiscriminato delle risorse marine. Una corretta tutela e
una gestione sostenibile dell’ambiente, in particolare quello marino, vengono oggi
avvertite, sia dai governi, sia dall’opinione pubblica come una necessità per il presente e
un dovere verso le generazioni future.
Le Aree Marine Protette (AMP), ad oggi, sono globalmente riconosciute come uno dei
migliori strumenti volti alla protezione della biodiversità, la conservazione degli
ecosistemi marini e la gestione degli stock ittici. All’interno delle AMP si riscontra,
infatti, un generale aumento di densità, taglia e biomassa corporea per specie target
della pesca, nonché un notevole miglioramento dell’output riproduttivo di tali specie.
Tutto ciò si traduce spesso in un incremento di biodiversità dell’area soggetta a
protezione. Le specie maggiormente soggette a prelievo, infatti, occupano spesso i
livelli più alti delle reti trofiche e il loro sovrasfruttamento può determinare gravi
conseguenze sull’intera comunità attraverso le cosiddette cascate trofiche con una
riduzione della diversità complessiva. Le AMP hanno dimostrato di essere in grado di
ristabilire le interazioni predatorie perdute e di promuovere il recupero a livello delle
comunità. Nonostante i risultati ottimistici ottenuti in molti studi, le risposte dei
popolamenti ittici alla protezione variano sia in direzione, che entità e le basi di questa
eterogeneità negli effetti delle AMP sono tuttora in gran parte sconosciuti. Le principali
sorgenti di eterogeneità nell’efficacia delle AMP sono: taglia ed età (cioè il tempo
trascorso dall’istituzione) della riserva, possibile inclusione in una rete di riserve, ciclo
vitale e caratteristiche ecologiche delle diverse specie soggette a protezione ed
eterogeneità spaziale nella distribuzione dei popolamenti protetti.
Il presente lavoro di tesi si propone valutare gli effetti di confusione che processi
spaziali legati all’eterogeneità ambientale (riassunti negli indici di eterogeneità del
substrato e della costa e dalle coordinate geografiche) possono introdurre nella
valutazione dell’effetto riserva, presso alcune isole del Parco Nazionale dell’Arcipelago

5
Toscano (Capraia, Giannutri, Montecristo e Pianosa). I dati sono stati raccolti attraverso
un campionamento della fauna ittica (effettuato tramite tecniche non distruttive di visual
census) appropriato per comparare la struttura dei popolamenti ittici, le densità e le
distribuzioni di taglia di singole specie target all’interno ed all’esterno delle AMP.
I risultati relativi alle modalità di distribuzione delle abbondanze delle specie censite
non hanno mostrato significative differenze tra le aree protette e le località esterne alle
AMP, mentre è emersa l’importanza degli effetti relativi alle coordinate spaziali e
all’indice di eterogeneità della costa. In particolare, le abbondanze delle specie ittiche
crescono in direzione Nord-Sud e sono massime per valori intermedi di eterogeneità
della costa.
Nonostante i fattori da considerare siano molteplici, dalle caratteristiche del ciclo vitale
proprie di ciascuna specie all’efficacia delle misure di controllo e tutela dell’area, i
risultati ottenuti nel presente lavoro di tesi suggeriscono l’importanza di includere, negli
studi sulla valutazione dell’effetto riserva, indici relativi a processi spaziali legati
all’eterogeneità ambientale.

6
1. INTRODUZIONE
1.1 Ecosistemi marini e attività umane: una visione di insieme Gli oceani si estendono per circa il 70,8% della superficie terrestre e, grazie
all’incredibile varietà di ecosistemi, organismi viventi e processi chimici, fisici,
geologici e biologici, rappresentano una delle più importanti risorse naturali a cui
l’uomo attinge da migliaia di anni.
Per secoli si è creduto che la vastità dell’oceano e la ricchezza delle sue forme di vita
avrebbero sopperito al prelievo attuato dall’uomo e che le sue risorse sarebbero state
praticamente illimitate. “I believe, then, [...] that probably all the great sea fisheries are
inexhaustible” rassicurava il biologo Thomas Huxley nel 1883: egli era realmente
convinto che le risorse alieutiche fossero inesauribili e che l’azione umana di prelievo
fosse assolutamente insignificante rispetto alla mortalità naturale e ininfluente sulla
consistenza degli stock per il loro enorme potenziale riproduttivo.
Sfortunatamente, quello che si può chiamare il “capitale” ittico non è illimitato. Anzi,
l’incapacità dell’uomo di gestire tali, seppur ingenti, risorse, aggravata da una forte
crescita demografica e da un insediamento costiero sempre più consistente, ha
determinato forme di impatto antropico a discapito degli ecosistemi sommersi a diversi
livelli (Carlton, 1999; Hixon et al., 2001). La maggior parte degli ambienti marini e
costieri, infatti, mostra alti livelli di deterioramento inequivocabilmente imputabili alle
attività umane in termini di sovrasfruttamento, alterazione fisica degli habitat,
introduzione di specie alloctone, cambiamenti climatici globali e inquinamento
(Peterson e Estes, 2001).
I sistemi più minacciati sono gli ambienti costieri, le barriere coralline e le comunità di
organismi il cui ciclo vitale è in parte, o totalmente, associato alla piattaforma
continentale (Hixon et al., 2001). Questi habitat sono più soggetti ad impatti indotti
dall’insediamento e dall’urbanizzazione costiera: attività di bonifica, disboscamento con
conseguente erosione del litorale, dragaggio, edificazione di porti, dighe foranee e
barriere di protezione della costa, costruzione di impianti di maricoltura e forme di
inquinamento organico, causate dallo sversamento di scarichi e acque reflue, che

7
modificano profondamente lo stato dei nutrienti negli ecosistemi marini, portando a
fenomeni di eutrofizzazione (Palumbi, 2007).
Per quanto riguarda, invece, gli ecosistemi pelagici, il passaggio dalla pesca artigianale
a nuove tecnologie, con più efficaci modalità di cattura, ha portato via via
all’impoverimento degli stock ittici e all’alterazione delle dinamiche di popolazione, dal
momento che le specie maggiormente sfruttate sono anche quelle che occupano,
all’interno delle reti trofiche, i livelli più alti (Pauly et al., 1998) e che quindi possono
determinare, attraverso quelle che vengono chiamate “cascate trofiche”, cambiamenti
nella struttura dell’intera comunità. Ad esempio, la pesca tramite dragaggio e quella a
strascico, attività di cui si hanno, purtroppo, pochi dati, danneggiano irreparabilmente
ambienti e comunità dei fondali (Dayton et al. 1995), non solo perché determinano
l’asportazione di tutto ciò che si trova sul fondo lungo il tratto battuto, ma anche a causa
della frequenza con cui tali pratiche vengono attuate.
Secondo la FAO (Food and Agricolture Organization of the United Nations), il 70% dei
principali stock commerciali presenta problemi di varia entità:
• 6% estinti,
• 44% sfruttati in modo intensivo,
• 16% sovrasfruttati,
• 31% sottosfruttati e moderatamente sfruttati,
• solo il 3% in lento recupero.
Tuttavia, sempre secondo la FAO, tra gli anni ’70 e ’90, le flotte pescherecce mondiali
sono aumentate di due volte rispetto al tasso di crescita delle popolazioni ittiche e oggi
la capacità di cattura della flotta europea sembra superi di 40 volte la risorsa prelevabile
in modo sostenibile, rendendo vani gli sforzi per preservare e gestire la sopravvivenza a
lungo termine delle specie ittiche. L’intensità e l’efficacia delle azioni di pesca non
permettono di sfruttare in modo sostenibile la risorsa alieutica, che progressivamente si
riduce (overfishing). Tristemente noto è il caso del merluzzo bianco dei banchi di
Terranova (Gadus morhua), che, negli anni ’70, a causa dell’eccessivo sfruttamento, era
quasi scomparso dalle acque canadesi (Kurlansky, 1999); lo stock si è parzialmente
ricostituito solo a seguito della riduzione forzata delle catture.
Un altro spinoso problema legato alla pesca è quello del cosiddetto “bycatch”, per cui,
animali come delfini, tartarughe e uccelli marini, vengono non intenzionalmente
catturati e in seguito scartati. A questi si aggiungono tutti gli esemplari di specie ittiche

8
pescati e successivamente buttati a mare o perché di scarso valore economico o perché
al di sotto dei limiti di taglia imposti dalla legge. Negli ultimi anni, il numero degli
esemplari rigettati, morti, a mare ha superato, in alcuni casi, quello delle specie catturate
per essere vendute (Palumbi, 2007).
Se quanto detto vale per le nazioni con un’economia ormai consolidata, altrettanto
grave risulta essere la situazione degli oceani su cui si affacciano paesi in via di
sviluppo: la pesca incontrollata e l’uso di dinamite, cianuro e altre sostanze velenose
finalizzate ad una pesca più efficace hanno posto le barriere coralline tra gli habitat più
a rischio (Attwood, 1997).
Altra seria minaccia per gli oceani risulta essere l’inquinamento dovuto a sversamenti
accidentali di greggio, presenza di metalli pesanti, idrocarburi alogenati (pesticidi come
il DDT, le diossine ed i PCB): la concentrazione di tali sostanze, ritrovate anche nei
tessuti di calamari, mammiferi e uccelli marini a causa di fenomeni di bioaccumulo e
biomagnificazione, è tale da necessitare una seria ed efficace politica di recupero
ambientale (Hixon et al., 2001).
La navigazione su scala mondiale, inoltre, ha inavvertitamente introdotto, nel corso dei
secoli, numerose specie alloctone. Tale fenomeno può determinare effetti devastanti
sulle specie e sulle comunità native, sia a causa di malattie importate dalle specie
invasive che sterminano le popolazioni originarie, sia perchè innesca processi di
competizione in cui le specie importate, di solito più resistenti, mostrano strategie
vincenti (Carlton, 1999).
Infine non va dimenticato l’impatto che le attività turistiche possono avere sugli
ecosistemi: danni alle comunità di coralli o kelp inavvertitamente provocati da
subacquei poco esperti o dalle ancore delle barche da diporto, scarichi e inquinamento
causato dalle navi da crocera, incidenti che provocano la morte di cetacei e tartarughe e
molte altre attività (Palumbi, 2007).
Alla luce di quanto brevemente riportato, nonostante l’incertezza riguardo ai
meccanismi di azione dei fenomeni che minacciano gli oceani e la scarsa conoscenza
degli ecosistemi marini, appare chiara la necessità di un’immediata ed efficace azione di
conservazione.

9
1.2. Le Aree Marine Protette come strumenti per la gestione delle risorse e degli ambienti marini
1.2.1. AMP e altri metodi gestionali di conservazione
In anni recenti, l’interesse per la biodiversità degli ambienti marini ed i processi
ecologici ad essi associati hanno messo in luce l’incapacità dell’uomo di gestire
l’immenso patrimonio custodito negli oceani e la totale mancanza di un approccio
lungimirante, teso a quantificare l’impatto provocato dall’uso indiscriminato delle
risorse marine.
La presa di coscienza di tale responsabilità ha dato il via ad un dialogo internazionale
volto a porre rimedio ai danni apportati all’ambiente, o almeno, finalizzato ad
identificare vie di compromesso tra progresso tecnologico e conservazione dei sistemi
naturali. Di conseguenza, i principi della conservazione non solo devono comprendere
progetti di protezione di specie in via di estinzione o di salvaguardia di aree di
particolare interesse naturalistico, bensì sono articolati su più livelli, dalla prevenzione
al risanamento, dalla gestione di riserve naturali all’educazione ambientale. Comincia
così a farsi strada il concetto di “sviluppo sostenibile”, il quale sottolinea che l’azione
dell’uomo, nonostante alteri l’ambiente per renderlo più adatto alle proprie esigenze,
non deve depauperare le risorse rinnovabili, in modo tale da consentire la loro
rigenerazione e da renderle disponibili in maniera illimitata.
Un altro passo importante nel processo di responsabilizzazione del genere umano nei
confronti dei danni ambientali è stato il riconoscimento e la successiva definizione dei
“servizi” offerti da mari ed oceani di cui l’uomo usufruisce, non solo indirettamente
attraverso fenomeni fisici passivi (ciclo idrologico, sottrazione di anidride carbonica
atmosferica, regolazione dei cambiamenti climatici), ma anche e soprattutto estraendo
attivamente risorse per il sostentamento e il progresso umano.
Tuttavia, fino agli anni ’70, le strategie di preservazione degli ambienti marini erano
finalizzate alla conservazione ed il ripristino degli stock ittici di valore commerciale e,
di conseguenza, si focalizzavano sulla protezione di singole specie. Le misure adottate,
basate sul concetto di Massimo Prelievo Sostenibile (MSY: Maximum Sustainable
Yield), prevedevano chiusure stagionali di alcune aree di pesca, limiti di taglia al

10
pescato e comprendevano anche forme di gestione volte a preservare il potenziale
riproduttivo degli stock o a proteggere gli stadi giovanili.
I progressi negli studi ecologici, che hanno via via messo in luce l’importanza delle
interazioni interspecifiche nella determinazione dei processi biologici alla base delle
catene trofiche oceaniche, hanno fatto intravvedere la necessità di una gestione
dell’ambiente marino a livello ecosistemico. I legami che esistono tra le diverse specie
all’interno di un ecosistema garantiscono che, se si verifica un impatto, questo non è
limitato alla specie interessata, ma si riverbera attraverso l’intero ecosistema in cui
quella specie è inserita (Palumbi, 2007).
Ad oggi, ambiente scientifico e organizzazioni internazionali, quali l’Unione
Internazionale per la Conservazione, sono concordi nell’affermare che le Aree Marine
Protette, consentendo una gestione delle risorse marine basata sugli ecosistemi, siano il
miglior strumento per conciliare attività antropiche e conservazione dell’ambiente e
delle specie sovrasfruttate o danneggiate.
1.2.2. AMP: definizioni, categorie e obiettivi
Varie definizioni sono state date per indicare un’area sottoposta a vincoli di protezione:
aree protette, riserve naturali, riserve di pesca, parchi, santuari, etc. Il termine Area
Marina Protetta (AMP), però, è emerso solo di recente e, secondo quanto sancito
dall’Unione Internazionale per la Conservazione, è definita come “area del dominio
marino (intertidale o subtidale), comprendente le acque, la flora, la fauna e le
caratteristiche storiche e culturali, che sia preservata da provvedimenti legislativi con
l’intento di proteggere parzialmente o totalmente l’ambiente in oggetto” (IUCN, 1988;
Kelleher e Recchia, 1998).
Questa definizione ben si adatta a includere la vasta terminologia applicata ad aree
marine sottoposte a specifici vincoli protezionistici, ma, proprio perché molto generica,
è stata supportata dall’istituzione di sei categorie (Riserve Naturali, Parchi Nazionali,
Monumenti Nazionali, Aree per la Gestione di Habitat o Specie, Aree per la Protezione
del Paesaggio Marino, Aree Protette per la Gestione delle Risorse), create dallo stesso
IUCN (Kelleher e Recchia, 1998), al fine di distinguere il diverso utilizzo e gli specifici
criteri gestionali applicati a una AMP.

11
Le AMP offrono una vasta gamma di potenziali funzioni che varia dalla conservazione
della biodiversità alla protezione di habitat particolarmente sensibili (Norse, 1993),
dalla conservazione degli ecosistemi più rilevanti al ripristino di stock ittici
intensamente sfruttati (Dungan e Davis, 1993; Friedlander, 2001), fino alla funzione
non meno importante di promuovere un turismo eco-compatibile (Ballantine, 1991;
Rowley, 1992). Data la vastità delle funzioni a cui le AMP posso adempiere, risulta più
facile operare una classificazione in base allo scopo per cui esse vengono istituite;
pertanto si distinguono tre principali tipologie:
1. riserve per il ripristino e la protezione degli stock ittici;
2. aree protette per la conservazione della diversità biologica;
3. riserve istituite per l’unicità geografica (Palumbi, 2001).
In ogni caso, in accordo con quanto riportato da Roberts et al. (2002), una riserva,
qualunque sia la sua funzione, deve tenere in considerazione:
• eterogeneità biogeografica: devono esservi incluse differenti zone
biogeografiche;
• eterogeneità a livello di habitat: devono esservi compresi diversi habitat;
• possibile esposizione ad attività antropiche non estrattive (es. inquinamento);
• possibile esposizione a eventi catastrofici di carattere naturale;
• idoneità delle dimensioni agli scopi per cui essa è istituita;
• connettività con altre riserve;
• possibile inclusione di habitat, popolazioni o stadi vitali più sensibili di altri,
specie sfruttabili dall’uomo e specie di particolare interesse naturalistico-
ambientale;
• possibile presenza di servizi ecologici per l’uomo.
1.2.3 AMP: vantaggi, efficacia e limiti Le AMP, da alcuni anni a questa parte, sono ampiamente utilizzate nella gestione e
nella conservazione dell’ambiente marino. Sono, infatti, considerate uno strumento
efficace per la gestione delle risorse alieutiche (Hastings and Botsford 1999, Roberts et
al. 2001, Pauly et al. 2002, Gõni et al. 2008, Harmelin-Vivien et al. 2008), per il
recupero delle risorse marine sovra sfruttate (Schrope 2001, Claudet et al. 2006a, 2008)
e per il ripristino della biodiversità (Halpern 2003, Micheli et al. 2004, Rodrigues et al.

12
2004, Claudet et al. 2008) su base ecosistemica. Inoltre, rispetto ai metodi tradizionali,
hanno il potenziale vantaggio di realizzare contemporaneamente una vasta gamma di
obiettivi, di promuovere e facilitare studi riguardo ai processi ecologici che determinano
il naturale andamento degli ecosistemi e, infine, sollecitare forme di educazione
ambientale (Boersma e Parrish, 1999).
Numerose sono le evidenze empiriche riguardo all’efficacia delle AMP dal punto di
vista ecologico e biologico.
È ampiamente riconosciuta la capacità delle AMP di portare ad un aumento della
densità, taglia e biomassa corporea delle specie ittiche all’interno dei propri confini,
specialmente per quelle target della pesca (Harmelin et al. 1995, Jennings 2004,
Mosquera et al. 2000, Micheli et al. 2004), nonché ad un notevole miglioramento
dell’output riproduttivo di tali specie, che si traducono in un incremento di biodiversità
dell’area soggetta a protezione.
Alcune indagini (Fujita, 2001; Halpern, 2003) riguardanti le AMP hanno rilevato che:
• il 90% delle riserve presenta valori di biomassa ittica superiori rispetto alle aree
non soggette a protezione;
• la densità dei pesci è maggiore del 63% nelle riserve;
• l’83% delle AMP presenta specie carnivore ed invertivore più grandi;
• il 59% delle AMP mostra una biodiversità più elevata.
Permettendo agli individui protetti di crescere maggiormente e vivere più a lungo, le
Aree Marine Protette potenzialmente consentono di raggiungere due obiettivi
sostanziali per la gestione delle risorse alieutiche: 1) sopperiscono al declino degli stock
ittici causato da fenomeni di overfishing; 2) sostengono la produzione di specie target
della pesca nelle aree adiacenti alla riserva e nelle quali la pesca è consentita. Queste
funzioni derivano principalmente da variazioni nei tassi di reclutamento e connettività,
caratteristiche peculiari delle popolazioni marine (Sale et al. 2005). Il reclutamento in
popolazioni di specie target della pesca è tipicamente molto variabile nel tempo e nello
spazio (Caley et al. 1996, Sale e Kritzer 2003). Le conseguenze di un grande evento di
reclutamento persistono per anni e risultano importanti nei processi di recupero a lungo
termine di una popolazione. Tuttavia, tassi di reclutamento variabili significano anche
che piccole popolazioni, come sono quelle sovra sfruttate, sono estremamente
suscettibili a drammatici crolli che possono portare anche ad estinzioni locali (Musick
1999, Foley 1994). Per questi motivi, le AMP, mantenendo livelli di densità piuttosto

13
elevati, possono costituire efficaci serbatoi contro il sovra sfruttamento (Zeller e Russ
2004). Le popolazioni marine sono interconnesse, caratterizzate cioè dallo scambio di
individui, che avviene principalmente attraverso la dispersione larvale. La connettività è
di cruciale importanza per il ruolo delle AMP nella gestione delle risorse alieutiche
perché rappresenta il meccanismo fondamentale che consente alle riserve marine di
migliorare la produttività nelle loro aree limitrofe (Hilborn et al. 2004, Botsford et al.
2001, Kritzer e Sale 2004). La capacità di una AMP di migliorare la pesca nelle aree
adiacenti dipende, infine, da due processi noti come recruitment subsidy e spillover. Le
popolazioni protette, caratterizzate da pesci di taglia maggiore e densità di popolazione
più elevate, come conseguenza, possono potenzialmente produrre più propaguli (uova e
larve) rispetto a quelle non protette e rifornire aree esterne alle AMP grazie alle
correnti, fenomeno conosciuto come recruitment subsidy (Russ e Alcala, 1996). Un
processo simile risulta favorito se le AMP proteggono specifici habitat in cui gli
individui di una data specie si riproducono (Kramer e Chapman, 1999). Gli aumenti di
densità e taglia conseguenti alla protezione, inoltre, tendono a favorire processi densità-
dipendenti (es. competizione per spazio e risorse), con potenziali benefici per la pesca
locale nelle aree adiacenti alle AMP, attraverso un fenomeno chiamato spillover,
letteralmente “traboccamento”, inteso come esportazione netta di biomassa adulta
attraverso i confini di una riserva verso le vicine aree di pesca (Roberts et al., 2001). Al
di là di concetti generali, i fenomeni di spillover ed il loro successo sono legati alle
specifiche caratteristiche dell’area in cui l’AMP viene collocata (es. tipologia e
distribuzione degli habitat all’interno ed all’esterno dell’AMP, presenza di barriere e
discontinuità, livello di pressione di pesca nei pressi dei confini delle AMP, dimensione
forma delle AMP), le quali sono a loro volta strettamente connesse a caratteristiche
tipiche di ogni specie ittica (es. dimensioni dell’home range, mobilità, preferenze
rispetto ad alcune tipologie di habitat).
Non ultimo, le AMP favoriscono la ricostituzione di habitat danneggiati con
conseguente ripristino delle comunità associate a quei determinati ambienti (Guidetti
2006, Guidetti e Sala 2007). Le comunità bentoniche del sublitorale roccioso
mediterraneo presentano due stadi alternativi: barren, fondali “spogli” dominati
dall’associazione a Rodoficee calcaree incrostanti, e letti di macroalghe (Sala et al.
1998). Quando la densità del riccio Paracentrotus lividus va oltre il valore soglia di 7-9
ricci per metro quadro, si assiste alla transizione da comunità macroalgali a barren

14
(Verlaque 1987, Benedetti-Cecchi et al. 1998, Hereu 2004), con gravi danni per l’intero
ecosistema. La scomparsa delle macroalghe erette, infatti, comporta una brusca
diminuzione della produzione primaria, della diversità biologica, della disponibilità di
detrito algale e della struttura tridimensionale presente, con una irreversibile riduzione
delle aree di nursery, cioè le aree di rifugio, foraggiamento ed accrescimento per stadi
giovanili di pesci ed invertebrati vagili che reclutano nell’infralitorale roccioso (Guidetti
e Sala, 2007). Le specie ittiche che predano i ricci sono numerose, ma quelle in grado di
controllarne effettivamente la densità sono gli sparidi Diplodus sargus, Diplodus
vulgaris e Sparus aurata (che si nutrono sia degli adulti che dei giovanili di P. lividus) e
grossi individui di labridi, come Coris julis e Thalassoma pavo, che predano solo i
giovani ricci, con diametro minore di 1 cm (Sala 1997, Guidetti 2006). La pesca,
rimuovendo gli esemplari più grandi delle specie predatrici di ricci, pertanto, tende a
favorire il passaggio da letti macroalgali a barren, con una perdita complessiva della
biodiversità dell’area. Recenti studi hanno evidenziato come le AMP, mediante la
ricostruzione delle interazioni predatorie perdute a seguito dell’eccessivo prelievo di
predatori di alto livello trofico, rappresentino un valido strumento per il ripristino di
habitat danneggiati. Bisogna però precisare che tali cambiamenti richiedono tempi più
lunghi per realizzarsi e pertanto possono non essere rilevati in studi effettuati pochi anni
dopo l’istituzione di una riserva (Russ e Alcala 2004, Russ et al. 2005). Come osservato
da diversi autori (es. Shears e Babcock 2003, Micheli et al. 2004) cambiamenti a livello
delle cascate trofiche all’interno delle riserve possono seguire complesse traiettorie di
successione ed essere caratterizzate da andamenti non lineari, che possono non essere
evidenziati in studi a breve termine.
E’ tuttavia evidente come le AMP da sole non possano essere sufficienti a garantire una
protezione totale all’ambiente, in quanto non isolate da certe forme di impatto (Allison
et al., 1998) che inevitabilmente oltrepassano i confini giurisdizionali delle riserve,
come l’inquinamento chimico (dovuto all’immissione idrocarburi, metalli pesanti, ma
anche acque reflue) e quello biologico (introduzione di specie alloctone e malattie
associate a maricoltura e acque di sentina). Inoltre, la scala su cui agiscono
fondamentali processi biologici, spesso, non è interamente rappresentabile all’interno di
una AMP (Allison et al., 1998): nella pratica è impossibile pensare di gestire riserve di
dimensioni tali da consentire la protezione di specie il cui ciclo vitale è interamente o in
parte caratterizzato da un vasto home range. Infine, non bisogna dimenticare che

15
l’istituzione di una AMP può causare un aumento della pressione antropica nelle aree
adiacenti la riserva, tale da compromettere l’efficacia della riserva stessa (Carr e Reed,
1993). Non solo, come afferma la teoria degli stati stabili alternativi (Knowlton, 2004),
il solo divieto di pesca può non essere garanzia di un recupero dell’ecosistema.
Nonostante la complessità e la variabilità dei sistemi naturali determinino incertezza
riguardo alla validità di questo strumento di protezione, le AMP, grazie ad un approccio
ecosistemico, rappresentano un metodo gestionale precauzionale in grado di preservare
interi ecosistemi marini.
1.2.4 Fattori che possono influenzare l’efficacia di una AMP Nonostante i risultati ottimistici ottenuti in molti studi, le risposte dei popolamenti ittici
alla protezione variano sia in direzione, che entità (Halpern, 2002) e le basi di questa
eterogeneità negli effetti delle AMP sono tuttora in gran parte sconosciuti. Approfondite
analisi volte a colmare tali lacune sono fondamentali per sviluppare una teoria generale
sull’efficacia delle aree marine protette (Claudet et al. 2008 pdf).
Le principali sorgenti di eterogeneità nell’efficacia delle AMP sono: taglia ed età (cioè
il tempo trascorso dall’istituzione) della riserva, possibile inclusione in una rete di
riserve, ciclo vitale e caratteristiche ecologiche delle diverse specie soggette a
protezione ed eterogeneità spaziale nella distribuzione dei popolamenti protetti.
1.2.4.1 Taglia ed età delle riserve Studi teorici suggeriscono che riserve grandi dovrebbero essere maggiormente efficaci
nell’adempiere a obiettivi di conservazione rispetto ad AMP piccole. Riserve grandi
ospitano grandi popolazioni di numerose specie; queste grandi popolazioni
risulterebbero meglio protette dall’estinzione, sia perché sono ampie, sia perché gli
individui sarebbero in grado di completare il loro ciclo vitale interamente entro i confini
della riserva, rendendo tale popolazione autosufficiente (Jones 1994, Botsford et al.
2001 e 2003, Hastings e Botsford 2003, Roberts et al. 2003). Tuttavia numerosi studi
empirici non hanno supportato questa ipotesi (Côté et al. 2001, Halpern 2003, Guidetti e
Sala 2007). Uno studio più recente (Claudet et al. 2008) e basato su dati di quaranta
studi su dodici AMP distribuite su un’area di 2500 km, dal Mediterraneo Nord-
occidentale all’Atlantico Centro-orientale, ha invece messo in evidenza che la risposta

16
delle specie target della pesca alla protezione dipende dalla taglia della riserva; in
particolare, un aumento delle dimensioni della no-take zone porta ad un aumento della
densità delle specie di valore commerciale rispetto all’esterno. Un possibile
meccanismo biologico che spieghi quanto osservato è che grandi zone interdette alla
pesca permettano a una frazione più grande di pesci con grandi home-range di rimanere
“protetti” entro l’AMP rispetto a riserve più piccole (Claudet et al. 2008). Grandi AMP,
inoltre, aumentano il cosiddetto self-recruitment (Botsford 2003). Al contrario,
sembrerebbe che un aumento della taglia della buffer zone determini effetti opposti:
nonostante presentino una regolamentazione più restrittiva in materia di pesca rispetto
alle aree non protette, queste zone sono sottoposte a una maggiore pressione di pesca
dal momento che attirano molti pescatori artigianali (Stelzenmüller et al. 2007).
Anche per quanto riguarda l’andamento della densità dei pesci target della pesca in
funzione degli anni passati dall’istituzione della riserva, i risultati degli studi condotti
sono eterogenei. Alcune review e singoli studi mostrano che la densità dei pesci di
valore commerciale e la ricchezza in specie aumentano dopo tre anni dall’inizio delle
misure di protezione (Halpern e Warner 2002, Russ et al. 2005, Claudet et al. 2006a);
altri invece affermano che è necessario un decennio (Micheli et al. 2004). Nello studio
di Claudet et al. 2008, invece, è emerso che, per ogni anno passato dall’istituzione, la
densità relativa dei pesci target aumenta mediamente dell’8.3 %. Tuttavia, non bisogna
dimenticare che, anche rimuovendo l’attività di pesca, che impatta alcune specie ed
alcune taglie, la grandezza della risposta all’istituzione di una riserva nel tempo è
strettamente correlata al valore commerciale e alla taglia dei pesci pescati (Claudet et al.
2006a). Inoltre, è importante proteggere i pesci più grandi, e quindi più vecchi, che
hanno il più grande potenziale riproduttivo e producono larve con tassi di sopravvivenza
maggiori rispetto a pesci più giovani (Birkeland e Dayton 2005). Il fatto che le AMP
europee più vecchie risultino maggiormente efficaci, può essere spiegato dal fatto che
molti specie di interesse commerciale hanno cicli vitali molto lunghi (es. Serranidae
possono vivere fino a quaranta anni) e, pertanto, il recupero delle comunità ittiche si
realizza molto lentamente (Claudet et al. 2008). Per questi motivi, un sistema di
rotazione temporale delle aree sottoposte a protezione può risultare inadeguato ad
adempiere a obiettivi di conservazione, tralasciando gli effetti sconosciuti dell’aumento
della varianza temporale della pressione di pesca sulla densità media delle popolazioni
ittiche (Benedetti-Cecchi et al. 2003).

17
1.2.4.2 Reti di riserve Modelli matematici di reti di AMP dimostrano l’importanza della vicinanza tra riserve,
nonostante la distanza ottimale tra due aree marine protette dipenda dagli obiettivi di
conservazione preposti, dai dati considerati e dalle assunzioni fatte (Sala et al. 2002,
Hastings e Botsford 2003). Lo studio di Claudet et al. 2008 è il primo a valutare
l’effetto della distanza tra riserve adiacenti sull’efficacia delle misure di protezione: non
si evidenzia però alcuna variazione nell’andamento della densità delle specie target e
della ricchezza di specie imputabile alla distanza tra AMP. Su scala regionale questo
fattore sembra non essere di fondamentale importanza; altri fattori, invece, come
discontinuità e frammentazione dell’habitat (Banks e Skilleter 2007, Moilanen e Wintle
2007), dispersione larvale (Cowen et al. 2006, Steneck 2006) e dinamiche di specie e di
disturbo (Cottenie 2005, Leroux et al. 2007) possono giocare un ruolo più importante
della connettività. Inoltre, se l’esportazione di biomassa dall’interno verso l’esterno di
una AMP si realizzano su piccola scala spaziale (Murawski et al. 2004, Goñi et al.
2006), le riserve del Mediterraneo potrebbero essere troppo lontane per fare parte di una
effettiva rete di AMP su scala regionale.
1.2.4.3 Caratteristiche del ciclo vitale ed ecologia delle specie protette É intuitivo pensare che gli effetti della protezione siano differenti da una specie ittica
all’altra. Ciò avviene sostanzialmente perché ogni specie possiede peculiari
caratteristiche del ciclo vitale, comportamento, legame con gli habitat, che la rendono
più o meno sensibile alla protezione. Per esempio, secondo Kramer e Chapman (1999),
densità e taglia media delle specie ittiche saranno maggiori all’interno di un’AMP
rispetto ad un’area adiacente aperta alla pesca per:
• specie con home range reltivamente piccoli;
• specie non migratrici;
• specie fortemente legate al substrato;
• specie i cui spostamenti sono limitati dalla presenza di barriere;
• specie che non mostrano una distribuzione densità-dipendente.
Anche modelli demografici spaziali hanno ipotizzato che pesci molto mobili non siano
influenzati dalla protezione (vedi ad esempio DeMartini 1993 e Walter et al. 1999).

18
Tuttavia, i pochi studi effettuati hanno fornito supporto limitato o nullo a tali previsioni
(Micheli et al. 2004, Blyth-Skyrme et al. 2006, Goñi et al. 2008).
Oltre alla mobilità, infatti, la risposta di una specie ittica alla protezione può dipendere
dalla taglia degli individui (Tupper 2007), e dalle caratteristiche dell’habitat
preferenziale (Garcia-Charton e Perez-Ruzafa 1999, Ashworth e Ormond 2005). Inoltre,
dal momento che molte attività di pesca sono altamente selettive sulla taglia degli
individui (es. pesca subacquea; Frisch et al. 2008), le specie ittiche che raggiungono le
maggiori dimensioni potrebbero rispondere maggiormente alla protezione (Mosquera et
al. 2000). Anche l’associazione con le diverse tipologie habitat bentonici può
influenzare la risposta di una specie alla protezione, dal momento che pesci legati al
substrato (bentonici o nectobentonici) vengono più facilmente catturati da alcuni tipi di
reti da pesca rispetto a peci pelagici (Hickford e Schiel 2008). Infine, oltre alle specie
target, le attività di pesca possono impattare anche specie di scarso o nullo valore
commerciale, vittime del cosiddetto bycatch. Pertanto, un’altra possibile fonte di
eterogeneità negli effetti della protezione potrebbero essere le caratteristiche che
rendono una specie non target vulnerabile al bycatch.
Uno studio molto recente (Claudet et al. 2010) ha indagato le variazioni nella risposta
alla protezione delle specie ittiche di dodici AMP mediterranee in relazione al ciclo
vitale e ad alcune caratteristiche ecologiche: taglia massima della specie, preferenze di
habitat (tipo e intervallo di profondità), comportamento gregario. È emerso che:
1)l’effetto protezione sui pesci dipende dal loro valore commerciale e in, in misura
minore, dal fatto che siano o meno vittime del by-catch. All’interno delle AMP si
riscontrano mediamente basse abbondanze di pesci bento-pelagici non sfruttati dalla
pesca. Nonostante la presenza di cascate trofiche sia stata ampiamente documentata per
pesci che predano invertebrati (Micheli et al. 2005, Guidetti 2006, Guidetti e Sala
2007), non ci sono chiare evidenze di cascate trofiche che coinvolgano altri pesci come
prede; quanto osservato, tuttavia, potrebbe riflettere l’instaurarsi di potenziali
interazioni preda-predatore, ipotesi confermata all’elevato numero di pesci di interesse
commerciale piscivori entro i confini delle AMP. Al contrario, specie bentoniche che
non sono direttamente influenzate dalla mortalità da pesca mostrano elevate densità
all’interno delle riserve, suggerendo che possano beneficiare indirettamente della
protezione, per esempio, a seguito del miglioramento della qualità dell’habitat entro le
AMP. 2)La risposta delle specie target della pesca dipende dalla taglia massima

19
raggiunta. Inoltre, le taglie piccole di queste specie sembrano non beneficiare della
protezione: dal momento che i giovanili sono troppo piccoli per essere pescati,
sarebbero state rilevate densità più elevate di questi se le riserve fossero realmente
efficaci nel trattenere le larve degli stock adulti protetti e/o nel favorire il reclutamento,
attraverso, per esempio, l’attrazione tra con specifici, processi realmente dimostrati solo
per alcune specie (rispettivamente Schmitt e Holbrook 1996, Lecchini et al. 2007 e
Swearer et al. 1999, Almany et al. 2007). 3)Specie di interesse commerciale in grado di
spostarsi su ampi intervalli di profondità, ma che si ritrovano anche a batimetriche
inferiori ai -10m, mostrano una più forte risposta alla protezione rispetto a quelli
caratterizzati da range di profondità più ristretti (compresi tra -50 e -10 m); quanto
osservato potrebbe dipendere dal fatto che la pesca artigianale e ricreativa esercitano
una maggiore pressione sui popolamenti ittici presenti alle quote minori. 4) Il risultato
più interessante, tuttavia, è che la protezione fornisce benefici alle specie target della
pesca molto vagili, sia bentoniche che bento-pelagiche, qualunque sia il loro home
range e tasso di spostamento annuale. Una possibile spiegazione per quanto osservato è
che la protezione aumenti la qualità dell’habitat entro la riserva, riducendo così la
tendenza dei pesci a uscire dalla riserva stessa (Rodwell et al. 2003). Inoltre, le
aumentate densità di pesci bentonici non target entro le AMP potrebbero agire da
sorgente di potenziali prede per le specie più mobili, aiutando a trattenerle nella riserva
(Steward e Jones 2001, Beukers-Steward e Jones 2004). Altri fattori oltre alle risorse,
infine, possono indurre complessi pattern comportamentali (Roshier et al. 2008) e la
pressione di pesca al di fuori della riserva può attrarre determinate specie al suo interno
(Eggleston e Parson 2008). Inoltre stimoli sensoriali provenienti da conspecifici
possono aumentare le probabilità che certi pesci molto vagili rimangano entro le AMP
piuttosto che muoversi verso altre aree soggette a pesca, dove la competizione può
essere minore, ma i tassi di mortalità sono più altri come conseguenza delle attività di
pesca.
1.2.4.5 Eterogeneità spaziale È ampiamente riconosciuto che le comunità ecologiche presentano una distribuzione a
chiazze, frammentata che si rende evidente su numerose e, spesso, simultanee scale
spaziali e temporali (Dayton e Tegner 1984, Kotliar e Wiens 1990). I fattori che
controllano le variazioni spaziali e temporali nella struttura di comunità (composizione
in specie e ricchezza specifica, organizzazione trofica) possono essere sia fisici

20
(struttura dell’habitat, disponibilità di luce e nutrienti, esposizione a correnti, onde e
maree), sia biologici (reclutamento, predazione, competizione, mutualismo, disturbo)
(García–Charton e Pérez–Ruzafa 1999). In particolare, variazioni nelle modalità di
distribuzione spaziale e temporale nelle abbondanze dei popolamenti ittici associati a
coste rocciose sono stati attribuiti a diversi fattori fisici e/o biologici (Jones 1991):
variazioni di profondità (Bell 1983, Harmelin 1990, McGehee 1994, García-Charton e
Pérez-Ruzafa 1998); strutturazione dell’habitat con conseguente differente disponibilità
di risorse come cibo e spazio (García-Charton et al. 2000); differenze climatiche
(Holbrook et al. 1997); predazione (Hixon 1991); competizione (Sale 1978, Gladfelter
et al. 1980); disturbo sporadico (Walsh 1983, Chabanet et al. 1995); dinamiche larvali
(Leis e McCormick 2002); variabilità nei tassi di reclutamento (Booth e Brosnan 1995).
Ciascuno di questi fattori agisce preferenzialmente su una sola scala spaziale (Sale
1998).
La frammentazione intrinseca agli ecosistemi, quindi, è il risultato dell’interazione tra le
variazioni dei fattori ambientali e le risposte degli organismi ad esse nello spazio e nel
tempo (Milne 1991). Questa eterogeneità ecologica ha il ruolo potenziale di modificare
le conseguenze di un processo ecologico (Wiens 1976, Levin 1992, Oksanen et al.
1992), influenzando così il modo con cui le diverse specie coesistono nello spazio e nel
tempo (Downing 1986) o come queste sono organizzate nelle reti trofiche (Schoener
1989, Polis e Strong 1996). Pertanto, lo studio dei pattern spaziali e temporali degli
organismi rappresenta un passo fondamentale sulla strada della comprensione delle
cause della distribuzione e dell’abbondanza degli organismi (Levin 1992) e permette di
gettare le basi per il monitoraggio a lungo termine dei cambiamenti dovuti sia al
disturbo naturale che a quello antropico (Underwood 1990).
Nel caso dello studio degli effetti della protezione sui popolamenti ittici costieri, si è
interessati soprattutto all’influenza dell’ambiente fisico, quello che Bell et al. (1991)
hanno definito “struttura dell’habitat”, che può essere fonte di confusione nel valutare il
cosiddetto “effetto riserva”. La struttura dell’habitat ha effetti su molteplici processi
ecologici, da quelli che agiscono sull’individuo, a quelli che interessano l’intera
comunità. A livello individuale, il comportamento (Lejeune 1984, Polunin e Klumpp
1989, Jan e Chang 1993), la fisiologia (Taborsky e Limberger 1980, Kotrschal 1983,
McFarland 1986, Boujard e Leatherland 1992) e la morfologia (Anderson 1990,
Turingan et al. 1995, Wainwright e Richard 1995) aiutano l’ecologo a capire come

21
l’individuo, che è limitato dalla propria struttura fisica (Schoener 1989), sceglie le prede
e il substrato per riprodursi, sfugge ai predatori, alle condizioni ambientali avverse e
alla pressione di pesca in presenza di variazioni su piccola scala dell’habitat. A livello
di popolazione, la struttura dell’habitat fornisce le basi meccanicistiche (Schoener 1986,
Inchausti 1994) per spiegare pattern spaziali e temporali su piccola scala di
distribuzione degli organismi. La frammentazione spaziale dell’habitat permette anche
di spiegare eventuali sovrapposizioni di nicchia e ripartizione delle risorse in termini
spaziali da parte di specie appartenenti a una stessa area. Numerosi studi sul
reclutamento di pesci di scogliere coralline, fenomeno cruciale nel determinare la
struttura di popolazione delle specie ittiche (Booth e Brosnan 1995), hanno dimostrato
che l’estrema variabilità su multi scale spaziali e temporali associata a tale processo è
strettamente correlata a cambiamenti nella struttura dell’habitat (Doherty 1991, Sale
1991 e 1999, Fowler et al. 1992, Planes et al. 1993, Caselle e Warner 1996). Infine, la
frammentazione dell’habitat introduce complessità nello studio di processi interattivi
come competizione e predazione, delle interazioni trofiche e degli effetti delle Aree
Marine Protette (García-Charton e Pérez-Ruzafa 1999). Difficoltà nel distinguere tra
variabilità naturale spaziale e/o temporale dei popolamenti ittici ed ”effetto riserva”
possono essere, pertanto, legate alla eterogeneità degli habitat marini costieri.
Analizzando il problema dal punto di vista dell’ecologia del paesaggio, l’ambiente
marino può essere considerato un sistema gerarchico, i cui componenti, organizzati in
livelli in base a scale funzionali specifiche, si rendono evidenti sottoforma di patch in
natura (Urban et al. 1987, Turner 1989). La frammentazione gerarchica dell’habitat,
pertanto, impone alcune regole, nel momento in cui si vogliano valutare gli effetti della
protezione nelle AMP. 1) È necessario scegliere le adeguate scale spaziali e temporali di
osservazione. 2) Ad ogni scala considerata, occorre misurare l’eterogeneità dell’habitat
in modo da controllare l’interferenza tra “effetto riserva” ed “effetto habitat” (García-
Charton e Pérez-Ruzafa 1999). Tuttavia, solo in pochi studi, volti a valutare gli effetti
delle AMP, l’eterogeneità spaziale è stata considerata come fattore di confusione.
Nel lavoro di García-Charton et al. 2004, ad esempio, emerge l’importanza della
struttura dell’habitat nella spiegazione della variabilità dei popolamenti ittici
mediterranei, in termini di abbondanza, ricchezza e composizione in specie, in
particolare quella su piccola scala spaziale. Tale variabilità rappresenta una fonte di
possibile confusione nello studio degli effetti della protezione sui popolamenti ittici, per

22
diverse ragioni: da un lato, molte AMP vengono istituite in aree che ospitano habitat
strutturalmente complessi, che favoriscono lo sviluppo una ricca fauna ittica (García-
Charton et al. 2004), dall’altro zone soggette a protezione possono escludere tratti di
costa caratterizzati da particolari caratteristiche ambientali.
Benedetti-Cecchi e altri autori (2003) hanno analizzato il problema di separare l’”effetto
protezione” dall’”effetto habitat” dal punto di vista della comunità bentonica nella AMP
dell’Arcipelago Toscano. Tale studio ha messo in luce che i pattern di variabilità
spaziale riscontrati erano nella maggior parte dei casi inconsistenti con l’estensione
della riserva marina; anzi supportavano il modello secondo il quale è l’eterogeneità
ambientale a determinare le modalità dei distribuzione degli organismi osservate.
Se consideriamo la struttura dell’habitat dal punto di vista funzionale, cioè definiamo
l’habitat come qualunque attributo fisico o biologico dell’ambiente che mette a
disposizione degli organismi oggetto di studio alcune risorse come cibo o spazio a una
data scala (McCoy e Bell 1991, Jones e Andrew 1992), è giusto chiedersi quali
caratteristiche dell’habitat sono importanti per questi organismi e quali sono le risposte
di tali organismi all’eterogeneità spaziale e/o temporale di uno degli attributi
dell’habitat. Pertanto, una reale verifica dell’influenza della variabilità dell’habitat deve
essere una componente fondamentale di ogni studio volto a discriminare gli effetti della
protezione da altre fonti di eterogeneità (García-Charton et al. 2000).
1.2.5 Aspetti sociali ed economici della conservazione Accurati studi biologici ed ecologici, profonda conoscenza delle aree da proteggere e
disegni sperimentali adeguati rappresentano i presupposti fondamentali per l’istituzione
e la gestione di AMP efficaci.
Bisogna però tenere in considerazione il fatto che le AMP rappresentano un esempio di
Sistema Socio-Ecologico (SSE): un sistema adattativo complesso che richiede una
gestione dinamica, flessibile ed adattativa (Levin, 1998). Si tratta, infatti, di un sistema
naturale ed antropico in cui l’uomo non è solo presente, ma gioca un ruolo da
protagonista nella sua gestione e fruizione. Nella gestione di questi sistemi e della loro
complessa interazione con i comparti sociali ed economici, diventano sempre più utili i
principi ispiratori della “Gestione Adattativa” introdotta da Holling nel 1978, un
processo attraverso cui le disposizioni istituzionali e le conoscenze ecologiche sono

23
testate attraverso una procedura dinamica di “learning by doing”, apprendere
dall’esperienza (Folke et al., 2002). Questo tipo di gestione può essere, quindi, definita
“adattativa” perché riconosce, da un lato, che le risorse gestite si modificano a causa
dell’intervento umano e, dell’altro, che la gestione avviene in condizioni di incertezza
dovendo continuamente affrontare le novità del sistema stesso (Gunderson, 1999).
Pertanto, altri fattori a carattere economico, culturale e sociale, contribuiscono in
maniera preponderante al successo di una AMP:
1. la coerenza tra gli obiettivi della gestione e le effettive disponibilità economiche
dell’Ente Gestore, da cui dipendono gli strumenti adottati allo scopo di garantire
una effettiva applicazione delle disposizioni messe in atto;
2. il supporto di leggi ed istituzioni: nonostante sia difficile coniugare leggi
regionali, nazionali e comunitarie spesso contradditorie o facilmente eludibili,
l’Ente Gestore deve costruire un Piano di Gestione chiaro e a disposizione di
tutti;
3. il coinvolgimento delle comunità costiere interessate dall’istituzione dell’AMP
mediante opere di sensibilizzazione ed educazione ambientale, intesa come
strumento teso a facilitare la comprensione del valore della riserva: i primi utenti
di un area protetta, ovvero i cittadini, non sono da vedere come una collettività
da persuadere in merito all’istituzione dell’AMP, bensì come membri di un
organo decisionale. È ormai evidente l’impossibilità di gestire un’AMP con la
forza, senza il consenso, il coinvolgimento e il supporto attivo di coloro che per
primi risentono della presenza di una riserva marina (Beaumont, 1997);
4. iniziative tese a compensare le perdite causate ai pescatori e a spiegare i
possibili benefici pratici derivanti da un’AMP quali aumento della resa del
pescato nelle zone adiacenti alla riserva, incremento delle attività ricreative
legate a un turismo ecocompatibile.
Le riserve marine, quindi, pur non rinunciando a raggiungere obiettivi di conservazione
e protezione ambientale, devono essere progettate tenendo conto:
delle attività economiche presistenti l’istituzione dell’AMP (ad esempio
pesche tradizionali e speciali);
dell’esistenza di particolari rilevanze storiche, archeologiche, spirituali o
culturali, la cui conservazione può essere coniugata con la salvaguardia
dell’ambiente marino;

24
della possibilità di associare la salvaguardia ambientale ad attività
ricreative sostenibili come immersioni subacquee e snorkeling,
adeguatamente controllate e monitorate.
Infine, le zone a protezione parziale possono diventare i laboratori per la
sperimentazione di sistemi di pesca sostenibili ed ecocompatibili: metodi di prelievo
delle risorse ittiche molto selettivi e poco invasivi nei confronti delle esigenze di
mantenimento degli stock. Spesso, queste attività ben si coniugano con le pesche
tradizionali operate per secoli sulle coste italiane e possono rappresentare la via di
riscatto dell’economia delle popolazioni costiere interessate dall’istituzione di un’AMP
e di riqualificazione dell’intera zona (Vierucci et al., 2006).
I pescatori, da considerare anch’essi ‘specie’ da proteggere in una riserva, possono
iniziare un nuovo “lavoro” nel rispetto dell’ambiente marino, delle sue forme di vita e
dell’antica tradizione marinara e peschereccia italiana. Queste iniziative, inoltre, hanno
enorme eco dal punto di vista dell’attrattiva turistica: in Italia vi sono già molti esempi
di “pesca-turismo”, attività che aumentano il prestigio di una località costiera agli occhi
della popolazione sensibile alle problematiche ambientali e alla salvaguardia del
patrimonio naturale.
La missione di un’AMP, quindi, non è solo la tutela di habitat e specie marine, ma
anche la valorizzazione di aspetti culturali, sociali ed economici delle realtà locali. Le
Aree Marine Protette, coniugando protezione ambientale, sviluppo economico
sostenibile e turismo ecocompatibile, devono diventare modelli di promozione
dell’educazione ambientale e delle culture tradizionali locali.

25
1.3 Caratteristiche dei popolamenti ittici protetti del Mediterraneo La maggior parte delle Aree Marine Protette del Mediterraneo comprende tratti di
fondale generalmente costituiti da un mosaico di substrato roccioso intersperso a
estensioni di sabbia e ciottoli e a patch di praterie di Posidonia oceanica. Pertanto, i
microhabitat che generalmente si incontrano sono rappresentati da:
− infralitorale roccioso: la cintura a Cystoseira, che risente di occasionali
emersioni, è caratterizzata dalla presenza di poche specie ittiche, tra le quali si
possono annoverare quelle appartenenti alle famiglie di blennidi, clinidi e
tripterigidi; nella colonna d’acqua antistante ai fondi ad alghe fotofile, invece,
sono presenti specie ittiche appartenenti a tutte le categorie trofiche, sia stadi
giovanili che adulti. Le specie di Labridi, Labrus viridis, Synphodus tinca, Coris
julis e Thalassoma pavo sono le più cospique. Molto comuni sono anche i
serranidi, come Serranus scriba e S. cabrilla, e diversi sparidi, come i saraghi
(Diplodus sargus, D. vulgaris, D. puntazzo), le occhiate (Oblada melanura) e le
salpe (Sarpa salpa).
− coralligeno e grotte: pesci caratteristici di questi ambienti scarsamente illuminati
sono cernie brune (Epinephelus marginatus) e grossi predatori come dentici
(Dentex dentex), corvine (Sciena umbra) e mostelle (Phycis phycis), ma anche re
di triglie (Apogon imberbis) e castagnole rosse (Anthias anthias).
− fondi sabbiosi: popolati da specie criptiche come sogliole (famiglia soleidi) e
rombi (famiglia botidi) e tracine (famiglia trachinidi) e da specie vagili in cerca
di cibo, come salpe, mormore (Lithognatus mormyrus) e orate (Sparus aurata).
− praterie di Posidonia oceanica: costituiscono ambienti tridimensionali che
rappresentano l’habitat ideale per numerose specie ittiche. Si possono
annoverare molti pesci planctonofaghi, come castagnole (Chromis chromis),
presenti nella colonna d’acqua di giorno e tra le fronde di notte, zerri (Spicara
smaris), menole (Spicara maena), boghe (Boops boops) e occhiate (Oblada
melanura). Tra i pesci che si possono riscontrare sulle praterie di P. oceanica vi
sono anche i cefali (come Liza aurata), caratterizzati da una dieta almeno
parzialmente costituita da detrito, e numerosi predatori come dentici (Dentex
dentex), barracuda (Sphyraena viridensis e S. Sphyraena), le ricciole (Seriola

26
dumerilii) e spigole (Dichentrarchus labrax). Molte specie ittiche vivono più
strettamente associate al manto fogliare di P. oceanica. Si tratta di specie che
nuotano poco al di sopra delle foglie e/o tra le foglie, come molti pesci
appartenenti alla famiglia dei labridi: tra questi il tordo nero (Labrus merula) e il
tordo marvizzo (Labrus viridis). A questi si aggiungono pesci come il tordo
pavone (Symphodus tinca), la donzella (Coris julis), il tordo ocellato
(Symphodus ocellatus) e la donzella pavonina (Thalassoma pavo). Un labride
piuttosto comune su P. oceanica è il tordo codanera (Symphodus melanocercus):
questo pesce di piccole dimensioni (lunghezza massima pari a 14 cm) è un
caratteristico pesce pulitore che interagisce con tantissime altre specie ittiche, tra
cui il tordo pavone, diversi saraghi (sparidi del genere Diplodus) e piccoli
serranidi come Serranus scriba e Serranus cabrilla. Altri labridi di piccole
dimensioni che comunemente si possono trovare su P. oceanica appartengono
sempre al genere Symphodus e sono essenzialmente pesci carnivori che si cibano
di piccoli invertebrati vagili (es. echinodermi, molluschi, policheti, crostacei)
che si trovano tra le foglie, sui rizomi e nel sedimento alla base dei fasci fogliari.
Tra questi ricordiamo il tordo fasciato (Symphodus doderleini), il tordo rosso (S.
mediterraneus), il tordo musolungo (S. rostratus) e il tordo verde (S. roissali). Il
tordo grigio (S. cinereus), per contro, lo si ritrova più comunemente nei tratti di
fondale sabbioso confinanti con P. oceanica o nei pressi delle praterie in cui si
accumula detrito fogliare. Molte sono le specie di sparidi che si possono trovare
associate al manto fogliare di P. oceanica. Prima di tutte va rammentata la salpa
(Sarpa salpa), che rappresenta la più importante specie ittica essenzialmente
erbivora (almeno da adulta) del sistema litorale mediterraneo. Le salpe, che
possono raggiungere fino a 50 cm di taglia massima, sono pesci che sovente
formano banchi costituiti anche da centinaia di individui; tale gregarismo si
riscontra sia nei giovanili (che hanno una dieta onnivora), sia negli adulti. Tra
gli sparidi del genere Diplodus (che include pesci comunemente definiti come
saraghi), lo sparaglione (D. annularis) è sicuramente quello che più di tutti si
trova associato a P. oceanica. Infine, a ridosso delle praterie nuotano orate
(Sparus aurata), tanute (Spondiliosoma cantharus) e pagelli fragolini (Pagellus
erythrinus).

27
1.4 Obiettivi Le AMP sono globalmente riconosciute come uno dei migliori strumenti volti alla
protezione della biodiversità, alla conservazione degli ecosistemi marini e alla gestione
delle risorse alieutiche. Alla luce di quanto discusso, tuttavia, l’efficacia dell’effetto
riserva deve essere valutata tenendo in considerazione alcuni fattori di fondamentale
importanza: la dimensione e l’età della riserva (e con essa l’effettiva messa in atto delle
misure di protezione), le caratteristiche ecologiche delle specie protette, l’inclusione o
meno in una rete di riserve e l’eterogeneità dell’habitat. In particolare, la
frammentazione degli habitat naturali rappresenta un importante fattore di confusione
che dovrebbe essere sempre considerato se si vogliono esaminare gli effetti di una
AMP; tuttavia, raramente figura negli studi presenti in letteratura.
Il presente lavoro di tesi si inserisce in questo contesto e si propone di analizzare ipotesi
multiple circa i rapporti tra processi spaziali, eterogeneità ambientale e protezione,
presso alcune isole del Parco Nazionale dell’Arcipelago Toscano (Capraia, Giannutri,
Montecristo e Pianosa), attraverso un campionamento della fauna ittica (effettuato
tramite tecniche non distruttive di visual census) appropriato per comparare la struttura
dei popolamenti ittici, le densità e le distribuzioni di taglia di singole specie target
all’interno ed all’esterno delle AMP. Le variabili predittive considerate nell’analisi
sono:
• per quanto riguarda la protezione, aree protette vs controlli e isole totalmente
protette vs isole parzialmente protette;
• coordinate geografiche, relative ai processi spaziali;
• indici di complessità del substrato e della costa, per tener conto dei processi
ambientali.
L’analisi è stata condotta avvalendosi di GAMM (Generalized Additive Mixed
Models), che permettono di modellizzare in maniera flessibile gli effetti delle covariate
(in questo caso le variabili predittive relative ai processi spaziali e ambientali)
utilizzando come predittore lineare una combinazione additiva di funzioni non
parametriche delle covariate ed effetti di fattori casuali (qui il tempo) (Zhang e Lin
2003).

28
Se l’AMP offre una protezione efficace ai popolamenti ittici, deve essere allora
possibile evidenziarne l’effetto anche dopo aver controllato statisticamente l’influenza
di processi ambientali e spaziali. In questo lavoro è stata considerata in modo esplicito
l’influenza della eterogeneità dell’habitat alle scale della costa (centinaia di metri) e del
substrato (decine di metri), oltre alla posizione geografica. Quest’ultima riassume
l’influenza di processi spaziali quali differenze climatiche tra isole, caratteristiche
oceanografiche e altre sorgenti di variabilità non identificate. E’ possibile che tali
processi siano più importanti degli effetti delle AMP nel determinare le modalità di
distribuzione e abbondanza delle specie ittiche costiere. In questo caso l’effetto riserva
verrebbe meno una volta che l’influenza degli altri processi è stata controllata nelle
analisi. Ciò indicherebbe la necessità di modificare gli schemi di protezione oggi
applicati.
Le isole oggetto di studio sono caratterizzate da due diversi regimi di tutela:
Montecristo e Pianosa sono interamente soggette a protezione, mentre Capraia e
Giannutri presentano un’alternanza di zone protette e non protette. Inoltre, Pianosa e
Montecristo sono protette da più tempo rispetto a Capraia e Giannutri, in quanto riserva
naturale la prima e isola adibita a carcere fino al 1998 la seconda. Pertanto, si propone
che l’alternanza di aree soggette a protezione e aree impattate dalla pesca generi
modalità di distribuzione nelle abbondanze dei popolamenti ittici che differiscono da
quelle che si originano nelle isole protette totalmente e da lungo tempo. In particolare, si
può ipotizzare che isole totalmente protette siano più efficaci di isole sottoposte a
differenti regimi di protezione, una volta isolati gli effetti di processi spaziali e
ambientali. E’ da notare, tuttavia, che, in questo studio, l’effetto dell’età della AMP non
può essere isolato dalla modalità di protezione (totale o parziale).

29
2. MATERIALI E METODI
2.1. L’Arcipelago Toscano
L’arcipelago toscano (42° 45’ N, 10° 15’ E) comprende sette isole di dimensioni e
storia geologica differenti. Queste isole rappresentano il più grande sistema di Aree
Marine Protette dell’Europa, con i loro 61.474 Km2 di acque protette compresi nel
perimetro del Santuario Internazionale per la tutela dei mammiferi marini, “Pelagos”. Il
parco nazionale è stato istituito nel 1996 (D.P.R. 22/7/96) con l’obiettivo di preservare
gli habitat naturali, ma anche promuovere un turismo eco-compatibile. Tuttavia, è bene
ricordare, che l’accesso al pubblico su Gorgona e Pianosa era proibito già da prima del
1996, in quanto sede di prigioni, mentre Montecristo è tutelata come riserva naturale dal
1971.
Le sette isole dell’Arcipelago Toscano sono caratterizzate da ambienti naturali
estremamente diversificati dovuti ad una storia geologica piuttosto complessa: Capraia
ha origine vulcanica; Gorgona è formata da rocce metamorfiche; Pianosa da rocce
Figura 1.1 – Cartina dell’Arcipelago Toscano

30
sedimentarie e da accumuli conchiliferi che racchiudono fossili marini; Giannutri da
sedimenti calcareo-dolomitici; Montecristo è interamente granitica, come quasi tutta
l’isola del Giglio, mentre l’Elba è la più varia dal punto di vista geomorfologico e
comprende, nella parte occidentale, il massiccio del Monte Capanne (m. 1018), la più
alta vetta dell’Arcipelago e, nella parte orientale, aree minerarie ricche di ferro.
L’assenza di corsi d’acqua e la dominanza di venti da ovest verso est garantiscono alle
acque dell’Arcipelago Toscano una bassa torbidità e il mantenimento di fluttuazioni
temporali nei valori di salinità entro i limiti determinati dai maggiori eventi
oceanografici. I fondali dell’arcipelago includono una vasta gamma di habitat lungo una
grande scala spaziale (Km). I fondali rocciosi sono la tipologia di habitat maggiormente
rappresentata, costituendo il 90% delle aree costiere. Spiagge sabbiose si incontrano al
Giglio e all’Elba e in quest’ultima sono presenti anche fondi fangosi. Caratteristiche del
subtidale sono grandi estensioni di praterie di Posidonia Oceanica che si alternano a
tratti di fondale roccioso ed aree sabbiose. Su alcune isole, Gorgona ad esempio, al di
sotto dei 40m di profondità si incontra il caratteristico habitat costituito da alghe
calcaree mobili, note con il nome di maerl. Gli organismi, inoltre, sperimentano una
elevata variabilità spaziale su piccola scala dovuta a cambiamenti nelle caratteristiche
fisiche e biologiche dell’ambiente. Per esempio, l’alternarsi di popolamenti algali
canopy-forming con alghe corallinacee incrostanti e turf-forming fornisce differenti
microhabitat per numerose specie di invertebrati così come per altre specie algali,
mentre substrati rocciosi continui alternati a franate rocciose supportano la presenza di
differenti assemblaggi di specie ittiche.
Per quanto riguarda la flora e la fauna, l’Arcipelago toscano rappresenta un hot-spot di
diversità poiché ospita numerosi endemismi e ricche popolazioni di specie altrove
diventate rare, come l’alga bruna Cystoseira sp., la Patella ferruginea, il gasteropode
predatore Stramonita haemastoma e il granchio Eriphia verrucosa.
31
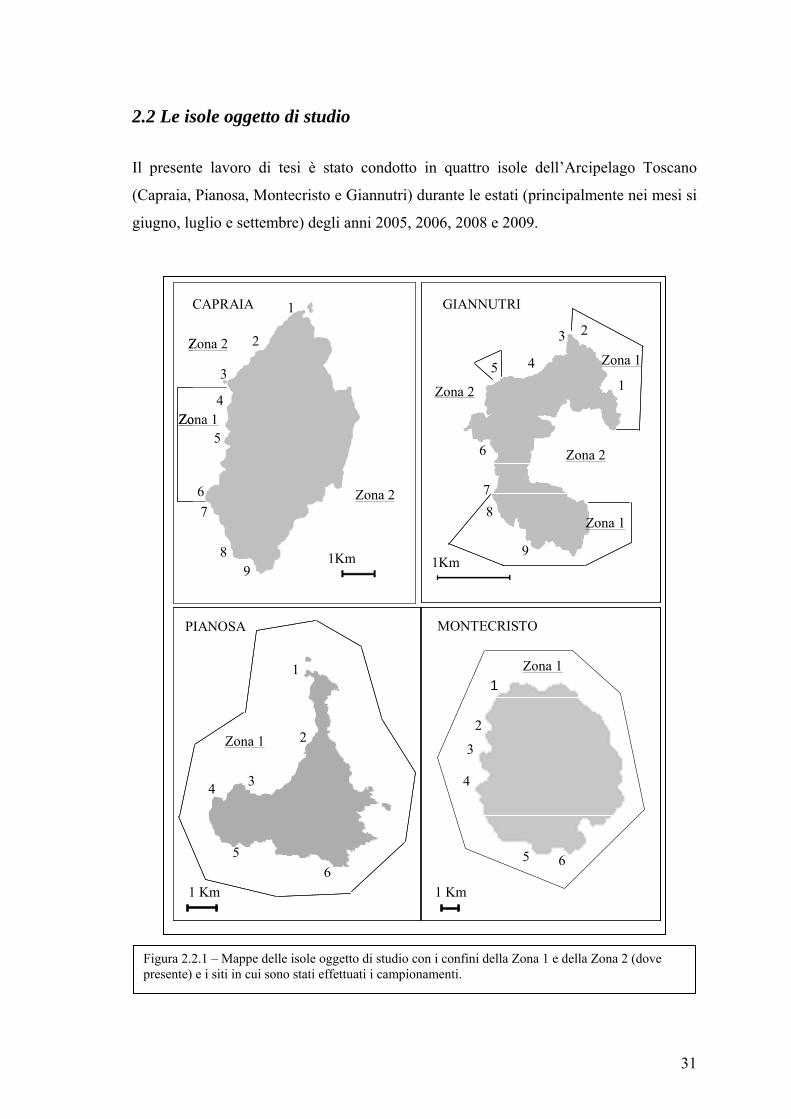
2.2 Le isole oggetto di studio
Il presente lavoro di tesi è stato condotto in quattro isole dell’Arcipelago Toscano
(Capraia, Pianosa, Montecristo e Giannutri) durante le estati (principalmente nei mesi si
giugno, luglio e settembre) degli anni 2005, 2006, 2008 e 2009.
1
23
45
6
78
91Km
Zona 2
Zona 1
Zona 1
1
2
3
4
5
67
89
1Km
Zona 1
Zona 2
Zona 2
Zona 2
CAPRAIA GIANNUTRI
1 Km
1
2
3
4
5 6
Zona 1
MONTECRISTOPIANOSA
1 Km
1
2
34
56
Zona 1
1
23
45
6
78
91Km
Zona 2
Zona 1
Zona 1
1
2
3
4
5
67
89
1Km
Zona 1
Zona 2
Zona 2
Zona 2
CAPRAIA GIANNUTRI
1
23
45
6
78
91Km
Zona 2
Zona 1
Zona 1
1
2
3
4
5
67
89
1Km
Zona 1
Zona 2
1
23
45
6
78
91Km
Zona 2
Zona 1
Zona 1
1
2
3
4
5
67
89
1Km
Zona 1
Zona 2
1
2
3
4
5
67
89
1Km
Zona 1
Zona 2
Zona 2
Zona 2
CAPRAIA GIANNUTRI
1 Km
1
2
3
4
5 6
Zona 1
MONTECRISTOPIANOSA
1 Km
1
2
34
56
Zona 1
1 Km
1
2
3
4
5 6
Zona 1
MONTECRISTOPIANOSA
1 Km
1
2
34
56
Zona 1
1 Km
1
2
3
4
5 6
Zona 1
MONTECRISTO
1 Km
1
2
3
4
5 6
Zona 1
MONTECRISTOPIANOSA
1 Km
1
2
34
56
Zona 1
PIANOSA
1 Km
1
2
34
56
Zona 1
Figura 2.2.1 – Mappe delle isole oggetto di studio con i confini della Zona 1 e della Zona 2 (dove presente) e i siti in cui sono stati effettuati i campionamenti.

32
Capraia
Capraia ha una superficie di 19,3 km2 ed è per estensione la terza isola dell'arcipelago.
E' fra le realtà insulari più marittime, essendo situata a ben 54 km dalla costa
continentale ad Est, mentre la Corsica dista circa 31 km ad Ovest. Capraia è l'unica
isola vulcanica dell'Arcipelago: si è formata circa nove milioni di anni fa con la nascita
di un cono eruttivo successivamente fratturatosi con lo sprofondamento della parte
occidentale, per cui oggi permane solo una scheggia della più antica struttura
magmatica. Quasi contemporaneamente avvenne lo sviluppo di un più piccolo camino
vulcanico presso l'odierna Punta dello Zenobito, dove le rocce rosse che circondano la
penisola rappresentano il susseguirsi delle antiche eruzioni.
Capraia Isola è un Comune autonomo in Provincia di Livorno con una popolazione di
circa trecento abitanti. Due sono i nuclei abitati che insistono presso la più vasta
insenatura isolana: il porto ed il paese.
Zone tutelate:
• A terra: quasi tutto il territorio (zona 2), eccetto il centro abitato. Sono tutelati anche
gli isolotti La Peraiola, Le Formiche, Lo Scoglione, lo Scoglio del Gatto, e lo Scoglio
della Manza.
• A mare: una parte di zona 1, compresa tra punta della Manza e Punta del Trattoio, il
resto zona 2 eccetto la zona libera compresa tra la Punta del Porto Vecchio e quella
della Bellavista (di fronte all'abitato). In zona 1 sono vietati l’accesso, la navigazione, la
sosta, l’ancoraggio, la pesca e l’immersione. In zona 2 la pesca è regolamentata
dall’Ente Parco.
Pianosa
Pianosa è la più bassa isola dell'arcipelago, praticamente un pianoro alto al massimo 29
metri sul livello del mare, situata a 14 km a S-SO dalle coste dell'Isola d'Elba e a circa
27 km a S-SE dalla più meridionale isola di Montecristo. L'isola ha una superficie di
10,25 km2, un'estensione costiera di 26 km ed è inserita nel territorio del Comune di
Campo nell'Elba. Nonostante sia poco elevata, presenta coste rocciose con presenza di
falesie, specialmente nel versante occidentale e pochissime spiagge. Le rocce sono di
origine sedimentaria, con uno strato inferiore argilloso risalente al Miocene e da strati di

33
calcari organogeni del Pliocene ricchissimi di fossili marini, testimonianza della
sedimentazione avvenuta su un antico fondale.
Dalla dismissione del carcere avvenuta nel 1997 l'isola è quasi disabitata: le vecchie
diramazioni carcerarie sono oggi vuote ed anche il piccolo nucleo urbano posto sul
versante orientale di Pianosa è privo di abitanti: vi rimane un piccolo presidio della
polizia penitenziaria e del corpo forestale dello stato.
Zone tutelate:
• A terra: il 100% del territorio (zona 1), inseriti nel perimetro del parco sono anche gli
isolotti La Scarpa e La Scola.
• A mare: il 100% fino a un miglio dalla costa (zona 1).
Montecristo
Montecristo, con una superficie di 10,4 km2 e uno sviluppo costiero di 16 km, è fra le
più meridionali isole dell'Arcipelago Toscano, annessa al territorio comunale di
Portoferraio. Montecristo dista circa 63 km dalle coste dell'Argentario ad Est, circa 45
km dalle coste dell'Elba meridionale situata a Nord, 43 dal Giglio (Est) e 27 km da
Pianosa (NN-O). Dalla pianta pressoché circolare, frastagliata da alcune insenature che
insistono in prossimità delle principali vallate, Montecristo è un cono granitico che
s'innalza fino 645 metri sul livello del mare culminando nel Monte della Fortezza, con
una piccola cresta d'alture che prosegue verso sud fino alla Cima dei Lecci a 563 metri.
Il paesaggio è caratterizzato dai grandi liscioni granitici che scendono ripidamente al
mare, con poca vegetazione ed alcune vallate scavate dall’azione di piccoli corsi
d'acqua dal regime stagionale. Dal punto di vista geologico l'isola è costituita quasi
interamente da un plutone granitico originatosi tra i sette e i cinque milioni di anni fa.
Sede fino al XVI secolo da una fiorente comunità monastica, l’isola è oggi quasi
disabitata, dal momento che è stata istituita Riserva Naturale Integrale con decreto
ministeriale del 4 marzo 1971 e Riserva Naturale Biogenetica diplomata dal consiglio
d'Europa dal 1988. L'odierna popolazione consiste nella famiglia del guardiano e da due
agenti del Corpo Forestale dello Stato, i cui alloggi sono situati nei pressi della Villa
Reale di Cala Maestra, unico approdo di Montecristo.
Zone tutelate:

34
• A terra: il 100% del territorio (zona 1); inserito nel perimetro del Parco è anche lo
Scoglio d'Africa o Formica di Montecristo, il cui territorio è tutelato al 100%.
• A mare: il 100% fino a 1 km dalla costa (zona 1).
Giannutri
E' la più meridionale delle isole di Toscana essendo situata a 18 km a S-SO da Porto
Ercole, lungo la costa meridionale del Monte Argentario, e a 14 km a SE dell'Isola del
Giglio. Risulta la penultima isola per estensione territoriale con una superficie di 2,6
km2. La sua lunghezza massima da Nord a Sud è di circa 2,5 km mentre la larghezza
massima è appena 500 m. Due sono gli approdi situati in prossimità delle principali
insenature dell'isola: Cala Spalmatoio, sul versante orientale dell'isola, esposta a Sud-
Est, e Cala Maestra, dalla parte opposta, esposta a Nord Ovest. Il territorio isolano
presenta una natura selvatica e rocciosa, caratterizzata da calcari cavernosi, antiche
rocce sedimentarie ricoperte da una fitta macchia mediterranea.
A Giannutri non esistono nuclei abitati ma solo insediamenti sparsi rappresentati per lo
più da residenze turistiche e seconde case frequentate per lo più nel periodo estivo.
Zone tutelate:
• A terra: il 100% del territorio (zona 2).
• A mare: in parte (zona 1 e zona 2), esclusi due corridoi di accesso per le imbarcazioni.
In zona 1 sono vietati l’accesso, la navigazione, la sosta, l’ancoraggio, la pesca e
l’immersione. In zona 2 la pesca è regolamentata dall’Ente Parco.
2.3 Disegno di campionamento
L’analisi di ipotesi formali in ecologia è complicata dalla elevata variabilità dei sistemi
naturali. Un corretto disegno sperimentale garantisce che la variabilità non controllata
(la variabilità che non è associata ad alcuna delle ipotesi in esame) sia equamente
distribuita tra i fattori presi in considerazione, in modo tale che il loro effetto possa
essere separato da altre sorgenti di variazione.
Due sono le condizioni necessarie affinché l’inferenza statistica produca dati
interpretabili:

35
1. i dati forniscono stime rappresentative della variabile di risposta;
2. l’esperimento non confonde l’effetto delle variabili predittive con altre sorgenti
di variabilità indesiderate ed incontrollate. Il disegno sperimentale stabilisce i
criteri necessari affinché tali condizioni siano garantite in ogni particolare
circostanza.
Una volta identificata la variabile (o le variabili) rilevanti per il problema affrontato,
siano esse misure di abbondanza di singole specie, di struttura di popolamenti oppure
misure di eterogeneità nello spazio e nel tempo, è opportuno identificare il contesto
spaziale e temporale nel quale le misurazioni devono essere condotte. A questo scopo è
di primaria importanza identificare l’area ritenuta esposta ad una qualche forma di
gestione umana (quindi anche un’area protetta) ed opportune aree di riferimento non
soggette alla particolare forma di gestione, ma con le stesse caratteristiche generali
(tipologia di popolamenti ed habitat) di quelle soggette ad influenza umana.
Le AMP, tuttavia, sono spesso scelte per le loro caratteristiche di unicità (ad esempio la
presenza di particolari specie, hot spot di diversità, aree di riproduzione) e
l’individuazione di aree di riferimento appropriate può essere difficoltosa (Glasby e
Underwood, 1998). La scelta delle aree di riferimento deve sottostare ai seguenti criteri
(Underwood, 1992; Chapman et al., 1995; Glasby, 1998):
− non devono essere influenzati dalla protezione;
− devono avere le stesse caratteristiche di habitat di quelle dell’area protetta;
− devono infine tenere conto di differenti scale spaziali nel caso in cui non sia
conosciuta la scala spaziale interessata dalla protezione.
La maggior parte dei disegni di campionamento alla base dei programmi di
monitoraggio sperimentale sono stati sviluppati per esaminare il possibile effetto
negativo di attività antropiche sui sistemi costieri (Underwood, 1993; Benedetti-Cecchi,
2001a). La denominazione “impatto”, tuttavia, può denotare due diverse tipologie di
effetto antropico: la prima, potenzialmente negativa, legata ad attività quali
frequentazione turistica, sviluppo urbano, inquinamento e pesca effettuata con mezzi
distruttivi; la seconda, potenzialmente positiva, conseguente l’istituzione di un’AMP. In
questo caso l’impatto, se esiste, è rappresentato dagli effetti che la protezione svolge nel
modificare le modalità di distribuzione spazio-temporale delle popolazioni e/o comunità
protette rispetto a quelle non soggette a vincoli di protezione (Benedetti-Cecchi, 2003).

36
Il presente lavoro, basato sul disegno di campionamento sopra schematizzato (fig.
2.3.2), si propone di valutare gli effetti di confusione che processi spaziali legati
all’eterogeneità ambientale possono introdurre nella valutazione dell’effetto riserva,
presso quattro isole del Parco Nazionale dell’Arcipelago Toscano (Capraia, Giannutri,
Pianosa e Montecristo).
Il disegno sperimentale prevede:
1. fattore TIPO DI ISOLA, fisso, con due livelli: PARZIALE (isole con aree
protette alternate ad aree non protette, ovvero Capraia e Giannutri) vs TOTALE
(isole totalmente protette, come Pianosa e Montecristo);
2. fattore Anno, random (i livelli non sono bilanciati, si veda Fig. 2.3.2);
3. fattore mese, random e gerarchizzato in Anno (i livelli non sono bilanciati, si
veda Fig. 2.3.2); fattore PROTEZIONE, fisso con due livelli (SI, NO);
4. fattore SITO, random (si veda la Fig. 2.3.2 per i livelli nelle diverse isole).
Le repliche sono rappresentate da cinque transetti effettuati per ciascun sito e data di
campionamento in esame.
I campionamenti sono stati effettuati sia all’interno dell’AMP, sia in aree non soggette
al regime di protezione (controlli), dove presenti, ovvero a Capraia e Giannutri. I
r1 … r5
Parziale
Capraia Giannutri
Giugno 2005 Giugno 2006
Si No
S1 … S3 S1 … S6
Si No
S1 … S3 S1 … S6
Settembre 2005
S1 … S5 S1 … S4
Si No
r1 … r5
Totale
Pianosa Montecristo
Luglio 2006 Luglio 2008
S1 … S6 S1 … S6
Maggio 2006
S1 … S6
Giugno 2009
S1 … S6
r1 … r5
Parziale
Capraia Giannutri
Giugno 2005 Giugno 2006
Si No
S1 … S3 S1 … S6
Si No
S1 … S3 S1 … S6
Settembre 2005
S1 … S5 S1 … S4
Si No
r1 … r5
Totale
Pianosa Montecristo
Luglio 2006 Luglio 2008
S1 … S6 S1 … S6
Maggio 2006
S1 … S6
Giugno 2009
S1 … S6
r1 … r5
Parziale
Capraia Giannutri
Giugno 2005 Giugno 2006
Si No
S1 … S3 S1 … S6
Si No
S1 … S3 S1 … S6
Settembre 2005
S1 … S5 S1 … S4
Si No
r1 … r5
Totale
Pianosa Montecristo
Luglio 2006 Luglio 2008
S1 … S6 S1 … S6
Maggio 2006
S1 … S6
Giugno 2009
S1 … S6
Figura 2.3.1 – Disegno di campionamento. Il numero di siti campionato per ciascun livello del fattore protezione cambia in relazione alle aree protette e non protette disponibili. Si noti, inoltre, che le isole oggetto di studio sono state campionate in anni differenti: Capraia nel giugno del 2005 e del 2006, Giannutri nel settembre del 2005, Pianosa nel luglio del 2006 e del 2008 e nel giugno del 2009, Montecristo nel maggio del 2006.

37
“controlli” sono stati scelti randomicamente da un set di possibili località non protette
caratterizzate in ogni caso da habitat comparabili a quelli campionati nell’AMP in
termini di tipologia generale, pendenza del substrato, esposizione al moto ondoso e
popolamenti presenti. L’unica differenza sostanziale è l’assenza dell’effetto della
protezione. Per questo motivo tutti i siti controllo sono situati sulla costa occidentale
delle due isole soggette a protezione parziale. In ciascun sito (che può essere
immaginato come un tratto di litorale di alcune centinaia di metri di lunghezza) i
popolamenti ittici sono stati campionati utilizzando cinque transetti.
2.4 Metodo di campionamento
Il metodo di campionamento utilizzato è basato sul visual census: una serie di tecniche
non distruttive che hanno trovato largo impiego nello studio e nel monitoraggio dei
popolamenti marini nelle Aree Marine Protette.
Altri importanti vantaggi del V.C. sono:
• la possibilità di campionamenti ripetuti nel tempo;
• l’acquisizione di dati quantitativi senza conseguenze sulle specie e sugli habitat;
• una conoscenza diretta delle caratteristiche dei siti e del comportamento e
dell’interazione sociale;
• campionamento più efficace delle specie abili nuotatrici, poiché comporta un
disturbo minimo.
Tuttavia presenta alcuni svantaggi, quali:
• limitazioni relative alla visibilità;
• limitazioni relative ai tempi e alle profondità di immersione;
• addestramento specifico degli operatori;
• interazioni “pesce-subacqueo” che portano a una sovrastima o sottostima dei dati
se, rispettivamente, l’animale è attratto o spaventato dalla presenza
dell’operatore.
In questo studio, la fauna ittica è stata censita tra gli 8 e i 10 m di profondità, in
immersione con autorespiratore lungo transetti di 25 m di lunghezza e 5 m di larghezza
(Fig. 2.3.1), secondo il metodo strip transect (Harmelin-Vivien & Harmelin, 1975), i cui
principi e limiti sono stati discussi da Harmelin-Vivien et al. (1985).

38
La tecnica del transetto ha il vantaggio, rispetto alla tecnica del punto fisso, di essere
particolarmente indicata per habitat uniformi, come quelli considerati in questo studio, e
permette di lavorare su una scala spaziale appropriata per molte delle specie ittiche
costiere.Tale metodo, inoltre, consente di ridurre il rischio di contare più volte gli
individui delle specie che vengono attratte dalla presenza dell’operatore (come accade
ad esempio con il labride Coris julis e le due specie di serranidi Serranus cabrilla e
Serranus scriba).
La tecnica consiste nel censire, nuotando lentamente in una direzione prestabilita lungo
i 25 m indicati dallo svolgimento della sagola legata all’operatore, tutti gli individui di
ciascuna specie ittica incontrati lungo il percorso. In genere, la velocità di esecuzione di
un transetto è di circa 8-10 minuti, durante i quali è consigliabile mantenere un’andatura
costante, in assetto leggermente negativo, curandosi di non sollevare detrito e di non
urtare organismi sessili del benthos. Percorrendo un “corridoio” di area (o volume) nota,
in un tempo preciso, è possibile, inoltre, rapportare i conteggi degli esemplari delle
varie specie a un valore di densità.
I dati sono stati raccolti sempre dagli stessi operatori e circa alla stessa ora (tra le 10.00
e le 14.00) onde evitare gli errori legati a fenomeni di migrazione o dispersione
giornaliera. Durante l’esecuzione del transetto, i dati in immersione vengono riportati su
una lavagnetta in materiale plastico a superficie ruvida, con una matita a mina morbida.
Gli esemplari delle specie di interesse per il presente lavoro sono stati censiti alla
migliore risoluzione tassonomica possibile (genere o specie) e sono stati contati
singolarmente per ciascun transetto; solo per individui di Chromis chromis sono state
usate classi di abbondanza, essendo tale specie fortemente gregaria.
Di ciascun individuo censito, inoltre, è stata valutata visivamente la lunghezza totale
(LT), secondo tre classi: piccoli, medi e grandi.
Fig. 2.4.1 – Transetto di 5m di larghezza e 25 m di lunghezza (Vivien
& Harmelin, 1975).

39
Alla fine di ciascun transetto un secondo operatore procedeva alla valutazione
dell’eterogeneità del substrato sottostante: una catena della lunghezza di 10 m viene
stesa sul fondale, avendo cura di seguirne il più possibile il profilo; successivamente,
con una bindella, viene misurata la proiezione orizzontale della catena. L’indice di
“rugosità” del substrato si ottiene, infine, dividendo la misura ottenuta con la bindella
per la lunghezza della catena.
L’eterogeneità della costa è stata valutata a posteriori misurando su una carta geografica
la variazione della linea di costa rispetto alla proiezione orizzontale di questa. L’indice
di eterogeneità è dato dalla varianze tra le misure di proiezione.
2.5 Analisi statistica dei dati
I dati raccolti sono stati analizzati mediante “Generalized Additive Mixed Models”
(GAMM), utilizzando la funzione gamm all’interno del pacchetto mgcv e nlme del
programma R 2.11.1 (R software project. http://www.r-project.org/).
2.5.1 Cenni teorici
2.5.1.1 Generalized Additive Models (GAM) Nei modelli statistici lineari le variabili predittive vengono tradizionalmente aggiunte
come funzioni lineari o trasformazioni parametriche linearizzate; in questi casi, viene
effettuata un’assunzione esplicita o implicita riguardo la forma parametrica della
funzione che deve essere “fittata” sui dati (quadratica, logaritmica, esponenziale,
logistica, reciproca o altro). In molti casi, tuttavia, si hanno una o più variabili
predittive, ma non si hanno a priori indizi che permettano di scegliere una particolare
forma parametrica rispetto a un’altra per descrivere la forma della relazione tra variabile
di risposta e variabili predittive. I GAM in questi casi risultano molto utili perché
permettono di cogliere la forma della relazione tra x e y senza pregiudicare il risultato
scegliendo a priori una particolare forma parametrica. Tali modelli, pertanto, estendono
il range di applicazione dei “Generalized Linear Models”, aggiungendo alle forme
parametriche, funzioni levigate non parametriche (smoothers o splines).

40
2.5.1.2 Mixed Effect Models (M&M) Questi modelli permettono di distinguere tra effetti di fattori fissi, che influenzano la
media di y, e di fattori random, che influenzano, invece, la varianza associata ad y.
Un fattore si dice “fisso” quando i livelli da includere nell’esperimento sono definiti
dall’ipotesi in esame e sono “fissati” dallo sperimentatore. Nel caso di un fattore fisso
tutti i livelli rilevanti all’analisi dell’ipotesi sono inclusi nell’esperimento. Un fattore si
dice “random” se i livelli inclusi nell’esperimento sono un sott’insieme di quelli
teoricamente possibili e la selezione avviene in modo casuale. Nel caso di un fattore
random solo un campione di possibili livelli è rappresentato nell’esperimento. Nel caso
di un fattore fisso gli effetti del trattamento sono misurati senza errore. Ciò deriva dal
fatto che per un tale fattore tutti i livelli rilevanti all’analisi dell’ipotesi sono stati inclusi
nell’esperimento. Le conclusioni dello studio sono tuttavia limitate ai soli livelli
esaminati. Nel caso di un fattore random, invece, gli effetti sono misurati con errore.
Esiste cioè una varianza associata a ciascun effetto in conseguenza del fatto che
l’esperimento include solo un campione di possibili livelli del fattore.
Alla luce di quanto esposto, i Mixed Effect Models sono da usare, per esempio, quando
il disegno di campionamento include effetti di fattori fissi, ma si ha una qualche forma
di non-indipendenza (dovuta ad esempio ad autocorrelazione tra misure ripetute nel
tempo negli stessi siti). La non-indipendenza tra osservazioni rappresenta un problema
per l’ecologo, in quanto una delle più importanti assunzioni dell’analisi statistica
standard è l’indipendenza degli errori. Riprendendo l’esempio di cui sopra, misure
ripetute nel tempo sugli stessi siti non hanno errori indipendenti poiché le peculiarità di
un dato sito si riflettono in tutte le misure fatte su di esso.
3.5.1.3 Generalized Additive Mixed Models (GAMM) I GAMM sono il risultato dell’unione delle potenzialità dei GAM e dei M&M e
risultano indicate per dati che mostrano sovradispersione e correlazione, tipici di studi
spaziali o temporali basati su disegni di campionamento gerarchizzati. Queste classi di
modelli permettono di modellizzare in maniera flessibile la dipendenza funzionale della
variabile di risposta a una o più covariate, utilizzando una regressione non parametrica
che tiene conto della correlazione tra osservazioni considerando gli effetti dei fattori
random. Anche in questi modelli, le non funzioni non parametriche vengono stimate
utilizzando delle funzioni levigate (splines).

41
2.5.1.4 Funzioni spline
A differenza dell'interpolazione polinomiale, che utilizza un unico polinomio per
approssimare la funzione su tutto l'intervallo di definizione, l'interpolazione spline è
ottenuta suddividendo l'intervallo in più sotto-intervalli (Ik=[xk,xk+1] con k=1,...,N-1)
e scegliendo per ciascuno di essi un polinomio di grado piccolo. Verrà poi imposto che
due polinomi successivi si saldino in modo liscio, cioè osservando la continuità di
qualche derivata. La funzione che si ottiene con un procedimento di questo genere si
chiama funzione spline. Pertanto, la spline è definita come una funzione, costituita da
un insieme di polinomi raccordati tra loro, il cui scopo è interpolare in un intervallo un
insieme di punti (detti nodi della spline), in modo da essere continua (almeno fino a un
dato ordine di derivate) in ogni punto dell'intervallo. La funzione interpolante ottenuta
con l’interpolazione spline, rispetto all’interpolante ottenuta con l'interpolazione lineare,
ad esempio con l’interpolazione polinomiale, presenta errori inferiori ed è più liscia, nel
senso che è la funzione interpolante con curvatura media minima.
Nel presente lavoro di tesi, sono state utilizzate delle spline cubiche nella
modellizzazione delle variabili latenti, “eterogeneità del substrato”, “eterogeneità della
costa” e “coordinate geografiche”. Questa funzione è a tratti una cubica ed è due volte
differenziabile nell'intero intervallo; inoltre, la relativa derivata seconda è zero nei punti
finali.
2.5.2 Variabili di risposta e modelli statistici
A partire dai dati di abbondanze dell’intero popolamento sono stati costruiti degli indici
cumulativi: abbondanza totale, abbondanza delle taglie piccole, medie e grandi,
abbondanza delle specie predatrici di ricci, abbondanza delle specie target della pesca e
ricchezza di specie. Nel calcolo dell’abbondanza dei predatori di ricci (Paracentrotus
lividus) sono state incluse quelle specie effettivamente in grado di controllarne la
densità, ovvero gli sparidi Diplodus sargus e D. vulgaris (che si nutrono sia degli adulti
che dei giovanili di P. lividus) e i labridi Coris julis e Thalassoma pavo, che predano
solo i giovani ricci, con diametro minore di 1 cm (Sala 1997, Guidetti 2006). Per quanto
riguarda, invece, l’abbondanza delle specie pescate, sono state considerate nel
conteggio quelle target delle tipologia di pesca permesse nelle zone 2 (tramagli, nasse,
palamiti, bolentino e lenze), cioè gli sparidi Dentex dentex, Diplodus sargus,

42
D.vulgaris, D. puntazzo, Lithognatus mormyrus, Oblada melanura, Spondiliosoma
cantharus; gli scorpenidi Scorpaena scrofa e S. porcus e i mugilidi (Mugil sp.). La
ricchezza di specie è stata calcolata come numero di specie per transetto.
Per analizzare l’effetto dei processi spaziali legati all’eterogeneità ambientale, sono
state incluse nell’analisi le coordinate geografiche relative ai siti oggetto di studio,
l’eterogeneità del fondo, misurata su ciascun transetto (scala delle decine di metri), e
l’eterogeneità della costa, relativa alla scala di variabilità delle centinaia di metri.
Successivamente si è proceduto all’esplorazione dei dati tramite dotcharts, che
permettono di visualizzare l’andamento dei dati in relazione a un fattore incluso
nell’analisi e la presenza di eventuali valori estremi e pair functions, che mostrano da un
lato l’andamento della variabile di risposta in funzione delle variabili predittive e
dall’altro la presenza di eventuali collinearità tra queste ultime. Tali rappresentazioni
grafiche consentono, quindi, di valutare la possibilità di trasformare i dati e l’impiego o
meno delle variabili predittive inizialmente prese in considerazione.
La procedura di analisi è continuata con la costruzione del generalized additive mixed
model, nel quale: sono stati inclusi gli effetti dei fattori fissi “tipo di isola” (TI) e
“protezione”(P); le covariate relative ai processi spaziali (“eterogeneità del substrato”
ES, “eterogeneità della costa” EC e “coordinate spaziali” LAT, LONG) sono state
modellizzate come funzioni spline; è bene precisare che i valori relativi alle variabili
“eterogeneità del substrato ed “eterogeneità della costa” sono stati trasformati con
secondo log(x + 0.1); il tempo (“anno” e “mese”) è stato invece incluso nella parte
random del modello.
η(μijk) = β0 + β1TIi + β2Pi + ƒ1(ESijk) + ƒ2(ECi) + ƒ3(LATi, LONGi) + αji + ϕkji (2.1)
In cui:
− η(μijk) rappresenta il valore medio atteso del predittore lineare per l’i-esimo sito,
il j-esimo anno e il k-esimo mese; η rappresenta la funzione di identità nel caso
di modelli gaussiani, la funzione logaritmica nel caso di modelli poissoniani o la
funzione esponenziale nel caso di modelli quasi-poisson,

43
− β0 indica l’intercetta del modello GAMM relativa all’andamento medio della
variabile di risposta nei siti considerati quando i valori delle altre variabili
predittive sono uguali a zero,
− β1TIi rappresenta il coefficiente relativo al fattore “tipo di isola” nell’i-esimo
sito;
− β2Pi rappresenta il coefficiente associato al fattore “protezione” nell’i-esimo
sito;
− ƒ1(ESijk) + ƒ2(ECi) + ƒ3(LATi, LONGi) indicano gli effetti delle covariate relative
a fattori ambientali e processi spaziali nell’i-esimo sito, modellizzate come
funzioni spline cubiche, non parametriche,
− αji rappresenta l’effetto casuale del j-esimo anno per l’i-esimo sito,
− ϕkji rappresenta l’effetto casuale associato al k-esimo mese nel j-esimo anno nel
per l’i-esimo sito.
Nel modello è possibile, infine, specificare la famiglia a cui far appartenere la
distribuzione dei dati: nel presente lavoro è stata preferita la distribuzione di Poisson (in
cui media e varianza risultano uguali), più adatta ai conteggi di individui rispetto a
quella gaussiana. Tuttavia, a causa dell’elevata sovradispersione dei dati (valutabile
attraverso il parametro scale inference), è stato necessario aggiustare tale distribuzione
rilasciando il vincolo per cui media e varianza siano uguali e correggendo gli errori
standard tramite un modello quasi-poisson, in cui la varianza deriva dal prodotto della
media (μ) per la dispersione (φ) del parametro. Il risultato è che rapporto tra media e
varianza è sempre più grande di 1 ma non è più un problema dal momento che si tiene
conto della sovradispersione dei dati.
A ciascuna variabile di risposta è stato applicato sia il modello completo sopra
esplicato, sia uno incompleto, in cui è stata eliminata la parte relativa alle coordinate
geografiche, al fine di testare l’importanza dei processi spaziali nella valutazione
del’effetto riserva.
La bontà dei modelli è stata valutata tramite “Akaike’s Information Criterion”,
conosciuto anche come penalized loglikelihood. In presenza di un modello per il quale è
possibile calcolare un valore di loglikelihood (Burnham and Anderson 2002), AIC = - 2
x loglikelihood +2(p +1), dove p è il numero di parametri del modello. In generale,

44
maggiore è il numero di parametri inclusi nel modello, migliore sarà il fit. Il fit perfetto
potrebbe essere raggiunto, infatti, aggiungendo un parametro diverso per ciascun dato,
tuttavia, un modello di questo tipo non avrebbe alcun potere esplicativo. Ci deve essere
quindi un giusto equilibrio tra bontà del modello e numero di parametri inclusi secondo
un principio di parsimonia. AIC è utile perché penalizza esplicitamente ogni parametro
superfluo presente nel modello aggiungendo il termine 2(p +1) alla devianza. Nel
comparare due modelli, pertanto, più piccolo è il valore di AIC, migliore è il modello.
Il modello selezionato è stato successivamente validato attraverso l’analisi dei residui.
Al fine di testare il modello secondo il quale l’alternarsi di zone soggette a protezione e
zone impattate dalla pesca su isole parzialmente protette potesse generare modalità di
distribuzione dei popolamenti ittici differenti da quelli che si originano su isole
totalmente protette, alle variabili di risposta sono stati applicati di volta in volta due
Generalized Additive Mixed Models con il fattore “Sito” gerarchizzato nel fattore “Tipo
di isola”, uno per le isole parzialmente protette e uno per quelle totalmente protette.
L’esistenza di differenze statisticamente significative tra i due modelli è stata testata
valutando la sovrapposizione degli intervalli di confidenza intorno alle varianze tra isole
e siti gerarchizzati in isola, calcolati tramite la funzione intervals del pacchetto nlme.
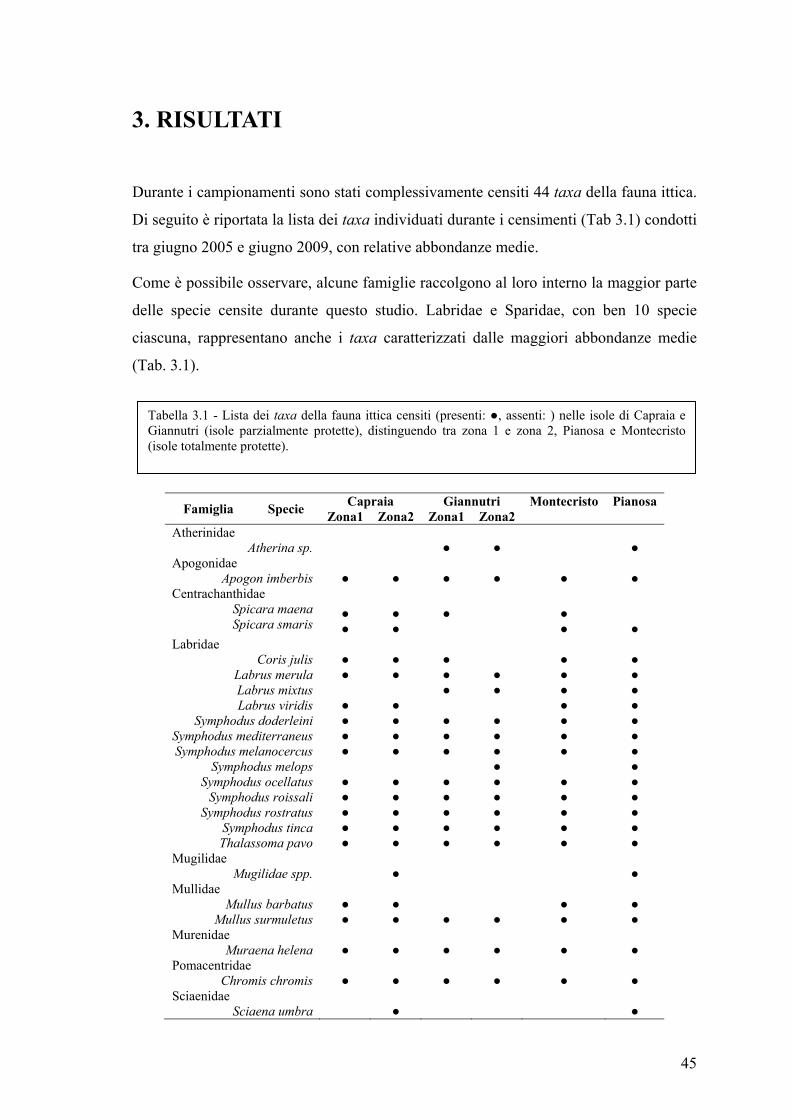
45
3. RISULTATI
Durante i campionamenti sono stati complessivamente censiti 44 taxa della fauna ittica.
Di seguito è riportata la lista dei taxa individuati durante i censimenti (Tab 3.1) condotti
tra giugno 2005 e giugno 2009, con relative abbondanze medie.
Come è possibile osservare, alcune famiglie raccolgono al loro interno la maggior parte
delle specie censite durante questo studio. Labridae e Sparidae, con ben 10 specie
ciascuna, rappresentano anche i taxa caratterizzati dalle maggiori abbondanze medie
(Tab. 3.1).
Famiglia Specie Capraia Giannutri Montecristo Pianosa Zona1 Zona2 Zona1 Zona2
Atherinidae Atherina sp. ● ● ●
Apogonidae Apogon imberbis ● ● ● ●
● ●
Centrachanthidae Spicara maena Spicara smaris
● ●
● ●
●
● ● ●
Labridae Coris julis
Labrus merula Labrus mixtus Labrus viridis
Symphodus doderleini Symphodus mediterraneus Symphodus melanocercus
Symphodus melops Symphodus ocellatus
Symphodus roissali Symphodus rostratus
Symphodus tinca Thalassoma pavo
● ● ● ● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ● ● ● ● ● ●
Mugilidae Mugilidae spp. ● ●
Mullidae Mullus barbatus
Mullus surmuletus ● ●
● ●
● ●
● ●
● ●
Murenidae Muraena helena ● ● ● ● ● ●
Pomacentridae Chromis chromis ● ● ● ● ● ●
Sciaenidae Sciaena umbra ● ●
Tabella 3.1 - Lista dei taxa della fauna ittica censiti (presenti: ●, assenti: ) nelle isole di Capraia e Giannutri (isole parzialmente protette), distinguendo tra zona 1 e zona 2, Pianosa e Montecristo (isole totalmente protette).

46
Scorpaenidae Scorpaena porcus Scorpaena scrofa
●
●
●
●
Serranidae Anthias anthias
Epinephelus marginatus Serranus cabrilla Serranus hepatus
Serranus scriba
● ● ● ●
● ● ●
● ● ● ●
● ● ● ●
● ● ● ● ●
● ● ● ●
Sparidae Boops boops
Dentex dentex Diplodus annularis Diplodus puntazzo
Diplodus sargus Diplodus vulgaris
Lythognatus mormyrus Oblada melanura
Pagellus acarne Sarpa salpa
Spondyliosoma cantharus
● ● ● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ●
● ● ● ● ● ● ●
● ● ● ● ● ● ● ● ●
● ● ● ● ● ● ● ● ● ●
Sphyraenidae Sphyraena viridensis ● ● ●
Trachinidae Trachinus draco ●
Nella maggior parte dei casi le medie delle abbondanze di ciascuna specie non sono
molto elevate. L’analisi, pertanto, non è stata condotta sull’intero popolamento, ma su
alcuni indici cumulativi: abbondanza totale, abbondanza di taglie piccole, medie e
grandi, abbondanza di specie predatrici del riccio Paracentrotus lividus (Tab 3.2),
abbondanza di specie pescate (Tab. 3.3) e ricchezza di specie.
Per quanto riguarda i predatori di ricci, sono state considerate solo le specie ittiche in
grado di controllare la densità delle popolazioni di Paracentrotus lividus di fondi
rocciosi costieri.
Famiglia Specie
Sparidae Diplodus sargus Si nutrono sia degli adulti che dei giovanili di
ricci (Paracentrotus lividus) Diplodus vulgaris
Labridae Coris julis Predano solo giovani ricci con diametro
minore di 1cm Thalassoma pavo
Tabella 3.2 – Lista delle specie predatrici di ricci.

47
Per quanto concerne i pesci pescati, sono state considerate le specie target delle pesche
permesse all’interno delle zone 2 delle isole parzialmente protette:
− tramagli (reti da posta fisse che lavorano ancorate sul fondo): le catture
riguardano specie bentoniche e nectobentoniche come scorpenidi e sparidi;
− nasse (trappole mobili): catturano principalmente organismi bentonici come
scorpenidi e gronghi;
− palamiti, lenze e bolentino sono da considerare attrezzi con ami che permettono
la cattura di sparidi, principalmente, ma anche grandi predatori pelagici (come
tonni e pesci spada) e pesce azzurro.
Famiglia Specie
Sparidae Dentex dentex
Diplodus puntazzo
Diplodus sargus
Diplodus vulgaris
Lithognatus mormyrus
Oblada melanura
Spondiliosoma
cantharus
Scorpaenidae Scorpaena porcus
Scorpaena scrofa
Mugilidae Mugil sp.
L’analisi è stata condotta procedendo secondo il seguente ordine: esplorazione dei dati;
fit e validazione del modello; inferenza e, infine, rappresentazione grafica dei risultati.
Tabella 3.3 – Lista delle specie pescate con le tecnica di pesca permesse all’interno delle zone 2 (tramagli, nasse, palamiti, lenze e bolentino).

48
3.1 Esplorazione dei dati
Uno strumento utile per visualizzare l’andamento dei dati è rappresentato dalle
cosiddette dotcharts, che permettono di “plottare” i valori delle variabili di riposta in
funzione di uno dei fattori inclusi nell’analisi. Questi grafici sono molto utili in quanto
consentono di evidenziare la presenza di strutturazione dei dati, così come di outliers.
Nel nostro caso, i dati relativi alle abbondanze (totale, delle taglie piccole, medie e
grandi, dei predatori di ricci e delle specie pescate) e alla ricchezza di specie sono stati
rappresentati separatamente per le quattro isole sito di studio.
Figura 3.1.1 – Dotchart relativa all’abbondanza totale, separatamente per ciascuna isola oggetto di studio.
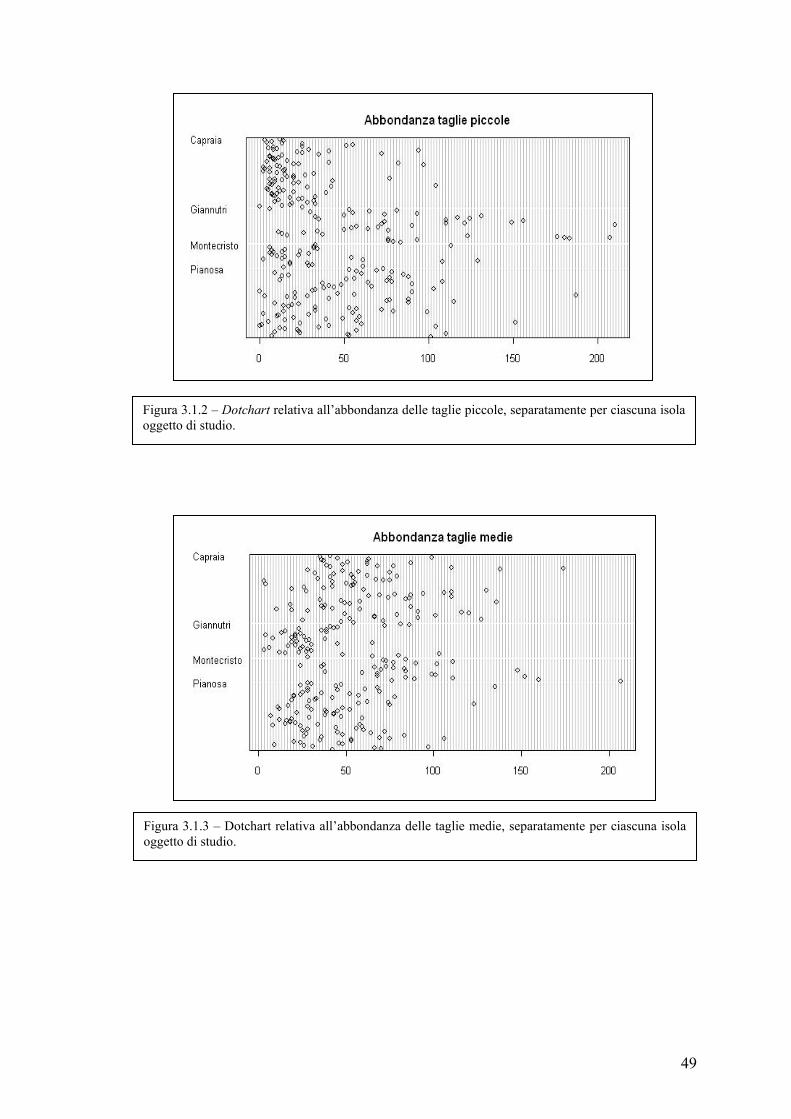
49
Figura 3.1.2 – Dotchart relativa all’abbondanza delle taglie piccole, separatamente per ciascuna isola oggetto di studio.
Figura 3.1.3 – Dotchart relativa all’abbondanza delle taglie medie, separatamente per ciascuna isola oggetto di studio.
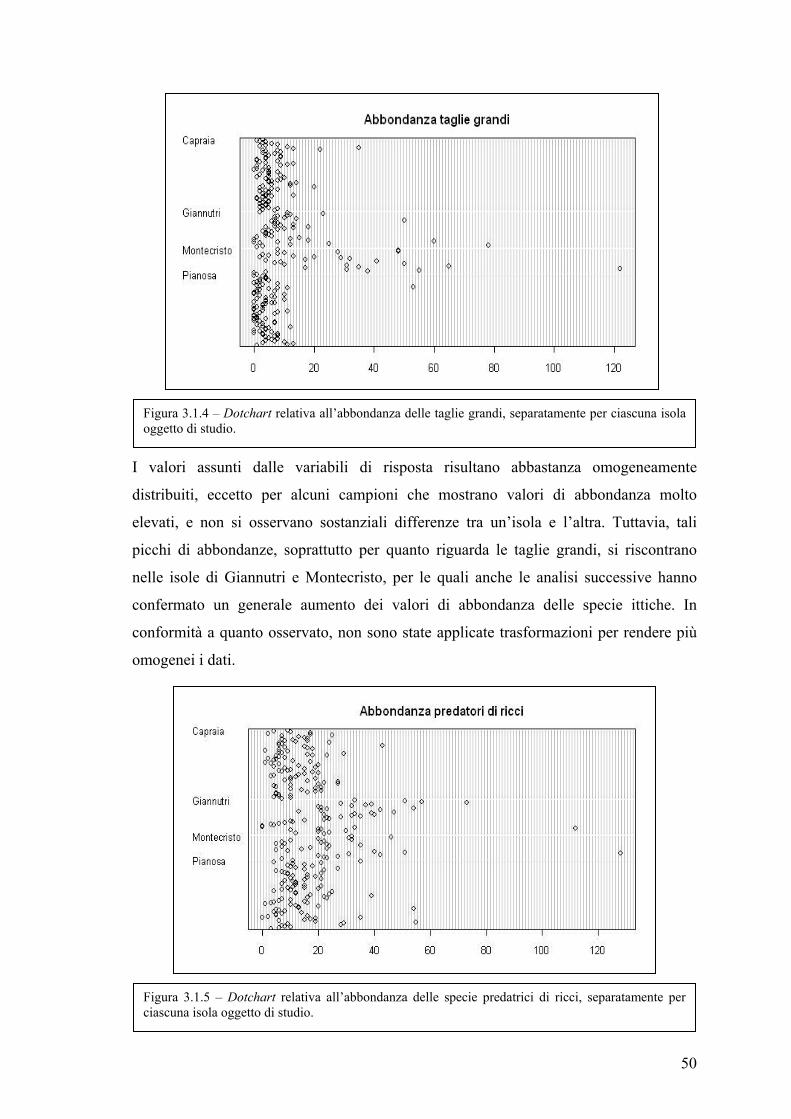
50
I valori assunti dalle variabili di risposta risultano abbastanza omogeneamente
distribuiti, eccetto per alcuni campioni che mostrano valori di abbondanza molto
elevati, e non si osservano sostanziali differenze tra un’isola e l’altra. Tuttavia, tali
picchi di abbondanze, soprattutto per quanto riguarda le taglie grandi, si riscontrano
nelle isole di Giannutri e Montecristo, per le quali anche le analisi successive hanno
confermato un generale aumento dei valori di abbondanza delle specie ittiche. In
conformità a quanto osservato, non sono state applicate trasformazioni per rendere più
omogenei i dati.
Figura 3.1.5 – Dotchart relativa all’abbondanza delle specie predatrici di ricci, separatamente per ciascuna isola oggetto di studio.
Figura 3.1.4 – Dotchart relativa all’abbondanza delle taglie grandi, separatamente per ciascuna isola oggetto di studio.
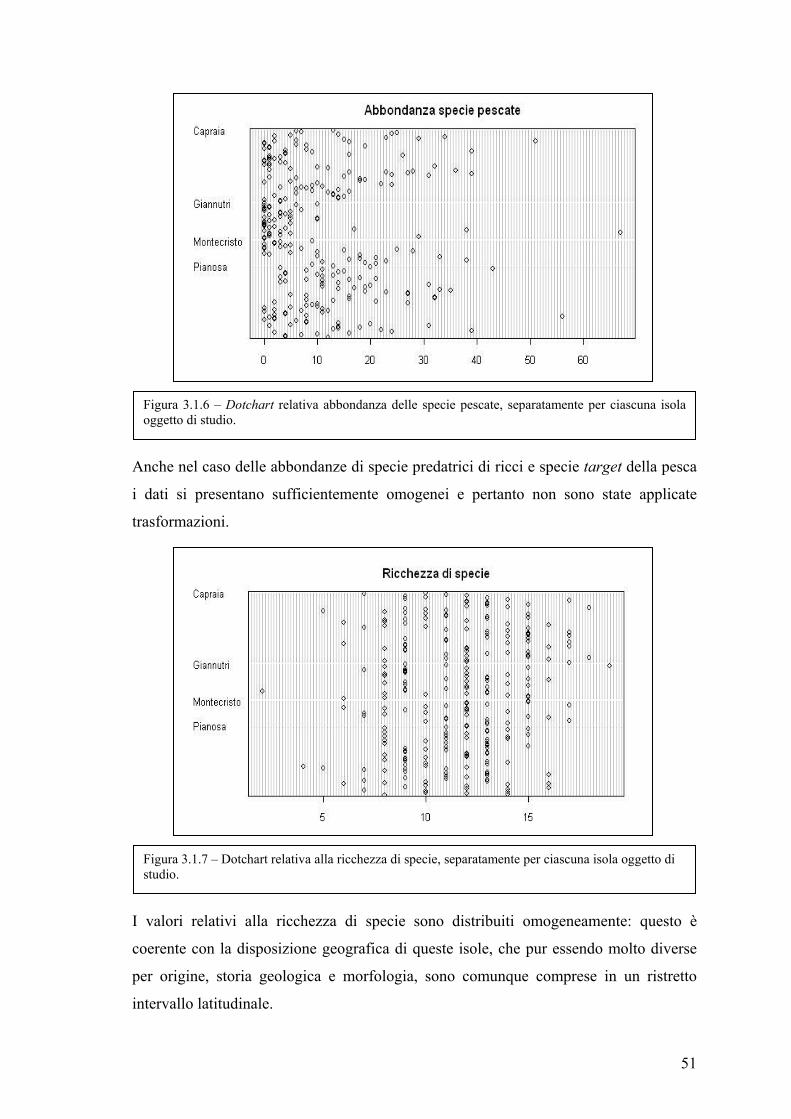
51
Anche nel caso delle abbondanze di specie predatrici di ricci e specie target della pesca
i dati si presentano sufficientemente omogenei e pertanto non sono state applicate
trasformazioni.
I valori relativi alla ricchezza di specie sono distribuiti omogeneamente: questo è
coerente con la disposizione geografica di queste isole, che pur essendo molto diverse
per origine, storia geologica e morfologia, sono comunque comprese in un ristretto
intervallo latitudinale.
Figura 3.1.7 – Dotchart relativa alla ricchezza di specie, separatamente per ciascuna isola oggetto di studio.
Figura 3.1.6 – Dotchart relativa abbondanza delle specie pescate, separatamente per ciascuna isola oggetto di studio.
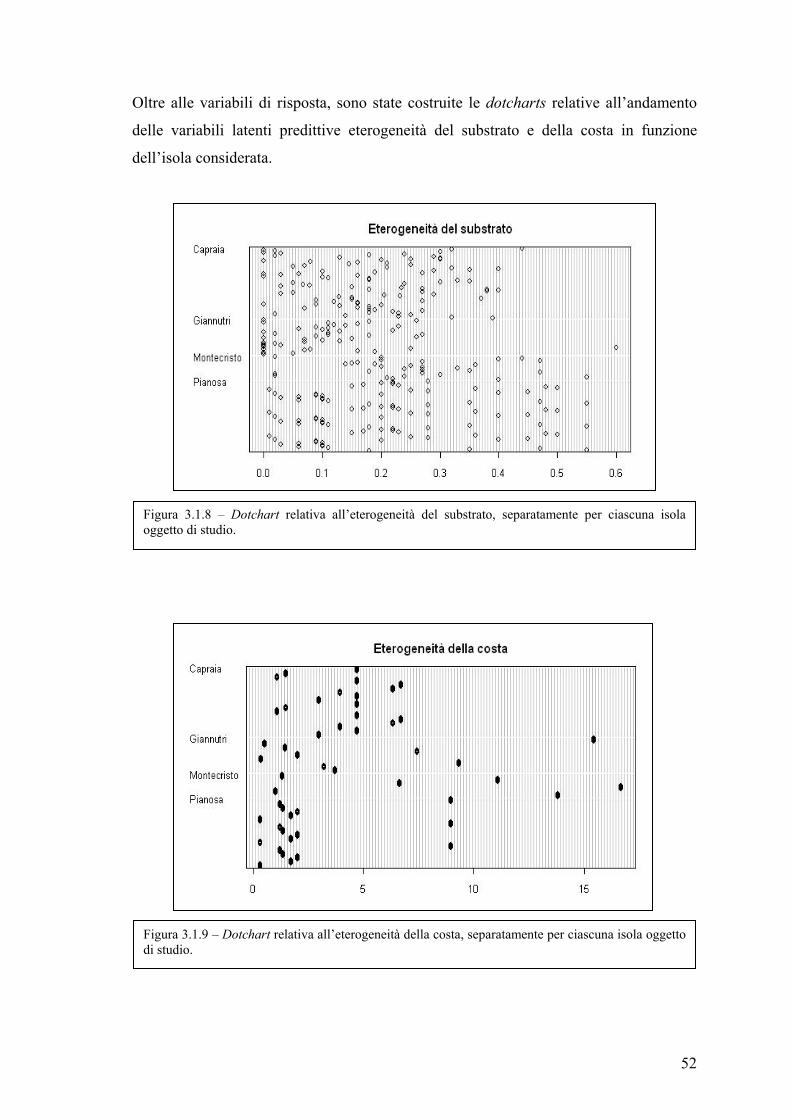
52
Oltre alle variabili di risposta, sono state costruite le dotcharts relative all’andamento
delle variabili latenti predittive eterogeneità del substrato e della costa in funzione
dell’isola considerata.
Figura 3.1.9 – Dotchart relativa all’eterogeneità della costa, separatamente per ciascuna isola oggetto di studio.
Figura 3.1.8 – Dotchart relativa all’eterogeneità del substrato, separatamente per ciascuna isola oggetto di studio.
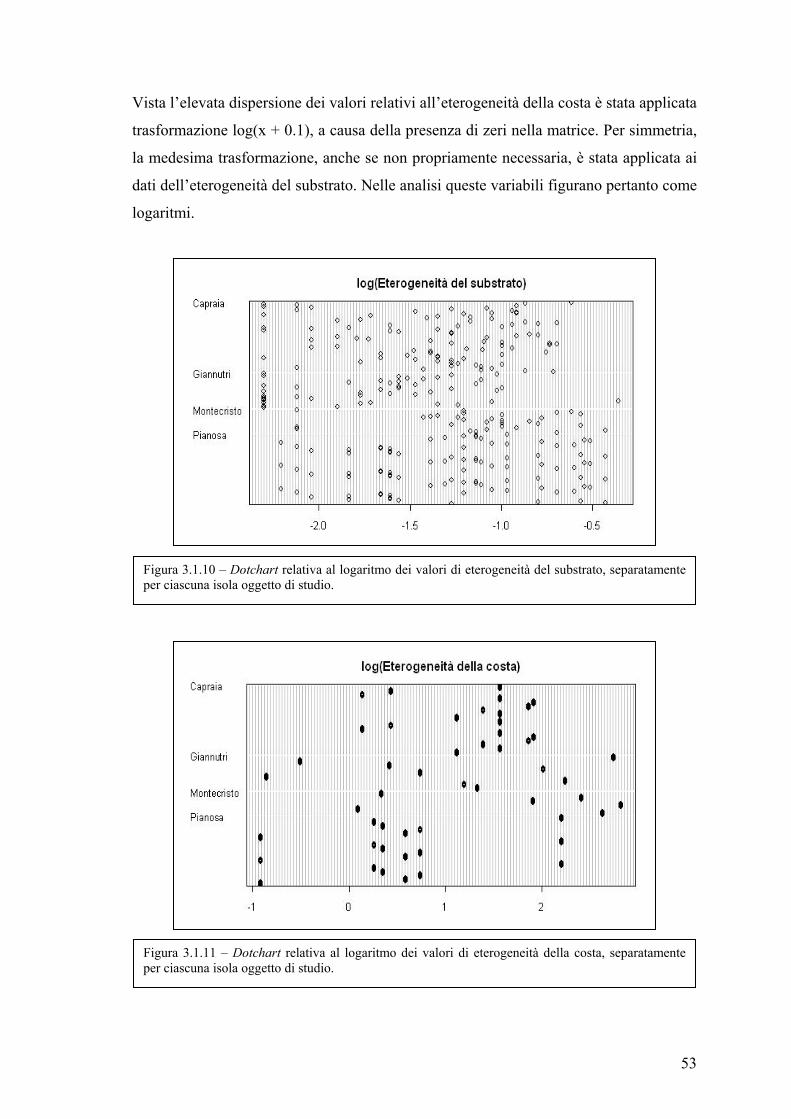
53
Vista l’elevata dispersione dei valori relativi all’eterogeneità della costa è stata applicata
trasformazione log(x + 0.1), a causa della presenza di zeri nella matrice. Per simmetria,
la medesima trasformazione, anche se non propriamente necessaria, è stata applicata ai
dati dell’eterogeneità del substrato. Nelle analisi queste variabili figurano pertanto come
logaritmi.
Figura 3.1.11 – Dotchart relativa al logaritmo dei valori di eterogeneità della costa, separatamente per ciascuna isola oggetto di studio.
Figura 3.1.10 – Dotchart relativa al logaritmo dei valori di eterogeneità del substrato, separatamente per ciascuna isola oggetto di studio.

54
Un altro valido strumento per l’esplorazione dei dati è rappresentato dalle cosiddette
pairs function, che permettono di visualizzare in un’unica schermata l’andamento della
variabile di risposta in funzione delle variabili predittive da includere nell’analisi e la
presenza di eventuali collinearità tra queste ultime (Fig. 3.1.12). Il risultato grafico è
costituito da una matrice rettangolare di grafici, divisa in due triangoli da una diagonale:
la prima colonna di grafici del triangolo inferiore restituisce l’andamento della variabile
di risposta (nell’esempio l’abbondanza totale) in funzione delle variabili predittive,
mentre nel triangolo superiore vengono visualizzati i valori di correlazione calcolati per
ciascuna coppia di variabili predittive. Sulla diagonale sono presenti gli istogrammi con
le distribuzioni dei dati relativi a ciascuna variabile.
Verificare la presenza di eventuali collinearità tra le variabili predittive è molto utile al
fine di decidere quali di queste è necessario includere nel modello che si vuole testare:
se due variabili sono correlate, infatti, sarebbe ridondante inserirle entrambe.
È stato utilizzato anche un altro indicatore di multicollinearità: il variance inflation
factor (fattore di inflazione della varianza), o VIF, calcolato per ciascuna variabile
predittiva del modello (se il coefficiente supera un determinato valore soglia, di solito 3
o 5, la variabile va esclusa dal modello). Nel caso del presente lavoro di tesi, nessun
coefficiente supera il valore soglia.
Protezione Tipo di
isola Log(Eterogeneità del
substrato) Log(eterogeneità della
costa VIF 1.735 1.855 1.071 1.072
Pairs function sono state costruite per ciascuna variabile di risposta inclusa nello studio,
al fine di visualizzare l’andamento di queste ultime in funzione delle variabili predittive.
Tuttavia, dal momento che il plot è, per la maggior parte, uguale per tutte le variabili di
risposta, di seguito sono riportate solo le prime colonne di grafici relative all’andamento
delle variabili di risposta in funzione delle variabili predittive (Fig. 3.1.13).
Tabella 3.1.1 – VIF (fattore di inflazione della varianza) calcolato per ciascuna variabile predittiva.

55
Figura 3.1.12 – Pairs function della variabile di risposta abbondanza totale.

56
Figura 3.1.13 – Andamento dei valori delle variabili di risposta in funzione delle variabili predittive incluse nello studio.
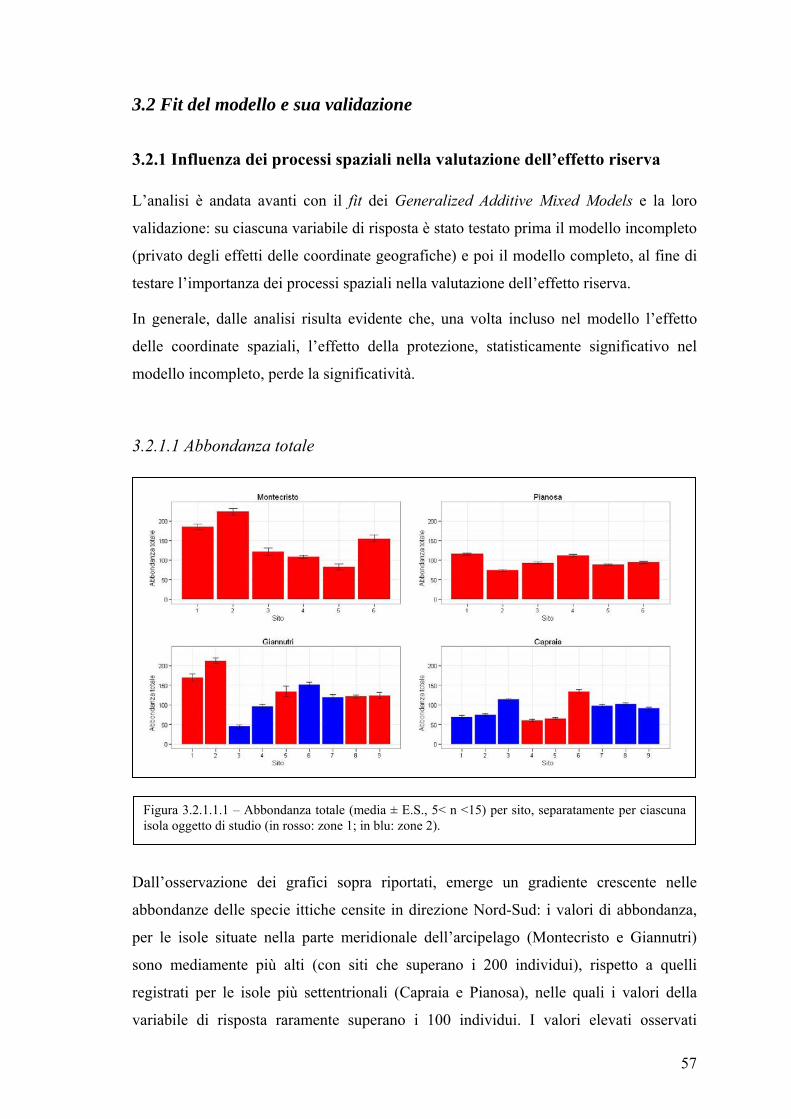
57
3.2 Fit del modello e sua validazione
3.2.1 Influenza dei processi spaziali nella valutazione dell’effetto riserva L’analisi è andata avanti con il fit dei Generalized Additive Mixed Models e la loro
validazione: su ciascuna variabile di risposta è stato testato prima il modello incompleto
(privato degli effetti delle coordinate geografiche) e poi il modello completo, al fine di
testare l’importanza dei processi spaziali nella valutazione dell’effetto riserva.
In generale, dalle analisi risulta evidente che, una volta incluso nel modello l’effetto
delle coordinate spaziali, l’effetto della protezione, statisticamente significativo nel
modello incompleto, perde la significatività.
3.2.1.1 Abbondanza totale
Dall’osservazione dei grafici sopra riportati, emerge un gradiente crescente nelle
abbondanze delle specie ittiche censite in direzione Nord-Sud: i valori di abbondanza,
per le isole situate nella parte meridionale dell’arcipelago (Montecristo e Giannutri)
sono mediamente più alti (con siti che superano i 200 individui), rispetto a quelli
registrati per le isole più settentrionali (Capraia e Pianosa), nelle quali i valori della
variabile di risposta raramente superano i 100 individui. I valori elevati osservati
Figura 3.2.1.1.1 – Abbondanza totale (media ± E.S., 5< n <15) per sito, separatamente per ciascuna isola oggetto di studio (in rosso: zone 1; in blu: zone 2).
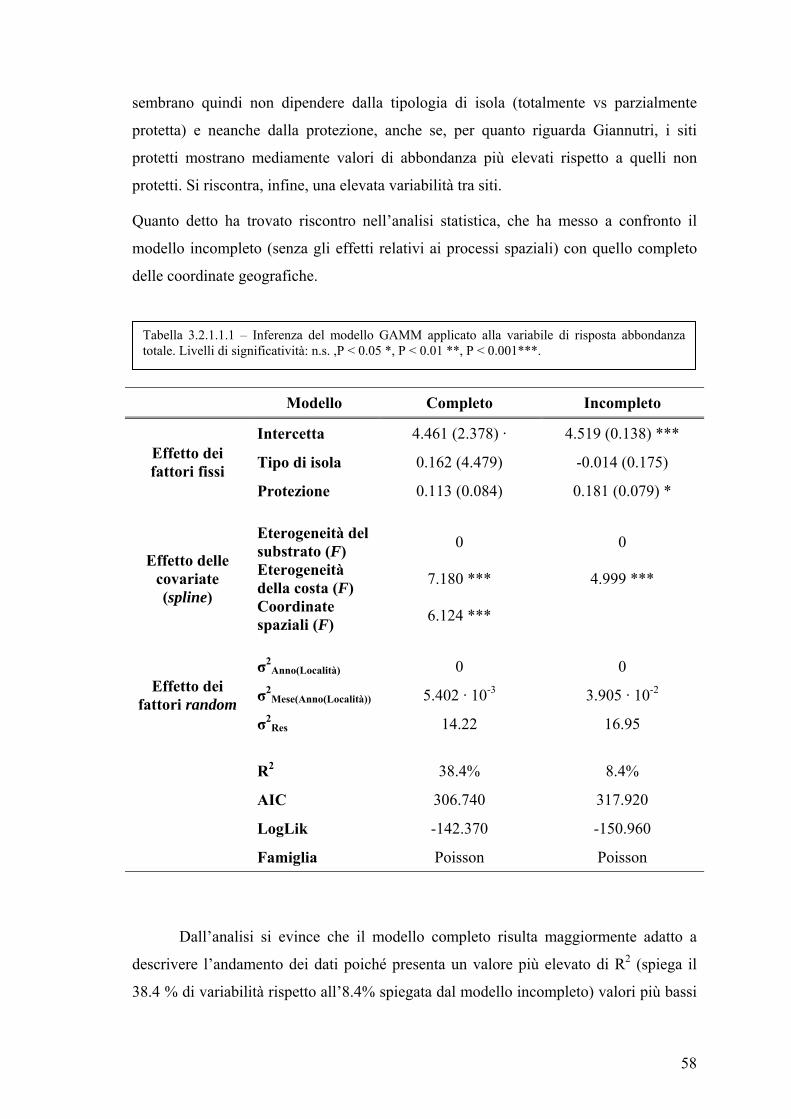
58
sembrano quindi non dipendere dalla tipologia di isola (totalmente vs parzialmente
protetta) e neanche dalla protezione, anche se, per quanto riguarda Giannutri, i siti
protetti mostrano mediamente valori di abbondanza più elevati rispetto a quelli non
protetti. Si riscontra, infine, una elevata variabilità tra siti.
Quanto detto ha trovato riscontro nell’analisi statistica, che ha messo a confronto il
modello incompleto (senza gli effetti relativi ai processi spaziali) con quello completo
delle coordinate geografiche.
Modello Completo Incompleto
Effetto dei fattori fissi
Intercetta 4.461 (2.378) · 4.519 (0.138) ***
Tipo di isola 0.162 (4.479) -0.014 (0.175)
Protezione 0.113 (0.084) 0.181 (0.079) *
Effetto delle covariate (spline)
Eterogeneità del substrato (F) 0 0
Eterogeneità della costa (F) 7.180 *** 4.999 ***
Coordinate spaziali (F) 6.124 ***
Effetto dei fattori random
σ2Anno(Località) 0 0
σ2Mese(Anno(Località)) 5.402 · 10-3 3.905 · 10-2
σ2Res 14.22 16.95
R2 38.4% 8.4%
AIC 306.740 317.920
LogLik -142.370 -150.960
Famiglia Poisson Poisson
Dall’analisi si evince che il modello completo risulta maggiormente adatto a
descrivere l’andamento dei dati poiché presenta un valore più elevato di R2 (spiega il
38.4 % di variabilità rispetto all’8.4% spiegata dal modello incompleto) valori più bassi
Tabella 3.2.1.1.1 – Inferenza del modello GAMM applicato alla variabile di risposta abbondanza totale. Livelli di significatività: n.s. ,P < 0.05 *, P < 0.01 **, P < 0.001***.

59
di LogLik e AIC (tab. 3.2.1.1.1). Anche i residui sono ragionevolmente conformi alle
assunzioni (vedi Fig. A.1, appendice A).
È bene notare che l’inclusione nel modello dell’effetto delle coordinate spaziali
(altamente significativo, P<0.001) determina la perdita di significatività dell’intercetta,
ma soprattutto dell’effetto della protezione.
Statisticamente significativo (P<0.001) è anche l’effetto della covariata “Eterogeneità
della costa”: in particolare, per valori intermedi di eterogeneità della costa, si registrano
le più alte abbondanze dei taxa del popolamento ittico (Fig. 3.2.1.1.2).
Figura 3.2.1.1.2 – Andamento della variabile di risposta abbondanza totale in funzione della spline relativa all’eterogeneità della costa; è riportato l’intervallo di confidenza.

60
3.2.1.2 Abbondanza delle taglie piccole
Il valori di abbondanza delle taglie piccole risultano generalmente bassi (mediamente al
di sotto dei 50 individui per sito) in tre delle quattro isole oggetto di studio; valori
decisamente più elevati sono stati registrati a Giannutri, con picchi superiori ai 150
individui. Pertanto, nonostante i bassi valori di Montecristo, l’esistenza di un gradiente
latitudinale nei valori di abbondanza, sembra essere confermato anche da questa
variabile di risposta. Anche in questo caso, il tipo di isola e la protezione non sembrano
influenzare le modalità di distribuzione delle taglie piccole del popolamento ittico.
Quanto osservato è stato confermato dall’inferenza: l’effetto delle coordinate spaziali e
dell’eterogeneità della costa sono altamente significativi, rispettivamente P<0.001 e
P<0.01, mentre si riscontra solo un debole effetto della protezione (Tab 3.2.1.2.1).
Anche in questo caso il modello completo è preferibile a quello privo degli effetti delle
coordinate spaziali, in base ai valori di AIC, LogLik ed R2 (Tab 3.2.1.2.1). L’analisi dei
residui è conforme alle assunzioni del modello (vedi Fig. A.2, appendice A)
Figura 3.2.1.2.1 - Abbondanza delle taglie piccole (media ± E.S., 5< n <15) per sito, separatamente per ciascuna isola oggetto di studio (in rosso: zone 1; in blu: zone 2).

61
Modello Completo Incompleto
Effetto dei fattori fissi
Intercetta 3.753 (0.233) *** 3.516 (0.268) ***
Tipo di isola -0.519 (0.351) -0.093 (0.356)
Protezione 0.238 (0.138) · 0.216 (0.138) ·
Effetto delle covariate (spline)
Log(Eterogeneità del fondo) (F) 0 0
Log(Eterogeneità della costa) (F) 6.873 ** 3.363 *
Coordinate spaziali (F) 43.180 ***
Effetto dei fattori random
σ2Anno(Località) 0 0
σ2Mese(Anno(Località)) 0 1.841 · 10-1
σ2Res 24.38 24.91
R2 34% 2.3%
AIC 638.629 650.887
LogLik -308.314 -317.443
Famiglia Poisson Poisson
Anche in questo caso, andando a valutare l’andamento della variabile di risposta in
funzione della covariata eterogeneità della costa, si osserva che i valori di elevati di
abbondanza delle taglie piccole si hanno per valori intermedi di eterogeneità (Fig.
3.2.1.2.2).
Tabella 3.2.1.2.1 - Inferenza del modello GAMM applicato alla variabile di risposta abbondanza taglie piccole. Livelli di significatività: n.s. ,P < 0.05 *, P < 0.01 **, P < 0.001***.
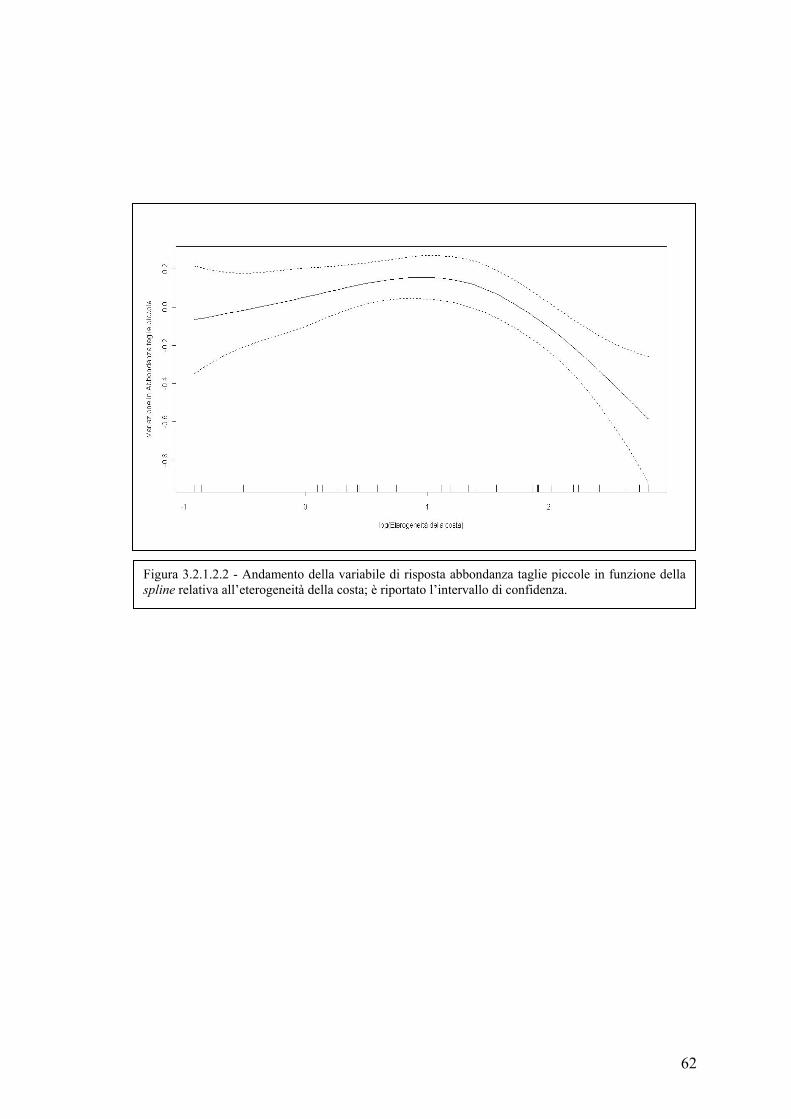
62
Figura 3.2.1.2.2 - Andamento della variabile di risposta abbondanza taglie piccole in funzione della spline relativa all’eterogeneità della costa; è riportato l’intervallo di confidenza.

63
3.2.1.3 Abbondanza delle taglie medie
Per quanto riguarda le taglie medie delle specie ittiche censite (Fig. 3.2.1.3.1), si
osservano valori di abbondanza mediamente più elevati nell’isola totalmente protetta di
Montecristo, con un picco di oltre 120 individui nel sito 1, rispetto alle altre isole
oggetto di studio; in particolare Capraia è caratterizzata dai più bassi valori di
abbondanza (attorno ai 45 individui per sito). Si riscontra, in generale, una marcata
variabilità tra siti, in particolare a Giannutri, indipendentemente dalle misure di
protezione applicate.
La tabella relativa all’inferenza del modello è stata omessa poiché i coefficienti ottenuti
non sono confrontabili con quelli delle altre variabili di risposta prese in esame. A causa
di problemi di convergenza nei dati, non è stato possibile applicare la distribuzione di
Poisson, la più indicata trattandosi di conteggi, ma è stata utilizzata la distribuzione
normale (gaussiana). Il modello ottenuto, presentando valori elevati di AIC, R2 molto
basso e residui non normali, non consente una corretta interpretazione dei risultati.
Figura 3.2.1.3.1 - Abbondanza delle taglie medie (media ± ± E.S., 5< n <15) per sito, separatamente per ciascuna isola oggetto di studio (in rosso: zone 1; in blu: zone 2).

64
3.2.1.4 Abbondanza delle taglie grandi Anche nel caso delle taglie grandi del popolamento ittico (Fig. 3.2.1.4.1), i valori più
elevati di abbondanze (mediamente 25 individui per sito, con un picco di 55 nel sito 2)
si registrano a Montecristo e a Giannutri (mediamente 10 individui per sito). Quanto
osservato sembra confermare l’ipotesi di un gradiente di abbondanze crescenti in
direzione Nord-Sud.
Il modello completo è maggiormente adatto a descrivere l’andamento dei dati poiché
presenta un valore più elevato di R2 (38.5 %) e valori più bassi di LogLik e AIC (Tab.
3.2.1.4.1). Anche i residui sono ragionevolmente conformi alle assunzioni (vedi Fig.
A.4, appendice A).
L’importanza della posizione delle isole nell’influenzare la distribuzione degli
organismi è stata confermata anche nel caso delle taglie grandi degli individui del
popolamento ittico: il coefficiente relativo agli effetti delle coordinate geografiche è
risultato altamente significativo (P<0.001). Lo stesso si può dire dell’indice di
eterogeneità della costa, statisticamente significativo anche per le taglie grandi
(P<0.01).
Figura 3.2.1.4.1 - Abbondanza delle taglie grandi (media ± E.S., 5< n <15) per sito, separatamente per ciascuna isola oggetto di studio (in rosso: zone 1; in blu: zone 2).

65
Modello Completo Incompleto
Effetto dei fattori fissi
Intercetta 5.153 (3.556) 1.707 (0.422) ***
Tipo di isola -6.140 (9.364) 0.231 (0.569)
Protezione 0.053 (0.201) 0.216 (0.199)
Effetto delle covariate (spline)
Eterogeneità del fondo (F) 0 0
Eterogeneità della costa (F) 10.09 ** 5.703 **
Coordinate spaziali (F) 14.00 ***
Effetto dei fattori random
σ2Anno(Località) 0 0
σ2Mese(Anno(Località)) 0 4.826 · 10-1
σ2Res 7.474 9.449
R2 38.5% 2%
AIC 805.913 830.936
LogLik -391.956 -407.468 Famiglia Poisson Poisson
Il grafico che mostra l’andamento delle abbondanze in funzione del logaritmo
dell’eterogeneità della costa (Fig. 3.2.1.4.2) conferma quanto osservato per
l’abbondanza totale e delle taglie piccole: valori massimi di abbondanza delle taglie
grandi si registrano per valori intermedi di eterogeneità della costa.
Tabella 3.2.1.4.1 - Inferenza del modello GAMM applicato alla variabile di risposta abbondanza taglie grandi. Livelli di significatività: n.s. ,P < 0.05 *, P < 0.01 **, P < 0.001***.
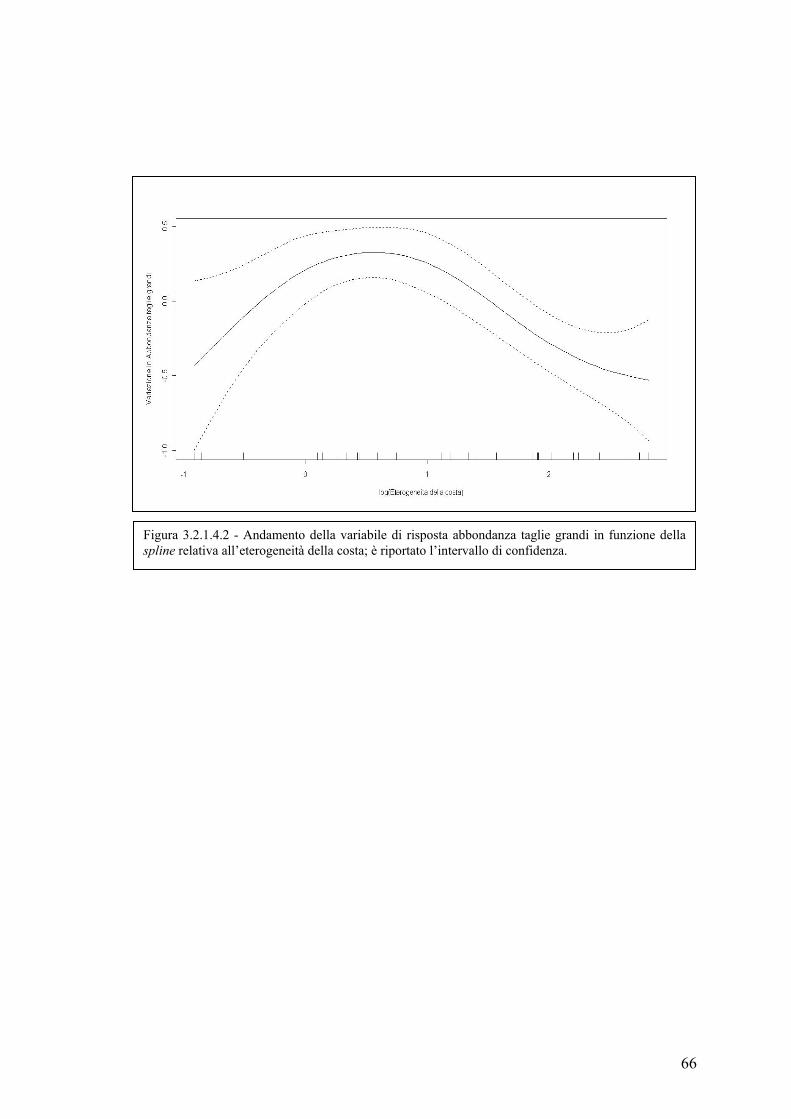
66
Figura 3.2.1.4.2 - Andamento della variabile di risposta abbondanza taglie grandi in funzione della spline relativa all’eterogeneità della costa; è riportato l’intervallo di confidenza.

67
3.2.1.5 Abbondanza delle specie predatrici di ricci
Anche nel caso dei predatori di ricci, i valori di abbondanza maggiori si riscontrano per
le isole situate a sud dell’arcipelago, Montecristo e Giannutri, mentre Pianosa e Capraia
si mantengono al di sotto dei individui per sito (Fig. 3.2.1.5.1). Il fattore tipo di isola
non ha alcuna influenza nel determinare la distribuzione delle specie predatrici di ricci,
mentre, almeno per quanto riguarda Giannutri, la protezione sembra avere un qualche
effetto sull’abbondanza di tali specie, dal momento che i siti inclusi nella zona 1 della
riserva mostrano valori mediamente più elevati di quelli facenti parte della zona 2.
Quanto osservato è stato confermato dalle analisi: il fattore tipo di isola non ha effetti
significativi sulle abbondanze dei predatori di ricci in nessuno dei modelli considerati,
mentre la significatività del fattore protezione permane anche una volta inclusi nel
modello gli effetti delle coordinate spaziali (Tab. 3.2.1.5.1).
Il modello completo è preferibile a quello privo degli effetti delle coordinate spaziali, in
base ai valori di AIC, LogLik e R2 (Tab 3.2.1.5.1). L’analisi dei residui è conforme alle
assunzioni del modello (vedi Fig. A.5, appendice A).
Figura 3.2.1.5.1 - Abbondanza delle specie predatrici di ricci (media ± E.S., 5< n <15) per sito, separatamente per ciascuna isola oggetto di studio (in rosso: zone 1; in blu: zone 2).
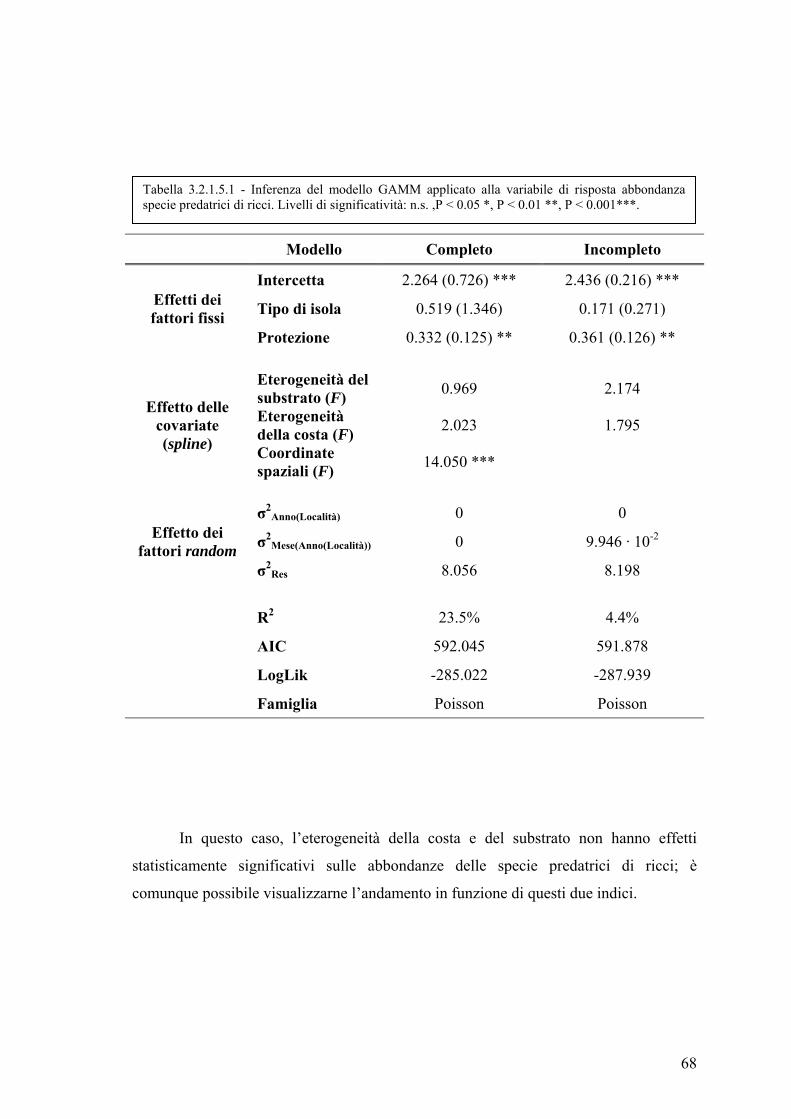
68
Modello Completo Incompleto
Effetti dei fattori fissi
Intercetta 2.264 (0.726) *** 2.436 (0.216) ***
Tipo di isola 0.519 (1.346) 0.171 (0.271)
Protezione 0.332 (0.125) ** 0.361 (0.126) **
Effetto delle covariate (spline)
Eterogeneità del substrato (F) 0.969 2.174
Eterogeneità della costa (F) 2.023 1.795
Coordinate spaziali (F) 14.050 ***
Effetto dei fattori random
σ2Anno(Località) 0 0
σ2Mese(Anno(Località)) 0 9.946 · 10-2
σ2Res 8.056 8.198
R2 23.5% 4.4%
AIC 592.045 591.878
LogLik -285.022 -287.939
Famiglia Poisson Poisson
In questo caso, l’eterogeneità della costa e del substrato non hanno effetti
statisticamente significativi sulle abbondanze delle specie predatrici di ricci; è
comunque possibile visualizzarne l’andamento in funzione di questi due indici.
Tabella 3.2.1.5.1 - Inferenza del modello GAMM applicato alla variabile di risposta abbondanza specie predatrici di ricci. Livelli di significatività: n.s. ,P < 0.05 *, P < 0.01 **, P < 0.001***.
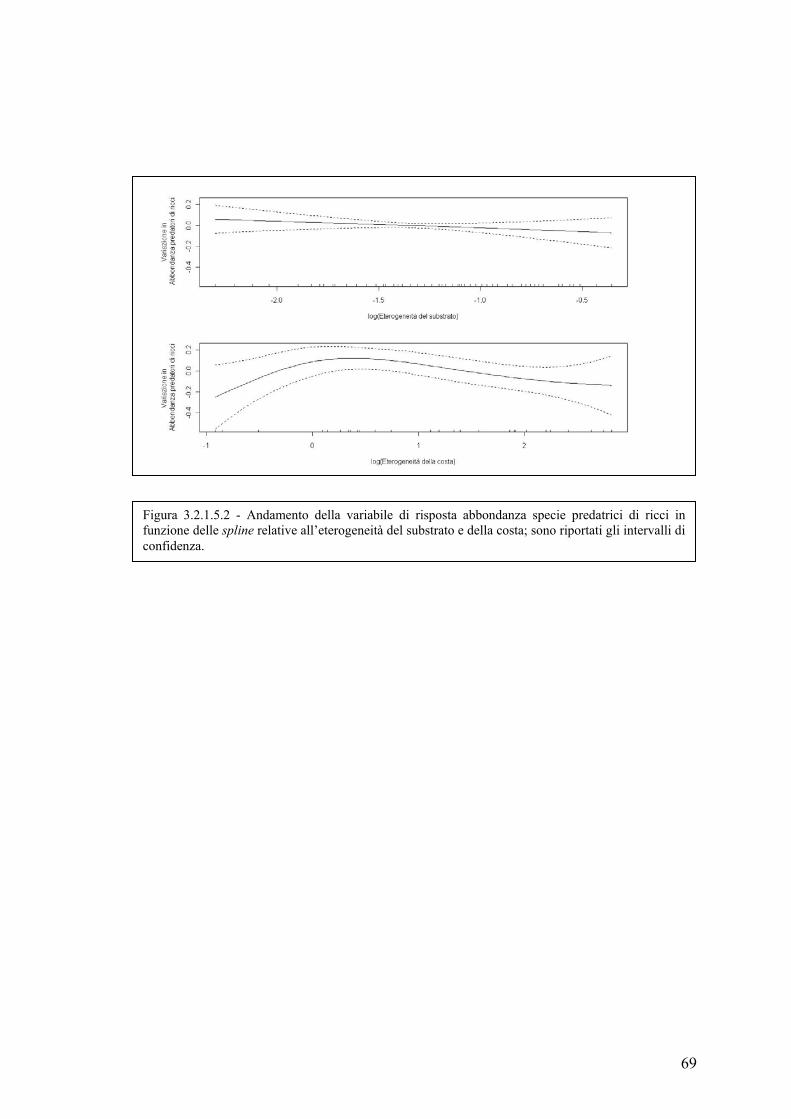
69
Figura 3.2.1.5.2 - Andamento della variabile di risposta abbondanza specie predatrici di ricci in funzione delle spline relative all’eterogeneità del substrato e della costa; sono riportati gli intervalli di confidenza.
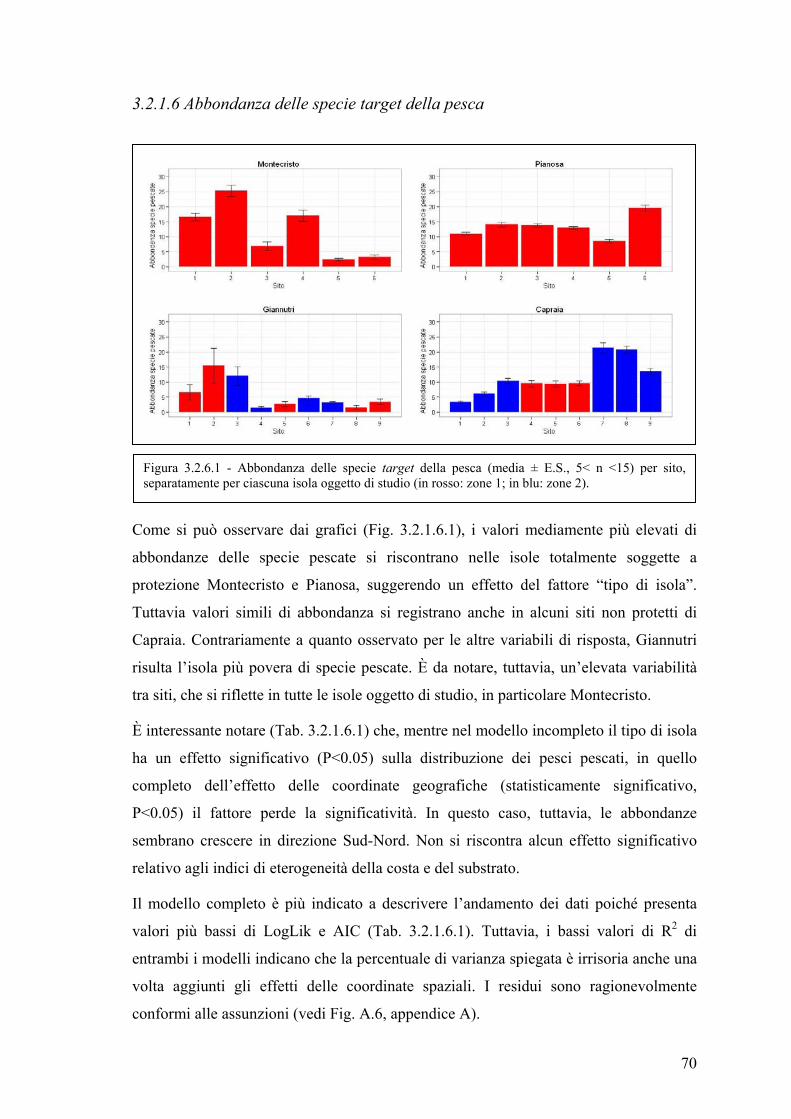
70
3.2.1.6 Abbondanza delle specie target della pesca
Come si può osservare dai grafici (Fig. 3.2.1.6.1), i valori mediamente più elevati di
abbondanze delle specie pescate si riscontrano nelle isole totalmente soggette a
protezione Montecristo e Pianosa, suggerendo un effetto del fattore “tipo di isola”.
Tuttavia valori simili di abbondanza si registrano anche in alcuni siti non protetti di
Capraia. Contrariamente a quanto osservato per le altre variabili di risposta, Giannutri
risulta l’isola più povera di specie pescate. È da notare, tuttavia, un’elevata variabilità
tra siti, che si riflette in tutte le isole oggetto di studio, in particolare Montecristo.
È interessante notare (Tab. 3.2.1.6.1) che, mentre nel modello incompleto il tipo di isola
ha un effetto significativo (P<0.05) sulla distribuzione dei pesci pescati, in quello
completo dell’effetto delle coordinate geografiche (statisticamente significativo,
P<0.05) il fattore perde la significatività. In questo caso, tuttavia, le abbondanze
sembrano crescere in direzione Sud-Nord. Non si riscontra alcun effetto significativo
relativo agli indici di eterogeneità della costa e del substrato.
Il modello completo è più indicato a descrivere l’andamento dei dati poiché presenta
valori più bassi di LogLik e AIC (Tab. 3.2.1.6.1). Tuttavia, i bassi valori di R2 di
entrambi i modelli indicano che la percentuale di varianza spiegata è irrisoria anche una
volta aggiunti gli effetti delle coordinate spaziali. I residui sono ragionevolmente
conformi alle assunzioni (vedi Fig. A.6, appendice A).
Figura 3.2.6.1 - Abbondanza delle specie target della pesca (media ± E.S., 5< n <15) per sito, separatamente per ciascuna isola oggetto di studio (in rosso: zone 1; in blu: zone 2).
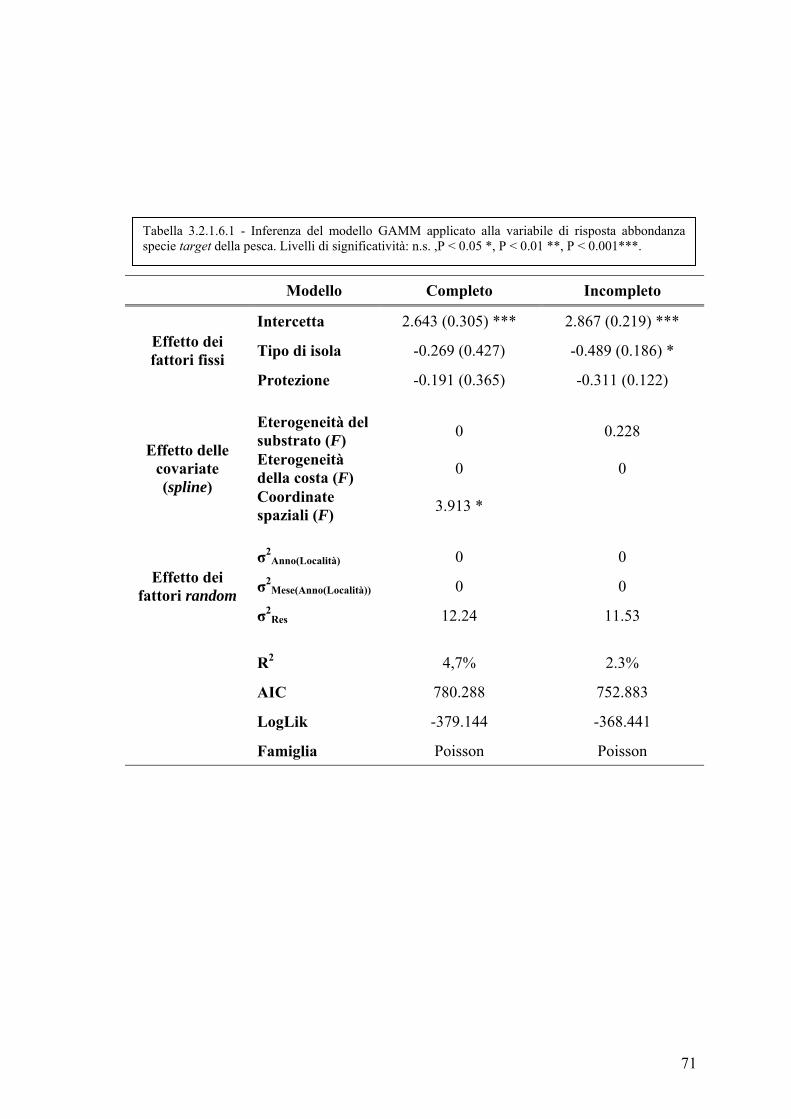
71
Modello Completo Incompleto
Effetto dei fattori fissi
Intercetta 2.643 (0.305) *** 2.867 (0.219) ***
Tipo di isola -0.269 (0.427) -0.489 (0.186) *
Protezione -0.191 (0.365) -0.311 (0.122)
Effetto delle covariate (spline)
Eterogeneità del substrato (F) 0 0.228
Eterogeneità della costa (F) 0 0
Coordinate spaziali (F) 3.913 *
Effetto dei fattori random
σ2Anno(Località) 0 0
σ2Mese(Anno(Località)) 0 0
σ2Res 12.24 11.53
R2 4,7% 2.3%
AIC 780.288 752.883
LogLik -379.144 -368.441
Famiglia Poisson Poisson
Tabella 3.2.1.6.1 - Inferenza del modello GAMM applicato alla variabile di risposta abbondanza specie target della pesca. Livelli di significatività: n.s. ,P < 0.05 *, P < 0.01 **, P < 0.001***.

72
3.2.1.7 Ricchezza specifica
I valori di ricchezza di specie si presentano omogenei in tutte le isole oggetto di studio,
con una bassa variabilità tra siti. Non si possono ipotizzare effetti dovuti ad alcuno dei
fattori presi in esame.
Figura 3.2.1.7.1 – Ricchezza specifica (media ± ± E.S., 5< n <15) per sito, separatamente per ciascuna isola oggetto di studio (in rosso: zone 1; in blu: zone 2).

73
L’analisi non evidenzia, infatti, alcun effetto significativo dovuto ai fattori inclusi nel
modello (tab 3.2.1.7.1).
Modello Completo Incompleto
Effetto dei fattori fissi
Intercetta 2.510 (0.084) *** 2.462 (0.054) ***
Tipo di isola -0.113 (0.119) 0.026 (0.056)
Protezione -0.045 (0.043) -0.049 (0.045)
Effetto delle covariate (spline)
Eterogeneità del substrato (F) 1.946 2.82 ·
Eterogeneità della costa (F) 0 0
Coordinate spaziali (F) 2.235
Effetto dei fattori random
σ2Anno(Località) 6.491 · 10-3 0
σ2Mese(Anno(Località)) 0 2.518 · 10-3
σ2Res 6.712 · 10-1 6.749 · 10-1
R2 1.7% 2.3%
AIC 30.034 24.314
LogLik -4.017 -4.157
Famiglia Poisson Poisson
Tabella 3.2.1.7.1 - Inferenza del modello GAMM applicato alla variabile di risposta ricchezza specifica. Livelli di significatività: n.s. ,P < 0.05 *, P < 0.01 **, P < 0.001***.

74
3.2.2 Valutazione degli effetti legati ai diversi regimi di protezione applicati
Al fine di testare il modello secondo il quale l’alternarsi di zone soggette a protezione e
zone impattate dalla pesca su isole parzialmente protette potesse generare modalità di
distribuzione dei popolamenti ittici differenti da quelli che si originano su isole
totalmente protette, la procedura di analisi è continuata testando le differenze tra due
modelli GAMM fittati sulle variabili di risposta.
Variabile di risposta Parzialmente protetta Totalmente protetta
Abbondanza totale 0.3123687
(0.2026386 – (0.4815184)
0.2407855
(0.1350659 – 0.4292546)
Abbondanza taglie piccole 0.5050329
(0.332402 – 0.767317)
0.2406786
(0.08549898– 0.6775076)
Abbondanza taglie medie 0.2779798
(0.1549602 – 0.4986621)
0.2408082
(0.1309230 – 0.4429213)
Abbondanza taglie grandi 0.333797
(0.1721135 – 0.647366)
0.5076149
(0.2794724 – 0.9219976)
Abbondanza predatori di ricci
0.4841298
(9.3029474 – 0.7736711)
0.4103107
(0.2377359 – 0.7081592)
Abbondanza pesci pescati 0.452103
(0.2401577 – 0.8510952) 0.1393757
Ricchezza di specie 0.006199124 0.06161307
(0.0180564 – 0.2102391)
Tabella 3.2.2.1 – Varianze medie stimate per ciascun tipo di isola (Parzialmente o Totalmente protetta) con l’intervallo di confidenza stimato. La significatività è valutata in base alla sovrapposizione degli intervalli di confidenza. In grassetto sono riportati i risultati significativi.

75
Non si riscontrano differenze statisticamente significative tra i due modelli in esame per
alcuna variabile di risposta (Tab. 3.2.2.1). Per quanto riguarda l’abbondanza dei pesci
pescati e la ricchezza specifica, sono riportate solo le varianze tra siti dal momento che
gli intervalli di confidenza non sono stimabili dal modello.

76
4. DISCUSSIONE E CONCLUSIONI
Gli ecosistemi marini costieri del Mediterraneo sono ambienti diversificati, complessi e
tridimensionali che mostrano eterogeneità strutturale su scale spaziali multiple. Nel
presente lavoro di tesi, si è ipotizzato che tale eterogeneità ambientale potesse costituire
un elemento di confusione nella valutazione dell’effetto riserva. I risultati ottenuti
hanno confermato questa ipotesi dal momento che, una volta introdotti nelle analisi gli
effetti di fattori ambientali e biologici variabili nello spazio (qui sintetizzati dagli indici
di eterogeneità di substrato e costa e dalla posizione geografica), l’effetto della
protezione perdeva la propria significatività statistica. Le analisi, infatti, non hanno
evidenziato alcun effetto dovuto né alla protezione né alla tipologia di isola, fattore che
doveva rendere conto delle differenze dovute ai diversi regimi di protezione applicati
alle isole oggetto di studio (protezione totale o parziale). Il fatto che non siano state
riscontrate differenze tra le aree soggette a protezione e quelle in cui la pesca è
consentita, per quanto riguarda l’abbondanza totale, ma soprattutto quella delle specie
pescate, suggerisce che presso le aree marine protette dell’Arcipelago Toscano non si
assista a risultati certo rimarchevoli relativamente alla protezione. Ci sono tuttavia
alcune eccezioni, soprattutto per quanto riguarda l’abbondanza delle specie predatrici di
ricci (gli sparidi Diplodus sargus e D. vulgaris e i labridi Coris julis e Thalassoma
pavo) la significatività della protezione rimane anche una volta aggiunti gli effetti delle
coordinate geografiche. Dal momento che il recupero delle popolazioni di sparidi
(specie di elevato valore commerciale), associato a basse densità di ricci e barrens poco
estesi (Guidetti e Sala 2007), è stato spesso osservato entro le AMP, la densità delle
specie di Diplodus è stata spesso assunta come indice della potenzialità di una riserva di
recuperare e successivamente mantenere rigogliosi popolamenti di macroalghe e con
essi l’intera pletora di funzioni ecosistemiche associate ai fondi duri costieri (Guidetti et
al. 2008). Quanto osservato fa quindi ben sperare riguardo a un potenziale
funzionamento delle AMP almeno negli anni a venire.
Molti sono i fattori che influenzano la risposta dei popolamenti ittici all’istituzione di
una riserva: quelli legati alla life-history delle specie presenti (disponibilità di
riproduttori, tassi di crescita individuali, età a cui la specie raggiunge la maturità
sessuale, quantità di uova emesse e frequenza di emissione, caratteristiche del ciclo
vitale, mobilità, dispersione e territorialità), la dimensione e l’età della riserva,

77
l’inclusione in una rete di AMP, l’eterogeneità degli habitat inclusi. Tuttavia, ciò che
realmente conta è l’efficacia delle misure di protezione: le considerazioni fatte, infatti,
valgono per AMP in cui le disposizioni di tutela sono effettivamente ed efficacemente
messe in atto. Un numero sempre maggiore di studi ha attestato che, dove si riscontra
una effettiva ripresa del popolamento ittico, il pattern di recupero di tale popolamento è
strettamente correlato all’efficacia delle misure di tutela messe in atto (Guidetti et al.,
2008). Pertanto, nonostante gli ottimi requisiti del Parco Nazionale dell’Arcipelago
Toscano (1. istituito nel 1996, con alcune isole già sotto regime di tutela, tra cui,
Montecristo dal 1971, come Riserva Naturale Biogenetica e Pianosa dal 1858, in quanto
colonia penale agricola; 2. le sette isole ivi comprese rappresentano il più grande
sistema di Aree Marine Protette dell’Europa, con i loro 61.474 Km2 di acque protette; 3.
i fondali dell’arcipelago includono una vasta gamma di habitat, tra cui praterie di
Posidonia oceanica, fondi duri costieri, coralligeno, grandi estensioni di sabbia e il
caratteristico ambiente costituito da alghe calcaree mobili, maerl), i risultati ottenuti in
questo studio suggeriscono che le misure di protezione non sono adeguatamente messe
in atto. Quanto osservato non è lontano dalla situazione generale di molte AMP italiane,
annoverate tra i cosiddetti “paper parks” (Mora et al., 2006), cioè riserve solo sulla
carta, nelle quali gli studi volti a testare l’efficacia della protezione sono di fatto inutili
(Guidetti et al., 2008). Per quanto riguarda le isole oggetto di studio, l’inefficacia delle
misure di protezione si coniugherebbe anche a una inadeguatezza delle scelte alla base
dell’istituzione delle AMP i cui confini sono stati tracciati non tenendo conto di criteri
scientifici bensì sulla base di compromessi economici e politici. Il fatto che, nelle isole
dell’arcipelago, sia l’eterogeneità ambientale uno dei principali fattori che governano le
abbondanze dei popolamenti ittici, conferma quanto ipotizzato.
Dai risultati del presente lavoro di tesi emerge in maniera preponderante, infatti,
l’importanza dei processi ambientali variabili nello spazio nel determinare le modalità
dei distribuzione dei popolamenti ittici. Nelle analisi sono stati inclusi gli effetti
dell’eterogeneità del substrato, della costa e della posizione geografica.
Per quanto riguarda l’eterogeneità del substrato, tale fattore è risultato statisticamente
significativo solo nel caso della ricchezza specifica. Bisogna tener presente, tuttavia,
che è stato calcolato come indice di rugosità, che tiene conto dei rilievi e delle asperità
del fondo, ma non dà indicazioni sulle tipologie di habitat incontrati nel transetto.

78
Gli effetti dell’eterogeneità della costa, invece, sono risultati statisticamente
significativi per tre variabili di risposta (abbondanza totale, delle taglie piccole e
grandi); in generale, le abbondanze maggiori si registrano per valori intermedi di
eterogeneità della costa. Questo indice è verosimilmente correlato con la tipologia degli
ambienti sommersi nelle vicinanze della riva. Si può supporre che a livelli bassi
dell’indice corrispondano ambienti poco diversificati e quindi poco adatti per la fauna
ittica. I valori più elevati dell’indice sono invece dovuti alla presenza di baie e cale,
anch’esse poco ospitali per i pesci. A valori intermedi l’indice dovrebbe invece riflettere
un livello di eterogeneità dell’ambiente sommerso appropriato per la fauna ittica.
L’importanza dell’eterogeneità della costa evidenziata da questa tesi legittima una
verifica diretta tra i valori assunti dall’indice e la tipologia di habitat lungo la costa. Ciò
rimane come prospettiva di indagine futura.
Il risultato più eclatante evidenziato in questa tesi è quello relativo agli effetti della
posizione geografica: è stato riscontrato un gradiente latitudinale di abbondanze
crescenti in direzione Nord-Sud. Le isole situate a Nord dell’arcipelago (Capraia e
Pianosa) hanno mostrato valori mediamente più bassi di abbondanze rispetto alle isole
poste a Sud (Montecristo e Giannutri). L’esistenza di un gradiente latitudinale nelle
abbondanze delle specie ittiche potrebbe essere legato alla oceanografia locale, a
differenze di microclima, alle tipologie di habitat presenti e alle preferenze tipiche di
ciascuna specie. Bisogna tenere presente, infatti, che le isole considerate in questo
studio presentano fondali molto eterogenei e caratteristiche geologiche assai differenti.
Nonostante, ad oggi, non ci sia evidenza sperimentale che descriva i meccanismi
specifici attraverso cui le caratteristiche fisico-chimiche delle rocce (composizione
mineralogica, grana, microtopografia) influenzino gli organismi marini ad esse
associati, un crescente numero di studi suggerisce tali caratteristiche del substrato siano
in grado di influenzare i popolamenti marini sia direttamente (es. l’epibenthos sessile:
Bavestrello et al. 2000, Cattaneo-Vietti et al. 2002), sia indirettamente (es. i pesci:
Guidetti e Cattaneo-Vietti 2002). Le caratteristiche mineralogiche del substrato
influenzano la colonizzazione di fondi molli da parte dell’infauna (Cerrano et al. 1998),
l’insediamento degli idroidi e la struttura delle comunità epibentoniche dei fondi duri
(Marini e Luna 2003), le modalità di distribuzione di molluschi vermetidi (Schiapparelli
et al. 2003) e lo sviluppo larvale di pesci d’acqua dolce (Maradonna et al. 2003).
L’attività di organismi endolitici (spugne clionidi e bivalvi), la cui attività litica di
esplica principalmente su substrati ricchi di carbonati e può essere ostacolata dalla

79
presenza di quarzo, dipende strettamente dalla composizione delle rocce del fondo. Lo
studio di Guidetti et al. (2004), infine, ha evidenziato l’esistenza di differenze a livello
di copertura algale e popolamenti ittici associati a fondi con differenti caratteristiche
mineralogiche. I fondi a granito, che presentano un elevato contenuto di quarzo, il quale
ha effetti fisiologici negativi sugli organismi a causa delle sue proprietà ossidanti,
mostrano popolamenti algali costituiti principalmente da alghe filamentose e
caratterizzati da un minor numero di specie rispetto a fondi calcarei. Anche i
popolamenti ittici presentano numerose differenze in termini di composizione in specie.
I fondi granitici ospitano un maggior numero di individui del serranide Serranus scriba
e dei labridi del genere Symphodus, notoriamente influenzati dalle caratteristiche del
popolamento macroalgale presente, nel quale trovano rifugio e predano i piccoli
invertebrati vagili di cui si nutrono (Ruitton et al. 2000, Letourneur et al. 2003, Garcia-
Rubies e Macpherson 2005). Molte specie ittiche e di invertebrati, infatti, sono associati
a fondi caratterizzati dalla presenza di specie strutturanti, principalmente macroalghe o
animali coloniali, che generano habitat complessi, in cui trovare riparo e cibo (Turner et
al. 1999, Chemello e Milazzo 2002). I fondi calcarei, invece, mostrano elevate
abbondanze del labride Thalassoma pavo, del serranide Serranus cabrilla e di specie
cripto bentoniche, come blennidi e gobidi. Nel caso di questi ultimi, come riportato di
Guidetti e Cattaneo-Vietti (2002), potrebbe essere il colore del substrato a determinarne
le elevate abbondanze: i colori chiari del calcare favoriscono il loro mimetismo col
fondo. Le isole oggetto di studio presentano composizione mineralogica molto
eterogenea: Capraia ha origine vulcanica; Pianosa è costituita da rocce sedimentarie e
da accumuli conchiliferi che racchiudono fossili marini; Giannutri da sedimenti
calcareo-dolomitici; Montecristo è interamente granitica. È plausibile supporre,
pertanto, che parte delle differenze in termini di abbondanza composizione in specie del
popolamento ittico siano imputabili alla differente composizione mineralogica e alla
conseguente differente copertura vegetale del substrato.
Un altro aspetto da considerare è l’eterogeneità del fondo in termini di tipologia di
habitat disponibile: i fondali mediterranei, e le isole oggetto di studio non fanno
eccezione, sono generalmente costituiti da un mosaico di substrato roccioso intersperso
a estensioni di sabbia e ciottoli e a patch di praterie di Posidonia oceanica. Tali habitat
ospitano generalmente popolamenti ittici differenti in termini di abbondanze, densità e
composizione in specie (Guidetti et al. 2000). In particolare, i fondi vegetati,
rappresentati nel Mediterraneo dalle praterie di fanerogame marine, supportano

80
l’esistenza di popolamenti ittici più ricchi, con un numero più elevato di giovanili,
rispetto ai fondi “spogli” adiacenti (Kikuchi 1966, Orth & Heck 1980, Pollard 1984,
Rozas e Odum 1988, Bell e Pollard 1989, Conrow et al. 1990, West & King 1996).
Questo fenomeno viene spiegato considerando l’elevata complessità strutturale e
tridimensionalità di questi sistemi vegetali che forniscono ombra, rifugio, cibo a un gran
numero di specie ittiche costiere. Inoltre, riguardo ai sistemi a fanerogame mediterranei,
molti autori (Harmelin-Vivien et al. 1995, Francour 1997, Francour e Le Direac’h
1994,) hanno riportato che praterie di P. oceanica, situate in aree riparate a bassa
profondità, hanno il ruolo cruciale di aree di nursery per molte specie ittiche. Specie
esclusive di questo habitat sono gli sparidi Diplodus annularis e Spondiliosoma
cantharus e il labride Symphodus ocellatus. Altrettanto importanti sono i fondi duri
costieri mediterranei, caratterizzati dalla presenza di popolamenti algali fotofili e
popolamenti ittici fondamentali per pesca: specie esclusiva di questi habitat è lo sparide
di interesse commerciale Diplodus sargus, oltre ai labridi Symphodus tinca, S. roissali e
S. mediterraneus. Infine, le distese sabbiose sommerse sono l’habitat caratteristico di
specie criptobentoniche come alcuni gobidi, trachinidi, e pesci target della piccola
pesca come mugilidi e mormore (Lithognatus mormyrus). Andando ad osservare la
distribuzione di questi habitat a livello dei siti considerati nel presente studio, si può
ipotizzare che parte della variabilità associata agli effetti della latitudine, potrebbe
derivare dall’inclusione o meno di una uguale proporzione di fondo roccioso, sabbioso
o di prateria di P. oceanica.
Possono essere fatte anche alcune considerazioni di carattere biogeografico. Alcuni
autori (vedi ad esempio Bianchi 2004) considerano l’isola d’Elba come il confine tra
due settori biogeografici del Mediterraneo: il bacino ligure – provenzale, caratterizzato
da numerose specie ad affinità boreale e poche ad affinità calda, e il bacino tirrenico, in
cui si registrano numerosi endemismi, la presenza di un numero elevato di specie ad
affinità sub-tropicale e in generale abbondanze maggiori delle specie ittiche. Quanto
affermato potrebbe trovare riscontro dal fatto che, a Capraia, l’isola situata più Nord, al
di sopra dell’Elba, si osservano i valori più bassi di abbondanze totale. Ulteriore
conferma potrebbe derivare anche dall’analisi della distribuzione della specie termofila
Thalassoma pavo, un piccolo labride proteroginico che abita i fondali rocciosi a bassa
profondità del Mediterraneo e dell’ oceano Atlantico Nord-orientale. Questa specie
risulta più abbondante nei settori meridionali del Mediterraneo dal momento la sua
distribuzione è limitata a Nord dalle temperature superficiali dell’acqua relativamente

81
più basse. Tuttavia, ad oggi, piccole popolazioni si ritrovano anche sulle coste della
Liguria, probabilmente a causa dell’innalzamento delle temperature terrestri e marine
(Guidetti et al. 2002). Nel presente studio, valori di abbondanze maggiori si riscontrano
a Giannutri (242), seguita da Pianosa (81) e infine Capraia (15). Anche Montecristo, a
dispetto della sua posizione, presenta solo 14 individui, ma in questo caso, il fattore
determinante potrebbe essere la mancanza di in habitat adatto. T. pavo predilige, infatti,
fondi rocciosi a bassa profondità con lieve pendenza (Velte 1998), ambiente
scarsamente rappresentato a Montecristo. Allo scopo di valutare la plausibilità
dell’ipotesi “biogeografia”, sarebbe, infine, interessante andare ad analizzare
l’andamento delle correnti e i pattern di dispersione di larve, propaguli e individui
adulti all’interno delle acque dell’arcipelago.
Tutte le ipotesi relative ai risultati osservati richiedono rigorose analisi statistiche per
essere confermate ed evidenziano la necessità di studi scientifici accurati non solo a
seguito dell’istituzione di una riserva, per valutarne l’efficacia, ma anche prima, al fine
di scegliere con cura la sua localizzazione e le misure di protezione da applicare.
Quanto osservato deve far riflettere in merito alle lacune che a tutt’oggi si hanno nella
conoscenza scientifica sulle Aree Marine Protette: il presente lavoro conferma quanto
affermato dallo studio Sale et al. 2005, che individua la mancanza di nozioni adeguate
su modalità di dispersione larvale, movimenti delle specie ittiche durante la fase
giovanile e adulta, impatti ecologici della pesca all’interno delle riserve, andamento
delle masse d’acqua in prossimità delle coste considerate e importanza dell’eterogeneità
spaziale.
In conclusione, si può affermare che, nel caso delle isole di Capraia, Pianosa
Montecristo e Giannutri, gli effetti dell’eterogeneità ambientale sono più importanti di
quelli della protezione nel determinare le modalità di distribuzione delle specie del
popolamento ittico e che i differenti regimi di protezione applicati nell’arcipelago (isole
totalmente protette vs isole parzialmente protette) non influenzano in alcun modo le
abbondanze delle specie ittiche. Indipendentemente dalla tipologia di misure protettive
applicate, le abbondanze sono elevate per valori intermedi di eterogeneità della costa e
crescono secondo un gradiente latitudinale: le isole situate a Nord dell’arcipelago
(Capraia e Pianosa) hanno mostrato valori mediamente più bassi delle isole poste a Sud
(Montecristo e Giannutri). Questo studio evidenzia l’importanza dell’eterogeneità
ambientale nell’influenzare i popolamenti ittici costieri, le sue implicazioni nella

82
pianificazione e gestione delle riserve e la necessità che questo fattore sia inserito negli
studi volti a valutare l’efficacia delle misure di protezione.
Al di là di obiettivi di carattere sociale (ad esempio, salvaguardia delle comunità di
pescatori) ed economico (turismo ad esempio), una AMP deve garantire la
conservazione di una vasta gamma di habitat e di specie, scelti sulla base di accurati
studi scientifici che identifichino il set di condizioni ambientali (valori di eterogeneità di
costa e substrato, posizione, esposizione) che massimizzino le abbondanze dei
popolamenti e la loro persistenza, e che non sono il risultato di compromessi economici
e politici. Studi rigorosi e ben progettati consentirebbero anche di colmare le lacune
presenti nella conoscenza scientifica, che impediscono lo sviluppo di una vera e propria
scienza delle Aree Marine Protette (Sale et al. 2005), requisito imprescindibile per la
creazione di criteri quantitativi attendibili da usare nel momento in cui si devono
istituire nuove AMP.

83
5. BIBLIOGRAFIA
Allison, G.W., J. Lubchenco, M.H. Carr, 1998. Marine reserves are necessary but not
sufficient for marine conservation. Ecological Application 8: 79-92.
Almany, G.R., M.L. Berumen, S.R. Thorrold, S. Planes, and G.P. Jones, 2007. Local
replenishment of coral reef fish populations in a marine reserve. Science 316:
742-744.
Ashworth, J.S., and R.F.G. Ormond, 2005. Effects of fishing pressure and trophic group
on abundance and spillover across boundaries of a no-take zone. Biological
Conservation 121: 333-344.
Attwood, C.G., J.M. Harris e A.J. Williams, 1997. International experience of marine
protected areas and their relevance to South Africa. S Afr J Mar Sci 18: 311-332.
Ballantine, W.J., 1991. Marine reserves for New Zealand. Leigh Laboratory Bullettin
Number 25. University of Auckland, Auckland, New Zealand.
Banks, S.A. & G.A. Skilleter, 2007. The importance of incorporating fine-scale habitat
data into the design of an intertidal marine reserve system. Biol Conserv 138: 13-
29.
Bavestrello G., C.N. Bianchi, B. Calcinai, R. Cattaneo-Vietti, C. Cerrano, C. Morri, S.
Puce, M. Sarà, 2000. Bio-mineralogy as a structuring factor for marine epibenthic
communities. Mar Ecol Prog Ser 193: 241-249.
Beaumont, J., 1997. Community participation in the establishment and management of
marine protected areas: a review of selected international experience. S Afr J Mar
Sci 18: 333-340.
Bell, J.D., 1983. Effects of depth and marine reserve fishing restrictions on the structure
of a rocky reef fish assemblage in the North-Western Mediterranean Sea. J Appl
Ecol 20: 357-369.
Bell, J. D. & D.A. Pollard, 1989. Ecology of fish assemblages and fisheries associated
with seagrasses. In The Biology of Seagrasses: An Australian Perspective
(Larkum, A.W.D., A.J. McComb & S.A. Shepherd, Eds). Elsevier, Amsterdam,
565-609.
Bell, S.S., E.D. McCoy, H.R. Mushinsky (Eds.), 1991. Habitat structure. The physical
arrangement of objects in space. Chapman & Hall, London.

84
Benedetti-Cecchi, L., F. Bulleri, F. Cinelli. 1998. Density dependent foraging in two
species of sea urchins in shallow subtidal reefs on the west coast of Italy (western
Mediterranean). Mar Ecol Prog Ser, 163: 203-211.
Benedetti-Cecchi, L., 2003. Disegno sperimentale ed analisi in ecologia. In Manuale di
metodologie di campionamento e studio del benthos marino Mediterraneo, Gambi
M. C., M. Dappino Eds. Biologia Marina Mediterranea 10 (1): 433-484.
Benedetti-Cecchi, L., I. Bertocci, F. Micheli, E. Maggi, T. Fosella & S. Vaselli, 2003.
Implications of spatial heterogeneity for management of marine protected areas
(MPAs): examples from assemblages of rocky coasts in the northwest
Mediterranean. Mar Environ Res 55: 429-458.
Beukers-Stewart, B. D., and G. P. Jones. 2004. The influence of prey abundance on the
feeding ecology of two piscivorous species of coral reef fish. Journal of
Experimental Marine Biology and Ecology 299: 155–184.
Bianchi, C.N., 2004. Proposta di suddivisione dei mari italiani in settori biogeografici.
Notiziario della società italiana di Biologia Marina 46: 57-59.
Birkeland, C. & P. Dayton, 2005. The importance in fishery management of leaving the
big ones. Trends Ecol Evol 20: 356-358.
Blyth-Skyrme, R.E., M.J. Kaiser, J.G. Hiddink, G. Edwards-Jones, and P.J.B. Hart,
2006. Conservation benefits of temperate marine protected areas: Variation
among fish species. Conservation Biology 20: 811–820.
Boersma, P.D. and J.K. Parrish, 1999. Limiting abuse: marine protected areas, a limited
solution. Ecol Econ 31: 287-304.
Botsford, L.W. et al., 2001. Dependence of sustainability on the configuration of marine
reserves and larval dispersal distance. Ecol Lett 4: 144-150.
Botsford, L.W., F. Micheli, A. Hastings, 2003. Principles for the design of marine
reserves. Ecol Appl 13: S25–S31.
Booth D.J., D.M. Brosnan, 1995. The role of recruitment dynamics in rocky shore and
coral reef fish communities. Adv Ecol Res 26: 309-385.
Boujard, T., J.F. Leatherland, 1992. Circadian rhythms and feeding time in fishes.
Environ Biol Fish 35: 109-131.
Burnham K.P. and D. Anderson, 2002. Model selection and multimodel inference: A
practical information – theroretic approach. Springer-Verlag, New York, 488 pp.
Caley, M.J., et al., 1996. Recruitment and the local dynamic of open marine population.
Annu Rev Ecol Syst 27: 477-500.

85
Carlton, J.T., 1999. The scale and ecological consequences of biological invasions in
the world’s oceans. In Invasive species and biodiversity management, O.T.
Sandlund, P.J. Schei and A. Viken Eds., Kluwer Publishers, Dordrecht,
Netherlands, 195-212.
Carr, M.H., D.C. Reed, 1993. Conceptual Issues Relevant to Marine Harvest Refuges:
Examples from Temperate Reef Fishes. Can J Fish Aquat Sci 50: 2019-2028.
Caselle, J.E., R.R. Warner, 1996. Variability in recruitment of coral reef fishes, the
importance of habitat at two spatial scales. Ecology 77 :2488-2504.
Cattaneo-Vietti, R., G. Albertelli, G. Bavestrello, C.N. Bianchi and 5 others, 2002. Can
rock composition affect sublittoral epibenthic communities? PSZN I: Mar Ecol
23: 65–77.
Cerrano, C., A. Arillo, G. Bavestrello, U. Benatti and 7 others, 1998. Organism–quartz
interactions in structuring benthic communities: towards a marine bio-
mineralogy? Ecol Lett 2: 1–3
Chabanet P., V. Dufour, R. Galzin, 1995. Disturbance impact on reef fish communities
in Reunion Island (Indian Ocean). J Exp Mar Biol Ecol 188: 29-48.
Chapman, M.G., A.J. Underwood, G.A. Skilleter, 1995. Variability at different spatial
scales between a subtidal assemblage exposed to the discharge of sewage and two
control assemblages. J Exp Mar Biol Ecol 189: 103-122.
Chemello R, M. Milazzo, 2002. Effect of algal architecture on associated fauna: some
evidence from phytal molluscs. Mar Biol 140: 981–990.
Claudet, J., D. Pelletier, J.-Y. Jouvenel, F. Bachet, and R. Galzin, 2006a. Assessing the
effects of marine protected area (MPA) on a reef fish assemblage in a
northwestern Mediterranean case study: identifying community-based indicators.
Biological Conservation 130:349–369.
Claudet, J., et al. 2008. Marine reserves: Size and age do matter. Ecology Letters
11:481–489.
Claudet, J., C.W. Osemberg, P. Dominici, F. Badalamenti, M. Milazzo, J.M. Falcón, I.
Bertocci, L. Benedetti-Cecchi, J. A. García-Charton, R. Goñi, P. Afonso, A. Brito,
I. Guala, L. Le Díreach, P. Sanchez-Jerez, P.J. Somerfield, S. Planes, 2010.
Marine reserves: fish life history and ecological traits matter. Ecological
Applications 20 (3): 830-839.

86
Conrow, R., A.V. Zale & R.W. Gregory, 1990 Distributions and abundances of early
life stages of fish in a Florida lake dominated by aquatic macrophytes.
Transactions of the American Fisheries Society 119: 521-528.
Côté, I.M., I. Mosqueira & J.D. Reynolds, 2001. Effects of marine reserve
characteristics on the protection of fish populations: a meta-analysis. J Fish Biol
59: 178-189.
Cottenie, K. (2005). Integrating environmental and spatial processes in ecological
community dynamics. Ecol Lett 8: 1175-1182.
Crawley, M.J., 2007. The R book. J. Wiley & Sons, Ltd.
Cowen, R.K., C.B. Paris & A. Srinivasan, 2006. Scaling of connectivity in marine
populations. Science 311: 522-527.
Dayton, P.K., M.J. Tegner, 1984. The importance of scale in community ecology: a
kelp forest example with terrestrial analogs. In: P.W. Price, C.N. Slobodehikoff,
W.S. Gaud (Eds.), A New Ecology. Novel Approaches to Interactive System.
Wiley, New York, 457-481.
Dayton, P.K., S.F. Thrush, M.T. Agardy, R.J. Hofman, 1995. Environmental effects of
marine fishing. Aquatic Conservation 5: 205-232.
DeMartini, E.E. 1993. Modeling the potential of fishery reserves for managing Pacific
coral reef fishes. Fisheries Bulletin 91:414–427.
Doherty, P.J., 1991. Spatial and temporal patterns in recruitment. In Sale P.F. (Ed.) The
ecology of fishes on coral reefs. Academic Press, New York, 261-293.
Downing, J.A., 1986. Spatial heterogeneity, evolved behaviour or mathematical
artifact? Nature 323:255-257.
Dungan, J.E. and G.E. Davis, 1993. Applicatios of marine refugia to coastal fisheries
management. Can J Fish Aquat Sc 50: 2029-2042.
Eggleston, D.B., and D.M. Parsons, 2008. Disturbance-induced ‘‘spill-in’’ of Caribbean
spiny lobster to marine reserves. Marine Ecology Progress Series 371:213–220.
FAO, 2002. Report and documentation of the International Workshop on Factors of
Unsustainability and Overexploitation in Fisheries (Bankok, Thailand, 4-8
Febraury, 2002). FAO Fishery report R672.
Foley, P., 1994. Predicting extinction times from environmental stochasticity and
carrying capacity. Conserv Biol 8: 124-137.
Fowler, A.J., P.J. Doherty, D.M. Williams, 1992. Multiscale analysis of recruitment of a
coral reef fish on the Great Barrier. Reef Mar Ecol Prog Ser 82:131-141.

87
Folke, C., S. Carpenter, T. Elmqvist, L. Gunderson, C.S. Holling, B. Walker, 2002.
Resilience and sustainable development: building adaptive capacity in a world of
transformations. Ambio 31: 437-440.
Francour, P., 1997 Fish assemblages of Posidonia oceanica beds at Port-Cros (France,
NW Mediterranean): assessment of composition and long-term fluctuations by
visual census. P.S.Z.N. I: Marine Ecology 18: 157-173.
Francour, P. & L. Le Direac’h, 1994. Recrutement de l’ichtyofaune dans l’herbier
superficiel à Posidonia oceanica de la réserve naturelle de Scandola (Corse,
Méditerranée nord-occidentale): données préliminaires. Travaux Scientifics du
Parc National et Régional Réserve Corse 46, 71–91.
Friedlander, A. M., 2001. Essential fish habitat and the effective design of marine
reserves: application for marine ornamental fishes. Aquarium Sci Conserv 3: 135-
150.
Frisch, A., R. Baker, J.P. Hobbs, and L. Nankervis. 2008. A quantitative comparison of
recreational spearfishing and linefishing on the Great Barrier Reef: implications
for management of multi-sector coral reef fisheries. Coral Reefs 27: 85–95.
Froese, R., D. Pauly, 2004. Fish Base. World Wide Web Electronic Publication.
<http://www.fishbase.org>.
Fujita, R.M., 2001. Why Marine Reserves? Environmental Defense, 1- 30.
García-Charton, J.A., and A. Pérez-Ruzafa, 1998. Correlation between habitat structure
and a rocky reef fish assemblage in SW Mediterranean. PSZN I, Mar Ecol 19:
111-128.
García-Charton, J.A., and A. Pérez-Ruzafa, 1999. Ecological heterogeneity and the
evaluation of the effects of marine reserves. Fisheries Research 42:1–20.
García-Charton, J.A., I. Williams, A. Pérez-Ruzafa, M. Milazzo, R. Chemello, C.
Marcos, M-S Kitsos, A. Koukouras, S. Riggio, 2000. Evaluating the ecological
effects of Mediterranean marine reserves: habitat, scale and the natural variability
of ecosystems. Environ Conserv 27: 159-178.
García-Charton, J.A., A. Pérez-Ruzafa, P. Sánchez-Jerez, J.T. Bayle-Sempere, O.
Reñones, D.Moreno, 2004. Multi-scale spatial heterogeneity, habitat structure and
the effect of marine reserves on Western Mediterranean rocky reef fish
assemblage. Mar Biol 144: 161-182.
Garcìa-Rubies A, E. Macpherson, 1995. Substrate use and temporal pattern of
recruitment in juvenile fishes of the Mediterranean littoral. Mar Biol 124: 35–42.

88
Gladfelter W.B., J.C. Ogden, E.H. Gladfelter, 1980. Similarity and diversity among
coral reef fish community: a comparison between tropical Western Atlantic
(Virgin Islands) and tropical Central Pacific (Marshall Islands) patch reef.
Ecology 61: 1156-1168.
Glasby, T.M., 1998. Estimating spatial variability in developing assemblages of
epibiota on subtidal hard substrata. Mar Freshw Resear 49: 429-437.
Glasby, T.M. e A.J. Underwood, 1998. Determining position for control locations in
environmental studies of estuarine marinas. Mar Ecol Progr Series 171: 1-14.
Goñi, R., A. Quetglas & O. Reñones, 2006. Spillover of lobsters Palinurus elephas
from a marine reserve to an adjoining fishery. Mar Ecol Prog Ser 308, 207-219.
Goñi, R., et al., 2008. Spillover from six western Mediterranean marine protected areas:
evidence from artisanal fisheries. Marine Ecology Progress Series 366: 159–174. Guidetti P., 2000. Differences Among Fish Assemblages Associated with Nearshore
Posidonia oceanica Seagrass Beds, Rocky–algal Reefs and Unvegetated Sand
Habitats in the Adriatic Sea. Estuarine, Coastal and Shelf Science 50: 515-529.
Guidetti P., C.N. Bianchi, G. La Mesa, M. Modena, C. Morri, G. Sara, M. Vacchi,
2002. Aboundance and size structure of Thalassoma pavo (Pisces: Labridae) in
the Western Mediterranean Sea: variability at different spatial scales. J Mar Biol
Ass U.K. 82: 495-500.
Guidetti P., R. Cattaneo-Vietti, 2002. Can mineralogical features influence distribution
patterns of fish? A case study in shallow Mediterranean rocky reefs. J Mar Biol
Assoc UK 82: 1043-1044.
Guidetti P., C.N. Bianchi, M. Chiantore, S. Schiaparelli, C. Morri, R. Cattaneo-Vietti,
2004. Living on the rocks: substrate mineralogy and the structure of subtidal
rocky substrate communities in the Mediterranean Sea. Mar Ecol Prog Ser 274:
57-68.
Guidetti, P., 2006. Marine reserves reestablish lost predatory interactions and cause
community changes in rocky reefs. Ecological Applications 16(3): 936-976.
Guidetti, P., 2006. Potential of Marine Reserves to Cause Community-Wide Changes
beyond Their Boundaries. Conservation Biology 21(2): 540-545.
Guidetti, P., E. Sala, 2007. Community-wide effects of marine reserves in the
Mediterranean Sea. Mar Eco Prog Ser 335: 43-56.
Guidetti, P., M. Milazzo, S. Bussotti, A. Molinari, M. Murenu, A. Pais, N. Spano`, R.
Balzano, T. Agardy, F. Boero, G. Carrada, R. Cattaneo-Vietti, A. Cau, R.

89
Chemello, S. Greco, A. Manganaro, G. Notarbartolo di Sciara, G. Fulvio Russo,
L. Tunesi, 2008. Italian marine reserve effectiveness: does enforcement matter?
Biological Conservation 141: 699-709.
Gunderson, L.H., 1999. Resilience, flexibility and adaptive management-antidotes for
spurious certitude? Conservation Ecology 3(1): 7.
Halpern, B.S. & R.R. Warner, 2002. Marine reserves have rapid and lasting effects.
Ecol Lett 5: 361–366.
Halpern, B.S., 2003. The impact of marine reserves: Do reserves work and does reserve
size matter? Ecological Applications 13: 117-137.
Harmelin J-G, 1990. Ichtyofaune des fonds rocheux de Méditerranée: structure du
peuplement du coralligène de l’île de Port-Cros (Parc National, France). Mésogée
50: 23-30.
Harmelin-Vivien, M., J.G. Harmelin, 1975. Présentation d’une méthode
d’évaluation “in situ ” de la faune ichtyologique. Travaux Scientifiques du Parc
National de Port-Cros 1: 47-52.
Harmelin-Vivien, M.L., J.G. Harmelin, C. Chauvet, C. Duval and 7 others, 1985.
Evaluation des peuplements et populations de poissons: méthodes et problèmes.
Revue Ecologie Terre Vie 40: 467-539.
Harmelin, J.G., F. Bachet e F. Garcia, 1995. Mediterranean marine reserves: fish indices
as tests of protection efficiency. Pubblicazioni della stazione zoologica di Napoli:
Marine Ecology 16: 223-250.
Harmelin-Vivien, M., J.G. Harmelin & V. Leboulleux, 1995. Microhabitat requirements
for settlement of juvenile sparid fishes on Mediterranean rocky shores.
Hydrobiologia 300–301, 309–320.
Harmelin-Vivien, M., L. Le Diréach, J. Bayle-Sempere, E. Charbonnel, J. A. García-
Charton, D. Ody, A. Pérez-Ruzafa, O. Reñones, P. Sánchez-Jerez, and C. Valle,
2008. Gradients of abundance and biomass across reserve boundaries in six
Mediterranean marine protected areas: evidence of fish spillover? Biological
Conservation 141: 1829-1839.
Hastings, A., and L.W. Botsford. 1999. Equivalence in yield from marine reserves and
traditional fisheries management. Science 284: 1537-1538.
Hastings, A. & L.W. Botsford, 2003. Comparing designs of marine reserves for
fisheries and for biodiversity. Ecol Appl 13: S65-S70.

90
Hickford, M.J.H., and D.R. Schiel, 2008. Experimental gillnetting of reef fish: species-
specific responses modify capture probability across mesh sizes. Journal of
Experimental Marine Biology and Ecology 358: 163-169.
Hixon M.A., 1991. Predation as a process structuring coral reef fish communities. In
Sale P.F. (Ed.) The ecology of fishes on coral reefs. Academic Press, New York,
475-508.
Hixon, M.A., P.D. Boersma, M.L. Hunter Jr, F. Micheli, E.A. Norse, H.P. Possingham,
P.V.R. Snelgrove, 2001. Oceans at Risks: Research Priorities in Marine
Conservation Biology. In Conservation Biology: Research Priorities for the Next
Decade, M.E. Soulé and G.H. Orians, Eds. Island Press, Covelo, CA.
Hereu B., 2004. The role of trophic interactions between fishes, sea urchins and algae in
northwestern Mediterranean rocky infralittoral. PhD Thesis, Universitat de
Barcelona.
Holbrook S.J., R.J. Schmitt, J.A. Stephens Jr, 1997. Changes in an assemblage of
temperate reef fishes associated with a climate shift. Ecol Appl 7: 1299-1310.
Holling, C.S. (a cura di), 1978. Adaptive environmental assessment and management.
John Wiley and Sons, Chichester, Ltd.
Inchausti, P., 1994. Reductionist approaches in community ecology. Am Nat 143: 201-
221.
IUNC, 1988. Proceedings of the 17h session of the General Assembly of IUCN and the
17h technical meeting. San Jose, Costa Rica, 1-10 February 1988. World
Conservation Union (IUNC), Gland, Switzerland.
Jan, R.-Q., K.-H. Chang, 1993. Substrate partitioning among non-territorial
damselfishes during spawning in Northern Taiwan. Bull Inst Zool, Academia
Sinica 32: 184-193.
Jennings S., 2004. The ecosystem approach to fishery management: a significant step
towards sustainable use of the marine environment? Marine Ecology Progress
Series, 274: 279-282.
Jones G.P., 1991. Post-recruitment processes in the ecology of coral reef fish
populations: a multifactorial perspective. In Sale P.F. (Ed.). The ecology of fishes
on coral reefs. Academic Press, New York, 294-328.
Jones G.P., N.L. Andrew, 1992. Temperate reefs and the scope of seascape ecology. In:
Battershill, C.N., D.R. Schiel, G.P. Jones, R.G. Creese, A.B. MacDiarmid (Eds.).

91
Proceedings of the Second International Temperate Reef Symposium, 7-10
January 1992, Auckland, New Zealand. NIWA Marine, Wellington, NZ, 29-45.
Jones, P.S.J., 1994. A review and and analysis for the objective of marine nature
reserves. Ocean Coastal Manage 24: 149-178.
Kelleher, G., C. Recchia, 1998. Editorial: Lessons from marine protected areas around
the world. PARKS, Internat J Protected Area Man 8: 1-4.
Knowlton, N., 2004. Multiple "stable" states and the conservation of marine
ecosystems. Progress in Oceanography 60: 387-396.
Kotliar, N.B., J.A. Wiens, 1990. Multiple scales of patchiness and patch structure: a
hierarchical framework for the study of heterogeneity. Oikos 59: 353-260.
Kikuchi, T. 1966 An ecological study on animal communities of the Zostera marina
belt in Tomioka Bay, Amakusa, Kyushu. Publications of Amakusa Marine
Biological Laboratory 1: 1-106.
Kotrschal, K., 1983. Northern Adriatic rocky reef fishes at low winter temperatures.
PSZN I: Mar Ecol 4: 275-286.
Kramer D. L., M.R. Chapman, 1999. Implication if fish home range size and relocation
for marine reserve function. Environmental Biology of Fishes 55: 65-79.
Kritzer, J.P., P.F. Sale, 2004. Metapopulation ecology in the sea: from Levin’s model
to marine ecology and fisheries science. Fish Fish 5: 131-140.
Kurlansky, M., 1999. Merluzzo: storia di un pesce che ha cambiato il mondo. Milano,
Mondadori.
Lecchini, D., C.W. Osenberg, J.S. Shima, C.M. St Mary, and R. Galzin. 2007a.
Ontogenetic changes in habitat selection during settlement in a coral reef fish:
ecological determinants and sensory mechanisms. Coral Reefs 26:423–432.
Lecchini, D., S. Planes, and R. Galzin. 2007b. The influence of habitat characteristics
and conspecifics on attraction and survival of coral reef fish juveniles. Journal of
Experimental Marine Biology and Ecology 341:85–90.
Leis J.M., M.I. McCormick, 2002. The biology, behavior and ecology of the pelagic
larval stage of coral reef fishes. In Sale P.F. (Ed.) The ecology of fishes on coral
reefs. Academic Press, New York, 171-199.
Lejeune, P., 1984. Étude Écoéthologique des comportements reproducteurs et sociaux
des labridés méditerranéens des genre Symphodus Rafinesque, 1810 et de Coris
Lacépède, 1802. Ph.D Thesis, Université de Liège.

92
Leroux, S.J., F.K.A. Schmiegelow, S.G. Cumming, R.B. Lessard & J. Nagy, 2007.
Accounting for system dynamics in reserve design. Ecol Appl 17: 1954-1966.
Letourneur Y., S. Ruitton, S. Sartoretto, 2003. Environmental and benthic factors
structuring the spatial distribution of a summer infralittoral fish assemblage in the
north-western Mediterranean Sea. J Mar Biol Assoc U.K. 83:193–204
Levin, S.A., 1992. The problem of pattern and scale in ecology. Ecology 73:1943-1967.
Levin, S.A., 1998. Ecosystems and the biosphere as complex adaptive systems.
Ecosystems 1: 431-436.
Lin, X., D. Zhang, 1999. Inference in Generalized Additive Mixed Models. J R Statistic
Soc B 61, Part 2: 381-400.
Manini E., G.M. Luna, 2003. Influence of the mineralogical composition on microbial
activities in marine sediments: an experimental approach. Chem Ecol 19: 399-
410.
Maradonna F., G. Bavestrello, M. Cardinali, I. Olivotto, C. Cerrano, M. Giovine, O.
Carnevali, 2003. Role of substrate on larval development of the freshwater teleost
Pelvicachromis pulcher. Mol Reprod Dev 66: 256-263
McCoy, E.D., S.S. Bell, 1991. Habitat structure, the evolution and diversification of a
complex topic. In: Bell, S.S., E.D. McCoy, H.R. Mushinsky (Eds.), 1991. Habitat
structure. The physical arrangement of objects in space. Chapman & Hall,
London, 3-27.
McFarland, W.N., 1986. Light in the sea – correlations with behaviors of fishes and
invertebrates. Am Zool 26: 389-401.
McGehee M.A., 1994. Correspondence between assemblages of coral reef fishes and
gradients of water motions, depth and substrate size of off Puerto Rico. Mar Ecol
Prog Ser 105: 243-255.
Micheli, F., B. Halpern, L.W. Botsford, and R.R. Warner, 2004. Trajectories and
correlates of community change in no-take marine reserves. Ecological
Applications 14: 1709–1723.
Micheli, F., L. Benedetti-Cecchi, S. Gambaccini, I. Bertocci, C. Borsini, G. C. Osio,
and F. Romano. 2005. Cascading human impacts, marine protected areas, and the
structure of Mediterranean reef assemblages. Ecological Monographs 75: 81-102.
Milne, B.T., 1991. Heterogeneity as a multiscale characteristic of landscapes. In:
Kolasa, J., S.T.A. Picket (Eds.), Ecological Heterogeneity. Springer, New York,
69-84.

93
Moilanen, A. & B.A. Wintle, 2007. The boundary-quality penalty: a quantitative
method for approximating species responses to fragmentation in reserve selection.
Conserv Biol 21: 355-364.
Mora, C., S. Andrefouet, M.J. Costello, C. Kranenburg, A. Rollo, J. Veron, K.J.
Gaston, R.A. Myers, 2006. Coral reefs and the global network of marine protected
areas. Science 312: 1750-1751.
Mosquera, I., I. M. Côté , S. Jennings, and J. Reynolds. 2000. Conservation benefits of
marine reserves for fish populations. Animal Conservation 4: 321-332.
Murawski, S.A., P. Rago & M. Fogarty, 2004. Spillover effects from temperate marine
protected areas. Am Fish Soc Symp 42: 167-184.
Norse, E.A., 1993. Global marine biological diversity. A strategy for building
conservation into decision making. Island, Washington D.C., USA.
Musick, J.A., 1999. Criteria to define extinction-risk in marine fishes. Fisheries, 24:6-
12.
Oksanen, T., L. Oksanen, M. Gyllemberg, 1992. Exploitation ecosystems in
heterogeneous habitat complexes II: impact of small-scale heterogeneity on
predator-prey dynamics. Evol Ecol 6: 383-398.
Orth, R.J. & K.L. Heck Jr, 1980. Structural components of the eelgrass (Zostera
marina) meadows in the lower Chesapeake Bay—fishes. Estuaries 3, 278–288.
Palumbi, S.R., 2001. The Ecology of marine protected areas. In: Bertness M.D., S.D.
Gaines and M.E. Hay (Eds). Marine Community Ecology. Sunderland, MA:
Sinauer Associates, 509-530.
Palumbi, S.R., 2007. Marine Reserves: a tool for ecosystem management and
conservation. Prepared for the Pew Oceans Commission, 1-52.
Pauly, D., V. Christensen, J. Dalsgaard, R. Froese, F. Torres Jr, 1998. Fishing down
marine food webs. Science 279: 860-863.
Pauly, D., V. Christensen, S. Guénette, T.J. Pitcher, U.R. Sumaila, C.J. Walters, R.
Watson, and D. Zeller. 2002. Towards sustainability in world fisheries. Nature
418: 689–695.
Peterson, C.H., J.A. Estes, 2001. Conservation and management of marine
communities. In Marine Community Ecology, M.D. Berness, S.D. Gaines, M.E.
Hay (Eds.), Sinauer Associates, Sunderland, MA.
Planes, S., A. Lefèvre, P. Legendre, R. Galzin, 1993. Spatiotemporal variability in fish
recruitment to a coral reef (Moorea, French Polynesia). Coral Reefs 12: 105-113.

94
Polis, G.A., D.R. Strong, 1996. Food web complexity and community dynamics. Am.
Nat. 147: 813-846.
Pollard, D., 1984 A overview of ecological studies on seagrass-fish communities, with
particular reference to recent studies in Australia. Aquatic Botany 18: 3-42.
Polunin, N.V.C., D.W. Klumpp, 1989. Ecological correlates of foraging periodicity in
herbivorous reef fishes of the Coral Sea. J Exp Mar Biol Ecol 126: 1-20.
Quaderni habitat, 2004. Ministero dell’Ambiente e della Tutela del Territorio e del
Mare. Museo friulano di storia naturale. Numeri: 7) Coste marine rocciose.
Capitolo “Il popolamento sommerso” (pp. 105-133) di C.N. Bianchi, F. Boero, S.
Fraschetti, C. Morri. 19) Praterie a fanerogame marine. Capitolo “La fauna: i
vertebrati” (pp. 91-111) di P. Guidetti.
Roberts, C. M., 1995. Effects of fishing on the ecosystem structure of coral reefs.
Conservation Biology, 9: 988-995.
R 2.11.1 (R software project. http://www.r-project.org/)
Roberts, C.M., J.A. Bohnsack, F. Gell, J.P. Hawkins, R. Goodridge, 2001. Effects of
marine reserves on adjacent fisheries. Science 294. 1920–1923.
Roberts, C.M., D.J. McClean, J.E.N. Veron, J.P. Hawkins, G.R. Allen, D.E. McAllister,
C.G. Mittermeier, F.W. Schueler, M. Spalding, F. Wells, C. Vynne, T.B. Warner,
2002. Marine biodiversity hotspots and conservation priorities for tropical reefs.
Science 295: 1280-1284.
Roberts, C.M., S. Andelman, G. Branch, R.H. Bustamante, J.C. Castilla, J.E. Dugan et
al., 2003. Ecological criteria for evaluating candidate sites for marine reserves.
Ecol Appl 13: S199-S214.
Rodwell, L.D., E.B. Barbier, C.M. Roberts, and T.R. McClanahan, 2003. The
importance of habitat quality for marine reserve–fishery linkages. Canadian
Journal of Fisheries and Aquatic Sciences 60: 171-181.
Rodrigues, A.S.L., et al. 2004. Effectiveness of the global protected area network in
representing species diversity. Nature 428: 640-643.
Roshier, D., V. Doerr, and E. Doerr. 2008. Animal movement in dynamic landscapes:
interaction between behavioural strategies and resource distributions. Oecologia
156: 465–477.
Rowley, R.J., 1992. Impact of marine reserves on fisheries: a report and review of the
literature. Science and Research series. Volume 51. Department of Conservation,
Wellington, New Zealand.

95
Rozas, L.P. & W.E. Odum, 1988. Occupation of submerged aquatic vegetation by
fishes: testing the roles of food and refuge. Oecologia 77: 101-106.
Ruitton S., P. Francour, C.F. Boudouresque, 2000. Relationships between algae, benthic
herbivorous invertebrates and fishes in rocky sublittoral communities of a
temperate sea (Mediterranean). Estuar Coast Shelf Sci 50: 217-230.
Russ, G.R., A.C. Alcala, 1996. Do marine reserves export adult fish biomass? Evidence
from Apo Island, central Philippines. Marine Ecology Progress Series, 132: 1-9.
Russ, G.R., A.C. Alcala, 2004. Marine Reserves: long-term protection is requie for full
recovery of predatory fish populations. Oecologia 138: 622-627.
Russ, G.R., B. Stockwell, A.C. Alcala, 2005. Inferring versus measuring rates recovery
in no-take marine reserves. Mar Ecol Prog Ser 292: 1-12.
Sala E., 1997. Fish predators and scavengers of the sea urchin Paracentrotus lividus in
protected areas of the north-western Mediterranean Sea. Mar Biol 129: 531-539.
Sala, E., C.F. Boudouresque, M.L. Harmelin-Vivien, 1998. Fishing, trophic cascades,
and the structure of algal assemblages: evaluation of an old but untested
paradigm. Oikos 82: 425–439.
Sala, E., O. Aburto-Oropeza, G. Paredes, I. Parra, J.C. Barrera & P.K. Dayton, 2002. A
general model for designing networks of marine reserves. Science 298: 1991-
1993.
Sale P.F., 1978. Coexistence of coral reef fishes – a lotteryfor living space. Environ
Biol Fish 3: 337-359.
Sale P.F., 1991. Habitat structure and recruitment in coral reef fishes. In : Bell, S.S.,
E.D. McCoy, H.R. Mushinsky (Eds.), Habitat structure : the physical arrangement
of objects in space. Chapman & Hall, London, 197-210.
Sale P.F., 1998. Appropriate spatial scales for studies on reef-fish ecology. Aust J Ecol
23: 202-208.
Sale P.F., 1999. Recruitment in space and time. Nature 397:25-27.
Sale, P.F., and J.P. Kritzer, 2003. Determining the extent and spatial scale of population
connectivity: decapods and coral reef fishes compared. Fish Res 65: 153-172.
Sale P.F., R.K. Cowen, B.S. Danilowicz, G.P. Jones, J.P. Kritzer, K.C. Lindeman, S.
Planes, N.V.C. Polunin, G.R. Russ, Y.J. Sadovy, R.S. Steneck, 2005. Critical
science gaps impede use of no-take fishery reserves. Trends in Ecology and
Evolution 20(2): 74-81.

96
Schiaparelli S., P. Guidetti, R. Cattaneo-Vietti, 2003. Can mineralogical features affect
the distribution patterns of sessile gastropods? The Vermetidae case in the
Mediterranean Sea. J Mar Biol Assoc U.K. 83: 1267–1268.
Schoener, T.W., 1986. Mechanicistic approaches to community ecology, a new
reductionism? Am Zool 26: 81-106.
Schoener, T.W., 1989. Food webs from the small to the large. Ecology 70: 1559-1589.
Schrope, M., 2001. Biologists urge US to build marine reserves. Nature 409: 971.
Shears N.T., R.C. Babcock, 2003. Continuing trophic cascade effects after 25 years of
no-take marine reserve protection. Mar Ecol Prog Ser 246: 1-16.
Schmitt, R.J., and S.J. Holbrook. 1996. Local-scale patterns of larval settlement in a
planktivorous damselfish: Do they predict recruitment? Marine and Freshwater
Research 47: 449–463.
Stelzenmüller, V., F. Maynou & P. Martín, 2007. Spatial assessment of benefits of a
coastal Mediterranean marine protected area. Biol Conserv 136 : 571-583.
Steneck, R.S. (2006). Staying connected in a turbulent world. Science 311: 480–481.
Stewart, B.D., and G.P. Jones, 2001. Associations between the abundance of
piscivorous fishes and their prey on coral reefs: implications for prey-fish
mortality. Marine Biology 138: 383-397.
Swearer, S.E., J.E. Caselle, D.W. Lea, and R.R. Warner. 1999. Larval retention and
recruitment in an island population of a coral-reef fish. Nature 402: 799-802.
Taborsky, M., D. Limberger, 1980. The activity rhythm of Blennius sanguinolentus
Pallas: an adaptation to its food source? PSZN I: Mar Ecol 1: 143-153.
Tupper, M.H., 2007. Spillover of commercially valuable reef fishes from marine
protected areas in Guam. Micronesia. Fisheries Bulletin 105: 527-537.
Turingan, R.G., P.C. Wainwright, D.A. Hensley, 1995. Interpopulation variation in prey
use and feeding biomechanics in Caribbean triggerfishes. Oecologia 102: 296-
304.
Turner, M.G., 1989. Landscape ecology : the effect of pattern and process. Ann Rev
Ecol Syst 20: 171-197.
Turner, S.J., S.F. Thrush, J.E. Hewitt, V.J. Cummings, G. Funnell, 1999. Fishing
impacts and the degradation or loss of habitat structure. Fish Manag Ecol 6: 401-
420.
Underwood, A.J., 1990. Experiments in ecology and management: their logics,
functions and interpretations. Aust J Ecol 15: 365-389.

97
Underwood, A.J., 1992. Beyond BACI: the detection of environmental impacts on
population in the real, but variable, world. J Mar Biol Ecol 161: 145-178.
Underwood, A.J., 1993. The mechanism of spatially replicated sampling programmes to
detect environmental impacts in a variable world. Austr J Ecol 18: 99-116.
Urban, D.L., R.V. O’Neill, H.H. Shugart Jr., 1987. Landscape ecology: A hierarchical
perspective can help scientists understand spatial patterns. BioScience 37: 119-
127.
Velte, F.,1998. Habitatanspruche und abundanzen von Thalassoma pavo (teleostei:
Labridae) bei Kreta, Greichenland. Zeitschrift für Fishkunde, Solingen 2: 47-58.
Verlaque M., 1987. Relations entre Paracentrotus lividus (Lamarck) et le phytobenthos
de Mediterranée occidentale. In Boudoresque CF (Ed) Colloque international sur
Paracentrotus lividus et les oursins comestibles. GIS Posidonie, Marseille, 5-36.
Vierucci, E., S. Bussotti, A. Ciccolella, P. Guidetti, 2006. La piccola pesca presso
l’Area Marina Protetta di Torre Guaceto: indicazioni per la gestione. Biol Mar
Medit 13(2): 298-299.
Wainwright, P.C., B.A. Richard, 1995. Predicting patterns of prey use from morphology
of fishes. Environ Biol Fish 44:97-113.
Walsh, W.J., 1983. Stability of a coral reef fish community following a catastrophic
storm. Coral Reefs 2:49-63.
Walters, C., D. Pauly, and V. Christensen. 1999. Ecospace: prediction of mesoscale
spatial patterns in trophic relationships of exploited ecosystems, with emphasis on
the impacts of marine protected areas. Ecosystems 2:539–554.
West, R. J. & King, R. J. 1996 Marine, brackish, and freshwater fish communities in the
vegetated and bare shallows of an Australian River. Estuaries 19, 31–41.
Wiens, J.A., 1976. Populations responses to patchy environments. Ann Rev Ecol Syst
7:81-120.
Zhang, D., X. Lin, 2003. Hypothesis testing in semiparametric additive mixed models.
Biostatistic 4(1): 57-74
Zeller, D. and G.R. Russ, 2004. Are fisheries “sustainable”? A counterpoint to Steele
and Hoagland. Fish Res 67: 241-245.
Zuur, A.F., E.N. Ieno, N.J. Walker, A.A. Saveliev, G.M. Smith, 2009. Mixed Effect
Models and extension in ecology with R. Springer.
www.islepark.it e www.wikipedia.org

98
APPENDICE A
Figura A.1 – Analisi dei residui del modello (GAMM) completo fittato sulla variabile di risposta ABBONDANZA TOTALE.
99
Figura A.2 – Analisi dei residui del modello (GAMM) completo fittato sulla variabile di risposta ABBONDANZA TAGLIE PICCOLE.
100
Figura A.3 – Analisi dei residui del modello (GAMM) completo fittato sulla variabile di risposta ABBONDANZA TAGLIE MEDIE.
101
Figura A.4 – Analisi dei residui del modello (GAMM) completo fittato sulla variabile di risposta ABBONDANZA TAGLIE GRANDI.

102
Figura A.5 – Analisi dei residui del modello (GAMM) completo fittato sulla variabile di risposta ABBONDANZA PREDATORI DI RICCI.
103
Figura A.6 – Analisi dei residui del modello (GAMM) completo fittato sulla variabile di risposta ABBONDANZA SPECIE PESCATE.

104
Figura A.7 – Analisi dei residui del modello (GAMM) completo fittato sulla variabile di risposta RICCHEZZA SPECIFICA.


















