I recettori per l’Ag sono di tre tipi - Omeroomero.farm.unipi.it/matdidFarm/74/Immuno4.pdf ·...
24
I recettori per l’Ag sono di tre tipi: • BCR, presente come proteina di membrana sui Linf. B e come molecola semplificata secreta dalle plasmacellule nei fluidi organici (BCR solubile=ANTICORPO). Il BCR lega il suo Ag (EPITOPO) direttamente in vivo e in vitro • TCR, presente come proteina di membrana sui Linf. T • Molecole MHC: presentano peptidi antigenici ai Linf. T
Transcript of I recettori per l’Ag sono di tre tipi - Omeroomero.farm.unipi.it/matdidFarm/74/Immuno4.pdf ·...

I recettori per l’Ag sono di tre tipi:
• BCR, presente come proteina di membrana sui Linf. B e come molecola semplificata secreta dalle plasmacellule nei fluidi organici (BCR solubile=ANTICORPO). Il BCR lega il suo Ag (EPITOPO) direttamente in vivo e in vitro
• TCR, presente come proteina di membrana sui Linf. T
• Molecole MHC: presentano peptidi antigenici ai Linf. T


BCR
• Il BCR contiene le proteine Iga e Igb: funzione di trasduzione del segnale del BCR. Eterodimero unito da ponti S=S
• Il linf B incontra un Ag, il BCR invia segnali interazione con altre celluleproduzione Ab e cellule memoria

• Linf B maturi prima dell’incontro con l’Ag esprimono mIgM e mIgD
• In seguito ad attivazione da parte dell’Ag avviene lo scambio isotipico (switch) e vengono espressi isotipi diversi: IgE, IgA, IgG
• Le cellule memoria possono esprimere questi isotipi


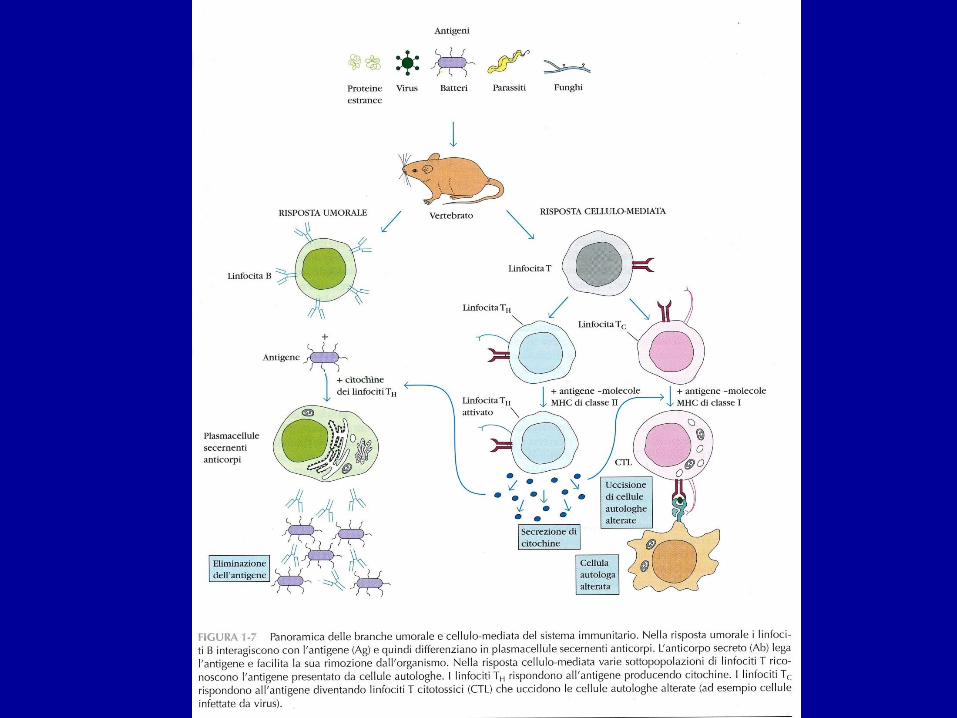
• I TCR DELLE CELLULE CD4+ RICONOSCONO SOLO SEGMENTI DI AG PROTEICI LEGATI A MOLECOLE MHC DI CLASSE II
• I TCR DELLE CELLULE CD8+ RICONOSCONO PORZIONI DI AG PROTEICI LEGATI A MOLECOLE MHC DI CLASSE I

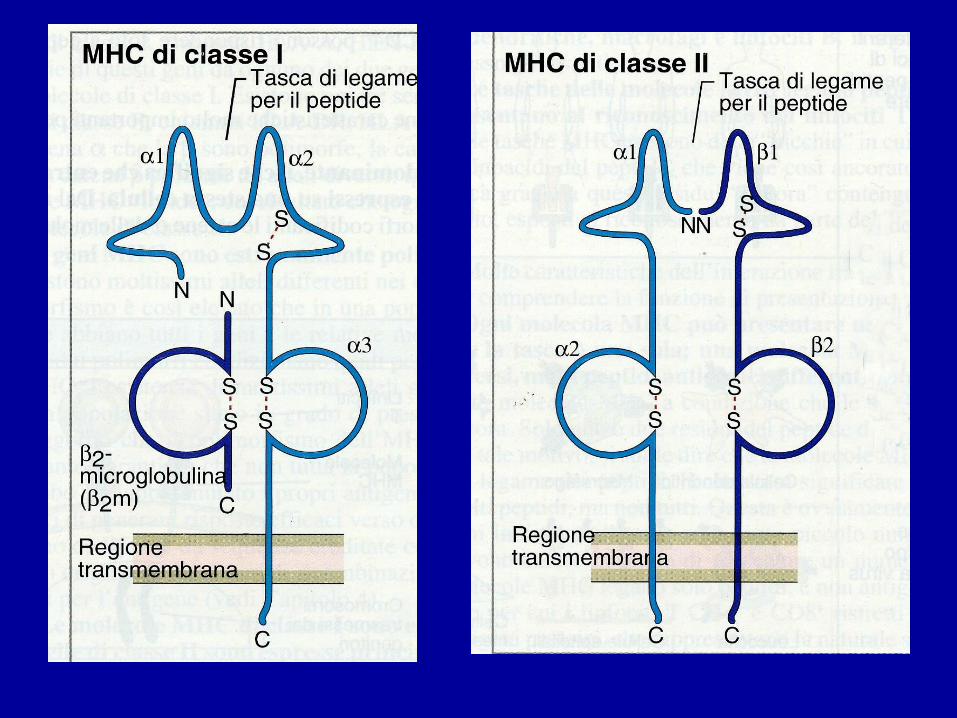
MOLECOLE MHC
• Le proteine codificate dai geni MHC (dette così per il ruolo svolto nel rigetto del trapianto) hanno struttura molecolare simile e sono:
• Di CLASSE Isu tutte le cellule nucleate
• Di CLASSE IIsu Linf B, macrofagi, cell
dendritiche
• Svolgono la funzione di presentare gli Ag peptidici ai Linf T



IL BCR RICONOSCE l’Ag COME TALE; NATURA CHIMICA e ASSOCIAZIONI MOLECOLARI NON CONTANO IL TCRab RICONOSCE l’Ag solo come PEPTIDE inserito in una MOLECOLA MHC SELF (è SELF-MHC-RISTRETTO)


COMPLEMENTO
• Costituito da circa 20 proteine presenti nel sangue o liquidi biologici in forma inattiva
• Attivato da microrganismi o altri fattori
• Meccanismo di attivazione “a cascata”

NOMENCLATURA
• Le proteine del complemento sono indicate con la lettera C seguita da un numero da 1 a 9
• Oppure con le lettere B, D, P
• I frammenti derivanti dalla scissione enzimatica sono indicati con lettere minuscole: C3b, C5a

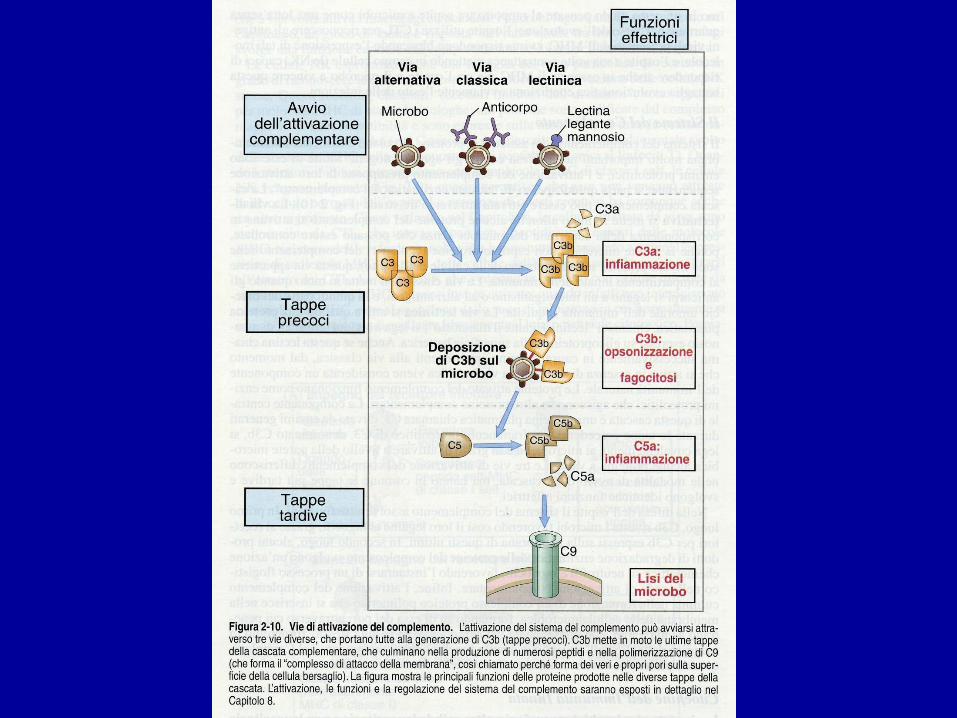
VIE DI ATTIVAZIONE
• VIA CLASSICA:
• È ATTIVATA DA INTERAZIONE TRA IMMUNOCOMPLESSO (Ag – Ab, di classe IgG o IgM) E PRIMO COMPONENTE C1q

• VIA ALTERNATIVA O PROPERDINICA:
• È ATTIVATA DA PRODOTTI DI ORIGINE BATTERICA O VIRALE CHE INTERAGISCONO CON C3 (attivazione spontanea, instabile).
• SI INNESCA IN ASSENZA DI AB, DIFESA ASPECIFICA

• VIA LECTINICA:
• Attivata dalla interazione tra la LECTINA LEGANTE IL MANNOSIO (MBL) e i residui di MANNOSIO presenti sulla superficie dei microrganismi
• MBL: famiglia delle collectine

FUNZIONI BIOLOGICHE
• LISI DI CELLULE, BATTERI, VIRUS (C5b-C9)
• OPSONIZZAZIONE (C3b, C4b)
• ATTIVAZIONE INFIAMMAZIONE attr. rilascio frammenti:
• 1)Chemiotattici: C5a (diapedesi e chemiotassi Neutrofili e Monociti)
• 2)anafilattici: C3a, C4a, C5a (degranulazione Mastociti e Basofili)
• RIMOZIONE IMMUNO COMPLESSI DALLA CIRCOLAZIONE (C3b)

• MECCANISMO TERMINALE AZIONE: COMPLESSO LITICO DI ATTACCO ALLA MEMBRANA (MAC) C5 – C9
• Il C3b lega la C3 convertasi che si trasforma in C5 convertasi
• C5 convertasi taglia C5 dando origine a C5a e C5b
• C5b funge da sito di assemblaggio del MAC


• La cascata del complemento porta all’attacco covalente del C3b sulla superficie estranea (fissazione del complemento):
• 1) la superficie estranea si trasforma in bersaglio per la fagocitosi
• 2) dà inizio alla formazione del MAC

CD4
CD8