FISIOLOGIA VEGETALE +...
20
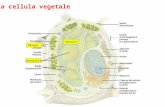
FISIOLOGIA VEGETALE + GLOSSARIO LA CELLULA VEGETALE Nella cellula vegetale si riconoscono: nucleo, reticolo endoplasmatico , golgi, mitocondri, plastidi (cloroplasti -clorofilla fotosintesi, cromoplasti-colorazione frutta/supporto scarsità luce, e leucoplasti (amiloplasti-accumulo amidi), vacuolo, micro corpi (perossisomi e gliossisomi) ed oleosomi (riserve lipidiche). Il vacuolo occupa gran parte dello spazio cellulare ha funzioni di riserva, digestione, omeostasi ionica, difesa da patogeni, contiene acqua, ioni inorganici, zuccheri, enzimi e metaboliti secondari. Cloroplasti e mitocondri sono organelli semiautonomi con proprio DNA e sintesi proteica. Microcorpi sono circondati da una singola unità di membrana come perossisomi (foto respirazione) e gliossisomi (accumulo grassi nei semi). Nelle cellule vegetali apparato del golgi è sito di sintesi delle emicellulose e pectine (componenti della parete cellulare). Parete cellulare: conferisce forza meccanica, mantiene la forma, controlla l’espansione,protegge dai patogeni, produce molecole segnale, immagazzina sostanze di riserva. La parete è costituita da (out>in): parete primaria (cellule giovani), lamella mediana e parete secondaria (cellule mature). TRASPORTO DEI SOLUTI ATTREAVERSO LA MEMBRANA: • PASSIVO (diffusione) - Alta concentrazione >> Bassa concentrazione: il movimento (spontaneo) della sostanza avviene nella direzione del suo gradiente di concentrazione. La Diffusione avviene: • le molecole non cariche si spostano secondo il loro gradiente di concentrazione , • le molecole cariche si spostano in base al gradiente elettrochimico • ATTIVO - Bassa concentrazione >> Alta concentrazione : il movimento della sostanza avviene contro il suo gradiente di concentrazione , si ha consumo di energia (es. ATP). Trasporto attivo Primario: il trasporto è accoppiato a una fonte di energia metabolica, idrolisi di ATP - reazione di ossidoriduzione, POMPE elettrogeniche H+-ATPasi (piante , funghi), Na + /K + - ATPasi (animali), elettroneutre H+/K+-ATPasi ( mucosa gastrica ) 26/01/13 Pag. 1 di20
Transcript of FISIOLOGIA VEGETALE +...

FISIOLOGIA VEGETALE + GLOSSARIO
LA CELLULA VEGETALENella cellula vegetale si riconoscono:nucleo, reticolo endoplasmatico , golgi, mitocondri, plastidi (cloroplasti -clorofilla fotosintesi, cromoplasti-colorazione frutta/supporto scarsità luce, e leucoplasti (amiloplasti-accumulo amidi), vacuolo, micro corpi (perossisomi e gliossisomi) ed oleosomi (riserve lipidiche). Il vacuolo occupa gran parte dello spazio cellulare ha funzioni di riserva, digestione, omeostasi ionica, difesa da patogeni, contiene acqua, ioni inorganici, zuccheri, enzimi e metaboliti secondari. Cloroplasti e mitocondri sono organelli semiautonomi con proprio DNA e sintesi proteica. Microcorpi sono circondati da una singola unità di membrana come perossisomi (foto respirazione) e gliossisomi (accumulo grassi nei semi). Nelle cellule vegetali apparato del golgi è sito di sintesi delle emicellulose e pectine (componenti della parete cellulare). Parete cellulare: conferisce forza meccanica, mantiene la forma, controlla l’espansione,protegge dai patogeni, produce molecole segnale, immagazzina sostanze di riserva. La parete è costituita da (out>in): parete primaria (cellule giovani), lamella mediana e parete secondaria (cellule mature).
TRASPORTO DEI SOLUTI ATTREAVERSO LA MEMBRANA:• PASSIVO (diffusione) - Alta concentrazione >> Bassa concentrazione: il movimento (spontaneo)
della sostanza avviene nella direzione del suo gradiente di concentrazione.La Diffusione avviene:• le molecole non cariche si spostano secondo il loro gradiente di concentrazione,• le molecole cariche si spostano in base al gradiente elettrochimico
• ATTIVO - Bassa concentrazione >> Alta concentrazione: il movimento della sostanza avviene contro il suo gradiente di concentrazione, si ha consumo di energia (es. ATP).Trasporto attivo Primario: il trasporto è accoppiato a una fonte di energia metabolica, idrolisi di ATP - reazione di ossidoriduzione, POMPE elettrogeniche H+-ATPasi (piante , funghi), Na+/K+ - ATPasi (animali), elettroneutre H+/K+-ATPasi ( mucosa gastrica )
26/01/13 Pag. 1 di20

Trasporto attivo secondario: la forza proton motrice può guidare il trasporto di altre sostanze contro il gradiente elettrochimico.
Il gradiente di H+ generato dalla H+-ATPasi viene utilizzato:• nel SIMPORTO (protone e molecola si muovono nella stessa direzione) per l’assorbimento di metabolici
come zuccheri, di anioni minerali, K+ quando è poco disponibile e l’influsso positivo tramite i canali non sarebbe sufficiente a mantenere l’omeostasi di questo ione, di ormoni come l’auxina. > >
• nell' ANTIPORTO (protone e molecola si muovono in direzioni opposte) per l’espulsione di Na+ dal citoplasma nelle piante tolleranti la salinità.
Trasporto dell’acqua:L’acqua si sposta dal terreno alla pianta e infine torna nell’atmosfera. La diffusione è processo tramite il quale le molecole si rimescolano a seguito della loro agitazione termica casuale.Velocità di diffusione è elevata a breve distanza ma è molto bassa a lunga distanza. Trasporto dell’acqua a lunga distanza (0,3 – 1,5 m/h) avviene tramite flusso di massa causato dalla pressione. Il potenziale idrico è potenziale chimico dell’acqua diviso il volume molale parziale dell’acqua, ovvero è la misura dell’energia libera dell’acqua per unità di volume. Potenziale del soluto o potenziale osmotico è l’effetto della presenza di soluti disciolti sul potenziale idrico. Pressione idrostatica positiva all’interno della cellula è detta pressione di turgore; pressione negativa si dice tensione. La componente gravitazionale dipende da altezza dell’acqua, densità e accelerazione di gravità. Potenziale di matrice è usato per tener conto della riduzione di energia libera dell’acqua quando esiste uno strato adsorbito sulla superficie di particelle di suolo relativamente secche.L’acqua cosi come entra allo stesso modo esce dalla cellula in risposta ad un gradiente di potenziale idrico. Conduttività idraulica esprime la facilità con la quale può spostarsi attraverso una membrana. Le acquaporine sono proteine integrali di membrana che agevolano spostamento dell’acqua attraverso le membrane cellulari . L’appassimento si verifica quando la pressione idrostatica all’interno delle cellule scende verso lo zero.La conduzione dell'acqua attraverso lo xilema avviene per:
• pressione radicale • potenziali osmotici molto negativi nelle foglie (evapotraspirazione fogliare) • flusso del floema che viene reintegrato prendendo acqua dallo xilema
Per spiegare il movimento dell'acqua nello xilema, dobbiamo pensare che l'acqua che si trova in cima agli alberi sviluppa una grande tensione (una pressione idrostatica negativa), che è in grado di aspirare l'acqua presente nello xilema. Tale meccanismo è solitamente definito la teoria della coesione-tensione della linfa ascendente, poiché richiede la presenta delle proprietà coesive dell'acqua per tollerare le grandi tensioni presenti nella colonna d'acqua dello xilema. Questa grande pressione negativa si sviluppa sulla superficie delle pareti cellulari delle foglie. La situazione è analoga a quella del suolo; all'inizio l'acqua evapora da una sottile pellicola che avvolge gli spazi aeriferi, man mano che l'acqua viene persa nell'aria, la superficie dell'acqua che rimane viene trascinata verso gli interstizi della parete cellulare, cioè le incrinature fra le cellule e fra le fibrille della parete.
L’acqua nel suolo:Capacità di campo dei suoli è capacità di trattenere l’umidità. La pressione idrostatica negativa dell’acqua del suolo ne abbassa il potenziale idrico. L’acqua si muove attraverso il suolo mediante il flusso di massa. Conduttività idraulica del suolo è la misura della facilità con la quale l’acqua si muove attraverso il suolo. Se diminuisce il contenuto idrico allora diminuisce anche la conduttività idraulica. Se un suolo è molto secco il potenziale idrico può cadere sotto il punto permanente di appassimento,e la pianta non può ripristinare la pressione di turgore. Radice assorbe acqua grazie ai suoi peli radicali. L’acqua si sposta nella radice tramite apoplasto, simplasto e la via trans membrana:• Apoplasto → sistema continuo di pareti cellulari e di spazi aeriferi intercellulari dei tessuti. Nella via
apoplastica l’acqua può muoversi attraverso le pareti cellulari e qualsiasi spazio extracellulare ripieno d’acqua (senza attraversare la membrana)
• Simplasto → continuità di tutto l’intreccio dei citoplasmi cellulari collegati dai plasmodesmi . L’acqua si muove attraverso il parenchima corticale (senza attraversare la membrana).
• Via trans membrana → Acqua entra da una parte di una cellula e ne esce dall’altra. L’acqua attraversa in ogni cellula almeno 2 membrane.
26/01/13 Pag. 2 di20

La banda di Casparay è formata da strisce di parete cellulare radiale dell’endodermide impregnate di suberina (sostanza idrofoba). Assorbimento dell’acqua diminuisce quando radici sono sottoposte a basse T° o condizioni anaerobiche.L’acqua dalla foglia all’atmosfera:Nel suo cammino dalla foglia all’atmosfera l’acqua è aspirata dallo xilema alle pareti cellulari del mesofillo dove evapora negli spazi cellulari fogliari. Tutta l’acqua persa dalle foglie esce per diffusione sotto forma di vapor d’acqua dai pori degli stomi. Chiusura o apertura degli stomi è regolata da cellule epidermiche specializzate(cellule di guardia). Apertura degli stomi è causata da un aumento della pressione di turgore nelle cellule di guardia.
LA FOTOSINTESIFotosintesi è processo di trasduzione dell’energia luminosa in energia chimica.Cloroplasto è sede della fotosintesi. Ci sono 4 tipi di clorofilla: clorofilla A(in tutti organismi con fotosintesi ossigenica), clorofilla B(negli organismi fotosintetici verdi), clorofilla C (pigmenti accessori nelle alghe), batterioclorofille (nei batteri con fotosintesi anossigenica). I carotenoidi fanno parte del complesso antenna e assicurano assorbimento della luce in ambiti spettrali non coperti dalle clorofilla, sono importanti anche nella protezione dai danni foto ossidativi. Energia dello stato di eccitazione che arriva al centro di reazione è < di quella alla periferia del complesso antenna. Fotosistemi sono pigmenti foto sintetici + proteine. Centro di reazione è sito del fotosistema dove si trova il pigmento foto ossidabile.Il tessuto fotosintetico delle piante superiore è mesofillo fogliare ricco di clorofille specializzate nell’assorbire la luce. La fotosintesi comprende le reazioni dei tilacoidi (granali e stomatici) e la fissazione del Carbonio . Clorofille A e B sono più diffuse nelle piante verdi mentre clorofille C e D sono diffuse nei protisti e cianobatteri. I carotenoidi sono pigmenti accessori che trasferiscono la luce assorbita alle clorofille. La fotosintesi avviene in complessi antenna che raccolgono la luce e trasferiscono energia al complesso del centro di reazione. Il processo generale della fotosintesi è la reazione redox nella quale gli elettroni sono rimossi da una specie chimica ossidandola ed aggiunti ad un’altra specie chimica riducendola. Reazioni chimiche nelle quali l’acqua si ossida a ossigeno, il NADP viene ridotto e l’ATP viene formato (reazioni tilacoidali). Reazioni di fissazione e riduzione di C sono reazioni stromatiche. Gli organismi che evolvono ossigeno possiedono 2 foto sistemi che lavorano in serie:• Foto sistema II (680 – rosso basso) → stimolato poco dalla luce nel rosso lontano, produce forte agente
ossidante (in grado di ossidare l’acqua) e un debole agente riducente rispetto a quello prodotto dal foto sistema I.
• Foto sistema I (700 – rosso + alto) → assorbe luce nel rosso lontano, produce forte agente riducente capace di ridurre NADP e un debole agente ossidante
I due foto sistemi sono legati da una catena di trasporto di elettroni; essi sono separati spazialmente: foto sistema I si trova solo nelle membrane stromatiche non impilate mentre il foto sistema II è localizzato nelle membrane impilate . Foto sistemi I e II ossidano l’acqua ad ossigeno molecolare e riducono NADP a NADPH. Fotosintesi è unico sistema biochimico capace di ossidare l’acqua. Fotoossidazione dell’acqua è rappresentata da un modello a 5 tappe definito meccanismo dello stato Z. Il flusso di elettroni durante la fotosintesi è accoppiato alla traslocazione di protoni attraverso la membrana tilacoidale, in questo evento lo stroma diventa + alcalino. E’ proprio il forte gradiente protonico a generare sintesi di ATP. NADPH e l’ATP formati forniscono l’energia per la riduzione del C.
Reazione alla luce → Fotosistema 1 è localizzato nelle lamelle stromatiche,foto sistema 2 è localizzato nelle lamelle dei grana.Il citocromo B6-F unisce i due foto sistemi. Proteine del “core”(PSI) legano il centro di reazione P700 ed i suoi accettori. Le proteine Redox che interagiscono con PSI sono plastocianina, ferredossina, ferredossina-NADP riduttasi. Il trasporto ciclico degli elettroni fornisce ATP, si attiva quando NADPH è in eccesso.CICLO DI CALVIN (Reazione del C): E’ ciclo riduttivo dei pentosi fosfati.Regolazione del ciclo di calvin avviene tramite 5 enzimi regolati alla luce tra cui rubisco. Il ciclo C2 recupera il 75% del carbonio perso originariamente dal ciclo di Calvin. IL ciclo di Calvin consuma NADPH e ATP sintetizzati durante le reazioni luminose nelle membrane tilacoidali.Tutti gli eucarioti fotosintetici riducono anidride carbonica con ciclo di Calvin che avviene nello stroma o fase solubile del cloroplasto. Ciclo di Calvin consuma 2 molecole NADPH e 3 molecole ATP per ogni molecola di anidride carbonica fissata, con efficienza termodinamica del 90%. I carboidrati sintetizzati per opera del ciclo di Calvin sono convertiti in saccarosio e amido. Saccarosio è
26/01/13 Pag. 3 di20

sintetizzato nel citosol e la sua sintesi è regolata dalla fosforilazione della saccarosio fosfato sintasi. Amido è sintetizzato nel cloroplasto.
Il ciclo C2 per l'ossidazione fotosintetica del carbonioUna proprietà caratteristica della rubisco è la sua capacità di catalizzare sia la carbossilazione che l'ossigenazione del ribulosio-1,5-difosfato. Questa ossigenazione dà inizio a una serie di eventi fisiologici in cui l'assorbimento dell'ossigeno dipendente dalla luce è associato allo sviluppo di CO2 in foglie fotosinteticamente attive. Questo processo, conosciuto come fotorespirazione, funziona in direzione diametralmente opposta alla fotosintesi, causando la perdita della CO2 fissata dal ciclo di Calvin. In dettaglio, l'incorporazione di una molecola di O2 nell'isomero 2,3-enediolo del ribulosio-1,5-difosfato genera un intermedio instabile che si divide rapidamente in 2-fosfoglicolato e 3-fosfoglicerato. La CO2 e l'O2 competono nella reazione con il ribulosio-1,5-difosfato, poiché la carbossilazione e l'ossigenazione avvengono all'interno dello stesso sito attivo. Il ciclo ossidativo fotosintetico C2 agisce come un'operazione di salvataggio per recuperare il carbonio perso durante la fotorespirazione dovuta alla reazione della rubisco ossigenasi. L'acido 2-fosfoglicolico, che si forma nel cloroplasto a seguito dell'ossigenazione del ribulosio-1,5-difosfato, viene rapidamente idrolizzato a glicolato attraverso l'azione di una fosfatasi specifica del cloroplasto. A questo punto, l'ulteriore metabolismo del glicolato implica la cooperazione di altri due organelli, i perossisomi e i mitocondri. Il glicolato esce dal cloroplasto tramite una proteina trasportatrice e diffonde in un perossisoma. Qui la glicolato ossidasi catalizza l'ossidazione del glicolato utilizzando O2 e producendo acido gliossilico e acqua ossigenata. Quest'ultima è demolita nel perossisoma per opera della catalasi, liberando O2, mentre l'acido gliossilico viene transamminato con il glutammato a formare l'amminoacido glicina. La glicina esce dal perossisoma ed entra nel mitocondrio, dove due enzimi, il complesso della glicina decarbossilasi (GDC) e la serina idrossimetiltrasferasi, catalizzano in sequenza la conversione di due molecole di glicina e di una di NAD+ in una di serina una di NADH, una di NH4+ e una di CO2. La serina neoformata diffonde dal mitocondrio verso il perossisoma dove p convertita in idrossipiruvato per mezzo di una transamminazione e a sua volta è trasformato in glicerato tramite una reduttasi NADH-dipendente. Infine, il glicerato ritorna nel cloroplasto dove è fosforilato per dare 3-fosfoglicerato. Parallelamente a queste reazioni l'NH4+ rilasciato dall'ossidazione della glicina diffonde rapidamente dalla matrice del mitocondrio verso i cloroplasti, dove la glutammina sintetasi catalizza l'incorporazione nel glutammato, formando glutammina. Quindi la glutammato sintasi ferredossina-dipendente catalizza la reazione della glutammina con il 2-oxoglutarato, che porta alla produzione di due molecole di glutammato.
26/01/13 Pag. 4 di20

In generale, comunque, durante la fotorespirazione gli atomi di carbonio, azoto e ossigeno, circolano simultaneamente attraverso tre cicli:
1. Nel primo ciclo il carbonio esce dal cloroplasto sottoforma di due molecole di glicolato e fa ritorno sottoforma di una molecola di glicerato, perdendo una molecola di CO2 nel mitocondrio;
2. Nel secondo ciclo l'azoto esce dal cloroplasto come molecola di glutammato e vi fa ritorno come molecola di NH4+, poi associata ad una molecola di 2-oxoglutarato. Di conseguenza l'azoto totale rimane invariato in quanto la formazione di azoto inorganico (NH4+) nel mitocondrio è bilanciata dalla sintesi di glutammina nel cloroplasto;
3. Nel terzo ciclo , quello legato all'ossigeno, le reazioni catalizzate dalla rubisco nel cloroplasto e dalla glicolato ossidasi nel perossisoma incorporano O2, dando un carattere ossidativo all'interno processo.
In generale, quindi, due molecole di 2-fosfoglicerato (quattro atomi di carbonio), che vengono persi dal ciclo di Calvin attraverso l'ossigenazione del ribulosio-1,5-difosfato vengono convertite in una molecola di 3-fosfoglicerato (tre atomi di carbonio) e una molecola di CO2. Il bilancio fra il ciclo di Calvin e il ciclo ossidativo fotosintetico C2 è principalmente determinato da tre fattori: uno relativo alla pianta (le proprietà cinetiche della Rubisco si modificano al variare della temperatura) e due legati all'ambiente (la temperatura e le concentrazioni di substrati CO2 e O2).
L'aumento della temperatura sposta progressivamente il bilancio lontano dal ciclo di Calvin e verso il ciclo ossidativo fotosintetico C2 .
La fotorespirazione, invece, risulta essere favorita da alte concentrazioni di O2 (l'aumento della temperatura aumenta il tasso di ossigenazione rispetto alla carbossilazione ) e basse di CO2, mentre è sfavorita da basse concentrazioni di O2 (calo della temperatura) e alte di CO2.
A umento della temperatura esterna >> modifica le costanti cinetiche della rubisco >> aumenta il tasso di ossigenazione rispetto alla carbossilazione >> si abbassa la concentrazione di CO2 in una soluzione con
l'aria più di quanto non avvenga all'O2).
L'aumento di temperatura diminuisce le proprietà di assorbimento della luce dei pigmenti fotosintetici
ECOLOGIA DELLA FOTOSINTESIL’anatomia fogliare è specializzata nell’assorbimento della luce,anche i movimenti dei cloroplasti all’interno delle cellule e l’inseguimento della luce da parte del lembo fogliare aiutano ad assorbire il max di luce. In zone temperate le rese quantiche di piante C3 (stomi aperti anche di giorno) sono superiori a quelle C4 (adattate a zone aride). Ad alti valori di flusso fotonico la fotosintesi è limitata dall’anidride carbonica ma questa limitazione è più bassa nelle piante C4 e CAM a causa dei loro meccanismi di concentrazione dell’anidride carbonica. La modulazione delle aperture stomatiche fornisce alla pianta un modo di controllare la perdita dell’acqua ed acquisto di anidride carbonica. A causa del ruolo che svolge la foto respirazione, la resa quantica è fortemente dipendente dalla T° nelle C3 ma è indipendente dalla T° in quelle C4.
METABOLISMO C4 e CAM il ciclo C4 ha 3 varianti:enzima malico(dipendente dal NADP),-enzima malico(dipendente dal NAD), fosfo-enolpiruvato-carbossi-chinasi(tipico del miglio). Piante C4 avendo bassa foto respirazione hanno una assimilazione positiva anche ad alte T°; la loro richiesta energetica è superiore a quelle C3. Meccanismo CAM è tipico delle piante desertiche.
Sintesi amido saccarosio →saccarosio è traslocato nel floema in tutta la pianta. Amido è carboidrato stabile ed insolubile di riserva in tutte le piante. Sia amido che il saccarosio sono sintetizzati dal trioso fosfato generato dal ciclo di calvin.
IL FLOEMA → Floema è un tessuto in grado di traslocare i prodotti della fotosintesi da foglie adulte ad aree di accrescimento ed accumulo. Floema è costituito da elementi dei tubi cribrosi nelle angiosperme , ricchi di proteina P e di plasmodesmi che collegano elementi del cribro alle cellule compagne. Nelle gimnosperme ci sono solo cellule cribrose. Nelle angiosperme, a differenza delle gimnosperme, ci sono placchi cribrosi. Le
26/01/13 Pag. 5 di20

cellule compagne sostengono metabolicamente gli elementi del cribro, sono anche coinvolte nel caricamento dei fotosintati. Velocità trasporto floematico varia da 0,3 a 1,5 m/h . Traslocazione avviene da aree di rifornimento (sorgenti) ad area di accumulo(pozzi). Le sorgenti (foglie mature) producono più fotosintati di quanti ne necessitano; i pozzi non producono fotosintati quindi devono importarli (radici, frutti in maturazione). Movimento del contenuto floematico è bidirezionale. Nelle sorgenti l’acqua si muove dallo xilema al floema con aumento della pressione di turgore. Nei pozzi l’acqua si muove dal floema allo xilema con diminuzione della pressione di turgore. Il caricamento di saccarosio nelle cellule compagne e negli elementi del cribro avviene lungo un gradiente di concentrazione in salita. Le giovani foglie funzionano da pozzi,quando raggiungono il 25% di espansione della foglia matura cominciano ad esportare saccarosio.L’apice della foglia inizia ad esportare mentre la parte basale continua ad importare. Allocazione è regolazione della quantità di carbonio fissato che viene incanalata tra le varie vie metaboliche, comprende la quantità che verrà metabolizzata,accumulata e trasportata. Ripartizione consiste nella distribuzione differenziata dai foto assimilati all’interno dell’intera pianta. Fattori che influenzano la ripartizione sono pressione di turgore e messaggi chimici. Scaricamento simplastico avviene (plasmodesmi) dove concentrazione degli zuccheri è bassa (tra foglie). Scaricamento apoplastico avviene in altri tessuti pozzi isolati simplasticamente (foglie radici).
Trasporto potenziale idrico: Potenziale idrico è l’energia per unità di volume necessaria per trasportare acqua reversibilmente e isotermicamente. Potenziale idrico serve per definire la direzione del flusso idrico e per valutare lo stato idrico della pianta. Acqua si muove sempre verso regioni a potenziale idrico più basso.
• Potenziale idrico dipende da concentrazione, pressione e gravità. • Pressione osmotica è la pressione che deve essere applicata ad una soluzione per controbilanciare la
diffusione dell’acqua verso soluzione + concentrata. Pressione osmotica negativa indica che i soluti disciolti riducono il potenziale idrico di una soluzione. Il potenziale osmotico è l’effetto sul potenziale idrico dei soluti disciolti nella soluzione.
• Potenziale di pressione si misura come differenza con la pressione atmosferica (è negativo nello xilema quando è attiva la traspirazione si dice tensione).
• Pressione di turgore è importante per distendere le pareti nella crescita e per aumentare la rigidità meccanica dei tessuti non lignificati.
• Potenziale di matrice è riferito a forze di adsorbimento ed è negativo o al max uguale a zero. Potenziale di matrice è una misura della tendenza della matrice ad assorbire altra acqua. Nel caso di cellule con pareti idratate il potenziale di matrice non si considera. Il potenziale idrico di una cellula è uguale a quello del liquido o vapore con cui la cellula è in equilibrio. Il movimento spontaneo dell’acqua può avvenire per:• diffusione molecolare (gradiente di concentrazione)• flusso di massa (gradiente di pressione)• osmosi (gradiente di potenziale).
TRASPORTO – TRASPIRAZIONE: L’acqua nelle piante si muove sempre verso potenziali idrici più negativi. La traspirazione è principale causa di risalita dell’acqua nella pianta,è evaporazione attraverso la superficie della pianta. Solo il 5% della traspirazione avviene con la cuticola,il resto avviene a livello degli stomi(foglie).La forza motrice della traspirazione è il gradiente di concentrazione del vapore acqueo. Fattori che regolano la perdita di acqua dalla foglia è gradiente di vapore acqueo e le resistenze alla diffusione del vapore. La traspirazione causa raffreddamento delle foglie perché evaporazione di acqua è associata alla perdita di calore. La resistenza stomatica è regolata da apertura e chiusura degli stomi. Forza motrice per la perdita d’acqua dalla foglia è gradiente di concentrazione assoluta (dipendente dalla T°). Perchè si abbia traspirazione il potenziale idrico dell'aria deve essere più basso di quello fogliare. (la perdita acqua sotto forma di vapore da parte di un tessuto biologico a contatto con l'ambiente esterno si ha prevalentemente a livello delle foglie ed è regolata al 90%, laddove sono presenti, dagli stomi. L'acqua si muove secondo gradienti di potenziale idrico - acqua libera potenziale = 0; nella pianta e nel terreno il potenziale è negativo, cioè è necessario esercitare un LAVORO per estrarre acqua libera), da potenziale idrico maggiore (dentro la foglia) a potenziale idrico minore (nell'atmosfera basta che l'umidità relativa sia poco sotto il 100% perché il potenziale idrico sia molto inferiore a quello fogliare).
26/01/13 Pag. 6 di20

TRASPOSTO FUSTO-RADICI-SUOLO: Forze di coesione/adesione delle molecole d’acqua consentono la trasmissione della tensione e la formazione di colonne d’acqua intatte che determinano risalita dell’acqua nello xilema. La tensione può trasmettere fino allo xilema delle piccole nervature. La coesione consente all’acqua di resistere ad una grande forza di tensione(-30 Mpa). La lignificazione delle pareti cellulari dello xilema consente la trasmissione della tensione.Tramite la guttazione alcune piante trasudano l’eccesso di linfa grezza durante la notte tramite gli idatodi. Il potenziale di matrice del suolo diminuisce durante la disidratazione. Infatti, quando il suolo si disidrata, l'acqua si ritrae negli interstizi tra le particelle del suolo, formando superfici aria-acqua la cui curvatura rappresenta l'equilibrio tra la tendenza a minimizzare la superficie dell'interfaccia aria-acqua e l'attrazione delle molecole d'acqua per le particelle di suolo. Il potenziale di matrice (τ) è una misura della tendenza del terreno ad assorbire ulteriori molecole d’acqua - La linfa xilematica è molto diluita, quindi il suo Ψs sarà solo poco negativo, ma essa si trova praticamente sempre in uno stato di tensione (P negativo), quindi il Ψ sarà più negativo nello xilema (-10) che nell’acqua circolante nel terreno (-5). Di conseguenza, l’acqua passerà dal terreno alla pianta. Affinché le piante possano sopravvivere deve esserci sempre questo gradiente di Ψ tra il terreno e le radici, anche quando il contenuto idrico nel terreno raggiunge valori molto bassi (es. valori compresi tra l’8 e il 5 % per siccità). Quindi se le soluzioni circolanti nel terreno si fanno troppo concentrate, c’è il rischio che venga richiamata acqua dalle radici verso il terreno, determinando la morte della pianta. Per ovviare a questo inconveniente, la natura ha selezionato delle piante (euxerofite) in grado di sintetizzare nelle proprie cellule radicali dei soluti che aumentano il Ψs del citoplasma. All’estremo opposto, tuttavia, la natura ha selezionato un’altra tipologia di piante, che sono in grado di sopravvivere quando l’acqua circolante nel terreno è estremamente diluita a causa di precipitazioni intense o per ricambi eccessivi. Tali piante vengono definite glicofite o anche calcifughe e hanno sopraffatto i competitori adattandosi a vivere in suoli paludosi acidi. L’elevata piovosità nell’ambiente in cui vivono determina in modo costante la lisciviazione degli ioni a Ca2+,
Mg2+, K+ ad una velocità molto più rapida della loro ricostituzione attraverso l’alterazione della frazione minerale, mentre gli ioni H+ e Al3+ diventano predominanti. La scarsità di calcio nei suoli acidi può anche limitare la crescita delle piante semplicemente perché in assenza di calcio, l’H+ è molto più tossico per le radici. Inoltre, la concentrazione relativamente alta di Al3+ presente in molti suoli acidi può inibire la crescita di alcune specie.)
L'ouabaina è un inibitore specifico della Pompa di Scambio Sodio / Potassio: questa molecola agisce dal lato esterno della membrana cellulare ed arresta il ciclo di trasporto nelle sue fasi terminali perché, competendo con il sito specifico per il Potassio, ne impedisce il legame. Di conseguenza viene a mancare la defosforilazione Potassio-dipendente della molecola trasportatrice, che dovrebbe restituirla allo stato iniziale del ciclo di trasporto, e ciò blocca sia l’attività di trasporto che quella enzimatica della pompa. Poiché l’affinità per il glucoside è molto elevata, esso può bloccare il funzionamento della pompa anche a concentrazioni molto basse.
26/01/13 Pag. 7 di20

GLOSSARIOACQUA NEL SUOLO Capacità di campo dei suoli è capacità di trattenere l’umidità. La pressione
idrostatica negativa dell’acqua del suolo ne abbassa il potenziale idrico. L’acqua si muove attraverso il suolo mediante il flusso di massa. Conduttività idraulica del suolo è la misura della facilità con la quale l’acqua si muove attraverso il suolo. Se cala il contenuto idrico allora scende anche la conduttività idraulica. Se un suolo è molto secco il potenziale idrico può cadere sotto il punto permanente di appassimento e la pianta non può ripristinare la pressione di turgore. Radice assorbe acqua grazie ai suoi peli radicali. L’acqua si sposta nella radice tramite apoplasto, simplasto e la via trans membrana:
ACQUAPORINE Proteine integrali di membrana che agevolano spostamento dell’acqua attraverso le membrane cellulari.
ANTIPORTO ANTIPORTO: protone e molecola si muovono in direzioni opposte (una entra e l'altra esce). Es. - nell' ANTIPORTO per l’espulsione di Na+ dal citoplasma nelle piante tolleranti la salinità. Trasferimento zuccheri in tessuti di riserva ad alta concentrazione (contro gradiente concentrazione). Trasporto attivo secondario.
APOPLASTO Nel'apoplasto la parete cellulare è permeabile all'acqua che può agevolmente fluire attraverso le pareti cellulari senza che debba necessariamente attraversare membrane ed entrare quindi nel protoplasto.
BANDA DI CASPARAY La banda di Casparay è formata da strisce di parete cellulare radiale dell’endodermide impregnate di suberina (sostanza idrofoba). L'assorbimento dell’acqua diminuisce quando radici sono sottoposte a basse T° o condizioni anaerobiche.
CARICAMENTO SACCAROSIO NEL FLOEMA (ES. BARBAB.)
Saccarosio nel floema avviene per trasporto attivo secondario: i tubi cribrosi assorbono saccarosio dall'apoplasto contro gradiente di concentrazione. Simporto H+ - saccarosio.
CAROTERNOIDI I carotenoidi sono pigmenti accessori che trasferiscono la luce assorbita alle clorofille in ambiti spettrali non coperti dalle clorofilla. Sono importanti anche nella protezione dai danni foto ossidativi.
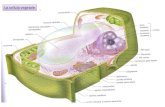
CELLULA VEGETALE I°
26/01/13 Pag. 8 di20

Nella cellula vegetale si riconoscono:
Nucleo, Reticolo endoplasmatico, Golgi: è sito di sintesi delle emicellulose e pectine componenti della parete cellulare. mitocondri:(coinvolti nel rilascio di CO2 e NH3), organelli semiautonomi con proprio dna e sintesi proteicaVacuolo: occupa gran parte dello spazio cellulare ha funzioni di riserva, digestione, omeostasi ionica, difesa da patogeni. Vacuolo contiene acqua, ioni inorganici, zuccheri, enzimi e metaboliti secondari.Plastidi: sono cloroplasti (contengono clorofilla ed operano fotosintesi – ciclo Calvin, RuBisco), cromoplasti (contengono carotenoidi che determinano colore dei frutti) e leucoplasti (amiloplasti - sono sede di accumulo di amido). Sono organelli semiautonomi con proprio dna e sintesi proteica. Microcorpi: circondati da una singola unità di membrana - perossisomi (foto respirazione) e gliossisomi (accumulo grassi nei semi). Parete cellulare: funzioni meccaniche, mantiene la forma, controlla l’espansione, protegge dai patogeni, produce molecole segnale, immagazzina sostanze di riserva; è costituita da: parete primaria > lamella mediana > parete secondaria (cellule adulte) > menbreana cellulare > Citoplasma Plasmodesmi canali x lo scambio acqua e nutrienti tra cellule.
CICLO CALVYN – RUBISCOalta temperatura = ciclo ossidativoalta temperatura =/// ciclo Calvinconcentrazioni O2 =fotorespirazioneconcentrazioni CO2 =ciclo calvin
E’ ciclo riduttivo dei pentosi fosfati. La regolazione del ciclo di Calvin avviene tramite 5 enzimi regolati alla luce tra cui Rubisco. Il ciclo C2 recupera il 75% del carbonio perso originariamente dal ciclo di Calvin. Consuma ATP e NADPH sintetizzati durante le reazioni luminose nelle membrane tilacoidali. Avviene nello stroma o fase solubile del cloroplasto e consuma 3 molecole ATP e 2 molecole NADPH per ogni molecola di CO2 fissata , con efficienza termodinamica del 90%. I carboidrati sintetizzati per opera del ciclo di Calvin sono convertiti in saccarosio e amido (sintetizzato nel citosol, la sua sintesi è regolata dalla fosforilazione della saccarosio fosfato sintasi). Amido è sintetizzato nel cloroplasto.
* RuBisCo interviene nella carbossilazione (ciclo calvin fotosintetico) * RuBisCo interviene ossidazione (fotorespirazione) con perdita della CO2 fissata dal ciclo di Calvin.
26/01/13 Pag. 9 di20

La CO2 e l'O2 competono nella reazione con il ribulosio-1,5-difosfato, poiché la carbossilazione e l'ossigenazione avvengono all'interno dello stesso sito attivo. Il ciclo ossidativo fotosintetico C2 agisce come un'operazione di salvataggio per recuperare il carbonio perso durante la fotorespirazione dovuta alla reazione della rubisco ossigenasi. La suddetta complessa attività metabolica avviene per il tramite di ulteriori due organelli i perossisomi, coinvolti nella fotorespirazione, ed i mitocondri, coinvolti nel rilascio di CO2 ed NH3. In generale, durante la fotorespirazione gli atomi di carbonio, azoto e ossigeno, circolano simultaneamente attraverso tre cicli:
• Nel primo ciclo il carbonio esce dal cloroplasto sottoforma di due molecole di glicolato e fa ritorno sottoforma di una molecola di glicerato, perdendo una molecola di CO2 nel mitocondrio;
• Nel secondo ciclo l'azoto esce dal cloroplasto come molecola di glutammato e vi fa ritorno come molecola di NH4+, poi associata ad una molecola di 2-oxoglutarato. Di conseguenza l'azoto totale rimane invariato in quanto la formazione di azoto inorganico (NH4+) nel mitocondrio è bilanciata dalla sintesi di glutammina nel cloroplasto;
• Nel terzo ciclo, quello legato all'ossigeno, le reazioni catalizzate dalla rubisco nel cloroplasto e dalla glicolato ossidasi nel perossisoma incorporano O2, dando un carattere ossidativo all'interno processo.
In sintesi: due molecole di 2-fosfoglicerato (4*C), che vengono persi dal ciclo di Calvin attraverso l'ossigenazione del ribulosio-1,5-difosfato vengono convertite in una molecola di 3-fosfoglicerato (tre atomi di carbonio) e una molecola di CO2. I l bilancio fra il ciclo di Calvin e il ciclo ossidativo fotosintetico C2 è principalmente determinato da tre fattori: >> uno relativo alla pianta (le proprietà cinetiche della rubisco)>> due legati all'ambiente (la temperatura e le concentrazioni di substrati CO2 e O2). L'aumento della temperatura esterna (modifica le costanti cinetiche della rubisco), aumenta il tasso di ossigenazione >> ciclo ossidativo C2, con conseguente riduzione del di carbossilazione (Ciclo di Calvin). La fotorespirazione, risulta essere favorita da alte concentrazioni di O2 e basse di CO2
26/01/13 Pag. 10 di20

FLOEMA E' un tessuto in grado di traslocare i prodotti della fotosintesi da foglie adulte ad aree di accrescimento ed accumulo. E' è costituito da elementi dei tubi cribrosi nelle angiosperme (cellule più specializzate impilate longitudinalmente caratterizzate dalla presenza di placche cribrose, pori più grandi disposti solitamente nelle regioni terminali, hanno vita di qualche anno e il nucleo è completamente riassorbito), sono ricchi di proteina P e di plasmodesmi che collegano elementi del cribro alle cellule compagne. Nelle angiosperme, placchi cribrosi, cellule compagne sostengono metabolicamente gli elementi del cribro, sono anche coinvolte nel caricamento dei fotosintati. Nelle gimnosperme troviamo le cellule cribrose nelle, meno evolute e specializzate, allungate e affusolate, i pori sono molto piccoli ed uniformemente distribuiti, sono di forma allungata con pareti celluloso pectiche, in genere sottili. Velocità trasporto floematico varia da 0,3 a 1,5 m/h. Traslocazione avviene da aree di rifornimento (sorgenti) ad area di accumulo (pozzi). Le sorgenti (foglie mature) producono più fotosintati di quanti ne necessitano; i pozzi non producono fotosintati quindi devono importarli (radici, frutti in maturazione). Movimento del contenuto floematico è bidirezionale. Nelle sorgenti l’acqua si muove dallo xilema al floema con aumento della pressione di turgore. Nei pozzi l’acqua si muove dal floema allo xilema con diminuzione della pressione di turgore. Il caricamento di saccarosio nelle cellule compagne e negli elementi del cribro avviene lungo un gradiente di concentrazione in salita. Le giovani foglie funzionano da pozzi,quando raggiungono il 25% di espansione della foglia matura cominciano ad esportare saccarosio. L’apice della foglia inizia ad esportare mentre la parte basale continua ad importare. Allocazione è regolazione della quantità di carbonio fissato che viene incanalata tra le varie vie metaboliche, comprende la quantità che verrà metabolizzata, accumulata e trasportata. Ripartizione consiste nella distribuzione differenziata dai foto assimilati all’interno dell’intera pianta. Fattori che influenzano la ripartizione sono pressione di turgore e messaggi chimici. Scaricamento simplastico avviene dove concentrazione degli zuccheri è bassa (fogie giovani). Scaricamento apoplastico avviene in altri tessuti pozzi isolati simplasticamente (tuberi, semi).
FOTORESPIRAZIONE
alta temperatura = ciclo ossidativoalta temperatura =/// ciclo Calvinconcentrazioni O2 =fotorespirazioneconcentrazioni CO2 =ciclo calvin
In presenza di una grande quantità di anidride carbonica, il ciclo di Calvin svolge in genere regolarmente; se di contro, nella foglia, la concentrazione CO2
è bassa, se paragonata a quella dell'ossigeno, l'enzima RuDP-carbossilasi catalizza una reazione tra RuDP e ossigeno invece che tra RuDP e anidride carbonica. Questa reazione è la prima tappa di un processo conosciuto come fotorespirazione, in cui i carboidrati vengono ossidati in presenza di luce con formazione di acqua e dell' anidride carbonica; in altre parole, la cellula si autoproduce la CO2 di cui necessita. E' un processo metabolico respirativo che le piante con ciclo C3 attuano alla luce, e continuano per un breve periodo anche al buio, per eliminare l'ossigeno in eccesso. Le alte pressioni di ossigeno atmosferico provocano uno stop della fotosintesi, al fine di prevenire la formazione di radicali liberi, dannosissimi alle cellule; via via che queste pressioni diminuiscono, in favore della pressione cellulare di anidride carbonica, il processo fotosintetico aumenta la sua attività.
FOTOSINTESI Il tessuto fotosintetico delle piante superiore è mesofillo fogliare ricco di clorofille specializzate nell’assorbire la luce. 4 tipi di clorofilla:• clorofilla A e B(in tutti organismi con fotosintesi ossigenica verdi),• clorofilla C (pigmenti accessori nelle alghe),• D batterioclorofille(nei batteri con fotosintesi anossigenica).Il Cloroplasto è la sede della fotosintesi.
26/01/13 Pag. 11 di20

Fotosistemi sono complessi costituiti da pigmento-proteina e svolgono l'azione di recettore: “ANTENNA” e collettore: “CENTRO DI REAZIONE” dove si trova il pigmento foto ossidabile.Fotosintesi è processo di trasduzione dell’energia luminosa in energia chimica attraverso reazioni redox; è l'unico sistema biochimico capace di ossidare l’acqua. Comprende le reazioni dei tilacoidi che raccolgono ioni H+ aumentando l'acidità agli estremi della membrana garantendo energia per la successiva fase: ATP sintetasi. Gli organismi che evolvono ossigeno possiedono 2 foto sistemi che lavorano in serie:
Foto sistema II° (680 rosso meno forte) produce un forte agente ossidante (in grado di ossidare l’acqua - lisi) e un debole agente riducente H2O > ADT > ATP (prodotto della fase luminosa, utilizzato nella fase oscura), è localizzato nelle membrane impilate (tilacoidi grana).
Foto sistema I° ( 700 rosso più forte ) produce un forte agente riducente capace di ridurre NADP > NADPH ed un debole agente ossidante. Si trova solo nelle membrane stromatiche non impilate.
Dalla lisi dell'acqua: ADP > ATP NADP > NADPH (trasporto energia H+)
MESOFILLO Insieme dei tessuti della foglia, esclusa l' epidermide, che l'avvolge.
METABOLISMO C3 C4 CAM
C 3 : Sono le più abbondanti . Nelle piante C3 invece il primo prodotto di fissazione stabile è il 3 fosfoglicerato. massimo tasso fotosintetico per mm2, max efficienza in condizioni luce e temperatura moderate, 400-1000gr di H2O per produrre 1 gr di materia secca.(stomi aperti anche di giorno), resa dipendente da T°; Le piante C3 quando vengono illuminate, gli stomi si aprono, entra la CO2 che viene fissata CO2 attraverso il ciclo di Calvin (RuBisCo - 5carbonio zucchero - in 3-fosfoglicerato) Tuttavia liberano anche anidride carbonica ed entrano in un meccanismo biochimico definito fotorespirazione (ciclo C2), processo ossigeno-dipendente.
C 4 : Sono adattate ad elevate intensità di luce e temperatura , <400gr di H2O per produrre 1 gr di materia secca; la loro richiesta energetica è superiore a quelle C3. La resa non dipende da T°, più efficienti delle C3 perchè minimizzano la fotorespirazione. Catturano la CO2 nel mesofillo e lo converte malato (acido carbonico a *C) poi riconvertito in CO2.
26/01/13 Pag. 12 di20

CAM : Il metabolismo acido delle crassulacee è un meccanismo di conversione della CO2 depositata nella la notte nei vacuoli nel primo prodotto stabile della fissazione che è l'acido bicarbossilico a 4 C. Questo processo permette di tenere chiusi gli stomi durante il giorno e quindi di conservare l'acqua, aprendoli di notte per l'ingresso della CO2. Durante la notte, gli stomi sono aperti consentendo l'ingresso della CO2 che viene convertito in malato e depositato nel mesofillo. Durante il giorno, quando gli stomi sono chiusi viene impedito l'ingresso della CO2 e il malato viene trasferito dai vacuoli e riconvertito in CO2 per essere usato dal RuBisCo per il ciclo di Calvbyn. Sono adatte ai climi aridi (organizzate per riserve di acqua ed acido crassulaceo); metabolismo intermittente; rapida fioritura;
OUABAINA L'ouabaina è un inibitore specifico della Pompa di Scambio Na/K: questa molecola agisce dal lato esterno della membrana cellulare ed arresta il ciclo di trasporto nelle sue fasi terminali perché, competendo con il sito specifico per il Potassio, ne impedisce il legame. Di conseguenza viene a mancare la defosforilazione Potassio-dipendente della molecola trasportatrice, che dovrebbe restituirla allo stato iniziale del ciclo di trasporto, e ciò blocca sia l’attività di trasporto che quella enzimatica della pompa.Poiché l’affinità per il glucoside è molto elevata, esso può bloccare il funzionamento della pompa anche a concentrazioni molto basse.
PEROSSIOMI Sono organelli caratterizzati dalla presenza di elevate concentrazioni di metaboliti secondari; sono considerati comparti metabolici specializzati, contenenti enzimi catalasi (detossificante), almeno 50 enzimi ossidativi.
PESSIONE RADICALE La pressione radicale è una delle forze responsabili della risalita della linfa all’interno delle piante. L'acqua del terreno passa per osmosi all'interno dei peli radicali attraverso la membrana costituita dalle cellule epidermiche. I peli radicali, assumendo dal terreno soluzioni di ioni inorganici e di piccole molecole di sali minerali, permettono l'instaurarsi di un gradiente concentrazione tra l'esterno (soluzioni poco concentrate nel terreno) e l'interno della pianta (soluzioni molto concentrate nelle cellule). Si genera così la cosiddetta "pressione radicale" che può essere sufficiente a spingere l'acqua fino ad altezze maggiori di quanto permetta la capillarità
PLASMOLISI Perdita di H2O , la membrana plasmatica si stacca dalla parete cellulare, opposto: turgore (eccesso di acqua), ok: isotonico.
POMPA SODIO POTASSIOTrasporto attivo primario
Detta anche pompa Na+/K+ ATP dipendente (Na+/K+ ATPasi), è un enzima che si trova nella membrana delle cellule . Questo tipo di pompa ionica è il più chiaro esempio di trasporto attivo primario di sostanze attraverso la membrana plasmatica . Si comporta da "antiporto", infatti, permette di trasportare contro gradiente di concentrazione tre ioni di Na + verso l'ambiente extracellulare e due ioni di K + verso l'ambiente intracellulare sfruttando l'energia derivante dall'idrolisi dell'adenosintrifosfato (ATP). All'inizio del processo di trasporto tre ioni di Na+ vanno a legarsi ai siti specifici ad alta affinità della proteina veicolo rivolti verso l'interno della cellula. Questo legame stimola la fosforilazione dipendente dall'ATP della pompa determinando un cambio conformazionale della proteina veicolo, la quale esporrà i siti di legame per gli ioni Na+ verso l'ambiente extracellulare, abbassando così la propria affinità per questi ioni cosicché essi vengano rilasciati al di fuori della cellula.Contemporaneamente alla fuoriuscita degli ioni Na+, due ioni K+ vanno a legarsi ai siti specifici esposti verso l'ambiente extracellulare stimolando la defosforilazione della proteina, la quale tornerà al suo stato conformazionale di partenza e rilascerà questi ioni all'interno della cellula. Una volta completata l'operazione, la proteina sarà pronta a ripetere il ciclo. Col progredire del
26/01/13 Pag. 13 di20

processo si determina una differenza di potenziale tra ambiente intracellulare ed ambiente extracellulare pari a circa -70 mV. Tale differenza è detta potenziale di membrana. La concentrazione del sodio è 143 mM all'esterno della membrana cellulare e 14 mM all' interno, mentre la concentrazione del potassio è dalle 10 alle 14 volte maggiore all'interno.Il contrasportatore Na-K-Cl (NKCC) è una proteina canale di membrana presente sul lato apicale dell' epitelio dell' ansa di Henle , permette il simporto attivo di uno ione potassio , uno ione sodio e due ioni cloruro (Na + /K + /2Cl - ) dal lume dell'ansa all'interno della cellula epiteliale , la sua azione è essenziale nella regolazione degli equilibri ionici e nell'osmolarità delle urine. Il trasporto è di tipo attivo secondario ed utilizza il sodio come ione motore, il gradiente del sodio è generato dalla pompa sodio potassio posizionata sulla membrana basolaterale della cellula stessa.
POTENZIALE DELL'ACQUA
Energia necessaria per spostare un'unità d'acqua. A pressione e temperatura ambiente Ψw = 0L'aggiunta di soluti rende meno negativa : Ψw < 0Pressioni positive innalzano Ψw Pressioni negative (tensione) riducono Ψw
Le tensioni negative assumono importanza sui lunghi tragittiΨ H2O = ΨPt + ΨS
ΨPt = pressione di turgore sempre positivaΨS = pressione osmotica sempre negativa del solutoH2O si muove verso regioni con Ψw più basso (Ψw delle piante << del Ψw suolo per consentire l'afflusso di acqua verso le foglie)
POTENZIALE DI MATRICE
Il potenziale di matrice (τ) è una misura della tendenza del terreno ad assorbire ulteriori molecole d’acqua
PRESSIONE NEGATIVA DELLO XILEMA
E' generata dall'evaporazione dell'acqua a livello delle foglie. Infatti se la traspirazione è bassa anche la tensione diminuisce.
PUNTO DI EQUILIBRIO Ψ Pb = - ( ΨPt – ΨS) = - Ψ H2O Ψ Pb + Ψ H2O= ΨS – ΨPt
Ψ Pb = pressione “artificiale” di equilibrio
SACCAROSIO TRASPORTO
Avviene tra:• le foglie mature superiori che forniscono gli assimilati ai meristemi apicali in via di sviluppo e alle foglie giovani che non producono saccarosio, mentre le foglie inferiori riforniscono prevalentemente il sistema radicale.
• foglie - frutto . Il frutto è un pozzo quindi riceve saccarosio dalla sorgente
SIMPLASTO Il simplasto è dato dalle cellule adiacenti il cui citoplasma (ovvero la matrice liquida in esse contenuta) è in comunicazione attraverso dei ponti (nello specifico mettono in comunicazione il reticolo endoplasmatico di due cellule adiacenti) chiamati plasmodesmi. L'acqua che è trasportata per via simplastica non può fluire liberamente come nell'apoplasto; segue due vie: quella attraverso la membrana o attraverso i plasmodesmi. Il simplasto rappresenta un rigoroso sistema di regolazione del flusso di acqua e delle sostanze che la pianta assorbe attraverso le radici. Trasferimento zuccheri in aree a bassa concentrazione (petali, gemme, apici radicali, giovani foglie, ecc.).
SINPORTO Protone e molecola si muovono nella stessa direzione. Trasporto attivo secondario.
SINTESI AMIDO - SACCAROSIO
Il s accarosio è traslocato nel floema in tutta la pianta. Amido è stabile ed insolubile di riserva in tutte le piante. Sia amido e saccarosio sono caboidrati sintetizzati dal trioso fosfato generato dal ciclo di Calvin.
26/01/13 Pag. 14 di20

TRASPIRAZIONE Nel suo cammino dalla foglia all’atmosfera l’acqua è aspirata dallo xilema alle pareti cellulari del mesofillo dove evapora negli spazi cellulari fogliari. Tutta l’acqua persa dalle foglie esce per diffusione sotto forma di vapor d’acqua dai pori degli stomi. Chiusura o apertura degli stomi è regolata da cellule epidermiche specializzate (cellule di guardia) ed è causata da un aumento della pressione di turgore nelle cellule di guardia.
L’acqua nelle piante si muove sempre verso potenziali idrici più negativi. La traspirazione è principale causa di risalita dell’acqua nella pianta,è evaporazione attraverso la superficie della pianta. Solo il 5% della traspirazione avviene con la cuticola (epidermide), il resto avviene a livello degli stomi (foglie). La forza motrice della traspirazione è il gradiente di concentrazione del vapore acqueo. Fattori che regolano la perdita di acqua dalla foglia è gradiente di vapore acqueo e le resistenze alla diffusione del vapore. La traspirazione causa raffreddamento delle foglie perché evaporazione di acqua è associata alla perdita di calore. La resistenza stomatica è regolata da apertura e chiusura degli stomi. Forza motrice per la perdita d’acqua dalla foglia è gradiente di concentrazione assoluta (dipendente dalla T°).
La linfa xilematica è molto diluita, quindi il suo Ψs sarà solo poco negativo, ma essa si trova praticamente sempre in uno stato di tensione (P negativo), quindi il Ψ sarà più negativo nello xilema (-10) che nell’acqua circolante nel terreno (-5). Di conseguenza, l’acqua passerà dal terreno alla pianta. Affinché le piante possano sopravvivere deve esserci sempre questo gradiente di Ψ tra il terreno e le radici. Quindi se le soluzioni circolanti nel terreno si fanno troppo concentrate, c’è il rischio che l'acqua venga richiamata dalle radici verso il terreno, con conseguente morte della pianta. Perchè si abbia traspirazione il potenziale idrico dell'aria deve essere più basso di quello fogliare. La perdita acqua sotto forma di vapore da parte di un tessuto biologico a contatto con l'ambiente esterno si ha prevalentemente a livello delle foglie ed è regolata al 90%, laddove sono presenti, dagli stomi. L'acqua si muove secondo gradienti di potenziale idrico:• acqua libera potenziale (Ψ= 0);• potenziale nel terreno (Ψ=-5);• potenziale nella pianta (Ψ=-10);• potenziale nell'atmosfera (da potenziale idrico maggiore - dentro la foglia - a potenziale idrico minore: nell'atmosfera basta che l'umidità relativa sia poco sotto il 100% perché il potenziale idrico sia molto inferiore a quello fogliare.)
TRASPORTO ATTIVO PRIMARIO E SECONDARIO
PRIMARIO: il consumo di kj/mol, normalmente sotto forma di idrolisi di molecole di adenosintrifosfato (ATP), è accoppiata direttamente al movimento della sostanza attraverso la membrana. Nella maggior parte dei casi il trasporto primario è eseguito da ATPasi. Un tipico trasportatore primario, presente universalmente nelle cellule è la pompa sodio-potassio (o ATPasi sodio-potassio), che contribuisce al mantenimento del potenziale di membrana.SECONDARIO: non viene speso direttamente ATP, ma viene sfruttata la differenza di potenziale elettrochimico creata dai trasportatori attivi che pompano ioni al di fuori della cellula. Questo significa che il trasporto secondario si basa comunque sul consumo di ATP, che permette di mantenere la differenza di potenziale senza la quale non ci sarebbe possibilità di trasporto secondario.Tre tipi di trasporto secondario:Uniporto : è il trasporto secondario di una sola sostanza che si muove sfruttando la differenza di potenziale elettrochimico creato da trasportatori primari.
26/01/13 Pag. 15 di20

Simporto: usa, analogamente all'antiporto, il flusso di un soluto secondo gradiente per muovere un'altra molecola contro gradiente ma il movimento avviene in questo caso attraversando la membrana nella stessa direzione. Un esempio è il simportatore di glucosio , che cotrasporta secondo gradiente due ioni sodio per ogni molecola di glucosio import ata nella cellula .
Antiporto: è il trasporto contemporaneo di due specie ioniche o di altri soluti che si muovono in direzioni diverse attraverso la membrana. Una delle due sostanze viene lasciata fluire secondo gradiente, da un compartimento ad alta concentrazione ad uno a bassa concentrazione. Questo genera l'energia entropica necessaria per guidare il trasporto dell'altro soluto contro gradiente, da bassa ad alta concentrazione. Un esempio di questo tipo di trasporto è lo scambiatore sodio-calcio , che mediano l'ingresso di ioni sodio e l'uscita del calcio . Molte cellule possiedono anche una calcio ATPasi che però opera molto più lentamente dello scambiatore e ha un diverso ruolo fisiologico.
TRASPORTO DEI SOLUTI ATTREAVERSO LA MEMBRANA
PASSIVO: il movimento (spontaneo) della sostanza avviene nella direzione del suo gradiente di concentrazione
Alta concentrazione >> Bassa concentrazionele molecole non cariche si spostano secondo il loro gradiente di concentrazione,le molecole cariche si spostano in base al gradiente elettrochimicoATTIVO:nel trasporto attivo è richiesta una spesa energetica (es. ATP) ed è sempre necessaria la mediazione di un trasportatore. In questa forma di trasporto le molecole si muovono contro un gradiente elettrico, chimico o elettrochimico.
Bassa concentrazione >> Alta concentrazione
TRASPORTO DELL'ACQUA
Velocità di diffusione è elevata a breve distanza ma è molto bassa a lunga distanza. Il trasporto dell’acqua a lunga distanza avviene tramite flusso di massa causato dalla pressione.
Potenziale idrico è potenziale chimico dell’acqua diviso il volume molale parziale dell’acqua (misura dell’energia libera dell’acqua per unità di volume)Potenziale del soluto o potenziale osmotico è l’effetto della presenza di soluti disciolti sul potenziale idrico. Potenziale di matrice = utilizzato per tener conto della riduzione di energia libera dell’acqua (es. Tessuti di riserva, suoli aridi).L’acqua cosi come entra allo stesso modo esce dalla cellula in risposta ad un gradiente di potenziale idrico.Pressione idrostatica positiva all’interno della cellula = pressione di turgore;Pressione idrostatica negativa = pressione di tensione.
Conduttività idraulica = facilità con la quale l'acqua può spostarsi attraverso una membrana. Acquaporine = proteine integrali di membrana che agevolano spostamento dell’acqua attraverso le membrane cellulari. L’appassimento si verifica quando la pressione idrostatica all’interno delle cellule scende verso lo zero. La conduzione dell'acqua attraverso lo xilema avviene per: * pressione radicale * potenziali osmotici molto negativi nelle foglie (evapotraspirazione) * flusso del floema reintegrato prendendo acqua dallo xilema
Movimento dell'acqua nello xilema (da radice a foglie: l'acqua che si trova in cima agli alberi sviluppa una grande tensione (una pressione idrostatica
26/01/13 Pag. 16 di20

negativa), che è in grado di aspirare l'acqua presente nello xilema. Tale meccanismo è solitamente definito la teoria della coesione-tensione della linfa ascendente, poiché richiede la presenta delle proprietà coesive dell'acqua per tollerare le grandi tensioni presenti nella colonna d'acqua dello xilema.
TRASPORTO FUSTO-RADICI-SUOLO
Forze di coesione/adesione delle molecole d’acqua consentono la trasmissione della tensione e la formazione di colonne d’acqua intatte che determinano risalita dell’acqua nello xilema. La tensione può trasmettere fino allo xilema delle piccole nervature. La coesione consente all’acqua di resistere ad una grande forza di tensione (-30 Mpa). La lignificazione delle pareti cellulari dello xilema consente la trasmissione della tensione. Tramite la guttazione alcune piante trasudano l’eccesso di linfa grezza durante la notte tramite gli idatodi.
TRASPORTO PASSIVO Il trasporto passivo consiste nel passaggio di molecole secondo gradiente, non è richiesto l’utilizzo di energia (come ATP). Può essere mediato da proteine di membrana. Può essere di due tipi differenti: diffusione semplice o facilitata. Diffusione semplice - è un tipo di trasporto che non necessita di nessun sistema proteico. Le molecole apolari come gli ormoni stereoidei sono capaci di diffondere facilmente attraverso il doppio strato lipidico. Le molecole apolari passano facilmente perché la zona interna della membrana è costituita de catene idrocarburiche. Anche piccole molecole polari come l'urea, il glicerolo, l'etanolo e l'acqua (quest'ultima nonostante la sua forte polarità riesce a passare perché molto piccola si insinua fra le molecole polari), riescono a passare. In questo tipo di trasporto, il gradiente di concentrazione è favorevole , ed è di segno positivo. Non c'è pertanto una spesa energetica in termini di ATP per spingere la molecola dall’altra parte della membrana.Diffusione facilitata - ha, come integrali della membrana, delle proteine canale (carrier) che facilitano e velocizzano il passaggio di una specifica molecola o di uno specifico ione, attraverso la formazione di legami deboli, necessari per facilitare il rilascio della molecola dall’altra parte della membrana. Zuccheri semplici e amminoacidi passano attraverso i carrier mentre gli ioni generalmente passano soprattutto attraverso i canali. Anche in questo tipo di trasporto, il gradiente di concentrazione è favorevole, ed è di segno positivo, pertanto non c’è una spesa energetica (in termini di ATP) per spingere la molecola dall’altra parte della membrana.
TRASPORTO POTENZIALE IDRICO
Potenziale idrico è l’energia per unità di volume necessaria per trasportare acqua reversibilmente e isotermicamente. Potenziale idrico definisce la direzione del flusso idrico e per valutare lo stato idrico della pianta. L'acqua si muove sempre verso regioni a potenziale idrico più basso. Potenziale idrico dipende da concentrazione, pressione e gravità. Pressione osmotica è la pressione che deve essere applicata ad una soluzione per controbilanciare la diffusione dell’acqua verso soluzione + concentrata; se negativa indica che i soluti disciolti riducono il potenziale idrico di una soluzione. Il potenziale osmotico è l’effetto sul potenziale idrico dei soluti disciolti nella soluzione. Potenziale di pressione si misura come differenza con la pressione atmosferica (è negativo nello xilema quando è attiva la traspirazione si dice tensione). Pressione di turgore è importante per distendere le pareti nella crescita e per aumentare la rigidità meccanica dei tessuti non lignificati. Potenziale di matrice è riferito a forze di adsorbimento ed è negativo o al max uguale a zero. Potenziale di matrice è una misura della tendenza della matrice ad assorbire altra acqua. Nel caso di cellule con pareti idratate il potenziale di matrice non si considera. Il potenziale idrico di una cellula è uguale a quello del liquido o vapore con cui la cellula è in equilibrio. Il movimento spontaneo dell’acqua può avvenire per diffusione molecolare (gradiente di concentrazione) flusso di massa (gradiente di pressione) osmosi (gradiente di potenziale).
26/01/13 Pag. 17 di20

Ciclo di CalvinLa riduzione del carbonio avviene nello stroma in una serie ciclica di reazioni che prende il nome dallo scienziato che l'ha osservata per primo, il chimico statunitense Melvin Calvin. Il ciclo di Calvin è analogo al ciclo di Krebs in quanto, a ogni giro, il composto iniziale viene rigenerato. Il composto iniziale (e anche quello finale) del ciclo di Calvin è costituito da uno zucchero a cinque atomi di carbonio legato a due gruppi fosfato, il
ribulosio difosfato (RuDP), che fu scoperto nelle alghe e nelle foglie di spinacio. Il ciclo comincia quando l'anidride carbonica si lega al RuDP. Per identificare il prodotto di questa reazione, Melvin Calvin e i suoi collaboratori fecero crescere per un breve periodo di tempo alcune alghe fotosintetiche in presenza di molecole di anidride carbonica i cui atomi di carbonio erano stati resi radioattivi (fondino arancione nella figura). Il primo prodotto in cui trovarono gli atomi di carbonio marcati fu l'acido fosfoglicerico (PGA) ; ciò indicava che, quando l'anidride carbonica si lega, al RuDP, la molecola si scinde immediatamente per formare due molecole di PGA. Questa reazione è catalizzata da un enzima specifico, la RuDP-carbossilasi (oggi nota come rubisco), che costituisce più del
15% del quantitativo proteico del cloroplasto; la RuDP-carbossilasi, in effetti, è ritenuta la proteina più abbondante sulla, Terra. Ogni molecola di PGA prodotta in quest'a reazione iniziale contiene tre atomi di carbonio; per questo motivo, il ciclo di Calvin è conosciuto anche come via del C3. Come nel ciclo di Krebs, ogni passaggio del ciclo di Calvin è catalizzato da un enzima specifico. A ogni giro completo una molecola di anidride carbonica entra nel ciclo, viene ridotta e si riforma una molecola di RuDP tre giri del ciclo fanno entrare tre molecole di anidride carbonica, sufficienti per ottenere uno zucchero a tre atomi di carbonio, la gliceraldeide 3-fosfato (G3P). Occorrono sei giri completi del ciclo, con l'introduzione di sei molecole di anidride carbonica, per produrre l'equivalente di una molecola di zucchero a sei atomi di carbonio, con il glucosio. L'equazione complessiva è la seguente: 6RuDP + 6CO2 + 12NADPH + 12H +
+18ATP → 6RuDP + glucosio + 12NADP +
+18ADP + 18P + 6H2O Partendo dalla scritta in alto a sinistra: sei molecole di ribulosiodifosfato (RuDP), un composto a cinque atomi di carbonio, si combinano con sei molecole di anidride carbonica. Ciò produce sei molecole di un composto intermedio instabile che si scinde immediatamente dando origine a dodici molecole di acido fosfoglicerico (PGA), un composto a tre atomi di carbonio. Nelle successive due tappe queste molecole sono ridotte a dodici molecole di gliceraldeide 3-fosfato. Dieci di queste molecole a tre atomi di carbonio si combinano e si riassemblano per riformare sei molecole di RuDP a cinque atomi di carbonio (ossia, il composto iniziale). Le due molecole «in più» di gliceraldeide 3-fosfato rappresentano il guadagno netto del ciclo di Calvin. La gliceraldeide 3-fosfato è la stessa molecola a tre atomi di carbonio che si forma quando il fruttosio difosfato si scinde nella quarta tappa della glicolisi. Usando questa molecola come punto di
26/01/13 Pag. 18 di20

partenza, la cellula può sintetizzare una certa varietà di zuccheri, amminoacidi e acidi grassi.
Una soluzione alla carenza di CO2: la fotorespirazione. In presenza di una grande quantità di anidride carbonica, il ciclo di Calvin svolge in genere regolarmente; invece, quando nella foglia la concentrazione di anidride carbonica è bassa, se paragonata a quella dell'ossigeno, l'enzima RuDP-carbossilasi catalizza una reazione tra RuDP e ossigeno invece che tra RuDP e anidride carbonica. Questa reazione è la prima tappa di un processo conosciuto come fotorespirazione, in cui i carboidrati vengono ossidati in presenza di luce con formazione di acqua e dell' anidride carbonica; in altre parole, la cellula si autoproduce la CO2 di cui necessita. Tuttavia, in modo diverso dalla respirazione dei mitocondri, la foto respirazione, infatti, non è un processo redditizio dal punto di vista energetico in quanto non produce né ATP né NADH. In alcune piante più del 50% del carbonio fissato nella foto sintesi può essere riossidato ad anidride carbonica durante il processo di fotorespirazione. Le condizioni che possono innescare la fotorespirazione sono piuttosto comuni. l'anidride carbonica non è sempre disponibile per le cellule fotosintetiche di una pianta. Come abbiamo visto, questo gas entra nella foglia attraverso gli storni, pori
26/01/13 Pag. 19 di20

specializzati che si aprono e si chiudono in seguito alla pressione dell'acqua e ad altri fattori. Quando una pianta si trova in un ambiente caldo e asciutto deve chiudere i suoi storni per conservare l'acqua; ciò impedisce il rifornimento di anidride carbonica e favorisce invece l'accumulo di ossigeno prodotto dalla fotosintesi. Come conseguenza si hanno concentrazioni basse di anidride carbonica e alte di ossigeno, fattori che inducono la fotorespirazione.
Una seconda soluzione: la via del C4.Il problema della carenza di CO2 viene risolto da alcune piante grazie a una via alternativa per catturare questo gas. In queste piante la prima tappa della fissazione del carbonio consiste nel legame tra l'anidride carbonica e un composto chiamato acido fosfoenolpiruvico (PEP), e nella formazione di un composto a quattro atomi di carbonio, l'acido ossalacetico. (L'acido fosfoenolpiruvico è un prodotto intermedio della glicolisi e l'acido ossalacetico è un prodotto intermedio del ciclo di Krebs). Inizialmente, nelle cellule del mesofillo della foglia, l'acido ossalacetico viene trasformato in acido malico. Attraverso i plasmodesmi l'acido malico passa nelle cellule della guaina vascolare, che formano un rivestimento intorno ai fasci vascolari e che, a differenza delle cellule della guaina delle piante C3, contengono i cloroplasti. Qui l'acido malico è demolito in acido piruvico e anidride carbonica che entra nel ciclo di Calvin l'acido piruvico passa attraverso i plasmodesmi verso le cellule del mesofillo, dove viene ritrasformato in acido fosfoenolpiruvico, pronto per ricevere un' altra molecola di anidride carbonica.Le piante che utilizzano questa via sono comunemente chiamate piante C4, per distinguerle dalle piante C3 in cui inizialmente il carbonio viene catturato per formare un composto a tre atomi di carbonio, l'acido fosfoglicerico (PGA). La PEP-carbossilasi, l'enzima che catalizza la sintesi di acido ossalacetico nelle piante C4, ha un' affinità maggiore per l'anidride carbonica rispetto alla RuDP-carbossilasi, l'enzima che catalizza la formazione di PGA. Se gli stomi devono rimanere chiusi gran parte del loro tempo, come accade, per esempio, nei climi caldi e secchi per conservare acqua, la pianta con metabolismo C4 prenderà più anidride carbonica a ogni, per così dire, «respiro», rispetto alla pianta con metabolismo C3. Le piante C4 si sono evolute principalmente nelle zone tropicali e si sono adattate particolarmente bene a un'alta intensità luminosa, alle alte temperature e alla siccità; la canna da zucchero e il granturco sono tra le piante C4 più conosciute.
Una terza soluzione: le piante CAM.Di solito gli stomi delle piante si aprono durante il giorno e si chiudono durante la notte, ma in alcune specie accade esattamente il contrario. Di notte, non solo la temperatura è più bassa, ma l'umidità è in genere più alta; entrambi questi fattori riducono la velocità di traspirazione. Le specie che aprono gli storni soltanto di notte comprendono una certa varietà di piante adattate ai climi caldi e secchi, come, per esempio, i cactus, gli ananas e i membri della famiglia delle crassulacee come, per esempio, il genere Sempervivum che cresce in montagna. Queste piante assorbono anidride carbonica di notte, trasformandola in acidi a quattro atomi di carbonio; durante il giorno, quando gli stomi si chiudono, l'anidride carbonica viene liberata da questi acidi organici e utilizzata immediatamente per la fotosintesi. Questo processo è detto metabolismo acido delle crassulacee e le piante che lo compiono sono dette piante CAM. Il metabolismo acido è analogo alla via del C4 della fotosintesi, sebbene si sia evoluto probabilmente in maniera indipendente.
26/01/13 Pag. 20 di20