Fisiologia respiratoria (meccanica, scambio e trasporto dei gasi, controllo nervoso).pdf
13
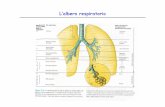
MECCANICA RESPIRATORIA Basi anatomo-funzionali Nell’apparato respiratorio abbiamo zone di conduzione e zone respiratorie. Le zone di conduzione vanno da naso/bocca, attraverso faringe, laringe, trachea e bronchi fino ai bronchioli terminali (in corrispondenza dei quali termina la circolazione bronchiale, di derivazione sistemica, che nutre i bronchi e il parenchima polmonare). Le funzioni delle zone di conduzione sono principalmente: • Riscaldamento (tramite ampio contatto con una ricca vascolarizzazione) e umidificazione (tramite evaporazione del velo liquido che ricopre costantemente l’epitelio respiratorio insieme al muco) dell’aria in transito • Ripartizione omogenea del flusso di aria tramite regolazione del calibro dei bronchi • Filtraggio delle particelle con diametro superiore ai 15 µ grazie alle vibrisse nasali • Protezione, tramite l’intrappolamento nel muco, da corpi estranei e conseguente clearance mucociliare o riflessi come tosse e starnuti. Dalla 18esima generazione di diramazioni bronchiali (dunque le ultime 7) si hanno le zone respiratorie, costituite da bronchioli respiratori, dotti alveolari e sacchi alveolari. Gli alveoli non sono costituiti da epitelio ciliato, né presentano muco; presentano 4 citotipi: • Pneumociti (cellule alveolari) di I° tipo. Cellule appiattite, tappezzano la superficie alveolare rappresentandone il 95% • Pneumociti di II° tipo. Cellule cuboidi che sintetizzano una sostanza fosfolipidica, il surfattante (dipalmitoil-lecitina), che riduce la tensione superficiale dell’alveolo. In caso di danni alle cellule di tipo I, i pneumociti di II tipo proliferano e possono rimpiazzarle (differenziandosi in pneumociti di I tipo, assicurando l’integrità e il normale spessore della superficie alveolare). • Pneumociti di III° tipo. Associate a terminazioni nervose, si pensa abbiano funzione chemocettiva. • Macrofagi alveolari. Sono macrofagi residenti nell’alveolo, con funzione classicamente fagocitaria e di pulizia dei detriti. Se ingeriscono sostanze non degradabili, i macrofagi migrano fino ai tratti distali della zona di conduzione per essere eliminati tramite il trasporto mucociliare. I capillari polmonari, interposti tra l’arteria polmonare (proveniente dal cuore destro) e le vene polmonari (che si immettono nel cuore sinistro), formano una fitta rete di vasi attorno agli alveoli. Si configura così la barriera aria-sangue, spessa tra 0,15 e 0,5 µ. Essa è costituita da: • Aria • Sottile velo liquido e surfattante (0,01 µ) • Superficie alveolare (cellule I tipo) • Membrana basale (spesso è la medesima per i pneumociti e i capillari e si trova a cavallo tra essi) • Endotelio capillare • Sangue (con eritrociti) L’impalcatura connettivale degli alveoli contiene molte fibre elastiche, che contribuiscono ai fenomeni di ritorno elastico che riducono il volume del polmone e degli alveoli dopo la dilatazione attiva ottenuta con la contrazione dei muscoli inspiratori. Pleure e liquido pleurico Le pleure sono membrane sierose (mesoteliali); la pleura parietale riveste internamente la parete toracica e il diaframma mentre la pleura viscerale riveste il polmone. La pleura parietale è in continuità con la pleura viscerale: a livello dell’ilo del polmone si ha la riflessione delle pleure. Tra i due foglietti pleurici (parietale e viscerale) vi è una cavità virtuale riempita dal liquido pleurico. Esso ha contenuto elettrolitico molto simile a quello del plasma mentre il contenuto proteico è nettamente inferiore. È prodotto per filtrazione dai capillari della pleura parietale. I capillari della pleura viscerale invece, insieme ai numerosi stomi linfatici, sono responsabili dell’assorbimento. L’attività di aspirazione del liquido è molto intensa e produce pressioni nettamente subatmosferiche che hanno un ruolo importante nel mantenere in stato di adesione i due foglietti pleurici: in tal modo il polmone è costretto a seguire i movimenti della gabbia toracica. Resistenze meccaniche alla respirazione Per mobilizzare aria nel e dall’apparato respiratorio, i muscoli respiratori devono vincere le resistenze al moto dipendenti dal polmone e dalle strutture della parete toracica. Vi sono resistenze elastiche che si oppongono all’espansione del sistema toraco-polmonare e resistenze non elastiche (viscose e inerziali).
-
Upload
bobbygustafson -
Category
Documents
-
view
239 -
download
5
Transcript of Fisiologia respiratoria (meccanica, scambio e trasporto dei gasi, controllo nervoso).pdf

MECCANICA RESPIRATORIA
Basi anatomo-funzionali Nell’apparato respiratorio abbiamo zone di conduzione e zone respiratorie. Le zone di conduzione vanno da naso/bocca, attraverso faringe, laringe, trachea e bronchi fino ai bronchioli terminali (in corrispondenza dei quali termina la circolazione bronchiale, di derivazione sistemica, che nutre i bronchi e il parenchima polmonare). Le funzioni delle zone di conduzione sono principalmente:
• Riscaldamento (tramite ampio contatto con una ricca vascolarizzazione) e umidificazione (tramite evaporazione del velo liquido che ricopre costantemente l’epitelio respiratorio insieme al muco) dell’aria in transito
• Ripartizione omogenea del flusso di aria tramite regolazione del calibro dei bronchi • Filtraggio delle particelle con diametro superiore ai 15 µ grazie alle vibrisse nasali • Protezione, tramite l’intrappolamento nel muco, da corpi estranei e conseguente clearance
mucociliare o riflessi come tosse e starnuti. Dalla 18esima generazione di diramazioni bronchiali (dunque le ultime 7) si hanno le zone respiratorie, costituite da bronchioli respiratori, dotti alveolari e sacchi alveolari. Gli alveoli non sono costituiti da epitelio ciliato, né presentano muco; presentano 4 citotipi:
• Pneumociti (cellule alveolari) di I° tipo. Cellule appiattite, tappezzano la superficie alveolare rappresentandone il 95%
• Pneumociti di II° tipo. Cellule cuboidi che sintetizzano una sostanza fosfolipidica, il surfattante (dipalmitoil-lecitina), che riduce la tensione superficiale dell’alveolo. In caso di danni alle cellule di tipo I, i pneumociti di II tipo proliferano e possono rimpiazzarle (differenziandosi in pneumociti di I tipo, assicurando l’integrità e il normale spessore della superficie alveolare).
• Pneumociti di III° tipo. Associate a terminazioni nervose, si pensa abbiano funzione chemocettiva.
• Macrofagi alveolari. Sono macrofagi residenti nell’alveolo, con funzione classicamente fagocitaria e di pulizia dei detriti. Se ingeriscono sostanze non degradabili, i macrofagi migrano fino ai tratti distali della zona di conduzione per essere eliminati tramite il trasporto mucociliare.
I capillari polmonari, interposti tra l’arteria polmonare (proveniente dal cuore destro) e le vene polmonari (che si immettono nel cuore sinistro), formano una fitta rete di vasi attorno agli alveoli. Si configura così la barriera aria-sangue, spessa tra 0,15 e 0,5 µ. Essa è costituita da:
• Aria • Sottile velo liquido e surfattante (0,01 µ) • Superficie alveolare (cellule I tipo) • Membrana basale (spesso è la medesima per i pneumociti e i capillari e si trova a cavallo tra essi) • Endotelio capillare • Sangue (con eritrociti)
L’impalcatura connettivale degli alveoli contiene molte fibre elastiche, che contribuiscono ai fenomeni di ritorno elastico che riducono il volume del polmone e degli alveoli dopo la dilatazione attiva ottenuta con la contrazione dei muscoli inspiratori. Pleure e liquido pleurico Le pleure sono membrane sierose (mesoteliali); la pleura parietale riveste internamente la parete toracica e il diaframma mentre la pleura viscerale riveste il polmone. La pleura parietale è in continuità con la pleura viscerale: a livello dell’ilo del polmone si ha la riflessione delle pleure. Tra i due foglietti pleurici (parietale e viscerale) vi è una cavità virtuale riempita dal liquido pleurico. Esso ha contenuto elettrolitico molto simile a quello del plasma mentre il contenuto proteico è nettamente inferiore. È prodotto per filtrazione dai capillari della pleura parietale. I capillari della pleura viscerale invece, insieme ai numerosi stomi linfatici, sono responsabili dell’assorbimento. L’attività di aspirazione del liquido è molto intensa e produce pressioni nettamente subatmosferiche che hanno un ruolo importante nel mantenere in stato di adesione i due foglietti pleurici: in tal modo il polmone è costretto a seguire i movimenti della gabbia toracica. Resistenze meccaniche alla respirazione Per mobilizzare aria nel e dall’apparato respiratorio, i muscoli respiratori devono vincere le resistenze al moto dipendenti dal polmone e dalle strutture della parete toracica. Vi sono resistenze elastiche che si oppongono all’espansione del sistema toraco-polmonare e resistenze non elastiche (viscose e inerziali).

Pressioni in meccanica respiratoria
• Pressione perduttale: differenza tra pressione alveolare e pressione atmosferica o barometrica. P=PA - PB. È quella realmente responsabile della direzione del flusso di aria, ci indica il gradiente pressorio. Per convenzione la pressione barometrica è 0; dunque se la pressione alveolare è negativa (inspirazione), il flusso d’aria è in entrata; se la pressione alveolare è positiva (espirazione), il flusso è in uscita. Quando la pressione alveolare è 0 non si ha flusso, come accade al termine della respirazione eupnoica (a capacità funzionale residua).
• Pressione transpolmonare: differenza tra pressione alveolare e pressione pleurica. PP = PA – Ppl. Esprime la forza di retrazione elastica del polmone; è spesso definita come pressione di espansione polmonare (mantiene espansi gli alveoli).
• Pressione transtoracica: differenza tra pressione pleurica e pressione barometrica. PT = Ppl - PB. Mantiene espansa la gabbia toracica. Essendo la pressione barometrica convenzionalmente 0, solitamente la pressione transtoracica è negativa. Questo accade perché la pressione pleurica è generalmente subatmosferica; però in condizioni di espirazione forzata la pressione pleurica può divenire positiva e di conseguenza anche la pressione transtoracica.
Relazione P/V del polmone isolato Insufflando aria in un polmone isolato, avremo che all’inizio per aumenti considerevoli della pressione corrisponderanno piccoli aumenti di volume; al contrario successivamente per modesti incrementi di pressione si avranno notevoli incrementi di volume; nell’ultimo tratto, la curva pressione/volume si comporta come il tratto iniziale. Completata la misurazione in insufflazione, eseguiamo la registrazione di pressione e volume svuotando il polmone ad intervalli. Sorprendentemente, a parità di volume i valori di pressione endoalveolare sono inferiori. Considerando le due curve, di insufflazione e di svuotamento, esse delimitano un’area, detta area di isteresi polmonare. Per isteresi si intende una reazione ritardata ad una sollecitazione; la reazione dipende anche dallo stato precedente. A parità di pressione, si hanno volumi diversi; a parità di volume, si hanno pressioni diverse. Se la stessa procedura viene effettuata con soluzione fisiologica invece che con aria, le due curve sono sovrapponibili e non v’è un’apprezzabile area di isteresi. Se ne deduce quindi che questo comportamento è dovuto alla tensione superficiale, che agisce solamente quando insuffliamo aria. Le caratteristiche elastiche del parenchima polmonare invece sono fattori che agiscono sia nel riempimento con aria che nel riempimento con soluzione fisiologica. Nel polmone collassato, a minimo volume (300 mL circa), gli alveoli hanno un raggio molto ridotto.
Considerando la legge di Laplace (� =��
�), se il raggio è piccolo la pressione necessaria a distendere
l’alveolo è molto maggiore (la tensione superficiale oppone resistenza alla distensione). Iniziata l’espansione, il raggio degli alveoli è maggiore e la resistenza da vincere è minore. Nelle ultime fasi di riempimento, l’ulteriore messa in tensione del tessuto polmonare richiede nuovamente un notevole incremento di pressione, stavolta per ragioni elastiche. Nella fase di svuotamento, gli alveoli partono da una posizione di distensione completa e la loro riduzione di volume avviene in modo uniforme, con tensione superficiale ridotta rispetto all’insufflazione, e dunque valori pressori minori a parità di volume. In vivo, la tensione superficiale (che tende a far collassare gli alveoli) è contrastata dal surfattante prodotto dagli pneumociti di II° tipo. I muscoli respiratori devono così vincere una resistenza più bassa, determinata perlopiù dal ritorno elastico del parenchima polmonare.
0
10
20
30
40
50
60
70
80
90
100
0 5 10 15 20
Curva P/V del polmone escisso
In ordinate il volume percentuale (100 = CPT); in ascisse la pressione in cm H20.
AREA DI ISTERESI

Relazione P/V del polmone in vivo Si misura la pressione endopleurica (con un palloncino esofageo) dopo un'inspirazione massimale, e si ottiene così una misura della massima forza di retrazione elastica del polmone; poi il soggetto espira volumi di aria progressivamente minori fino al volume residuo, quando la pressione endopleurica è di poco inferiore alla pressione atmosferica, essendo diminuita di molto la forza di retrazione. Si ottiene così una curva da cui si evince che la compliance del polmone aumenta progressivamente man mano che ci si avvicina al volume residuo (è piatta per volumi vicini alla capacità polmonare totale, più ripida per volumi vicini al volume residuo). Ne risulta che la compliance polmonare è massima per volumi corrispondenti alla CFR. Relazione P/V della gabbia toracica Ipoteticamente asportiamo il polmone e ricostituiamo la cavità toracica, con la parete toracica e il diaframma. Insufflando aria e quindi aumentando il volume si produce un aumento della pressione, che si manifesta come la tendenza della parete toracica a tornare nella condizione di partenza. Alla massima espansione la pressione è piuttosto bassa (8-10 cm H2O), a dimostrazione della notevole distensibilità della parete toracica. Se invece di immettere aria la aspiriamo, la pressione diminuisce e diventa negativa; con una espirazione massima può arrivare a -40 cm H2O. Dunque se la gabbia toracica è portata, per insufflazione o svuotamento, a valori di volume diversi da quello di equilibrio, tenderà sempre a tornare al volume di equilibrio (rispettivamente tramite riduzione di volume o tramite espansione). Differenze tra gabbia toracica e polmone Sostanzialmente il polmone tende sempre al collasso, mentre la gabbia toracica tende al collasso o all’espansione in relazione al proprio volume rispetto a quello d’equilibrio. Relazione P/V del sistema toraco-polmonare La pressione esercitata dal sistema toraco-polmonare è la somma algebrica della pressione transpolmonare e della pressione transtoracica. Dunque PP+T = PP + PT. Ma se PP = PA – Ppl e PT = Ppl - PB, allora PP+T = PA – Ppl + Ppl - PB. Semplificando si ha PP+T =PA - PB. Ricordando che PB è convenzionalmente 0, la pressione di distensione del sistema toraco-polmonare sarà quella misurata negli alveoli. Questa pressione esprime per ogni volume la forza esercitata dalle forze elastiche del sistema toraco-polmonare sull’aria contenuta e può essere positiva o negativa in relazione al volume del sistema. A valori di CPT, la pressione endoalveolare è di circa +49 cm H2O, ovvero la somma di +40 cm H2O data dalla retrazione del polmone e +9 cm H2O dalla pressione della gabbia toracica. Dunque sia il polmone (come sempre) che la gabbia toracica a questi valori volumetrici tendono a collassare. A fine di una inspirazione normale, la gabbia toracica è in equilibrio e dunque la pressione è +9 H2O, dovuta solamente alla retrazione del polmone. Al termine di una espirazione normale (ovvero raggiunto il volume di capacità funzionale residua), si ha il punto d’equilibrio di tutto il sistema: il volume è equivalente a circa il 35% della CV e la pressione è 0. La tendenza del polmone al collasso (+5 H2O) è controbilanciata dal valore pressorio negativo (ovvero la tendenza all’espansione) sviluppato dalla parete toracica (-5 H2O). La CFR è dunque il volume che permette l’equilibrio pressorio del sistema, il volume a cui il sistema tende spontaneamente a muscoli rilasciati. A valori inferiori della CFR, la pressione endoalveolare è sempre negativa: la tendenza all’espansione della gabbia toracica sarà sempre maggiore della tendenza del polmone al collasso. A VR si avrà -44 cm H2O per la gabbia toracica e +2 cm H2O per il polmone, con pressione endoalveolare risultante di 42 cm H2O. Compliance durante respirazione a riposo: 200 mL/cm H2O

Resistenze viscose e inerziali Oltre alle forze statiche che contrastano la variazione del sistema torace-polmone, vi sono forze dinamiche dovute alle resistenze che l’aria incontra nelle vie aeree (resistenze viscose) e all’attrito interno tra i tessuti (resistenze inerziali, molto minori). Il flusso d’aria nelle vie aeree può essere laminare (cilindri concentrici) e osservare la legge di Poiseuille, di transizione (vortici in corrispondenza della biforcazione delle vie aeree) o turbolento (vortici in ogni direzione con notevole attrito interno; si instaura quando il numero di Reynolds supera 2000). Nella trachea e nei grossi bronchi il moto è turbolento (alta velocità); nelle vie respiratorie intermedie il flusso è probabilmente transizionale; solo nelle piccole vie respiratorie il moto è laminare. In ottemperanza alla legge di Ohm, le resistenze al flusso d’aria sono via via minori con la ramificazione delle vie aeree poiché aumenta la sezione trasversa (condotti in parallelo). La porzione delle vie aeree che oppone la maggiore resistenza al flusso sono le ramificazioni fino all’ottava generazione, con picco alla quarta. Nella respirazione a riposo il 70% delle resistenze si hanno nelle vie aeree superiori fino ai bronchi di medio calibro; solo le cavità nasali offrono il 50% delle resistenze totali. I fattori che determinano la resistenza delle vie aeree sono:
• Volume polmonare. Essendo la resistenza proporzionale alla quarta potenza del raggio, è chiaro che piccole riduzioni del raggio comportano significativi incrementi di resistenza; dunque a CPT la resistenza sarà minima, mentre a VR la resistenza sarà massima.
• SNV: le modificazioni ortosimpatiche (dilatazione) e parasimpatiche (costrizione) del tono della muscolatura liscia dei bronchi e dei bronchioli determinano una variazione del calibro di queste vie aeree e dunque rispettivamente ne diminuiscono o aumentano le resistenze.
Variazioni delle pressioni nel ciclo respiratorio (durata = inspirazione 1,7 s, espirazione 2,3 s)
• Fase precedente l’inspirazione: pressione alveolare = pressione atmosferica, dunque flusso d’aria = 0, muscoli rilasciati, sistema torace-polmone in equilibrio (CFR), pressione endopleurica = -5 cm H2O
• Inspirazione: contrazione del diaframma (aumento del diametro cranio-caudale) e degli intercostali esterni (aumento del diametro antero-posteriore per il movimento delle prime coste, aumento del diametro trasverso per le ultime coste) causa un’espansione della cavità toracica, che trascina con sé anche i polmoni; la pressione endopleurica è ancora più negativa di prima (-6 cm H2O, a causa dell’aumentata tendenza del polmone alla retrazione) e l’endoalveolare è negativa (- 1 cm H2O = - 0,73 mm Hg); la pressione perduttale risulta negativa (gradiente pressorio favorevole in entrata) e il flusso è in entrata; gli alveoli si espandono
• Termine dell’inspirazione: cessazione della contrazione dei muscoli inspiratori; essendo l’aria già presente nell’alveolo e non essendoci ulteriore distensione, il gradiente pressorio si annulla (pressione alveolare = pressione atmosferica = 0); dunque si ha
• Espirazione: in condizioni di riposo, l’espirazione è un processo passivo poiché la forza di retrazione del polmone è sufficiente a ridurre il volume dell’alveolo e dunque ad aumentare la pressione endoalveolare fino a valori positivi (+1 cm H2O); con la retrazione (anche se parziale) del polmone la pressione endopleurica torna a valori di -5 H2O.
Nei casi di atti respiratori massimali e non a volume corrente, l’inspirazione è coadiuvata anche dalla contrazione di muscoli inspiratori accessori quali lo sternocleidomastoideo (solleva sterno e clavicole facendo perno sul processo mastoideo) e gli scaleni (sollevano le prime due coste facendo perno sui processi trasversi delle vertebre cervicali). L’espirazione è normalmente un processo passivo, ma in atti massimali è potenziata dall’azione di muscoli espiratori quali gli intercostali interni e i muscoli addominali (trasverso dell’addome, retto dell’addome, obliquo interno e obliquo esterno; esercitano tutti pressione sui visceri addominali incrementando la spinta che questi esercitano sul diaframma). Relazione flusso/volume del ciclo respiratorio La registrazione simultanea del flusso nelle vie aeree e dei volumi polmonari è di grande utilità clinica. Si collega allo spirometro uno pneumotacografo (flussimetro) in grado di misurare il flusso istantaneo (L/s) che esce o entra dalla bocca del soggetto.
• Espirazione massimale: il picco del flusso (in uscita) si raggiunge attorno al 70-80% della CV. • Inspirazione massimale: il picco del flusso (in entrata) si raggiunge al 20% della CV e si mantiene
tale fino al 75% della CV per poi decrescere rapidamente fino al riempimento completo (100% CV).

Tale andamento della curva flusso/volume si spiega molto facilmente: all’inizio dell’espirazione massimale, a capacità vitale = 100%, l’attività dei muscoli espiratori è elevata, la forza di retrazione del polmone è massima, la pressione endopleurica è oltremodo negativa e le vie aeree essendo distese offrono la minima resistenza al flusso. Nella fase iniziale dell’inspirazione invece, a partire dal volume residuo, la tendenza all’espansione del sistema toraco-polmonare agisce in maniera consensuale all’azione dei muscoli inspiratori; la pressione endopleurica è negativa e lo è anche quella alveolare, per cui la pressione transmurale delle vie intratoraciche è nettamente a favore dell’aumento di calibro. Sul calibro delle vie respiratorie, nel corso dell’inspirazione, agiscono due meccanismi opposti: dilatandosi sempre di più, le resistenze viscose al flusso diminuiscono; di contro il continuo aumento del volume polmonare fa sì che i muscoli inspiratori contratti perdano il vantaggio meccanico in base alla relazione tensione-lunghezza. Il risultato è un flusso elevato e quasi costante per buona parte dell’inspirazione. *porzione sforzo-indipendente: superato, in espirazione, il 50% della CV, la curva flusso-volume si stabilizza a certi valori indipendentemente dallo sforzo espiratorio osservato. Ciò è dovuto alla particolare struttura degli alveoli e delle piccole vie aeree, privi di un’impalcatura propria che li mantenga pervi; accade dunque che durante uno sforzo espiratorio, queste vie riducono il loro calibro, facendo aumentare la velocità del flusso e di conseguenza riducendo la pressione. Ma se la pressione si riduce, viene favorita l’ulteriore compressione degli alveoli e delle piccole vie aeree: si instaura un circolo vizioso per cui anche aumentando lo sforzo espiratorio non si ha un aumento del flusso, anzi questo ne risulta ostacolato dalla progressiva e proporzionale compressione che porta ad un aumento delle resistenze. È come se l’espirazione forzata ostacoli se stessa (compressione dinamica delle vie aeree).
Patologie ostruttive (maggiore resistenza vie aeree): Il soggetto ha difficoltà ad espirare (↑ CPT e ↑ VR). Il flusso espiratorio massimo diminuisce e la parte sforzo dipendente della curva è alterata, perché le vie aeree collassano più facilmente. Tiff ↓ - Motley ↑ (gravi) Patologie restrittive (maggiore resistenza elastica): Il soggetto ha difficoltà ad espandere il polmone (↓ CPT e ↓ VR). Il flusso espiratorio massimo diminuisce perché i volumi raggiunti in inspirazione sono minori, ma la parte sforzo dipendente della curva è praticamente normale.
Asma: patologia ostruttiva, iperreattività bronchiale con broncocostrizione, edema della sottomucosa e ipersecrezione di muco per ipersensibilità di I tipo. Enfisema: patologia ostruttiva, dovuta alla distruzione dei setti interalveolari a causa dell’intervento di proteasi. Aumenta il volume polmonare (raccolte aeree intraparenchimali) e aumenta la compliance. Atelectasia: compattamento del parenchima di un’area del polmone priva d’aria, che non si espande. È solitamente dovuta ad un’ostruzione bronchiale di varia natura (tappi di muco, corpi estranei, tumori). Fibrosi: patologia restrittiva, il parenchima polmonare è sostituito da tessuto fibroso per cui si avrà una distensibilità alterata, minore e dunque una compliance ridotta.

SPIROMETRIA: VOLUMI E CAPACITÀ
Volume/capacità/ventilazione Definizione Corrispondenze
VOLUME CORRENTE Volume di aria che entra ed esce nell’apparato respiratorio ad ogni singolo atto.
VC = 500 mL
VOLUME DI RISERVA
INSPIRATORIA
Volume di aria che può essere introdotto con un’inspirazione forzata massimale effettuata dopo un’inspirazione normale a volume corrente.
VRI = 2,5 L
VOLUME DI RISERVA
ESPIRATORIA
Volume di aria che può essere espulso con un’espirazione forzata massimale effettuata dopo un’espirazione normale a volume corrente.
VRE = 1,5 L
VOLUME RESIDUO Volume di aria presente nell’apparato respiratorio dopo un’espirazione forzata massimale. È il volume di aria sempre presente all’interno dell’apparato che in pratica non può essere mobilizzato.
VR = 1,5 L Si misura, durante la spirometria, col metodo di diluizione con elio. Vi x C i = Vf x Cf
CAPACITÀ VITALE Volume massimo di aria mobilizzabile dall’apparato respiratorio. Equivale al volume di aria inspirato ed espirato con un’ispirazione e un’espirazione massimali (anche in ordine inverso).
CV = VC + VRI + VRE CV = 4,5 L (proporzionale a statura e allenamento, fino a 8 L)
CAPACITÀ POLMONARE
TOTALE
Volume di aria contenuta nel polmone al termine di una massima inspirazione forzata.
CPT = 5-6 L CPT = CV + VR CPT = VC + VRI + VRE + VR
CAPACITÀ FUNZIONALE
RESIDUA
Volume di aria contenuto nei polmoni dopo una normale espirazione. Equivale al volume di aria che non partecipa allo scambio durante una respirazione normale a volume corrente.
CFR = 3 L CFR = VR + VRE
VOLUME ESPIRATORIO
MASSIMO AL PRIMO
SECONDO
Volume d’aria espulso nel primo secondo di un’espirazione forzata e massimale preceduta da una profonda e lenta inspirazione.
VEMS = 80% CV VEMS = 3,6 L
FREQUENZA RESPIRATORIA Numero di atti respiratori al minuto. FR = 10-18 / min
VENTILAZIONE POLMONARE Flusso di aria che entra ed esce dall’apparato respiratorio in un minuto.
VP = 6 - 9 L VP = FR x VC
SPAZIO MORTO ANATOMICO
Volume di aria contenuto nelle zone di conduzione dell’apparato respiratorio che non partecipa agli scambi gassosi.
Vm = 150 mL Se sommiamo anche il volume degli alveoli poco perfusi, si ha lo spazio
morto fisiologico.
VENTILAZIONE DELLO SPAZIO
MORTO
Volume d’aria che entra ed esce nello spazio morto in un minuto.
VSP = FR x Vm VSP = 150 mL x 15/min = 2,25 L/min
VENTILAZIONE ALVEOLARE Volume d’aria che entra ed esce nello spazio alveolare al minuto.
VA = FR x (Vc – Vm) VA = 350 mL x 15/min = 5,25 L/min
INDICE DI TIFFENAU Quota di capacità vitale espirata nel 1° s IT = VEMS / CV(F) ; IT = 0,7 Eventualmente espresso in %
INDICE DI MOTLEY Quota di aria non mobilizzabile sul volume totale di aria dopo inspirazione massima.
IM = VR / CPT ; IM =< 0,3 Eventualmente espresso in %

SCAMBI E TRASPORTO DEI GAS RESPIRATORI
SCAMBI GASSOSI ALVEOLOCAPILLARI
• Leggi fondamentali e composizione dell’aria I gas respiratori, ovvero ossigeno, anidride carbonica e azoto, seguono l’equazione di stato dei gas, espressa da PV = nRT. Secondo la legge di Dalton, un gas in una miscela esercita una pressione parziale che è proporzionale alla sua concentrazione nella miscela; questa pressione equivale a quella che eserciterebbe se nello stesso volume fosse presente solo quel determinato gas. La somma delle diverse pressioni parziali dei componenti della miscela è uguale alla pressione totale. Pgas = Ptot x Fgas. Secondo la legge di Henry, la quantità di gas disciolto in un liquido è proporzionale alla pressione parziale del gas in equilibrio con la fase liquida. La proporzione tra quantità disciolta e pressione parziale è espressa dal coefficiente di solubilità α. Esso è inversamente proporzionale alla temperatura ed è pari a 0 alla temperatura di ebollizione del liquido.
A 37°C, αC02 = 0,032 - α02 = 0,0013 - αN2 = 0,0007. Secondo la legge di diffusione di Graham, la diffusione di un gas da una miscela gassosa alla fase liquida è inversamente proporzionale alla radice quadrata del peso molecolare del gas considerato. Dunque la velocità di diffusione di un gas in un liquido è direttamente proporzionale al coefficiente di solubilità del gas ed inversamente proporzionale alla radice quadrata del PM del gas. Ne risulta che la C02 è 24 volte più solubile dell’02 e diffonde circa 20 volte più velocemente. L’aria atmosferica comprende il 20,95% di 02, il 79,01% di N2 e 0,04% di C02. È l’aria che viene inspirata. L’aria alveolare, che ha viaggiato nelle vie aeree, è stata riscaldata a 37°C ed è stata saturata di vapore acqueo. In queste condizioni, la pressione totale della miscela gassosa è sempre 760 mm Hg, ma dobbiamo sottrarvi il valore di pressione parziale esercitata dal vapore acqueo, ovvero 47 mm Hg. Avremo dunque 713 mm Hg dovuti ad ossigeno, azoto e anidride carbonica; su questo valore baseremo i calcoli delle pressioni parziali all’interno dell’alveolo. Le percentuali dei gas respiratori nell’aria alveolare sono diverse da quelle dell’aria atmosferica: infatti l’ossigeno diffonde nel sangue dei capillari polmonari mentre l’anidride carbonica viene dal sangue rilasciata nell’aria alveolare. Avremo dunque il 15% di 02 e il 5% di C02; N2 sarà circa l’80%. La composizione dell’aria alveolare non è una costante fisiologica, ma varia secondo il ciclo respiratorio e la localizzazione polmonare. L’aria alveolare è continuamente mescolata con l’aria appena inspirata, e parte della miscela risultante viene espulsa durante l’espirazione. L’aria espirata ha composizione peculiare, con maggiore quantità d’ossigeno e minore di anidride carbonica rispetto all’aria alveolare, poiché, a volume corrente, 350 ml provengono dagli alveoli mentre 150 ml provengono dallo spazio morto (aria che non partecipa agli scambi gassosi). I gas alveolari possono essere valutati con degli analizzatori rapidi, alla bocca del soggetto. All’inizio avremo l’aria espirata dello spazio morto, con composizione sostanzialmente analoga a quella dell’aria inspirata. In seguito avremo l’espirazione dell’aria alveolare, con P02 di 100 mm Hg e PC02 di 40 mm Hg.
Riepilogo pressioni totali/parziali e composizione percentuale: Pressione
totale P parziale ossigeno
P parziale C02 P parziale N2 P parziale H20
Aria inspirata 760 mm Hg 158 (21%) 0,3 (0,04%) 596 (79%) 5,7 Aria alveolare 760 mm Hg 100 (15%) 40 (5%) 573 (80%) 47 (a 37°C) Aria espirata 760 mm Hg 116 32 565 47 Arterie polmonari (sangue venoso misto)
/// 40 46 573 47
Vene polmonari (sangue arterioso)
/// 95 40 573 47
Per l’aria alveolare, considerare 713 mm Hg come Ptot per ossigeno, anidride carbonica e azoto. Si noti come il sangue arterioso sia in equilibrio con l’aria alveolare, ad eccezione dell’ossigeno (a causa degli shunt anatomici e della disomogenea distribuzione del rapporto ventilazione/perfusione).

• Scambi attraverso la membrana alveolocapillare La membrana è costituita da uno strato liquido (con il surfattante) che riveste l’alveolo, l’epitelio alveolare (pneumociti I e II tipo), la membrana basale dell’epitelio alveolare, la membrana basale endoteliale (spesso fusa con quella dell’epitelio) e l’endotelio capillare. È mediamente spessa 0,5 µ. I capillari ricoprono gli alveoli come una rete; gli scambi gassosi tra aria alveolare e sangue avvengono per diffusione; è un processo spontaneo, senza alcun consumo di energia. I gas passano da un compartimento a pressione parziale maggiore ad un altro in cui la pressione parziale è minore. Seguono dei gradienti pressori di diffusione. Il movimento dei gas, oltre a seguire i gradienti pressori, è determinato dalla solubilità del gas nel liquido, dal peso molecolare e dalla natura chimica della membrana.
È regolato dalla legge di Fick per la diffusione: ����� = ���� ∙
� ∙ � � − ��. V è il volume di gas che
diffonde nell’unità di tempo, D il coefficiente di diffusione del gas, d lo spessore della membrana, S l’area di scambio (superficie della membrana), P1-P2 è la differenza di pressione parziale dei gas ai due lati della membrana. Dunque sono molteplici i fattori che determinano le quantità di ossigeno e anidride carbonica che si trasferiscono nell’unità di tempo; a questi fattori presenti nella legge di Fick vanno aggiunti il tempo di contatto tra i due compartimenti (tempo di permanenza del sangue) e il volume di sangue esposto sulla superficie di scambio. Il tempo in cui un eritrocita attraversa i capillari polmonari è normalmente 0,75 s; questo intervallo è fisiologicamente ridondante, in quanto normalmente le pressioni parziali di ossigeno ai due lati della membrana alveolocapillare si equilibrano in soli 0,25-0,35 secondi. La quantità di sangue di norma presente nei soli capillari polmonari è circa 60-140 ml a riposo, fino a 1 litro durante esercizio fisico. Nel momento in cui il sangue viene a trovarsi nel capillare polmonare, affacciato alla membrana alveolocapillare, inizia l’equilibrazione secondo il gradiente pressorio per diffusione passiva. La P02 del sangue venoso dell’arteria polmonare è 40 mm Hg a riposo (dunque con possibili variazioni a seconda dello stato di attività metabolica dei vari distretti corporei). La P02 alveolare è invece 100 mm Hg; mediamente vi è un gradiente pressorio favorevole di circa 60 mm Hg. A causa degli shunt anatomici (vene bronchiali che confluiscono nelle vene polmonari e vasi di Tebesio che sfociano nel ventricolo sinistro) vi è una naturale commistione del sangue arterioso/venoso; inoltre vi è anche un diverso rapporto ventilazione/perfusione in ogni porzione dei polmoni: per questi due fattori la P02 arteriosa non è esattamente uguale a quella alveolare ma circa 5-10 mm Hg inferiore. La costante di diffusione della C02 è di molto superiore alla costante di diffusione dell’02 per cui l’equilibrazione tra le PCO2 ai lati della membrana di scambio, nonostante il gradiente sia solo 6 mm Hg, è molto veloce (0,15 s) e sempre presente (a differenza dell’equilibrazione dell’ossigeno che può, in casi di diminuzione della superficie di scambio per senescenza o di ispessimento della membrana alveolocapillare per ipertensione polmonare, essere deficitaria persino negli 0,75 s di transito del sangue). Solo in casi gravissimi di notevole ispessimento patologico della membrana l’equilibrazione dell’anidride carbonica può risultare incompleta. TRASPORTO DEI GAS NEL SANGUE Nel sangue l’ossigeno e l’anidride carbonica possono essere trasportati in soluzione fisica, legati a molecole trasportatrici o in forma combinata. È importante sottolineare che ciascuna molecola di ossigeno e anidride carbonica scambiata a livello alveolare o tissutale deve passare prima in forma disciolta in soluzione fisica prima di legarsi a proteine plasmatiche o acqua. Inoltre sono queste molecole in soluzione fisica che giocano un ruolo determinante nella regolazione chimica della ventilazione.
• Trasporto dell’ossigeno A causa della sua bassa solubilità, solo il 2% dell’ossigeno è disciolto nel plasma e negli eritrociti; il 98% è trasportato dall’emoglobina. L’emoglobina (Hb) è una cromoproteina costituita da 4 catene polipeptidiche, 2 α e 2 β, che presentano ognuna un gruppo eme. Questi gruppi eme sono anelli tetrapirrolici con al centro uno ione ferroso (Fe2+). È proprio l’atomo di ferro che media il legame reversibile con l’02. 1 grammo di Hb può trasportare fino a 1,36 ml di 02. Essendo la concentrazione ematica media di emoglobina circa 15 g/100 ml, in 100 ml di sangue possono essere trasportati 20 ml di 02. L’ossigeno si lega velocemente e reversibilmente all’emoglobina secondo la reazione 4 02 + Hb <-> Hb(02)4. La quantità di Hb02 (ossiemoglobina) formata è funzione della P02 nel sangue. Nei capillari polmonari la reazione è spostata verso destra e l’emoglobina viene caricata di ossigeno; al contrario nei capillari

tissutali la reazione è spostata verso sinistra e l’ossigeno si scinde dall’emoglobina e può diffondere verso i tessuti. Il comportamento dell’Hb a diverse pressioni parziali di ossigeno è descritto dalla curva di saturazione dell’emoglobina. Essa presenta un andamento sigmoide; l’affinità per l’ossigeno aumenta con l’aumentare della P02. Si pensa che questo fenomeno sia dovuto al fatto che l’ossigenazione di un gruppo eme influenzi quello dell’eme successivo. Nel sangue arterioso la saturazione dell’Hb è circa 97% mentre in quello venoso è 72-73%. Più fattori influenzano l’affinità tra Hb e 02 e di conseguenza la curva di saturazione. Nello specifico, l’aumento della temperatura, della concentrazione di 2,3-bifosfoglicerato (2,3-BFG), della concentrazione di anidride carbonica e la diminuzione del pH diminuiscono l’affinità tra Hb e ossigeno e spostano la curva di saturazione verso sinistra (a parità di pressione parziale dell’ossigeno si ha meno Hb legata). Il cambiamento di affinità causato da aumento della [C02] e diminuzione del pH prende il nome di effetto Bohr. Uno spostamento a destra della curva si osserva nei capillari tissutali, in cui con l’aumento della [C02] e diminuzione del pH l’Hb può liberare grandi quantità di 02 anche senza grandi variazioni della PO2. Nel polmone invece si osserva uno spostamento a sinistra della curva, perché il pH aumenta e diminuisce la [C02] permettendo l’aumento di affinità dell’Hb per l’02; si ha in questo modo il “caricamento” della molecola. Si definisce P50 il valore di P02 per cui il 50% dell’Hb è ossigenata; normalmente equivale a 27 mm Hg.
• Trasporto dell’anidride carbonica Nel sangue l’anidride carbonica viene veicolata in 3 forme diverse:
o Disciolta in soluzione fisica (7%) o Come ione bicarbonato (70%). La C02 reagisce con l’acqua formando acido carbonico,
specialmente nell’eritrocita in cui è presente l’enzima anidrasi carbonica; l’acido carbonico si dissocia velocemente in idrogenione + ione bicarbonato. Lo ione bicarbonato per gradiente chimico diffonde nel plasma ma per preservare il gradiente elettrico viene scambiato con ioni cloruro (scambiatore di anioni/shift dei cloruri).
o Come composto carbaminico (23%). La C02 può legarsi principalmente all’Hb ma anche ad altre proteine plasmatiche attraverso una reazione coi loro gruppi amminici liberi (-NH2) per formare un gruppo carbaminico terminale (-NHCOOH).
Nel sangue arterioso la concentrazione di C02 è circa 48 ml/100 ml; in quello venoso circa 52 ml/100 ml. La curva di equilibrio della C02 è lineare e ripida: l’anidride carbonica può essere ceduta o acquisita facilmente anche per piccole variazioni della PCO2. È da sottolineare come, similmente all’affinità tra Hb e 02, l’affinità del sangue per la C02 possa cambiare: per l’effetto Haldane, alte pressioni parziali di 02 diminuiscono l’affinità del sangue per la C02. Ciò assume grande importanza nei capillari perialveolari, in cui la PO2 è alta: la minore affinità permette alla C02 di diffondere nell’aria alveolare. Al contrario nei tessuti, con la caduta della PO2, il sangue aumenta l’affinità per la C02 e così quest’ultima diffonde nel plasma e negli eritrociti. RESPIRAZIONE TISSUTALE Per respirazione tissutale s’intende lo scambio di gas fra cellule e ambiente circostante. Il limite al metabolismo ossidativo dei tessuti è dato più dall’approvvigionamento di ossigeno che dalla rimozione di anidride carbonica. A livello tissutale gli scambi gassosi sono governati dagli stessi principi che li governano nel polmone. Il sangue arterioso, con PO2 di 95 mm Hg e PCO2 di 40 mm Hg, diventa sangue venoso equilibrandosi con i tessuti e raggiungendo quindi una PO2 di 40 mm Hg e una PCO2 di 46 mm Hg. L’anidride carbonica diffonde con molta più facilità: il fattore limitante degli scambi gassosi è infatti la diffusione dell’ossigeno. Quest’ultima è però assicurata da una vastissima superficie di scambio (1000 m2) e da una piccola distanza di diffusione (10-25 µ).

RAPPORTO VENTILAZIONE/PERFUSIONE Nel polmone la ventilazione alveolare e il flusso sanguigno capillare non sono distribuiti in maniera omogenea: esistono aree di parenchima ben ventilate ma poco perfuse, come aree poco ventilate ma perfettamente irrorate. Ciò è molto più accentuato in molte malattie polmonari. L’indice che tiene conto della ventilazione alveolare e dell’irrorazione polmonare è il rapporto ventilazione/perfusione. Per ventilazione intendiamo il flusso di aria negli alveoli in un minuto, per perfusione il flusso ematico nei capillari perialveolari sempre in un minuto (corrisponde alla gittata cardiaca). In condizioni di riposo la ventilazione alveolare è di circa 4 L di aria al minuto, mentre la perfusione corrisponde a 5 L di sangue al minuto: il rapporto è 4/5 = 0,8. La ventilazione, per motivi dipendenti dalla diversa distensibilità degli alveoli, è maggiore alla base che non all’apice dei polmoni. Il flusso ematico polmonare, per motivi gravitazionali, è anch’essa maggiore alla base che non all’apice. L’entità della variazione è però maggiore nel flusso ematico; ne deriva che il rapporto ventilazione/perfusione aumenta progressivamente dalla base all’apice dei polmoni.

CONTROLLO DELLA RESPIRAZIONE
I meccanismi di controllo della ventilazione devono svolgere essenzialmente due compiti: fornire lo schema motorio per i muscoli respiratori e assecondare le richieste metaboliche dell’intero organismo. CONTROLLO NERVOSO DELLA RESPIRAZIONE I muscoli respiratori sono innervati da motoneuroni situati nel midollo cervicale (nervi cervicali e nervo frenico, C3-C8), toracico (nervi intercostali) e lombare (segmento L1: nervi ileoipogastrico e ileoinguinale per i muscoli addominali). Questi motoneuroni ricevono afferenze da fibre che provengono dal tronco encefalico, nello specifico da neuroni dei circuiti respiratori. Si individuano 3 fasi di attività neuronale nella respirazione: fase I (inspirazione), fase PI (postinspirazione), fase E2 (espirazione). Circuito respiratorio troncoencefalico Si distinguono 2 strutture, entrambe a livello bulbare:
• Gruppo respiratorio ventrale (GRV), vicino al nucleo ambiguo. È costituito da neuroni di più tipi, ovvero sia inspiratori che espiratori. Comprende il complesso preBötzinger.
• Gruppo respiratorio dorsale (GRD), nella regione del NTS. È costituito da neuroni ad attività inspiratoria.
Sulla base del pattern di scarica in relazione all’attività del nervo frenico, si sono individuati diversi tipi di neuroni:
• Inspiratori precoci. Hanno una rapida depolarizzazione con diversi potenziali d’azione all’inizio o poco prima dell’attività del nervo frenico. La frequenza di scarica poi decresce ed è poi nulla nella seconda fase dell’inspirazione e nelle fasi PI e E2.
• Inspiratori (a rampa). Mantengono la depolarizzazione per tutta l’inspirazione. Sono poi inibiti durante PI e E2.
• Inspiratori tardivi. Cominciano a scaricare solo nella seconda metà della fase I e sono inibiti durante PI e E2.
• Postinspiratori. Scaricano dopo la fine della fase I, diminuiscono la loro frequenza nella seconda parte di PI e sono inibiti in E2.
• Espiratori (a rampa). Danno pochi potenziali durante PI, vengono inibiti e poi la loro attività raggiunge il massimo durante E2. Vengono poi inibiti.
• Preinspiratori. Iniziano a depolarizzarsi alla fine di E2 e continuano a scaricare fino all’inizio di I. Ritmogenesi respiratoria La ritmogenesi segue questa sequenza:
• Attivazione dell’inspirazione • Genesi del pattern inspiratorio • Inattivazione reversibile dell’inspirazione • Inattivazione irreversibile dell’inspirazione • Attivazione dell’espirazione • Inattivazione dell’espirazione.
In base a diverse evidenze sperimentali, vi sono differenti modelli interpretativi per spiegare la sequenza di questi eventi. Secondo il modello a rete, le connessioni eccitatorie (glutammatergiche) e inibitorie (GABAergiche e glicinergiche) tra i diversi nuclei (il circuito di base è considerato essere il complesso preBötzinger nel GRV) determinano, a partire da variazioni di probabili stimolazioni toniche dall’esterno (ad esempio chemocettori centrali o periferici), l’esatta sequenza di eventi respiratori. Secondo il modello a segnapassi, vi sarebbero neuroni pacemaker che per caratteristiche intrinseche e peculiari della membrana avrebbero una capacità di depolarizzazione ritmica. Neuroni di questo tipo sono stati identificati nel nucleo dell’ipoglosso (ad attività modulatoria sulle efferenze motorie respiratorie) e nel complesso preBötzinger. Vi sono infine modelli ibridi che cercano di integrare tra modelli a cellule segnapassi e modelli basati sulle connessioni sinaptiche tra differenti popolazioni di neuroni. Attualmente è evidente che le strutture con cellule segnapassi siano comprese in circuiti neurali che possono ampiamente influenzare i meccanismi generatori di base.

Qualunque sia il modello che si consideri, lo schema motorio determinato dalla ritmogenesi centrale del respiro è il seguente:
• Inspirazione – Attività “a rampa” dei motoneuroni che innervano i muscoli inspiratori, con graduale aumento dell’intensità della contrazione di diaframma e intercostali esterni.
• Postinspirazione – Diminuisce il grado di contrazione dei muscoli inspiratori e inizia la fase di “espirazione passiva”, poiché non vi è contrazione dei muscoli espiratori ma semplicemente viene utilizzato il lavoro del ritorno elastico del sistema toraco-polmonare.
• Espirazione attiva – I muscoli intercostali interni e addominali si contraggono per ridurre il volume della gabbia toracica.
Controllo esercitato dai centri pontini I centri bulbari risentono di un controllo di centri più rostrali del tronco cerebrale:
• Centro apneustico. Situato nella parte caudale del ponte, sotto stimolo determina profonde inspirazioni intervallate da brevi espirazioni. Questo tipo di respiro viene definito apneusi.
• Centro pneumotassico. Probabilmente corrispondente ai nuclei parabrachiale mediale e di Köllicher-Fuse (ponte rostrale). Inibisce l’apneusi e si pensa controlli soprattutto la profondità del respiro sulla base delle variazioni della PCO2.
CONTROLLO CHIMICO DELLA RESPIRAZIONE Grazie a recettori centrali e periferici, variazioni dei valori ematici di PCO2, PO2 e pH regolano l’azione dei centri respiratori troncoencefalici sulla frequenza e la profondità della ventilazione. Chemocettori centrali Nel bulbo si distinguono 2 principali zone bilaterali in area ventrolaterale: l’area rostrale, subito al di sotto della giunzione bulbopontina, e l’area caudale, mediale rispetto all’emergenza del nervo ipoglosso. Le due aree proiettano poi ad una zona intermedia in cui vengono integrate le informazioni con altre afferenze neurovegetative. Le cellule di queste aree sono estremamente sensibili all’aumento della PCO2 (ipercapnia), che si manifesta nell’ambiente extracellulare come acidosi respiratoria (aumento pH). Piccolissime variazioni (3-5 mm Hg) della PCO2 attivano i chemocettori centrali. Si pensa che questi chemocettori rispondano dunque ad un solo stimolo, ovvero alla concentrazione idrogenionica nel liquor (che è funzione della PCO2 delle arterie cerebrali). Infatti la CO2 diffonde facilmente attraverso la barriera ematoencefalica ed ematoliquorale; di contro, gli ioni bicarbonato diffondono più difficilmente. Dunque la scarsità di bicarbonato e l’assenza di emoglobina determinano la minore capacità tampone del liquor rispetto al sangue. I chemocettori centrali si trovano in una regione in cui il pH è influenzato dalla PCO2 ematica ma è diverso (più basso) dal pH arterioso. Chemocettori periferici A differenza di quelli centrali, i chemocettori periferici modificano la ventilazione in base a variazione di PO2, PCO2 e pH nel sangue. I chemocettori periferici si trovano nei glomi aortici e carotidei. I glomi aortici sono irrorati da collaterali dell’aorta e dalla succlavia destra; i glomi carotidei sono irrorati da collaterali dell’arteria occipitale. I capillari dei glomi sono fenestrati. Per innescare una risposta iperventilatoria, l’ipossia registrata dai chemocettori periferici deve essere abbastanza grave (PO2 sotto 50-60 mm Hg). Le cellule dei glomi hanno origine neuroectodermica, hanno un alto indice metabolico e il loro flusso ematico è estremamente elevato. Sono cellule dopaminergiche. I chemocettori aortici inviano afferenze nella compagine del nervo vago, quelli carotidei nel contesto del nervo glossofaringeo. Entrambe le afferenze terminano nel NTS (che fa parte del GRD). Sembra che gli stimoli causati da ipossia consistano nell’inibizione di canali del potassio (la normale attività di questi canali dovrebbe essere legata alla presenza di quantità fisiologiche di ossigeno) a cui segue una depolarizzazione della membrana, apertura dei canali voltaggio-dipendenti del calcio e in seguito liberazione di dopamina. Gli stimoli invece dell’acidosi e dell’ipercapnia consisterebbero nell’aumento della concentrazione idrogenionica all’interno delle cellule glomiche, che determinerebbe un incremento dell’attività dello scambiatore Na+/H+ con introito di sodio ed estrusione di idrogenioni. Di riflesso si avrebbe un inversione dell’attività dello scambiatore 3 Na+/Ca2+ con estrusione di sodio e ingresso di calcio (a concentrazioni normali, questo scambiatore estrude calcio e introduce sodio, sfruttando il gradiente del sodio), aumento di concentrazione del calcio e avvio dell’esocitosi sinaptica con liberazione di dopamina.

INFLUENZE NON CHIMICHE SULLA RESPIRAZIONE Sistemi a feedback
• Stiramento. Recettori da stiramento (tensocettori), ad adattamento lento, misurano il grado d’insufflazione del polmone; le fibre Aα e Aβ afferenti determinano un riflesso di inibizione dell’inspirazione. Sono responsabili del riflesso inibitorio di Hering-Breuer. Durante la desufflazione i recettori da stiramento non sono stimolati e viene meno il loro effetto inibitorio sul centro inspiratorio troncoencefalico (riflesso eccitatorio di Hering-Breuer). Il riflesso scompare in seguito a vagotomia; nell’eupnea dell’uomo tale riflesso riveste un’importanza modesta (è evocato per valori di volume corrente superiori a 800 ml).
• Stimoli irritativi. I recettori che rispondo a stimoli irritativi sono presenti nel polmone e a diversi livelli delle vie aeree; a questo tipo di recettori appartengono recettori delle vie aeree superiori sensibili alla polvere (che innescano il riflesso della tosse) e recettori dell’albero bronchiale sensibili a irritanti chimici.
• Variazioni della perfusione polmonare. I recettori J, presenti nel parenchima polmonare tra i capillari e gli alveoli, rispondono a stimoli chimici e farmacologici e a variazioni della perfusione polmonare. Determinano un’iniziale apnea seguita da un respiro rapido e superficiale.
Sistemi non a feedback
• Afferenze dai centri nervosi superiori (centri corticali volontari, sistema limbico e ipotalamo in seguito a forti emozioni di paura, dolore o gioia)
• Afferenza della mucosa bronchiale (riflesso di deflazione di Head, per cui all’aumento della frequenza respiratoria si ha una diminuzione del volume polmonare)
• Afferenze dalle fibre C • Afferenze da pressocettori (alta pressione conduce a ipoventilazione, bassa pressione a
iperventilazione) • Dolore (risposta iperventilatoria) • Recettori per il caldo e per il freddo (periferici e centrali; il calore stimola l’iperventilazione) • Influenze ormonali (catecolamine e progesterone determinano iperventilazione) • Sonno (in fase NREM la regolazione della respirazione dipende solamente dalla regolazione
metabolica)