di Nadrian C. Seemandownload.kataweb.it/mediaweb/pdf/espresso/scienze/2004_431_8.pdf · tificata da...
5
Il DNA non è solamente il segreto della vita: è anche un componente versatile per la costruzione di strutture e dispositivi grandi un miliardesimo di metro di Nadrian C. Seeman el 2003 è stato celebrato il cinquan- tesimo anniversario della scoperta della struttura a doppia elica del DNA da parte di James D. Watson e Francis H. Crick, una scoperta che ha messo in evidenza le basi chimiche della genetica e ha po- sto le premesse per il successivo mezzo secolo di ricerca biologica. Oggi migliaia di scienziati lavorano per decifrare gli innumerevoli modi con cui i geni controllano lo svilup- po e il funzionamento degli organismi viventi. Tutti questi geni sono scritti su un supporto di DNA. Eppure questa straordinaria molecola ha altre applicazioni oltre a quelle biochimiche. Impiegando i meto- di delle moderne biotecnologie, possiamo costruire lunghe moleco- le di DNA con una sequenza di componenti scelta a piacere, imboc- cando così percorsi nuovi, mai seguiti dalla natura nel corso dell'e- voluzione biologica. Nel 1994, per esempio, Leonard M. Adleman della University of Southern California dimostrò in che modo si pos- sa sfruttare il DNA per realizzare un particolare tipo di computer. In questo articolo illustrerò un altro impiego non biologico del DNA, os- sia la costruzione di strutture e dispositivi i cui elementi e mecca- nismi essenziali hanno dimensioni variabili da 1 a 100 nanometri: in una parola, nanotecnologia. FILAMENTI DI DNA SI AUTOASSEMBLANO in una struttura complessa quando le loro sequenze di basi sono progettate in modo da appaiarsi a specifici filamenti complementari. Qui si è formato un modello a bastoncini di un ottaedro tronco, che ha sei facce quadrate e otto facce esagonali. Gli spigoli hanno una lunghezza di circa 20 nanometri. Un breve «spillo» di DNA sporge da ciascun vertice. Queste prominenze potrebbero essere modificate per poter legare insieme diversi ottaedri tronchi e formare una struttura tridimensionale regolare. q ( (
Transcript of di Nadrian C. Seemandownload.kataweb.it/mediaweb/pdf/espresso/scienze/2004_431_8.pdf · tificata da...

Il DNA non è solamente il segreto
della vita: è anche un componente
versatile per la costruzione
di strutture e dispositivi grandi
un miliardesimo di metro
di Nadrian C. Seeman
el 2003 è stato celebrato il cinquan-
tesimo anniversario della scoperta della
struttura a doppia elica del DNA da parte
di James D. Watson e Francis H. Crick,
una scoperta che ha messo in evidenza
le basi chimiche della genetica e ha po-
sto le premesse per il successivo mezzo
secolo di ricerca biologica. Oggi migliaia di scienziati lavorano per
decifrare gli innumerevoli modi con cui i geni controllano lo svilup-
po e il funzionamento degli organismi viventi. Tutti questi geni sono
scritti su un supporto di DNA. Eppure questa straordinaria molecola
ha altre applicazioni oltre a quelle biochimiche. Impiegando i meto-
di delle moderne biotecnologie, possiamo costruire lunghe moleco-
le di DNA con una sequenza di componenti scelta a piacere, imboc-
cando così percorsi nuovi, mai seguiti dalla natura nel corso dell'e-
voluzione biologica. Nel 1994, per esempio, Leonard M. Adleman
della University of Southern California dimostrò in che modo si pos-
sa sfruttare il DNA per realizzare un particolare tipo di computer. In
questo articolo illustrerò un altro impiego non biologico del DNA, os-
sia la costruzione di strutture e dispositivi i cui elementi e mecca-
nismi essenziali hanno dimensioni variabili da 1 a 100 nanometri:
in una parola, nanotecnologia.
FILAMENTI DI DNA SI AUTOASSEMBLANO in una struttura complessa
quando le loro sequenze di basi sono progettate in modo da appaiarsi a
specifici filamenti complementari. Qui si è formato un modello a
bastoncini di un ottaedro tronco, che ha sei facce quadrate e otto facce
esagonali. Gli spigoli hanno una lunghezza di circa 20 nanometri. Un
breve «spillo» di DNA sporge da ciascun vertice. Queste prominenze
potrebbero essere modificate per poter legare insieme diversi ottaedri
tronchi e formare una struttura tridimensionale regolare.
q
(
(

LA STRUTTURA DEL DNA
I l DNA è costituito da due
catene di molecole di
zucchero e gruppi fosfato,
tenute insieme da «ponti»
formati da coppie
complementari di basi [A e
T, Ce 6] unite da legami
deboli [a sinistra). La forma
più comune è il B-DNA (al
centro], che si awolge in
una doppia elica destrorsa
di circa due nanometri di
diametro. Un awolgimento
completo dell'elica è lungo
circa 3,5 nanometri, ossia
10-10,5 coppie di basi. In
circostanze speciali il DNA
può formare una doppia
elica sinistrorsa, chiamata
Z-DNA (a destra].
B-DNA Z-DNAGruppo fosfato
Molecola di zucchero(desossiribosio) Nucleotide
2,0 nanometri
Basi
Elica
l_ Elicadestrorsa
T
Legami debolifra le basi
Scheletrozucchero-fosfato
Scheletro zucchero-fosfato
Basi
CGCGTAATATCGGC
4's(v11141Wri v11)er i t› i vCACGCATGTGCGTA
eg,
gy,e
C
ATGCGTG CACGTACGCAC GT GC
CG C.TATAAT
A T
GCGCTA
CGTGGCAC
GCTAATCGCG
ATAT
T A
CGGC
CGGCCGATATTACGCG
GCACGCAT GACTACCGCG TG CGTA CTGATGGC
1
TA
TAC GA TA T•T A•G CC G
L'AUTOASSEMBLAGGIO DI STRUTTURE DI DNA è possibile perché i filamenti con
sequenze di basi complementari tendono a legarsi l'uno all'altro formando una doppia
elica. Le estremità coesive (a), brevi tratti di filamenti spaiati che sporgono a
un'estremità della molecola, possono congiungere unità specifiche. Un secondocomponente chiave è il DNA ramificato (b) , in cui tre o più eliche si uniscono in un
punto. In natura, questo punto può spostarsi (c), perché le sequenze di basi sui
quattro bracci sono simmetriche. Il DNA ramificato artificiale, privo di questa
simmetria, ha un punto di diramazione fisso (d). Copie di DNA ramificato con
estremità coesive complementari (e) si autoassemblano in una struttura a reticolo.
Strutture di questo genere hanno molte potenziali applica-zioni. Reticoli regolari costituiti da DNA potrebbero conteneremacromolecole biologiche in una matrice ordinata, adatta alladeterminazione strutturale per mezzo della cristallografia con iraggi X: un passo importante nella progettazione «razionale» difarmaci. In alternativa, i reticoli potrebbero servire da impalca-ture per componenti nanoelettronici, che potrebbero essere siadispositivi funzionanti sia fasi intermedie nella costruzione diun altro strumento. Si potrebbero realizzare materiali - costitui-ti da DNA o sintetizzati tramite esso - con strutture esattamentepianificate a livello molecolare. Macchine di DNA con parti inmovimento potrebbero essere usate come sensori nanomeccani-ci, interruttori e pinzette, o svolgere funzioni robotiche anchepiù complesse.
DNA ramificato
La nanoscala è la scala delle molecole. Un tipico legame inte-ratomico ha una lunghezza di circa 0,15 nanometri (un nano-metro è un miliardesimo di metro). Il diametro dell'elica delDNA è di circa due nanometri, ed essa descrive un avvolgimen-
Nel DNA ricombinante, il punto di ramificazione cade doveciascuno dei quattro filamenti si scambia il compagno. Il puntodi ramificazione si sposta a causa della particolare simmetria del-le sequenze di basi che lo fiancheggiano. Questa simmetria im-plica che ciascun filamento si possa appaiare con l'uno o l'altrodi altri due filamenti. Nel 1979, mentre lavoravo con Bruce H.Robinson, ora all'Università di Washington, per descrivere lanatura di questo spostamento, mi resi conto che molecole sinte-tiche di DNA prive di simmetria bilaterale potevano formaremolecole ramificate con punti di biforcazione fissi. Per costruireuna simile intersezione, si dovrebbero sintetizzare quattro fila-menti di DNA. Per ciascuno di essi, la sequenza lungo una metàdel filamento dovrebbe corrispondere a quella di metà di un se-condo filamento e la rimanente metà del primo corrispondereb-be a metà di un terzo filamento (si veda l'illustrazione in bassonella pagina a fronte).
La struttura preferita del DNA è la classica doppia elica iden-tificata da Watson e Crick. Ciò che determina questa preferenzaè una grandezza che prende il nome di energia libera. In gene-rale, quest'ultima stabilisce se una reazione chimica procede inuna direzione o nell'altra, e definisce anche la conformazione -
I filamenti di DNA interagiscono in modi facilmente programmabili.
La loro variabilità si presta perfettamente alla
to completo più o meno ogni 3,5 nanometri: una distanza pari acirca 10 coppie delle basi che costituiscono i «pioli» della scaladel DNA (si veda l'illustrazione nella pagina a fronte). Un brevesegmento di DNA può interagire in modo altamente specificocon altre sostanze, a seconda della sequenza delle sue coppie dibasi. Si può immaginare di utilizzare simili segmenti come son-de per riconoscere particolari molecole o come catalizzatori percontrollare la composizione di un materiale. Da molti anni i bio-logi impegnati nell'ingegneria genetica sfruttano le proprietà diriconoscimento del DNA, e soprattutto le sue «estremità coesive»(in inglese, sticky ends). Quando un filamento della doppia elicasporge, con diverse basi spaiate, al di là del filamento comple-mentare, allora si forma un'estremità coesiva (si veda l'illustra-zione in basso nella pagina a fronte). Questo strano nome indicala tendenza del filamento più lungo a legarsi a un filamentocompatibile, che possiede cioè le basi complementari nell'ordinecorretto: l'adenina di un filamento si appaia con la timina del-l'altro, e la citosina si lega con la guanina.
A prima vista, non si direbbe che il DNA possa generarestrutture interessanti. In natura esso forma una catena lineare,come un lungo pezzo di spago, e quindi si potrebbe pensare chese ne possano ricavare solo segmenti o cerchi, tutt'al più defor-mati o annodati in qualche modo. Ma la catena lineare non èl'unica forma possibile per il DNA. In alcuni processi cellulari,esso esiste per breve tempo come molecola ramificata. Ciò acca-de durante la duplicazione del DNA (in preparazione della divi-sione cellulare) e la ricombinazione (il processo in cui materialegenetico è scambiato fra coppie corrispondenti di cromosomi,come accade quando vengono prodotti spermatozoi e ovociti).
Le «ramificazioni» si formano quando la doppia elica si svol-ge parzialmente in due filamenti. Nella duplicazione, da ciascu-no di essi si ottiene una nuova doppia elica aggiungendo nu-cleotidi complementari lungo tutto il filamento. (Un nucleotide èla combinazione di una base con la sezione corrispondente del-lo scheletro dell'elica.) Più interessante è ciò che avviene nellaricombinazione, allorché due segmenti di DNA si spezzano e sisvolgono parzialmente, e i quattro filamenti che ne risultano siuniscono in una forma che ricorda molto un incrocio stradale.
IN SINTESI
• ll DNA è una molecola ideale per la costruzione di strutture
a scala nanometrica. Si possono programmare filamenti
di DNA perché si autoassemblino secondo schemi complessi
sintetizzandoli grazie alla combinazione di basi
complementari, che si legano preferenzialmente e formano
tratti di doppia elica.
• Impalcature di DNA potrebbero trattenere molecole
«ospiti» in reticoli ordinati per le analisi cristallografiche.
Potrebbero anche includere dispositivi elettronici di
dimensioni molecolari, oppure essere utilizzate per la
costruzione di materiali con precise configurazioni
molecolari.
• Macchine di DNA a scala nanometrica possono funzionare
grazie al passaggio da una conformazione a un'altra di parti
della loro struttura. Queste transizioni possono essere
controllate con metodi chimici o con l'uso di speciali filamenti
di DNA.
gli avvolgimenti e le biforcazioni - di macromolecole come ilDNA, l'RNA e le proteine. Un sistema chimico tende sempre amodificarsi in direzione dello stato a cui è associata la minimaenergia libera. Per due filamenti di nucleotidi complementari,questo stato viene raggiunto quando si appaiano e formanouna doppia elica.
I quattro filamenti della nostra intersezione fissa possonounirsi e dar luogo alla massima quantità di DNA convenzionalea doppia elica solo formando una molecola ramificata. In gene-rale, un punto di biforcazione non è favorito - perché aumental'energia libera della molecola - ma questo incremento è com-pensato dal risparmio molto maggiore di energia nei quattrobracci composti da normale DNA a doppia elica. Oggi è sempli-
a
i›41Yri i 'ATAG C „ri bz-11-159Iniz • I Estremità coesiva
ce sintetizzare simili filamenti e realizzare nella pratica l'idea diuna molecola ramificata stabile di DNA, ma nel 1979 si trattavadi chimica d'avanguardia e io ero un cristallografo, non un chi-mico organico, sicché mi limitai a elucubrazioni teoriche. (Fusolo nel 1982 che imparai a sintetizzare il DNA.)
Ispirato da Escher
Immaginai che dovesse essere possibile costruire intersezionicon molti bracci, e non solo quattro, in molecole di DNA. Ungiorno, nell'autunno 1980, andai al bar dell'università per ri-flettere con calma sulla possibilità di realizzare intersezioni asei bracci. Per qualche motivo, mi venne in mente la xilografia
Profondità, di Maurits Cornelis Escher (si veda l'illustrazione ap. 104). Mi resi conto che il centro di ciascun pesce dell'imma-gine era una rappresentazione ideale del punto di ramificazionein un'intersezione a sei bracci. Sei strutture si estendono dalpunto centrale del pesce: testa e coda, pinna superiore e pinnainferiore, pinna sinistra e pinna destra. I pesci sono organizzaticome le molecole di un cristallo molecolare, con «celle» regolar-mente ripetute in avanti e all'indietro, in alto e in basso, a sini-stra e a destra. All'improvviso ebbi l'idea che, se avessi tenutounite le intersezioni sfruttando le estremità coesive, avrei potu-to organizzare la materia a scala nanometrica nello stesso mo-do in cui Escher aveva disposto il suo banco di pesci usando lafantasia.
CGGCTATAATGCGC
sinistrorsa
102 LE SCIENZE 431/ luglio 2004
www.lescienze.it 103

LA XILOGRAFIA DI ESCHER Profondità (a sinistra) ha ispirato l'autore a
considerare un reticolo di intersezioni a sei bracci unite a formare un
cristallo molecolare tridimensionale (sotto). Il centro di ciascun pesce ècome il punto di diramazione delle intersezioni. Anziché bracci, nei pesci
sporgono da esso testa e coda, pinna superiore e pinna inferiore, pinna
sinistra e pinna destra. Le impalcature molecolari potrebbero trattenere
altre molecole in reticoli regolari. Per esempio, gabbie di DNA che ospitino
macromolecole biologiche potrebbero servire per analisi cristallografiche.
Scheletro del DNA
Coppie di basi
UN CUBO DI DNA (a sinistra) costruito con sei anelli del più famoso
acido nucleico è la dimostrazione che si possono realizzare
strutture in tre dimensioni. Lo scheletro dei filamenti di DNA è
rappresentato da sfere colorate (di colore diverso per ciascunfilamento) e le basi sono indicate con sfere bianche. Ogni spigolo
comprende 20 coppie di nucleotidi, corrispondenti a circa due
avvolgimenti della doppia elica. Ciascun vertice è un'intersezione
a tre bracci. Lo schema semplificato a destra mostra come sono
collegati i filamenti di DNA, ma omette gli avvolgimenti dell'elica.
Le prime strutture che avevamo costruito sembravano quei giochi
che si fanno da bambini con gli stuzzicadenti piantati nella plastilinaMacromolecola
Ci sono parecchie buone ragioni per cercare di costruirestrutture del genere. Prima di tutto si può pensare di realizzareaggregati macroscopici di materia costituiti da molecole appo-sitamente progettate, unite in una struttura controllata conprecisione nanoscopica. Con questo procedimento si potrebbe-ro realizzare materiali con proprietà nuove. Per esempio, si po-trebbero produrre cristalli fotonici o altri materiali con specifi-che caratteristiche ottiche, costruendo reticoli esattamente de-fmiti, le cui unità si ripetono a una distanza specifica.
Un altro obiettivo è utilizzare il DNA come «impalcatura»per organizzare altre molecole in un reticolo, comprese quelleche di per sé non formano strutture cristalline regolari. In que-sto modo si potrebbero ottenere cristalli adatti per esperimentidi cristallografia sintetizzando gabbie di DNA che racchiudanomacromolecole biologiche come le proteine. Queste gabbiepermetterebbero di determinare con metodi cristallografici lastruttura tridimensionale delle molecole imprigionate: un re-quisito fondamentale per la progettazione razionale di farmaciche si adattino perfettamente a recettori specifici di una mole-cola bersaglio. (Peraltro questa applicazione è la motivazionepiù forte del mio interesse per questo campo di ricerca.) Oggiinfatti molti recettori che potrebbero essere eccellenti obiettiviper farmaci non possono essere studiati con i metodi conven-zionali della cristallografia. In maniera analoga, si potrebberoorganizzare componenti nanoelettronici in minuscoli disposi-tivi di memoria, come Robinson e io proponemmo già nel1987.11 mio gruppo di ricerca non ha ancora utilizzato il DNAcome impalcatura, ma i molti successi ottenuti ci avvicinanodi continuo a questo obiettivo.
Perché usare il DNA per questi scopi? La ragione principaleè che i filamenti dell'acido nucleico interagiscono in modo fa-cilmente programmabile e prevedibile. Un'estremità coesiva
della lunghezza di N basi ha una delle 4" sequenze di basi pos-sibili. Questa enorme variabilità e la tendenza dell'estremità alegarsi solo a una sequenza corrispondente aprono un campovastissimo alla progettazione di molecole consistenti in ungran numero di filamenti di DNA uniti l'uno all'altro in modototalmente specificato. Sappiamo anche che due estremità coe-sive, interagendo, formano la classica struttura del DNA, e chequesti segmenti a elica sono relativamente rigidi. Pertantosappiamo non solo quali filamenti tendano a legarsi fra loro,ma anche quale sia, in dettaglio, la forma dei segmenti che nerisultano. Riguardo alle proteine e agli anticorpi, che sono ul-teriori possibilità per realizzare elementi funzionali, non ab-biamo informazioni così specifiche. Anche queste molecolehanno una variabilità enorme, ma determinare quale confor-mazione assume una proteina o come si legano due proteine odue anticorpi è un problema arduo, che deve essere risolto se-paratamente per ogni singolo caso.
Un altro motivo per lavorare con il DNA è la semplicità dellasua sintesi, grazie agli strumenti delle biotecnologie. Possiamomanipolarlo con molti enzimi, come gli enzimi di restrizione(che tagliano il DNA in particolari siti) o le ligasi (che catalizza-no l'unione di due molecole con legami covalenti, solidi legamichimici che comportano la condivisione di coppie di elettronifra gli atomi coinvolti). Questi strumenti possono servire per sin-tetizzare e modificare il DNA convenzionale, ma anche i suoiderivati esotici, che includono basi diverse dalle solite quattro, onei quali molecole aggiuntive sono fissate all'esterno dello sche-letro del DNA. La ricerca biomedica ha prodotto molte di questevarianti, nella speranza di utilizzare gli acidi nucleici (DNA eRNA) a scopi terapeutici. E il DNA si presta particolarmente be-ne alla sintesi di questi derivati, perché ciascun nucleotide lungol'elica ha siti a cui possono legarsi altre molecole.
Infine, come vedremo più avanti, si può spingere il DNA aformare strutture diverse dalla doppia elica. Così si potrebberocostruire dispositivi nanomeccanici con parti che si muovono -per esempio pinzette che si chiudono, o un albero che ruota -sfruttando le transizioni del DNA da una struttura a un'altra.Un punto a sfavore è che gli oggetti di DNA devono essere co-struiti in soluzione acquosa. Non è un problema, tuttavia,asciugare le strutture risultanti (su una base di mica, per esem-pio), come facciamo per realizzare immagini microscopichedelle nostre creazioni.
Modelli appiccicosi
11 primo passo in qualsiasi nuovo programma di ricercascientifica è di verificarne la reale fattibilità. Nel 1991 JunghueiChen, ora all'Università del Delaware, e io la dimostrammo rea-lizzando una molecola di DNA con la forma di un cubo com-posto da bastoncini (si veda l'illustrazione in alto a destra). Cia-scuno spigolo del cubo è un segmento di DNA a doppia elica;ciascun vertice è un'intersezione a tre bracci. Dato che un verti-ce è collegato ad altri tre, si dice che la connettività del cubo èpari a tre. Gli studiosi di ingegneria genetica hanno ottenutomolti costrutti lineari di DNA, ma questa è stata la prima mole-cola con una connettività maggiore di due. 11 cubo si autoas-sembla a partire da segmenti di DNA progettati in modo daaderire l'uno all'altro, ma non di unirsi alle estremità. Le ligasipossono legare queste estremità libere, formando sei anelli chiu-si, uno per ogni faccia del cubo. A causa della struttura elicoi-dale del DNA, ogni anello è attorcigliato attorno agli anelli chelo fiancheggiano, cosicché il cubo non potrebbe disassemblarsineppure se tutti i legami che tengono unite le coppie di basi ve-nissero in qualche modo spezzati.
Yuwen Zhang, ora alla Baxter Healthcare, e io costruimmoun'altra forma, un ottaedro tronco, simile a un cubo ma piùcomplesso (si veda l'illustrazione a p. 109). Per costruire sin-goli ottaedri tronchi sarebbero sufficienti intersezioni a trebracci, ma decidemmo di realizzarle a quattro bracci. Nelle no-stre intenzioni, il braccio supplementare che sporge da ciascunvertice doveva servire per unire fra loro gli ottaedri tronchi inuna struttura più grande: ma alla fine abbandonammo questadirezione di ricerca.
Avevamo creato soltanto una minuscola quantità di ottae-dri tronchi - abbastanza per caratterizzarne la struttura, manon per tentare di unirli insieme - e anche questo infimo cam-pione ci aveva impegnati al limite delle nostre possibilità sen-za potenziare sostanzialmente le nostre procedure (per esem-pio automatizzando le fasi ripetitive). Decidemmo invece di ri-volgerci a componenti più semplici.
Ma c'era un'altra ragione a farci cambiare direzione. A uncerto punto ci rendemmo conto che i poliedri a bastoncini cheavevamo realizzato non erano rigidi. 11 DNA non è una mole-cola flessibile: un segmento della lunghezza di due o tre avvol-gimenti (come quelli che usiamo per gli spigoli dei nostri polie-dri) può piegarsi intorno all'asse dell'elica non più di quantopossa farlo un pezzo di spaghetto cotto lungo due o tre milli-metri. Questa assenza di flessibilità garantiva che gli spigolidelle nostre strutture fossero rigidi, ma scoprimmo che gli an-goli ai vertici erano assai variabili. I primi poliedri che avevamocostruito sembravano quei giochi da bambini fatti con gli stuz-zicadenti piantati in pezzetti di plastilina. Strutture del generepotrebbero avere qualche applicazione, ma certo non servono acostruire un reticolo regolare. È molto più semplice autoassem-blare un aggregato di materia ordinato, cristallino, partendo dacomponenti simili a mattoni che non a pezzi di plastilina.
104. LE SCIENZE 431/ luglio 2004
www.lescienze.it 105

DX+ J (double crossover + junction)
RETICOLI RIGIDI DI DNA
S
i possono ottenere cristalli
Jbidimensionali a partire da mattoni
rigidi di DNA. Questi ultimi (a) sono unità
DX e DX + J, che non possono ondeggiare
nei punti di giunzione come invece
accade nelle intersezioni a molti bracci.
a
Estremità coesive
Estremità coesive
411> IL,,,,,,,up111 , OPPI> e41,n.1~1b41'
DX (double crossover)
Ogni mattone ha quattro distinte
estremità coesive per legarsi ad altre
unità. Il lungo filamento verde sull'unità
DX + J sporge dal piano. Le unità hanno
lati di circa 4 nanometri per 16. Per
semplicità, entrambi i tipi sono
raffigurati schematicamente, con forme
geometriche che rappresentano le
estremità coesive (b). In soluzione,
queste ultime si legano fra loro e le unità
si autoassemblano in uno schema
bidimensionale (c). La configurazione a
strisce è ben visibile in un'immagine del
cristallo (che per la visualizzazione
viene depositato su una superficie
piana di mica) al microscopio a forza
atomica (d). Le strisce chiare,
distanziate di circa 32 nanometri, sono
i segmenti di DNA che sporgono dalle
unità DX + J. Possono autoassemblarsi
in configurazioni bidimensionali anche
parallelogrammi di DNA (e," ).
AIEVEstremità coesive • Estremità coesive
d—deiÌ
A•è§.~~41~~
~~~~4.~~M~.~4n4~»
L'Z.M1~4.1,
6201%3
V t/
e ,QC 'Lì2
j"Z-DNA
B-DNA
1— Molecola di colorante
e
E
3
Aggiunta della molecolai i r effettriceRimozione della molecola
effettrice 4)
Per risolvere il problema, il mio gruppo prese in considera-zione un altro schema di ramificazione che si osserva nei siste-mi biologici ricombinanti, vale a dire la molecola «a doppio in-crocio» o DX (double crossover), consistente in due doppie eli-che allineate affiancate, con filamenti che passano trasversal-mente da un'elica all'altra, legandole saldamente insieme (si ve-da la finestra nella pagina a fronte). Caratterizzammo questamolecola, e stabilimmo che è rigida. E dimostrammo anche cheuna molecola DX contenente un'altra piccola regione a doppiaelica (chiamata DX + J) è ancora più rigida. La regione supple-mentare a doppia elica forma una gobba sulla molecola DX,che funge da marcatore: l'equivalente nanotecnologico di unapennellata di colore.
In collaborazione con Eric Winfree, del California Institute ofTechnology, Furong Liu e Lisa A. Wenzler del mio gruppo allaNew York University hanno utilizzato combinazioni di moleco-le DX e DX + J come tasselli per realizzare cristalli bidimensio-nali aventi configurazioni definite. I tasselli sono tenuti insiemedalle estremità coesive di ciascuna elica. Una disposizione incui colonne di tasselli DX si alternano con colonne di tasselliDX + J produce uno schema di strisce distanziate fra loro dicirca 12 nanometri. Abbiamo depositato il cristallo su una su-perficie piatta di mica per esaminarlo con un microscopio aforza atomica e confermare che la struttura avesse le dimensio-ni corrette. Abbiamo anche stabilito che lo schema non eraemerso casualmente sintetizzando un secondo cristallo con tas-selli modificati in cui si hanno tre colonne DX per ogni colon-na DX + J, e abbiamo prodotto strisce con una separazionedoppia della precedente.
Di recente, il gruppo di John H. Reif alla Duke University ha
Un obiettivo cruciale
per la nanotecnologia del DNA
è riuscire ad ampliare
alle tre dimensioni
i successi ottenuti
con i sistemi bidimensionali
lizzato configurazioni bidimensionali a partire da parallelo-grammi di DNA simili ai nostri poliedri a bastoncini. Copie diquesta unità possono essere congiunte per costituire un cri-stallo che si estende come un waffle (le classiche cialde ameri-cane cotto in uno stampo di ferro quadrato) in due dimensioni.È possibile regolare le dimensioni delle cavità del reticolo va-riando quelle dei parallelogrammi. Anche se le singole interse-zioni fra i bracci non sono rigide, sistemandone quattro ai ver-tici di un parallelogramma si ottiene un'unità che si comportacorrettamente nel reticolo.
Costruire nanomacchine
Un aspetto fondamentale della nanotecnologia sono le mac-chine a scala molecolare. 11 DNA si è rivelato molto utile nellaloro costruzione. Abbiamo realizzato diversi dispositivi a parti-re da questa molecola, ma qui mi limiterò a descriverne due chehanno strutture ben definite. In entrambi i casi, il meccanismoè basato su una transizione strutturale delle molecole di DNA:vale a dire il passaggio da una conformazione (come la norma-le doppia elica) a un'altra.
11 DNA convenzionale ha un'elica destrorsa. Immaginate dirisalire una scala a chiocciola tenendo la mano sinistra sul cor-rimano interno e la destra su quello esterno: ecco, una scala co-sì è un'elica destrorsa. Questo DNA per così dire «convenziona-le» è chiamato B-DNA, ed è la struttura favorita dal punto divista energetico in un tipico mezzo acquoso.
11 DNA a doppia elica, però, può anche assumere strutturedifferenti, a seconda della sua sequenza di basi e delle speciechimiche presenti nella soluzione in cui è immerso. Una di que-
dimostrato che utilizzando simili configurazioni si possono ot-tenere «codici a barre» di DNA. In queste tassellature le posi-zioni delle strisce erano programmate in modo da formare unaconfigurazione che rappresentasse il numero «01101» (conmolecole analoghe alle nostre DX e DX + J che corrispondeva-no rispettivamente a O e 1). Lo schema veniva programmatousando un filamento iniziale di DNA la cui sequenza codifi-casse la successione 01101. Gli analoghi dei «mattoni» DX eDX + J si autoassemblavano sulle sezioni del filamento diDNA corrispondenti a O e 1. Molte di queste sequenze di cin-que mattoni si univano poi in parallelo, generando lo schema01101 con strisce distanziate di circa 15 nanometri. Quando siesaminano queste ultime con un microscopio a forza atomica,a tutti gli effetti si utilizza il codice a barre per leggere i datiche erano stati codificati sul filamento di DNA iniziale. Questometodo visivo per leggere la sequenza del DNA potrebbe acce-lerare notevolmente la fase di lettura negli elaboratori a basedi DNA, e potrebbe essere sfruttato anche per costruire mappedi mutazioni.
Chengde Mao, ora alla Purdue University, e io abbiamo rea-
UN DISPOSITIVO NANOMECCANICO B-Z che dimostra la fattibilità di
movimenti controllati è formato da due unità DX (in blu e arancione) unite
da un albero di 20 coppie di basi (in viola). Due molecole di colorante
(sfere bianca e gialla) evidenziano le posizioni delle molecole DX.
Nello stato B (in alto) l'albero è normale B-DNA destrorso, ed entrambe
le molecole DX sono sullo stesso lato. Quando si aggiunge alla soluzione
una molecola effettrice, l'albero si trasforma in Z-DNA sinistrorso,
e una delle unità DX compie tre rotazioni e mezza rispetto all'altra: così,
nella configurazione finale esse si trovano su lati opposti dell'albero.
ste strutture è lo Z-DNA, che fu caratterizzato per la prima vol-ta nel 1979 da Alexander Rich e colleghi del Massachusetts In-stitute of Technology (si veda l'illustrazione in alto a destra ap. 103). Lo Z-DNA ha una struttura sinistrorsa.
Per sintetizzare lo Z-DNA normalmente occorre una se-quenza di basi alternate citosina-guanina. Lo scheletro dellamolecola contiene gruppi fosfato, con carica negativa, chevengono a trovarsi molto vicini nella struttura dello Z-DNA.Questa disposizione è favorita solo se le cariche sui fosfati pos-sono essere reciprocamente scherniate da un ambiente acquo-
so che contiene elevate concentrazioni di sali oppure una par-ticolare specie «effettrice», che produce lo stesso risultato inconcentrazione molto più bassa. 11 vincolo della sequenza cito-sina-guanina ci permette di controllare in che punto della mo-lecola di DNA avviene la transizione B-Z (e quindi che cosa fala nostra macchina), e il vincolo ambientale ci consente dicontrollare quando avviene la transizione (e quindi quando lamacchina funziona).
Con due colleghi della New York University, Weiciiong Sun eZhiyong Shen, Mao e io abbiamo costruito un dispositivo costi-tuito da due molecole DX collegate da un «albero» di DNA adoppia elica (si veda l'illustrazione nella pagina a fronte). Alcentro dell'albero vi è una sequenza di 20 coppie di basi che, indeterminate condizioni, può assumere la struttura Z. Normal-mente ciascuna parte del dispositivo è costituita da B-DNA e ledue molecole DX sono entrambe sullo stesso lato rispetto al-l'asse dell'albero. Se si aggiunge alla soluzione una specie effet-trice, la parte centrale dell'albero si trasforma in Z-DNA e unamolecola DX compie circa tre rotazioni e mezza rispetto all'al-tra: la mezza rotazione fa sì che alla fine esse si trovino su lati
opposti rispetto all'asse dell'albero. L'eliminazione della mole-cola effettrice riporta il dispositivo alla struttura di partenza.Abbiamo dimostrato che il moto avviene realmente mediantetecniche spettroscopiche, utilizzando due molecole di colorantelegate alle molecole DX.
Questo dispositivo B-Z è piuttosto affidabile, ma non è pri-vo di difetti. Se si riuscisse a incorporare un certo numero didispositivi B-Z diversi in una struttura più grande (per esempioimo dei reticoli bidimensionali di cui abbiamo parlato in pre-cedenza), l'intero sistema avrebbe solo due stati: o tutte lemacchine sono nello stato B oppure tutte sono nello stato Z.Per controllare individualmente una serie di macchine occor-rono attivatori indipendenti. C'è un modo del tutto naturaleper raggiungere questo obiettivo: utilizzare come attivatori fi-lamenti di DNA, facendo sì che ciascuna macchina sia aziona-ta da una sequenza di basi differente.
Per mettere in pratica questa idea, Hao Yan, ora alla DukeUniversity, Xiaoping Zhang della New York University, Shen eio abbiamo ideato un sistema in grado di cambiare formaquando a esso si legano filamenti diversi. 11 dispositivo è costi-
106 LE SCIENZE 431/ luglio 2004
www.lescienze.it 10?
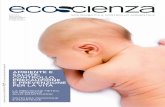
a
Filamentidi settaggio
Filamenticomplementari
Stato PX
1
6 <
3 PX PX PX
3
(-n r`
-... All"
I,—+.--— -.,•Zry 'T'..-
....,Stato JX
JX JX JX
v
I I n dispositivo a DNA controllabile può alternarsi fra due stati
(a, passi 1-8) grazie all'aggiunta e al distacco di specifici
segmenti di DNA chiamati «filamenti di settaggio». Il
dispositivo «nudo» consiste di quattro doppie eliche unite nel
centro da due filamenti spaiati di DNA (1). Quando si
aggiungono i filamenti di settaggio azzurri (2), essi si legano ai
filamenti spaiati in modo da costringere il dispositivo ad
assumere lo stato JX, ojuxtaposed ( 3). In questo stato, le
eliche rossa e verde sono sullo stesso lato, in alto e in basso. I
filamenti azzurri si staccano quando vengono aggiunti i loro
filamenti complementari (4), lasciando di nuovo il dispositivo
nudo (5).A questo punto si aggiungono i filamenti di settaggio
viola (6), che si legano in modo diverso e portano il dispositivo
nella cosiddetta configurazione PX, o paranemic crossover [7].Questo processo fa ruotare la parte inferiore del dispositivo,
collocando su lati opposti le eliche rosse e verdi. Il ciclo della
macchina può proseguire con il distacco dei filamenti viola (8)
e il ripristino di quelli azzurri.
Il funzionamento di questo dispositivo è stato verificato
collegandone numerose copie, ognuna dotata di un grande
«marcatore» trapezoidale di DNA, a formare una catena.
Quando i dispositivi sono nello stato PX (b, sotto], tutti i
marcatori sono sullo stesso lato, mentre, quando sono nello
stato JX, i marcatori sono disposti in maniera alterna (c]. Con
un microscopio a forza atomica si osserva perfettamente
questo comportamento (a, d].
USARE IL DNA COME ATTIVATORE
PER APPROFONDIRE
ANDREW J. TURBERFIELD, ALLEN P. MILLS, JR., FRIEDRICH C.SIMMEL, JENNIFER L. NEUMANN e BERNARD YURKE,A DNA-Fuel-led Molecular Machine Made of DNA, in «Nature», Vol. 406, pp.605-608, 10 agosto 2000.CHENGDE MAO, THOMAS H. LABEAN, JOHN H. REIF e NADRIAN C.SEEMAN, Logica! Computation Using Algorithmic Self-Assembly0f DNA Triple Crossover Molecules, in «Nature», Vol. 407, pp.493-496,28 settembre 2000. (Erratum: «Nature», Vol. 408, p.750, 7 dicembre 2000.)HAO YAN, XIAOPING ZHANG, ZHIYONG SHEN e NADRIAN C. SEE-MAN, A Robust DNA Mechanical Device Controlled by Hybridiza-tion Topology, in «Nature», Vol. 415, pp. 62-65, 3 gennaio2002.NADRIAN C. SEEMAN, DNA in a Materia! World, in «Nature», Vol.421, pp. 427-431, 23 gennaio 2003.ANDREW TURBERFIELD, DNA as an Engineering Materia!, in«Physics World», Vol. 16, n. 3, pp. 43-46, marzo 2003.WILLIAM M. SHIH, JOEL D. QUISPE e GERALD F. JOYCE, A 1.7-Ki-lobase Single-Stranded DNA That Folds into a Nanoscale Oc-tahedron, in «Nature», Vol. 427, pp. 618-621, 12 febbraio2004.Ulteriori informazioni sulle nanomacchine a DNA si possono tro-vare sul sito Internet del laboratorio di Nadrian C. Seeman:http://seemanlab4.chem.nyu.edu/
tuito da due doppie eliche parallele di DNA, ciascuna dellequali si riduce a un singolo filamento in una regione centraledi incrocio. Questa può assumere due stati differenti a secondadei particolari filamenti che sono stati aggiunti alla soluzioneaffinché si leghino alle sezioni a filamento singolo (si veda lafinestra qui sopra). I due stati del dispositivo sono chiamati PX(paranemic crossover) e JX (juxtaposed). Nel primo stato, le dueeliche su un lato dell'incrocio centrale sono spostate di circamezza rotazione rispetto alla loro posizione nello stato JX.
L'aggiunta alla soluzione di una particolare coppia di fila-menti (chiamati set strand, o «filamenti di settaggio») pone il di-spositivo nello stato JX, formando un legame senza incrociocon la regione centrale. Per passare allo stato PX occorre primarimuovere questi filamenti. Nel 2000 Bernard Yurke e colleghidella Lucent Technologies hanno dimostrato che si può asporta-re un filamento dal DNA legandolo al suo filamento comple-mentare. Perché questo processo sia possibile, i nostri filamentisono dotati di brevi estremità che sporgono e restano spaiate.Quando aggiungiamo alla soluzione un filamento complemen-tare per tutta la sua lunghezza a un filamento di settaggio, dap-
prima esso si lega alle estremità spaiate e poi stacca il resto delfilamento dal dispositivo.
Avendo rimosso dalla struttura i primi filamenti di settaggio,possiamo ora inserirne altri, che si leghino alla regione centraleformando un incrocio. Questo legame fa ruotare le due doppieeliche e porta il dispositivo nello stato PX. Il processo può esse-re capovolto ripristinando il primo gruppo di filamenti; in que-sto modo le doppie eliche possono essere fatte ruotare avanti eindietro a volontà. Se si hanno molti dispositivi PX-JX diversi,è possibile farli funzionare in modo indipendente inserendo erimuovendo filamenti di settaggio progettati per adattarsi alleloro specifiche regioni di legame.
Per verificare i movimenti del nostro dispositivo abbiamoutilizzato il microscopio a forza atomica. A questo scopo abbia-mo costruito una lunga catena di dispositivi e collegato unagrande unità di DNA a forma di trapezio a un lato di ciascundispositivo. Quando questi sono tutti nello stato PX, le unitàtrapezoidali si trovano tutte sullo stesso lato della catena, men-tre quando tutti i dispositivi sono nello stato JX le unità adotta-no una disposizione a zig-zag.
Nel 2000 Yurke e colleghi hanno dimostrato il funzionamentodi «pinzette» nanoscopiche costituite da tre filamenti di DNA. Fi-lamenti di settaggio che Yurke chiama «filamenti combustibile»provocavano l'apertura e la chiusura del dispositivo. Altri ricer-catori hanno usato metodi simili per attivare ribozimi (enzimi co-stituiti da RNA). Nel 1998 Michael P. Robinson e Andrew D. El-lington dell'Università del Texas ad Austin riuscirono a incre-mentare di 10.000 volte l'attività di un ribozima grazie all'ag-giunta di un particolare filamento di settaggio che si legava al ri-bozima stesso, alterandone la conformazione.
Il futuro
Un obiettivo cruciale per la nanotecnologia che si basa sulDNA è quello di ampliare alle tre dimensioni i successi ottenuticon sistemi bidimensionali. Quando sarà raggiunto, avremo di-mostrato la possibilità di progettare materiali solidi specificandouna serie di sequenze di DNA e poi combinandole. Se i sistemisaranno altamente ordinati, sarà possibile eseguire le analisi cri-stallografiche di molecole trattenute all'interno di reticoli regola-
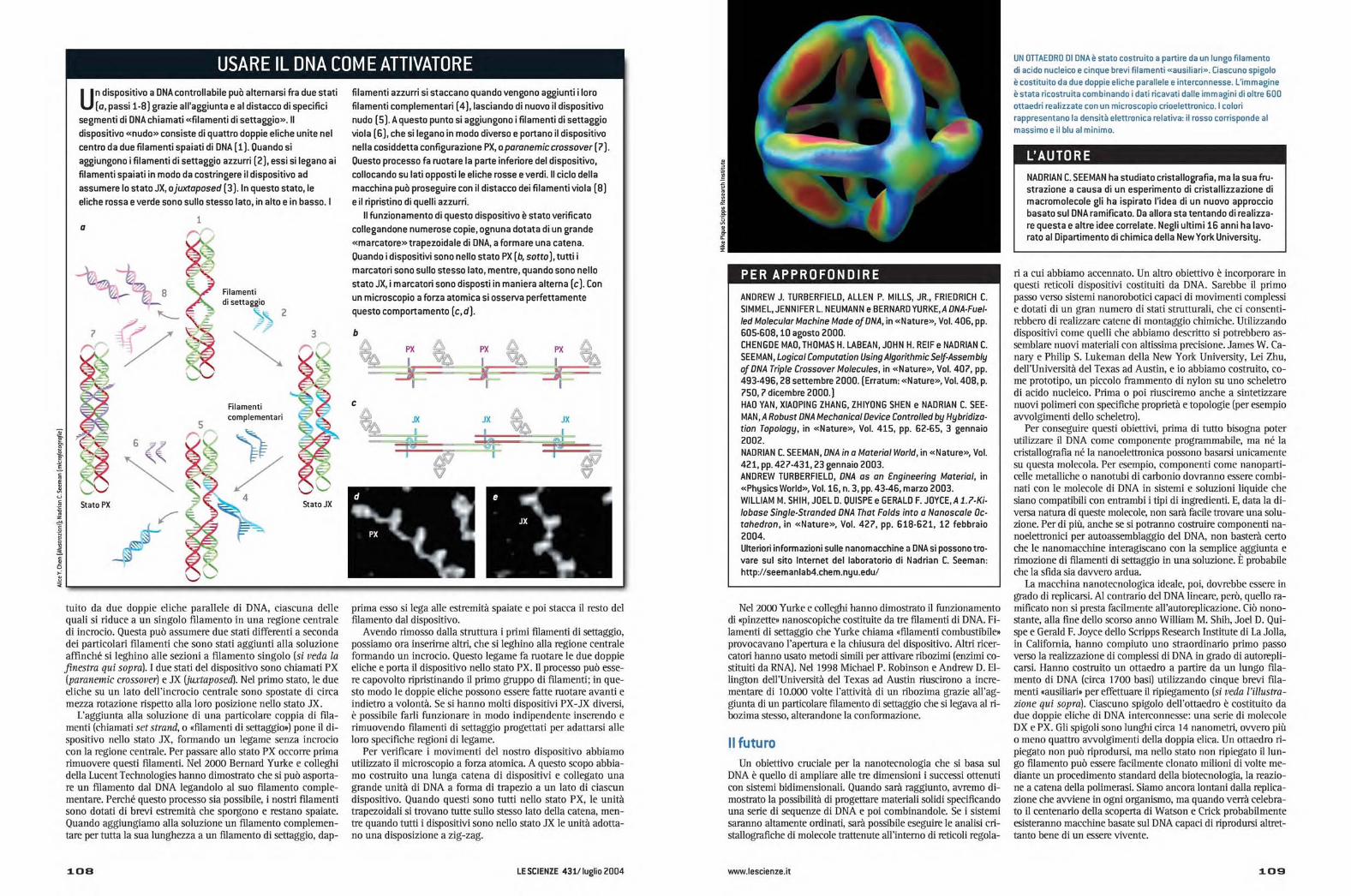
IN OTTAEDRO DI DNA è stato costruito a partire da un lungo filamento
di acido nucleico e cinque brevi filamenti «ausiliari». Ciascuno spigolo
è costituito da due doppie eliche parallele e interconnesse. L'immagine
è stata ricostruita combinando i dati ricavati dalle immagini di oltre 600
ottaedri realizzate con un microscopio crioelettronico. I colori
rappresentano la densità elettronica relativa: il rosso corrisponde al
massimo e il blu al minimo.
L'AUTORE
NADRIAN C. SEEMAN ha studiato cristallografia, ma la sua fru-strazione a causa di un esperimento di cristallizzazione dimacromolecole gli ha ispirato l'idea di un nuovo approcciobasato sul DNA ramificato. Da allora sta tentando di realizza-re questa e altre idee correlate. Negli ultimi 16 anni ha lavo-rato al Dipartimento di chimica della New York University.
ri a cui abbiamo accennato. Un altro obiettivo è incorporare inquesti reticoli dispositivi costituiti da DNA. Sarebbe il primopasso verso sistemi nanorobotici capaci di movimenti complessie dotati di un gran numero di stati strutturali, che ci consenti-rebbero di realizzare catene di montaggio chimiche. Utilizzandodispositivi come quelli che abbiamo descritto si potrebbero as-semblare nuovi materiali con altissima precisione. James W. Ca-nary e Philip S Lukeman della New York University, Lei Zhu,dell'Università del Texas ad Austin, e io abbiamo costruito, co-me prototipo, un piccolo frammento di nylon su uno scheletrodi acido nucleico. Prima o poi riusciremo anche a sintetizzarenuovi polimeri con specifiche proprietà e topologie (per esempioavvolgimenti dello scheletro).
Per conseguire questi obiettivi, prima di tutto bisogna poterutilizzare il DNA come componente programmabile, ma né lacristallografia né la nanoelettronica possono basarsi unicamentesu questa molecola. Per esempio, componenti come nanoparti-celle metalliche o nanotubi di carbonio dovranno essere combi-nati con le molecole di DNA in sistemi e soluzioni liquide chesiano compatibili con entrambi i tipi di ingredienti. E, data la di-versa natura di queste molecole, non sarà facile trovare una solu-zione. Per di più, anche se si potranno costruire componenti na-noelettronici per autoassemblaggio del DNA, non basterà certoche le nanomacchine interagiscano con la semplice aggiunta erimozione di filamenti di settaggio in una soluzione. È probabileche la sfida sia davvero ardua.
La macchina nanotecnologica ideale, poi, dovrebbe essere ingrado di replicarsi. Al contrario del DNA lineare, però, quello ra-mificato non si presta facilmente all'autoreplicazione. Ciò nono-stante, alla fine dello scorso anno William M Shih, Joel D. Qui-spe e Gerald F. Joyce dello Scripps Research Institute di La Jolla,in California, hanno compiuto uno straordinario primo passoverso la realizzazione di complessi di DNA in grado di autorepli-carsi. Hanno costruito un ottaedro a partire da un lungo fila-mento di DNA (circa 1700 basi) utilizzando cinque brevi fila-menti «ausiliari» per effettuare il ripiegamento (si veda l'illustra-zione qui sopra). Ciascuno spigolo dell'ottaedro è costituito dadue doppie eliche di DNA interconnesse: una serie di molecoleDX e PX. Gli spigoli sono lunghi circa 14 nanometri, ovvero piùo meno quattro avvolgimenti della doppia elica. Un ottaedro ri-piegato non può riprodursi, ma nello stato non ripiegato il lun-go filamento può essere facilmente donato milioni di volte me-diante un procedimento standard della biotecnologia, la reazio-ne a catena della polimerasi. Siamo ancora lontani dalla replica-zione che avviene in ogni organismo, ma quando verrà celebra-to il centenario della scoperta di Watson e Crick probabilmenteesisteranno macchine basate sul DNA capaci di riprodursi altret-tanto bene di un essere vivente.
08 LE SCIENZE 431/luglio 2004
www.lescienze.it 109