
CORSO DI PERFEZIONAMENTO IN MEDICINA...
127
UNIVERSITA’ “G. d’ANNUNZIO” -CHIETI- PESCARA DIPARTIMENTO DI SCIENZE BIOMEDICHE Sezione di Terapia Medica CORSO DI PERFEZIONAMENTO IN MEDICINA BIOINTEGRATA Prof. Dott. Francesco Borghini
Transcript of CORSO DI PERFEZIONAMENTO IN MEDICINA...

UNIVERSITA’ “G. d’ANNUNZIO” -CHIETI-PESCARA
DIPARTIMENTO DI SCIENZE BIOMEDICHE Sezione di Terapia Medica
CORSO DI PERFEZIONAMENTO IN
MEDICINA BIOINTEGRATA
Prof. Dott. Francesco Borghini

UNIVERSITA’ “G. d’ANNUNZIO” -CHIETI-PESCARA
DIPARTIMENTO DI SCIENZE BIOMEDICHE Sezione di Terapia Medica
CORSO DI PERFEZIONAMENTO IN
MEDICINA BIOINTEGRATA
www.centrostellamaris.it

Programma AA.2010-2011 • Definizione di PNEI. Il Sistema nervoso: le cellule del SN e loro
connessioni; il tronco encefalo, diencefalo, SNV. Sistema endocrino: gli ormoni, loro funzioni e comunicazioni.
Controllo cerebrale sul Sistema Endocrino. Assi neuroendocrini: ipotalamo-ipofisi, ipotalamo-ipofisi-surreni, ipotalamo-ipofisi-tiroide, ipotalamo-ipofisi-gonadi. Sistema immunitario: cenni di immunologia (teoria della selezione clonale, immunità naturale ed acquisita, sistema HLA, sistema immunitario come organo di senso, produzione di ormoni da parte del sistema immunitario che fungono da mediatori tra cervello e sistema endocrino). Cervello e psiche. La grande connessione: Sistema nervoso ed immunità; sistema dello stress; sistema endocrino e cervello. Endocrinosenescenza e P.N.E.I. DCA prototipo di PNEIG.
Controllo cerebrale sul
Sistema Endocrino

il bioritmo

La Cronobiologia studia i fenomeni biologici condizionati dal tempo e gli effetti delle loro
interazioni con gli organismi viventi.
La cronobiologia

Tutta la materia vivente presenta una ritmicità, determinata biologicamente, ereditaria e specie specifica, che necessita però di una influenza ambientale per la sua sincronizzazione. Esistono ritmi biologici circadiani (circa-dies = nell'arco di un giorno ), ultradiani (si verificano più volte al giorno, ogni 90-120 min.) e infradiani (che richiedono più di un giorno, meno di 1 ciclo in 28 ore). I ritmi circadiani sono il frutto delle interazioni tra un orologio interno (NSC) e i segnali esterni del magnetismo terrestre, dei campi elettrici, dei campi gravitazionali e delle radiazioni cosmiche influenzate dal movimento di altri pianeti, per un totale di 24 ore .
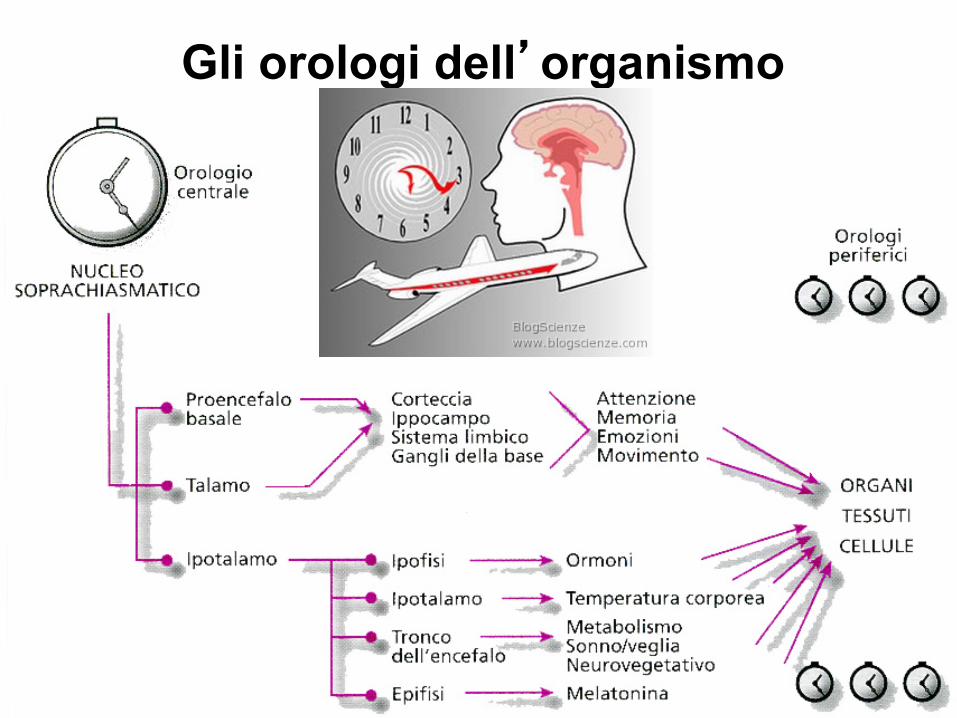
Gli orologi dell’organismo

Functional significance of circadian rhythms
� Fundamental property of living organisms
� “Predictive homeostasis”: Matching of rest/activity patterns and functioning to geophysical day
� Virtually every physiological and mental function in human beings varies as a function of time of day
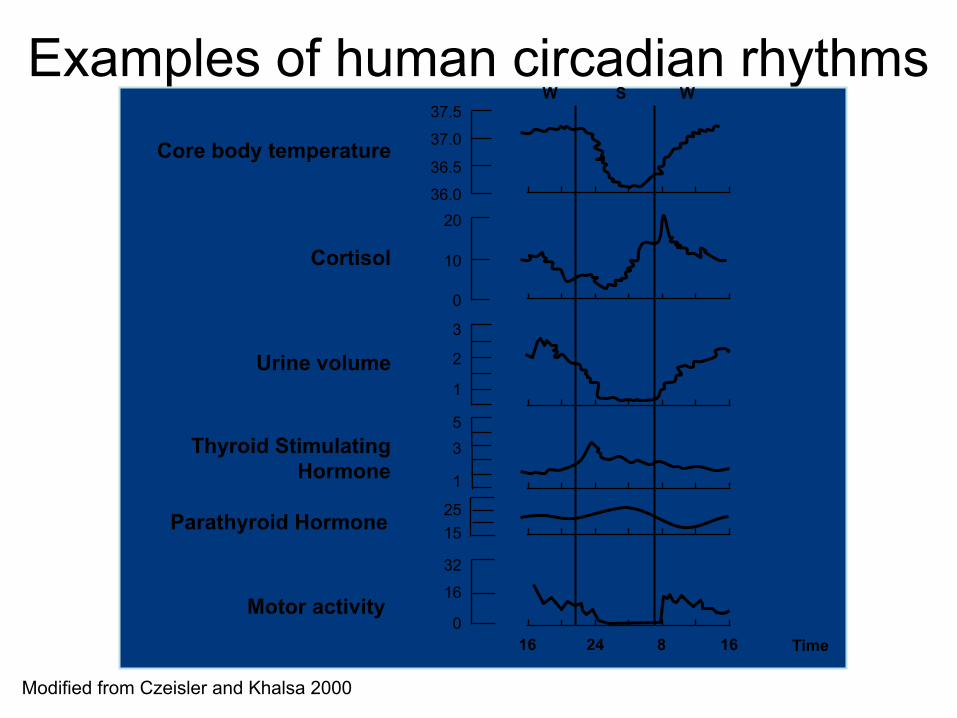
Examples of human circadian rhythms
Modified from Czeisler and Khalsa 2000
Time
Urine volume 3
2
1
Cortisol
20
10
0
Core body temperature
37.5
37.0
36.5
36.0
W W S
Thyroid Stimulating Hormone 1
3 5
Parathyroid Hormone 25 15
Motor activity 16 24 8 16
16
0
32
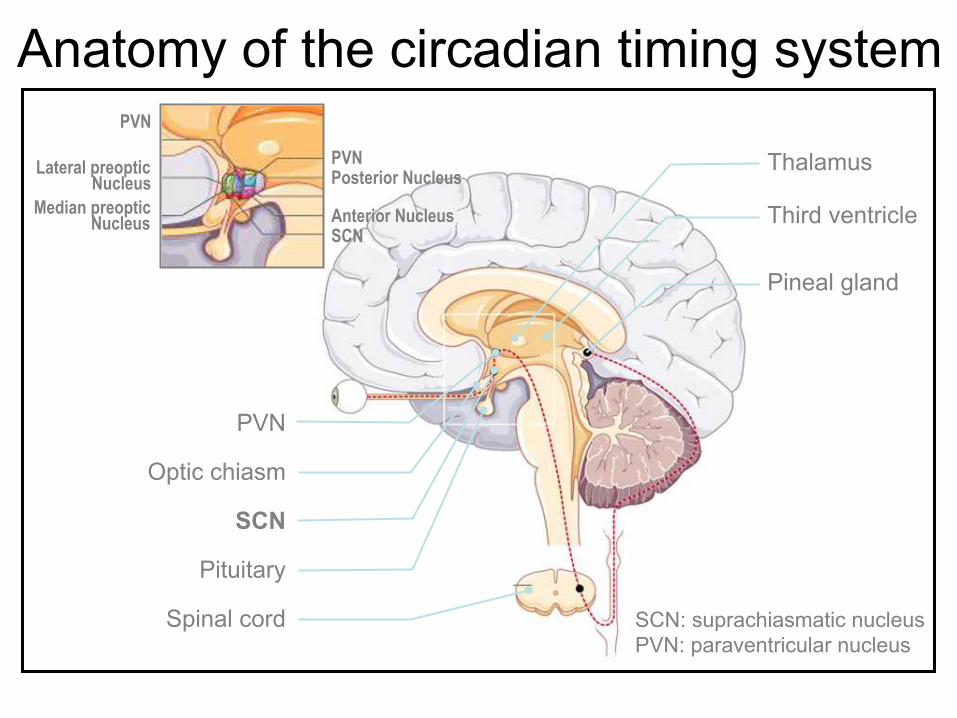
Anatomy of the circadian timing system
Corpus callosum
Ventromedial Nucleus
Thalamus
Third ventricle
Pineal gland
PVN
Optic chiasm
SCN
Pituitary
Spinal cord SCN: suprachiasmatic nucleus PVN: paraventricular nucleus
PVN Posterior Nucleus
Anterior Nucleus SCN
PVN
Lateral preoptic Nucleus
Median preoptic Nucleus

L’orologio centrale ipotalamico imprime il ritmo all’organismo; tuttavia, per evitare sfasamenti, questo ha bisogno di sincronizzatori esterni (Zeitgebers): primari: ciclo luce-buio; caldo/freddo, latitudine/longitudine; cicli stagionali; cicli lunari secondari: derivanti dall’organizzazione della nostra vita: ritmi lavorativi; orario dei pasti

NASA 23/04/2010 Solar Dynamics Observatory
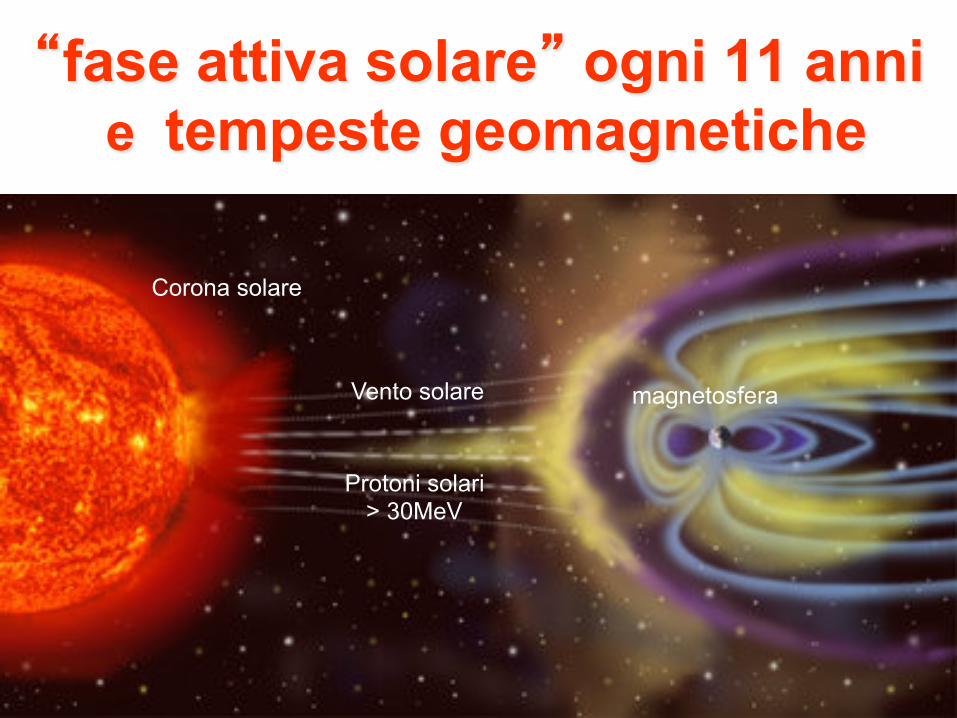
“fase attiva solare” ogni 11 anni e tempeste geomagnetiche
magnetosfera Vento solare
Corona solare
Protoni solari > 30MeV
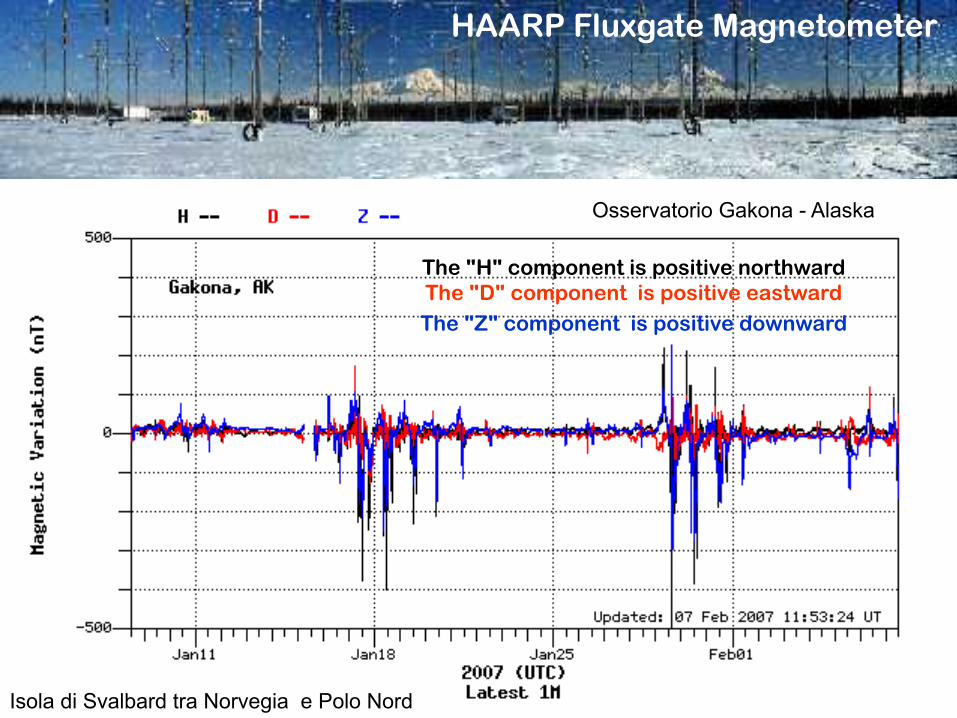
HAARP Fluxgate Magnetometer
The "H" component is positive northward The "D" component is positive eastward The "Z" component is positive downward
Isola di Svalbard tra Norvegia e Polo Nord
Osservatorio Gakona - Alaska

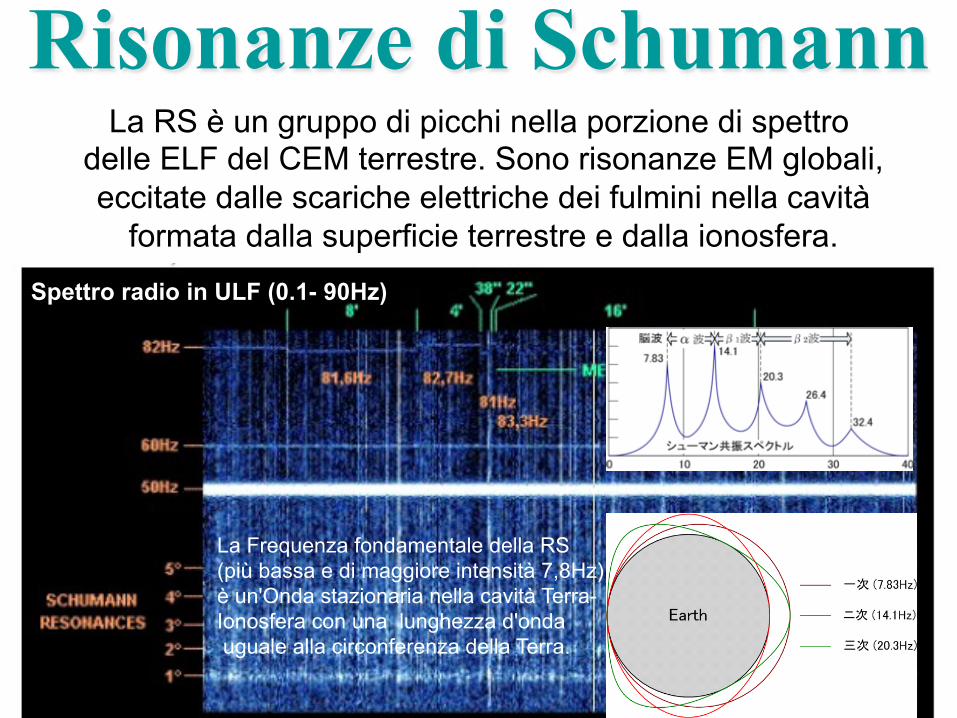
Risonanze di Schumann La RS è un gruppo di picchi nella porzione di spettro
delle ELF del CEM terrestre. Sono risonanze EM globali, eccitate dalle scariche elettriche dei fulmini nella cavità
formata dalla superficie terrestre e dalla ionosfera.
Spettro radio in ULF (0.1- 90Hz)
La Frequenza fondamentale della RS (più bassa e di maggiore intensità 7,8Hz) è un'Onda stazionaria nella cavità Terra-Ionosfera con una lunghezza d'onda uguale alla circonferenza della Terra.
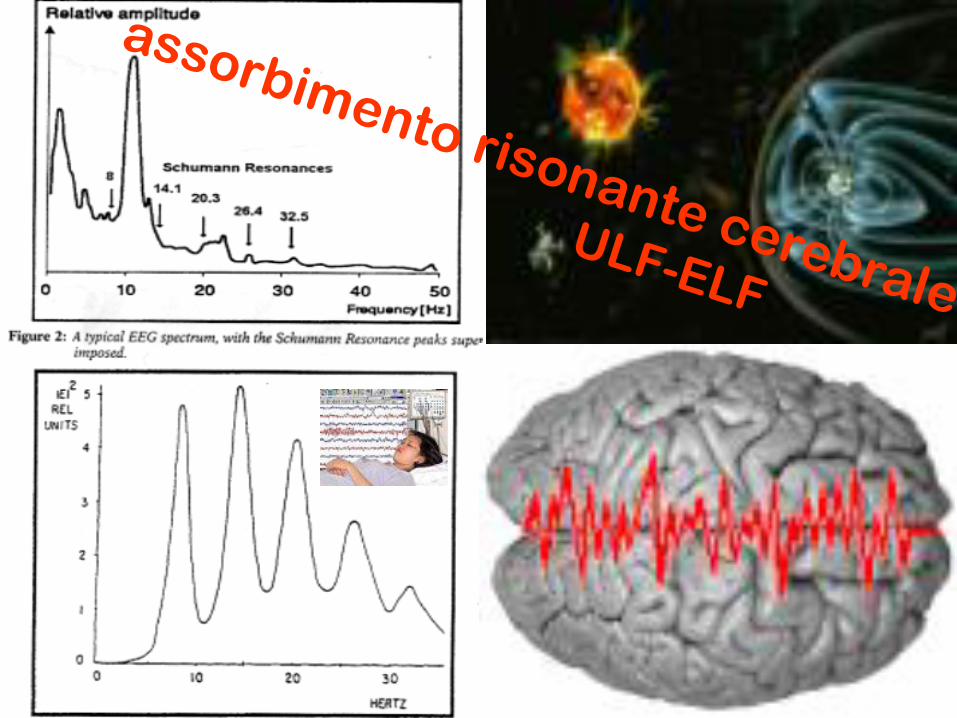
assorbimento risonante cerebrale
ULF-ELF

ECG
E E G
10-1 V/cm
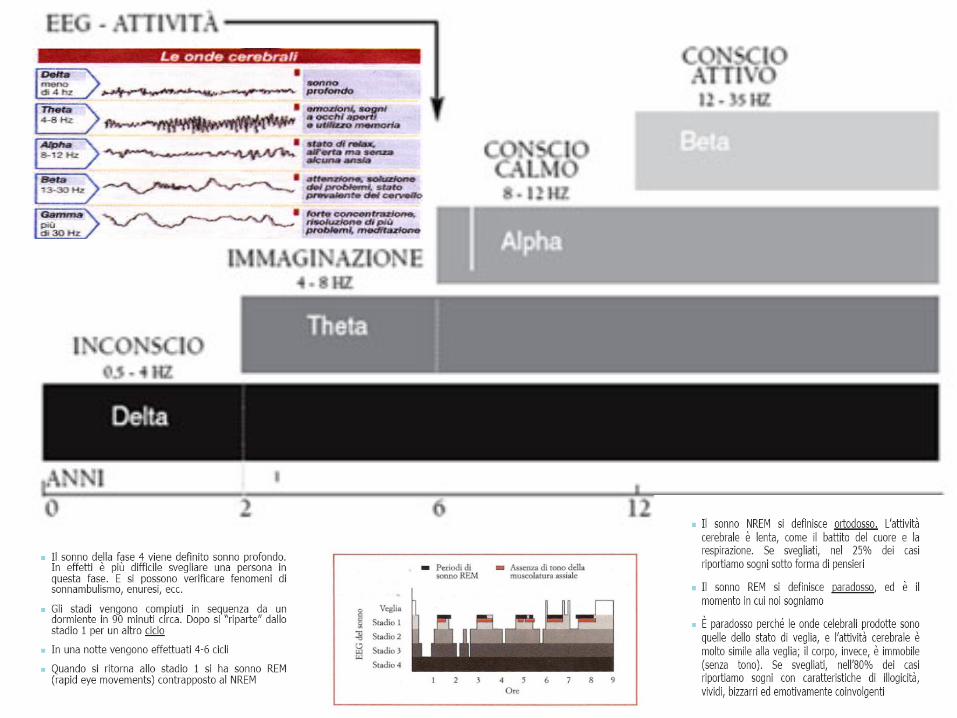

• Il passaggio dallo stadio 1 allo stadio 4 è caratterizzato da onde sempre più lente THETA /DELTA ma di un voltaggio sempre più elevato nella corteccia cerebrale, da un rallentamento del respiro e da un approfondimento del sonno. Questi stati detti SONNO SINCRONO, in quanto i ritmi elettrico corticali, cioè della corteccia cerebrale sono sincronizzati: la SEROTONINA contenuta nei neuroni provoca una induzione al sonno SINCRONICO e lo mantiene. Dopo circa 90 minuti di sonno sincrono, pur continuando a dormire, avviene, quasi improvvisamente, una trasformazione dei ritmi che da sincroni diventano desincronizzati.

• Il tono muscolare, ridotto durante il sonno sincrono, cade completamente a zero, ed cominciano a comparire dei movimenti oculari molto rapidi RAPID EYES MOVIMENTS. LA FASE del sonno REM o desincronizzato è la FASE DEL SOGNO
• Il sonno REM è caratterizzato da onde ALFA; il loro primo ciclo dura circa 10 minuti, ed aumenta di 2-3 minuti ad ogni ciclo, sino a diventare di 30 minuti. Queste fasi di onde ALFA sono dette fasi REM, che diventano sempre più lunghe col trascorrere della notte, fino ad arrivare al risveglio in onde BETA.

• Il sonno REM è caratterizzato da una desincronizzazione dei ritmi corticali, paradossalmente simili a quelli della veglia, per cui questa fase di sonno è chiamata SONNO PARADOSSO. Durante questa fase si notano movimenti oculari e atonia muscolare. Dal punto di vista neurovegetativo si hanno invece delle vere e proprie burrasche neurovegetative che si manifestano in variazione della pressione sanguigna, aritmie cardiache ed irrorazione cardiaca.

TEORIE SUL SONNO
• La teoria CIRCADIANA: sostiene che seguiamo il ritmo luce–buio e durante la notte possiamo rilassarci. I nostri antenati avevano maggiori probabilità di sopravvivenza se riparati durante la notte.
• La teoria RISTORATIVA: sostiene che dormiamo per recuperare le forze mentali e fisiche, una sorta di riparatore delle attività eseguite durante la veglia


leptina

I releasing hormons ipotalamici, oltre all’effetto potenziante o inibente sull’ipofisi sono essi stessi
influenzati da neurotrasmettitori e ormoni: nell’IPAA, la secrezione del CRH è influenzata dalla
serotonina (+) dal GABA (-) nell’asse tiroideo il TRH, che stimola la produzione del
TSH ipofisario, è contrastato dalla dopamina. NB –possono infine esserci inserimenti laterali tra
assi neuroendocrini diversi (feed-sideward)
La regolazione neuroendocrina può dunque essere meglio resa passando da una visione sequenziale
ad una visione di funzionamento a rete
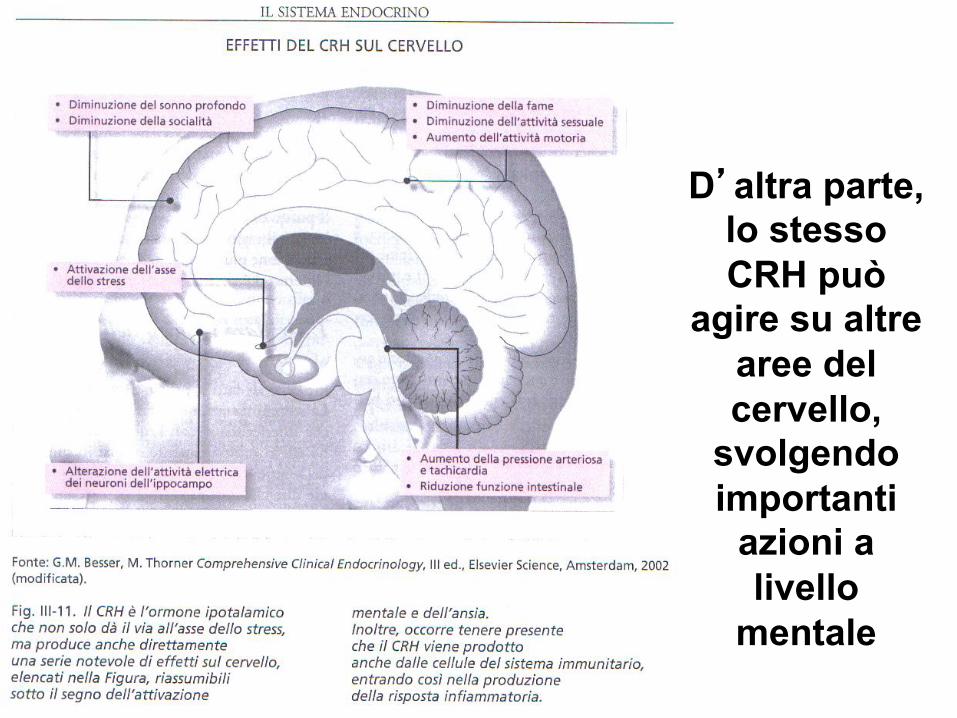
D’altra parte, lo stesso CRH può
agire su altre aree del cervello,
svolgendo importanti
azioni a livello
mentale
epifisi

• La ghiandola pineale o epifisi è una ghiandola endocrina grande come una nocciola, sporge all'estremità posteriore del 3° ventricolo. Appartiene all‘epitalamo ed è collegata mediante alcuni fasci nervosi pari e simmetrici , alle circostanti parti nervose. I "pinealociti" derivati dalla nevroglia, sono situati in sede perivascolare e producono l‘ormone melatonina antagonista degli ormoni gonadotropi ipofisari, infatti all'inizio della pubertà i suoi livelli calano notevolmente e nell'epifisi si accumula la “sabbia pineale” o "cristalli di microcalcite“, concrezioni calcaree di fosfati e carbonato di calcio, soggetti alle alterazioni dei campi elettromagnetici.
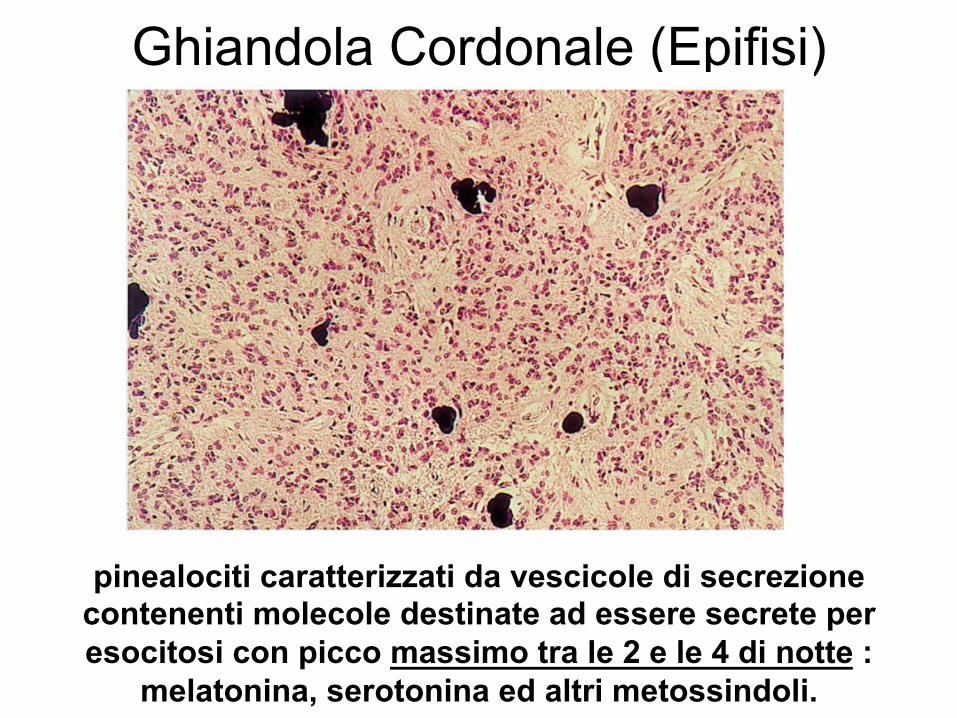
Ghiandola Cordonale (Epifisi)
pinealociti caratterizzati da vescicole di secrezione contenenti molecole destinate ad essere secrete per esocitosi con picco massimo tra le 2 e le 4 di notte :
melatonina, serotonina ed altri metossindoli.

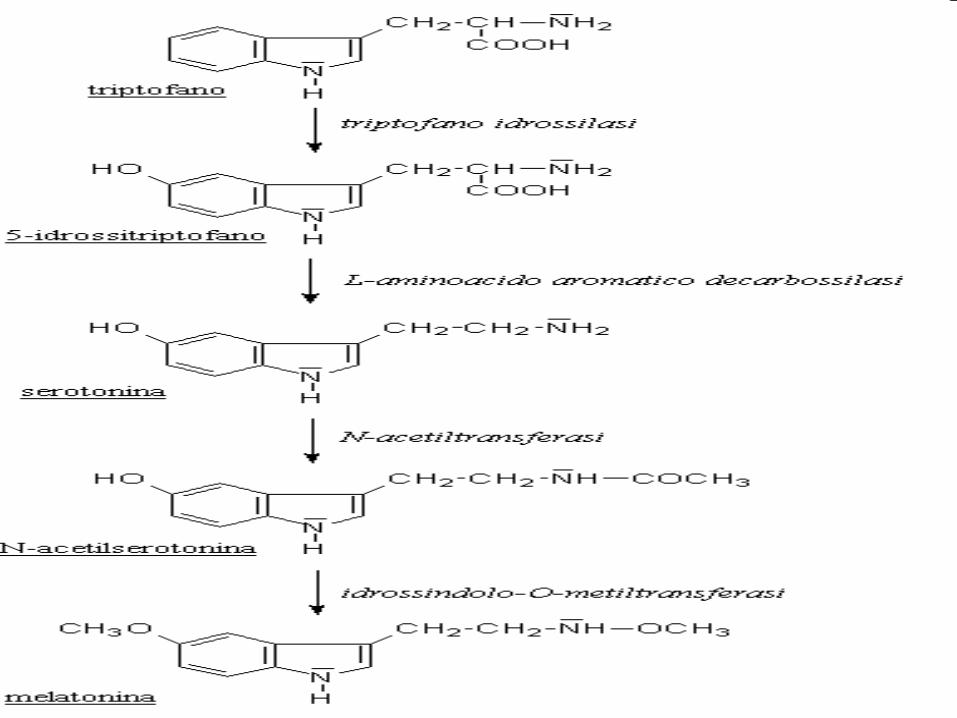
La produzione di melatonina non dipende dalla concentrazione intracellulare del suo precursore, il triptofano, ma da quella degli
enzimi che catalizzano le ultime due reazioni della biosintesi dell’ormone: la
serotonin N acetil transferasi (SNAT) e la idrossindol o metil transferasi (HIOMT).
Entrambi controllati dalle catecolamine, che, stimolando direttamente i pinealociti, attivano la trascrizione dei geni che li codificano aumentandone la sintesi.

Serotonin concentrations are higher in the pineal than in any other organ or in any brain region. They exhibit a striking diurnal rhythm remaining at a maximum level during the daylight hours and falling by more than 80% soon after the onset of darkness as the serotonin is converted to melatonin, 5-hydroxytryptophol and other methoxyindoles. The activities of both pineal enzymes SNAT e HIOMT rise soon after the onset of darkness because of the enhanced release of norepinephrine from sympathetic neurons terminating on the pineal parenchymal cells

Serotonin and Melatonin • "Melatonin is manufactured from serotonin.
When melatonin levels increase, serotonin levels usually decrease, since more serotonin is converted to melatonin. On the other hand, exposure to light lowers melatonin levels and increases serotonin levels. Consequently, serotonin levels are lower, while melatonin levels are higher, in the winter as compared to the spring and summer, especially in people with SAD. (Page 143)"
• Book: Somer, Elizabeth, M.A., R.D. Food & Mood. Henry Holt and Company, LLC, 1999.

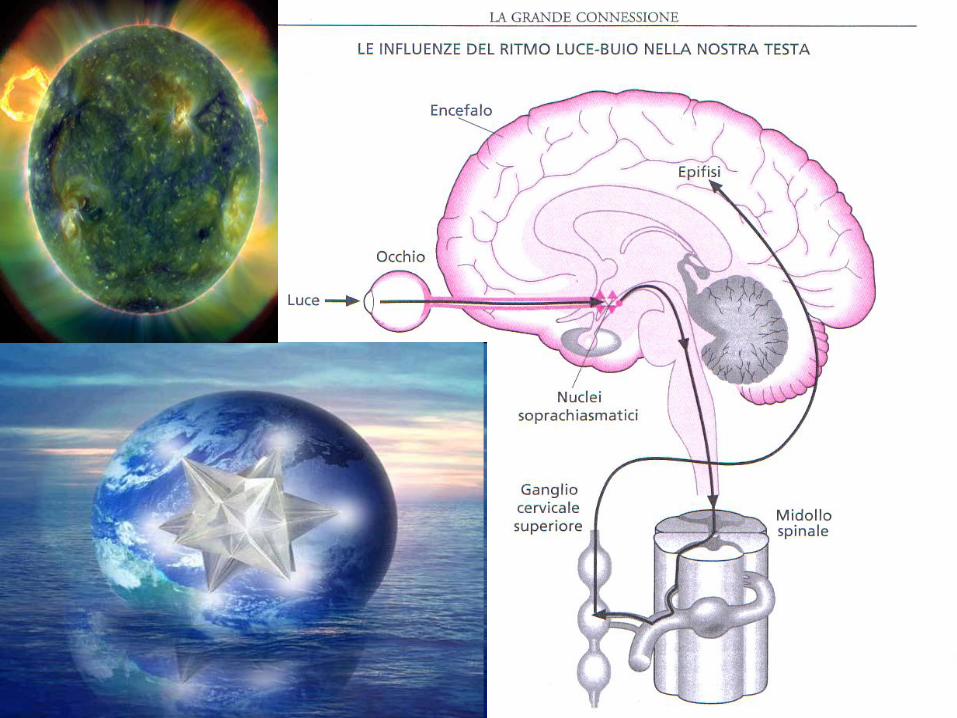
• la melatonina e' prodotta dall’epifisi trasduttore neuroendocrino con ritmo circadiano di 24 ore dalle ore 20 al picco delle 3-4 del mattino, per poi scendere ai normali valori diurni. Luce→ cell. ganglionari retina ant.→ n.ottico→ n.soprachiasmatico ipot→ nn.paraventricolari→ midollo allungato→ganglio cervicale sup. → inibizione pineale. La luce iperpolarizzado i fotorecettori inibisce il rilascio di noradrenalina (NA) a livello del pinealocita.Viceversa, al buio vi è un aumento sia del numero di recettori a1- e b1- adrenergici all'interno della ghiandola, sia della secrezione di NA che attiva la sintesi e secrezione di melatonina, ridotta da 200 lux e bloccata >2500 lux. Antistress per antagonismo del cortisolo, modula i ritmi circadiani e del SNV; riduce i livelli circolanti di NE e la PA.

Biosintesi della melatonina
La MT, sintetizzata di notte dall’epifisi, deriva dalla 5HT prodotta dai pinealociti, per N-acetilazione e
metilazione. La MT infine agisce su recettori MT1-2-3. La luce iperpolarizzando i fotorecettori inibisce il rilascio di NA a livello del pinealocita. Viceversa, il buio aumenta sia il numero di recettori alfa1- e beta1- adrenergici all'interno della ghiandola, sia la secrezione di NA, che stimola, attraverso recettori beta1 e l’cAMP, l’enzima N-acetiltransferasi.
TP idrossilasi
TP decarbossilasi
HIOMT
SNAT
STOP alla luce iperpolarizzante

Mentre la serotonina aumenta l’energia e la coscienza, la melatonina tende a diminuire l’attività delle onde cerebrali e prepara il corpo per il sonno. Induce il “sonno a onde lente”, profondamente ristoratore durante il quale avviene il ripristino dei tessuti danneggiati grazie alla capacità della MT di stimolare la riparazione del DNA. La produzione di melatonina, si riduce anche in condizioni di stress cronico per la caduta dei livelli di corticoidi surrenali che impedisce la trasformazione di serotonina in melatonina .

• J Neuroendocrinol. 2009 Feb;21(2):90-7. Epub 2008 Dec 6. • Local corticosterone infusion enhances nocturnal pineal
melatonin production in vivo. • Fernandes PA, Bothorel B, Clesse D, Monteiro AW, Calgari C, Raison S, Simonneaux V,
Markus RP.Laboratório de Cronofarmacologia, Departamento de Fisiologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, Brazil. [email protected]
• Melatonin, an important marker of the endogenous rhythmicity in mammals, also plays a role in the body defence against pathogens and injuries. In vitro experiments have shown that either pro- or anti-inflammatory agents, acting directly in the organ, are able to change noradrenaline-induced pineal indoleamine production. Whereas corticosterone potentiates melatonin production, incubation of the gland with tumour necrosis factor-alpha decreases pineal hormonal production. In the present study, we show that nocturnal melatonin production measured by intra-pineal microdialysis is enhanced in pineals perfused with corticosterone at concentrations similar to those measured in inflamed animals. In vitro experiments suggest that this enhancement may be due to an increase in the activity of the two enzymes that convert serotonin to N-acetylserotonin (NAS) and NAS to melatonin. The present results support the hypothesis that the pineal gland is a sensor of inflammation mediators and that it plays a central role in the control of the inflammatory response.

• La melatonina e la serotonina sono note per il loro effetto sulle cellule nervose e sulla attività psichica; ma questa funzione è solo il 5% della loro attività, poiché il 95% viene prodotta dalle cellule enterocromaffini dell'intestino e qui regolano la motilità intestinale. La serotonina stimola spasmo intestinale, produzione di muco, secrezione di enzimi digestivi sia direttamente che indirettamente attraverso il parasimpatico. Inoltre è anche un mediatore chimico dell’infiammazione preformato nei mastociti, provoca vasodilatazione e vasocostrizione a seconda dell’integrità dell'endotelio (emicranie serotoninergiche).

• la serotonina, contenuta nelle piastrine e viene rilasciata al momento della loro attivazione (coagulopatie). La melatonina antiinfiammatoria per blocco delle citochine proinfiammatorie (IL1-IL6) e attivazione di quelle antiinfiammatorie (IL10). Neutralizza i radicali liberi, aumentando l'attività del glutatione. Entra nel nucleo cellulare regolando l’espressione genica (EPIGENETICA). Un eccesso di serotonina attiverebbe gli enzimi che la trasformano in melatonina; che poi per una risposta pronta alle noxae si ritrasforma in serotonina.



Si produce una sincronizzazione tra variazioni dell'ambiente e variazioni nell'interno dell'organismo, sempre
attraverso ipotalamo, ipofisi ed epifisi. Essi regolano poi cuore, fegato, reni,
intestino e ghiandole surrenali, elaborando una notevole quantità di messaggi ormonali.
Seguono questo ritmo anche la temperatura, il sonno ed in modo particolare
i livelli serici di serotonina, che si presentano molto più bassi durante
i mesi invernali.

Nel DSM IV° si classifica il SAD (seasonal affective disorder ),
come disturbo affettivo o emotivo stagionale, che si evidenzia con alterazione patologica
duratura del tono dell'umore, caratterizzato da fasi alterne di depressione ed eccitazione,
di norma depressione in autunno-inverno ed eccitazione in primavera-estate.

Il ruolo chiave nella SAD, dell'attività dell'epifisi, che reagisce a seconda dell'intensità degli stimoli luminosi
provenienti dall’esterno. In primavera sia l'intensità come la durata della luce
del sole sono in continuo aumento. Qualsiasi tipo di luce, sia essa naturale od artificiale, influenza i livelli ormonali, la temperatura corporea, l'attività del
cervello e perfino l'umore dell'individuo.

Le Sindromi Affettive Stagionali dipendono, oltre che dalle ore di luce solare,
dall’evoluzione delle perturbazioni atmosferiche, in modo particolare tutte
quelle in arrivo dall’Atlantico nei mesi di febbraio e marzo. Esse portano una
notevole diminuzione delle ore di sole e della luminosità del cielo, con conseguente azione depressiva più acuta e persistente. La SAD è più frequente nelle femmine in
età riproduttiva ed in determinati gruppi familiari

I livelli serici di serotonina aumentano in primavera e cadono poi in inverno
proporzionalmente alle ore di luce solare. La melatonina, sintetizzata di notte
dall’epifisi, deriva dalla serotonina prodotta dai pinealociti, per N-acetilazione e
metilazione. La sua sintesi è regolata dal fotoperiodo ed è sotto il controllo della NA che, nelle ore di buio, stimola, attraverso
recettori Beta1 e l’cAMP, l’enzima N-acetiltransferasi. La melatonina infine
agisce su recettori MT1-2-3

Melatonergic pathway controls suprachiasmatic nucleus activity
� The human SCN contains high density of melatonergic and 5-HT2C receptors
� Melatonin feedback on MT1 and MT2 receptors participates in the regulation of SCN activity
– MT1 receptors mediate the amplitude of the SCN rhythm
– MT2 receptors mediate phase shifting of the SCN rhythm
� 5-HT2C receptors are involved in the regulation of deep sleep
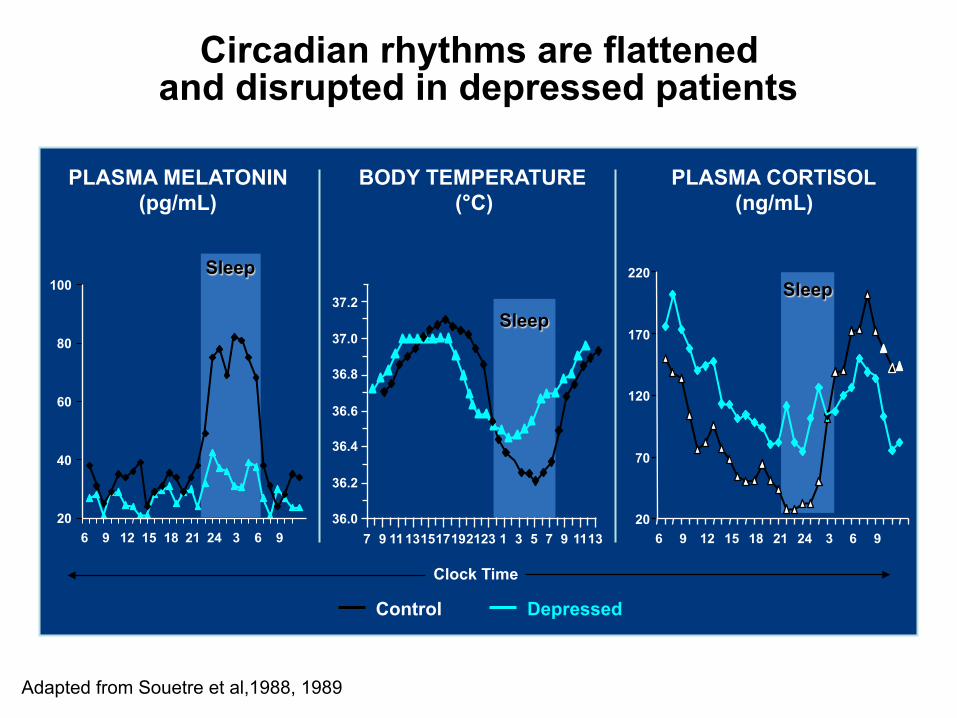
Circadian rhythms are flattened and disrupted in depressed patients
Adapted from Souetre et al,1988, 1989
Control Depressed
20
40
60
80
100
6 9 12 15 18 21 24 3 6 9
Sleep
PLASMA MELATONIN (pg/mL)
PLASMA CORTISOL (ng/mL)
Sleep
20
70
120
170
220
6 9 12 15 18 21 24 3 6 9
BODY TEMPERATURE (°C)
Sleep
7 9 11 13 15 17 19 21 23 7 9 11 13 5 3 1
37.2
37.0
36.8
36.6
36.4
36.2
36.0
Clock Time

� Depressed mood is the most salient feature of depression
� Depressed mood is the first listed criterion required for the diagnosis of MDD (DSM-IV)
Symptomatic circadian disturbances in MDD patients: depressed mood
Depressed mood, a circadian core symptom of depression
Diagnostic and Statistical Manual of Mental Disorders, Fourth Revision (DSM-IV)
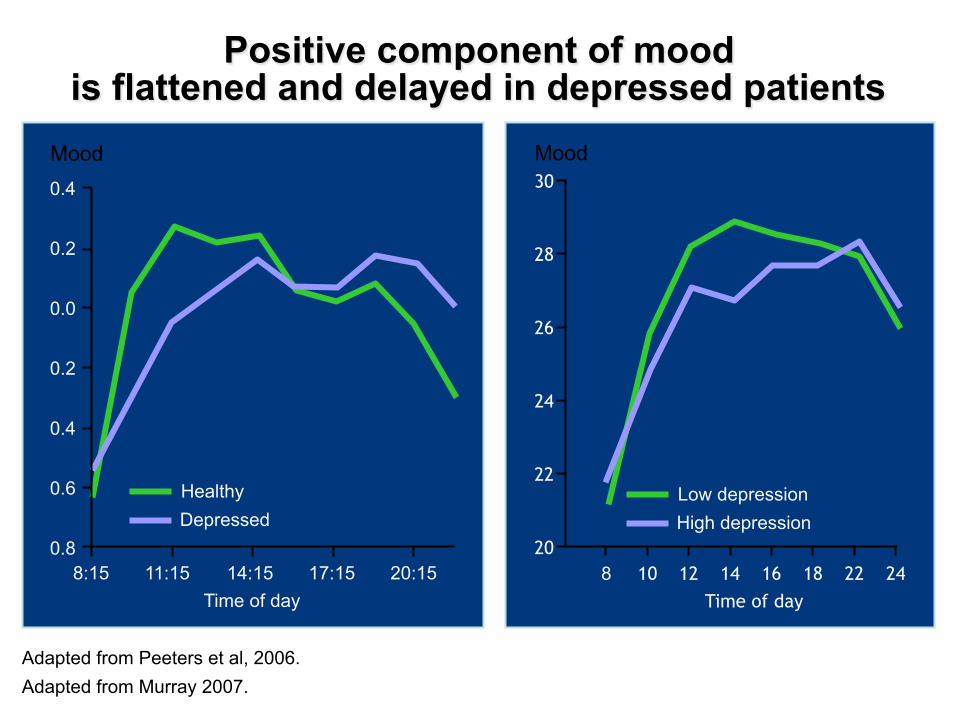
Adapted from Peeters et al, 2006.
Positive component of mood is flattened and delayed in depressed patients
Adapted from Murray 2007.
Mood
Time of day
0.4
0.2
0.0
0.2
0.4
0.6
0.8 8:15 11:15 14:15 17:15 20:15
Healthy Depressed
Mood 30
28
26
24
22
20 8 10 12 14 16 18 22 24
Time of day
Low depression High depression
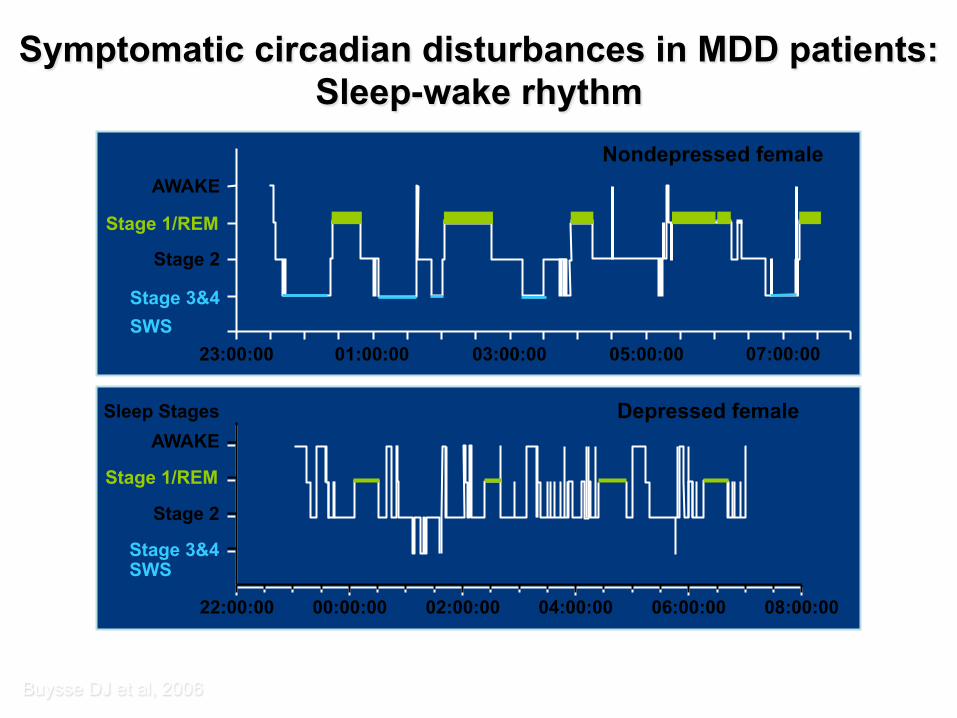
Buysse DJ et al, 2006
AWAKE
Stage 1/REM
Stage 2
Stage 3&4 SWS
23:00:00 01:00:00 03:00:00 05:00:00 07:00:00
Nondepressed female
22:00:00 00:00:00 02:00:00 04:00:00 06:00:00 08:00:00
AWAKE
Stage 1/REM
Stage 2
Stage 3&4 SWS
Sleep Stages Depressed female
Symptomatic circadian disturbances in MDD patients: Sleep-wake rhythm

Depression and circadian rhythms
� Circadian rhythms play a fundamental role in the regulation of human behavior and physiology
� The key structure for the organization of circadian rhythms is the Suprachiasmatic nucleus, which contains high density of melatonergic MT1 /MT2 and 5HT2C receptors
� Core symptoms of depression, i.e., depressed mood and sleep disturbances, follow circadian variations and are experienced by the depressed patients
Conclusion
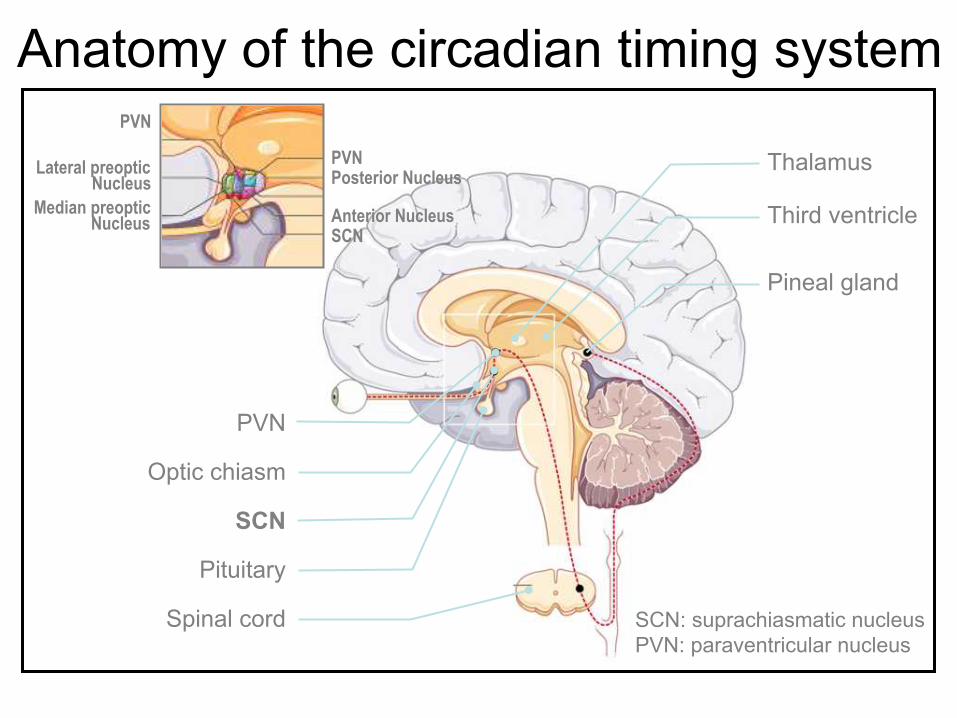
Anatomy of the circadian timing system
Corpus callosum
Ventromedial Nucleus
Thalamus
Third ventricle
Pineal gland
PVN
Optic chiasm
SCN
Pituitary
Spinal cord SCN: suprachiasmatic nucleus PVN: paraventricular nucleus
PVN Posterior Nucleus
Anterior Nucleus SCN
PVN
Lateral preoptic Nucleus
Median preoptic Nucleus

Agomelatine A unique pharmacology

Receptor profile of agomelatine
� Negligible affinity for other receptors or transporters
� No down regulation of 5HT1A receptors
� No influence on extracellular serotonin levels
IC50 1.3x10-10 MT1
IC50 4.7x10-10 MT2
5-HT2C receptors Melatonergic receptors
IC50 2.7x10-7 5-HT2C
Antagonist Agonist
Millan et al, 2003; Hanoun et al, 2004; Audinot et al, 2003
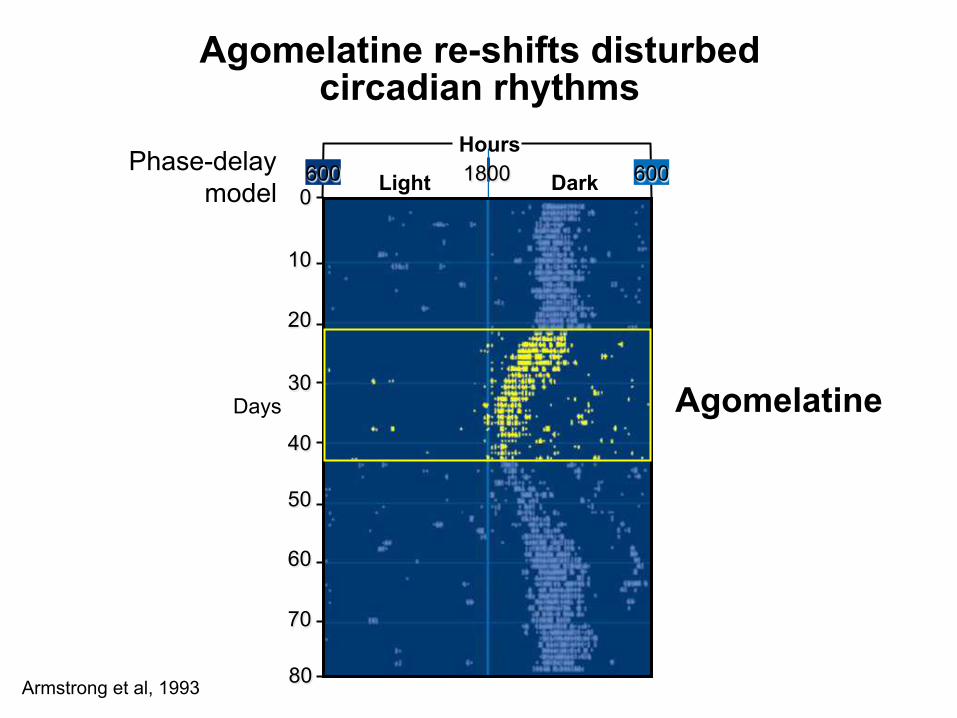
Agomelatine re-shifts disturbed circadian rhythms
Armstrong et al, 1993
Agomelatine Days
80
0
10
20
60
70
50
30
40
Phase-delay model Dark Light 1800
Hours 600 600
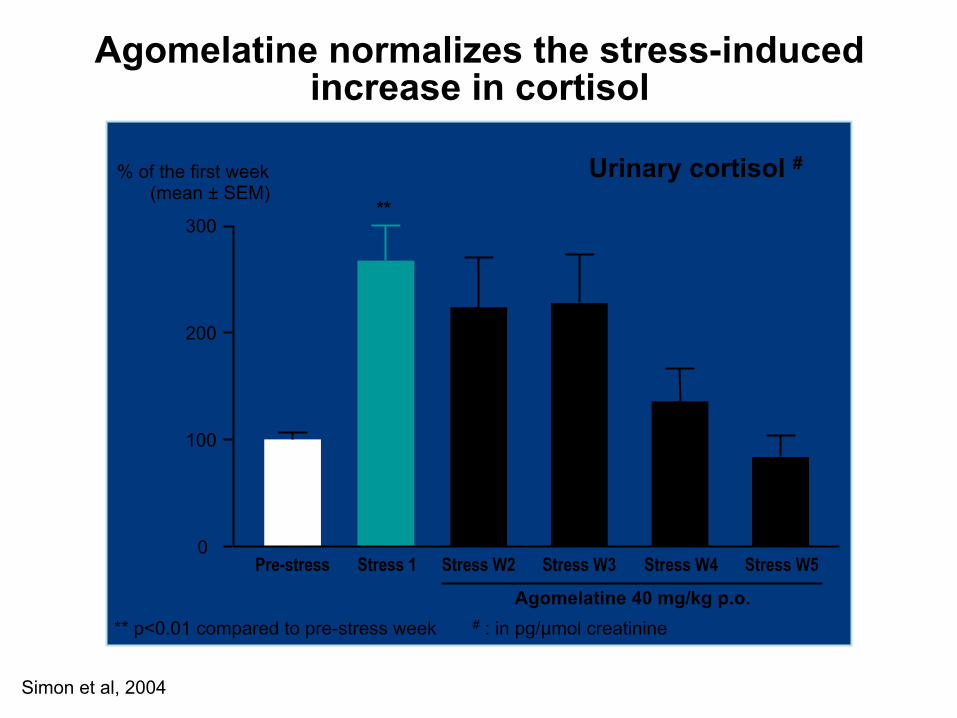
Simon et al, 2004
Agomelatine normalizes the stress-induced increase in cortisol
** p<0.01 compared to pre-stress week # : in pg/µmol creatinine
Urinary cortisol #
Pre-stress Stress W2 Stress W3 Stress W4 Stress W5 0
100
200
300
Stress 1
**
% of the first week (mean ± SEM)
Agomelatine 40 mg/kg p.o.
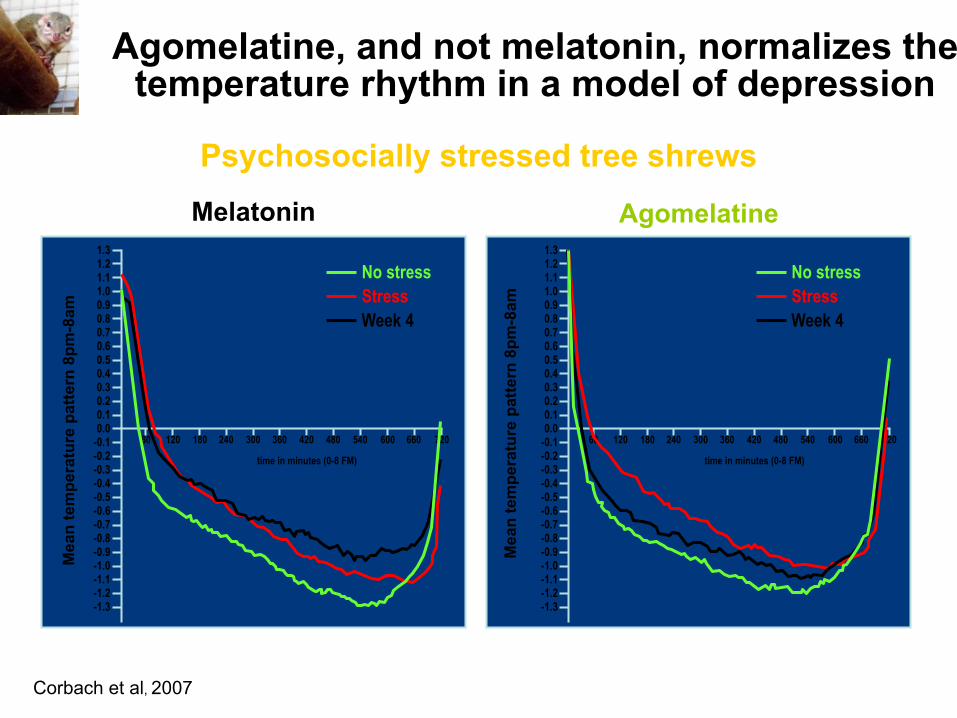
Corbach et al, 2007
Melatonin Agomelatine
60 120 180 300 240 420 360 480 540 600 660 720
time in minutes (0-8 FM)
No stress Stress Week 4
1.3 1.2 1.1 1.0 0.9 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0.0
-0.1 -0.2 -0.3 -0.4 -0.5 -0.6 -0.7 -0.8 -0.9 -1.0 -1.1 -1.2 -1.3
Mea
n te
mpe
ratu
re p
atte
rn 8
pm-8
am
60 120 180 300 240 420 360 480 540 600 660 720
time in minutes (0-8 FM)
1.3 1.2 1.1 1.0 0.9 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0.0
-0.1 -0.2 -0.3 -0.4 -0.5 -0.6 -0.7 -0.8 -0.9 -1.0 -1.1 -1.2 -1.3
No stress Stress Week 4
Mea
n te
mpe
ratu
re p
atte
rn 8
pm-8
am
Agomelatine, and not melatonin, normalizes the temperature rhythm in a model of depression
Psychosocially stressed tree shrews

� All three receptors show a high density in the suprachiasmatic nucleus1-3
� All three receptors are involved in circadian rhythm regulation4-6
� MT1 and 5-HT2C receptors are expressed in a circadian manner7,8
� Melatonergic receptors have a beneficial effect on sleep and antagonism at 5-HT2C receptors promotes slow-wave sleep9,10
1 Boess and Martin, 1994; 2 Moyer and Kennaway, 1999; 3 Dubocovich et al, 1996; 4 Kennaway and Moyer, 1998; 5 Varcoe et al 2003; 6 Barret et al, 2003 minireview; 7 Holmes et al, 1997; 8 Masana et al, 2000; 9 Dugovic and Wauquier, 1987; 10 Dugovic et al, 1989; * Papp et al, 2003
Antidepressant efficacy of agomelatine involves melatonergic MT1/MT2, and 5-HT2C receptors*
Potential synergy of melatonergic (MT1/ MT2) and 5-HT2C receptors
Agomelatine, a new pharmacological approach

A+: agonist A-: antagonist ↓: desensitization +: reuptake inhibition
TCA
mirtazapine
escitalopram
fluoxetine
Agomelatine
paroxetine
(weak)
+
Histaminergic
H1
ACh
M1
Dopaminergic
Reuptake Inhibition
Noradrenergic
Reuptake Inhibition α2 α1
Serotonergic
Reuptake inhibition 5-HT3 5-HT2A 5-HT2C
Melatonergic
MT2
+
+
+
+
+
A− A− A− A−
A−
A− A− A− A−
A+ A−
A−
A−
A− A−
A+
5HT1A
venlafaxine + +
duloxetine + + +
MT1
↓
↓
↓
↓
↓
↓
Agomelatine, a unique receptor profile: MT1/MT2 agonist and 5-HT2C antagonist
Adapted from Stahl 2008

Central distribution of melatonergic MT1/MT2 and 5-HT2C receptors*
* in situ hybridation, immunocytochemistry in rodents or in human
MT1 (Mel1A) MT2 (Mel1B) 5-HT2C
SCN SCN SCN
Hippocampus Hippocampus Hippocampus
Frontal cortex Hypothalamus Frontal, temporal cortex
Hypothalamus Retina Striatum Cerebellum Cerebellum Olfactory bulb Retina Septum Thalamus Thalamus
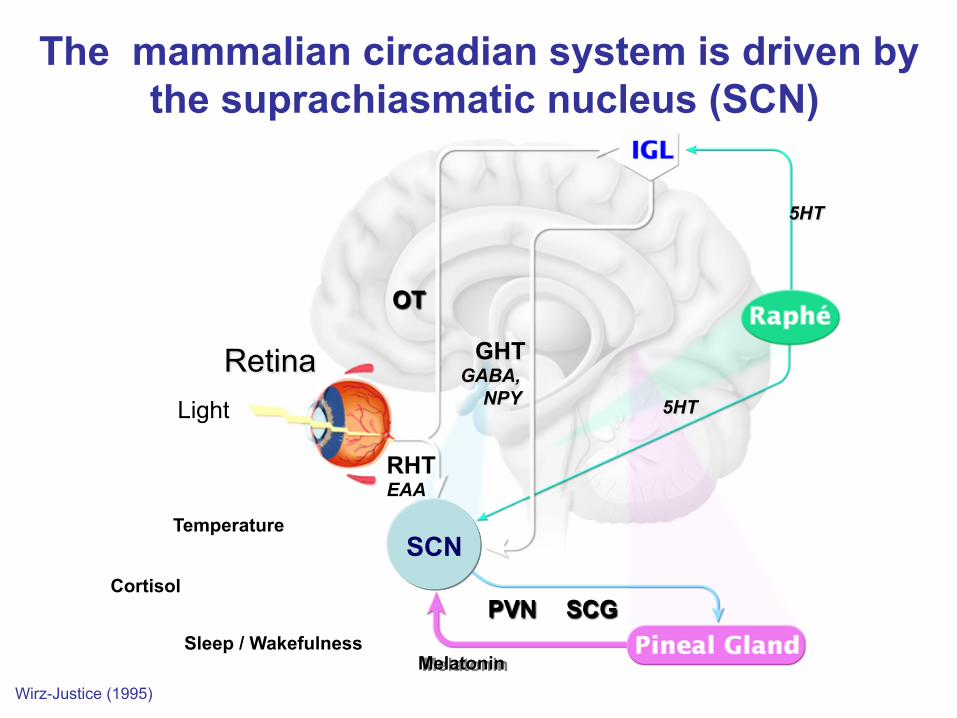
The mammalian circadian system is driven by the suprachiasmatic nucleus (SCN)
Wirz-Justice (1995)
Light
Retina
RHT EAA
OT
GHT GABA,
NPY 5HT
PVN SCG
Melatonin
5HT
Temperature
Sleep / Wakefulness
Cortisol
SCN

Evidence for circadian rhythm disturbances in depression
Wirz-Justice, Int Clin Psychopharmacol. 2006; Turek, Int Clin Psychopharmacol. 2007; McClung, Pharmacol Ther. 2007
Depression severity is correlated with circadian dysregulation
Emens et al, Psychiatry Research (2009)
• Diurnal mood changes, anxiety
• Impaired day time functioning, energy level, concentration
• Sleep disorders: difficulties initiating and maintaining sleep, early awakening
• Physiological functions: loss of appetite, core body temperature
• Biological parameters : prolactin, cortisol, growth hormone, melatonin …

• Dissection of the effects of agomelatine:
– Glutamatergic system
– Neurotrophic factor: BDNF
– Early genes: Arc
– Neuronal survival
Evidence of synergy between MT1, MT2 and 5HT2C receptors

↑ DA ↑ NA
Resynchronization of circadian rhythms
Agomelatine MT1/ MT2 receptor
agonist 5HT2C receptor
antagonist
MT1 MT2
Resynchronization of circadian rhythms at intracellular level
Racagni et al. in preparation
Circadian expression
↓ Glutamate release ↑ BDNF/Arc
↑ Neurogenesis
D2 α2
Antidepressant efficacy

• Agomelatine has a unique receptorial profile: melatonergic MT1 and MT2 receptor agonist and 5HT2C receptor antagonist
• MT1, MT2 and 5HT2C receptors act in synergy, demonstrated at the intracellular signaling levels
• By its unique profile, agomelatine provides antidepressant efficacy through the resynchronization of circadian rhythms of depressed patients
Conclusion

Lo stress è una risposta fisiologica normale e positiva, necessaria all’evoluzione della specie e individuale. E’ un insieme di reazioni che, scatenate da uno stressor esterno, partendo dall’ "alto“, cioè dalla percezione intellettuale di un pericolo o di una situazione sconosciuta, promuove una reazione a catena che coinvolge le funzioni inferiori, il SNV e infine il sistema endocrino.

Suo scopo principale: mettere l’individuo in “condizione di lotta o di fuga”.
Si distinguono due tipi di stress : l’eustress positivo o di adattamento e quello negativo o distress. Condizioni di distress sono quelle che generano uno stato di preallarme ma non consentono di giungere a una risoluzione del conflitto: la perdita di un congiunto, la perdita del lavoro, un trasloco. Esistono stress fisici (uno shock elettrico, l’esposizione al freddo, ecc…), metabolici (ipoglicemia), psicologici (una prova d’esame), psicosociali (un evento di perdita o lutto).

Oltre alla natura dello stressor sono molto importanti anche l’intensità, la frequenza e la durata dello stimolo nel condizionare l’entità della risposta: stressor troppo potenti, frequenti e prolungati sono in grado di superare la possibilità di resistenza dell’organismo, e di iniziare un processo patologico. Altro aspetto dello stressor è il grado di novità, prevedibilità ed evitabilità dello stimolo. Qualcosa di NON RICONOSCIBILE o imprevedibile e inevitabile induce una risposta più ampia.

• La reazione risulta dal patrimonio genetico individuale e dall’ "imprinting psicobiologico" dovuto a precedenti esposizioni a stressor di varia natura. Determinanti età, sesso, attività dei sistemi nervoso e immunitario e il profilo di personalità. L’invecchiamento è una fase di ridotta energia adattativa e quindi di aumentata risposta allo stress. L’ambiente è sorgente di stimoli stressogeni, sia interni che esterni : geoclimatici, di interazione sociale ecc.

Tra stimolo e reazione non c’è un rapporto di tipo lineare. L’attivazione avrebbe inizio con la percezione dell’evento stressante da parte del SNC, che sulla base di precedenti esperienze, proprio come per gli stimoli dolorifici, dà inizio a modificazioni che non sarebbero legate strettamente allo stressor. Il modello recente di Herman e Cullinan (1997) discrimina il tipo di risposta a seconda della capacità dei circuiti seguiti dallo stressor per accedere al nucleo dell’ipotalamo in maniera diretta o indiretta. .

• Le ultime ricerche evidenziano implicati nella cascata della risposta di stress numerosi neuro peptidi (Beta endorfina, Met-enkefalina, GH-RH, Somatostatina, Vasopressina, Ossitocina, Prolil-Lysin-Glicinamide (PLG), Sostanza P, Angiotensina II, LH-RH, CCK, Bombesina-Gastrin Releasing Peptide, NeuropeptideY (NPY), Atriopeptide, NAF, Diazepam Binding Inhibitor e TRH che , in cronico, partecipano nell’eziopatogenesi di patologie somatiche.

Ipotalamo e bioritmo metabolico all’interno del SNC, l’ipotalamo possiede
un ruolo centrale nel controllo dell’appetito e del metabolismo energetico. La sua attività deriva dalla complessa integrazione di differenti impulsi modulatori centrali e periferici, quali diversi nutrienti, la vista, l’olfatto, l’esperienza, lo stile di vita e le condizioni ambientali.

Clin Physiol Funct Imaging. 2008 Jul;28(4):270-6. – Association of serotonin transporter promoter
regulatory region polymorphism and cerebral activity to visual presentation of food. Kaurijoki S, Kuikka JT, Niskanen E, Carlson S, Pietiläinen KH, Pesonen U, Kaprio JM, Rissanen A, Tiihonen J, Karhunen L.Department of Clinical Nutrition, University of Kuopio, Finland.
– Recent functional magnetic resonance imaging (fMRI) studies have revealed links between genetic polymorphisms and cognitive and behavioural processes. Serotonin is a classical neurotransmitter of central nervous system, and it is connected to the control of appetite and satiety. In this study, the relationship between the functional variation in the serotonin transporter gene and the activity in the left posterior cingulate cortex (PCC), a brain area activated by visual food stimuli was explored. Thirty subjects underwent serial fMRI studies and provided DNA for genetic analyses. Subjects homozygous for the long allele exhibited greater left PCC activity in the comparison food > non-food compared with individuals heterozygous or homozygous for the short allele. The association between genotype and activation was linear, the subjects with two copies of the long allele variant having the strongest activation. These results demonstrate the possible genetically driven variation in the response of the left PCC to visual presentation of food in humans.

• Il corpo sperimenta i vantaggi della disintossicazione e la crisi di guarigione ad ogni cambiamento di stagione (ci si ammala più frequentemente in quei periodi dell'anno).
• Ci sentiamo così bene, infatti, che il corpo decide di scavare ancora un po' più in profondità per eliminare alcune delle tossine più vecchie, e quindi ci sentiamo peggio. Questo è il ciclo continuo del processo di guarigione.

• Ricerche nel settore del bioritmo hanno dimostrato che il corpo ha bioritmi di 14 giorni per l'autoregolazione dello stato fisico, 28 giorni per lo stato emotivo e 33 giorni per lo stato mentale. Il corpo mentre conduce il suo processo di disintossicazione può avere una serie di alti e bassi. Un giorno può avviare una consistente eliminazione di tossine e ci sentiamo malissimo. Il giorno successivo, le tossine sono state eliminate e ci sentiamo eccezionalmente bene.

Sindrome metabolica L’Organizzazione Mondiale della Sanità (OMS) definisce “globesity” una sempre
più diffusa “epidemia di obesità e diabete”.
Alla sua base c’è un’alimentazione troppo spesso caratterizzata dal consumo
eccessivo di proteine, grassi saturi, zuccheri e sale, a cui si aggiunge una crescente sedentarietà e una sempre
minore attitudine al movimento e all’attività fisica, sia sul luogo di lavoro
sia nel tempo libero.

la “sindrome metabolica” è:
una pericolosa associazione di sintomi che vede coesistere
ipertensione, obesità, diabete e dislipidemia in soggetti che
diventano perciò ad altissimo rischio di eventi cardiovascolari.

“rischio” di sindrome metabolica se presenti tre o più dei seguenti sintomi:
1) indice di massa corporea (rapporto altezza/ peso) superiore a 30; 2) girovita superiore a 102 cm per gli uomini e a 88 cm per le donne; 3) ipertensione arteriosa superiore a 130/ 90;
4) glicemia a digiuno superiore a 110 mg/dl; 5) colesterolemia superiore a 200 mg/dl; 6) trigliceridi superiori a 150 mg/dl.

Le conseguenze della sindrome metabolica: • oltre la metà della popolazione italiana (adulta e
giovanile) è in sovrappeso • 4 milioni di persone (il 25% in più rispetto a 10
anni fa) risultano obese. • casi di diabete insulino-resistente raddoppiati
(6-8% degli italiani), • 1 italiano su 3 soffre di ipertensione arteriosa e
1 su 5 di ipercolesterolemia, • i decessi per malattie cardiovascolari
ammontano ogni anno a circa 250 mila, prima causa di morte (40% del totale)

• La policistosi ovarica, o sindrome dell’ovaio policistico, è caratterizzata da resistenza all’insulina con iperinsulinemia compensatoria. Se si associa la sindrome metabolica compaiono iperandrogenemia, più bassi livelli sierici di SHBG (Sex hormone binding globulin) e più alta prevalenza di acantosi nigricans, tutte caratteristiche che possono riflettere una più grave resistenza all’insulina.

• Il metabolismo corporeo si divide in due fasi :
• nella mattina fino al primo pomeriggio si ha la fase catabolica con produzione di energia
• nel tardo pomeriggio, serata e durante la notte si ha la fase anabolica in grado di recuperare danni cellulari avvenuti durante la giornata, di fare la sintesi delle proteine e limitare al massimo la domanda di energia (snc?).
• All'alba lo stomaco produce un ormone la ghrelina, che stimola l'assunzione di carboidrati. La colazione deve contenere almeno 25-30 grammi di carboidrati per bloccare l'effetto della ghrelina.

• Se non si fa colazione con questa dose di carboidrati, la ghrelina rimane attiva spingendo verso una disordinata alimentazione giornaliera fino alla grande abbuffata della cena. Mangiare molto a cena crea difficoltà alla qualità del sonno. Pertanto durante la notte non si dorme bene, si tende a russare. Il tutto porta ad una maggiore produzione di ghrelina durante la parte terminale della notte, che spinge a mangiare carboidrati. Questa situazione viene ripetuta tutti i giorni con evidenti danni cellulari e aumento di peso corporeo.
• Una buona qualità del sonno fa diminuire la secrezione della ghrelina, con una minore fame verso i carboidrati. Per dimagrire occorre fare un profondo sonno restauratore e combattere l'insonnia.

• Un eccesso di carboidrati ingeriti a cena (pane, pasta, dolci.... ) aumentano la glicemia e la secrezione di insulina, con un’azione diretta sull'enzima attivo nel fegato per la produzione di colesterolo endogeno, attorno alle 2-3 del nuovo giorno. L'eccesso di insulina nella notte, nella fase anabolica del metabolismo cellulare, porta ad un maggiore rilascio delle lipoproteine VLDL da parte del fegato, con un netto incremento del colesterolo totale e –LDL, favorente l'aterosclerosi e malattie cardio-vascolari. A cena meglio meno carboidrati e più proteine, assecondando la fase anabolica, ma la loro associazione favorirebbe i primi.


Programma AA.2010-2011 • Definizione di PNEI. Il Sistema nervoso: le cellule del SN e loro
connessioni; il tronco encefalo, diencefalo, SNV. Sistema endocrino: gli ormoni, loro funzioni e comunicazioni.
Controllo cerebrale sul Sistema endocrino. Assi neuroendocrini: ipotalamo-ipofisi, ipotalamo-ipofisi-surreni, ipotalamo-ipofisi-tiroide, ipotalamo-ipofisi-gonadi. Sistema immunitario: cenni di immunologia (teoria della selezione clonale, immunità naturale ed acquisita, sistema HLA, sistema immunitario come organo di senso, produzione di ormoni da parte del sistema immunitario che fungono da mediatori tra cervello e sistema endocrino). Cervello e psiche. La grande connessione: Sistema nervoso ed immunità; sistema dello stress; sistema endocrino e cervello. Endocrinosenescenza e P.N.E.I. DCA prototipo di PNEIG.

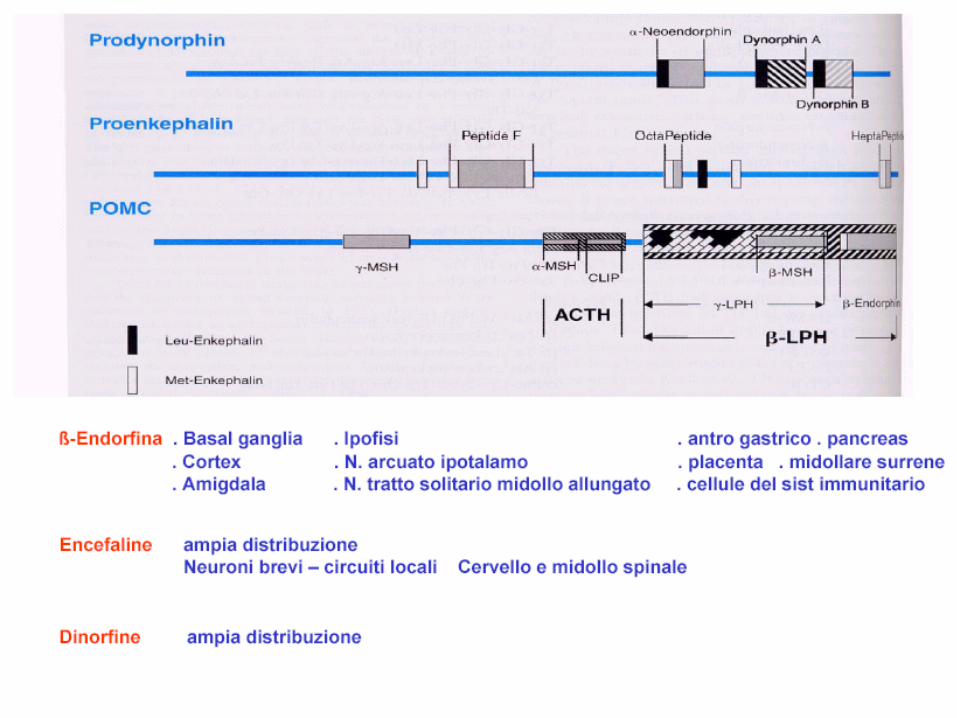
• Negli anni ‘70 la scoperta di recettori specifici soprattutto nelle aree deputate alla percezione dolorifica del SNC confermò la capacità dell’organismo di sintetizzare "morfine endogene" che furono chiamate endorfine. Questi peptidi, sintetizzati nel SNC, nell'ipofisi, nei surreni e in alcuni tratti dell'apparato digerente, sono implicati nella reazione allo stress e dotati di una potente attività analgesica ed eccitante, simile a quella della morfina e di altre sostanze oppiacee.
• Attualmente si conoscono quattro distinte classi di endorfine, dette rispettivamente "alfa", "beta", "gamma" e "delta".

• I neuroni del sistema oppiode si dividono in : • neuroni ad assoni lunghi (soma nel n. arcuato
dell’ipotalamo medio-basale), che sintetizzano la proopiomelanocortina (POMC) di 241 aa
• neuroni ad assoni corti che sintetizzano proenkefalina, precursore delle enkefaline deputate alla percezione del dolore (lamine I e II del midollo spinale, trigemino, s. grigia periacqueduttale), ed alla modulazione del comportamento affettivo ( amigdala, ippocampo, locus coeruleus).
• neuroni ad assoni corti dinorfinergici, che sintetizzano prodinorfina precursore della dinorfina

• La porzione terminale dell’ ormone ipofisario POMC (residuo 61-91 AA.) costituisce la Beta lipotropina da cui originano il beta MSH e le Beta-endorfine (da cui poi le alfa e le gamma)
- dalla porzione media della POMC per scissione proteolitica derivano la metenkefalina e l’ACTH da cui l’Alfa MSH.
• Le Enkefaline: si dividono in met-enkefalina RF (eptapeptide) e met-enkefalina RGL (octopeptide).
• Le dinorfine derivano dalla prodinorfina che da origine a 7 peptidi da cui derivano la dinorfina A = met-enkefalina + arg-arg-lle-arg-pro-lys-leu-lys-trp-asp-asn-gln e la dinorfina B.
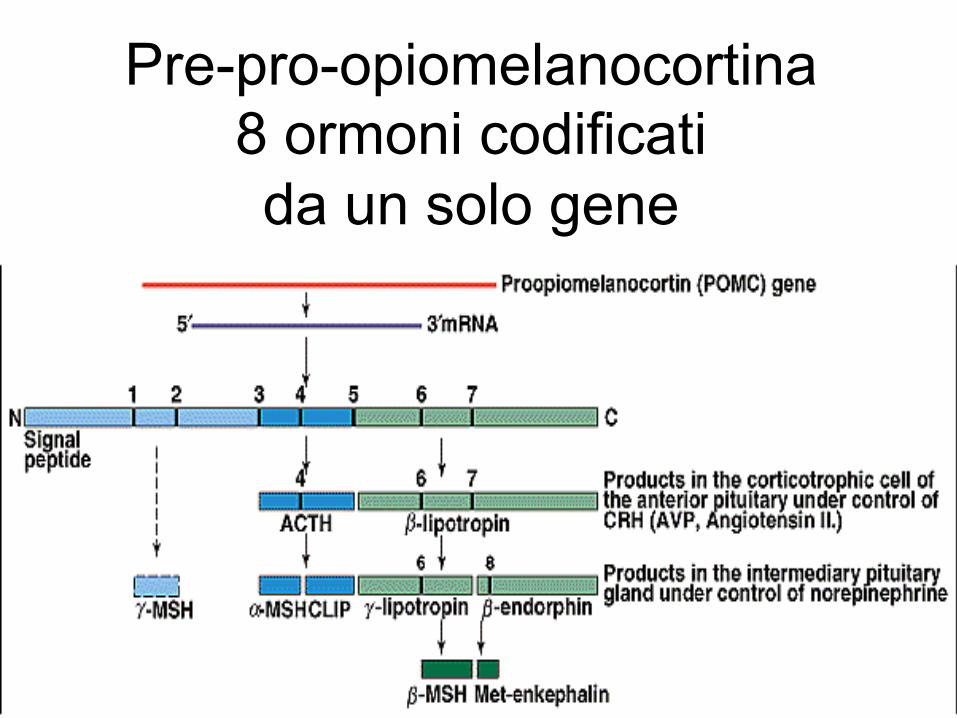
Pre-pro-opiomelanocortina 8 ormoni codificati da un solo gene

LE ENDORFINE Le endorfine originano dalla PROOPIOMELANO
CORTINA (POMC) un polipeptide di 239 aa
La porzione terminale (91 AA) della POMC costituisce la β-lipotropina (β-LPH), peptide che ha la capacità di liberare acidi grassi dai tessuti adiposi Dalla β-lipotropina si forma la β-endorfina e da questa, successivamente la α-endorfina e la γ-endorfina Dalla β-lipotropina deriva inoltre un peptide chiamato ormone β-melanoforo (β-MSH)

La porzione n-terminale della POMC contiene anche un gruppo di peptidi chiamati γ-MSH, γ1 MSH, γ2 MSH, per analogie strutturali con l’α-MSH ed il β-MSH. La loro attività biologica è ancora sconosciuta ma sembra che abbiano un’attività modulatoria nei confronti dell’ACTH e del MSH o su quella delle endorfine La porzione centrale o media della POMC dà origine all’ACTH e all’α-MSH che sono due peptidi non dotati di attività intrinseca di tipo oppioide, ma possono interagire in senso funzionale con le endorfine modulandone l’azione

La massima parte di α-MSH si forma nell’ipofisi intermedia dall’ACTH formato preferenzialmente nell’ipofisi anteriore. La POMC contenuta a livello ipofisario e quella contenuta nel S.N.C. costituiscono due “pools” indipendenti. 1) Le Endorfine ipofisarie hanno un ruolo principalmente ormonale 2) Le Endorfine di origine centrale hanno un ruolo di neurotrasmettitore o neuromodulatore Ci può essere tuttavia un ruolo di “controllo” da parte delle endorfine ipofisarie sulle funzioni centrali, direttamente o indirettamente, mediante l’immissione di questi peptidi dall’ipofisi nel cervello attraverso un flusso portale retrogrado

Le endorfine sono contenute nelle cellule corticotrope dell’Ipofisi anteriore e intermedia, cioè quelle che contengono ACTH con cui condividono il meccanismo di liberazione . Nel S.N.C. sono contenute in neuroni lunghi il cui soma è concentrato soprattutto nell’ipotalamo medio-basale a livello del nucleo arcuato. Dall’ipotalamo i neuroni endorfinergici si proiettano anteriormente e caudalmente terminando in diverse aree cerebrali:
1) Setto anteriore 2) Nucleo paraventricolare 3) Nucleo paraventricolare del talamo 4) Nucleo parabrachiale 5) Materia grigia periacqueduttale

DISTRIBUZIONE DELLE ENKEFALINE
a) Globus pallidus a) Ipofisi posteriore b) Gangli della base b) Ganglio celiaco c) Amigdala c) Ganglio superiore cervicale d) Ipotalamo d) Ghiandole salivari e) Cervelletto e) Tratto gastro-intestinale f) Substantia nigra f) Midollare del surrene g) Talamo g) Sangue h) Corteccia h) Liquor
Le enkefaline sono contenute in neuroni ad assone corto o interneuroni che hanno il corpo cellulare e le terminazioni
all’interno dello stesso nucleo cerebrale
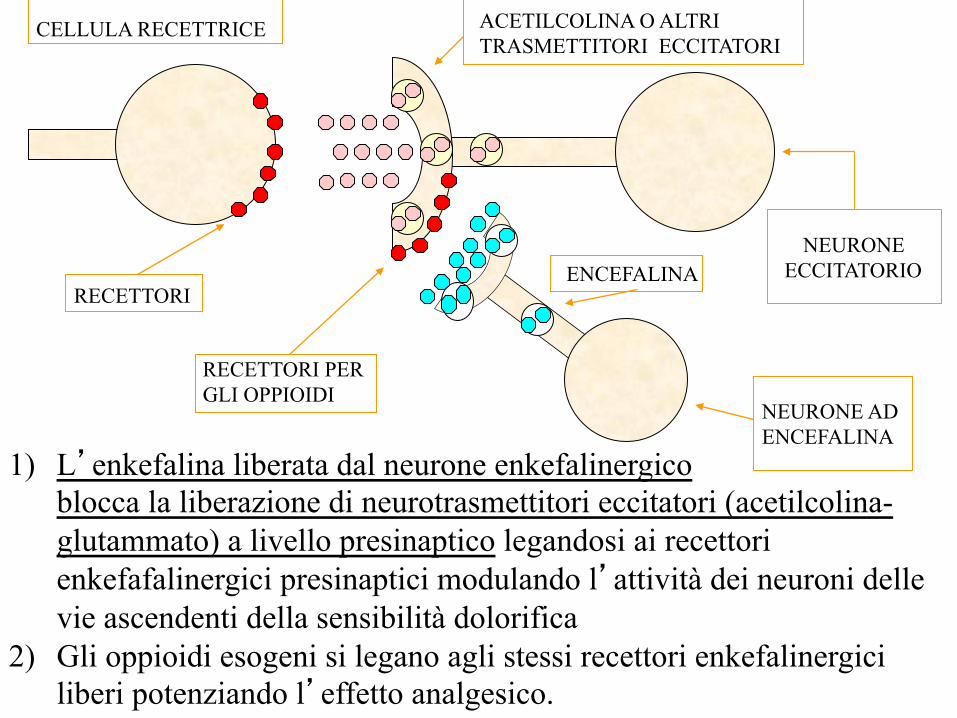
ENCEFALINA NEURONE
ECCITATORIO
NEURONE AD ENCEFALINA
ACETILCOLINA O ALTRI TRASMETTITORI ECCITATORI
CELLULA RECETTRICE
RECETTORI
RECETTORI PER GLI OPPIOIDI
1) L’enkefalina liberata dal neurone enkefalinergico blocca la liberazione di neurotrasmettitori eccitatori (acetilcolina-glutammato) a livello presinaptico legandosi ai recettori enkefafalinergici presinaptici modulando l’attività dei neuroni delle vie ascendenti della sensibilità dolorifica
2) Gli oppioidi esogeni si legano agli stessi recettori enkefalinergici liberi potenziando l’effetto analgesico.

LE DYNORFINE Derivano da un precursore, la prodynorfina che contiene nella sua struttura, a partire dalla porzione n-terminale, la sequenza di tre peptidi:
α-neoendorfina (10AA) a) Neoendorfina β-neoendorfina (9 AA) b) Dynorfina A (17 AA)
c) Dynorfina B (13 AA)
Tutte le dynorfine contengono la sequenza della leu-enkefalina nella porzione n-terminale che è alla base della loro attività oppioide

DISTRIBUZIONE DELLE DYNORFINE
1) IPOFISI POSTERIORE
2) S.N.C. a) Ipotalamo nucleo magno cellulare b) Striato via nigro-striatale c) Mesencefalo d) Ippocampo e) Ponte f) Midollo allungato
corno dorsale g) Midollo spinale zona marginale
lamine profonde h) Corteccia i) Cervelletto
3) TESSUTI PERIFERICI a) Tratto gastro-intestinale b) Pancreas c) Polmoni d) Cuore e) Muscolatura scheletrica f) Fegato g) Reni h) Testicoli i) Ovaie

APPARATO RECETTORIALE del SISTEMA OPPIOIDE:
AGONISTI RECETTORI
enkefaline δ-delta endorfine µ-mu dinorfine κ-kappa

RECETTORI OPPIOIDI E AZIONI DA QUESTI MEDIATE RECETTORI AZIONI MEDIATE µ1 Analgesia a livello sopraspinale: area grigia periacqueduttale, nucleo del rafe
Rilascio dell’ormone prolattina. Turnover di acetilcolina Catalessia.
µ2 Depressione respiratoria. Rilascio dell’ormone della crescita
Turnover della Dopamina nel cervello Inibizione delle contrazioni dell’ileo di cavia Motilità gastrointestinale. Effetti cardiovascolari Controllo dell’appetito.
δ Analgesia a livello spinale. Turnover della dopamina nel cervello
Inibizione delle contrazioni del dotto deferente di topo Rilascio dell’ormone della crescita. Controllo dell’appetito.
κ Analgesia a livello spinale. Inibizione del rilascio di vasopressina.
Sedazione. Controllo dell’appetito. σ Inibizione delle contrazioni del dotto deferente del topo ε Effetti psicotomimetici. Possibile interazione con i recettori per il
N-metil-D-aspartato

LOCALIZZAZIONE DEI RECETTORI PER GLI OPPIOIDI
RECETTORI CHE MEDIANO L’ANALGESIA corna posteriori del midollo spinale
grigio periacqueduttale talamo
RECETTORI CHE MEDIANO GLI EFFETTI SU RESPIRAZIONE, TOSSE, VOMITO E DIAMETRO PUPILLARE parte ventrale del tronco encefalico RECETTORI CHE MODULANO LA SECREZIONE ENDOCRINA ipotalamo RECETTORI CHE MEDIANO GLI EFFETTI SULL’UMORE E SUL COMPORTAMENTO strutture limbiche
(ippocampo,amigdala, etc.)

INTERAZIONE TRA RECETTORI OPPIOIDI
Lo studio delle interazioni tra recettori oppioidi nella modulazione delle risposte agli agonisti, ha evidenziato: Una cooperazione tra recettori µ e recettori δ per l’azione analgesica e respiratoria degli agonisti oppioidi I recettori µ e κ possono invece mediare reazioni opposte

SISTEMA OPPIOIDE: MODALITA’ D’AZIONE
intervento Localizzazione presinaptico postsinaptico
Meccanismo riduzione incremento influx Ca2+ conduttanza k+
Effetto diminuzione riduzione
release risposte mediatori al mediatore

Diversi effetti dell’attivazione dei recettori oppioidi presenti in sede pre- o post-sinaptica. L’attivazione dei recettori µ, κ, δ sui terminali presinaptici delle fibre afferenti nocicettive, riduce il rilascio di trasmettitori eccitatori coinvolti nel dolore (glutammato, sostanza P, etc.). L’attivazione dei recettori µ in sede postsinaptica, aumenta la conduttaza al K+ determinando l’insorgenza di potenziali postsinaptici inibitori (IPSP) e riduzione conseguente della scarica dei neuroni diretti ai centri superiori.

SIGNIFICATO BIOLOGICO del SISTEMA OPPIOIDE: 1) Modulazione delle afferenze nocicettive: a) a livello spinale: trasmissione, b) a livello sovratalamico: integrazione. 2) Comparsa, per attivazione protratta, della dipendenza fisica
e psichica. 3) Modulazione a livello centrale di funzioni vegetative
(respiratoria, cardio- circolatoria, gastrointestinale) e di funzioni neuroendocrine.
4) Modulazione a livello periferico della funzione
immunologica

ENDOMORFINE solo nel SN?
Endomorfina 1 Neuropeptidi di più recente scoperta Anni 80
Endomorfina 2 Presentano l’aminoacido tirosina in posizione 1 e la fenilalanina in posizione 3 o 4, requisito fondamentale per il legame ai recettori oppioidi. Le endomorfine mostrano altissima affinità e selettività per i recettori µ

DISTRIBUZIONE DELLE ENDOMORFINE
Corna dorsali del midollo spinale Nucleo spinale del trigemino Nucleo ambiguo Nucleus accumbens Setto Nuclei talamici Ipotalamo Amigdala Locus coeruleus Sostanza grigia periacqueduttale Sono assenti: Corteccia frontale Striato Ippocampo

NOCICETTINA
La nocicettina (17 AA) detta anche orfanina FQ, OFQ, è un peptide correlato alle dinorfine più per analogie strutturali che funzionali. La nocicettina non possiede in posizione n-terminale la tirosina ed il legame di questo peptide al suo recettore (ORL1) non è antagonizzato dal naloxone così come non lo sono i suoi effetti farmacologici Non esistono ad oggi antagonisti selettivi del recettore ORL1 per studiare le funzioni del sistema OFQ.

DISTRIBUZIONE DELLA NOCICETTINA
Neuroni centrali degli strati superficiali delle corna dorsali del midollo Complesso sensorio del trigemino Sostanza grigia periacqueduttale Nucleo del rafe Il precursore della nocicettina contiene un altro eptadecapeptide bioattivo chiamato nocistatina che attenua vari tipi di sensazioni dolorose pur non agendo sui recettori oppioidi e sul recettore ORL1

SISTEMA OPPIOIDE E CONTROLLO DEL DOLORE L’informazione nocicettiva nasce nei sensori periferici (nocicettori) e arriva ai neuroni del corno posteriore del midollo spinale. Il dolore vero e proprio è la presa di coscienza delle informazioni nocicettive integrate al livello corticale e limbico. Strutture coinvolte nel dolore: Corteccia anteriore cingolata Corteccia somatico sensoriale (SI,SII) Corteccia prefrontale Corteccia insulare Ipotalamo Talamo Sostanza grigia periacqueduttale Cervelletto Gangli della base

Meccanismi di controllo del dolore
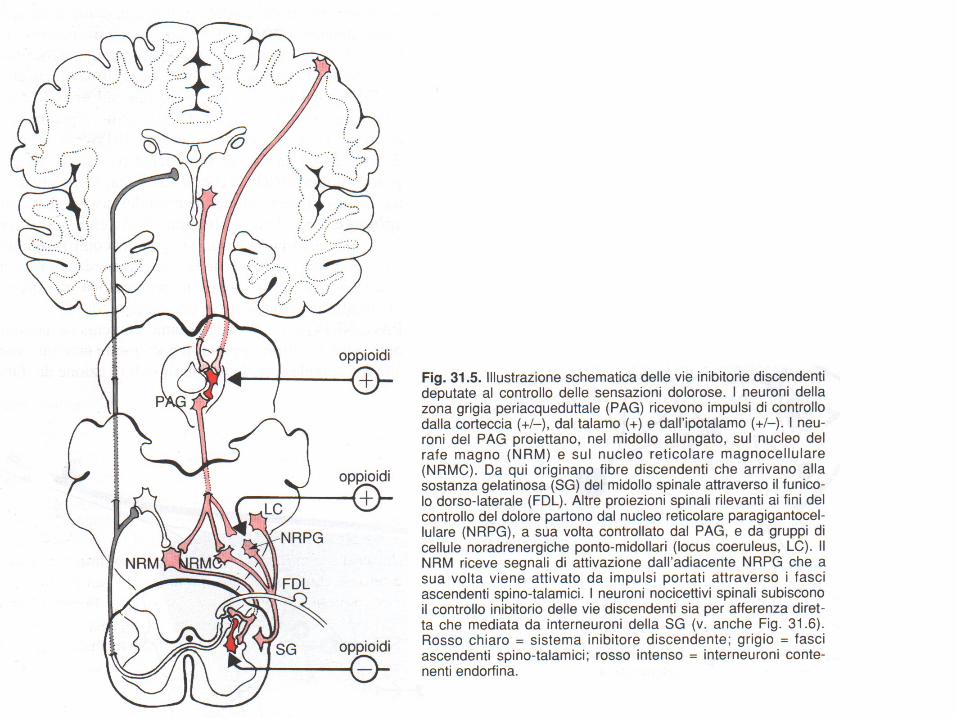
L’informazione dolorifica è sottoposta ad un complesso sistema di controllo endogeno affidato a neurotrasmettitori rilasciati da neuroni spinali e ai fasci nervosi discendenti dai centri superiori.

Controllo spinale del dolore
A livello delle corna dorsali del midollo spinale, nell’area definita sostanza gelatinosa, si trova la prima
stazione di controllo delle afferenze nocicettive.
In questa sede si realizza un vero e proprio cancello di accesso ( gate control ) alle afferenze nocicettive.
Un ruolo di primo piano è esercitato dagli interneuroni oppioidergici (soprattutto encefalinergici) che inibiscono il rilascio di neurotrasmettitori eccitatori dalle terminazioni delle fibre A δ e C.

Vie discendenti dai centri superiori
Vie serotoninergiche e noradrenergiche provenienti dalla corteccia e da aree subcorticali come ipotalamo, talamo, area periacqueduttale e diversi nuclei reticolari, hanno un ruolo fondamentale nel controllo del dolore sia attraverso l’attivazione degli interneuroni oppioidergici della sostanza gelatinosa, sia attraverso l’inibizione diretta de i neuroni de l le v ie neo-e pa leo-spinotalamiche.

• Oltre ad aumentare la tolleranza al dolore le endorfine sono coinvolte nel già ricordato senso di benessere che insorge al termine di un rapporto sessuale
• nella secrezione di GH, ACTH, prolattina, catecolamine e cortisolo implicati nella risposta allo stress
• nella regolazione del sonno • nel controllo dell'appetito e dell'attività
gastrointestinale • nella termoregolazione • nella regolazione del ciclo mestruale
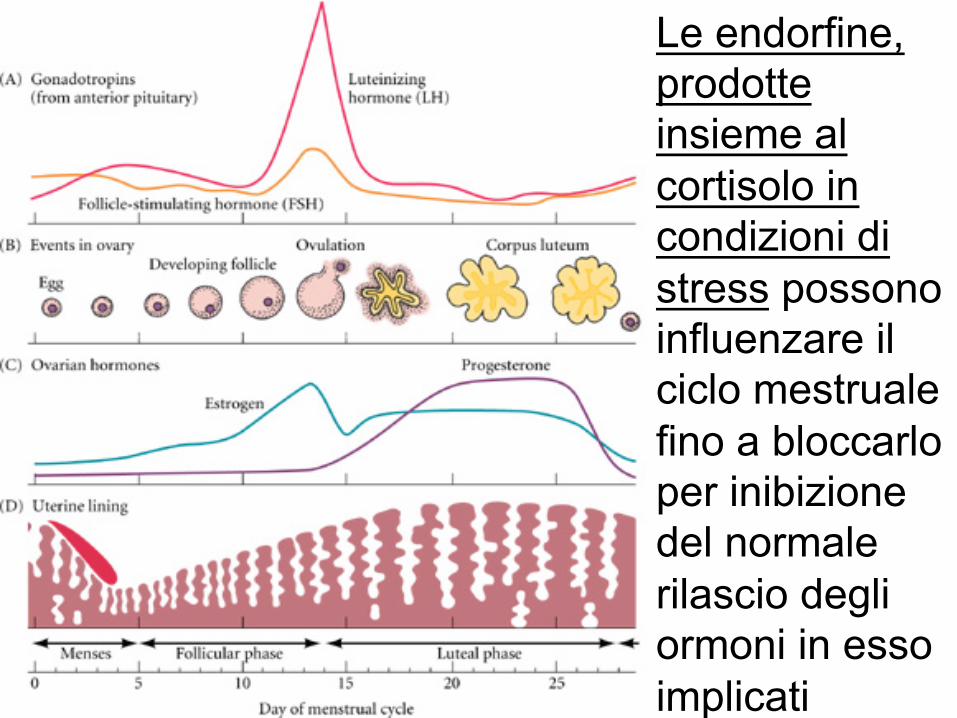
Le endorfine, prodotte insieme al cortisolo in condizioni di stress possono influenzare il ciclo mestruale fino a bloccarlo per inibizione del normale rilascio degli ormoni in esso implicati

• Durante situazioni particolarmente stressanti, per difesa, il nostro organismo rilascia endorfine che da un lato aiutano a sopportare meglio il dolore e dall'altro influiscono positivamente sullo stato d’animo.
• L'interazione di queste sostanze con altri ormoni e neurotrasmettitori è alla base di numerosi aspetti della sfera psicologica e sessuale dell'uomo. Esiste una forte correlazione tra situazioni come tradimenti, maternità, amore, sesso, infatuazione e la quantità di endorfine presente nel sangue.

• La dipendenza da sostanze da abuso, come l'eroina, si spiega proprio nell'inibizione della produzione endogena di endorfine, a cui esse si sostituiscono. In astinenza, i ridotti livelli plasmatici di endorfine producono stanchezza, insoddisfazione e quel malessere generale responsabili del craving. L’esercizio fisico aumenta la sintesi di oppiodi endogeni mediamente del 500%. L’allenamento porta a una più lenta degradazione degli oppiodi endogeni aumentando la tolleranza alla fatica prolungata. Le endorfine migliorano la coordinazione motoria e il reclutamento delle fibre muscolari.

• Il MSH agisce sui melanociti cutanei, inducendo la sintesi della melanina. E’ ormai certo che l’α-MSH è presente nel SNC, specialmente nell’area dell’ipotalamo in connessione con l’ipofisi. Sia nell’uomo che negli animali l’α-MSH può giocare il ruolo di neuropeptide in tutta una serie di funzioni fisiologiche non correlate con la pigmentazione, come l’aumento della motivazione e della capacità di apprendere, lo stato di veglia, la crescita del feto, l’eccitamento sessuale.

• Il rilascio di α-MSH, sia nei mammiferi che negli anfibi, si trova sotto il controllo di uno specifico inibitore di origine ipotalamica, MIF o MSH-release inhibiting factor. Pare esista anche un fattore di rilascio di origine anch’esso ipotalamico, detto MRF. L’imbrunimento delle palpebre che può rapidamente far seguito a uno stress emotivo potrebbe essere dovuto a una brusca liberazione di MSH e di ACTH, in conseguenza della mobilitazione dell’asse ipofiso-surrenale, che fa parte delle primissime fasi della sindrome d’adattamento.

• la melanotropina insieme alla prolattina, all'ormone della crescita e alla tireotropina è uno dei quattro ormoni di cui agli inizi degli anni '70 furono scoperti i prodotti ipotalamici con azione inibente la secrezione. Dunque, era già disponibile un dettaglio piuttosto ricco di conoscenze sui meccanismi con cui il sistema nervoso centrale presiede alle funzioni endocrine.

• Le ultime ricerche evidenziano Beta endorfina e Met-enkefalina implicate nella cascata della risposta di stress con numerosi altri neuro peptidi (GH-RH Somatostatina, Vasopressina, Ossitocina, Prolil-Lysin-Glicinamide (PLG), Sostanza P, Angiotensina II, LH-RH, CCK, Bombesina-Gastrin Releasing Peptide, Neuropeptide (NPY), Atriopeptide, NAF, Diazepam Binding Inhibitor e TRH che, in cronico, partecipano nell’eziopatogenesi di patologie somatiche.


















