
Corso di Immunologia A.A. 2009-10 Il Sistema del...
29
Corso di Immunologia A.A. 2009-10 1 Il Sistema del Complemento
Transcript of Corso di Immunologia A.A. 2009-10 Il Sistema del...

Corso di Immunologia
A.A. 2009-10
1
Il Sistema del Complemento

IL COMPLEMENTO: BackgroundIl plasma dei vertebrati contiene diverse proteine che,
attivandosi sequenzialmente, svolgono una determinata
funzione (sistemi di attivazione). Particolarmente
importanti per lo sviluppo e controllo di una risposta
infiammatoria sono il sistema della coagulazione (in cui
una serie di proteine plasmatiche si attivano con
conseguente formazione del coagulo che blocca il flusso
2
ematico in tessuti danneggiati), il sistema fibrinolitico (che
scioglie il coagulo che non e' più necessario e che
potrebbe danneggiare la circolazione sanguigna per la
formazione di piccoli trombi), il sistema delle bradichinine
(che aumenta la permeabilità vasale agendo sulla
muscolatura liscia dei piccoli vasi), ed il sistema
complementare (per la distruzione di microrganismi in
collaborazione diretta o indiretta con gli anticorpi). .

Proprietà sistemi di attivazione
Tutti questi sistemi hanno in comune tre proprietà:
• sequenzialità di attivazione dei vari componenti,
• inattivazione a causa di inibitori,
• amplificazione da parte di un singolo componente
3
di numerosi componenti sequenziali dello stesso e
di altri sistemi.

Storia (1)
Alla fine del 1800 Nuttal aveva capito che era presente
qualcosa di litico nel siero degli animali o delle
persone immunizzate contro qualche agente
patogeno. Si è però dovuto aspettare un po’ di tempo
per capire che nel siero c’era un’attività termolabile, in
grado di lisare i batteri, quando gli animali erano
sensibilizzati contro quei batteri. Termolabile significa
4
sensibilizzati contro quei batteri. Termolabile significa
che, prendendo il siero, e scaldandolo, ad esempio,
per mezz’ora a 56°C, veniva persa la capacità di
uccidere i batteri. Ciò significa che nel siero è
presente una componente termolabile, unitamente ad
una componente termostabile (ovvero gli anticorpi)

Storia (2)
L’esperimento era più o meno fatto in questo modo:
venivano presi dei batteri, e del siero iperimmune
(ovvero da un animale immunizzato più volte) verso i
batteri, tale siero uccideva i batteri. Tale siero perdeva
la sua attività litica con un riscaldamento a 56°C per
mezz’ora. Un altro siero proveniente da un animale
non immunizzato non uccide ovviamente i batteri, ma
5
non immunizzato non uccide ovviamente i batteri, ma
è capace di "complementare" l’attività litica del siero
scaldato.

Storia (3)
Questo dimostrava come nel siero fossero presenti due
componenti:
- termolabile, aspecifica
- termostabile, specifica
Quella termolabile è data appunto dalle proteine del
complemento, quella termostabile è data invece dagli
anticorpi.
6
anticorpi.

Il complemento
Il termine Complemento deriva dal concetto classico di
"fattore complementare all'azione degli anticorpi". Il
Complemento e' composto da una serie di molecole
che si attivano a cascata. Alcuni dei componenti
complementari (C4, C2 e fattore B) sono codificati
all'interno del MHC (antigeni di classe III). I componenti
7
all'interno del MHC (antigeni di classe III). I componenti
complementari hanno alcune caratteristiche in comune:
• interagiscono a livello delle membrane cellulari;
• posseggono attivita' localizzata nella zona di innesco;
• hanno una emivita nell'ordine dei millisecondi;
• sono regolati da inibitori.

Nomenclatura
I componenti della via classica sono designati con una C seguita
da un numero (p. es. C1, C3). A causa dell'ordine con il
quale sono stati via via identificati, i primi quattro
componenti sono numerati come C1, C4, C2 e C3.
I componenti della via alternativa sono designati con una lettera
(p. es. B, P, D). Alcuni componenti vengono denominati
8
fattori (p. es. fattore B, fattore D).
I frammenti di clivaggio sono designati con una lettera minuscola
dopo il nome del componente da cui derivano (p. es. C3a e
C3b sono frammenti del C3). Il C3b inattivo è designato
come iC3b.
I recettori della membrana cellulare per il Complemento sono
abbreviati come CR1, CR2, CR3 e CR4.
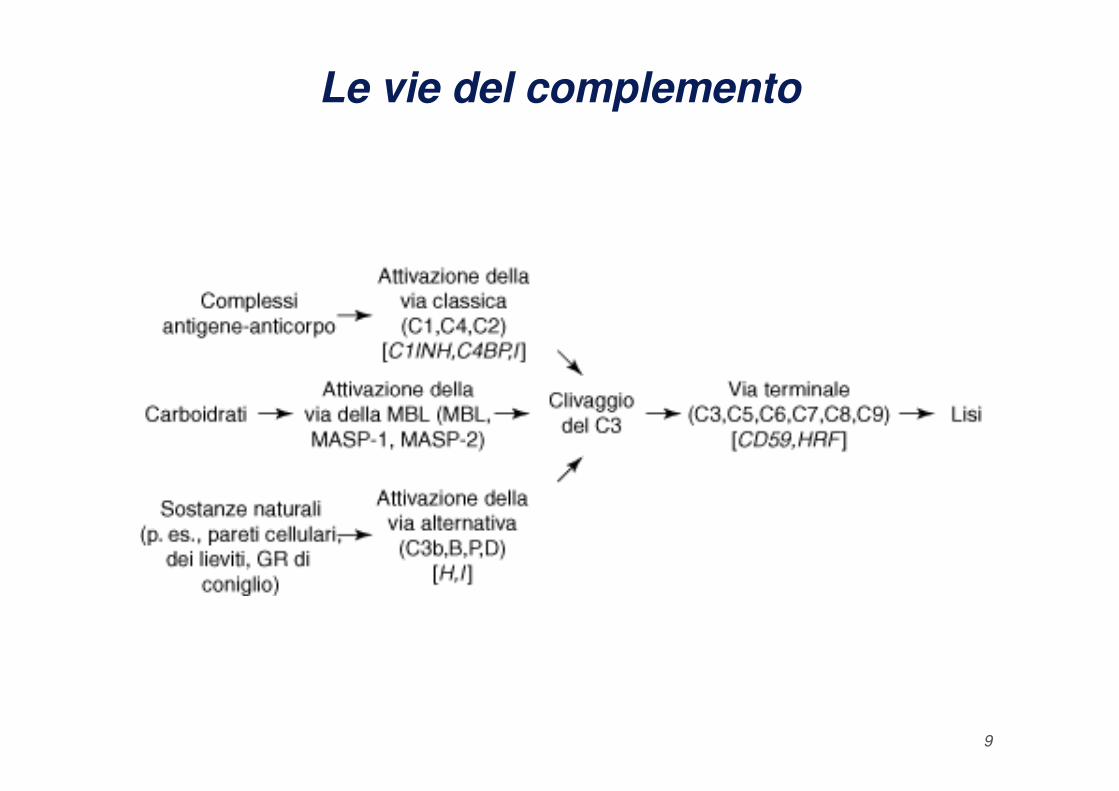
Le vie del complemento
9

La via classica (1)
La Via Classica (specifica) di attivazione del complemento
richiede l'intervento del complesso immune (Ag + 2 IgG
o 1 IgM) o complessi formati da Ig aggregate o
denaturate. Il meccanismo iniziale di attivazione
consiste nell'aggregazione di più molecole Ig che
quindi presentano più punti recettoriali per i filamenti di
10
quindi presentano più punti recettoriali per i filamenti di
C1q con conseguente modificazione conformazionale
e attivazione di tale proteina. L'alto contenuto di
aminoacidi "collageno-simili" (glicina-prolina) di C1q
facilita la formazione di un complesso con le altre due
molecole C1r e C1s.

La via classica (2)
A parte C1q, tutte le proteine complementari svolgono attività
enzimatica: nella forma inattiva (proenzima) il sito
dell'enzima che si deve legare al substrato é nascosto
all'interno della molecola, ma viene esposta in seguito al
taglio della catena peptidica determinato dal fattore
precedente. Il C1s attivato taglia un piccolo frammento di
11
C4 (C4a) mentre il rimanente C4b rimane attaccato alla
membrana (tramite ponte tioestere, C4A:proteine,
C4B:carboidrati). In assenza di Ig o di membrane C4b si
inattiva con l’H2O (mC4b).Contemporaneamente il C1s
agisce su C2 tagliandolo in C2b e C2a che si attacca al
C4b formando il complesso C4b2a (C3 convertasi della
via classica).

La via classica (3)
Questo complesso attiva la molecola C3 (la più importante
e rappresentativa del Complemento, legata da C4b e
clivata da C2a) che quindi forma il complesso
C4b2a3b che rappresenta la C5 convertasi della via
classica (tagliando il C5 in C5a e C5b). In assenza di
12
classica (tagliando il C5 in C5a e C5b). In assenza di
Ig o di membrane C4b si inattiva con l’H2O (mC3b).

La via classica
13

La via alternativa (1)
La Via Alternativa (aspecifica) dipende dall'azione diretta di
molecole microbiche (LPS, polisaccaridi) con azione diretta
sul C3 mediante attivazione sequenziale dei Fattori D, B e
P. Il Fattore B lega il C3b basale e viene scisso dal Fattore
D in Ba e Bb, formando il complesso C3bBb (C3 convertasi
della via alternativa). Il Fattore P (properdina) ha consentito
14
della via alternativa). Il Fattore P (properdina) ha consentito
di scoprire la via alternativa ma non svolge un ruolo
importante, essendo la sua funzione di stabilizzare il
complesso C3bBb sulla membrana.

La via alternativa (2)
In particolare, C3 può attivarsi spontaneamente a bassi livelli
(per idrolisi del tioestere con H2O) formando C3i. Questo
lega il fattore B formando una C3 convertasi iniziale (in fase
liquida) della via alternativa (C3iBb) stabilizato dal fattore P.
Il complesso C3iBb cliva il C3 formando mC3b, il quale in
soluzione si inattiva con H2O ma in presenza di membrane
15
soluzione si inattiva con H2O ma in presenza di membrane
si lega alle proteine di membrana e lega a sua volta il fattore
B dando origine al complesso C3bBb (C3 convertasi della
via alternativa).
In seguito, il complesso C3bBb cliva il C3 e lega un nuovo
frammento di C3b, dando origine al complesso C3bBb3b
(C5 convertasi della via alternativa)

La via delle lectine
La via della lectina legante il mannano (MBL) dipende
per la sua attivazione dal riconoscimento innato di
sostanze estranee (cioè, carboidrati). Questa via
presenta analogie strutturali e funzionali con la via
classica. La MBL è simile al C1q e la MASP-1 e la
16
classica. La MBL è simile al C1q e la MASP-1 e la
MASP-2 sembrano essere simili rispettivamente al
C1r e al C1s della via classica. Pertanto, la MASP-
2 potrebbe clivare il C4 e portare alla formazione di
una C3 convertasi derivata dalla via della MBL.

Le tre vie
1.1.1.1. LaLaLaLa ViaViaViaVia ClassicaClassicaClassicaClassica: Prende origine dal complesso antigene-
anticorpo.
2.2.2.2. LaLaLaLa ViaViaViaVia AlternativaAlternativaAlternativaAlternativa: Innescata da molecole presenti sulla
superficie di agenti patogeni.
3.3.3.3. LaLaLaLa ViaViaViaVia delladelladelladella MBLMBLMBLMBL: Ovvero: MANNAN BINDING LECTINE,
17
3.3.3.3. LaLaLaLa ViaViaViaVia delladelladelladella MBLMBLMBLMBL: Ovvero: MANNAN BINDING LECTINE,
ovvero lectine che legano il mannosio. Tali lectine, che
sono normali costituenti del siero, sono capaci di legare
in maniera specifica il mannosio, presente in alcuni
batteri capsulati.

Convergenza delle tre vie: C5b e MAC
Le vie di attivazione del complemento portano alla formazione di
quello che viene definito “Complesso di Attacco alla
Membrana": MAC.
La fase litica del complemento inizia con la produzione del
fattore C5b da parte delle C5 convertasi e la formazione del
complesso C5bC6C7 che si lega alla membrana e fissa una
18
complesso C5bC6C7 che si lega alla membrana e fissa una
molecola di C8 e sei molecole di C9. Il complesso
C5bC6C7C8C9 forma un cilindro con zone in grado di
legare ed inserirsi nel doppio strato lipidico causando dei
fori di circa 10 nm di diametro che permettono il passaggio
di acqua e sali con conseguente lisi osmotica della cellula o
del batterio.
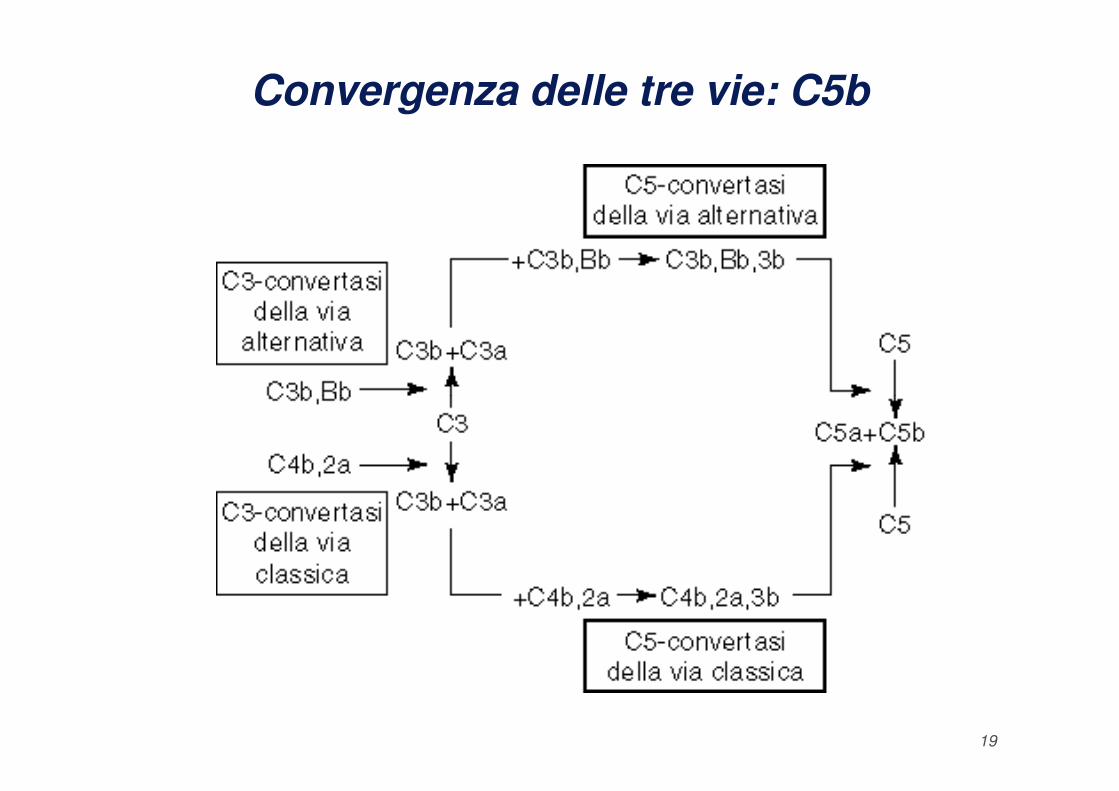
Convergenza delle tre vie: C5b
19
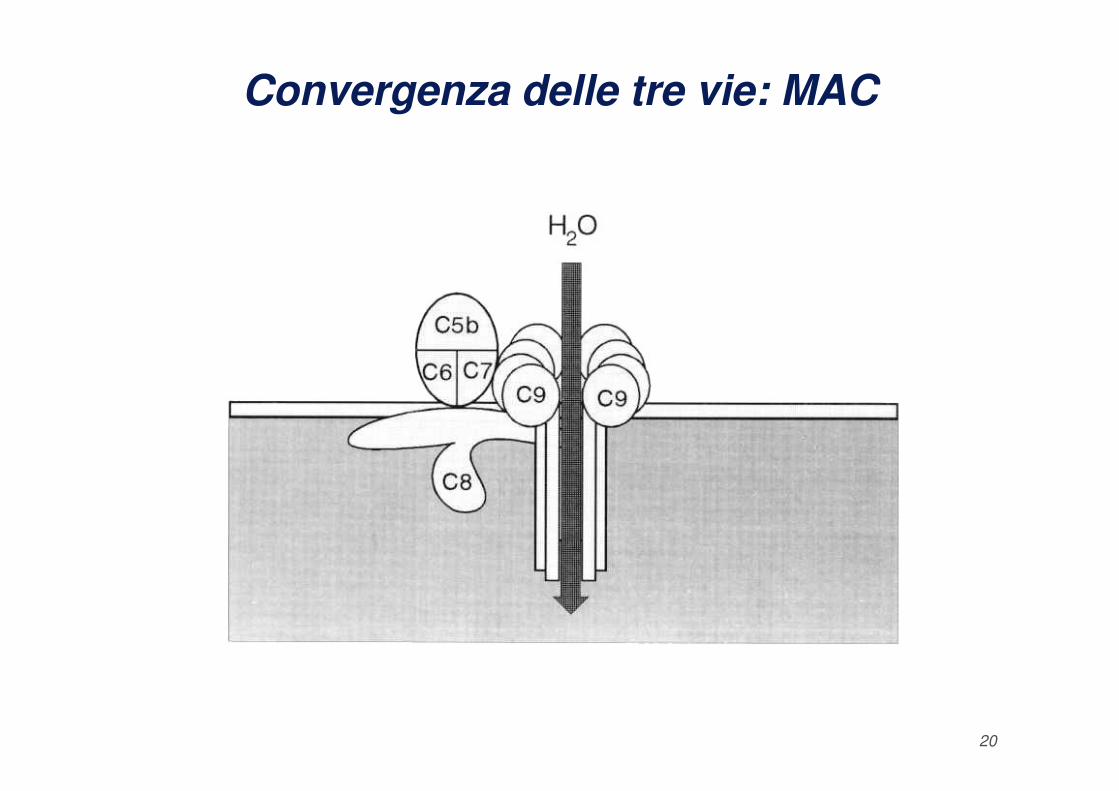
Convergenza delle tre vie: MAC
20

Attività biologiche del complemento (1)
Innanzitutto, la capacità di provocare la lisi delle cellule
bersaglio.
Ovviamente non si tratta solo di cellule batteriche, ma il
complemento interviene soprattutto nei meccanismi di
citotossicità anticorpo-mediata (ADCC): ad esempio,
nella glomerulo-nefrite, nelle cardiopatie post-
21
nella glomerulo-nefrite, nelle cardiopatie post-
streptococciche, o nella miastenia grave. In tutti
questi casi si ha l’attivazione del complemento, e il
problema è dato dal fatto che in questo caso l’anticorpo
reagisce contro componenti proprie dell’organismo, e si
parlerà, come vedremo, di auto-immunità. Nella
emoglobinuria parossistica notturna l’emolisi è
dovuta ad assenza di DAF.

Attività biologiche del complemento (2)
Alcune componenti del complemento, poi, sono in grado di
indurre attivazione di alcune cellule come neutrofili, o
fagociti mononucleati.
Altre componenti del complemento sono in grado di
opsonizzare o gli immuno-complessi o i batteri e
22
indurre la rimozione o la fagocitosi, rispettivamente.
Quindi, possiamo riassumere le tre attività espresse
finora, che sono:
1. Lisi
2. Attivazione
3. Opsonizzazione

Attività biologica dei fattori complementari
Le attivita' biologiche dei vari fattori complementari
possono essere cosi' riassunte:
- aumento della permeabilita' vasale (C2b, C4b)
- opsonizzazione (C3b, C4b)
- neutralizzazione virale (C4b)
- chemiotassi (C5a, C5b,6,7)
23
- chemiotassi (C5a, C5b,6,7)
- attivita' anafilotossinica (C3a, C4a, C5a)
- liberazione enzimi lisosomiali (C5,6,7)
- attivita' litica (C8 e C9)
Va ricordato che sia linfociti B, i macrofagi ed i neutrofili
hanno recettori per alcuni componenti complementari. I
più conosciuti sono quelli che legano il C3.

Molecole inibitorie del complemento
Esistono alcune molecole inibitorie del Complemento:
• C1r-INH e C1s-INH agiscono al livello del C1 (esche);
• C4bp (C4 binding protein) si lega al C4b;
• C3b-INA (o fattore I) blocca il legame del fattore B al C3b;
24
• il fattore H inibisce la C3 convertasi;
• Il DAF (membrana) accelera il processo degradativo delle
C3/C5 convertasi;
• CD59 (membrana) previene l’inserimento e la
polimerizzazione di C9
• fattore S si lega al complesso C5b-9 bloccando l'attivita'
litica.

Recettori per il complemento (CR)CR1: lega il C3b e C4b, rappresenta un sistema di "clearance" dei
complessi immuni ed é presente con alta densità sui
monociti, neutrofili, eosinofili e linfociti B e con bassa densità
sui linfociti T, macrofagi tissutali ed eritrociti. Il ruolo
fisiologico di tale recettore può essere così suddiviso:
1. come recettore ad attività opsonizzante favorisce
25
l'endocitosi;
2. la presenza sugli eritrociti o piastrine può favorire il trasporto
dei complessi immuni o dei batteri dal circolo alle sedi delle
cellule fisse mononucleari fagocitiche (milza);
3. sui linfociti B, può servire insieme al CR2 per l'attivazione
linfocitaria.

Recettori per il complemento (2)
CR2 conosciuto anche come CD21, é il recettore per
il virus di Epstein Barr, lega il iC3b ed é distribuito
sui linfociti B e sulle cellule dendritiche follicolari.
Per quanto il suo ruolo non sia ben caratterizzato,
sembra essere importante per l'attivazione e
26
sembra essere importante per l'attivazione e
sintesi delle Ig dei linfociti B.

Recettori per il complemento (3)
CR3 (CD11b/CD18) lega il C3b ed é presente sui
neutrofili, eosinofili, monociti, linfociti T citotossici e
cellule NK. Il CR3 interviene nella fagocitosi,
mediando l'adesione delle particelle rivestite di C3b
destinate a essere fagocitate.
CR4 lega il C3b, è distribuito sulle cellule della linea
27
CR4 lega il C3b, è distribuito sulle cellule della linea
mieloide e linfoide con la massima espressione sui
monociti tissutali dove può funzionare da recettore per
le particelle opsonizzate con C3b (fagocitosi).

Recettori per il complemento (4)
C5a-R e C3a/C4a-R presenti sui mastociti, sui
neutrofili e monociti giocano un ruolo importante
nelle risposte infiammatorie e nel danno tissutale ad
esse legato.
28

Controllo genico del complementoC'e' almeno un locus per ogni componente
complementare. I loci complementari sono suddivisi in
almeno quattro gruppi di cui il più interessante è quello
inserito nel MHC come molecole di classe III (fattore B,
C2 e C4).
Le singole catene di molti componenti polipeptidici (C3,
C4, C5) sono codificati dal medesimo locus, trascritti in
29
C4, C5) sono codificati dal medesimo locus, trascritti in
un singolo mRNA e tradotti in una singola catena che
viene successivamente scissa in due o tre catene.
Delezione o inattivazione di un particolare gene
complementare puo' causare la non produzione di un
fattore complementare con conseguente
immunodeficienza.


















