
Corso di aggiornamento sulle Biotecnologie CusMiBio · Corso di aggiornamento sulle Biotecnologie...
44
Corso di aggiornamento sulle Biotecnologie CusMiBio Regolazione dell’espressione genica dal DNA alle proteine Prof. Paolo Plevani, Aula 200, 22/09/2014
Transcript of Corso di aggiornamento sulle Biotecnologie CusMiBio · Corso di aggiornamento sulle Biotecnologie...

Corso di aggiornamento sulle Biotecnologie CusMiBio
Regolazione dell’espressione genica dal DNA alle proteine
Prof. Paolo Plevani, Aula 200, 22/09/2014

LEARNING WEEK
Qualche novità e/o stimolo sul controllo dell’espressione genetica…

Figura 3.1 Schema originale di Francis Crick che descrive per la prima volta il “dogma centrale”.
IL DOGMA CENTRALE DELLA BIOLOGIA

Trascrizione, trasporto etc.
DNA
RNA precursori
RNA maturi
Proteine
Funzionalità delle proteine
Livelli di regolazione
Oltre la sequenza: organizzazione cellulare del DNA, traffico, etc.
Processing, stabilità etc.
Regolazione a livello di traduzione
Regolazione post-‐traduzionale

Figura 4.15A Organizzazione del DNA in fibre cromaEniche. (A) In assenza dell’istone H1 la cromaPna appare, al microscopio eleQronico, come una serie di grani su un filo che cosPtuisce la fibra da 10 nm di diametro, chiamata anche “collana di perle”. (B) In presenza di H1 la fibra da 10 nm si organizza in una struQura superiore, chiamata fibra da 30 nm; (C e D) mostrano, rispeXvamente, una rappresentazione schemaPca della fibra da 10 nm e uno dei possibili modelli per la fibra da 30 nm che la descrive come struQura elicoidale (“solenoide”) con sei nucleosomi per giro.


Trascrizione, trasporto etc.
DNA
RNA precursori
RNA maturi
Proteine
Funzionalità delle proteine
Livelli di regolazione
Oltre la sequenza: organizzazione cellulare del DNA e traffico…..
Processing, stabilità etc.
Regolazione a livello di traduzione
Regolazione post-‐traduzionale
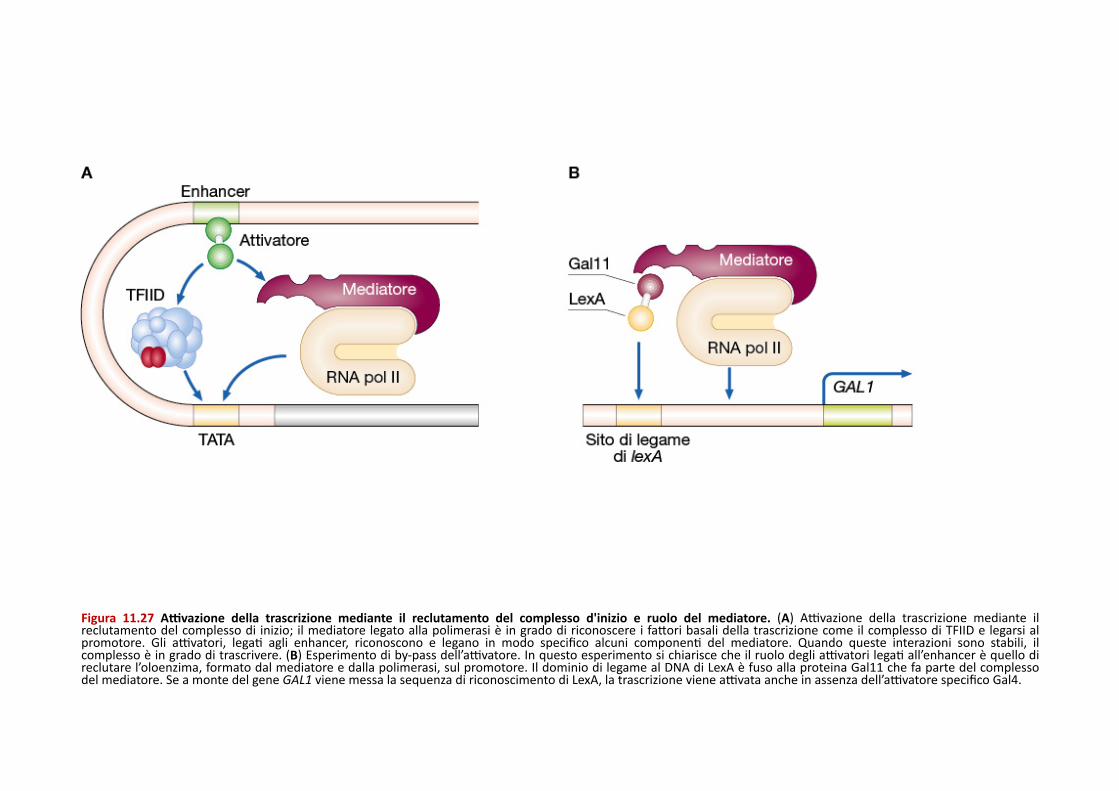
Figura 11.27 AIvazione della trascrizione mediante il reclutamento del complesso d'inizio e ruolo del mediatore. (A) AXvazione della trascrizione mediante il reclutamento del complesso di inizio; il mediatore legato alla polimerasi è in grado di riconoscere i faQori basali della trascrizione come il complesso di TFIID e legarsi al promotore. Gli aXvatori, legaP agli enhancer, riconoscono e legano in modo specifico alcuni componenP del mediatore. Quando queste interazioni sono stabili, il complesso è in grado di trascrivere. (B) Esperimento di by-‐pass dell’aXvatore. In questo esperimento si chiarisce che il ruolo degli aXvatori legaP all’enhancer è quello di reclutare l’oloenzima, formato dal mediatore e dalla polimerasi, sul promotore. Il dominio di legame al DNA di LexA è fuso alla proteina Gal11 che fa parte del complesso del mediatore. Se a monte del gene GAL1 viene messa la sequenza di riconoscimento di LexA, la trascrizione viene aXvata anche in assenza dell’aXvatore specifico Gal4.

Figura 11.28 Ruolo degli aIvatori nella regolazione della struMura della cromaEna. Oltre a interagire con il mediatore e reclutare il complesso di inizio, alcuni aXvatori hanno un ruolo aXvo nella regolazione della cromaPna. (A) Un promotore è inaccessibile a causa della struQura compaQa della cromaPna. Un aXvatore specifico recluta sulla regione una istone acePl transferasi, che rende meno compaQa la cromaPna modificando le code N-‐terminali degli istoni. (B) Il promotore diventa accessibile quando un aXvatore specifico recluta nella regione un rimodellatore della cromaPna che, modificando la struQura della cromaPna (nucleosomi gialli), rende disponibile il promotore per la formazione del complesso di inizio.

Figura 11.29A Visione globale dei meccanismi di aIvazione della trascrizione. (A) I contaX tra gli aXvatori, il mediatore e i TAF portano alla formazione di un complesso stabile che permeQe alP livelli di trascrizione. (B) Il reclutamento delle istone acePl transferasi apre la regione della cromaPna e permeQe la formazione del complesso di inizio. (C) Il reclutamento di una aXvità di istone deacePlasi rende più compaQa la cromaPna e reprime la trascrizione rendendo inaccessibile il promotore.

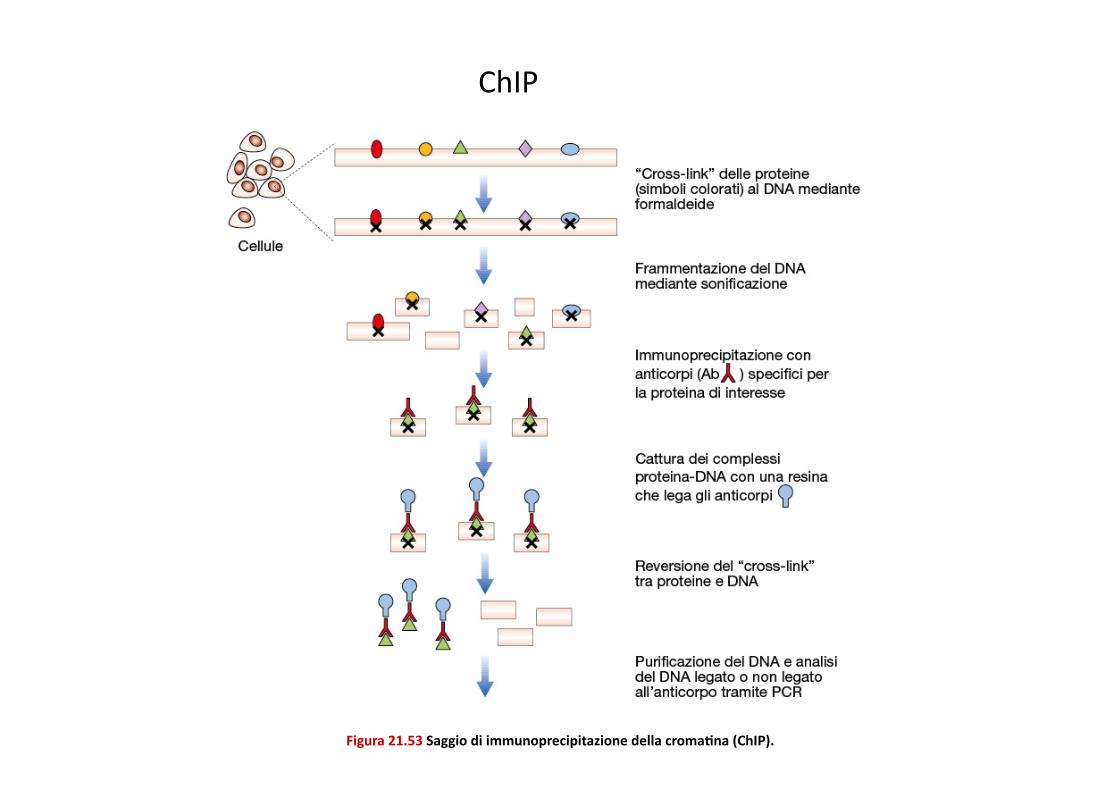
Figura 21.53 Saggio di immunoprecipitazione della cromaEna (ChIP).
ChIP

Mappaggio del legame di numerosi faQori trascrizionali tramite ChIP indica che quesP si legano su migliaia di siP, ma l’occupazione dei siP è quanPtaPvamente differente e solo il legame con alP livelli può essere biologicamente rilevante
Però………


Trascrizione, trasporto etc.
DNA
RNA precursori
RNA maturi
Proteine
Funzionalità delle proteine
Livelli di regolazione
StruQura stessa del DNA, topologia, modificazioni, cromaPna, etc.
Processing, stabilità etc.
Regolazione a livello di traduzione
Regolazione post-‐traduzionale


Figura 14.15 Possibili evenE elementari che concorrono alla generazione di più trascriI a parEre da uno stesso gene. Uno stesso gene può esprimere numerosi trascriX disPnP aQraverso l’uPlizzo di siP alternaPvi di inizio (a) e terminazione (b) della trascrizione (AAUAAA è il Ppico segnale di poliadenilazione), e lo splicing alternaPvo che comprende diversi evenP elementari (c-‐g).

Figura 14.17 Regolazione dello splicing. La regolazione dello splicing comporta l’interazione tra proteine in grado di legare specifici moPvi presenP nel pre-‐mRNA sia negli esoni (ESE, ESS) che negli introni (ISE, ISS). All’azione delle proteine SR che legano le sequenze enhancer negli esoni (ESE) e negli introni (ISE), e che aXvano lo splicing (frecce rosse e verdi a punta), si oppongono le proteine hnRNP, che invece riconoscono le sequenze silencer negli esoni (ESS) e negli introni (ISS) (frecce viola a punta piaQa). Il meccanismo di aXvazione o repressione normalmente comporta l’interazione con altre componenP dello spliceosoma.

Trascrizione, trasporto etc.
DNA
RNA precursori
RNA maturi
Proteine
Funzionalità delle proteine
Livelli di regolazione
StruQura stessa del DNA, topologia, modificazioni, cromaPna, etc.
Processing, stabilità etc.
Regolazione a livello di traduzione
Regolazione post-‐traduzionale
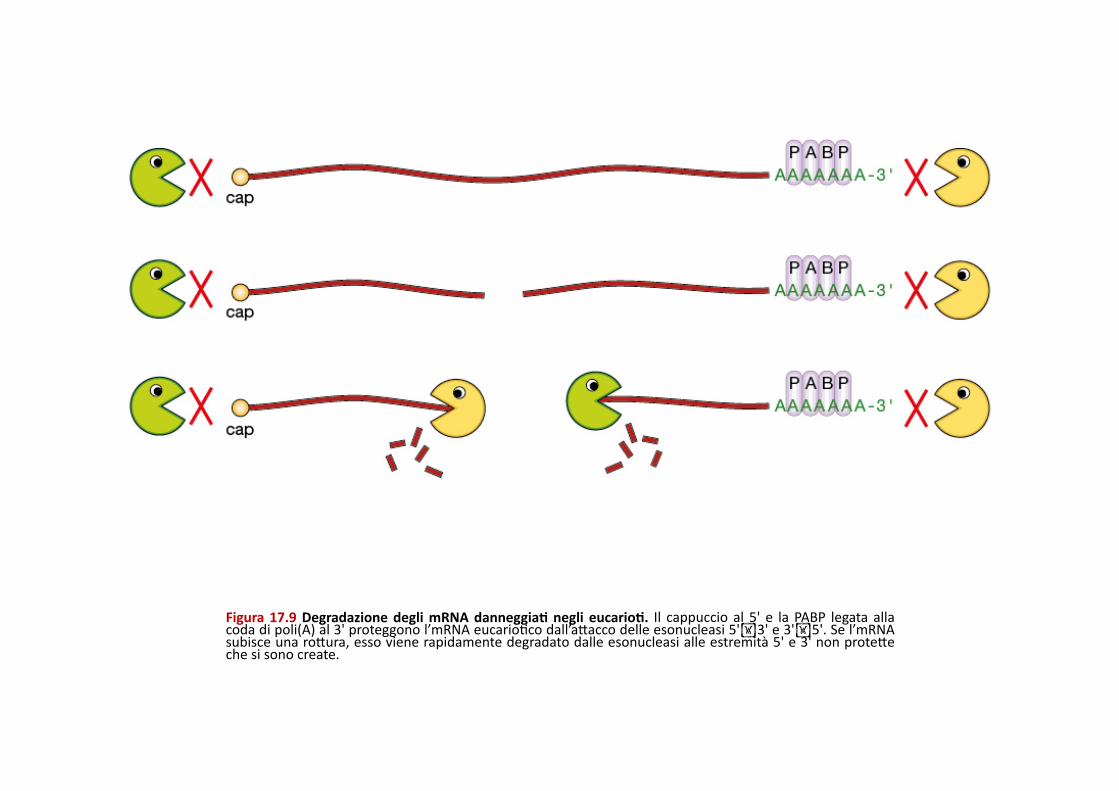
Figura 17.9 Degradazione degli mRNA danneggiaE negli eucarioE. Il cappuccio al 5' e la PABP legata alla coda di poli(A) al 3' proteggono l’mRNA eucarioPco dall’aQacco delle esonucleasi 5'3' e 3'5'. Se l’mRNA subisce una roQura, esso viene rapidamente degradato dalle esonucleasi alle estremità 5' e 3' non proteQe che si sono create.

Figura 17.10 Sequenze ARE e controllo della stabilità degli mRNA. (A) Una sequenza ARE nella 3'UTR di un mRNA lo rende instabile accelerandone la deadenilazione e quindi la degradazione. (B) La proteina AUF1 lega la sequenza ARE che si trova nella 3'UTR dell’mRNA c-‐myc, accelerandone la degradazione. (C) La proteina Hu-‐R lega l’elemento ARE che si trova nella 3'UTR di vari mRNA bersaglio, stabilizzandoli.
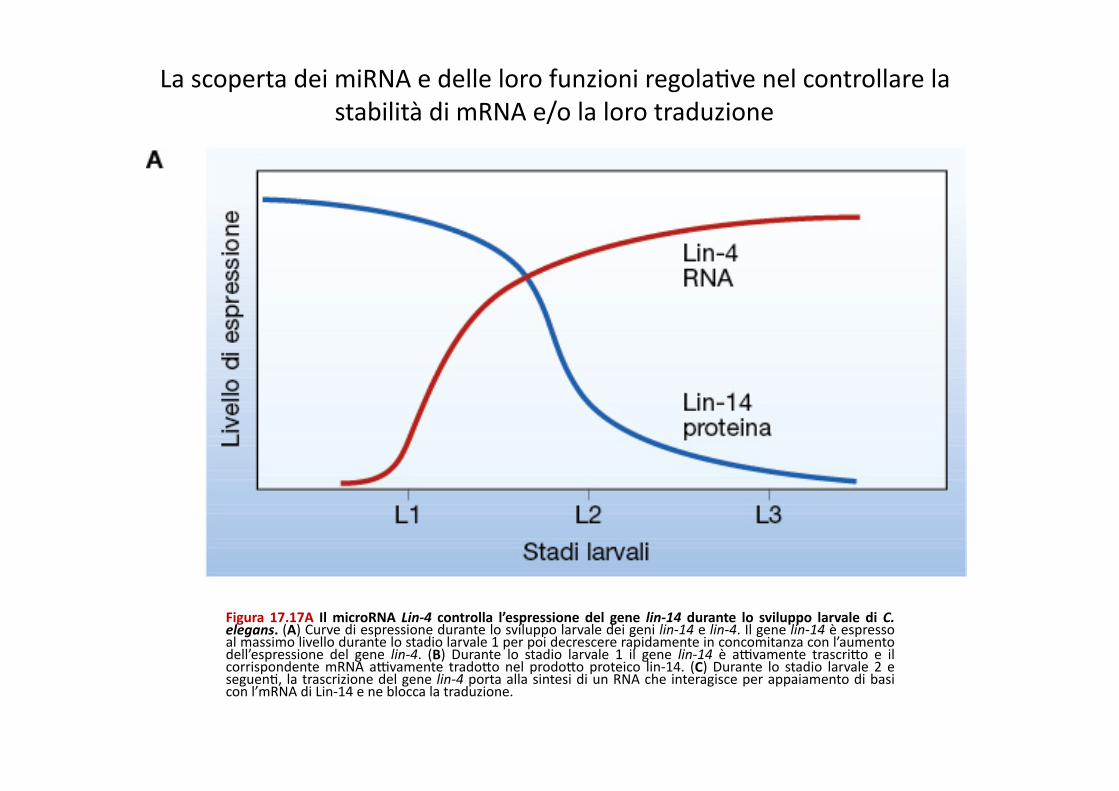
Figura 17.17A Il microRNA Lin-‐4 controlla l’espressione del gene lin-‐14 durante lo sviluppo larvale di C. elegans. (A) Curve di espressione durante lo sviluppo larvale dei geni lin-‐14 e lin-‐4. Il gene lin-‐14 è espresso al massimo livello durante lo stadio larvale 1 per poi decrescere rapidamente in concomitanza con l’aumento dell’espressione del gene lin-‐4. (B) Durante lo stadio larvale 1 il gene lin-‐14 è aXvamente trascriQo e il corrispondente mRNA aXvamente tradoQo nel prodoQo proteico lin-‐14. (C) Durante lo stadio larvale 2 e seguenP, la trascrizione del gene lin-‐4 porta alla sintesi di un RNA che interagisce per appaiamento di basi con l’mRNA di Lin-‐14 e ne blocca la traduzione.
La scoperta dei miRNA e delle loro funzioni regolaPve nel controllare la stabilità di mRNA e/o la loro traduzione

Stadio L1
Stadio L2

Figura 12.8 Biogenesi dei microRNA. I miRNA vengono trascriX dalla RNA polimerasi II soQo forma di trascriX primari di lunghezza variabile (pri-‐miRNA), che vengono riconosciuP e maturaP dal complesso enzimaPco DROSHA in molecole di precursore che presentano una struQura a forcina denominate pre-‐miRNA. I pre-‐miRNA vengono quindi esportaP nel citoplasma per mezzo dell’esporPna 5, e qui interviene un altro complesso proteico, denominato DICER, che produce un duplex di RNA di lunghezza compresa tra 19 e 24 nt. Solo uno dei due filamenP di RNA cosPtui-‐sce il miRNA maturo (l’altro filamento è denominato miRNA*) che viene incorporato nel complesso denominato RISC (RNA-‐Induced Silencing Complex). Il complesso RISC-‐miRNA riconosce la sequenza bersaglio, Ppicamente nella regione 3'UTR di un mRNA, e può indurre la repressione della traduzione o la degradazione del-‐l’mRNA, in funzione del grado di complementarietà con la sequenza bersaglio (vedi par. 17.4).

Figura 17.18 Rappresentazione schemaEca del ruolo dei micro-‐RNA (miRNA). Il miRNA porta il complesso RISC a legarsi al-‐l’mRNA bersaglio mediante appaiamento di basi, con due possibili esiP. (A) Un appaiamento di basi non perfeQo tra il miRNA e la sequenza bersaglio nella 3'UTR di un mRNA risulta in una repressione (reversibile) della traduzione dell’mRNA stesso. (B) Un appaiamento perfeQo di basi tra il miRNA e una regione qualsiasi dell’mRNA bersaglio, perlopiù nella regione codificante, risulta nella degradazione di quest’ulPmo. In genere nella regolazione di un mRNA sono implicaP più miRNA ma, per semplicità, nella figura ne è mostrato uno solo.

Figura 21.50 Inibizione dell’espressione di geni specifici. Descrizione schemaPca delle tecniche di inibizione di geni specifici: (A) mediante RNA anPsenso e (B) mediante l’interferenza da RNA (RNAi).
Silencing

Figura 17F2.1 Funzioni biologiche dei lncRNA. La figura riassume i vari ruoli che possono assumere i long non coding RNA: (A) sequestrare faQori di trascrizione; (B) competere e sequestrare come “spugne” i microRNA; (C) possono essere componenP di complessi RNA-‐Proteine (RNP); (D) reclutare rimodellatori e modificatori della cromaPna, come nel caso di Xist; (E) modulare lo splicing; (F) inibire la traduzione bloccando l’mRNA; e infine (G) portare l’mRNA verso la degradazione.
I long non coding RNA (lncRNA) e i loro ruoli regolaPvi
Xist (17 kb) X inacPvaPon; Tsix si appaia a Xist e inibisce la sua azione di silenziamento
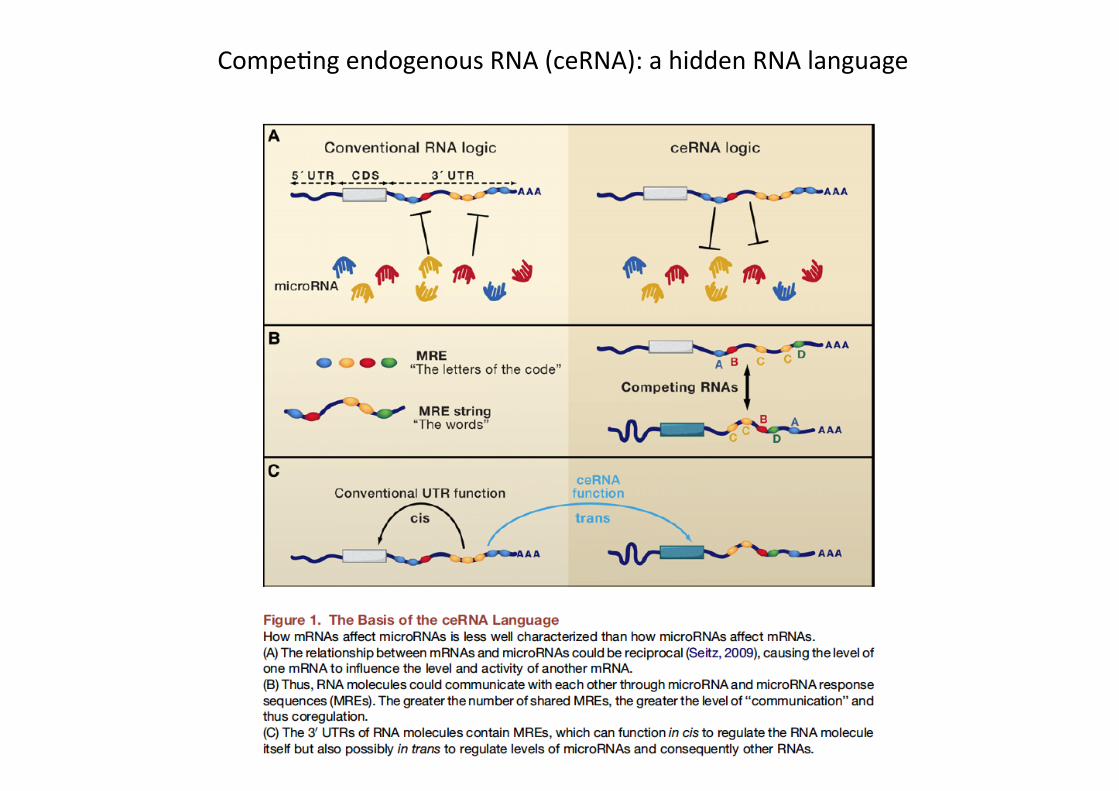
CompePng endogenous RNA (ceRNA): a hidden RNA language

Trascrizione, trasporto etc.
DNA
RNA precursori
RNA maturi
Proteine
Funzionalità delle proteine
Livelli di regolazione
StruQura stessa del DNA, topologia, modificazioni, cromaPna, etc.
Processing, stabilità etc.
Regolazione a livello di traduzione
Regolazione post-‐traduzionale
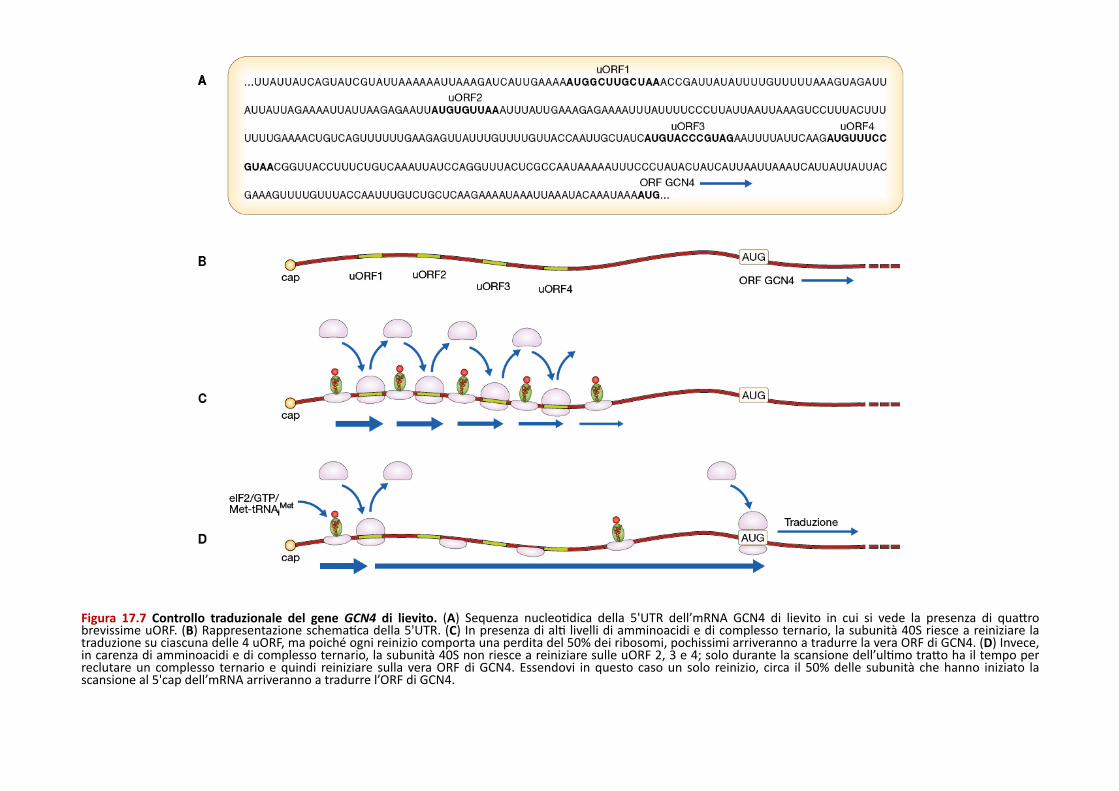
Figura 17.7 Controllo traduzionale del gene GCN4 di lievito. (A) Sequenza nucleoPdica della 5'UTR dell’mRNA GCN4 di lievito in cui si vede la presenza di quaQro brevissime uORF. (B) Rappresentazione schemaPca della 5'UTR. (C) In presenza di alP livelli di amminoacidi e di complesso ternario, la subunità 40S riesce a reiniziare la traduzione su ciascuna delle 4 uORF, ma poiché ogni reinizio comporta una perdita del 50% dei ribosomi, pochissimi arriveranno a tradurre la vera ORF di GCN4. (D) Invece, in carenza di amminoacidi e di complesso ternario, la subunità 40S non riesce a reiniziare sulle uORF 2, 3 e 4; solo durante la scansione dell’ulPmo traQo ha il tempo per reclutare un complesso ternario e quindi reiniziare sulla vera ORF di GCN4. Essendovi in questo caso un solo reinizio, circa il 50% delle subunità che hanno iniziato la scansione al 5'cap dell’mRNA arriveranno a tradurre l’ORF di GCN4.

Figura 17.12 Il sistema di sorveglianza NMD (Non-‐sense Mediated Decay) degli eucarioE. (A) Dopo lo splicing i siP dell’mRNA da cui sono staP rimossi gli introni restano marcaP dalla presenza di complessi proteici EJC. QuesP subiscono poi dei cambiamenP struQurali, tra cui il legame delle proteine Upf, che, se non rimosse, porteranno l’mRNA a degradazione. (B) Al primo ciclo di traduzione il ribosoma che scorre su un mRNA normale rimuove le proteine Upf rendendo così l’mRNA stabile. (C) Se l’mRNA conPene un codone di stop prematuro (PTC), questo causa il rilascio del ribosoma prima che esso abbia raggiunto e rimosso la seguente proteina Upf, che quindi causerà la degradazione dell’mRNA.
Non-‐sense mediated decay lega splicing con traduzione

Trascrizione, trasporto etc.
DNA
RNA precursori
RNA maturi
Proteine
Funzionalità delle proteine
Livelli di regolazione
StruQura stessa del DNA, topologia, modificazioni, cromaPna, etc.
Processing, stabilità etc.
Regolazione a livello di traduzione
Regolazione post-‐traduzionale
TROPPE…. fosforilazione, ubiquiPnazione etc.etc.

Figura 23.6 Esperimento di “distruzione genica” in lievito. (A) Un gene X (in giallo) su uno dei 16 cromosomi di S. cerevisiae può venire inaXvato mediante l’introduzione all’interno del nucleo di cellule di lievito di un frammento di DNA in cui parte del gene X (in giallo) è stata sosPtuita dal marcatore seleXvo (in azzurro), la cui presenza conferisce alle cellule che lo contengono un fenoPpo facilmente individuabile (vedi testo). Questo frammento di DNA può sosPtuire il gene X sul cromosoma determinando l’inaXvazione della sua funzione (distruzione genica) e l’espressione del marcatore seleXvo. (B) L’esperimento descriQo nel pannello (A) viene eseguito su un ceppo diploide di lievito in modo da provocare la distruzione di una soltanto delle due copie del gene X. Dopo sporificazione e dissezione degli aschi per analizzare il genoPpo delle spore, si oQengono quaQro spore vive se il gene X inaXvato non svolge una funzione essenziale per la vitalità cellulare e due di queste (in azzurro) presenteranno il fenoPpo associato alla presenza del marcatore. Se il gene X codifica per una funzione essenziale, solo le due spore di ogni asco che non hanno integrato il marcatore saranno vitali.
Knockout e genome ediPng

Figura 23.23 Produzione di cellule staminali embrionali (ES) da blastocisE di topo e loro uElizzazione. Tre giorni dopo l’incrocio, vengono prelevate dalla femmina gravida le blastocisP che sono colPvate su piastre Petri. Le cellule della massa interna delle blastocisP che crescono su piastra vengono rimosse e dissociate in singole cellule mediante traQamento con enzimi proteoliPci. Queste cellule staminali, chiamate ES, possono differenziarsi in vari tessuP o possono essere mantenute come cellule staminali embrionali indifferenziate a seconda dei terreni di coltura uPlizzaP. DNA esogeno può essere inserito nelle cellule ES tramite eleQroporazione o veQori virali e le cellule contenenP tale DNA esogeno sono riconosciute e selezionate per essere inieQate in una blastocisP che viene poi impiantata nell'utero di una femmina pseudogravida. ES manipolate in vitro derivano, normalmente, da topi con un colore del pelo diverso da quello del topo nella cui blastocisP vengono impiantate le cellule ES manipolate, così che i topi transgenici possono venire riconosciuP come topi chimerici con un mantello di colori diversi.
Topi transgenici

Figura 23.24 UElizzo di vari marcatori per la selezione di topi knock-‐out che derivano da evenE di ricombinazione omologa. Come descriQo nel testo, un transgene contenente il gene neo in un esone del gene che si vuole inaXvare e il gene HSV-‐TK a valle del costruQo, può essere selezionato in quanto resistente all’anPbioPco G418 e al ganciclovir.
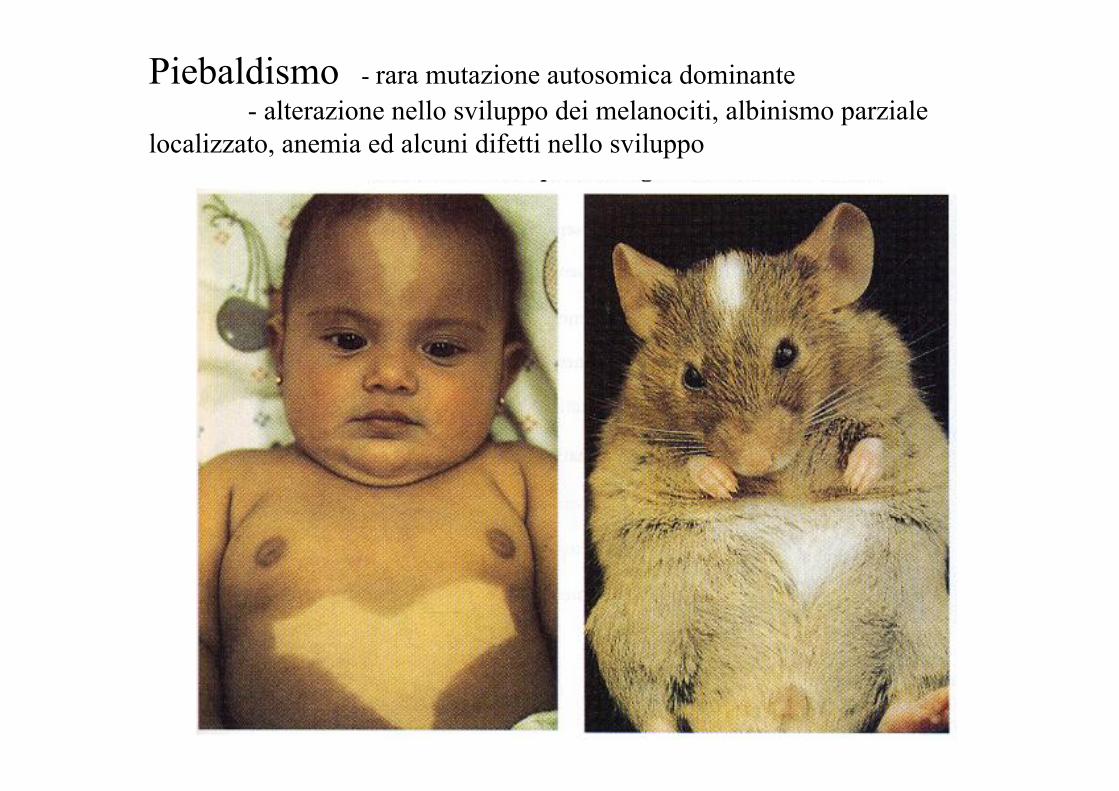
Piebaldismo - rara mutazione autosomica dominante - alterazione nello sviluppo dei melanociti, albinismo parziale
localizzato, anemia ed alcuni difetti nello sviluppo

ZFN: Zinc Ginger Nucleases


The CRISPR-‐Cas system


Si possono fare sempre più cose…che sono sempre migliorabili… …..e non solo dal punto di vista tecnico……



















