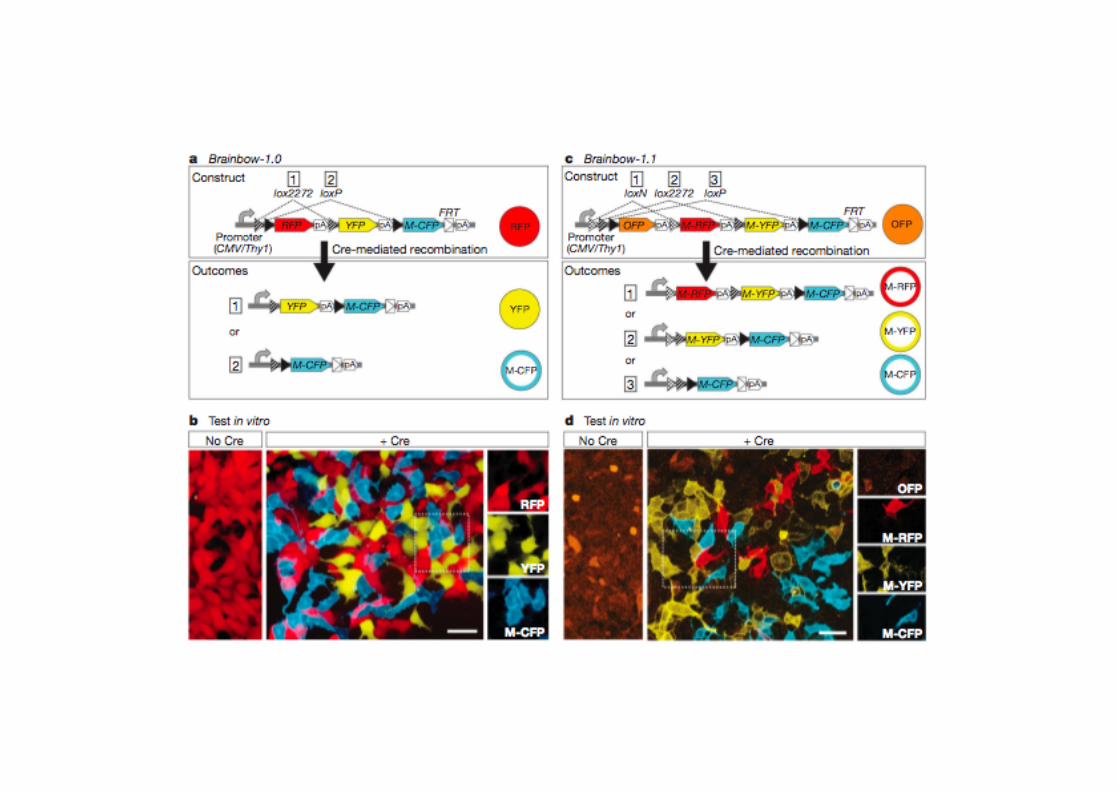
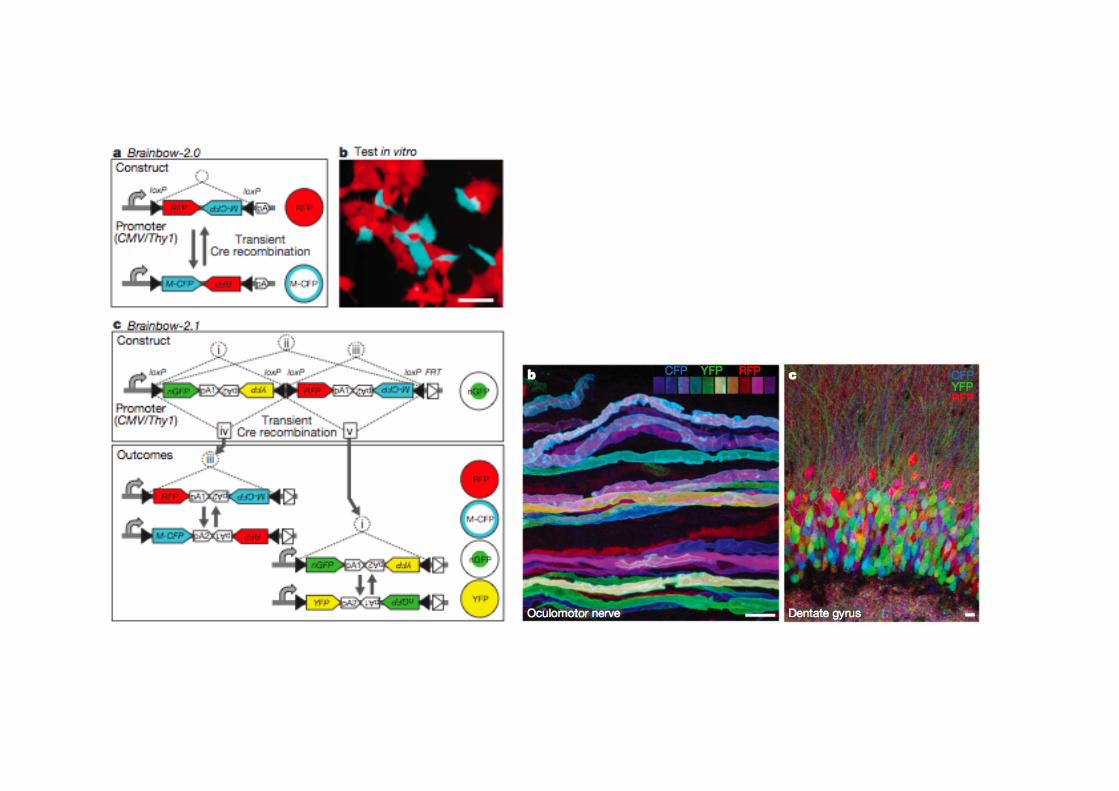
COME%STUDIARE%LA%FUNZIONE% … - Montagner.pdf · Post-genomic era the “PHENOTYPE GAP”...
93
COME STUDIARE LA FUNZIONE GENICA IN EUCARIOTI
Transcript of COME%STUDIARE%LA%FUNZIONE% … - Montagner.pdf · Post-genomic era the “PHENOTYPE GAP”...

COME STUDIARE LA FUNZIONE GENICA IN EUCARIOTI

Post-genomic era
the “PHENOTYPE GAP”
L’identificazione di nuovi geni è molto più rapida della capacità di studiare le loro funzioni.
Necessità di sviluppare approcci di mutagenesi sistematica dell’intero genoma per lo
studio della funzione genica.
FUNCTIONAL GENOMICS

GAIN OF FUNCTION Understand the role of a protein by its
overexpression
LOSS OF FUNCTION: Deduce the relevance of a gene by its absence

GAIN OF FUNCTION Understand the role of a protein by its overexpression
By mutations: - proteins that are active all the time (oncogenes) = proteins that are active in a constitutive way
- new functions (es: new interactions with proteins or DNA) - typically genetically DOMINANT (one allele is sufficient)
By overexpression: - amplification of the gene - mutation of regulatory elements (promoters, enhancers)
Mimicked in research with:
- transgenesis (extra gene copies) - Retrovirus/Lentivirus (stable) - Transfection (transient)
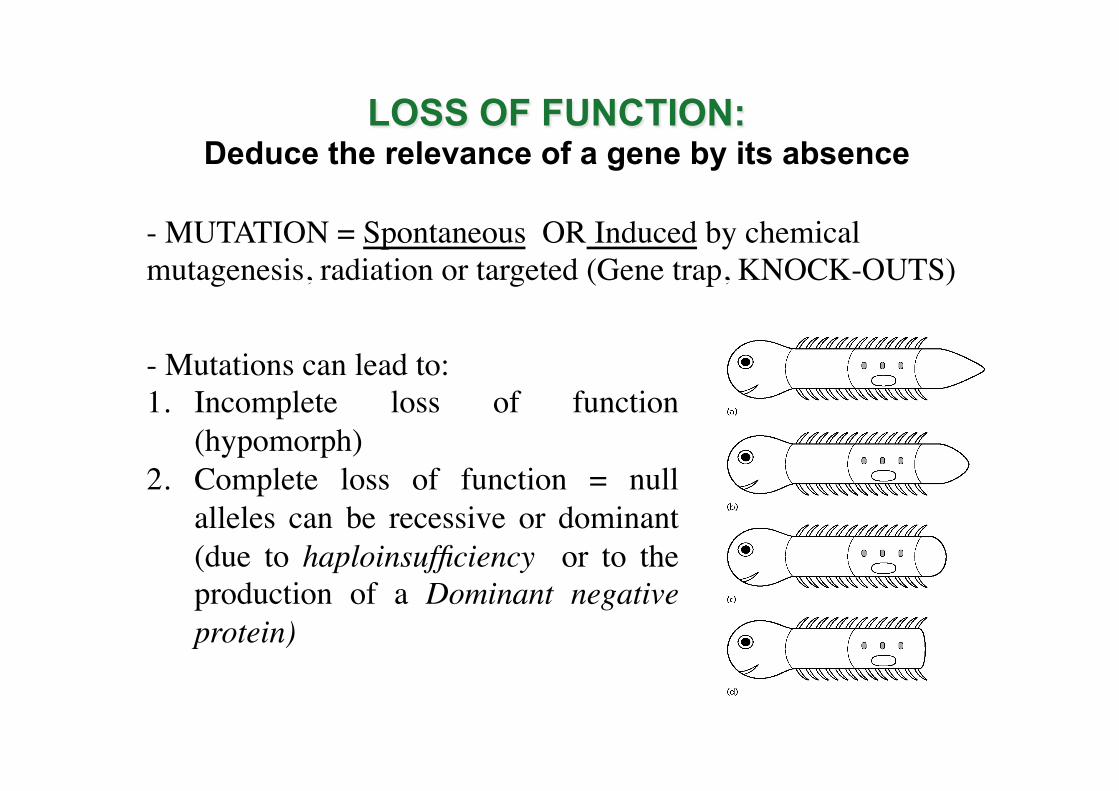
LOSS OF FUNCTION: Deduce the relevance of a gene by its absence
- MUTATION = Spontaneous OR Induced by chemical mutagenesis, radiation or targeted (Gene trap, KNOCK-OUTS)
- Mutations can lead to: 1. Incomplete loss of function
(hypomorph) 2. Complete loss of function = null
alleles can be recessive or dominant (due to haploinsufficiency or to the production of a Dominant negative protein)

LOSS OF FUNCTION
- Transient ways to inhibit gene function:
1. Design DOMINANT NEGATIVE PROTEINS 2. RNAi = dsRNA, (small 21 bp) 3. Neutralizing antibodies
- Stable ways to inhibit gene function: affect the genome
4. Deletion of a gene (knock-out)

La manipolazione sperimentale dell'espressione genica (esperimenC di gain e loss of funcCon) passa quasi invariabilmente per la capacita' di introdurre DNA o RNA in cellule eucarioC DNA Plasmidico (esprimente un gene soJo un promotore forte; esprimente un gene reporter soJo un promotore di un gene eucarioCco oppure soJo un promotore sensore di una certa via di trasduzione; un shRNA per loss of funcCon etc) mRNA microRNA siRNA

Uno sguardo ad un Cpico veJore di espressione per cDNA in eucarioC

L'introduzione di acidi nucleici nella cellula bersaglio aJraverso la membrana plasmaCca avviene aJraverso: 1. TRASFEZIONE (transfec)on) (DNA complessato con Calcio fosfato o liposomi, oppure RNA con liposomi). 2. MICROINIEZIONE 3. VETTORI VIRALI, Tipicamente (90%) retrovirus o lenCvirus ma anche adenovirus
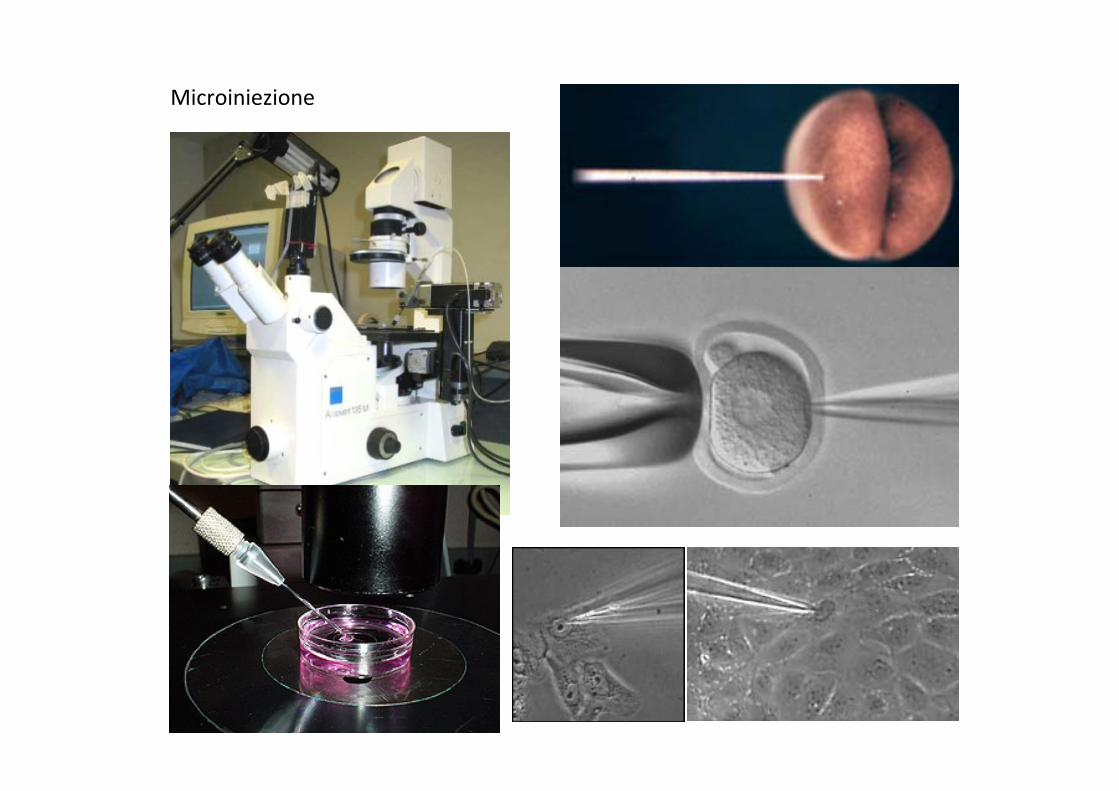
Microiniezione

La Microiniezione Vantaggi: • Consente di introdurre nella cellula qualunque molecola • Consente di introdurre quanCta’ ben definite di materiale nelle cellule • In grandi cellule consente di introdurre degli RNA o delle proteine in modo localizzato • Consente di individuare le cellule che hanno ricevuto l’iniezione, mediante la co-‐iniezione di lineage tracers (ad es. sostanze fluorescenC) • Consente di fare penetrare del materiale geneCco in vivo in modo localizzato (ad e. in singoli miotubi di un muscolo)
Svantaggi: • Richiede una apparecchiatura costosa • Richiede molto tempo per inieJare un numero elevato di cellule • Le cellule inieJate possono morire in conseguenza della perdita del citoplasma dal foro prodoJo dall’ago. Alcune cellule, come i neuroni, sono ipersensibili alla microiniezione. • Cellule troppo piccole non possono essere inieJate senza indurne la morte

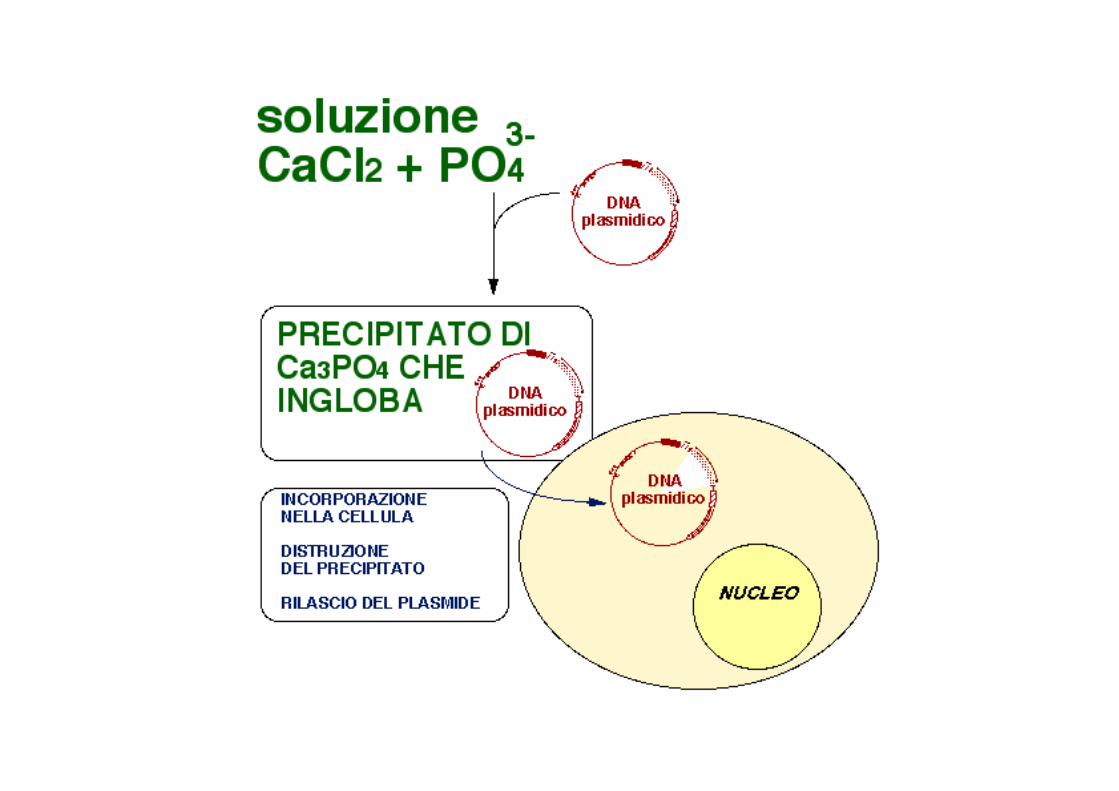
TRASFEZIONE I principali metodi trasfezione consistono nell’”impaccheJare” le macromolecole di interesse (DNA, RNA o proteine) in complessi con delle sostanze “carrier”. Le macromolecole cosi’ complessate vengono somministrate alle cellule, che le inglobano tramite gli endosomi. Qui le macromolecole vengono liberate dal “carrier”. La membrana dell’endosoma viene poi ad essere distruJa dalla cellula, liberando le macromolecole nel citoplasma.
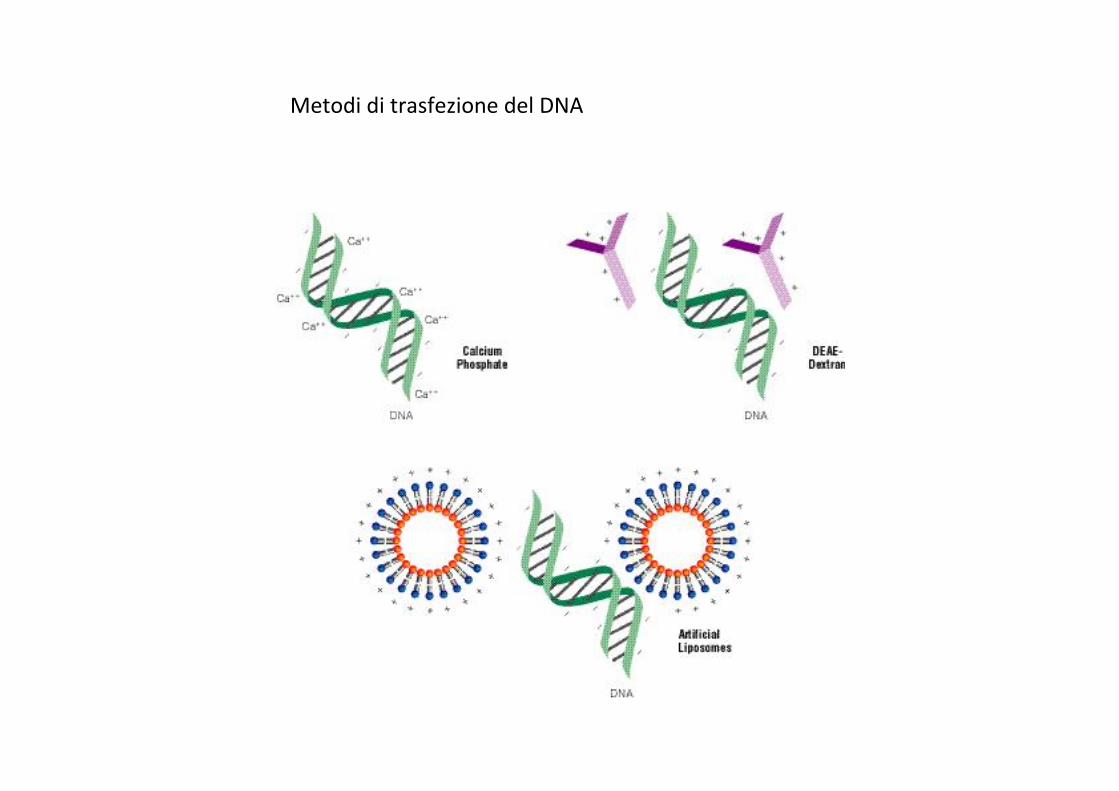
Metodi di trasfezione del DNA
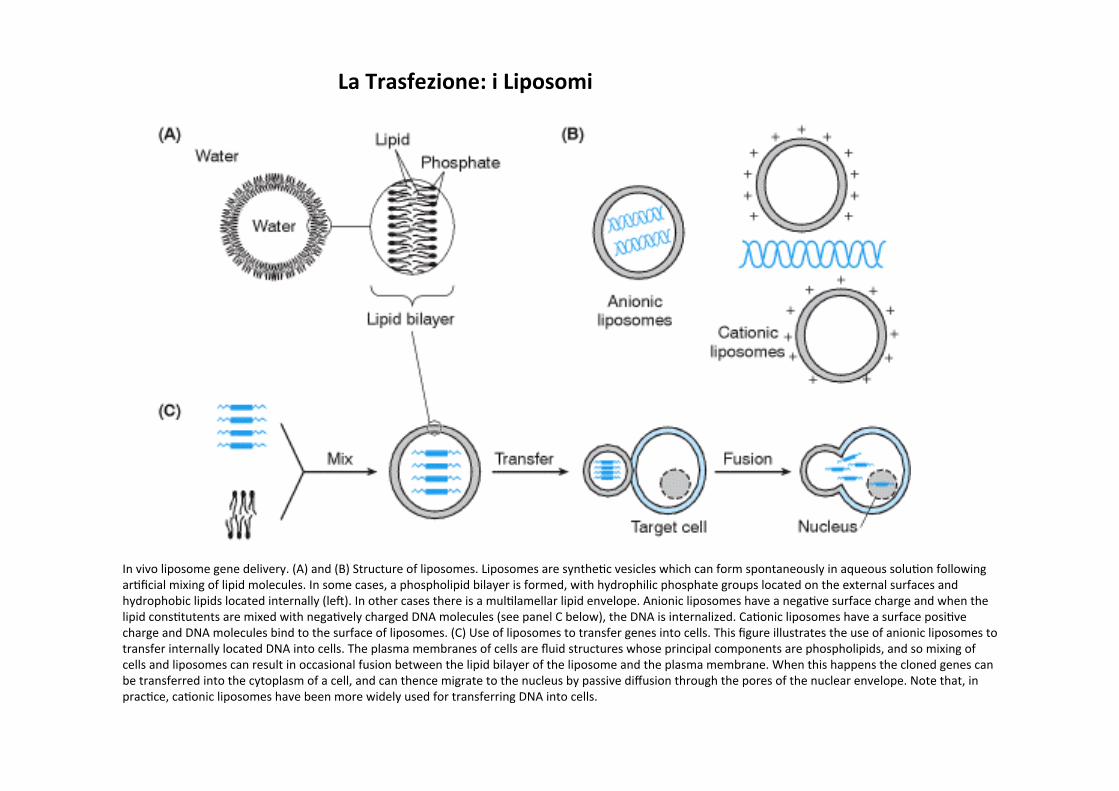
In vivo liposome gene delivery. (A) and (B) Structure of liposomes. Liposomes are syntheCc vesicles which can form spontaneously in aqueous soluCon following arCficial mixing of lipid molecules. In some cases, a phospholipid bilayer is formed, with hydrophilic phosphate groups located on the external surfaces and hydrophobic lipids located internally (lea). In other cases there is a mulClamellar lipid envelope. Anionic liposomes have a negaCve surface charge and when the lipid consCtutents are mixed with negaCvely charged DNA molecules (see panel C below), the DNA is internalized. CaConic liposomes have a surface posiCve charge and DNA molecules bind to the surface of liposomes. (C) Use of liposomes to transfer genes into cells. This figure illustrates the use of anionic liposomes to transfer internally located DNA into cells. The plasma membranes of cells are fluid structures whose principal components are phospholipids, and so mixing of cells and liposomes can result in occasional fusion between the lipid bilayer of the liposome and the plasma membrane. When this happens the cloned genes can be transferred into the cytoplasm of a cell, and can thence migrate to the nucleus by passive diffusion through the pores of the nuclear envelope. Note that, in pracCce, caConic liposomes have been more widely used for transferring DNA into cells.
La Trasfezione: i Liposomi
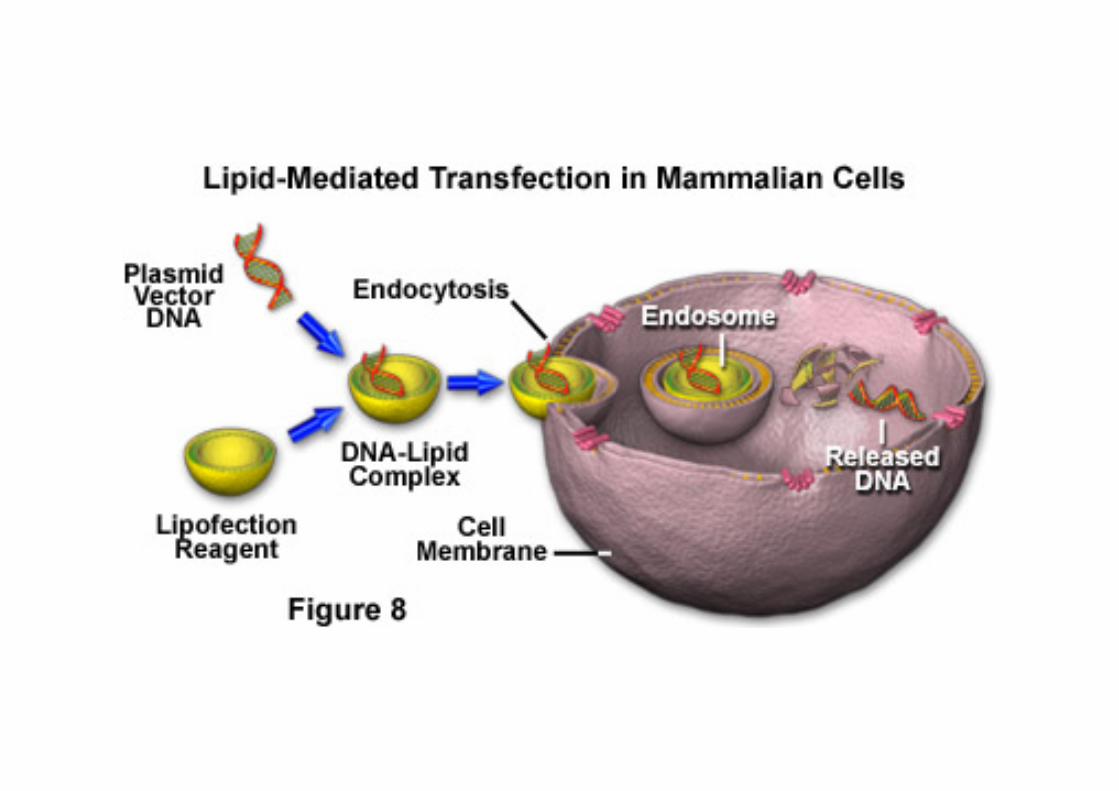

La Trasfezione funziona in molC Cpi cellulari, ma richiede che le cellule advamente incapsulino I materiali da trasfeJare (DNA, RNA o proteine) negli endosomi. Se questo non avviene (es. Le cellule ES) e’ necessario ricorrere a tecniche alternaCve.
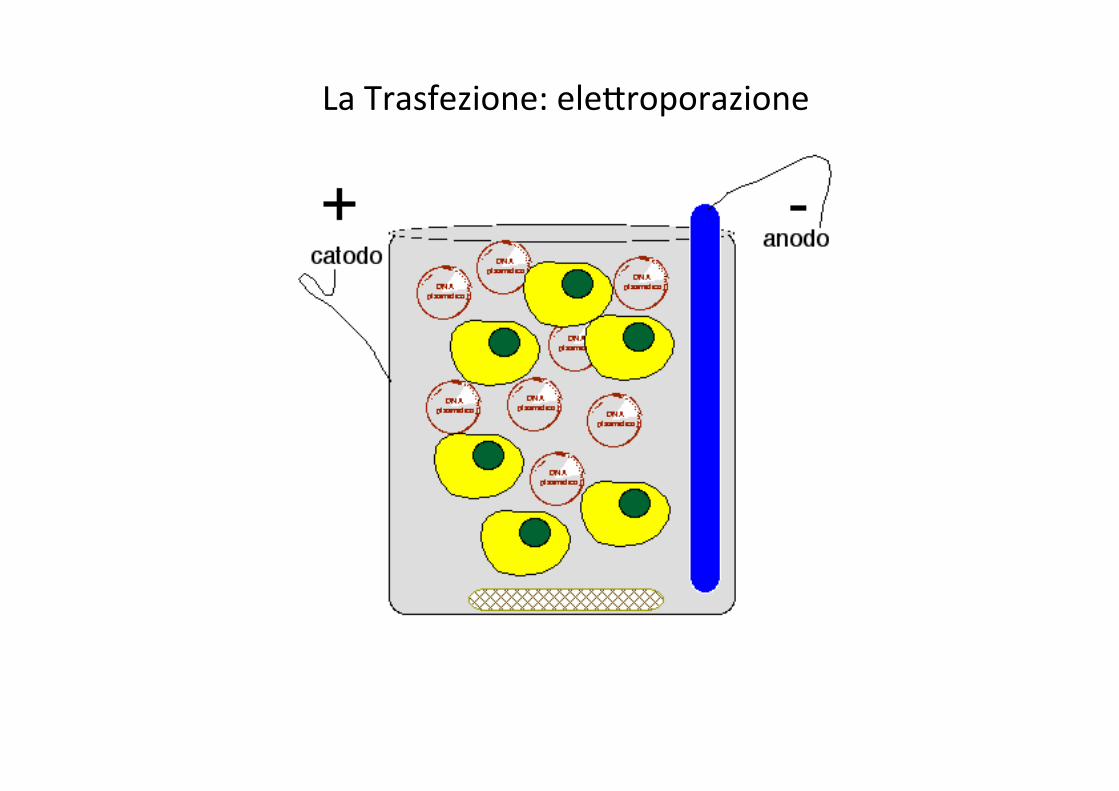
La Trasfezione: eleJroporazione
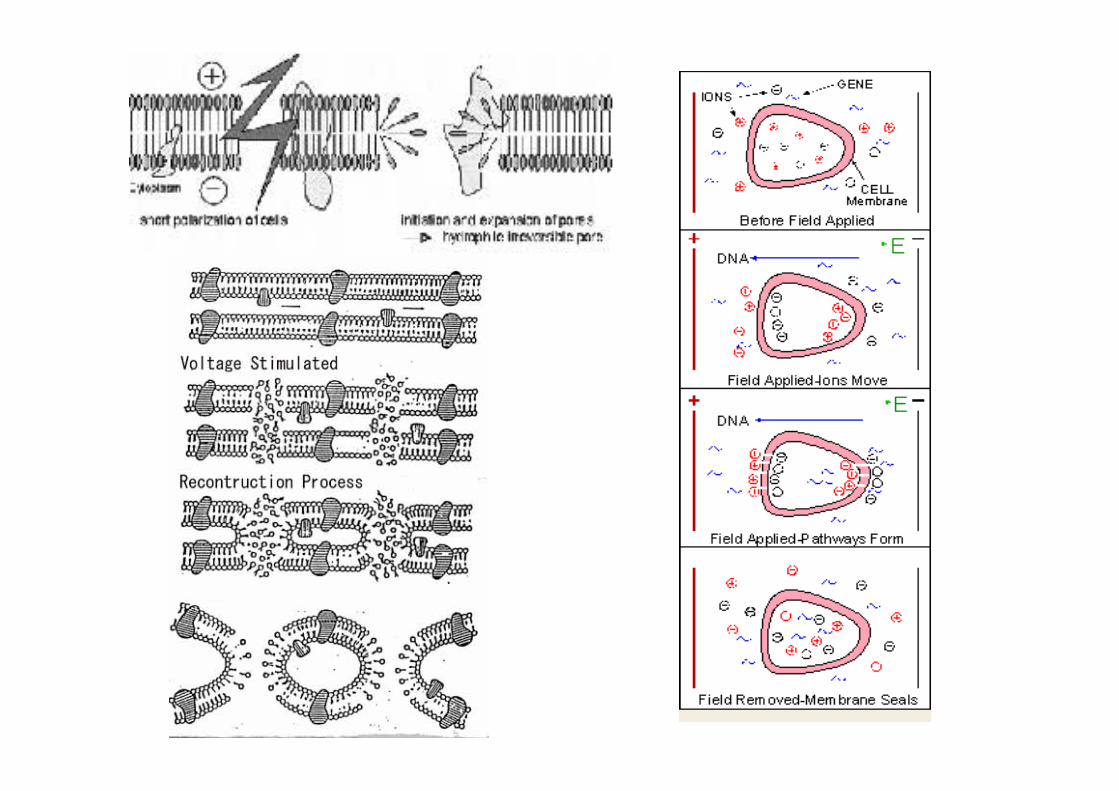

https://www.youtube.com/watch?v=noNJjOthtJ8

Transgenic and “knock-‐out” Animals
= Exogenous DNA sequences are stably integrated in the genome and transmiJed by germ cells -‐ Gain of funcCon or Loss of FuncCon

Applica)ons Study of gene expression and funcMons in the right biological context
Gain informaCons about gene funcCons in pathological contexts
Recombinant protein producCon Experimental model for diseases Genes and cellular therapies

• organismo modello migliore per malattie umane e per lo studio dello sviluppo dei mammiferi
- il topo ha in comune diverse caratteristiche anatomiche e fisiologiche con l’uomo;
- la gran parte dei geni umani ha omologhi funzionali in topo;
- in molti casi il fenotipo di un difetto genetico è molto simile fra uomo e topo;
- le dimensioni, la capacità riproduttiva e l’estensiva storia di studi di genetica classica e molecolare rendono il topo l’organismo modello di scelta per studiare la funzione genica nelle malattie umane.
• unico mammifero adatto per studi di funzione genica in grande scala
• il topo è attualmente l’unico organismo nel quale è possibile indurre, caratterizzare e mantenere mutazioni in cellule ES
VANTAGGI DEL TOPO PER STUDI DI GENOMICA FUNZIONALE

Transgenic produc)on
• Transgenic mice are oaen generated to 1. characterize the ability of a promoter to direct Cssue-‐specific gene expression – e.g. a promoter can be aJached to a reporter gene such as LacZ or GFP
2. examine the effects of overexpressing and misexpressing endogenous or foreign genes at specific Cmes and locaCons in the animals
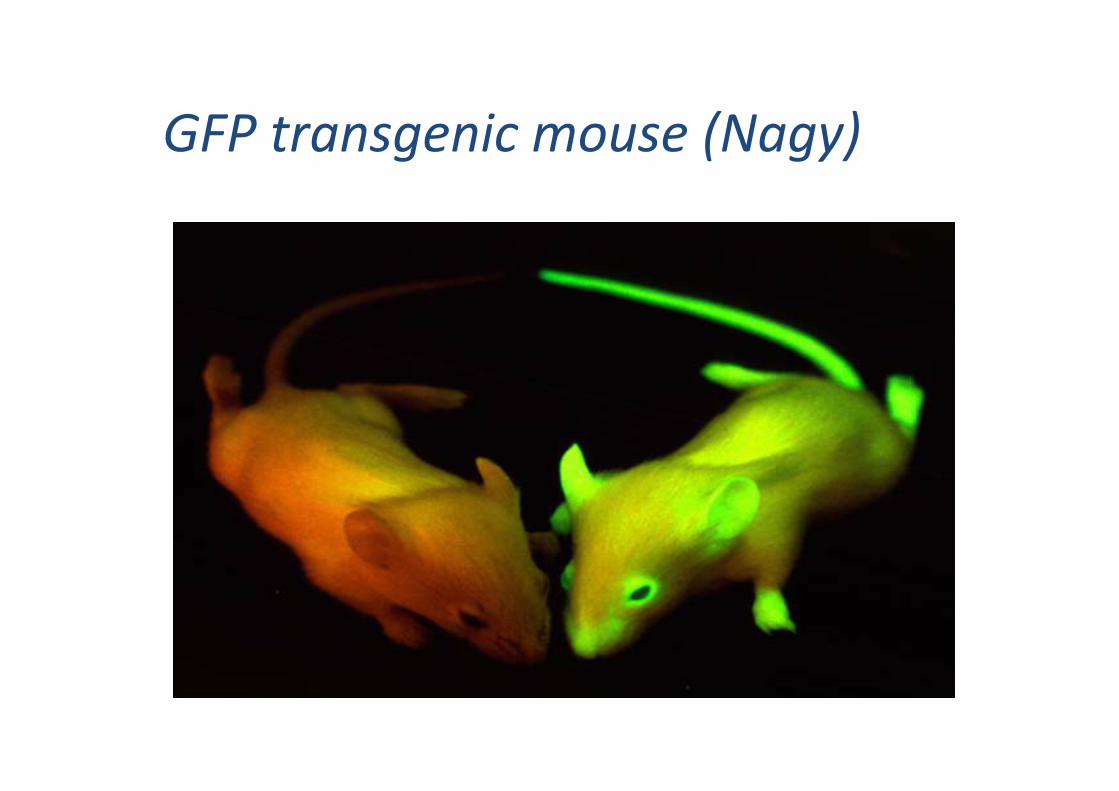
GFP transgenic mouse (Nagy)
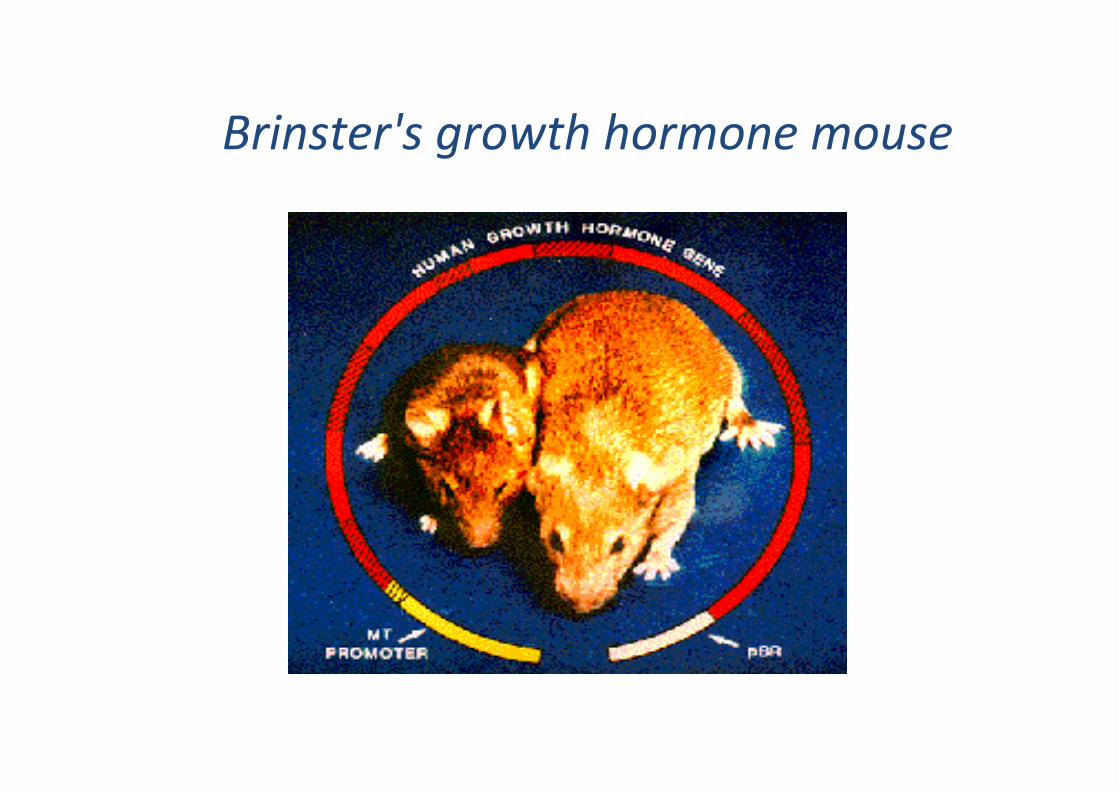
Brinster's growth hormone mouse
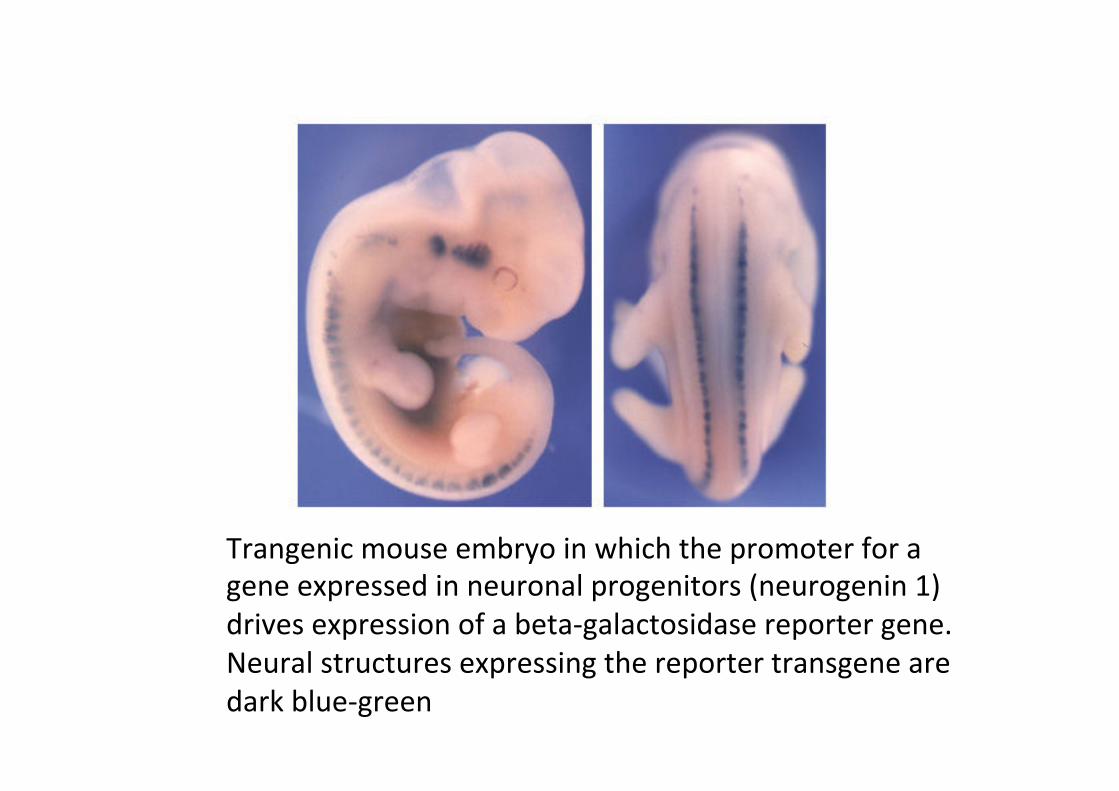
Trangenic mouse embryo in which the promoter for a gene expressed in neuronal progenitors (neurogenin 1) drives expression of a beta-‐galactosidase reporter gene. Neural structures expressing the reporter transgene are dark blue-‐green

Transgenic mice as tools Polio virus receptor Normal mice can't be infected with polio virus. They lack the cell-‐surface molecule that, in humans, serves as the receptor for the virus.
Tg mice expressing the human gene for the receptor can be infected by polio virus and even develop paralysis and other pathological changes characterisCc of the disease in humans
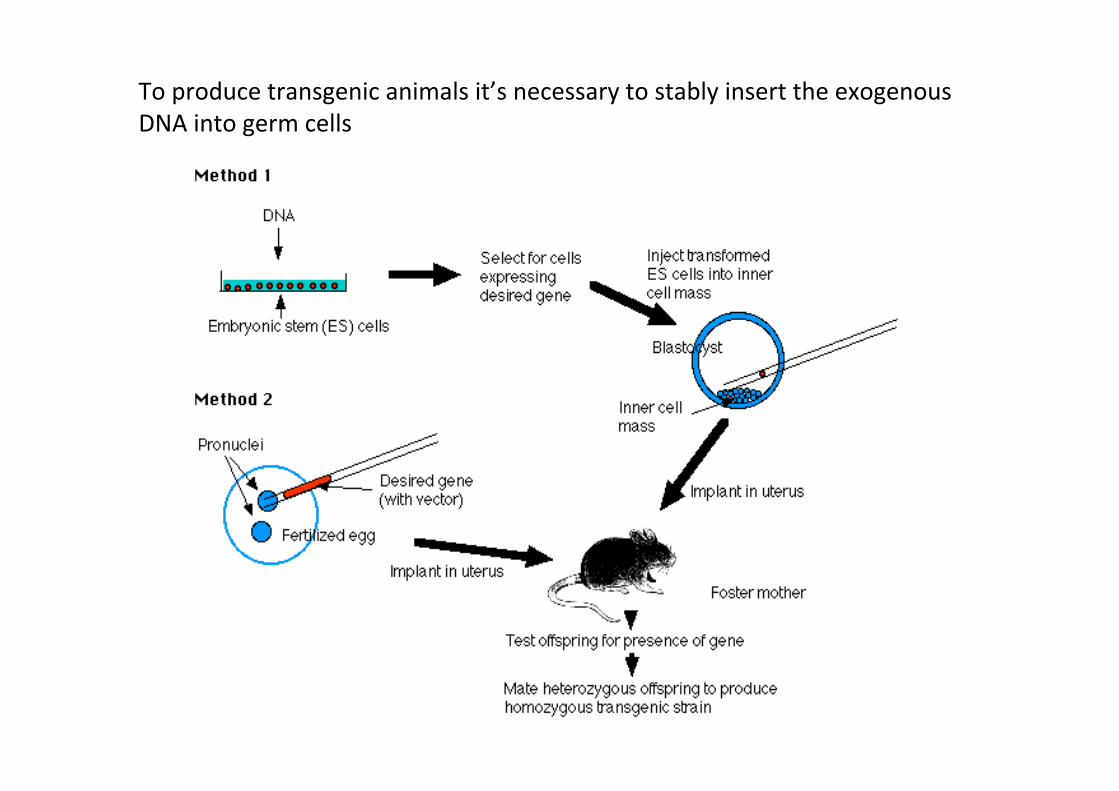
To produce transgenic animals it’s necessary to stably insert the exogenous DNA into germ cells
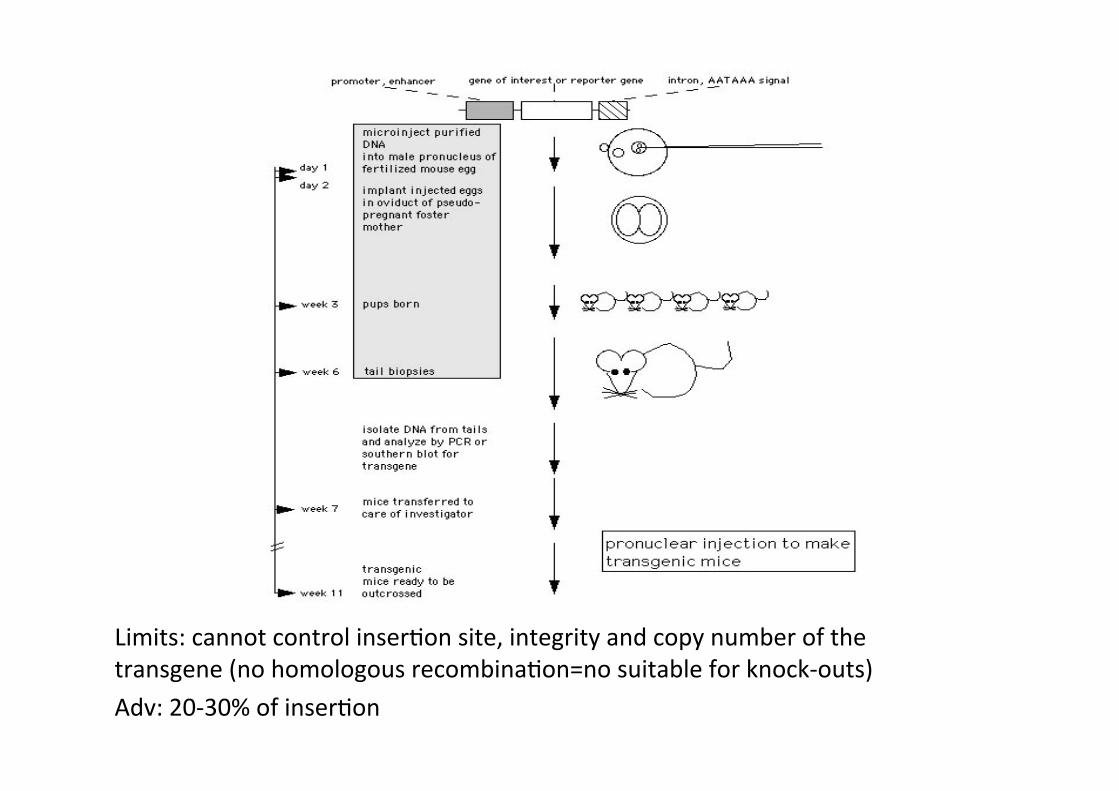
Limits: cannot control inserCon site, integrity and copy number of the transgene (no homologous recombinaCon=no suitable for knock-‐outs) Adv: 20-‐30% of inserCon
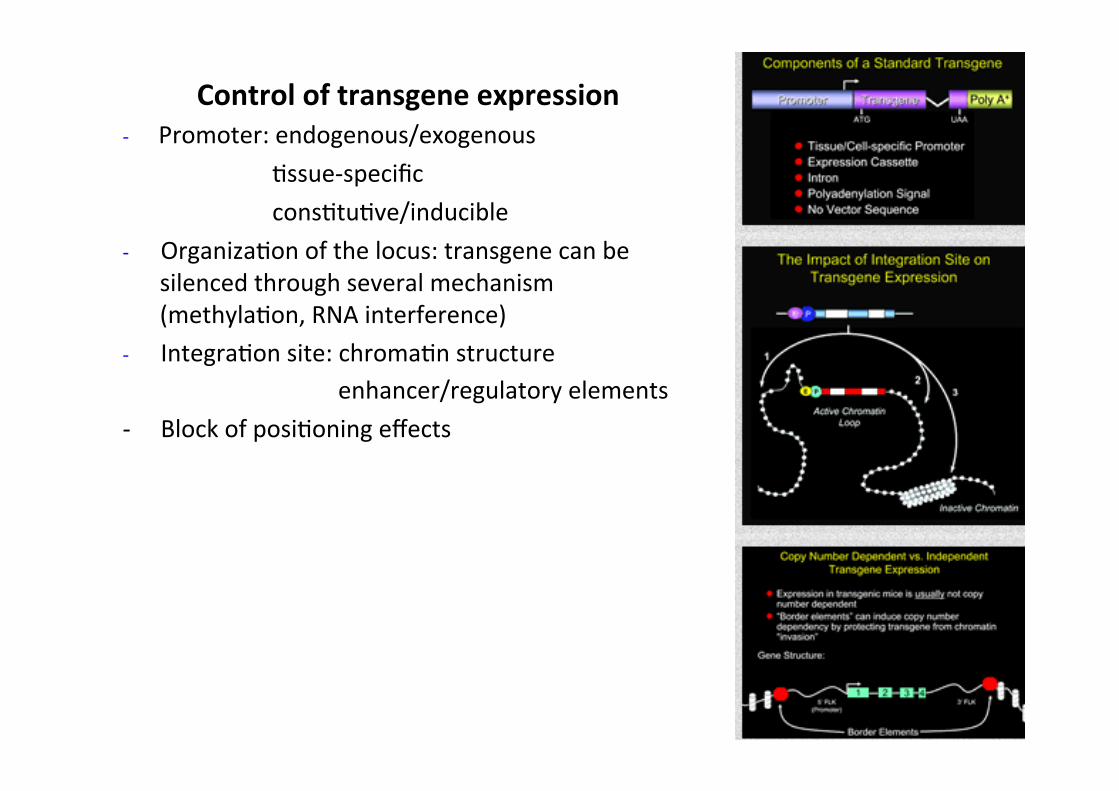
Control of transgene expression -‐ Promoter: endogenous/exogenous
Cssue-‐specific consCtuCve/inducible
-‐ OrganizaCon of the locus: transgene can be silenced through several mechanism (methylaCon, RNA interference)
-‐ IntegraCon site: chromaCn structure enhancer/regulatory elements
-‐ Block of posiConing effects

PREMIO NOBEL per la MEDICINA 2007 MoCvazione: for their work on "principles for introducing specific gene modificaCons in mice by
the use of embryonic stem cells and gene targeCng.”
GENE TARGETING
Mario R. Capecchi
MarCn J. Evans Oliver Smithies

GENE TARGETING
Mutagenesi MIRATA mediante ricombinazione omologa in cellule ES
PermeJe la creazione di una mutazione in un gene PREDETERMINATO
Questa mutagenesi può avere come risultato: 1) INATTIVAZIONE dell’espressione di un gene (KNOCK-‐OUT) 2) DIMINUZIONE dell’espressione di un gene (KNOCK-‐DOWN) 3) INSERZIONE di un gene difedvo/selvaCco 4) (correzione) (KNOCK-‐IN)
IL GENE TARGETING SFRUTTA LA RICOMBINAZIONE OMOLOGA

Caratteristiche ed applicazioni
GENE TARGETING
• Mutagenesi mirata: introduzione di modificazioni sito-specifiche nel genoma mediante ricombinazione omologa
• I topi "knockout" costituiscono un tool estremamente potente per: - l'analisi di diversi aspetti della funzione genica in vivo - dissezionare la funzione di componenti specifici di processi biologici complessi - ottenere modelli murini di malattie ereditarie umane
• È possibile produrre alterazioni geniche specifiche, da mutazioni puntiformi fino a estesi riarrangiamenti cromosomici
• Recenti sviluppi: è possibile generare knockout condizionali o gene targeting inducibili in precisi tessuti o in stadi definiti di sviluppo
• Basato su costrutti contenenti cassette selezionabili, introdotti in cellule ES

GENE TARGETING

Gene TargeMng Technology
-‐ ES Cells take up exogenous DNA -‐ frequency is very low -‐ can be induced to higher frequency
-‐ chemical, electrical, injecCon
-‐ In Dividing Cells: -‐ some DNA incorporated into genome -‐ Random integraCon -‐rare event -‐ Homologous RecombinaCon -‐ even more rare

Transgenic vs. “knock-‐out”
Transgenic: an organism that has had DNA introduced into one or more of its cells arCficially
gene(s) added can be: -‐ a foreign gene -‐ extra copies of endogenous gene -‐ mutated endogenous gene
IntegraCon: -‐ Random (approx.. 10% disrupt an endogenous gene important for normal development), Gene trap -‐ Targeted -‐ Single or mulCple copies
Transgenic

“knockout”: one or more endogenous genes are specifically inacCvated
DNA is introduced first into embryonic stem (ES) cells. ES cells that have undergone homologous recombinaCon are idenCfied and injected into a 4 day old mouse embryo -‐ a blastocyst • targeted inserCon
Transgenic vs. “knock-‐out” Knock-‐out

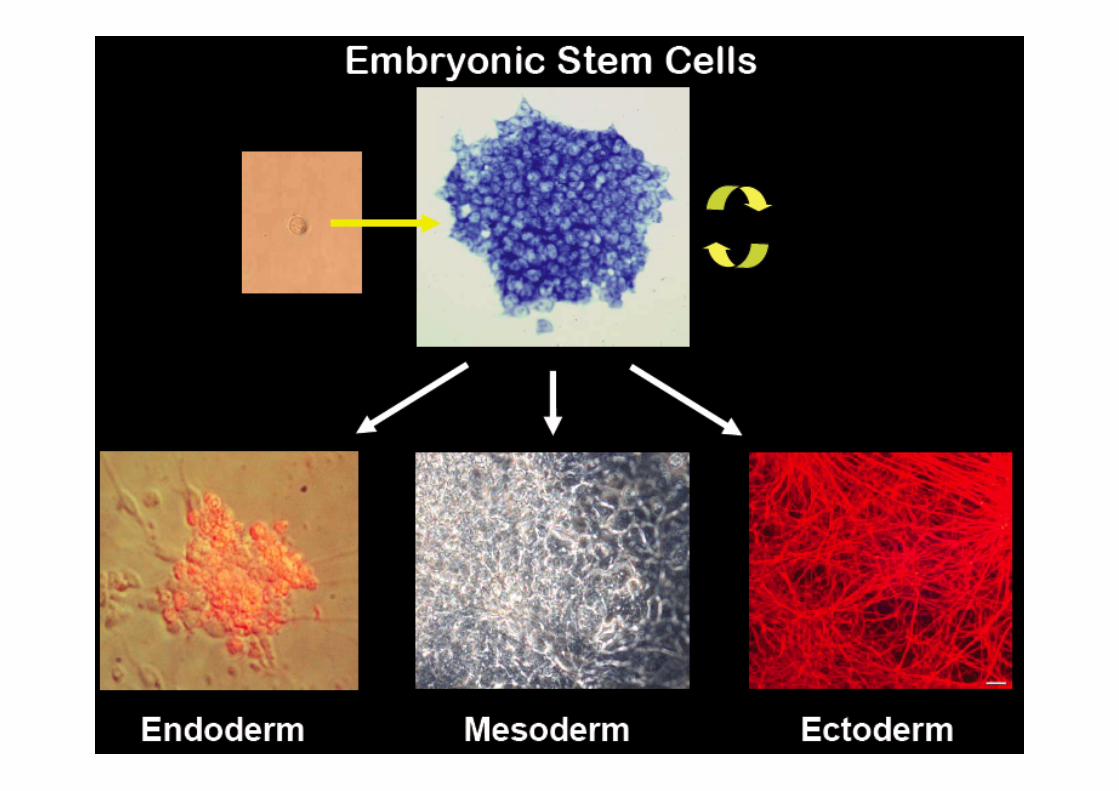
Embryonic Stem (ES) Cells
-‐ isolated from a pre-‐implantaCon embryos -‐ from inner cell mass of blastula stage embryo -‐ cells are undifferenCated -‐ cells are pluripotent -‐ able to differenCate into many (all?) different
cell types in embryo -‐ most importantly germ cells -‐ grown in culture -‐ need cells to divide but not differenCate -‐ longer Cme in culture = more differentaCon -‐ available for mouse, human, rat
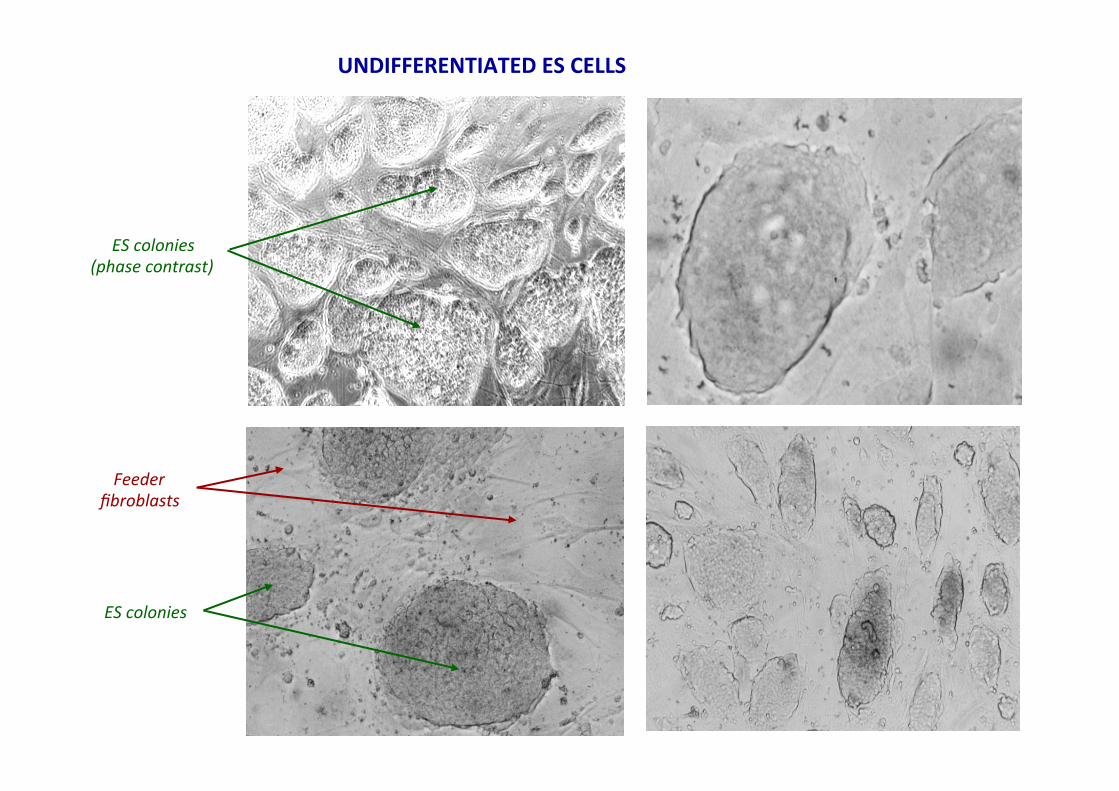
UNDIFFERENTIATED ES CELLS
ES colonies (phase contrast)
ES colonies
Feeder fibroblasts
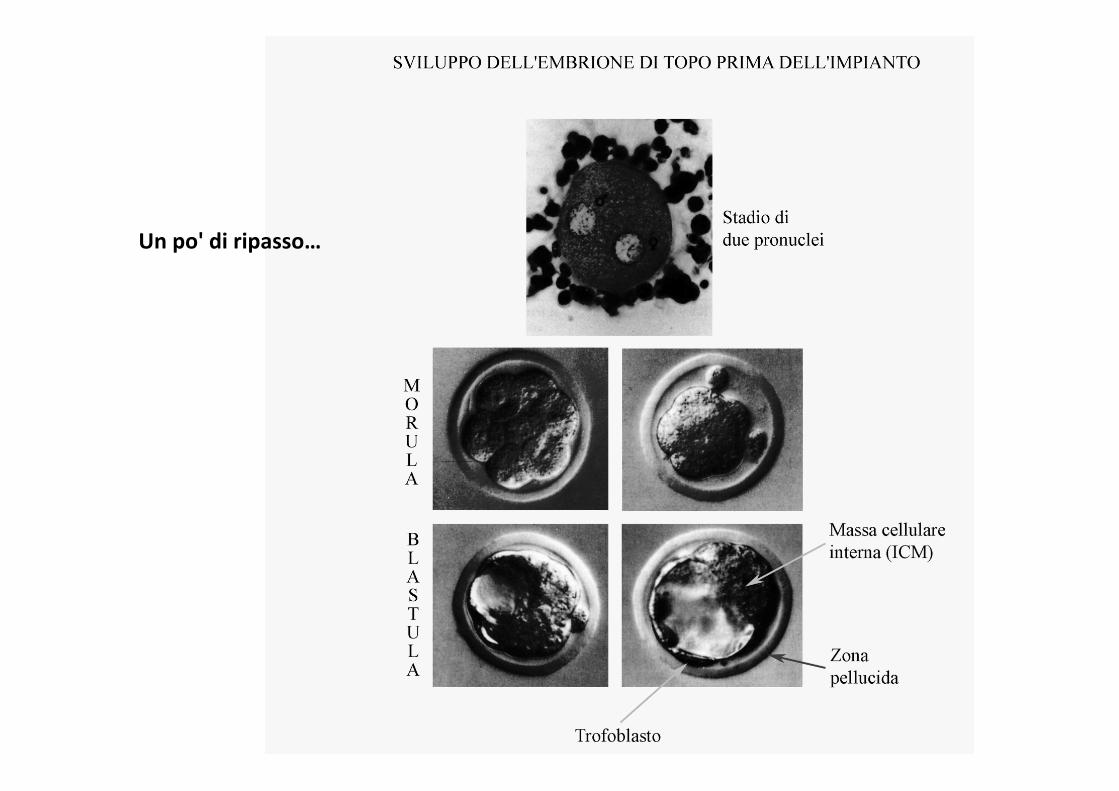
Un po' di ripasso…
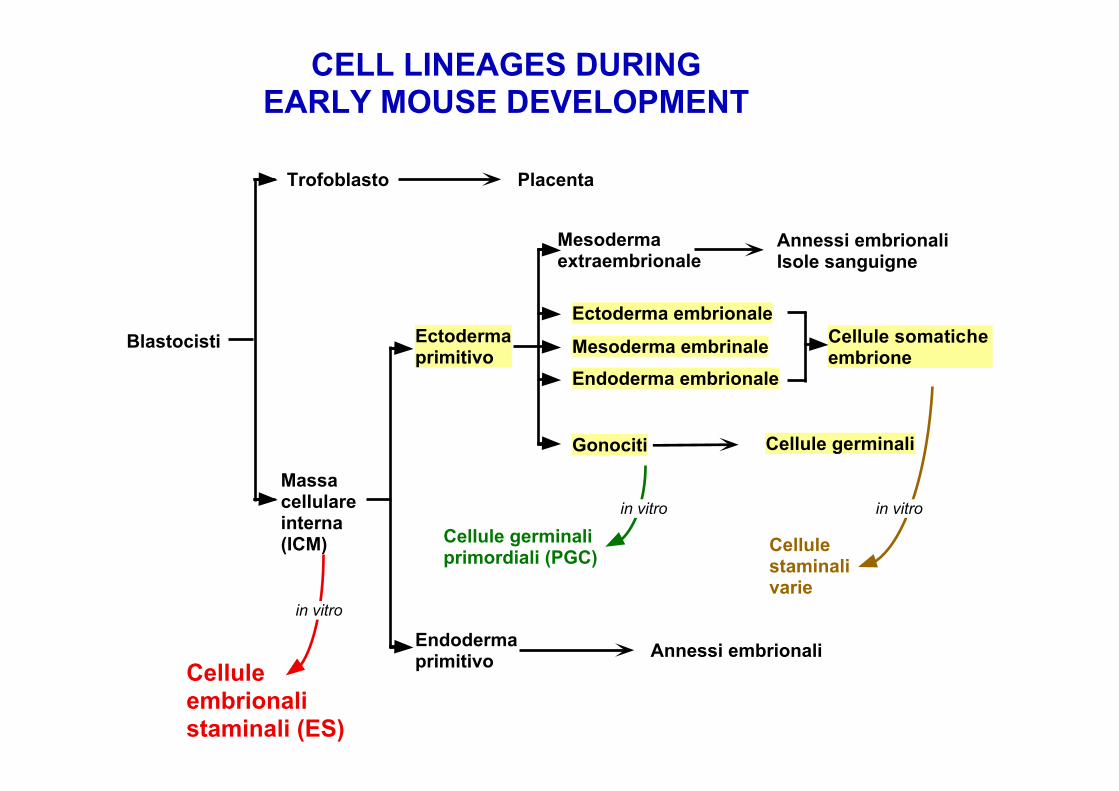
Cellule germinaliprimordiali (PGC)
Cellulestaminalivarie
in vitro in vitro
Blastocisti
Trofoblasto Placenta
Massacellulareinterna(ICM)
Mesodermaextraembrionale
Annessi embrionaliIsole sanguigne
Endodermaprimitivo Annessi embrionali
Celluleembrionalistaminali (ES)
in vitro
CELL LINEAGES DURINGEARLY MOUSE DEVELOPMENT
Ectodermaprimitivo
Ectoderma embrionale
Mesoderma embrinale
Endoderma embrionale
Cellule germinali
Cellule somaticheembrione
Gonociti
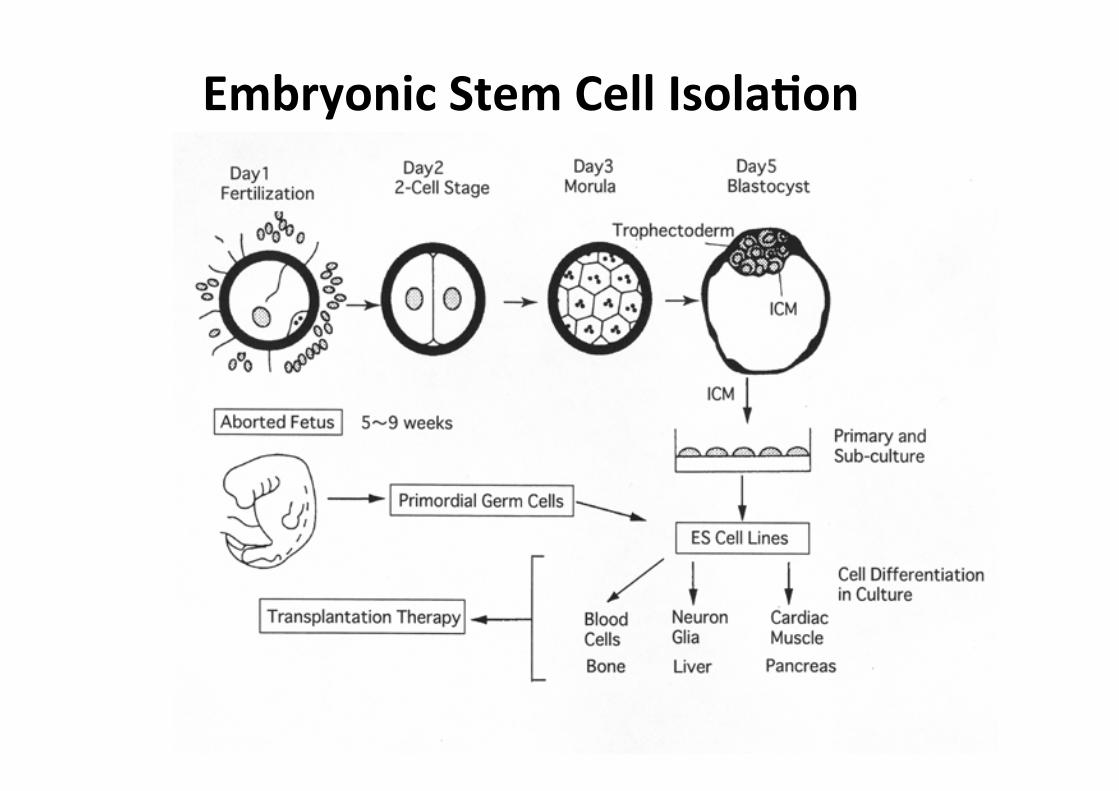
Embryonic Stem Cell IsolaMon

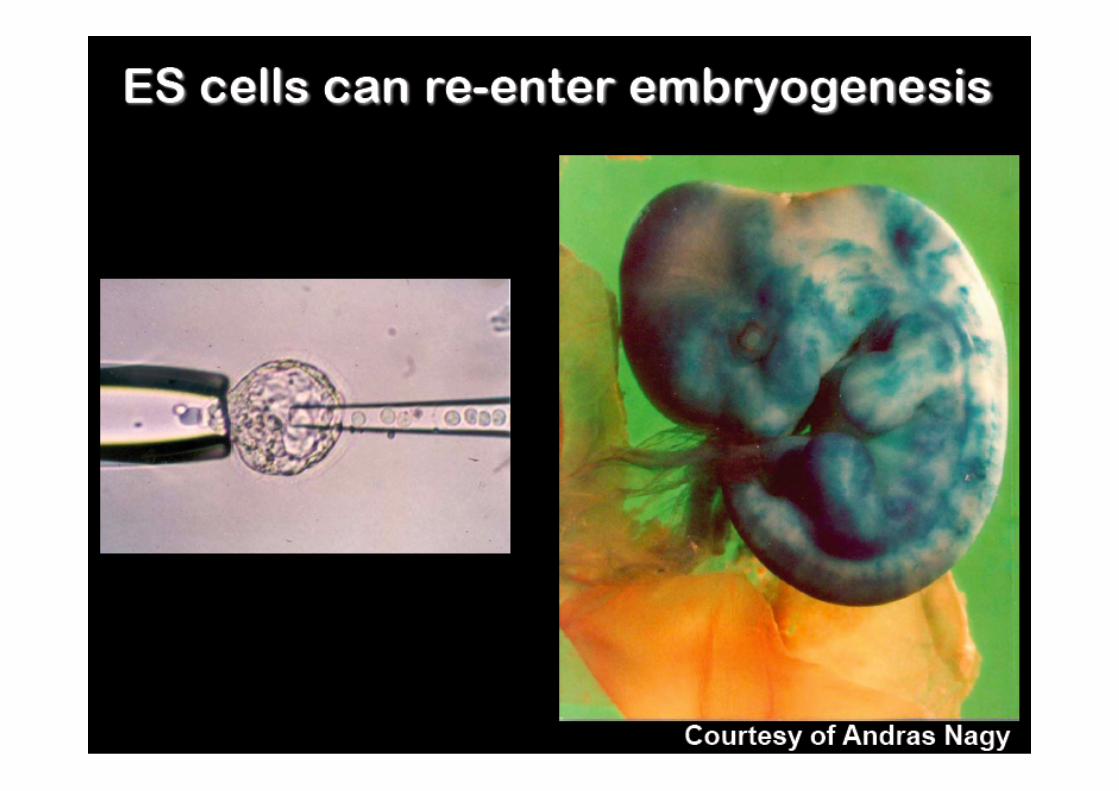
• Cellule totipotenti indifferenziate derivate dalla ICM di blastocisti
• Sono capaci di ricolonizzare ad alta efficienza tutti i tessuti, compresa la linea germinale, se reintrodotte in vivo (morula aggregation, blastocyst injection)
• Queste capacità dipendono in maniera critica da condizioni di coltura che mantengano le cellule ES in uno stato indifferenziato (LIF, feeder fibroblasts)
• Possibilità di manipolazione genetica in vitro (gene targeting, gene trapping)
• Possibilità di differenziamento delle cellule ES in vitro in maniera controllata verso vari tipi di lineages cellulari (embryoid bodies, growth factors)
ES CELLS

Una volta che il DNA è entrato nella cellula ES può seguire diversi desCni: -‐ No integraCon -‐ Random integraCon -‐ Homologous ricombinaCon

Random IntegraMon:
-‐ DNA incorporated anywhere in genome (not targeted)
-‐ copy number can be very high
-‐ disrupts the endogenous DNA at inserCon site
-‐ transgenic animals are usually random integraCons
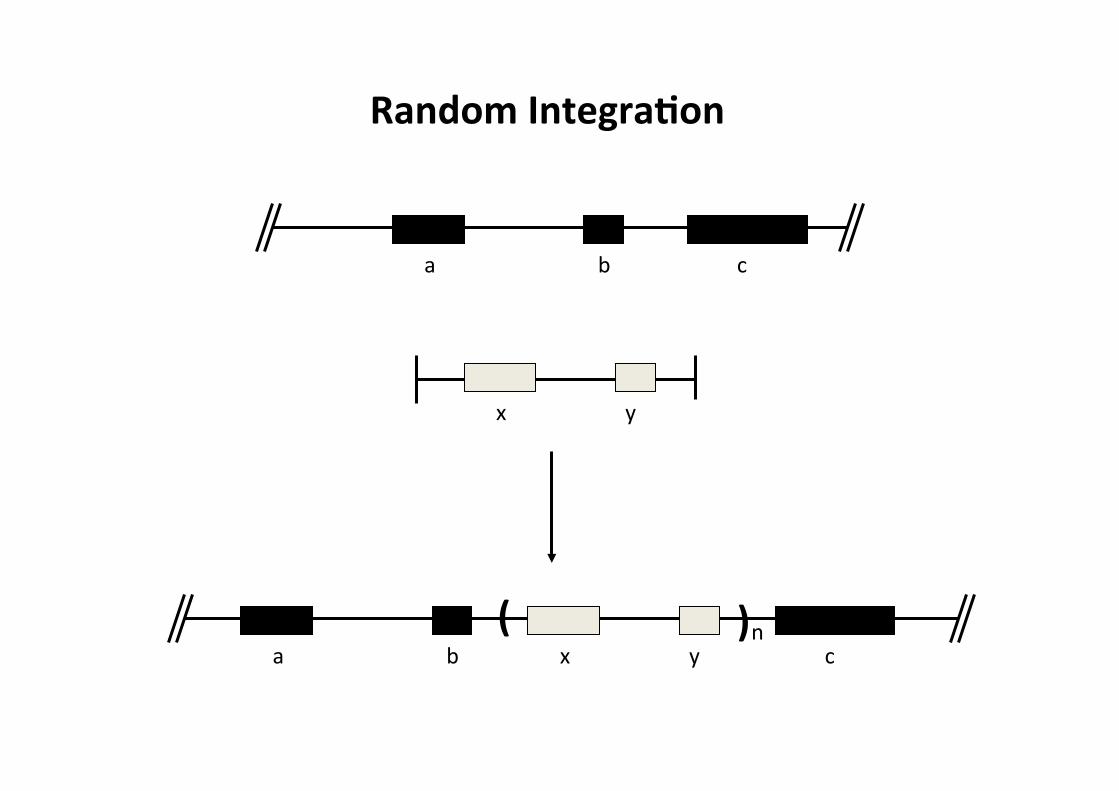
a b c
x y
a b x y c
Random IntegraMon
( ) n

Homologous RecombinaMon:
-‐ driven by the DNA sequences (targeted) -‐ rare (1000X less frequent than random)
-‐ increased efficiency: -‐ isogenic DNA -‐ longer stretches of homology
-‐ results in: -‐ endogenous DNA replaced with exogenous -‐ specific -‐ targeted -‐ copy number = 1
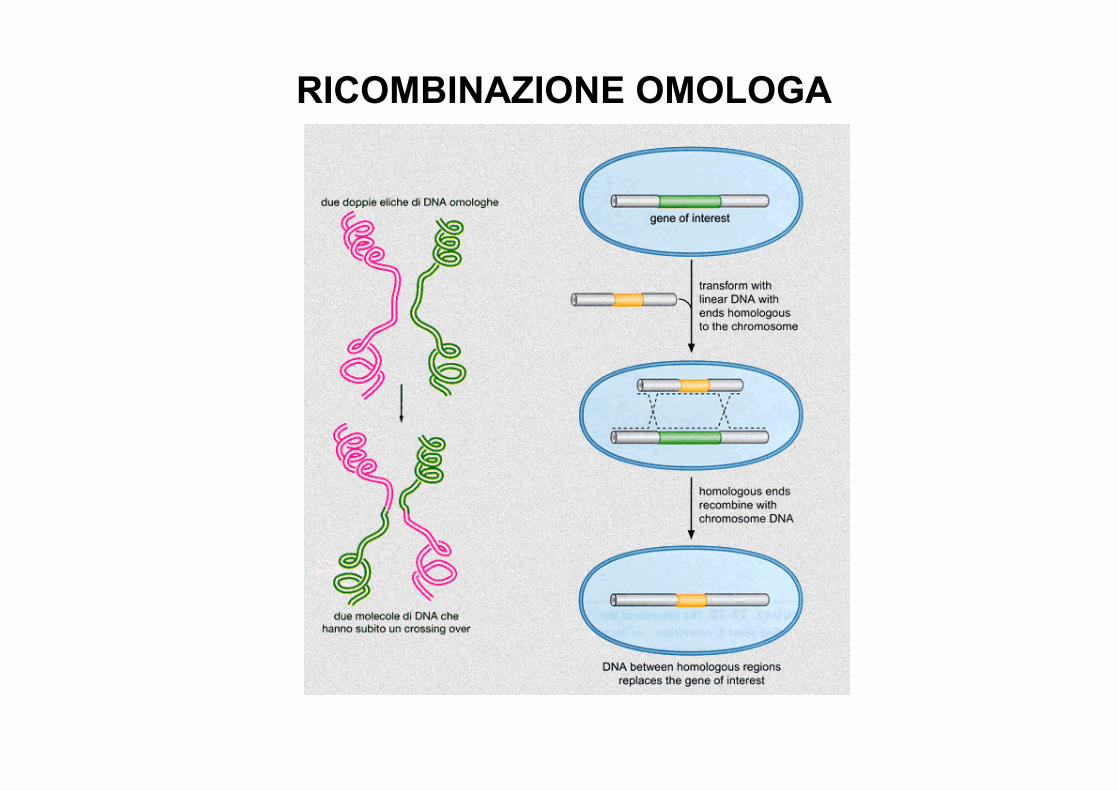
RICOMBINAZIONE OMOLOGA
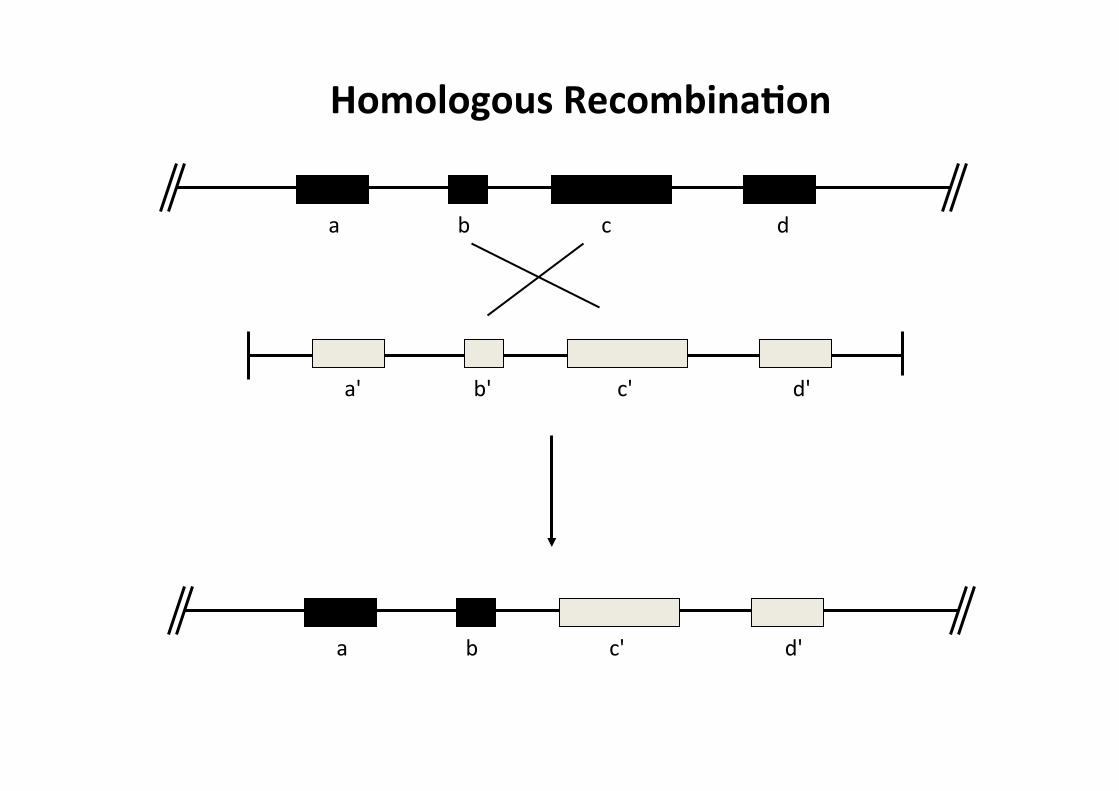
a b c d
a' b' c' d'
a b c' d'
Homologous RecombinaMon
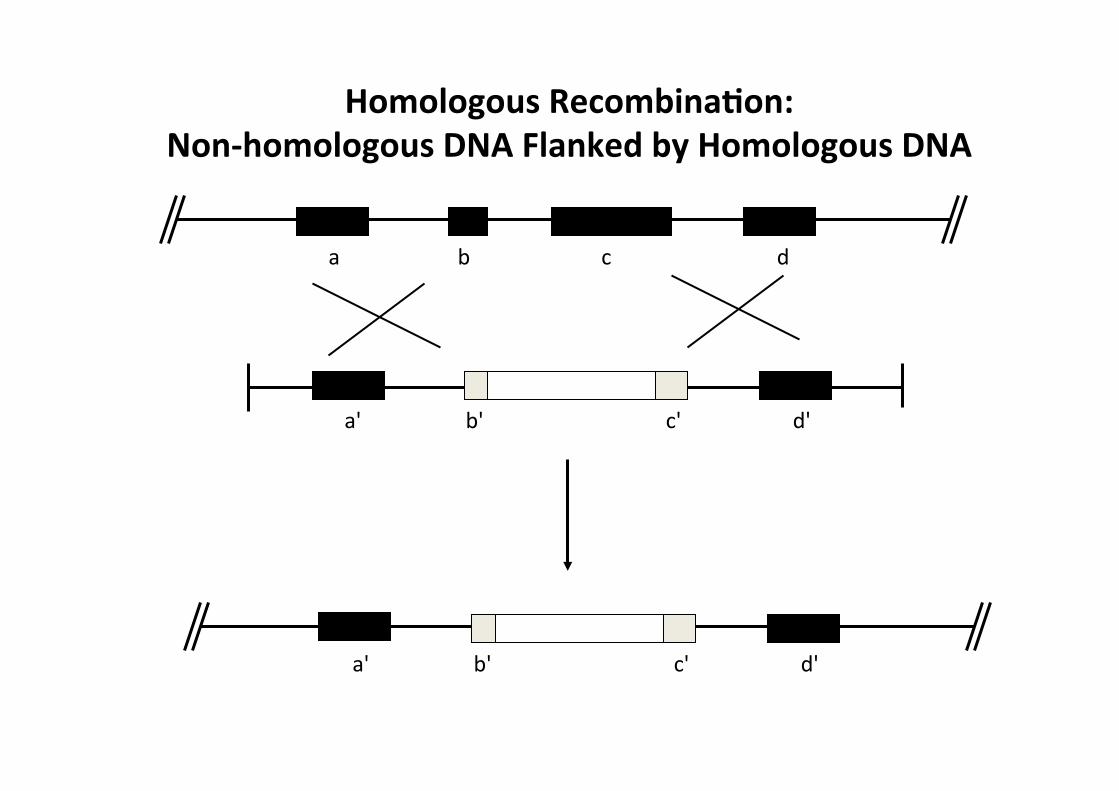
a b c d
a' b' c' d'
a' b' c' d'
Homologous RecombinaMon: Non-‐homologous DNA Flanked by Homologous DNA

How to disMnguish between random integraMon Vs Homologous ricombinaMon?
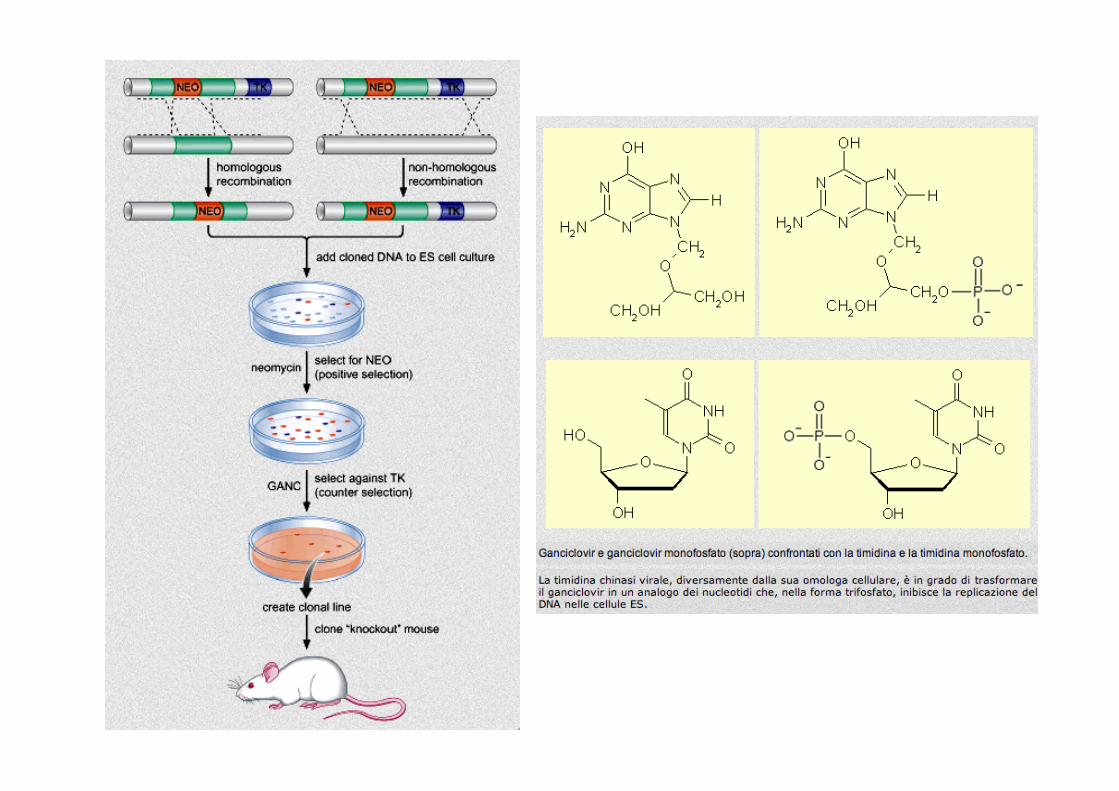
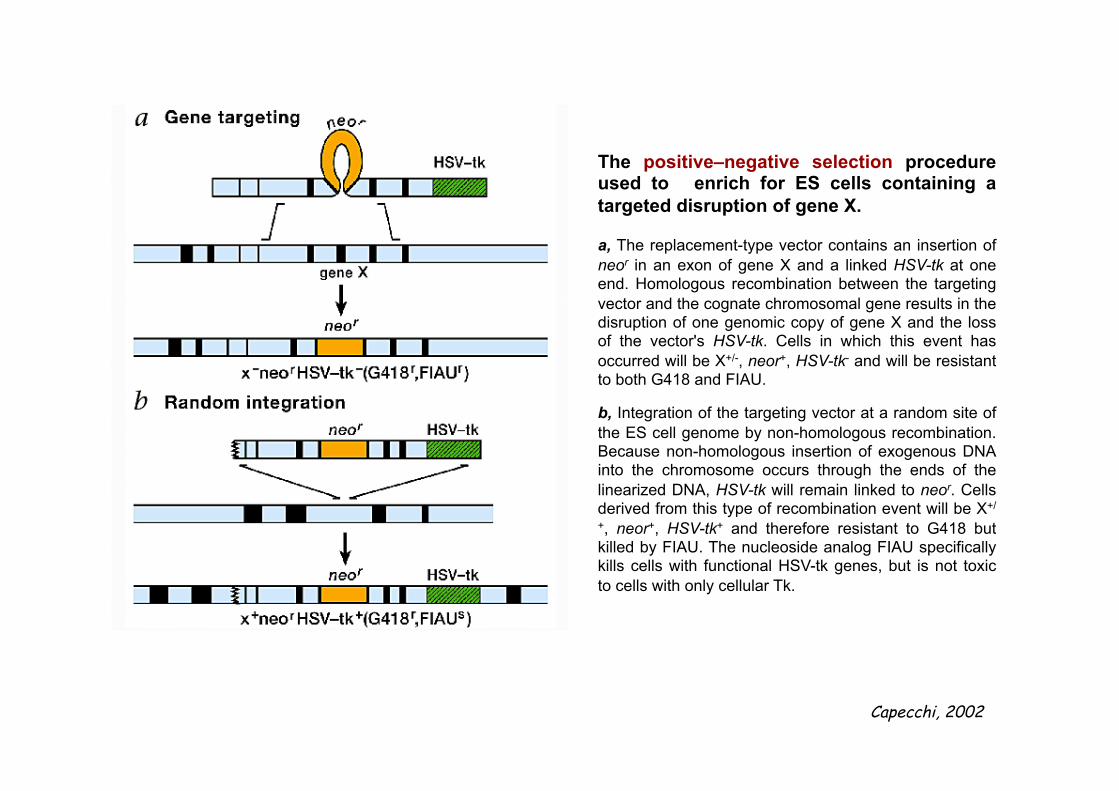
The positive–negative selection procedure used to enrich for ES cells containing a targeted disruption of gene X. a, The replacement-type vector contains an insertion of neor in an exon of gene X and a linked HSV-tk at one end. Homologous recombination between the targeting vector and the cognate chromosomal gene results in the disruption of one genomic copy of gene X and the loss of the vector's HSV-tk. Cells in which this event has occurred will be X+/-, neor+, HSV-tk- and will be resistant to both G418 and FIAU. b, Integration of the targeting vector at a random site of the ES cell genome by non-homologous recombination. Because non-homologous insertion of exogenous DNA into the chromosome occurs through the ends of the linearized DNA, HSV-tk will remain linked to neor. Cells derived from this type of recombination event will be X+/
+, neor+, HSV-tk+ and therefore resistant to G418 but killed by FIAU. The nucleoside analog FIAU specifically kills cells with functional HSV-tk genes, but is not toxic to cells with only cellular Tk.
Capecchi, 2002
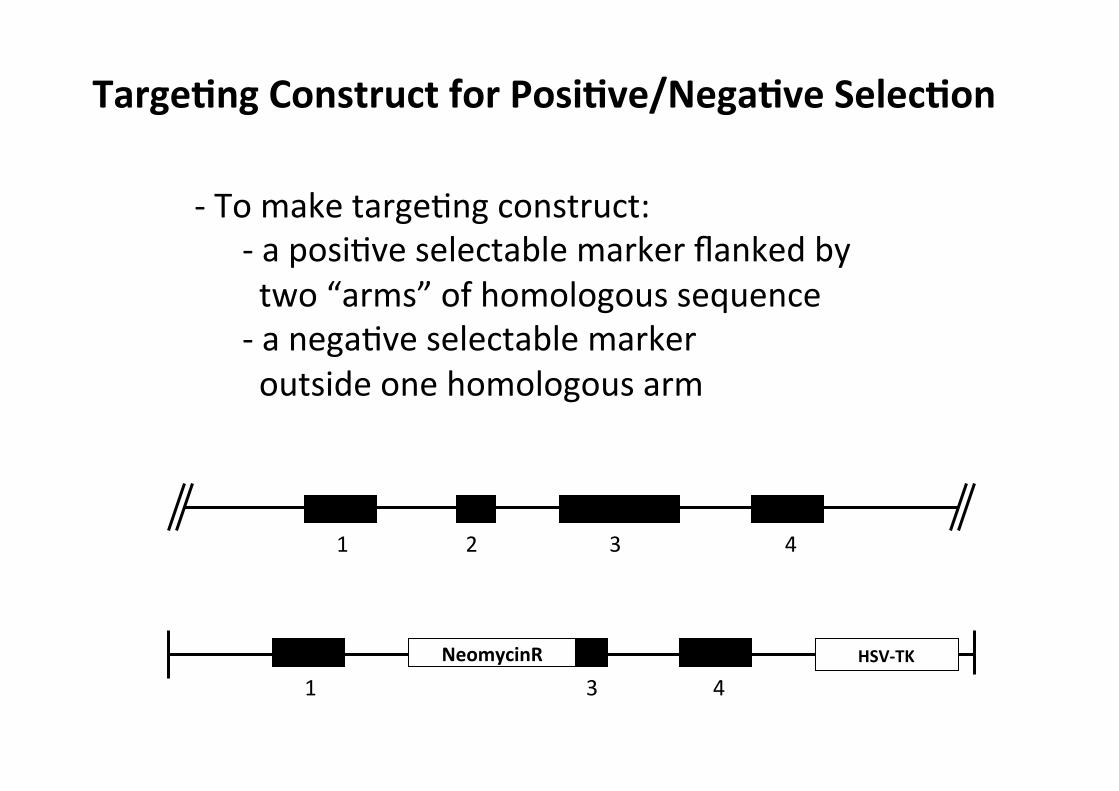
TargeMng Construct for PosiMve/NegaMve SelecMon
-‐ To make targeCng construct: -‐ a posiCve selectable marker flanked by two “arms” of homologous sequence -‐ a negaCve selectable marker outside one homologous arm
1 3 4 NeomycinR HSV-‐TK
1 2 3 4
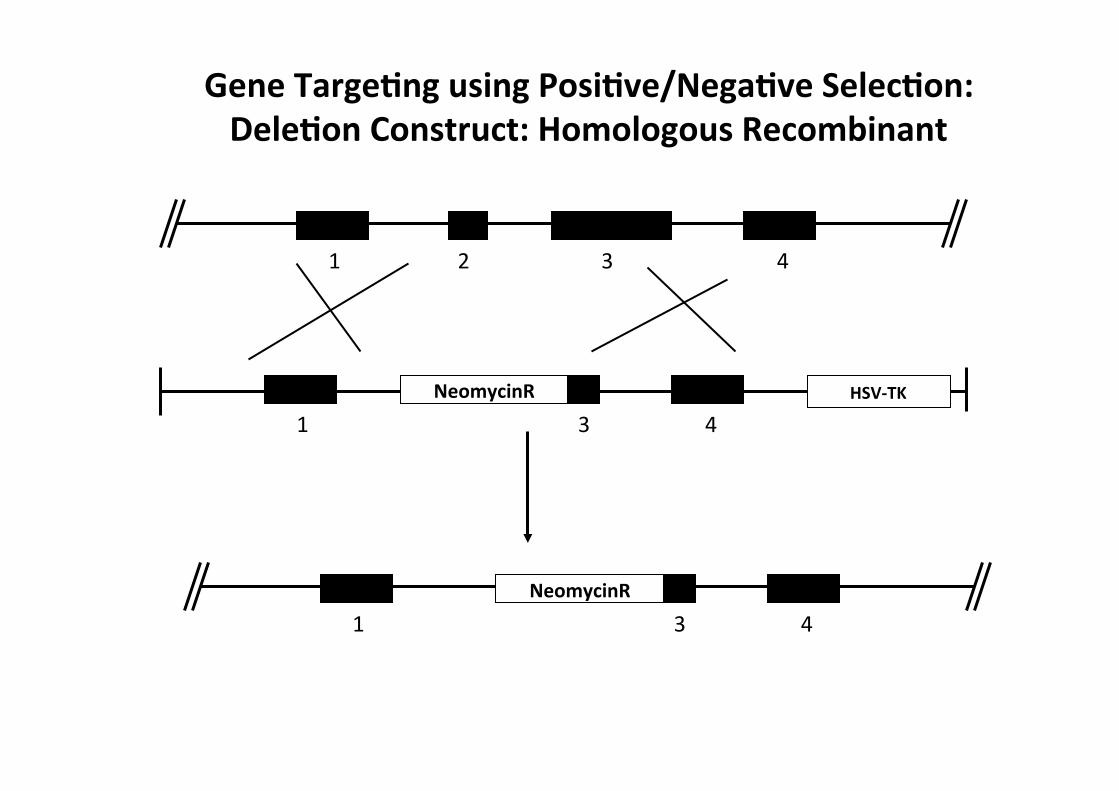
1 2 3 4
1 3 4
1 3 4 NeomycinR HSV-‐TK
NeomycinR
Gene TargeMng using PosiMve/NegaMve SelecMon: DeleMon Construct: Homologous Recombinant
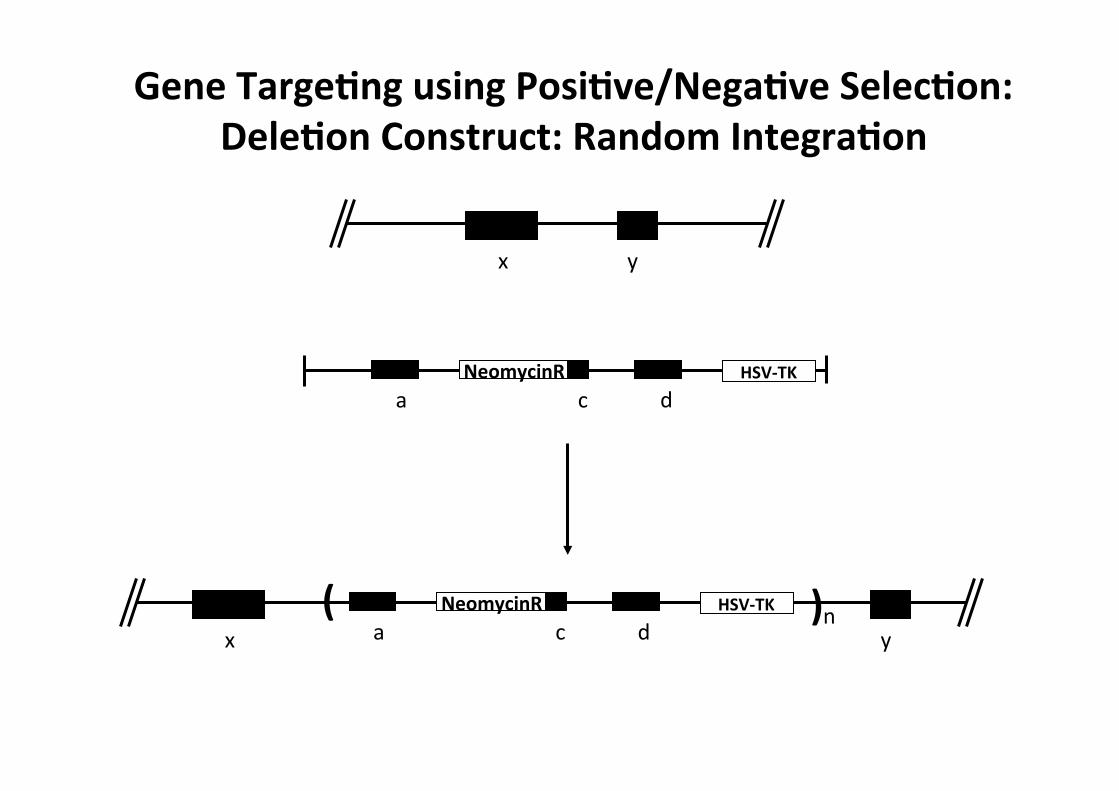
x y
Gene TargeMng using PosiMve/NegaMve SelecMon: DeleMon Construct: Random IntegraMon
x y
a c d NeomycinR HSV-‐TK
a c d NeomycinR HSV-‐TK ( ) n

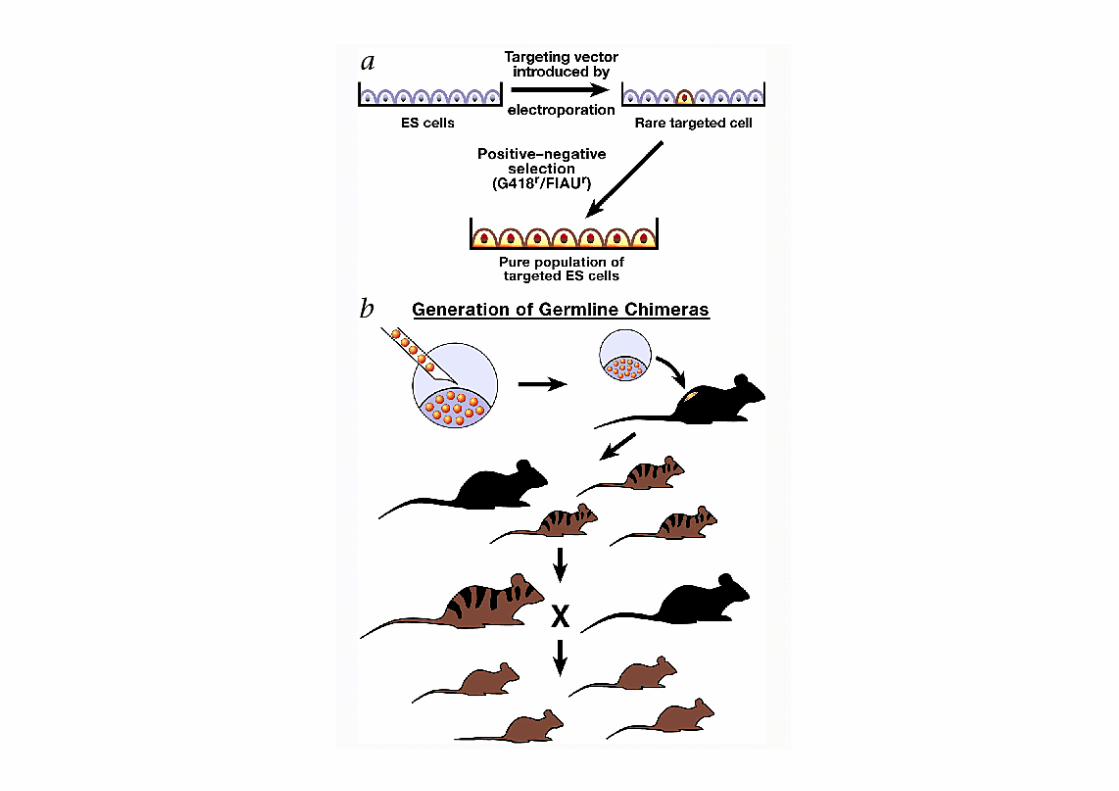
SelecMon Strategy: -‐ PosiMve SelecMon – G418 -‐ Neomycin Resistance gene
-‐ confers resistance to G418 -‐ G418 selects for both: -‐ homologous and random integraCons -‐ kills cells that have not taken up DNA
-‐ NegaMve SelecMon -‐ Gancyclovir -‐ Herpes Simplex Virus Thymidine Kinase (HSV-‐TK) -‐ sensiCve to gancyclovir -‐ selects against random integrants
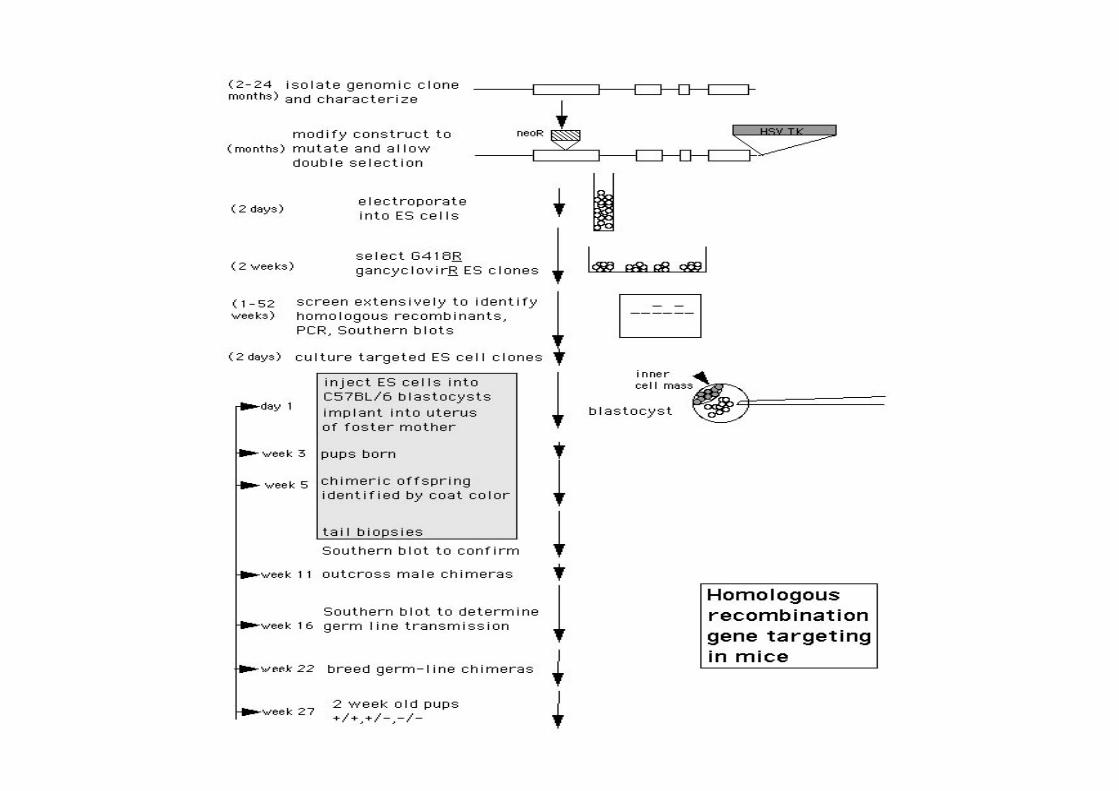

TargeMng: -‐ Electroporate ES cells with TargeCng Construct -‐ Select in G418 and Gancyclovir -‐ enriches for homologous recombinaCon
-‐ -‐ Pick individual colonies of resistant ES cells (100s) -‐ Screen for properly targeted cells -‐ PCR and/or Southern blot
-‐ Use targeted cells to make a mouse
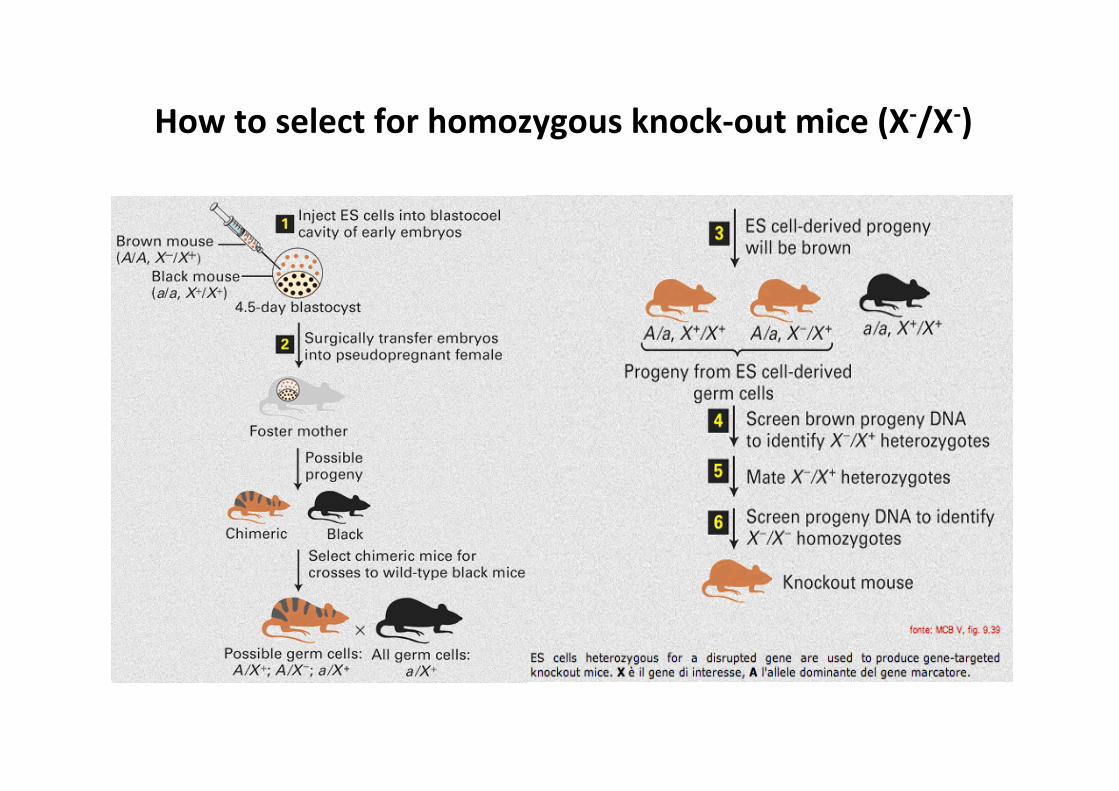
How to select for homozygous knock-‐out mice (X-‐/X-‐)
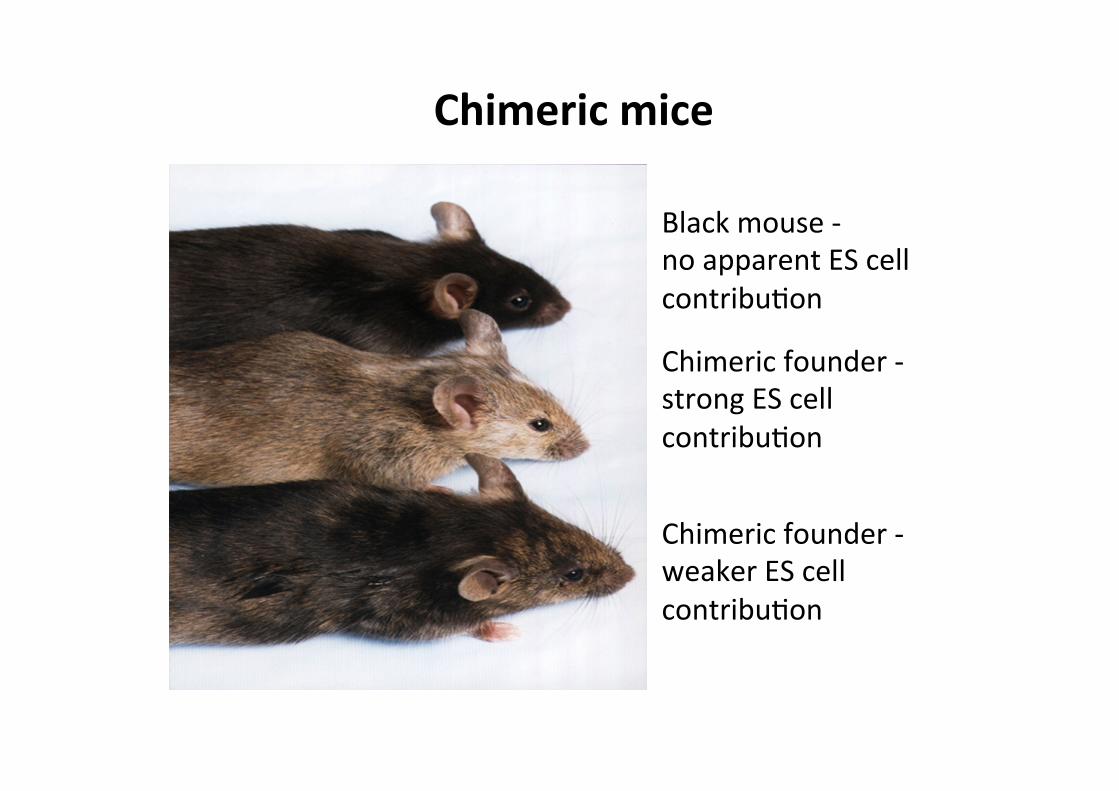
Black mouse -‐ no apparent ES cell contribuCon
Chimeric founder -‐ strong ES cell contribuCon
Chimeric founder -‐ weaker ES cell contribuCon
Chimeric mice
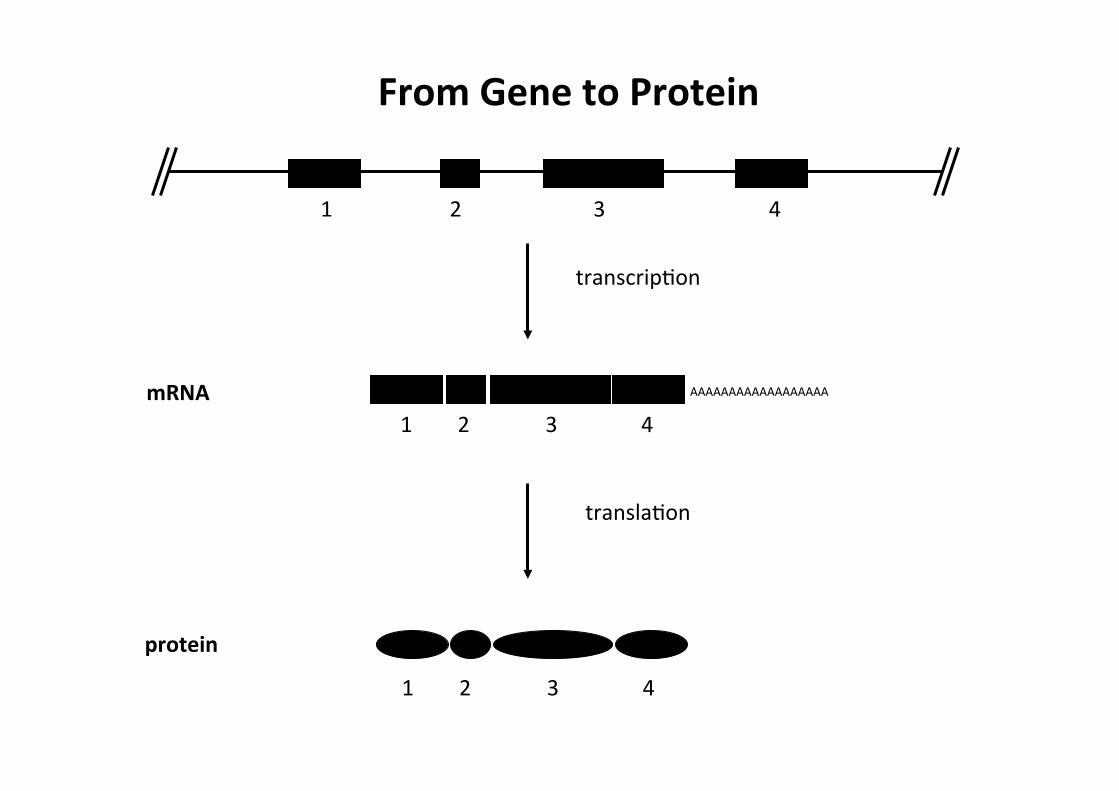
1 2 3 4
1 2 3 4 AAAAAAAAAAAAAAAAAA
protein
mRNA
transcripCon
translaCon
From Gene to Protein
1 2 3 4


Analysis of Mice: Possible Outcomes -‐ Phenotype: happy -‐ No homozygous knockouts seen
-‐ embryonic lethal -‐ No heterozygous knockouts seen
-‐ haploinsufficiency -‐ No phenotype. -‐ funcConal redundancy -‐ unable to detect (wrong assay) -‐ incomplete protein inacCvaCon -‐ Unexpected phenotype. -‐ loss of protein (happy) -‐ abnormal protein
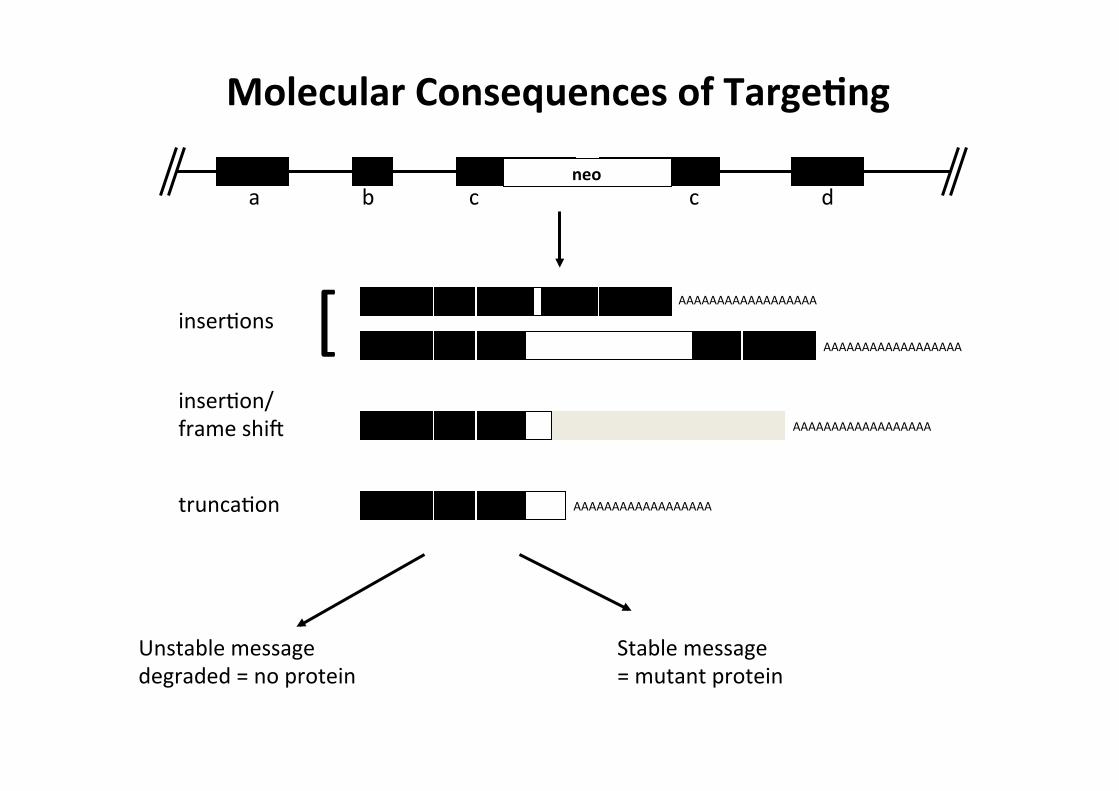
a d b c neo
c
AAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAAA
AAAAAAAAAAAAAAAAAA
[ inserCons
truncaCon
inserCon/ frame shia
Unstable message degraded = no protein
Stable message = mutant protein
Molecular Consequences of TargeMng

Prima di analizzare il fenoCpo mutante, si deve verificare che non vi sia proteina residua espressa dal locus inadvato. 1-‐ "leaky mutaCon" con espressione residua di proteina (hypomorphs)
(promotori cripCci/alternaCvi, read-‐through transcripCon, traduzione da AUG cripCci, aberrant splicing) 2-‐ presenza di polipepCdi tronchi (possono acquisire nuove proprietà, es. dominant negaCve) Alcuni di quesC knockout incompleC cosCtuiscono varianC alleliche con fenoCpi interessanC (es.
rescue parziale di un fenoCpo letale, possono rivelare funzioni di un gene inaspeJate; es. N-‐myc)
Possibili soluzioni: Rimozione di sequenze codificanM o di elemenM regolatori di altri geni La rimozione di tud gli esoni codificanC di un gene da inadvare può evitare il problema di espressione residua di proteina. Però la generazione di ampie delezioni genomiche può provocare effed inaJesi:
-‐ perdita di geni non idenCficaC (all'interno di introni, o codificaC dallo strand opposto) -‐ perdita di elemenC. regolatori che controllano l'espressione di altri geni (es. MRF4 e MRF5, CD3 e Oct-‐1)
No phenotype – Incomplete Knockout

• Assenza di feno-po o feno-po lieve: Ridondanza e meccanismi compensatori Assenza di feno)po "the gene does not play a role in the process" !? Il gene non è essenziale per un certo processo. Il gene può essere coinvolto nel processo, ma
in sua assenza la funzione viene svolta da altri geni ("ridondanza"). Proteine correlate possono funzionalmente compensare l'assenza di un singolo membro di
una famiglia genica (es. src-‐related tyrosine kinase, myogenic factors, neurotrophic factors, dystrophin-‐utrophin, cadherins).
Due possibili spiegazioni:
1. Overlapping func-ons Ridondanza funzionale di diversi pathways geneCci, con funzioni parzialmente overlapping
(es. IFN-‐receptors).
2. Compensa-on Il gene normalmente svolge una certa funzione ma in sua assenza un secondo gene
(normalmente non coinvolto nella stessa funzione) lo sosCtuisce: upregula-on
Generazione di “knockout doppi o mulMpli” elimina possibilità di overlapping funcCons e meccanismi compensatori fra due o più geni (fenoCpo più grave).
No phenotype – Redundancy

Embryonic Lethal -‐ Now What?
-‐ Conclusion: -‐ important for embryonic development -‐ Is protein important later in life? -‐ CondiMonal Knockouts -‐ Cssue specific -‐ temporally regulated

Gene targeCng standard: inadvazione di un gene in tud i tessuC del corpo dall'inizio dello sviluppo e per tuJa la vita.
Gene targeMng condizionale: permeJe di controllare inabvazione genica in modo definito per un certo tessuto o un certo stadio di sviluppo.
ParCcolarmente uCle quando inadvazione completa determina fenoCpo letale. Se un gene ha un paJern di espressione complesso, inadvazione tessuto-‐specifica permeJe di definire il ruolo del gene in un certo tessuto, senza compromeJere le altre funzioni nell'organismo.
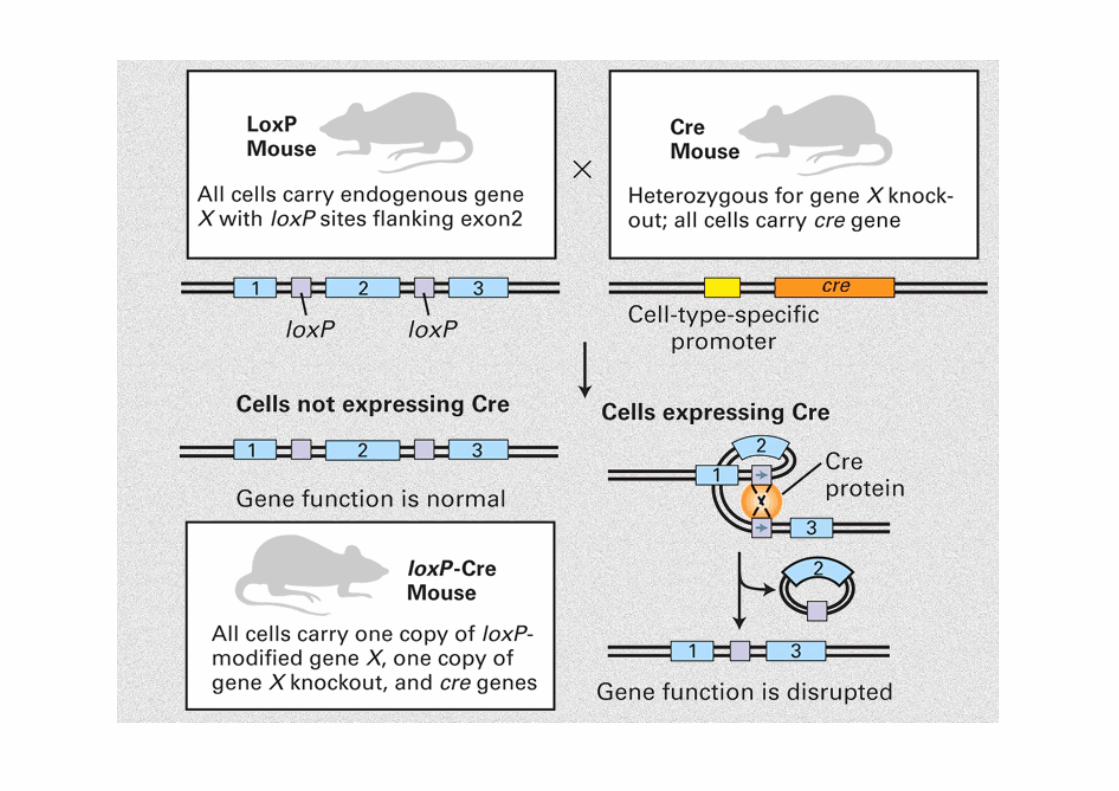
• Tissue-‐specific knockout a) generazione di topi transgenici che esprimono Cre ricombinasi sodo il controllo di promotore
tessuto-‐specifico. b) generazione di topi con siM loxP fiancheggianM il gene da inabvare. Incrocio topi a) x b) : topi dove gene di interesse è stato inabvato solo nelle cellule in cui Cre è
espresso.
Database di linee di topi transgenici Cre: hNp///www.mshri.on.ca/develop/Nagy/Cre.htm • Inducible gene targeMng Controllo della funzione genica in maniera tempo-‐dipendente. Uso di sistema di ricombinazione basato su Cre, in cui l'espressione di Cre è sodo il controllo di un
promotore inducibile oppure va al nucleo solo in risposta ad uno sMmolo preciso . Esempi: Cre ER: ricombinazione solo con tamoxifene tet system: silenziamento di Cre con tetraciclina reverse tet system: induzione di Cre con tetraciclina
CondiMonal Gene TargeMng

Cre/loxP System
-‐ Cre Recombinase -‐ site-‐specific recombinase -‐ P1 bacteriophage -‐ recombinaCon between two loxP sites -‐ no other cofactors required -‐ LoxP sites -‐ 34bp sequence -‐ two inverted 13bp repeats surrounding 8bp core
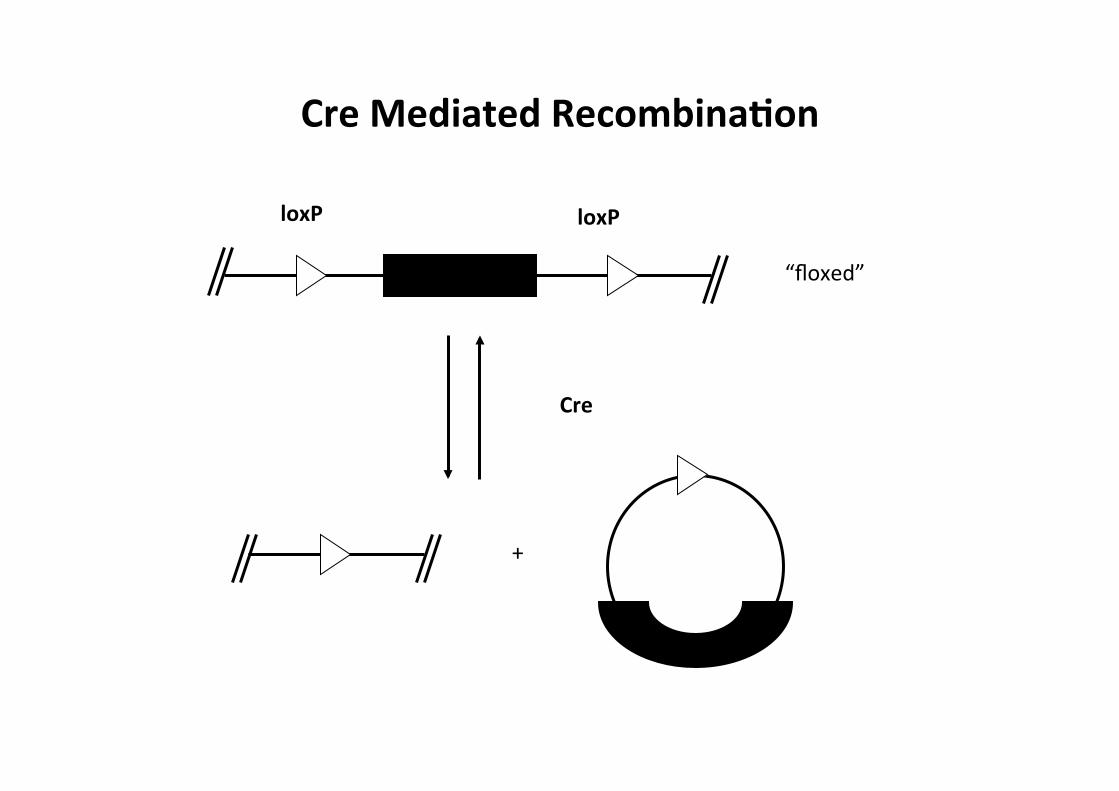
Cre Mediated RecombinaMon
+
Cre
loxP loxP
“floxed”
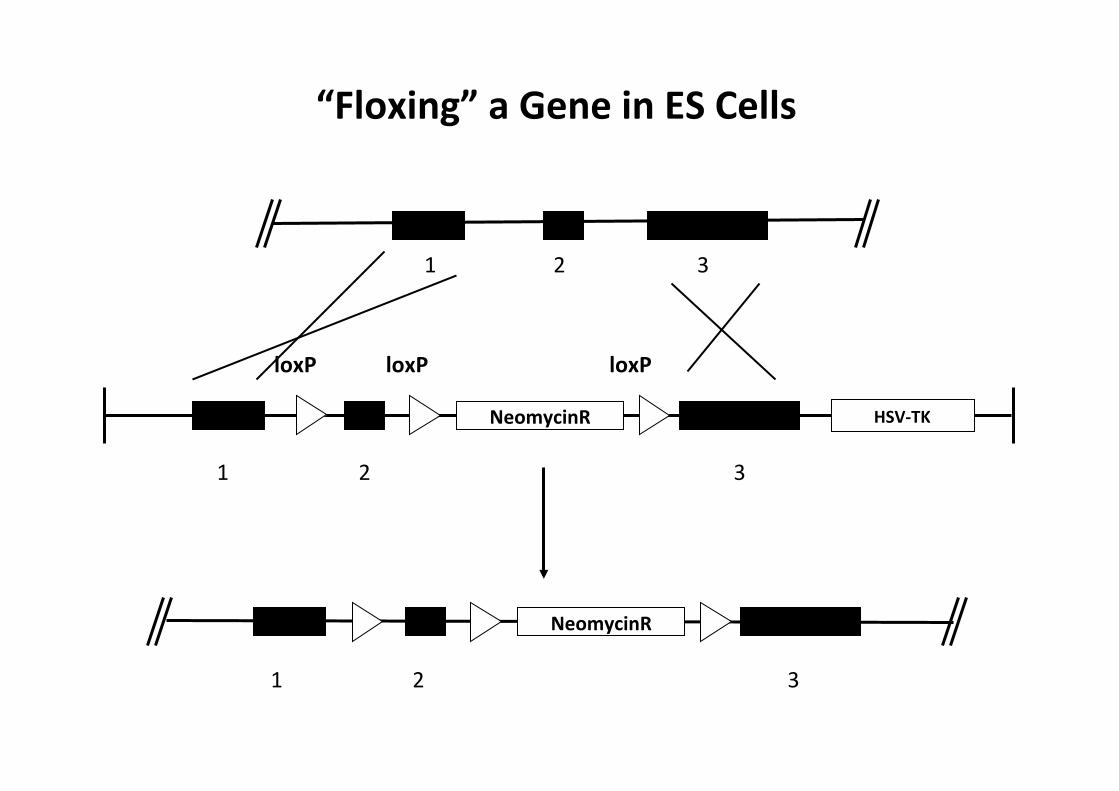
1 2 3
HSV-‐TK NeomycinR
“Floxing” a Gene in ES Cells
NeomycinR
1 2 3
1 2 3
loxP loxP loxP
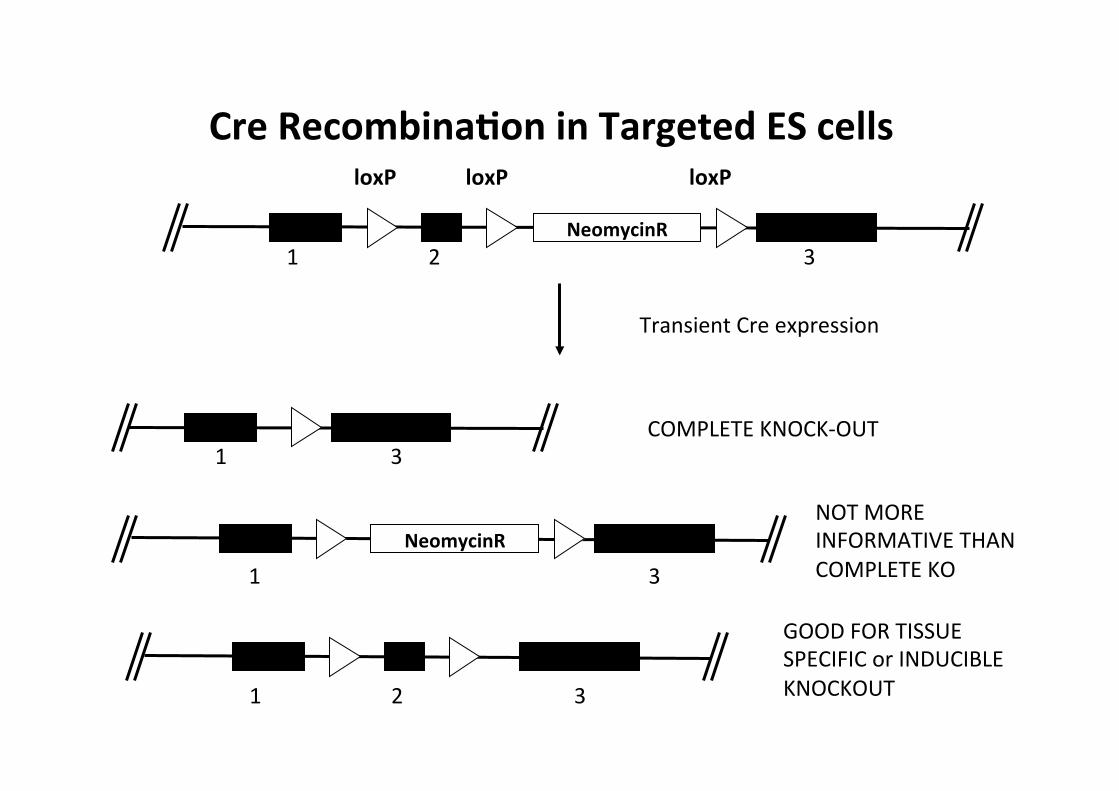
Cre RecombinaMon in Targeted ES cells
NeomycinR 1 2 3
Transient Cre expression
1 3
NeomycinR
1 3
1 2 3
COMPLETE KNOCK-‐OUT
NOT MORE INFORMATIVE THAN COMPLETE KO
GOOD FOR TISSUE SPECIFIC or INDUCIBLE KNOCKOUT
loxP loxP loxP

Tissue Specific Cre-‐Expressing Mouse
Transgenic Mouse
Cre Tissue Specific Promoter
Expresses Cre in a subset of Mssues Note: many Cssue-‐specific Cre-‐expressing mice have been made, more are being made.
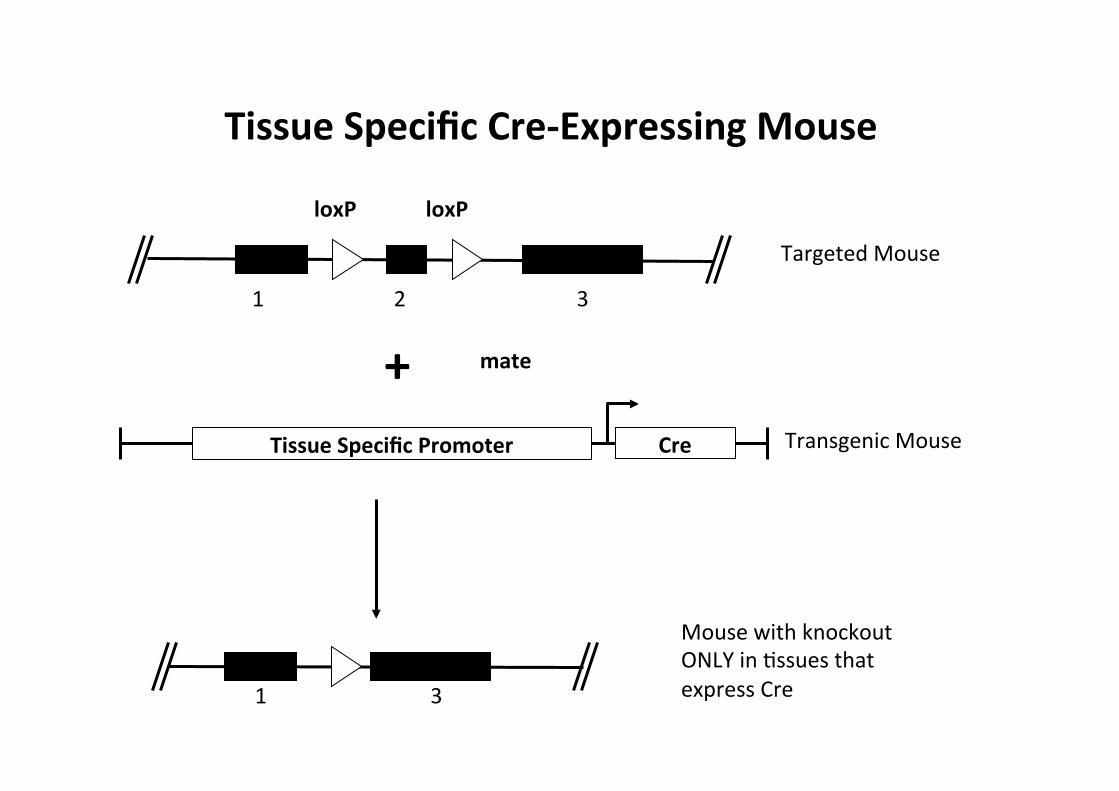
Tissue Specific Cre-‐Expressing Mouse
1 2 3
Cre Tissue Specific Promoter
1 3
Targeted Mouse
Transgenic Mouse
+
Mouse with knockout ONLY in Cssues that express Cre
mate
loxP loxP
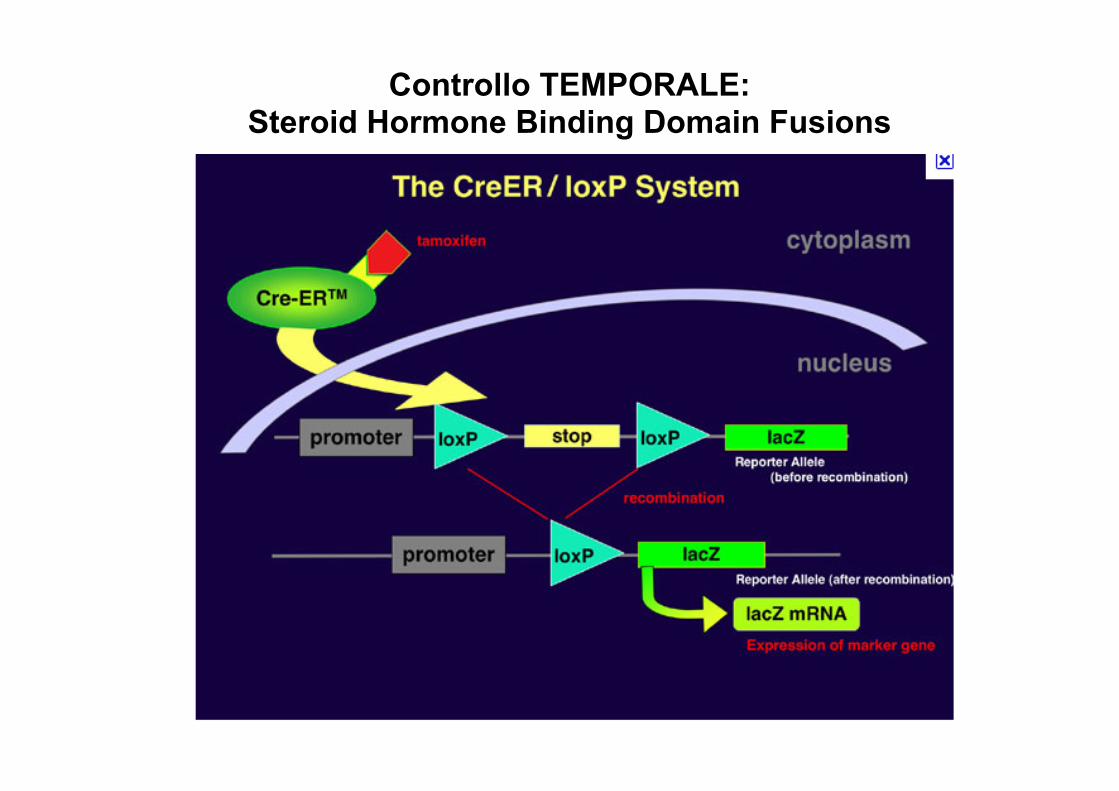
Controllo TEMPORALE: Steroid Hormone Binding Domain Fusions

Temporal + SpaMal RegulaMon of Cre: Steroid Hormone Binding Domain Fusions
ERT-‐Cre Tissue Specific Promoter
ERT-‐Cre
cytoplasm
nucleus
ERT-‐Cre
cytoplasm
nucleus
No hormone + Tamoxifen

L'uso di topi knockout e' ovvio per studiare la funzione di un gene in tuJa la vita, in tud i tessuC e quando mi interessa il suo LOSS OF FUNCTION. L'uso di Knockout condizionali permeJe di focalizzare l'aJenzione su parCcolari tessuC oppure a parCre da un momento ben preciso definito dal ricercatore. DeJo ciò, molte malade sono dovute a mutazioni punCformi, alleli neomorfici etc etc, basC pensare al ruolo di oncogeni advaC nel cancro… KNOCK-‐OUT Vs KNOCK-‐IN NON esiste alcuna differenza praCca. Cambia lo scopo: con il KO INATTIVO un gene. Con il Knock-‐in realizzo una ricombinazione omologa con lo scopo di introdurre variazioni piu' fini, per esempio mutazioni a livello di
specifici aminoacidi, piccole delezioni di enhancers etc
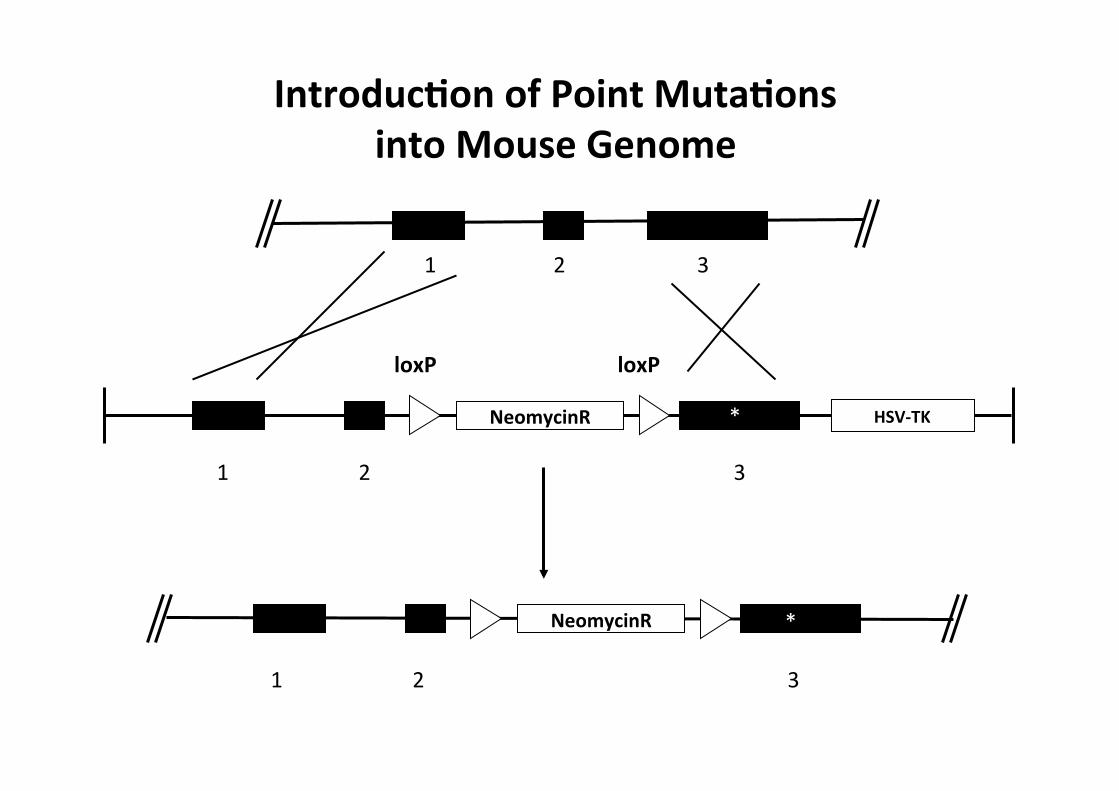
1 2 3
HSV-‐TK NeomycinR
IntroducMon of Point MutaMons into Mouse Genome
NeomycinR
1 2 3
1 2 3
*
*
loxP loxP
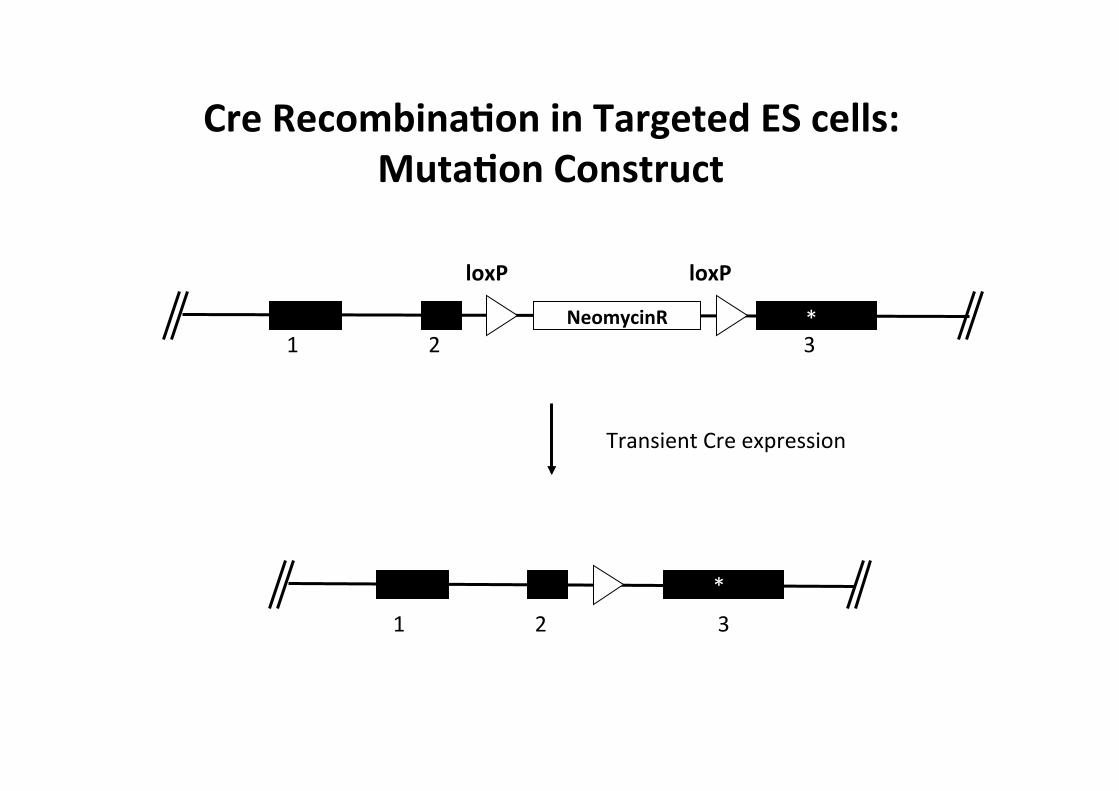
Cre RecombinaMon in Targeted ES cells: MutaMon Construct
NeomycinR 1 2 3
Transient Cre expression
1 2 3
*
*
loxP loxP

• How can we drive Cssue specific point mutaCon?
=> TranscripConal STOP casseJe

• DOMANDA: Con il sistema cre-lox inattivo un gene (ovvero ne “faccio un Knockout”, se condizionale, in un tessuto a piacere etc). Ma sei io volessi attivare l’espressione in una forma mutante di gene endogeno in certo tessuto?
• Risposta: E' possibile, realizzanto il knock-in della forma mutata, con l’accortezza di utilizzare delle cassette di STOP trascrizionali
• transcriptional stop cassette (che sono delle sequenze contenti codoni stop in tutti i frames di lettura)
SPAT
IAL CO
NTR
OL OF KN
OCK
-‐OUT
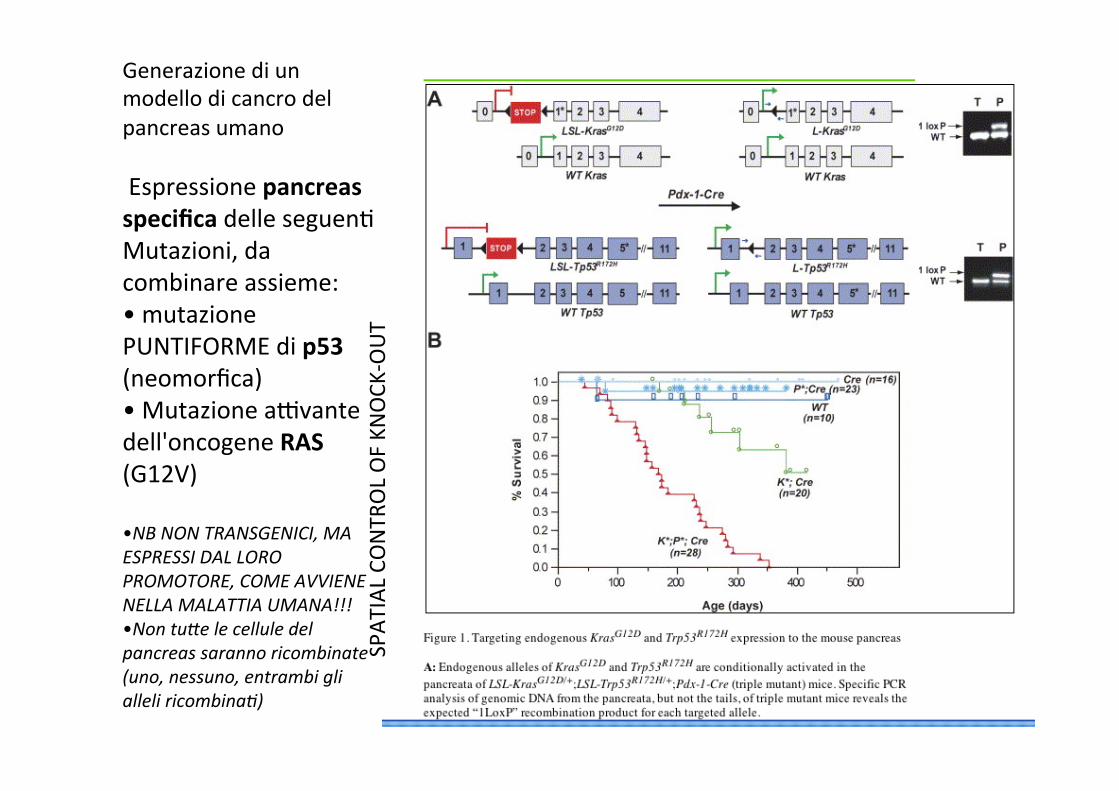
Generazione di un modello di cancro del pancreas umano Espressione pancreas specifica delle seguenC Mutazioni, da combinare assieme: • mutazione PUNTIFORME di p53 (neomorfica) • Mutazione advante dell'oncogene RAS (G12V)
• NB NON TRANSGENICI, MA ESPRESSI DAL LORO PROMOTORE, COME AVVIENE NELLA MALATTIA UMANA!!! • Non tuNe le cellule del pancreas saranno ricombinate (uno, nessuno, entrambi gli alleli ricombina))
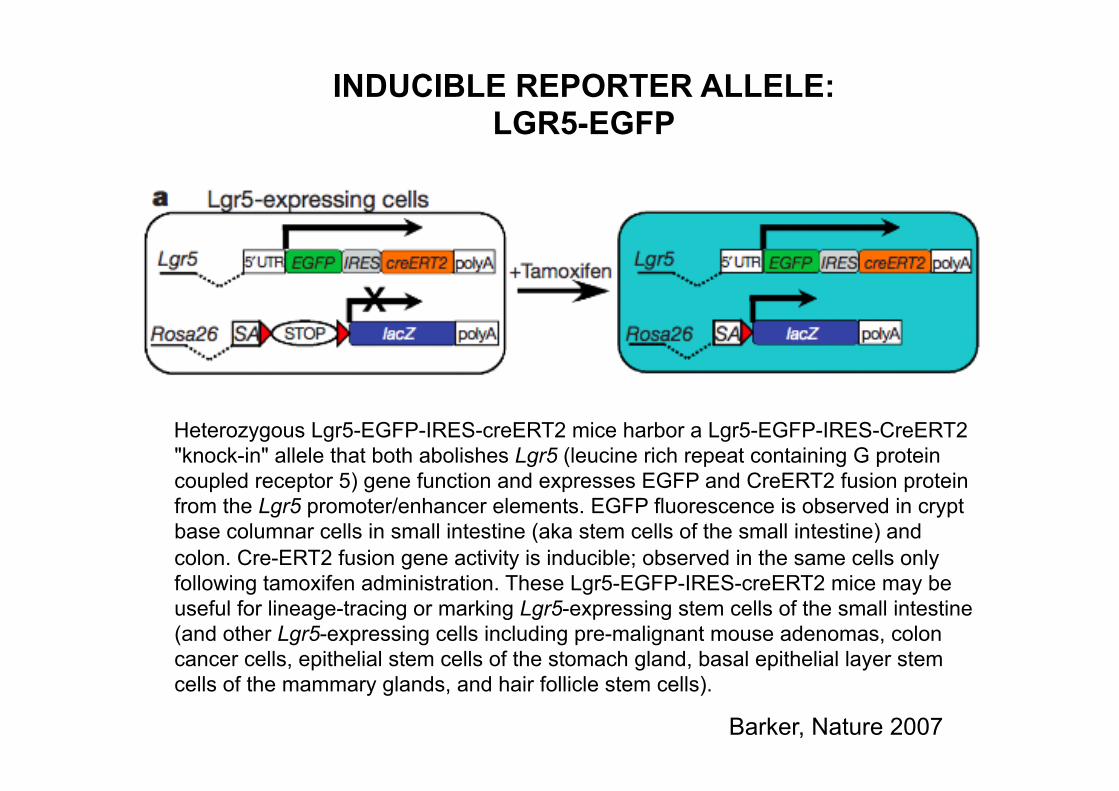
Heterozygous Lgr5-EGFP-IRES-creERT2 mice harbor a Lgr5-EGFP-IRES-CreERT2 "knock-in" allele that both abolishes Lgr5 (leucine rich repeat containing G protein coupled receptor 5) gene function and expresses EGFP and CreERT2 fusion protein from the Lgr5 promoter/enhancer elements. EGFP fluorescence is observed in crypt base columnar cells in small intestine (aka stem cells of the small intestine) and colon. Cre-ERT2 fusion gene activity is inducible; observed in the same cells only following tamoxifen administration. These Lgr5-EGFP-IRES-creERT2 mice may be useful for lineage-tracing or marking Lgr5-expressing stem cells of the small intestine (and other Lgr5-expressing cells including pre-malignant mouse adenomas, colon cancer cells, epithelial stem cells of the stomach gland, basal epithelial layer stem cells of the mammary glands, and hair follicle stem cells).
INDUCIBLE REPORTER ALLELE: LGR5-EGFP
Barker, Nature 2007
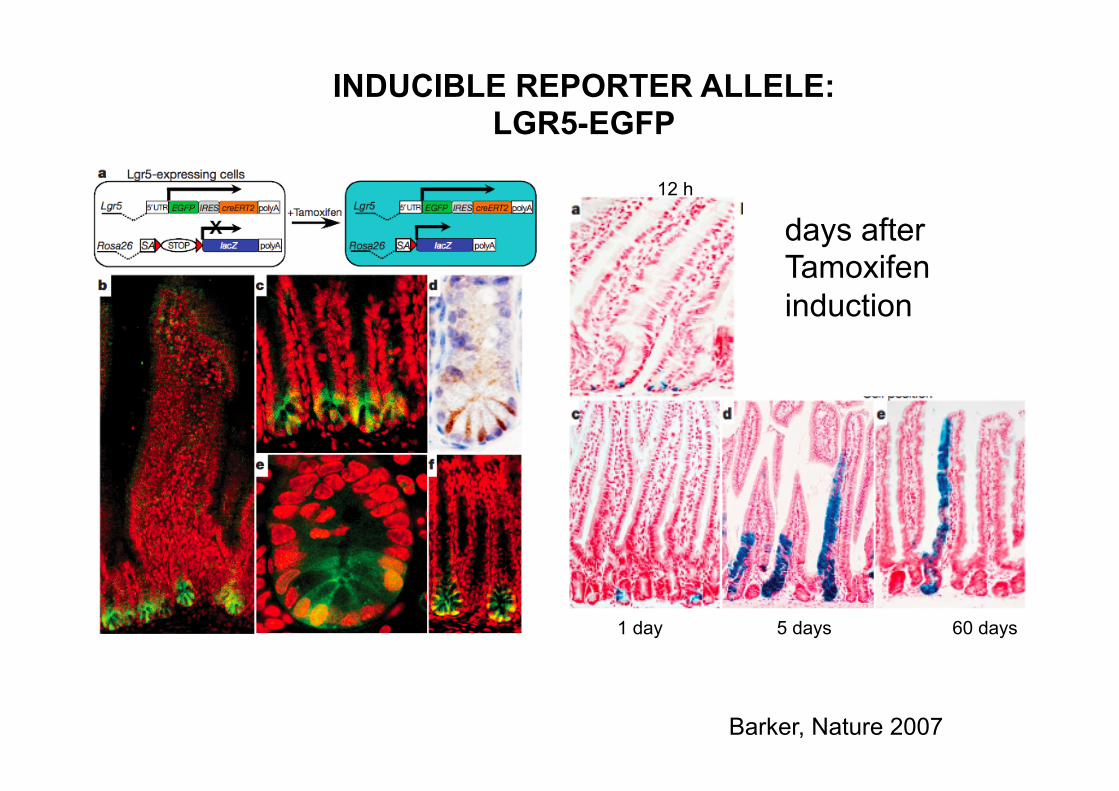
12 h
1 day 5 days 60 days
days after Tamoxifen induction
INDUCIBLE REPORTER ALLELE: LGR5-EGFP
Barker, Nature 2007