Cervelletto, nuclei della base, controllo della postura, sistema ventricolare
26
Lezione 15 (CERVELLETTO) 10/05/05 Questo è uno schema generale di come sono organizzate le vie in ingresso e le vie in uscita. È uno schema estremamente semplificato, però ci dà l’idea di quello che succede, di qual è il circuito interno alla corteccia cerebellare (di cui abbiamo visto la volta scorsa una descrizione più particolareggiata e cellulare). La figura ci dice anche che le vie in ingresso sono di due tipi: quella delle cellule rampicanti che arrivano dal nucleo olivare inferiore, che portano una afferenza eccitatoria direttamente sui grossi neuroni del Purkinje. Mentre le altre afferenze che arrivano con la via delle fibre muscoidi hanno sulle cellule del Purkinje un significato inibitorio. Quindi abbiamo afferenze di due tipi sulla stessa cellula, vedremo che non solo c’è una differenza tra il significato eccitatorio o inibitorio dell’afferenza sulla cellula del Purkinje, ma ci sono anche differenti modalità di eccitazione della cellula del Purkinje da parte delle due afferenze. In questo modo la modulazione è molto diversa a seconda che arrivi dalla fibra rampicante oppure dalle fibre muscoidi. Tutto, comunque, converge sulle cellule del Purkinje e queste sono le uniche cellule efferenti dalla corteccia cerebellare ed hanno un’attività inibitoria sui nuclei profondi cerebellari. Le vie in uscita dal cervelletto, come troverete su molti libri, in realtà non escono dal cervelletto, non ci sono delle vie efferenti “lunghe” che escono dall’insieme corteccia-nuclei cerebellari. Le vie in uscita dalle cellule del Purkinje (via 1
-
Upload
api-3704348 -
Category
Documents
-
view
9.863 -
download
5
Transcript of Cervelletto, nuclei della base, controllo della postura, sistema ventricolare

Lezione 15 (CERVELLETTO) 10/05/05
Questo è uno schema generale di come sono organizzate le vie in ingresso e le vie in uscita. È uno schema estremamente semplificato, però ci dà l’idea di quello che succede, di qual è il circuito interno alla corteccia cerebellare (di cui abbiamo visto la volta scorsa una descrizione più particolareggiata e cellulare). La figura ci dice anche che le vie in ingresso sono di due tipi: quella delle cellule rampicanti che arrivano dal nucleo olivare inferiore, che portano una afferenza eccitatoria direttamente sui grossi neuroni del Purkinje. Mentre le altre afferenze che arrivano con la via delle fibre muscoidi hanno sulle cellule del Purkinje un significato inibitorio. Quindi abbiamo afferenze di due tipi sulla stessa cellula, vedremo che non solo c’è una differenza tra il significato eccitatorio o inibitorio dell’afferenza sulla cellula del Purkinje, ma ci sono anche differenti modalità di eccitazione della cellula del Purkinje da parte delle due afferenze. In questo modo la modulazione è molto diversa a seconda che arrivi dalla fibra rampicante oppure dalle fibre muscoidi. Tutto, comunque, converge sulle cellule del Purkinje e queste sono le uniche cellule efferenti dalla corteccia cerebellare ed hanno un’attività inibitoria sui nuclei profondi cerebellari.Le vie in uscita dal cervelletto, come troverete su molti libri, in realtà non escono dal cervelletto, non ci sono delle vie efferenti “lunghe” che escono dall’insieme corteccia-nuclei cerebellari. Le vie in uscita dalle cellule del Purkinje (via efferente) proiettano sui nuclei cerebellari e da qua l’informazione verrà smistata altrove. L’effetto inibitorio che si ottiene decerebellando (togliendo la corteccia cerebellare) viene portato sui nuclei cerebellari profondi e da qui a varie zone, sia corticali superiori che spinali. Vediamo più nel dettaglio come si realizza questo circuito, con uno schema che riassume come sono posti fra loro i neuroni cerebellari (dei quali ricordiamo la tipizzazione cellulare fatta l’altra volta).
1

Distinguiamo tre strati: strato molecolare che presenta: cellule dei canestri; cellule stellate di tipo A e B (che qui non
sono indicate) che hanno attività uguale alle cellule dei canestri. Le cellule di questo strato hanno tutte, più o meno, la stessa funzione. Vi sono, inoltre, le fibre parallele.
strato delle cellule del Purkinje: qui ci sono le cellule del Purkinje, le più grandi cellule del SNC, con le loro efferenze sui nuclei centrali cerebellari.
strato dei granuli che presenta: cellula del granulo e cellula di Golgi.
Vediamo come sono messe le afferenze rispetto a queste cellule: fibra rampicante: arriva dall’oliva inferiore. Ha effetto eccitatorio sia sui nuclei centrali
cerebellari che direttamente sulla cellula del Purkinje. Ad ognuna di queste cellule arriva una sola fibra rampicante, contraendo una sinapsi eccitatoria. Il potenziale che si genera nella cellula del Purkinje, quando viene eccitata da un’informazione portata tramite la fibra rampicante, prende il nome di POTENZIALE COMPLESSO. L’eccitazione che arriva è molto forte, tale da indurre sempre eccitazione sulla fibra del Purkinje. Ogni volta che l’informazione entra tramite la fibra rampicante proveniente dal nucleo olivare inferiore porta sempre e comunque ad un’eccitazione e, quindi, ad una risposta della cellula del Purkinje. Questo potenziale si dice complesso perché si verifica in più fasi, è abbastanza lungo, porta sempre a eccitazione. È importante ricordarsi che la stimolazione è molto specifica: ho una cellula rampicante, una fibra del Purkinje, la risposta da quella cellula del Purkinje (disposizione 1:1).
fibre muscoidi: eccitano una cellula del granulo e, contemporaneamente, un dendrite della cellula di Golgi.
fibre parallele: assone della cellula del granulo che sale verso la corteccia, si infila nello strato del Purkinje, arriva allo strato molecolare dove la cellula si divide a T. La lunghezza di questa fibra è di circa 1,5 cm per lato. La cellula del granulo eccitata, porta la sua eccitazione sia sulla
2

cellula del Purkinje che sulle altre cellule che si trovano con il loro dendrite localizzato nello strato molecolare, cioè le cellule dei canestri e la cellula del Golgi. Effetto della fibra parallela sulla cellula del Purkinje : eccitatorio, anche se è uno stimolo
molto più disperso perché qui le fibre muscoidi contraggono sinapsi con molte cellule dei granuli nel formare il complesso del glomerulo. Le afferenze che arrivano dal granulo alla cellula del Purkinje, lungo le fibre parallele, non sono specifiche (come abbiamo visto per le fibre rampicanti), ma convogliano un tipo d’informazione più estesa, più integrata. Inoltre su ogni cellula del Purkinje non arriva un’unica fibra parallela, ma ne arrivano migliaia. Ognuna di queste porta un’eccitazione sulla fibra del Purkinje che causa un fenomeno detto POTENZIALE SEMPLICE. Non è detto che questo raggiunga la soglia: ci vogliono molte stimolazioni da parte di molte fibre parallele e quindi da molte cellule del granulo (integrazione ampia di molte informazioni afferenti) perché le fibre parallele portino a soglia la cellula del Purkinje e perché questa dia una risposta. Abbiamo un dualismo: informazioni specifiche dalle fibre rampicanti – informazioni più vaghe, di controllo, integrate tramite le fibre muscoidi.
Effetto della fibra parallela sulla cellula dei canestri e sulla cellula del Golgi : eccitatorio. Queste cellule, che vengono eccitate, sono inibitorie. Le cellule dei canestri, così come quelle stellate di tipo A e B, sono cellule che inibiscono la cellula del Purkinje. L’informazione delle fibre muscoidi può, forse, eccitare la cellula del Purkinje, se integrata con molte altre informazioni similari, ma nel contempo ha un’attività inibitoria sulla cellula stessa. C’è un doppio grado di controllo sulla risposta che la cellula del Purkinje dà all’afferenza che giunge tramite le fibre muscoidi.La cellula del Golgi inibisce la cellula del granulo.L’informazione che sale dalle fibre muscoidi, una volta che è passata, viene subito inibita. Questo sembra sia uno stratagemma della corteccia cerebellare per far sì che la cellula del Purkinje sia sempre pronta a ricevere informazioni nuove. Sembra che il cervelletto rimuova le informazioni ogni decimo di secondo, così da poter ricevere nuove informazioni per il controllo motorio in tempi molto rapidi. Questo è possibile grazie al circuito della cellula del Golgi, il quale serve sia a modulare che a preparare la cellula del Golgi ad un’ulteriore stimolazione.
L’integrazione delle informazioni si verifica a livello delle cellule del Purkinje, è l’unica strada di uscita, l’unica via efferente. Questa cellula ha un’attività inibitoria sui nuclei cerebellari profondi.Questo è il CIRCUITO CEREBELLARE, che serve per il controllo dei movimenti fini. Esso è integrato con circuiti molto più complessi, situati nei nuclei della base, che illustreremo brevemente (vedi dopo).
Visto che l’integrazione cerebellare si avvale, come unica via d’uscita, delle efferenze delle cellule del Purkinje sui nuclei centrali, vediamo dove finiscono queste fibre.
3

Ricordiamo che il nucleo dentato è uno dei principali nuclei cerebellari, inoltre abbiamo nuclei della reticolare mesencefalica e nel tronco dell’encefalo.Le efferenze che escono dal cervelletto possono andare verso l’alto ai centri sovrassiali superiori, tipicamente della corteccia. Questa è la via CEREBELLO-TALAMO-CORTICALE: partono proiezioni delle cellule del Purkinje che arrivano al nucleo dentato, quindi al peduncolo cerebellare superiore, al talamo, al nucleo ventrale laterale e anteriore e infine alla corteccia motoria.Una stazione importante della via è il nucleo rosso, punto di partenza della via RUBRO-SPINALE (extrapiramidale), che riceve componenti cerebellari per comporre le vie di controllo extrapiramidali. Dal nucleo rosso le proiezioni scenderanno.Un’altra componente importante è quella che parte dal paleocervelletto, va ai nuclei del tetto presenti nel tronco dell’encefalo (non sono cerebellari), giunge infine al midollo spinale. Il nucleo del tetto dà origine alla via extrapiramidale detta via TETTO-SPINALE. C’è un duplice controllo sulla zona spinale. Il nucleo del tetto riceve dal paleocervelletto ed e quindi particolarmente implicato in quei processi che controllano la postura, anche se in realtà l’organo principale per il controllo della postura è il vestibolo.
Proiezioni delle tre porzioni della corteccia cerebellare sui nuclei cerebellari: Paleocerebello: è la zona più interna e antica, quella del verme.
Con il tratto spino cerebellare arrivano le afferenze somatiche dalle parti corporee prossimali, tipicamente quelle che compongono anche l’asse corporeo, legato maggiormente alla postura. Queste arrivano al verme, che proietta in particolare al nucleo fastigiale (cerebellare profondo) e, con minore importanza al nucleo globoso. Qui si divide in due proiezioni: una passa dal mesencefalo, arriva al talamo e proietta alla corteccia cerebrale motoria (via cerebello-talamo-corticale); l’altra scende alla reticolare mesencefalica e arriva al nucleo vestibolare (tratti cortico-spinali ventrali, anch’essi vanno a modificare l’atteggiamento posturale a seguito della stimolazione dai tratti mediali prossimali).Schema:
parti corporee prossimalitratto spino-cerebellare
vermeNUCLEO FASTIGIALE
mesencefalo reticolare mesencefalicatalamo NUCLEO VESTIBOLAREcorteccia
4

Parte intermedia: zona più lateralizzata. Riceve proiezioni dagli arti, non più dalla parte mediale. Sono proiezioni di tipo discriminativo, anche sensoriali che riguarderanno in parte le afferenze tattili, ma soprattutto quelle propriocettive. Le proiezioni arrivano alla zona intermedia e possono poi avere delle efferenze che riguardano i nuclei globoso ed emboliforme, dai quali partono delle afferenze che di nuovo passano nel tronco encefalico, in particolare nel nucleo rosso (molto importante in queste proiezioni cerebello corticali), quindi abbiamo il passaggio al talamo e, infine, alla corteccia cerebellare.Ricordiamo che dal nucleo rosso origina la via rubro-spinale; dalla corteccia origina la via cortico-spinale, proiezione piramidale che si associa a quelle extrapiramidali. Per esempio, in questo caso, il tratto cotico-spinale laterale è ben distinto dalla via extrapiramidale del tratto rubro-spinale.Le vie piramidali sono quelle che originano dalla corteccia. Le vie extrapiramidali sono quelle che proiettano nelle stesse zone di proiezione del fascio cortico-spinale, cioè i motoneuroni ά e γ, ma portano l’integrazione di altre vie (vestibolari, cerebellari e dei nuclei della base).Schema:
artiNUCLEI GLOBOSO ed EMBOLIFORME
tronco encefalico, soprattutto al NUCLEO ROSSOtalamo
corteccia Neocerebello:
Non arrivano afferenze sensoriali dalla periferia, ma qualcosa che ha già subito un’elaborazione subcorticale da parte dei nuclei pontini, dopo che i nuclei pontini sono stati eccitati da una via che proviene dalla corteccia. L’emisfero laterale ha proiezioni specifiche al nucleo dentato, che raccoglie le efferenze delle cellule del Purkinje e del cerebrocerebello, quindi arriviamo al talamo e infine alla corteccia.Schema:
via corticaleNUCLEI PONTINI
NUCLEO DENTATOtalamo
corteccia
Funzioni delle tre porzioni della corteccia: Controllo del movimento fine. Sede di deposito della memoria motoria, del quale parleremo facendo i centri superiori
(linguaggio e memoria). Controllo del tono muscolare. Una lesione delle vie discendenti dal cervelletto può provocare un
ipertono.
N:B: le tre porzioni cerebellari proiettano su nuclei diversi ed hanno funzioni diverse.C’è una stretta associazione (scambio di informazioni) tra cervelletto, corteccia e periferia.
Oltre all’attività di corteccia motoria e cervelletto esistono altre due componenti importanti per il controllo motorio che sono i nuclei della base e il vestibolo, che hanno un significato completamente diverso.
5

NUCLEI DELLA BASE.
Sono nuclei profondi, intermedi alla capsula interna, a ridosso della scissura laterale (sono bilaterali). Costituiscono un importante punto di controllo del movimento, la loro lesione dà luogo a molte malattie e deficit motori gravi. Sono collegati strettamente all’amigdala, nucleo coinvolto negli aspetti emotivi, espressione delle attività superiori (sentimenti, sensazioni, emozioni). Molti tipi di lesioni dei nuclei della base, ad esempio il Parkinson, hanno un’importante componente affettiva perché i nuclei della base sono strettamente correlati alle proiezioni da e per l’amigdala.
Distribuzione dei nuclei della base: capsula internaNucleo caudato, putamen, globo pallido, sostanza nigra, nucleo subtalamico.
Sui libri di fisiologia troverete poco a riguardo dei nuclei della base perché sono strutture profonde e la loro interazione è estremamente complessa. Per questo si sa pochissimo di definitivo e la materia è in corso di studio.
6

I nuclei della base sono stati estratti dalla corteccia. Questa controlla il movimento tramite delle efferenze (vie piramidali, vie extrapiramidali che scendono al tronco dell’encefalo).Dalla corteccia partono efferenze che vanno ai nuclei della base, dai quali partono delle afferenze che entrano nella corteccia. Cercheremo di distinguere queste afferenze e il loro ruolo.I nuclei della base sono una stazione di circuitazione interna dei centri sovrassiali, per cui un’informazione che parte dalla corteccia ritorna alla corteccia. Non ci sono grosse efferenze dai nuclei della base. Tutto ciò che viene elaborato nei nuclei della base, ad eccezione di poche informazioni, rimane nella corteccia. Questi nuclei servono per modulare un’attività efferente della corteccia.La corteccia impartisce un ordine motorio, invia una seria di spike lungo la via piramidale, attiva i nuclei del tronco encefalico, attiva le vie extrapiramidali la muscolatura si contrae e il tono aumenta. Simultaneamente la corteccia informa i nuclei della base i quali rimodulano il segnale, passando dal talamo (che è sempre implicato nelle faccende che riguardano gli ingressi nella corteccia). In questo modo si riesce a controllare il segnale in uscita.Mentre nel caso del cervelletto il controllo si dedica a raffinare i movimenti fini, nel caso dei nuclei della base sembra che il controllo si dedichi a far sì che il movimento venga modificato durante l’esecuzione. Se i nuclei della base sono lesionati c’è un’incapacità nel correggere il movimento mentre lo si sta compiendo.Esempio: se stiamo giocando a tennis e ci sembra che la pallina abbia una certa traiettoria, iniziamo ad impostare la risposta in un certo modo. Se poi la pallina subisce una deviazione, immediatamente correggiamo la postura e la posizione del braccio per colpire bene la pallina. Questo non potremmo farlo se avessimo lesionato i nuclei della base. Non è un movimento fine, ma dobbiamo essere in grado di controllarlo e cambiarlo in itinere.
7

MACCANISMO:È necessario che i nuclei della base ricevano informazioni dall’encefalo su quello che la corteccia motoria ha ordinato.Dalla corteccia, non solo quella motoria, ma anche dalle zone limitrofe (aree associative e corteccia sensoriale), partono delle proiezioni che vanno allo striato ed hanno attività eccitatoria. Dalla corteccia motoria parte l’immagine speculare di ciò che essa manda ai centri sottostanti, ma ciò che succede in periferia (ad esempio la posizione del mio braccio) non può saperlo la corteccia motoria, lo saprà la corteccia sensoriale che riceve le afferenze sensitive. Per questo è importante ricordare che le proiezioni partono, oltre che dalla corteccia motoria, anche dalle zone vicine.Questi segnali vanno allo striato che il punto di raccolta dei segnali provenienti dalla corteccia. Lo striato proietta al talamo. Il talamo proietta alla corteccia prefrontale che informa lo striato.Quindi, nel contempo, lo striato è informato:
di come il movimento è stato pianificato. di come il movimento è stato realizzato. del fatto che il movimento sia stato realizzato nelle maniera opportuna oppure no.
La via che parte dallo striato e va verso il talamo può seguire due strade: via diretta: determina una inibizione sul globo pallido (sia sul nucleo esterno che su quello
interno), portata mediante il GABA. L’inibizione, quindi, passa al talamo.Si parla di via strio-pallidale diretta. È il primo contingente della via cortico-strio-pallido-talamo-corticale.Questa via INIBISCE.
Via indiretta: è un altro contingente della via cortico-stri-pallido-talamo-corticale. Questa non solo passa dal globo pallido esterno, ma, tramite un circuito retrogrado, fa in modo che l’inibizione portata sul nucleo pallido esterno, determini poi un’inibizione del nucleo subtalamico. Quest’ultimo proietta con un’attività eccitatoria, per cui l’inibizione dell’esterno induce un’inibizione dell’inibizione che produce un’eccitazione.Questa via ECCITA.
Alla fine la via cortico-strio-pallido-talamo-corticale finisce sui neuroni corticali, sui quali ha un’attività differente a seconda che faccia parte del contingente della via strio pallidale diretta oppure di quello della via indiretta: la via diretta ha un’attività ECCITATORIA SUI NEURONI CORTICALI.
La via è eccitatatoria fino al globo pallido, la proiezione al talamo è inibitoria, quindi abbiamo l’inibizione dell’inibizione = eccitazione che viene mandata ai neuroni corticali. Se determiniamo un’eccitazione dei neuroni corticali (che sono inibitori sul midollo spinale), abbiamo un aumento dell’attività INIBITORIA SUI MOTONEURONI. Quindi un’eccitazione della via diretta produce una diminuzione del tono muscolare (se decortichiamo, cioè rendiamo impossibile l’attività inibitoria dei neuroni corticali sul midollo spinale, abbiamo un aumento del tono muscolare).
la via indiretta ha un’attività INIBITORIA SUI NEURONI CORTICALI.La via, grazie al circuito retrogrado, inibisce la componente in uscita dal globo pallido interno. Avremo una componente inibitoria, perché eccita un’inibizione, sui neuroni corticali; la componente sarà ECCITATORIA SUI MOTONEURONI.
A seconda che l’informazione passi attraverso la via diretta o indiretta, la componente motrice corticale potrà essere un po’ più eccitata o inibita. In questo modo si realizza il controllo, che comunque non è completo perché mancano quelle vie che sono in uscita e che non ritornano alla corteccia (la via qui non è indicata perché la figura vuole solamente evidenziare l’aspetto di circuitazione interna tra corteccia-nuclei della base-talamo-corteccia). Queste vie “scappano fuori” dal globo pallido e dalla sostanza nigra formando la via cortico-strio-pallido-mesencefalo-spinale. È una proiezione discendente che ha delle stazioni ai nuclei mesencefalici, gli stessi che appartengono alle vie extrapiramidali e che in questo modo
8

regolano non solo l’attività dell’encefalo (via piramidale), ma anche quella dei nuclei sottocerebrali del tronco encefalico che sono coinvolti nel controllo motorio (vie extrapiramidali).La sostanza nigra è una componente particolare dei nuclei della base. Da essa esce un contingente della via cortico-strio-pallido-mesencefalo-spinale; inoltre riceve proiezioni inibitorie dallo striato (viene quindi informata dell’attività della corteccia motoria) e poi fornisce un circuito riverberante che è eccitatorio o inibitorio sullo striato. Questo dipende dal fatto che la sostanza nigra è formata da vari stipiti cellulari raggruppati in:
pars compacta: produttori di DOPAMINA, che ha attività eccitatoria sullo striato. pars reticolata: produttori di GABA, che ha attività inibitoria sullo striato.
Non diciamo qual è al netto l’effetto della sostanza nigra sullo striato e quindi sull’attività motoria, ma sappiamo che induce una modificazione può essere sia di tipo eccitatorio che di tipo inibitorio sullo striato. La via conduce anche una mediazione allo striato in risposta agli stati emozionali, in quanto la sostanza nigra riceve afferenze dall’amigdala. Essa può influire sul comportamento motorio modificando le informazioni che arrivano allo striato proprio mediante questa proiezione che parte dalla sostanza nigra, detta proiezione nigro-striatale. Nel controllo motorio operato dai gangli della base interviene anche un controllo motorio di tipo emozionale.
Fenomeni dovuti a lesioni dei nuclei della base (alcuni esempi): Corea di Huntington TesosiPatologie legate a disturbi della motilità, in particolare ipotonia e ipercinesia (nel senso di incapacità di mantenere una posizione stabile, che quindi provoca movimenti incontrollati spesso associati a tremore). Morbo di Parkinson. Determina ipertonia, ipocinesia e tremore.Queste patologie sono spesso associate a disturbi mentali nella fase più tardiva perché il riverbero dei nuclei della base su tutta la corteccia è molto esteso e, soprattutto, c’è una compromissione emozionale da parte dell’amigdala. Lesioni dei nuclei della base, quindi, comportano una perdita del controllo motorio e della capacità di mantenere un corretto tono muscolare.
Porzione della corteccia interessata all’interazione con i nuclei della base e con il cervelletto:
9

Cervelletto: interessa una porzione abbastanza limitata della corteccia, cioè quella motoria primaria e quella sensoriale.Dai nuclei pontini le informazioni arrivano al cervelletto, passano attraverso i nuclei ventro-laterali del talamo, quindi tornano in corteccia. Inoltre il cervelletto riceve informazioni dal midollo e da tronco encefalico.
Nuclei della base: è coinvolta praticamente tutta la corteccia in diverso grado. Tutte le proiezioni corticali arrivano ai gangli della base, da qui si portano ai nuclei ventrali anteriori e laterali del talamo.
Incrocio del controllo motorio afferente ed efferente mediato da area motoria primaria, cervelletto e nuclei della base:
Dal muscolo parte un’informazione che sale a vari livelli (cervelletto, formazione reticolare). I muscoli ricevono una efferenza che parte dalla formazione reticolare del tronco encefalico e che fa parte delle vie extrapiramidali ultime.Come sono integrate le attività del cervelletto, dell’area motoria e i nuclei della base sul controllo dei motoneuroni ά e γ (cioè del muscolo)?Il cervelletto riceve informazioni. Dal cervelletto parte un’efferenza che passa da talamo e arriva al nucleo caudato (controllo diretto sul nucleo caudato da parte delle informazioni che arrivano dal cervelletto). Quindi i nuclei della base, non solo di quanto succede in uscita dalla corteccia, ma anche di quanto è successo in uscita dal cervelletto.Il nucleo caudato riceve informazioni dal cervelletto, dalla corteccia motoria, dalle aree motorie. Queste informazioni, direttamente o passando attraverso il putamen, vengono convogliate verso la
10

via d’uscita che è rappresentata dal globo pallido e, soprattutto, dalla sostanza nigra (che è uno dei punti di partenza della via discendente verso il tronco encefalico). La sostanza nigra riceve informazioni dal globo pallido e dalla corteccia motoria. Proietta verso la formazione reticolare, quindi verso i muscoli.Il nucleo rosso riceve dal cervelletto e proietta alla formazione reticolare.Dall’oliva inferiore arrivano le fibre rampicanti, quelle la cui eccitazione determina un’attivazione specifica e univoca delle cellule del Purkinje. Riceve dalla corteccia motoria, dal muscolo, dai nuclei della base.
RICORDA: Vie piramidali: output che va direttamente dalla corteccia ai motoneuroni. Questa via non è
soggetta ad un controllo, se non quando arriva sul motoneurone. Ciò che permette il controllo del movimento e fa sì che possiamo modificare, raffinare,
correggere il movimento volontario è il contemporaneo arrivo, sugli stessi motoneuroni, delle componenti extrapiramidali che correggono il messaggio iniziale dopo averlo integrato con cervelletto e nuclei della base. Tutto questo comprende anche aspetti di tipo affettivo provenienti dall’amigdala.
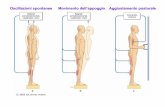
Controllo della postura.Il controllo della postura è un aspetto importante del controllo motorio perché integra informazioni che ci dicono qual è la “posizione dell’organismo nello spazio”, anche se, in realtà, questa dicitura non è corretta. Il controllo della postura mantiene la postura avvalendosi di tre fondamentali tipi di informazioni: visive. che arrivano dai fusi neuromuscolari e dai propriocettori articolari del collo. che arrivano dal vestibolo (organo preposto alla valutazione di questo aspetto, localizzato
nell’orecchio interno).
Il meccanismo di controllo della postura consente di controllare la posizione dell’organismo rispetto alla direttrice che passa per il baricentro ed è diretto secondo la forza gravitazionale (cioè verso il basso).
Noi possiamo spostare l’asse del capo rispetto alla direttrice oppure spostare tutto l’asse del capo e del tronco rispetto alla direttrice. In entrambe i casi verrà attivato il sistema di controllo della postura che è quello che ci consente di mantenere la posizione del capo rispetto alla direttrice; che il capo sia spostato insieme ad un altro segmento corporeo è un dettaglio. È importante considerare la posizione del capo rispetto alla direttrice perché ci siamo evoluti in un campo gravitazionale ed abbiamo sviluppato sistemi per avvertire questo campo gravitazionale e per modificare la nostra posizione. Se mi inclino sempre di più rispetto alla direttrice, dopo un po’ il
11

centro di massa del baricentro cadrebbe al di fuori della base dei piedi e io cadrei per terra. Per mantenere la postura occorre che il baricentro rimanga entro il perimetro segnato dalla base di appoggio.
Il controllo della postura viene mantenuto grazie alla contrazione dei muscoli estensori del dorso, del collo, degli arti inferiori e dalla muscolatura addominale. Questi gruppi muscolari hanno un tono muscolare che consente al soggetto di mantenere una certa posizione, una modifica di questo tono muscolare consente di modificare la postura.Il controllo della postura prevede che venga registrata la posizione della testa rispetto all’asse con tre sistemi recettoriali specifici: componente visiva, spinale e vestibolare.Queste componenti mandano le loro afferenze ai centri sovrassiali i quali risponderanno modificando, con le metodiche già viste, il tono muscolare. Basta modificare l’output motorio (ad opera dei nuclei della base del cervelletto, del controllo extrapiramidale operato dai nuclei vestibolari) per modificare lo stato di contrazione dei muscoli estensori e quindi la postura. Le modifiche della postura ci consentono non solo di non cadere, ma anche di assumere atteggiamenti posturali automatici (in discesa portiamo il busto indietro, in salita lo portiamo avanti).
Componente spinale: REAZIONI IMMEDIATE. Reazione di piazzamento. Riflesso di stiramento. Riflessi tonici del collo. Riflesso estensorio crociato.
Esempio: se sto parlando con un amico e mi capita di inciampare, non cado, ma avverto la presenza di un ostacolo e modifico la postura in modo da allungare il passo e superare l’ostacolo. Questo è il riflesso di piazzamento che permette di correggere automaticamente il movimento in modo da superare un ostacolo. Poi ci sono i riflessi da stiramento; riflessi tonici del collo per i quali la tonicità e quindi la posizione del collo rispetto al busto viene modificata; riflesso estensorio crociato che mi permette di sostenere un attimo in più il peso del corpo sulla sola gamba destra, permettendomi di superare l’ostacolo e di rimanere in piedi mentre c’è una reazione di piazzamento. Sono tutti riflessi che si basano sul circuito spinale arco riflesso che abbiamo visto, sul riflesso monosinaptico o sul riflesso estensorio crociato.
Componente visiva: FACILITAZIONE delle AFFERENZE SOMATOSENSORIALI SPINALI.un soggetto a cui viene rimossa la componente visiva ha più difficoltà nel mantenimento della postura, non solo nella deambulazione.
Componente vestibolare: COMPENSAZIONE LENTA.La sua risposta riguarda un interessamento molto esteso della muscolatura.
Per il mantenimento della postura la componente più importante è quella spinale, seguita dalla vestibolare e infine da quella visiva.
VESTIBOLO.L’orecchio interno, grazie ad un particolare sistema recettoriale, ci permette di percepire le onde pressorie causate dal suono. Però c’è un’altra componente dell’orecchio interno che non ha significato di percezione dei suoni, questa componente è il vestibolo.Il vestibolo ci consente di percepire due grandezze fisiche: l’accelerazione lineare e quella angolare (si può quindi dire che il vestibolo è un accelerometro).A questo proposito riconosciamo due tipi di strutture. Il sacculo e l’utricolo che sono due zone nelle quali abbiamo il sistema recettoriale per la percezione dell’accelerazione lineare, in particolare questi sistemi sono soggetti all’accelerazione gravitazionale. L’altra struttura importante è quella rappresentata dai tre canali semicircolari che sono cavità scavate in un’intercapedine ossea, cioè
12

intorno c’è una struttura rigida dentro la quale c’è l’endolinfa. I tre canali semicircolari sono messi in modo da valutare una variazione di accelerazione angolare nelle tre dimensioni dello spazio. Essi sono posti tra loro con un’angolatura di circa 90° (in pratica nelle tre dimensioni spaziali); uno in proiezione orizzontale, con un’angolatura di circa 30° rispetto alla proiezione antero-posteriore, uno a 90° rispetto al piano orizzontale e in posizione antero-posteriore, uno in posizione verticale.
Il vestibolo è differenziato per rilevare accelerazioni angolari nei tre canali semicircolari e accelerazioni lineari nel sacculo e nell’utricolo.In entrambe le strutture ci sono cellule ciliate con un funzionamento assolutamente simile a quello che abbiamo visto per le cellule cigliate dell’organo del Corti, sfruttano le stesse caratteristiche dell’endolinfa sfruttate dall’organo del Corti per percepire i suoni. La differenza consiste nel fatto che nell’organo del Corti l’attivazione delle cellule cigliate è legata alle onde pressorie a cui è soggetta la membrana basilare, invece in questo caso è legata a movimenti delle cellule cigliate dovuti, nel caso di sacculo e otricolo, allo spostamento degli otoliti, nel caso dei canali semicircolari, allo spostamento dell’endolinfa. Nel sacculo e nell’utricolo le cellule cigliate sono sovrastate da uno strato gelatinoso in cui ci sono delle formazioni (otoliti) con densità superiore allo strato gelatinoso, sono come dei sassolini di carbonato di calcio che possono muoversi in una direzione piuttosto che in un’altra, sollecitati dallo spostamento. Per esempio se sposto la testa, gli otoliti si spostano di conseguenza e peseranno deformando le ciglia da un lato piuttosto che dall’altro a seconda di come viene spostato il capo. La risultante della forza di questo spostamento, della forza peso degli otoliti sulle cellule ciliate, è l’elemento che costituisce l’attivazione delle cellule sensoriali e dà una risposta che viene interpretata come variazione di spostamento. In realtà è una variazione di una componente di forza risultante dall’accelerazione lineare. Sappiamo che F = ma m: è quella degli otoliti, a: è quella gravitazionale, F: ha una direzione d’applicazione perché è una grandezza vettoriale. Se sposto la testa, l’accelerazione rimane la stessa, ma F avrà una grandezza diversa rispetto alla cellula cigliata perché è variato il vettore forza a seconda della direzione dello spostamento rispetto alla direttrice.I canali semicircolari sono riempiti di endolinfa, però nell’ampolla c’è la cresta ampollare, una formazione in cui ci sono cellule di sostegno e cellule cigliate le cui ciglia protrudono in una massa gelatinosa che prende forma di una cupola, questa volta senza otoliti. Questa cupola, con inserite le ciglia, potrà essere spostata da una parte o dall’altra a seconda del movimento dell’endolinfa nel canale semicircolare. Lo spostamento delle ciglia, a seconda della direzione dello spostamento, sarà il segnale di stimolazione per i recettori.
Le cellule cigliate non sono di per sé cellule nervose, ma cellule recettrici che sono in comunicazione con una via afferente. La porzione apicale, oltre a possedere le ciglia, presenta
13

anche il chinociglio. Questo vale per macula e utricolo che sopra hanno la componente gelatinosa con gli otoliti, ma vale anche per l’ampolla con la sua cupola gelatinosa. Le cellule cigliate dell’utricolo sono poste tutte nella stessa direzione, così anche quelle dell’ampolla che hanno una posizione particolare e sono tutte rivolte verso l’utricolo.C’è una distinzione tra chinociglio e ciglia. A seconda che le ciglia vengano avvicinate o allontanate dal chinociglio, avremo rispettivamente una depolarizzazione o una iperpolarizzazione. E’ lo stesso meccanismo visto per le cellule cigliate della coclea a riguardo del potassio (che si comporta un po’ come il sodio in questo caso) che invece di uscire normalmente dalla cellula, entra a causa dell’elevata concentrazione di potassio nell’endolinfa. Se porto le ciglia verso il chinociglio si assiste ad una depolarizzazione, se le allontano c’è una iperpolarizzazione; il fenomeno è di tipo oscillatorio se il movimento è protratto e rimane se sopprimo il chinociglio.
L’aumento dell’attività della cellula cigliata, quando si depolarizza, determina un aumento della frequenza di eccitazione. Questo fa parte della risposta di ogni singolo recettore che tanto più viene stimolato, tanto più aumenterà la frequenza. Se sposto le ciglia verso il chinociglio ho una depolarizzazione che viene mantenuta e quindi la frequenza di scarica viene mantenuta lungo tutta la durata dello spostamento. Questo vuol dire che questi sono recettori che non si adattano. Se rimango per 40 minuti con la testa reclinata, i recettori continuano ad accorgersene e continuano a scaricare informando sempre il SNC del fatto che ho la testa reclinata. Questo è un comportamento abbastanza atipico perché la maggior parte dei recettori si assiste, dopo un’iniziale eccitazione, ad una adattamento del recettore più o meno lento o marcato. Normalmente il recettore tende ad adattarsi, mentre in questo caso non ci sono segni di adattamento perché è importante essere costantemente informati sulla posizione di questi recettori.
14

Qui abbiamo la macula a riposo, con la testa messa in posizione lineare rispetto alla componente gravitazionale e con sopra gli otoliti. Se reclino la testa da un lato o dall’altro è chiaro che gli otoliti si sposteranno di conseguenza rispetto al chinociglio. Gli otoliti si trascinano dietro la massa gelatinosa: in un caso le ciglia saranno spostate verso il chinociglio e ci sarà una depolarizzazione, nell’altro saranno allontanate dal chinociglio e ci sarà un’iperpolarizzazione.
Guardando la cellula cigliata dall’alto vediamo il chinociglio con un pool di ciglia. Il grafico dice che se sposto la testa in direzione B o in direzione A, avvicino o allontano le ciglia dal chinociglio. Se sposto la testa in direzione C, non ho attività. Quindi ci sarà un pool di cellule orientate nelle varie direzioni dello spazio in modo da assicurare che qualsiasi movimento del capo venga recepito, sia in un senso che nell’altro. Altrimenti succederebbe che se io muovessi la testa a destra e a sinistra il sistema vestibolare se ne accorgerebbe, se la muovessi avanti e indietro non se ne accorgerebbe. Questo non succede grazie al differente orientamento delle cellule nella macula e nell’utricolo.
15

Vediamo in particolare le figure disegnate dentro la testa. Stiamo parlando di macula e otricolo (rilevatori dell’accelerazione lineare). Se ruotiamo la testa gli otoliti si spostano di lato, ma abbiamo lo stesso movimento degli otoliti anche se spostiamo la testa unitamente al colo e alla prima porzione del tronco. Quello che importa non è l’angolo tra testa e collo, ma quello tra testa e direzione di applicazione della forza gravitazionale.
Cerchiamo ora di capire cosa succede nei canali semicircolari contenenti endolinfa. Ricordiamo che nell’ampolla c’è la cresta ampollare (cupola di gel) con dentro le propaggini delle cellule ciliate.La cresta ampollare non si muove quando la testa è soggetta ad una accelerazione lineare che non intacca i recettori dei canali semicircolari. Se cammino in avanti i sensori del vestibolo in questione non se ne accorgono, rimangono fermi. Ma se muovo la testa in un senso o nell’altro i recettori vestibolari se ne accorgono, così come se mi mettono su una sedia e mi fanno girare. Questo è un metodo per sballare il sistema vestibolare, la quale sono sottoposti gli astronauti (centrifughe).Consideriamo l’ampolla in posizione normale, circondata da endolinfa. Se faccio ruotare il disegno nella direzione della forza esterna, la parete esterna del canale semicircolare, che è ossea, si gira solidalmente al movimento che compio. Ma l’endolinfa che c’è dentro è un liquido viscoso soggetto alla forza inerziale, quindi gira dopo rispetto alla parete ossea, all’inizio del movimento l’endolinfa rimane ferma. Quindi c’è una dissociazione tra il movimento della testa e quello dell’endolinfa per cui l’ampolla, che è di gel, risente di questo ritardo e viene deflessa. È come se il liquido, invece di seguire il movimento dell’intercapedine ossea, tornasse indietro, deflettendo la cupola che è associata al movimento del liquido. La cresta ampollare si deforma nella direzione opposta al movimento, rimane indietro, e altera la posizione delle ciglia spostandole rispetto al chiniciglio.
F = K δ s + µ v + m a K: coefficiente di elasticità µ: viscosità dell’endolinfaδ: coefficiente di frizione µ direttamente proporzionale a F
La forza con cui la cresta ampollare viene deflessa ha varie componenti. Una legata allo spostamento (s), una legata alla velocità (v), una è la componente inerziale legata all’accelerazione angolare a cui il liquido viene sottoposto (a) e alla massa dell’endolinfa (m).Sottoposta a questa forza risultante, l’ampolla si deflette e, insieme a questa, anche le ciglia rispetto al chinociglio.
16

In clinica, per valutare la funzionalità del sistema vestibolare, si induce un’eccitazione dei recettori vestibolari, non con lo spostamento, ma registrando i potenziali evocati, cioè dei potenziali che si possono registrare dall’esterno in risposta all’eccitazione di un gruppo di neuroni, ad esempio di quelli vestibolari. Per simulare il movimento dei canali semicircolari, si riscalda l’orecchio esterno con acqua calda, in questo modo si riscalda un po’ l’endolinfa del canale. Come tutti i fluidi, anche l’endolinfa, quando viene scaldata, diminuisce la sua densità e tende ad andare verso l’alto. Questa tendenza delle porzioni più calde dell’endolinfa, rispetto a quelle più distanti dalla sorgente di riscaldamento, genera un ricircolo dell’endolinfa che non è legato al movimento del capo, ma permette comunque di vedere se al movimento dell’endolinfa corrisponde una risposta dei recettori vestibolari.Se si fa la prova ruotando veramente il capo, la cellula cigliata del canale semicircolare ha una sua frequenza di scarica tonale di base (basale). Se ruotiamo il capo da un lato la scarica aumenta (aumenta lo stato di eccitazione), quando terminiamo la rotazione anche la cellula smette di ruotare, torna al suo valore basale e si inibisce (otterremo l’effetto opposto).
A differenza dei recettori della macula, i recettori dei canali semicircolari sono di tipo FASICO, cioè percepiscono la variazione di accelerazione angolare. Infatti scaricano quando si inizia la rotazione,poi si zittiscono; terminano di scaricare e vengono inibiti quando si arresta la rotazione, che è come stimolare il recettore con accelerazione opposta.I recettori di macula e utricolo sono di tipo tonico (scaricano sempre quando viene presentato lo stimolo, cioè la variazione di accelerazione lineare), quelli dei canali semicircolari sono di tipo fasico (scaricano solo quando lo stimolo viene dato o tolto).
Per la loro struttura, i canali semicircolari rispondono alle accelerazioni angolari, non a quelle lineari.Risposta del canale semicircolare ad un movimento breve (rapido) della testa:
17

La cupola al variare della velocità angolare, la capsula con sopra gli otoliti si sposta e l’aumento della scarica (fasica) persiste durante tutto l’aumento della velocità angolare.Risposta del canale semicircolare ad un movimento protratto (lento) della testa:
Il movimento protratto è quello di una accelerazione angolare che aumenta, con una velocità che viene mantenuta protratta. Esempio: siamo su legati su una sedia che ruota su se stessa. All’inizio l’accelerazione aumenta per dare origine alla rotazione, poi la velocità di rotazione della sedia viene mantenuta costante e quindi l’accelerazione è zero (a=0). La cupola ha uno spostamento inizialmente marcato, poi la sua deformazione cala, ma si mantiene un po’ superiore rispetto al livello basale. In questo caso scarica molto all’inizio, rimane un po’ più alta durante la scarica basale. Quando cesso il movimento, cioè ritorno ad una velocità uguale a zero, l’accelerazione angolare varia in senso opposto e quindi c’è un’inibizione.
18