
Capitolo_10
11
1 Attivazione delle cellule B e produzione di anticorpi 1.1 Caratteristiche generali della risposta umorale • Il processo di attivazione delle cellule B è sequenziale. I linfociti B maturi e in grado di rispondere agli antigeni si sviluppano da precursori midollari prima di incontrare l’antigene e popolano poi i tissuti linfoidi periferici, sede della futura interazione antigenica. Gli antigeni legano le IgM e IgD di membrana sui linfociti naive e li attivano; l’attivazione può avvenire sia in dipendenza che in indipendenza dai linfociti T. L’attivazione può portare a proliferazione, cioè all’espansione del clone antigene specifico, e a differenziazione, cioè alla generazione di plasmacellule e cellule della memoria. Alcune cellule B attivate iniziano a produrre anticorpi diversi da IgM e IgD in un fenomeno detto switching (switching dell’isotipo della catena pesante). I linfociti in grado di produrre anticorpi con la più alta affinità per l’antigene vengono inoltre preferenzialmente espansi: si parla di maturazione dell’affinità. Un singolo linfocita può, in sette giorni, dare origine a quattromila plasmacellule che producono oltre 10 12 anticorpi al giorno. • Le risposte anticorpali agli antigeni proteici richiedono i linfociti T helper CD4 + che riconoscono gli antigeni e hanno ruolo fondamentale nell’attivazione dei B. Per questo motivo le proteine sono classificate come antigeni timo dipendenti. • Le risposte anticorpali agli antigeni multivalenti, con epitopi polisaccaridici e lipidici, non richiedono i linfociti helper. Per questo motivo gli antigeni polisaccaridici e lipidici sono definiti timo indipen- denti. • I linfociti attivati differenziano in plasmacellule, alcune delle quali continuano a produrre an- ticorpi per anni, e in cellule della memoria. Le risposte umorali originano agli organi linfoidi periferici, ma alcune plasmacellule migrano da questi al midollo osseo dove si stabiliscono per anni producendo bassi livelli di anticorpi che forniscono protezione immediata per i microbi da essi riconosciuti. • Lo switching degli isotipi e la maturazione dell’affinità sono tipici delle risposte T-dipendenti agli antigeni proteici. Lo switching è stimolato direttamente dai segnali in arrivo dalle cellule T, tra i quali la molecola CD40L e varie citochine. La maturazione riguarda la generazione di mutazioni somatiche ad alta frequenza in geni Ig V riarrangiati e la consequente selezione delle cellule B con grande affinità per l’antigene originale. La natura della risposta umorale varia inoltre in funzione del distretto anatomico: ad esempio i tessuti linfoidi mucosali sono adattati a produrre grandi quantità di IgA. • Le risposte anticorpali primarie e secondarie differiscono quantitativamente e qualitativamente. Le risposte primarie derivano dall’attivazione di cellule B naive, le secondarie dalla stimolazione di cloni espansi delle cellule della memoria; per questo motivo le risposte secondarie sono più rapide e legate a quantità maggiori di anticorpi. In aggiunta a questo sia lo switching che la maturazione aumentano con ripetute esposizioni allo stesso antigene. • Set differenti di linfociti rispondono preferenzialmente a diverse tipologie di antigene. Le cellule B follicolari degli organi linfoidi periferici preferiscono gli antigeni proteici; le cellule della zona marginale della milza riconoscono antigeni multivalenti. 1.2 Riconoscimento dell’antigene e attivazione antigene-indotta I linfociti circolano attraverso i follicoli degli organi linfoidi periferici in cerca del loro antigene. L’in- gresso nei follicoli è guidato dalla chemochina CXCL13 prodotta dalle cellule dendritiche follicolari e da quelle stromali; questa molecola si lega al recettore CXCR5 e attrae i linfociti nella giusta sede. La sopravvivenza dei linfociti follicolari dipende dai segnali in arrivo dal B cell receptor (BCR) ma anche da quelli mediati da una citochina detta BAFF (appartenente alla famiglia del TNF); BAFF e il ligando correlato, APRIL, possono attivare altri due recettori, TACI e BCMA, che hanno ruolo nelle fasi più tardive della maturazione. Gli antigeni entrano negli organi linfoidi tramite le APC o in forma solubile e attivano i linfociti grazie all’interazione con il BCR. Questo recettore ha due ruoli nella fase di attivazione: 1
-
Upload
sebastiano -
Category
Documents
-
view
530 -
download
2
Transcript of Capitolo_10

1 Attivazione delle cellule B e produzione di anticorpi
1.1 Caratteristiche generali della risposta umorale
• Il processo di attivazione delle cellule B è sequenziale. I linfociti B maturi e in grado di rispondereagli antigeni si sviluppano da precursori midollari prima di incontrare l’antigene e popolano poii tissuti linfoidi periferici, sede della futura interazione antigenica. Gli antigeni legano le IgMe IgD di membrana sui linfociti naive e li attivano; l’attivazione può avvenire sia in dipendenzache in indipendenza dai linfociti T. L’attivazione può portare a proliferazione, cioè all’espansionedel clone antigene specifico, e a differenziazione, cioè alla generazione di plasmacellule e celluledella memoria. Alcune cellule B attivate iniziano a produrre anticorpi diversi da IgM e IgD inun fenomeno detto switching (switching dell’isotipo della catena pesante). I linfociti in grado diprodurre anticorpi con la più alta affinità per l’antigene vengono inoltre preferenzialmente espansi:si parla di maturazione dell’affinità. Un singolo linfocita può, in sette giorni, dare origine aquattromila plasmacellule che producono oltre 1012 anticorpi al giorno.
• Le risposte anticorpali agli antigeni proteici richiedono i linfociti T helper CD4+ che riconosconogli antigeni e hanno ruolo fondamentale nell’attivazione dei B. Per questo motivo le proteine sonoclassificate come antigeni timo dipendenti.
• Le risposte anticorpali agli antigeni multivalenti, con epitopi polisaccaridici e lipidici, non richiedonoi linfociti helper. Per questo motivo gli antigeni polisaccaridici e lipidici sono definiti timo indipen-denti.
• I linfociti attivati differenziano in plasmacellule, alcune delle quali continuano a produrre an-ticorpi per anni, e in cellule della memoria. Le risposte umorali originano agli organi linfoidiperiferici, ma alcune plasmacellule migrano da questi al midollo osseo dove si stabiliscono peranni producendo bassi livelli di anticorpi che forniscono protezione immediata per i microbi daessi riconosciuti.
• Lo switching degli isotipi e la maturazione dell’affinità sono tipici delle risposte T-dipendenti agliantigeni proteici. Lo switching è stimolato direttamente dai segnali in arrivo dalle cellule T, tra iquali la molecola CD40L e varie citochine. La maturazione riguarda la generazione di mutazionisomatiche ad alta frequenza in geni Ig V riarrangiati e la consequente selezione delle celluleB con grande affinità per l’antigene originale. La natura della risposta umorale varia inoltre infunzione del distretto anatomico: ad esempio i tessuti linfoidi mucosali sono adattati a produrregrandi quantità di IgA.
• Le risposte anticorpali primarie e secondarie differiscono quantitativamente e qualitativamente.Le risposte primarie derivano dall’attivazione di cellule B naive, le secondarie dalla stimolazione dicloni espansi delle cellule della memoria; per questo motivo le risposte secondarie sono più rapidee legate a quantità maggiori di anticorpi. In aggiunta a questo sia lo switching che la maturazioneaumentano con ripetute esposizioni allo stesso antigene.
• Set differenti di linfociti rispondono preferenzialmente a diverse tipologie di antigene. Le celluleB follicolari degli organi linfoidi periferici preferiscono gli antigeni proteici; le cellule della zonamarginale della milza riconoscono antigeni multivalenti.
1.2 Riconoscimento dell’antigene e attivazione antigene-indotta
I linfociti circolano attraverso i follicoli degli organi linfoidi periferici in cerca del loro antigene. L’in-gresso nei follicoli è guidato dalla chemochina CXCL13 prodotta dalle cellule dendritiche follicolari eda quelle stromali; questa molecola si lega al recettore CXCR5 e attrae i linfociti nella giusta sede. Lasopravvivenza dei linfociti follicolari dipende dai segnali in arrivo dal B cell receptor (BCR) ma ancheda quelli mediati da una citochina detta BAFF (appartenente alla famiglia del TNF); BAFF e il ligandocorrelato, APRIL, possono attivare altri due recettori, TACI e BCMA, che hanno ruolo nelle fasi piùtardive della maturazione.
Gli antigeni entrano negli organi linfoidi tramite le APC o in forma solubile e attivano i linfociti grazieall’interazione con il BCR. Questo recettore ha due ruoli nella fase di attivazione:
1

1. L’accumulo di recettori antigene-indotto rende possibile la segnalazione biochimica
2. Il recettore lega e internalizza l’antigene per processarlo in peptidi per la presentazione ai linfocitiT helper
1.2.1 Trasduzione del segnale
I recettori dei linfociti B naive, cioè IgM e IgD, hanno code citoplasmatiche troppo corte per trasdurreil segnale, il compito viene infatti svolto da altre due molecole dette Igα e Igβ. Queste molecole sonotra loro legate da ponti disolfuro e sono associate in modo non covalente alla membrana; sono ancherichieste per l’espressione superficiale delle molecole Ig e insieme ad esse formano il complesso recet-toriale delle cellule B (complesso BCR). Igα e Igβ sono dunque analoghe a CD3 e ζ per i linfociti T. Idomini citoplasmatici di queste due molecole contengono i motivi ITAM già visti per le cellule T e sonoanche blandamente associati a tirosin chinasi della famiglia Src.
Il cross-linking delle Ig di membrana porta le chinasi citoplasmatiche ad avvicinarsi e questo le attivafacendo loro fosforilare i domini ITAM. La fosforilazione di ITAM fornisce un sito di attacco per i dominiSH2 della tirosin chinasi Syk, l’equivalente nelle cellule B di ZAP-70 dei linfociti T. Syk attivata va afosforilare dei residui di tirosina su una proteina adattatrice detta SLP-65 facilitando il reclutamentosu questa di altri domini SH2 di vari enzimi. Le principali molecole che interagiscono con SLP-65 sono:
1. Il fattore SOS viene reclutato da SLP-65 e catalizza la sostituzione di GDP in GTP sulla proteinaRAS e sulla proteina RAC. Queste proteine in forma GTP-legata attivano la via della chinasiJNK-MAP.
2. Una fosfolipasi, PLCγ2, viene attivata quando si lega a SLP-65 e viene fosforilata da Syk e Btk.Questo enzima si porta a demolire il fosfatidilinositolo di membrana (PIP2) generando inositolo3-fosfato e diacilglicerolo. L’inositolo mobilita il calcio, il DAG in presenza di calcio attiva laprotein chinasi C che fosforila varie altre proteine.
3. La protein chinasi C fosforila una proteina detta CARMA1 contentente un dominio CARD che nemedia le interazioni con le altre proteine. Le attività di CARMA1 culminano infine con l’attivazionedel complesso IKK (IκB chinasi); il complesso è critico per l’attivazione di NF-κB in quanto è ingrado di fosforilare IκBα, un inibitore di NF-κB, e destinarlo al proteasoma: in questo modo NF-κBè libero di entrare nel nucleo.
4. Questa serie di cascate porta all’attivazione di fattori di trascrizione che inducono l’espressione digeni i cui prodotti sono richiesti per le risposte delle cellule B.
Queste vie di segnalazione funzionano con qualsiasi recettore Ig, in quanto tutti si associano ad Igα eIgβ per poter trasdurre il segnale.
2

1.2.2 Ruolo dei recettori CR2/CD21 come corecettori per le cellule B
I linfociti B esprimono un recettore per la proteina del complemento C3d che prende il nome diCR2 o CD21. Il complesso C3d+antigene o quello C3d+antigene+anticorpo lega il linfocita in modo chel’Ig riconosca l’antigene e CR2 riconosca la proteina del complemento. CR2 è espresso sotto forma dicomplesso con altre due proteine, CD19 e CD81: questo complesso viene spesso chiamato complessocorecettoriale delle cellule B perchè lega C3d allo stesso momento in cui BCR lega l’antigene. Illegame di C3d al corecettore porta CD19 in prossimità delle chinasi associate al BCR e la coda di CD19diventa in questo modo fosforilata; la fosforilazione risulta nel reclutamento della chinasi Lyn, che puòamplificare il segnale di BCR fosforilando direttamente i domini ITAM. CD19 fosforilata attiva anchealtre vie di segnalazione, tra le quali una legata a PIP3 , che aumentano ulteriormente i pathway apertidalle Ig. Il risultato netto è un grande stimolo della risposta della cellula B stimolata.
1.2.3 Risposte funzionali dei linfociti B agli antigeni
Il riconoscimento dell’antigene stimola l’ingresso delle cellule nella fase G1 del ciclo cellulare, uscendocosì dalla precedente fase G0. La sopravvivenza delle cellule B viene migliorata grazie all’induzione divari geni anti apoptotici. I linfociti B attivati mostrano un’aumentata espressione di molecoleMHCII e di costimolanti (B7-2 prima e B7-1 dopo, ed è questa la ragione per cui sono in grado diattivare i linfociti helper). L’espressione dei recettori per le citochine derivanti dai linfociti T vieneanch’essa aumentata in modo da rendere le cellule B recettive, inoltre cambia anche l’espressione deirecettori per le chemochine in modo da permettere la mobilitazione.
L’importanza del BCR nelle risposte è diversa a seconda dell’antigene. Gli antigeni multivalentihanno di solito parecchi epitopi uguali sulla stessa molecola e sono quindi in grado di stimolare inmodo efficace il linfocita. Gli antigeni peptidici sono invece spesso dotati di un solo epitopo e quindinon sono in grado di stimolare il linfocita: in questo caso il BCR si limita ad internalizzare l’antigeneper presentarlo al linfocita helper, il quale poi si occuperà di attivare la cellula B.
1.3 Risposte anticorpali helper-dipendenti ad antigeni proteici
Le prime fasi delle risposte helper-dipendenti avvengono ai bordi delle zone T e dei follicoli primarie risultano nella proliferazione delle cellule B, nella secrezione di anticorpi iniziale e in un limitatoswitching. Le fasi più tardive avvengono invece nei centri germinativi all’interno dei follicoli linfoidi erisultano nella maturazione dell’affinità, nella generazione di cellule della memoria e nello switchingpiù evidente.
1.3.1 Sequenza degli eventi nelle risposte anticorpali T-dipendenti
1. Assunzione dell’antigene dalle cellule dendritiche e presentazione ai linfociti T helper.
3

2. Attivazione degli helper e espressione di CD40L e citochine.
3. Migrazione degli helper verso il follicolo grazie alle chemochine.
4. Attivazione delle cellule B da antigeni solubili o presentati dalle cellule dendritiche.
5. Processamento e presentazione dell’antigene delle cellule B e migrazione verso la zona T grazie airecettori per chemochine.
6. Interazione tra cellule B e T e attivazione delle prime grazie a CD40L e citochine.
7. Inizio dello switching e della secrezione di Ig.
8. Migrazione delle cellule B attive verso il follicolo, formazione di centri germinali nel follicolo.Nei centri germinali si ha marcato switching, mutazioni somatiche, maturazione dell’affinità egenerazione delle cellule della memoria.
9. Generazione di plasmacellule a lunga vita che migreranno poi nel midollo osseo.
1.3.2 Attivazione degli helper
Le cellule che per prime riconoscono l’antigene sono le cellule dendritiche, che lo processano e locaricano sulle molecole MHCII per farlo riconoscere ai linfociti CD4+ naive. Le cellule dendritiche sonoinoltre stimolate a produrre B7-1 e B7-2 che forniranno secondi segnali per l’attivazione degli helper.I linfociti attivati dalle cellule dendritiche sono indotti a proliferare, esprimere CD40L e secernerevarie citochine. Queste cellule modificano inoltre il loro set di recettori per chemochine, aumentandol’espressione di CXCR5 e diminuendo quella di CCR7; in questo modo la cellula segue il gradiende diconcentrazione di CXCL13, prodotto dalle cellule dendritiche follicolari, che la porta nel follicolo.
1.3.3 Presentazione dell’antigene dalle cellule B e migrazione
Il BCR è un recettore ad alta affinità in grado di internalizzare efficacemente l’antigene per endocitosirendendolo disponibile al processamento e al caricamento sull’MHCII. Sono molti i segnali secondariche permettono ad un linfocita B di rispondere ad antigeni proteici dannosi e non a quelli innocui:
1. L’aiuto dagli helper viene fornito solo da linfociti T che hanno risposto a cellule dendritiche cheesprimono B7; l’espressione di B7 è a sua volta indotta dal coinvolgimento del TCR che riconoscestrutture non-self.
2. I linfociti B percepiscono direttamente i patogeni grazie alla presenza di uno o più TLR, tra i qualiTLR4, TLR5 e TLR9.
I linfociti B antigene attivati downregolano l’espressione di CXCR5 e aumentano l’espressione di CCR7:questo comportamento, opposto a quello dei linfociti T CD4+, li attira verso l’interfaccia T-B dellinfonodo.
In una qualsiasi risposta umorale le cellule B specifiche per l’antigene che ha iniziato la rispostavengono attivate preferenzialmente; esistono varie ragioni per giustificare questo fatto:
1. Solo i linfociti B le cui molecole Ig legano l’antigene possono ricevere i segnali di attivazione.
2. I linfociti B sono in grado di presentare il loro antigene a concentrazioni anche 106 volte minoririspetto all’antigene che non riconoscono in quanto l’internalizzazione via BCR è estremamenteefficiente.
3. I linfociti B nei coniugati cellula T-cellula B sono esposti ai segnali portati da CD40L e da alteconcentrazioni di citochine T-derivate, in parte per via della formazione delle sinapsi immuno-logiche.
4

1.3.4 Effetto aptene-carrier
Gli apteni, come il dinitrofenolo, sono piccole molecole che possono legare anticorpi specifici ma nonsono immunogeni da soli. Se un aptene si lega ad una proteina carrier il loro complesso diventa peròimmunogeno. Tre sono le caratteristiche importanti delle risposte anticorpali verso questi complessi:
1. Sono necessarie sia cellule B specifiche per l’aptene che per il carrier.
2. Per avere risposta carrier e aptene devono essere fisicamente associati, la somministrazione sep-arata non fornisce reazione.
3. L’interazione è MHCII ristretta, cioè gli helper collaborano solo con i linfociti B che esprimonoqueste molecole che vengono riconosciute come self dai T.
I linfociti aptene-specifici legano l’antigene attraverso il determinante dell’aptene, lo internalizzano epresentano i peptidi derivati dalla proteina carrier ai linfociti T carrier-specifici: i due linfociti cooperantiriconoscono dunque due epitopi diversi dello stesso antigene. L’effetto carrier-aptene è alla base dellosviluppo dei vaccini coniugati.
1.3.5 Attivazione delle cellule B helper-dipendente
I linfociti helper attivati esprimono una molecola detta CD40L, il cui recettore è CD40, espresso suilinfociti B che presentano l’antigene: l’interazione tra i due è alla base dell’attivazione. CD40 è unmembro della famiglia dei recettori TNF; CD40L è una proteina trimerica. CD40 è espresso in modocostituivo dalle cellule B mentre il suo ligando viene espresso dagli helper solo dopo che questi sonostati attivati. Il legame tra ligando e recettore induce alterazioni conformazionali dei trimeri di CD40 equesto causa l’associazione di una proteina citosolica detta TRAF; TRAF inizia una cascata enzimaticache porta all’attivazione e alla traslocazione nucleare dei fattori di trascrizione, tra i quali NF-κB eAP-1. L’induzione dei fattori di trascrizione CD40-dipendente è cruciale per la formazione dei centrigerminativi e anche per l’espressione di un gene codificante la deaminasi attivazione-indotta (AID),un enzima critico per lo switching e le mutazioni somatiche. Questo sistema di risposta cellularecontatto-mediata è il meccanismo generale di attivazione di cellule bersaglio da parte degli helper e nonè dunque unico per la produzione di anticorpi.Il virus di Epstein-Barr (EBV) infetta i linfociti B e ne induce proliferazione che può portare a linfoma.La coda citoplasmatica di una proteina trasformante del virus, LMP1, si associa alle stesse molecoleTRAF attivate da CD40 e in questo modo stimola la proliferazione dei linfociti.
I linfociti helper attivati secernono citochine che agiscono insieme a CD40L per stimolare la prolif-erazione e la produzione di anticorpi di diversi isotipi. Le citochine servono per due scopi principali inambito di risposte anticorpali:
1. Aumentano la proliferazione e la differenziazione delle cellule B
2. Promuovono lo switching verso differenti isotipi delle catene pesanti
Il riconoscimento dell’antigene nei linfociti B aumenta l’espressione dei recettori per le citochine, molecolepresenti ad alte concentrazioni nelle sedi di contatto con il linfocita helper. Le citochine helper-derivate,soprattutto IL-2, IL-4 e IL-21, potenziano proliferazione e differenziazione dei linfociti B, allo stessomodo delle citochine BAFF e APRIL della famiglia del TNF. La citochina IL-6, prodotta da macrofa-gi, linfociti T e altre cellule, è invece un fattore di crescita per cellule B già differenziate e secernentianticorpi.
L’attivazione contribuisce all’iniziale formazione di foci extrafollicolari di cellule B attivate che pos-sono andare incontro a differenziazione e switching dell’isotipo. Ognuno dei foci formati contienequalche centinaio di plasmablasti e plasmacellule, i cui anticorpi prodotti possono contribuire a for-mare immunocomplessi che hanno un ruolo nell’iniziare la reazione di formazione del centro germinale.
1.3.6 Reazione del centro germinativo
L’iniziale risposta agli antigeni dei linfociti B si ha nella zona tra i follicoli linfoidi e le zone T; dopoquattro-sette giorni dall’esposizione alcuni dei linfociti B attivati migrano in profondità del follicolo e
5
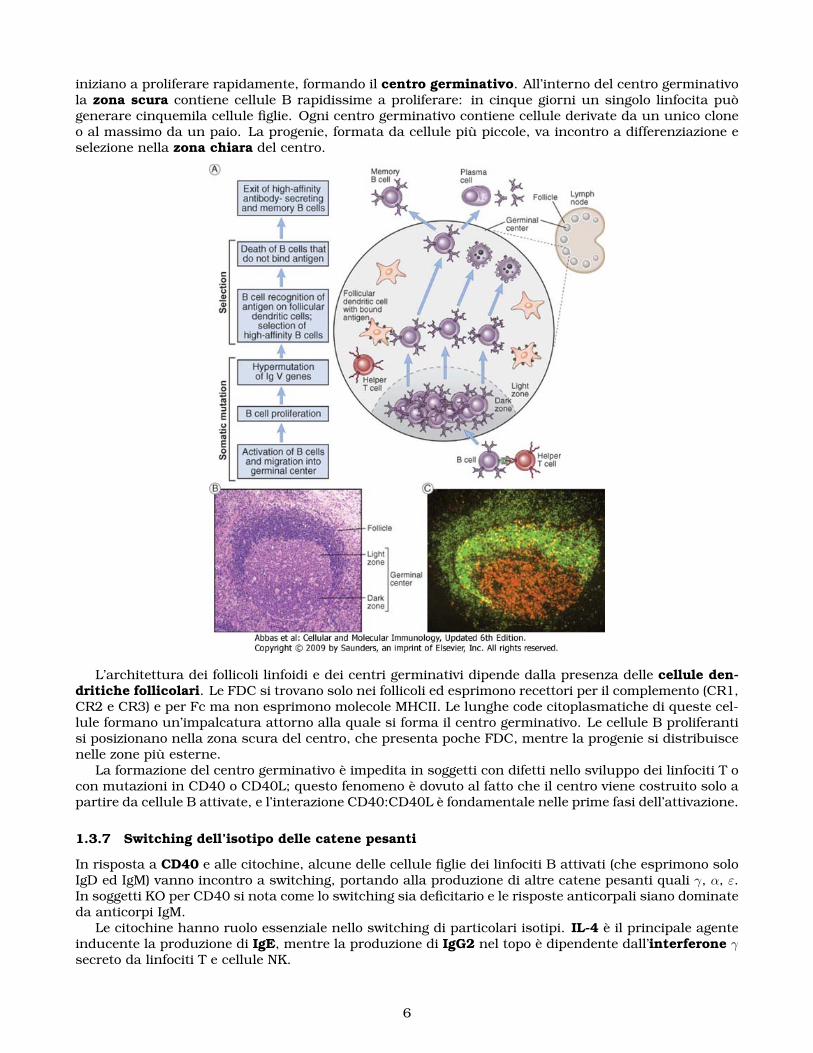
iniziano a proliferare rapidamente, formando il centro germinativo. All’interno del centro germinativola zona scura contiene cellule B rapidissime a proliferare: in cinque giorni un singolo linfocita puògenerare cinquemila cellule figlie. Ogni centro germinativo contiene cellule derivate da un unico cloneo al massimo da un paio. La progenie, formata da cellule più piccole, va incontro a differenziazione eselezione nella zona chiara del centro.
L’architettura dei follicoli linfoidi e dei centri germinativi dipende dalla presenza delle cellule den-dritiche follicolari. Le FDC si trovano solo nei follicoli ed esprimono recettori per il complemento (CR1,CR2 e CR3) e per Fc ma non esprimono molecole MHCII. Le lunghe code citoplasmatiche di queste cel-lule formano un’impalcatura attorno alla quale si forma il centro germinativo. Le cellule B proliferantisi posizionano nella zona scura del centro, che presenta poche FDC, mentre la progenie si distribuiscenelle zone più esterne.
La formazione del centro germinativo è impedita in soggetti con difetti nello sviluppo dei linfociti T ocon mutazioni in CD40 o CD40L; questo fenomeno è dovuto al fatto che il centro viene costruito solo apartire da cellule B attivate, e l’interazione CD40:CD40L è fondamentale nelle prime fasi dell’attivazione.
1.3.7 Switching dell’isotipo delle catene pesanti
In risposta a CD40 e alle citochine, alcune delle cellule figlie dei linfociti B attivati (che esprimono soloIgD ed IgM) vanno incontro a switching, portando alla produzione di altre catene pesanti quali γ, α, ε.In soggetti KO per CD40 si nota come lo switching sia deficitario e le risposte anticorpali siano dominateda anticorpi IgM.
Le citochine hanno ruolo essenziale nello switching di particolari isotipi. IL-4 è il principale agenteinducente la produzione di IgE, mentre la produzione di IgG2 nel topo è dipendente dall’interferone γsecreto da linfociti T e cellule NK.
6

Il principale meccanismo grazie al quale CD40 induce lo switching è lo stimolo alla trascrizione delgene AID. Il gene AID viene dunque trascritto dietro stimolo di CD40, sono però le varie citochinea indurre i fattori di trascrizione che identificano quale catena pesante sarà il target dello switchingmediato da AID.
Lo switching in risposta a diverse tipologie di microbo è regolato dal tipo di cellula helper che vieneattivata dai microbi stessi, ad esempio:
• Batteri con capsule ricche in polisaccaridi stimolano la produzione di IgM i quali poi favorisconoil complemento, la fagocitosi e l’opsonizzazione.
• Gli antigeni polisaccaridici, che non necessitano l’aiuto degli helper, stimolano IgM.
• Molti virus e batteri stimolano la produzione di IgG, che bloccano l’ingresso dei patogeni nellacellula e ne facilitano la fagocitosi. Virus e batteri attivano gli helper del sottogruppo TH1 cheproducono interferone γ, il principale induttore di switching a catena γ nelle cellule B.
• I parassiti elmintici generano risposte di tipo principalmente IgE, anticorpi che partecipano al-l’uccisione eosinofilo-mediata dei patogeni. Gli anticorpi IgE sono anche alla base delle reazioniallergiche. Gli elminti attivano gli helper del sottogruppo TH2 i quali producono IL-4, principaleinduttore di switching verso la catena pesante ε.
In aggiunta a questo meccanismo, anche la sede anatomica influenza lo switching. I linfociti B dellemucose producono soprattutto IgA, l’anticorpo più efficace nell’essere trasportato attraverso gli epiteli;lo switch è stimolato dal transforming growth factor β (TGF-β ) prodotto da parecchie cellule nellemucose. Il recettore TACI (substrato sia per APRIL che per BAFF) ha anch’esso un ruolo critico nelloswitch verso IgA.
Il principale meccanismo molecolare di switching è un processo detto ricombinazione switch incui il segmento genico riarrangiato VDJ di una cellula B si ricombina con un gene della regione Ca valle mentre il DNA in mezzo viene eliminato. Questi eventi ricombinatori coinvolgono sequenzenucleotidiche dette regioni switch poste negli introni J-C alle estremità 5’ di ogni locus CH ; questeregioni sono lunghe 1-10kb, contengono numerose ripetizioni di GC e si trovano a monte di ogni genecodificante catene pesanti ad eccezione del gene δ. A monte di ogni regione di switch c’è un piccoloesone detto esone I (per iniziatore della trascrizione) preceduto da un promotore. CD40 e le citochinestimolano lo switching rendendo più accessibile il DNA di una specifica regione C e inducendo poi latrascrizione attraverso l’esone I, la regione di switch e l’esone CH . Questi trascritti, detti trascrittigerminali, non codificano proteine ma hanno un ruolo fondamentale nello switch.
La trascrizione germinale è accompagnata dall’accessibilità di un particolare gene C a rotture eriparazioni del DNA; come risultato l’esone riarrangiato VDJ giusto a monte della regione di switch µ siaccoppia con la regione C a valle trascrizionalmente attiva.
L’enzima chiave richiesto per lo switching è la deaminasi attivazione-indotta (AID). AID è unaDNA deaminasi che converte la citosina in uracile all’interno di template di DNA a singolo filamento.La trascrizione produce sempre una piccola bolla di DNA a singolo filamento mentre il complesso dellapolimerasi scorre lungo il filamento codificante; dato che il DNA nella bolla è a singolo filamento eccoche può subire l’azione di AID. Un enzima detto uracil N-glicosilasi rimuove a questo punto i residuidi uracile creati da AID generando siti abasici che vengono eliminati dall’endonucleasi Ape1. I buchisu entrambi i filamenti contribuiscono alle rotture sia alla regione Sµ che al locus a valle coinvolto nelloswitching di quel particolare isotipo. L’esistenza di rotture nelle due regioni di switch causa la delezionedel DNA interposto e l’unione delle due giunzioni da parte dei sistemi di riparazione di questo tipo didanno.
7

1.3.8 Maturazione dell’affinità
Il processo di maturazione dell’affinità genera anticorpi con crescente capacità di legare gli antigenie quindi di neutralizzare i microbi. I linfociti helper e le interazioni CD40:CD40L sono richieste perprocedere e quindi la maturazione avviene solo in risposta ad antigeni proteici T-dipendenti.
Nella zona scura proliferativa dei centri germinativi i geni IgV vanno incontro a mutazioni puntifor-mi ad un tasso di una ogni 103 coppie di geni, cioè da mille a diecimila volte più frequentemente delnormale: questo significa che ci sarà una mutazione nelle regioni V in media ogni divisione cellulare.Le mutazioni nel gene continuano anche nella progenie, quindi ogni clone di cellula B può accumulareparecchie mutazioni nella sua vita al centro germinativo.
I meccanismi di mutazione somatica sono poco conosciuti. Si sa che il DNA Ig VDJ diventa altamentemutabile probabilmente a seguito di legame con fattori mutageni. Non si sa se i centri germinativiforniscano segnali contatto-mediati o citochine per stimolare le mutazioni ma si sa che l’enzima AID èfondamentale. I residui di uracile creati da AID possono essere convertiti a residui di timina o possonoessere eliminati dalla glicosilasi, in ogni caso favorendo la mutazione.
In sostanza si crede che le ripetute esposizioni all’antigene generino parecchie mutazioni, di cuila maggior parte inutili mentre alcune effettivamente portano ad un anticorpo più efficace: il passosuccessivo è dunque la selezione delle cellule che producono gli anticorpi migliori.
Le cellule dendritiche follicolari dei centri germinativi presentano gli antigeni, e le cellule B che sonoin grado di legarli con alta affinità vengono selezionate per la sopravvivenza. La prima fase di rispostaall’antigene è la produzione di anticorpi, alcuni dei quali formano complessi con l’antigene e attivano ilcomplemento. Le FDC hanno recettori per la porzione Fc dell’anticorpo e per i prodotti di attivazionedel complemento; questi recettori legano e presentano gli antigeni complessati con anticorpi o prodottidel complemento. Nel frattempo i linfociti B dei centri germinativi che hanno subito le mutazionimigrano verso la zona ricca di FDC: queste cellule moriranno di apoptosi se non verranno salvate dal
8

riconoscimento dell’antigene. In questo modo le cellule che riconoscono in maniera specifica l’antigenemostrato sulle FDC sono selezionate per vivere. L’aumento della produzione di anticorpi va di paripasso con l’eliminazione dell’antigene che sarà sempre più raro sulle FDC: si ha dunque necessità dilinfociti B sempre più specifici per la sopravvivenza perchè dovranno avere un’affinità sempre più altaper legare i pochi antigeni rimasti.
Le mutazioni somatiche avvengono nella zona scura basale del centro germinativo nei centroblastiche contengono l’enzima AID; terminata la mutazione le cellule migrano verso la zona chiara apicaledove vengono selezionate dalle FDC e possono andare incontro ad un ulteriore switching. Le celluleescono infine dal centro germinativo e diventano cellule della memoria o plasmacellule ad altissimaaffinità.
1.3.9 Differenziazione dei linfociti B in plasmacellule secernenti anticorpi
La sintesi degli anticorpi, come la proliferazione dei linfociti B, è stimolata da segnali mediati da CD40e dalle citochine. Molte citochine, tra le quali IL2, IL4 e IL6, sono state individuate come stimolatrici diquesto processo.
All’interno degli organi linfoidi le plasmacellule si trovano soprattutto in sede extrafollicolare, comela polpa rossa della milza o la midollare del linfonodo. Le plasmacellule sono linfociti B differenziati inmaniera terminale e destinate alla sola produzione di anticorpi. Lo sviluppo di queste cellule dipendeall’induzione di un fattore di trascrizione detto BLIMP-1. Esistono due tipologie di plasmacellula. Leplasmacellule a vita breve vengono ritrovate negli organi linfoidi secondari e nei tessuti non linfoidiperiferici. A seguito della reazione del centro germinale alcune plasmacellule ottengono la capacità didirigersi al midollo osseo dove vengono mantenute grazie al recettore BCMA, e queste cellule prendono ilnome di plasmacellule a vita lunga. Tipicamente dopo due o tre settimane dall’infezione da parte di unantigene T-dipendente il midollo diventa una sede chiave di produzione di anticorpi. Le plasmacellulemidollari continuano a secernere antibodi per mesi o anni dopo la scomparsa dell’antigene per fornireprotezione immediata in caso di nuovo incontro; quasi la metà degli anticorpi circolanti di un adulto èprodotto da plasmacellule a vita lunga. Gli anticorpi secreti entrano nella circolazione e nelle secrezionidelle mucose ma le cellule che li hanno prodotti non sono circolanti.
Le molecole Ig secrete e di membrana differiscono per via della loro regione carbossi terminale. Adesempio nelle IgM secrete il dominio Cµ4 è seguito da una coda contenente aminoacidi polari. Nelle IgMdi membrana invece lo stesso dominio è seguito da una sequenza più corta e idrofobica transmembranae da una coda citoplasmatica di tre aminoacidi. La transizione da Ig di membrana a secreta rifletteuna variazione nel processing dell’RNA messaggero per la catena pesante. Il trascritto primario ditutte le cellule B produttrici di IgM contiene infatti VDJ, i quattro esoni Cµ per i domini costanti edue esoni per i domini citoplasmatici e transmembrana. Il processing dell’RNA determina se gli esonitransmembrana e citoplasmatico saranno o meno inclusi nell’mRNA finale. In sostanza tutti i linfociti Bpossono sintetizzare anticorpi sia di membrana che di secrezione; con il procedere della differenziazionela quantità di anticorpi di secrezione tende però ad aumentare. I segnali che regolano il processo dellosplicing alternativo non sono conosciuti. Nota: la forma secretoria della catena pesante δ è raramenteespressa, infatti le IgD sono tipicamente proteine di membrana.
1.3.10 Generazione di cellule della memoria e risposte umorali secondarie
Alcuni dei linfociti B attivati acquisiscono l’abilità di sopravvivere per lunghi periodi apparentementesenza stimolazione antigenica: sono le cellule della memoria. Alcune di queste cellule possono ri-manere negli organi linfoidi mentre altre ricircolano tra la milza ed i linfonodi. Le cellule della memoriatipicamente portano recettori antigenici ad alta affinità e molecole Ig di isotipi switch con più frequenzadei linfociti naive.
Molte delle caratteristiche delle risposte umorali secondarie riflettono la precedente attivazione deilinfociti B da parte degli helper CD4+. Lo switching delle catene pesanti è tipico delle risposte secondariein quanto indotto dai linfociti helper e dalle loro citochine. La maturazione dell’affinità è anch’essa sec-ondaria all’attivazione T-dipendente dei linfociti B. Per neutralizzare molti microbi e le loro tossine sonorichiesti anticorpi ad alta affinità; un vaccino effettivo contro tali microorganismi deve dunque indurrematurazione dell’affinità e produzione di cellule della memoria: entrambi questi processi avverrannosolo in caso di attivazione degli helper. Nel caso di infezioni batteriche in cui l’antigene bersaglio è un
9

polisaccaride (incapace di stimolare i linfociti T), si sfrutta il sistema aptene-carrier e si parla di vacciniconiugati.
1.4 Risposte anticorpali ad antigeni T-indipendenti
Gli anticorpi prodotti in questo tipo di risposte hanno generalmente bassa affinità e sono soprattuttoIgM con un limitato switch verso alcuni sottotipi di IgG.
I più importanti antigeni TI sono polisaccaridi, glicolipidi ed acidi nucleici, tutti in grado diindurre produzione specifica di anticorpi in animali privi di linfociti T. Tutti questi antigeni non possonoessere processati sulle molecole MHC e quindi essere riconosciuti dagli helper. La maggior parte degliantigeni TI è polivalente, e questo induce cross-linking massimale del complesso BCR sui linfociti B,portando ad attivazione senza aiuto degli helper. In aggiunta molti polisaccaridi attivano il complementoseguendo la via alternativa, generando C3d che lega l’antigene e aumenta l’attivazione dei linfociti B.Le risposte delle cellule B dipendono infine dai segnali in arrivo dai recettori della famiglia BAFF cherispondono a fattori di crescita prodotti dalle cellule dendritiche, dai macrofagi e dai TLR.
Le risposte agli antigeni TI sono diverse a seconda del sito anatomico; possono avere inizio nellamilza, nel midollo, nel peritoneo o nelle mucose. I macrofagi in associazione alla milza sono particolar-mente efficienti nell’intrappolare polisaccaridi. Le cellule B della zona marginale sono un sottogruppodelle cellule B che risponde soprattutto ai polisaccaridi producendo IgM. Un’altra linea di cellule B cherisponde bene agli antigeni TI è quella delle cellule B B-1, in gran parte derivate dalle cellule staminalidel fegato fetale e sono esposte all’antigene principalmente nel peritoneo e nelle mucose.
Il senso pratico degli antigeni TI è che molti polisaccaridi parietali dei batteri vi appartengono;individui con deficienze congenite o acquisite dell’immunità umorale sono infatti molto suscettibili adinfezioni di batteri capsulati, quali Pneumococcus, Meningococcus e Haemophilus. In aggiunta gliantigeni TI contribuiscono alla generazione degli anticorpi naturali, normalmente presenti in circoloe apparentemente indotti senza esposizione ai patogeni. La maggior parte degli anticorpi naturali habassa affinità ed è prodotta dalle cellule B di tipo B-1 del peritoneo stimolate dai batteri del tratto GI.
Alcuni antigeni TI inducono isotipi diversi da IgM. Nell’uomo l’anticorpo principale indotto dalpolisaccaride di capsula del pneumococco è IgG2. In assenza di cellule T, BAFF e APRIL possonoindurre nelle cellule di origine mieloide (cellule dendritiche, macrofagi) la sintesi di AID per dar luogoallo switch.
Nonostante l’incapacità di attivare gli helper, molti vaccini polisaccaridici producono immunità pro-tettiva di lunga durata. Risposte secondarie rapide e ampie tipiche della memoria si sviluppano peresposizione secondaria a questi antigeni. Il fenomeno della memoria IgM è stato dimostrato e in topie uomo è possibile evidenziare le cellule B della memoria per antigeni TI; nell’uomo queste celluleesprimono alti livelli di CD27 e IgM o IgD.
1.5 Feedback anticorpale: regolazione della risposta umorale da parte dei re-cettori Fc
Gli anticorpi secreti inibiscono la continua attivazione delle cellule B formando complessi antigene-anticorpo che si legano in simultanea ai recettori antigenici ed ai recettori Fc sui linfociti B antigenespecifici: questa è la spiegazione del fenomeno di feedback anticorpale, cioè della downregolazionedella produzione di anticorpi da parte delle IgG secrete. Gli anticorpi IgG inibiscono i linfociti for-mando appunto complessi che si legano ad un recettore per la forzione Fc della molecola chiamatorecettore FcγII (FcγRIIB o CD32). Il dominio citoplasmatico del recettore contiene un dominio a seiamminoacidi condiviso con altri recettori di questo tiipo che mediano segnali negativi; per analogiacon gli ITAM questo dominio viene chiamato ITIM (Immunoreceptor Tyrosin-based Inhibition Motif).Quando il recettore viene stimolato il dominio ITIM viene fosforilato formando un sito di attacco perl’inositolo 5-fosfatasi SHIP; SHIP idrolizza un fosfato su PIP3 e in questo modo termina la risposta dellinfocita all’antigene. Il complesso antigene anticorpo interagisce simultaneamente sia con il recettoreantigenico che con quello per la porzione Fc, portando la fosfatasi inibitoria vicina al recettore anti-genico da bloccare. L’importanza dell’inibizione attraverso FcγRIIB è dimostrata nei topi KO per questogene. Un polimorfisfmo in questo gene è stato collegato al lupus eritematoso sistemico nell’uomo.
I linfociti B esprimono un altro recettore inibitorio detto CD22, una lectina che lega acido sialico. Illigando naturale non è conosciuto e non si sa come si attivi ma si sa che topi KO mostrano una enorme
10
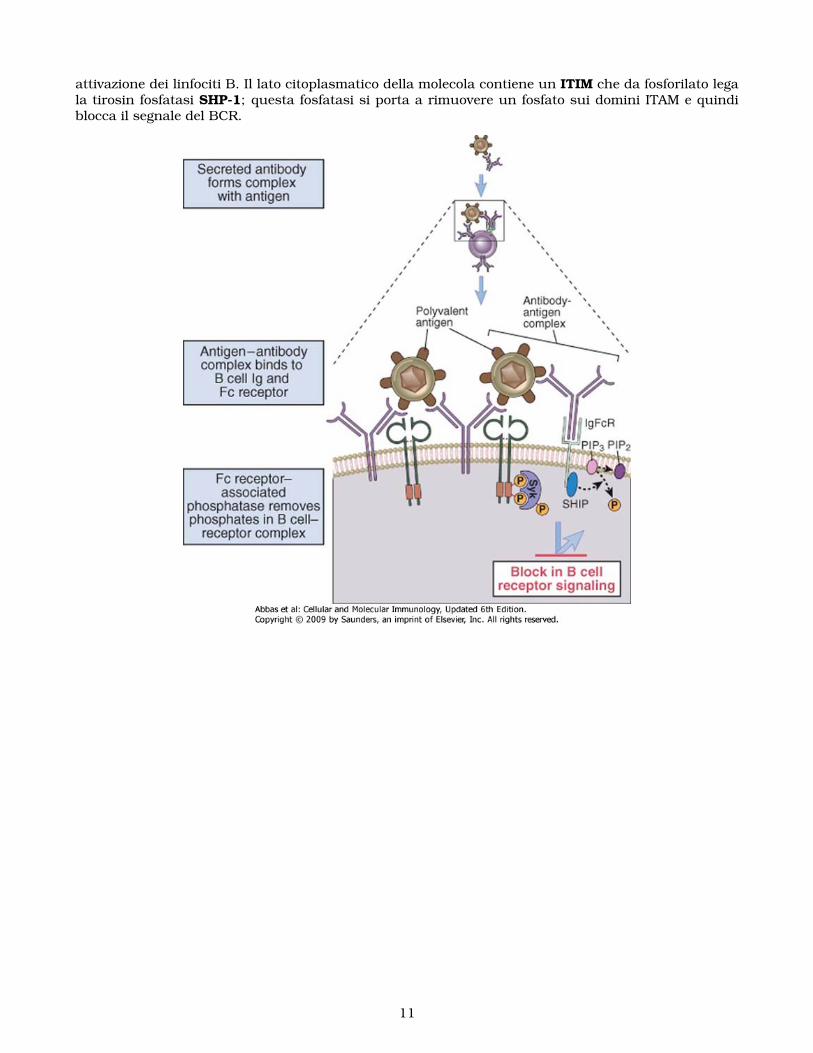
attivazione dei linfociti B. Il lato citoplasmatico della molecola contiene un ITIM che da fosforilato legala tirosin fosfatasi SHP-1; questa fosfatasi si porta a rimuovere un fosfato sui domini ITAM e quindiblocca il segnale del BCR.
11

















