CAPITOLO VI - I Canali di Membranaxoomer.virgilio.it/luciotroise/estetica/sistema nervoso.pdf ·...
33
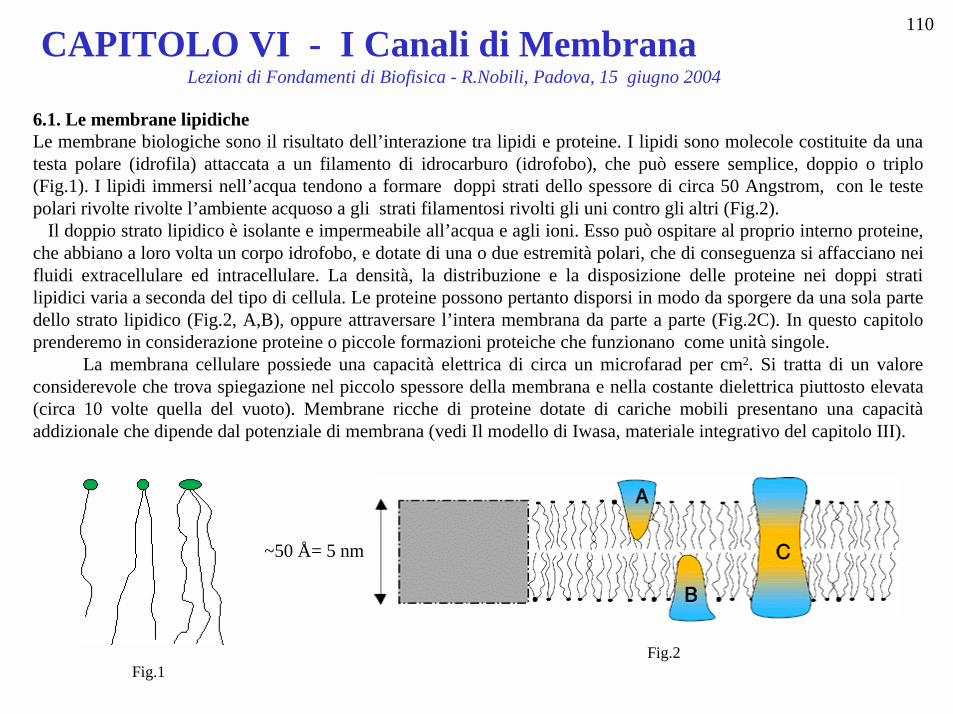
CAPITOLO VI - I Canali di Membrana Lezioni di Fondamenti di Biofisica - R.Nobili, Padova, 15 giugno 2004 110 6.1. Le membrane lipidiche Le membrane biologiche sono il risultato dell’interazione tra lipidi e proteine. I lipidi sono molecole costituite da una testa polare (idrofila) attaccata a un filamento di idrocarburo (idrofobo), che può essere semplice, doppio o triplo (Fig.1). I lipidi immersi nell’acqua tendono a formare doppi strati dello spessore di circa 50 Angstrom, con le teste polari rivolte rivolte l’ambiente acquoso a gli strati filamentosi rivolti gli uni contro gli altri (Fig.2). Il doppio strato lipidico è isolante e impermeabile all’acqua e agli ioni. Esso può ospitare al proprio interno proteine, che abbiano a loro volta un corpo idrofobo, e dotate di una o due estremità polari, che di conseguenza si affacciano nei fluidi extracellulare ed intracellulare. La densità, la distribuzione e la disposizione delle proteine nei doppi strati lipidici varia a seconda del tipo di cellula. Le proteine possono pertanto disporsi in modo da sporgere da una sola parte dello strato lipidico (Fig.2, A,B), oppure attraversare l’intera membrana da parte a parte (Fig.2C). In questo capitolo prenderemo in considerazione proteine o piccole formazioni proteiche che funzionano come unità singole. La membrana cellulare possiede una capacità elettrica di circa un microfarad per cm 2 . Si tratta di un valore considerevole che trova spiegazione nel piccolo spessore della membrana e nella costante dielettrica piuttosto elevata (circa 10 volte quella del vuoto). Membrane ricche di proteine dotate di cariche mobili presentano una capacità addizionale che dipende dal potenziale di membrana (vedi Il modello di Iwasa, materiale integrativo del capitolo III). Fig.1 Fig.2 ~50 Å= 5 nm
Transcript of CAPITOLO VI - I Canali di Membranaxoomer.virgilio.it/luciotroise/estetica/sistema nervoso.pdf ·...
CAPITOLO VI - I Canali di MembranaLezioni di Fondamenti di Biofisica - R.Nobili, Padova, 15 giugno 2004
110
6.1. Le membrane lipidicheLe membrane biologiche sono il risultato dell’interazione tra lipidi e proteine. I lipidi sono molecole costituite da unatesta polare (idrofila) attaccata a un filamento di idrocarburo (idrofobo), che può essere semplice, doppio o triplo(Fig.1). I lipidi immersi nell’acqua tendono a formare doppi strati dello spessore di circa 50 Angstrom, con le testepolari rivolte rivolte l’ambiente acquoso a gli strati filamentosi rivolti gli uni contro gli altri (Fig.2). Il doppio strato lipidico è isolante e impermeabile all’acqua e agli ioni. Esso può ospitare al proprio interno proteine,che abbiano a loro volta un corpo idrofobo, e dotate di una o due estremità polari, che di conseguenza si affacciano neifluidi extracellulare ed intracellulare. La densità, la distribuzione e la disposizione delle proteine nei doppi stratilipidici varia a seconda del tipo di cellula. Le proteine possono pertanto disporsi in modo da sporgere da una sola partedello strato lipidico (Fig.2, A,B), oppure attraversare l’intera membrana da parte a parte (Fig.2C). In questo capitoloprenderemo in considerazione proteine o piccole formazioni proteiche che funzionano come unità singole. La membrana cellulare possiede una capacità elettrica di circa un microfarad per cm2. Si tratta di un valoreconsiderevole che trova spiegazione nel piccolo spessore della membrana e nella costante dielettrica piuttosto elevata(circa 10 volte quella del vuoto). Membrane ricche di proteine dotate di cariche mobili presentano una capacitàaddizionale che dipende dal potenziale di membrana (vedi Il modello di Iwasa, materiale integrativo del capitolo III).
Fig.1Fig.2
~50 Å= 5 nm

Fig.3
6.1.1. La varietà dei lipidi.Il grande gruppo dei lipidi comprende composti caratterizzati da una relativa insolubilità in acqua e dalla solubilità insolventi organici come benzolo, etere di petrolio e cloroformio. Questa proprietà generale dei lipidi e dei composti adessi simili è dovuta al fatto che nella loro molecola predominano lunghe catene di idrocarburi alifatici (catene dicarbonio lineari) talvolta anche con anelli benzoici: strutture entrambe apolari e idrofobiche. In molti lipidi un’estremitàdi queste catene è attaccata a un gruppo polare che rende la molecola idrofila e quindi in grado di legare l’acqua permezzo di ponti idrogeno (Capitolo II). I lipidi costituiscono una varietà di molecole piuttosto numerosa. Essi possonoessere classificati come segue:
Lipidi semplici (esteri alcolici grassi).Si formano per combinazione di un alcol con un acido grasso edeliminazione di una molecola di acqua (Fig.3). Tra questi vi sono igrassi naturali (trigliceridi), e le cere (es. cera d’api) che si formanoper combinazione di acidi grassi con alcoli diversi dal glicerolo(glicerina). Se ne conoscono circa 100 tipi diversi.
Steroidi.Possiedono il nucleo del ciclo-pentano-peridrofenantrene. Essicomprendono una serie di sostanze di enorme importanza perl’organismo (ormoni sessuali, ormoni corticosurrenali, vitamina D,acidi biliari). Gl steroidi che possiedono un ruppo -OH sono dettisteroli. Il colesterolo è uno sterolo molto diffuso nell’organismo(Fig.4). E’ presente nella bile, nell’encefalo, nelle ghiandole surrenali ein altri organi. Spesso lo si trova esterificato con acidi grassi.
Lipidi coniugati.Questo gruppo comprende: lecitine, cefaline, inositidi, plasmalogeni(acetilfosfatidi). Sottoposti ad idrolisi, questi lipidi danno origine, oltreche ad alcol e acidi, ad altri composti. Per il fatto di essere più o menosolubili, insieme agli steroidi, essi sono detti lipoidi. Sono uncomponente importante delle membrane cellulari.
111

Fig.4
Fra i lipdi coniugati annoveriamo: i fosfatidi (fosfolipidi) che sono diesteri dell’acido fosforico, che può essereesterificato con glicerolo, colina, etanolamina, serina, ecc; i glicolipidi e gli sfingolipidi, che sono caratterizzati dalfatto che che il glicerolo è sostituito da un aminoalcol, la sfingosina, a questo gruppo appartengono ad esempio lesfingomieline che si trovano soprattutto nelle guaine mieliniche dei nervi; i cerebrosidi, che possiedono galattosio eglucosio nella loro molecola; i solfatidi, che contengono acido solforico esterificato col galattosio; i gangliosidi,queste molecole di grande polarità sono presenti nelle membrane cellulari dove sono coinvolte nel trasporto ionicoattraverso le membrane e dove svolgono anche il ruolo di recettori di virus. I lipidi sono un componente importante dell’alimentazione anche per le loro proprietà organolettiche (Fig.5). Essisono i componenti principali dei grassi animali e vegetali, oli vari, cacao, latte ecc.
Fig.5
112

6.1.2. I lipidi delle membrane cellulari.La figura 6 rappresenta tre classi di lipidi che contribuiscono a formare le membrane cellulari. Le loro diversitàstrutturali riflettono le diversità funzionali della membrana, sia per quanto riguarda la capacità di ospitare vari tipidi proteine sia per la capacità d’interagire con le molecole esterne alla membrana.
113
Fig.6

Fig.7 Fig.8
La figura 7 rende un’idea approssimativa della complessità e varietà della composizione lipidica e proteica di unamembrana cellulare. Un glicolipide come il glicosil-fosfatitidil-inositolo (GPI) svolge un ruolo importante nell’ancoraggio di proteineche non possono entrare nella membrana lipidica per la mancanza di un segmento idrofobo sufficientemente esteso. Le proteine che si ancorano al GPI formano una diversa famiglia di molecole che comprende enzimi associati allamembrana, molecole adesive, antigeni attivanti, marcatori di differenziazione, componenti dell’involucro protettivo deiprotozoi, e una miscellanea di altre glicoproteine. Nei reni sono state identificate diverse proteine ancorate al GPI tra lequali l’uromodulina (glicoproteina di Tamm-Horsfall), l’anidrasi carbonica del IV tipo, la fosfatasi alcalina, Thy-1,BP-3, l’aminopeptidasi P e la dipeptidilpeptidasi. La figura 8 mostra come ha luogo il processo di ancoraggio di unaproteina al glicolopide. Una proteina temporaneamente agganciata alla membrana ad un terminale lipidico idrofobo(terminale C) si trova in prossimità di un glicolipide che porta un radicale amminico H2N. Il legame peptidico -CO-NH- della proteina viene successivamente rimpiazzato da un legame glicoproteico dello stesso tipo.
114

Fig.9
6.2. Le proteine di membrana e le loro funzioni.
Le membrane cellulari ospitano un grande varietà di proteine che hanno diversi ruoli funzionali (Fig.9): adesioneintercellulare; trasporto attivo o passivo di ioni e piccole molecole; ricezione di segnali di varia natura;riconoscimento di gruppi funzionali e distribuzioni di cariche molecolari molecolari ecc. La macchina cellulare èun sistema straordinariamente complesso caratterizzato da una molteplicità di livelli funzionali. Essa è nello stessotempo: 1) un sistema chimico che utilizza certe sorgenti di energia per sintetizzare nuove molecole; 2) un sistemaelettrochimico che promuove processi di scambio ionico e molecolare con l’ambiente esterno; 3) un sistemacibernetico che provvede alla regolazione dei propri processi interni; 4) un sistema di comunicazione che riceve edemette segnali da e verso il mondo esterno; 5) un sistema informatico che elabora informazione in modi complessi(Fig.10). In The Theory of Self-Reproducing Automata (1965) John von Neumann ha caratterizzato i sistemi viventicome macchine dotate di capacità costruttive e computazionali universali. Secondo il grande fisico-matematicoqueste macchine possiedono un apparato chimico-fisico capace di produrre qualunque tipo di struttura materiale,sulla base di adeguati programmi, e di apparati di calcolo che, come i moderni calcolatori, sono dotati di capacitàalgoritmiche universali.
115
Fig.10

116
dove R1, R2 rappresentano i radicali degli aminoacidi costituenti. Il legame -NH-CO- è conosciuto col nome di legamepeptidico (Fig.11, lato destro). Le molecole che si formano conservano il loro carattere anfotero perché contengonosempre un gruppo basico ad una estremità e un gruppo acido all’altra, e in più hanno residui laterali (radicali) chepossono essere acidi o basici. I 20 aminoacidi si distinguono per le diverse proprietà meccaniche e chimico-fisiche deiloro radicali che sono così suddivisi:
• Acidi: -CH2COOH- (acido aspartico); -CH2CH2COO- (acido glutammico).• Basici: -(CH2)4NH3
+ (lisina); -(CH2)3NHC(NH2)NH3+ (arginina).
• Polari non carichi:-H (glicina); -CH2SH (cisteina); -CH2OH (serina); -CH(OH)CH3 (treonina); -CHC6H4OH (tirosina);-CH2CONH2 (asparagina); -CH2CH2CONH2 (glutammina); -CH2C(NHCHNCH) (istidina).• Non polari: -CH3 (alanina); -CH(CH3)2 (valina); -CH2CH(CH3)2 (leucina); -CH(CH3)CH2CH3 (isoleucina); -CH2C6H5
(fenilalanina); -CH2CH2CH2N (prolina); -CH2C(CHNH)C6H4 (triptofano); -CH2CH2SCH3 (metionina).
Una catena polipeptidica lineare formata da aminoacidi è conosciuta come struttura primaria della molecola proteica.Essa costituisce la struttura specifica e determina, entro certi limiti, le strutture dette secondarie e terziarie.
Fig.11
6.2.1. Strutture primaria, secondaria, terziaria e quaternaria di una proteina.Gli elementi costitutivi delle proteine sono 20 differenti tipi di aminoacidi. Un aminoacido deriva essenzialmente da unacido organico in cui un atomo di idrogeno del carbonio vicino al gruppo carbossilico -COOH è sostituito da un gruppoaminico -NH2. Al carbonio vicino è legata una catena laterale, o radicale, (Fig.11, lato sinistro) che conferiscono agliaminoacidi varie proprietà importanti per la formazione delle proteine. La presenza contemporanea di un gruppoaminico (basico) e di uno carbossilico (acido) conferiscono agli aminoacidi un carattere anfotero (in soluzione essipresentano cariche ioniche di segno opposto). Gli aminoacidi liberi possono provenire dalla demolizione di proteine o dal siero che circonda la cellula. Da essi lacellula trae gli elementi necessari per la sintesi delle proteine. La condensazione di aminoacidi avviene in modo che ilgruppo acido di un aminoacido si combina col gruppo aminico di un aminoacido vicino con la perdita di una molecolad’acqua (Fig.11).

Fig.12
117Struttura secondaria di una proteina.In una proteina formata da parecchie centinaia di aminoacidi la catena può talvolta essere lineare, ma più di frequenteessa assume forme diverse che costituiscono la cosiddetta struttura secondaria. Per le proteine di membrana sonostati classificati due tipi di strutture secondarie: le α-eliche e i β-foglietti (Fig.12). La struttura ad α-elica si forma quando la catena polipeptidica assume l’andamento di una spirale avvolta attornoad un cilindro immaginario, in modo che i ponti di idrogeno si stabiliscono all’interno della molecola e non tramolecole adiacenti (Fig.12A). I β-foglietti sono formati da catene di aminoacidi ripiegate in modo da formare una struttura pieghettata nella qualei residui laterali degli aminoacidi sporgono perpendicolarmente rispetto al piano della catena principale (Fig.12B). Lecatene principali sono legate tra loro da ponti idrogeno e formano una rete peptidica. La figura 13 illustra le strutture di due proteine di membrana in cui sono nettamente prevalenti le α-eliche(batteriorodopsina) e i β-foglietti (purina).
A
B
Fig.13

118
Struttura quaternaria di una proteina.Aggregati di unità proteiche organizzate in unità secondarie e terziarie possono dar luogo a forme più complesse diorganizzazione, dette strutture quaternarie (Fig.14C). Queste sono formate da due o più catene peptidiche, che possonoessere uguali o differenti e che sono legate fra loro da legami deboli (non covalenti). Le strutture quaternarie si formano spontaneamente per autoaggregazione grazie alle distribuzioni complementarialtamente specifiche delle cariche superficiali proprie delle strutture secondarie e terziarie che costituiscono icomponenti della struttura quaternaria.
Struttura terziaria di una proteina.Le catene polipeptidiche con struttura secondaria sono in generale intervallate da tratti lineari che favoriscono ilripiegamento delle struttura secondaria (Fig.14A) e la formazione di uno stato di aggregazione nel quale tratti distrutture secondarie sono legati o agganciati tra loro da legami chimici relativamente forti, ad esempio ponti -S-S-(disolfuro) che si formano per eliminazione di due ioni idrogeno dai radicali della cisteina, e legami polari o covalentitra i radicali di aminoacidi vicini (Fig.14B). La rottura di questi legami, causata da temperatura elevata o eccessivevariazioni di pH, e il conseguente disfacimento della struttura terziaria, è noto come processo di denaturazione delleproteine. Talvolta questo processo è reversibile e la proteina può riacquistare la sua configurazione originaria(rinaturazione).
Fig.14

6.3. I canali di membrana.
Le strutture proteiche, generalmente quaternarie, che condizionano il passaggio di piccole molecole o ioni attraverso lemembrane cellulari sono generalmente note come canali di membrana. Le funzioni esercitate da queste proteine sonospesso altamente specifiche, ma talvolta una stessa proteina può svolgere funzioni diverse. La varietà delle funzioniche sono espletate dai canali di membrana è grandissima e la loro conoscenza si amplia di mese in mese. Pertanto èchiaro che, allo stato presente, una classificazione sistematica dei canali di membrana sarebbe provvisoria e indefinitiva poco sensata. Possiamo tuttavia ripartire i canali di membrana in almeno tre grandi gruppi funzionali: 1)trasportatori metabolici; 2) attivatori e regolatori dello stato elettrochimico cellulare; 3) recettori e trasportatori disegnali. Al primo gruppo appartengono ad esempio i trasportatori di acqua (acquaporine), di glucosio (cotrasportatorisodio-glucosio), le giunzioni intercellulari (gap-junctions) ecc. Al secondo appartengono ad esempio i canali chepossono essere controllati dal potenziale di membrana (canali del potassio, del sodio, del calcio ecc); i canali attivati daioni (come i canali del potassio o del calcio attivati dal calcio) o da altri ligandi; i canali cotrasportatori; quelliantitrasportatori, ecc. Al terzo gruppo appartengono i recettori dei segnali nervosi eccitatori e inibitori, che si apronosotto l’azione di vari tipi di neurotrasmettitori e sono soggetti a vari tipi di regolazione da parte dei potenziali dimembrana o di molecole di varia specie. Citiamo ad esempio i recettori nicotinici che lasciano passare sodio epotassio sotto l’azione dell’acetilcolina rilasciata dalle terminazioni nervose dei neuroni eccitatori; i recettori delglutammato, che lasciano passare anch’essi sodio e potassio, ma in modo più veloce, sotto l’azione del glutammatorilasciato da neuroni eccitatori; i recettori dell’acido gamma-amino-butirrico (GABA), che lasciano passare ioni cloroquando subiscono l’azione del GABA rilasciato dai terminali assonici dei neuroni inibitori, ecc. Poiché in questo capitolo il nostro interesse è principalmente rivolto allo studio dei fenomeni elettrochimici,focalizzeremo la nostra attenzione solo sui canali del secondo gruppo; in particolare su quelli che intervengono inmodo essenziale nella generazione dei potenziali di azione delle cellule eccitabili.
6.3.1. Processi di trasporto attivi e passivi.La differenza di potenziale che si stabilisce attraverso la membrana cellulare di un organismo animale dipende in modoessenziale dalla presenza di un duplice meccanismo di trasporto, uno attivo e l’altro passivo: la pompa del sodio-potassio e il canale di co-trasporto sodio-glucosio. La prima è conosciuta anche come Na-K-ATPasi: nome la cuidesinenza -asi serve ad indicare che questa proteina funziona anche come un enzima che catalizza l’idrolisidell’adenosintrifosfato (ATP), processo questo che fornisce energia alla proteina.
119

I due canali sono rappresentati insieme nella figura 15. La pompa del sodio-potassio trasporta 3 ioni sodio fuori dallacellula e 2 ioni potassio dentro la cellula per ogni molecola di ATP idrolizzata. Quest’attività di pompaggio, presente intutte le cellule animali, determina considerevoli aumenti relativi del sodio extracellulare e del potassio intracellulare e laconseguente formazione di una differenza di potenziale tra i due lati della membrana. Poiché le membrane cellulari sonoin generale piuttosto permeabili al potassio ma assai poco al sodio, il potenziale di Nernst corrispondente al rapporto trale concentrazioni intracellulare ed extracellulare del potassio si avvicina al potenziale della membrana; mentre invece,per la ragione opposta, il potenziale di Nernst del sodio assume, in generale, valori lontani dal potenziale di membrana. Il secondo tipo di canale sfrutta l’elevata concentrazione relativa di sodio extracellulare, causata delle pompe sodio-potassio, per introdurre nella cellula il glucosio necessario per la sintesi dell’ATP. Questo canale lascia passare uno ionesodio solo se questo si trova associato ad una molecola di glucosio. Questo meccanismo è definito di co-trasporto. Conun meccanismo complementare, i canali di antiporto sodio-calcio, sfruttano il gradiente del sodio per espellere il calciodall’interno delle cellule.
120
Fig.15

1216.3.2. Le pompe sodio-potassio.La figura 16 illustra come il pompaggio del sodio e del potassio sia effettuato da una transizione conformazionale dellaNa-K-ATP-asi. In una prima conformazione la proteina si apre in modo da esporre al citoplasma una cavità che rendepossibile la cattura di tre ioni sodio da parte di tre siti proteici elettronegativi. Successivamente, grazie all’energiafornita dall’ATP, il canale passa ad una seconda conformazione (metastabile) che comporta la chiusura della primacavità, la rimozione dei siti del sodio e la formazione di una seconda cavità esposta verso l’esterno della cellula. Inseguito a questo processo il sodio viene rilasciato nello spazio extracellulare mentre il potassio extracellulare entra nellacavità ancora aperta legandosi a due siti elettronegativi. Appena gli ioni potassio sono stati catturati, la proteina ritornaspontaneamente nella configurazione originaria, rilasciando il potassio nello spazio intracellulare e dando pertantoinizio a un nuovo ciclo di pompaggio.
Fig.16
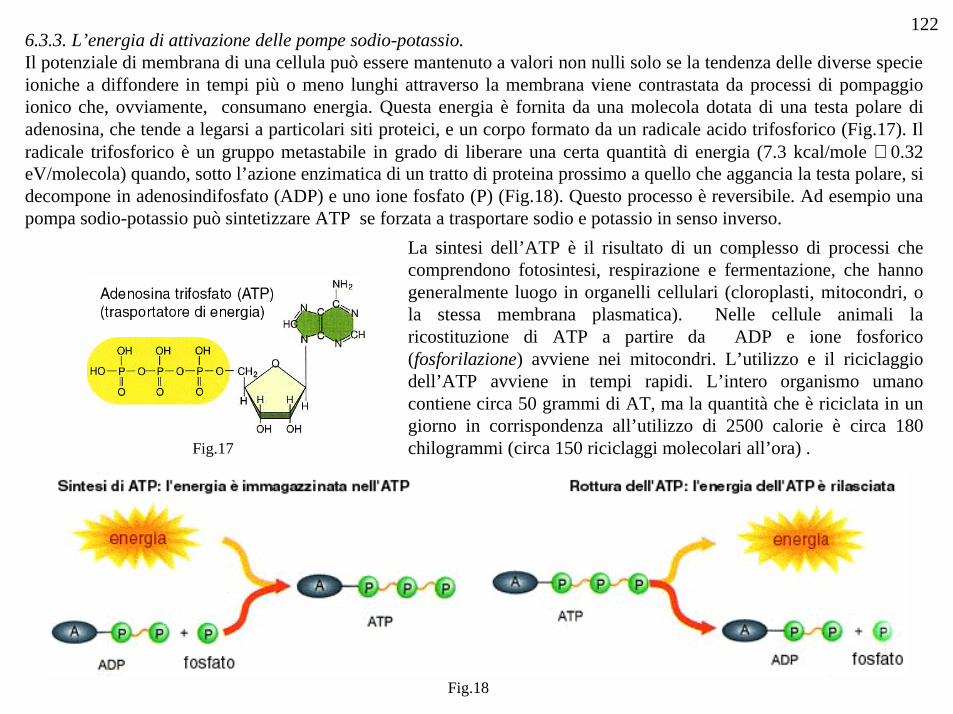
Fig.18
1226.3.3. L’energia di attivazione delle pompe sodio-potassio.Il potenziale di membrana di una cellula può essere mantenuto a valori non nulli solo se la tendenza delle diverse specieioniche a diffondere in tempi più o meno lunghi attraverso la membrana viene contrastata da processi di pompaggioionico che, ovviamente, consumano energia. Questa energia è fornita da una molecola dotata di una testa polare diadenosina, che tende a legarsi a particolari siti proteici, e un corpo formato da un radicale acido trifosforico (Fig.17). Ilradicale trifosforico è un gruppo metastabile in grado di liberare una certa quantità di energia (7.3 kcal/mole ≅ 0.32eV/molecola) quando, sotto l’azione enzimatica di un tratto di proteina prossimo a quello che aggancia la testa polare, sidecompone in adenosindifosfato (ADP) e uno ione fosfato (P) (Fig.18). Questo processo è reversibile. Ad esempio unapompa sodio-potassio può sintetizzare ATP se forzata a trasportare sodio e potassio in senso inverso.
La sintesi dell’ATP è il risultato di un complesso di processi checomprendono fotosintesi, respirazione e fermentazione, che hannogeneralmente luogo in organelli cellulari (cloroplasti, mitocondri, ola stessa membrana plasmatica). Nelle cellule animali laricostituzione di ATP a partire da ADP e ione fosforico(fosforilazione) avviene nei mitocondri. L’utilizzo e il riciclaggiodell’ATP avviene in tempi rapidi. L’intero organismo umanocontiene circa 50 grammi di AT, ma la quantità che è riciclata in ungiorno in corrispondenza all’utilizzo di 2500 calorie è circa 180chilogrammi (circa 150 riciclaggi molecolari all’ora) .Fig.17

Fig.19
6.3.4. Struttura interna dei canali ionici.La figura 19 illustra il meccanismo di funzionamento del canale del potassio scoperto da Roderick MacKinnon nel1998. Gli ioni (potassio, sodio, calcio ecc.) assieme al loro guscio di idratazione vengono accolti nel poro idrofilo (A).Questo comunica con un tratto di canale (B) tappezzato da atomi di ossigeno che si trovano a distanze uguali a quelledegli atomi di ossigeno nel guscio idratazione del potassio. Per questa ragione solo gli ioni potassio riescono adattraversare il canale ossigenato, che pertanto funziona come un filtro ionico. La figura 20 illustra il canale di acquaporina scoperto da Peter Agree nella metà degli anni 80. Lo stesso autore hadefinitivamente chiarito il meccanismo di funzionamento nel 2000. Questo canale lascia passare molecole d’acqua. Lacarica positiva posta nel centro del canale impedisce il passaggio di ioni positivi, in particolare dell’idronio (H3O
+),impedendo così il passaggio dei protoni. Per queste loro scoperte i due autori hanno ricevuto il premio Nobel per la chimica nel 2003.
123
Fig.20
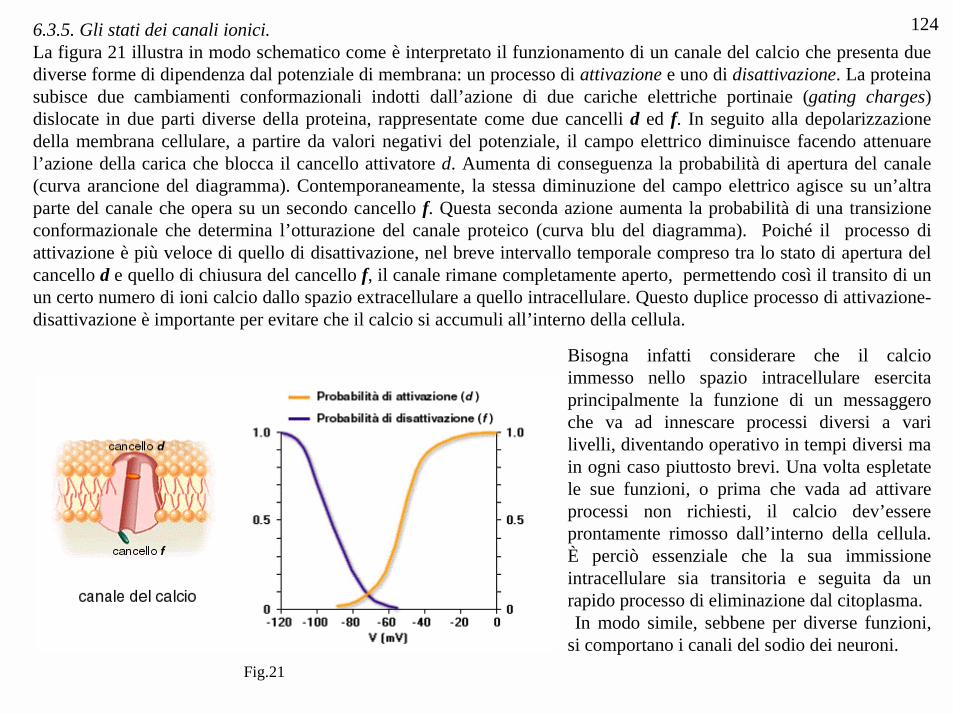
Fig.21
1246.3.5. Gli stati dei canali ionici.La figura 21 illustra in modo schematico come è interpretato il funzionamento di un canale del calcio che presenta duediverse forme di dipendenza dal potenziale di membrana: un processo di attivazione e uno di disattivazione. La proteinasubisce due cambiamenti conformazionali indotti dall’azione di due cariche elettriche portinaie (gating charges)dislocate in due parti diverse della proteina, rappresentate come due cancelli d ed f. In seguito alla depolarizzazionedella membrana cellulare, a partire da valori negativi del potenziale, il campo elettrico diminuisce facendo attenuarel’azione della carica che blocca il cancello attivatore d. Aumenta di conseguenza la probabilità di apertura del canale(curva arancione del diagramma). Contemporaneamente, la stessa diminuzione del campo elettrico agisce su un’altraparte del canale che opera su un secondo cancello f. Questa seconda azione aumenta la probabilità di una transizioneconformazionale che determina l’otturazione del canale proteico (curva blu del diagramma). Poiché il processo diattivazione è più veloce di quello di disattivazione, nel breve intervallo temporale compreso tra lo stato di apertura delcancello d e quello di chiusura del cancello f, il canale rimane completamente aperto, permettendo così il transito di unun certo numero di ioni calcio dallo spazio extracellulare a quello intracellulare. Questo duplice processo di attivazione-disattivazione è importante per evitare che il calcio si accumuli all’interno della cellula.
Bisogna infatti considerare che il calcioimmesso nello spazio intracellulare esercitaprincipalmente la funzione di un messaggeroche va ad innescare processi diversi a varilivelli, diventando operativo in tempi diversi main ogni caso piuttosto brevi. Una volta espletatele sue funzioni, o prima che vada ad attivareprocessi non richiesti, il calcio dev’essereprontamente rimosso dall’interno della cellula.È perciò essenziale che la sua immissioneintracellulare sia transitoria e seguita da unrapido processo di eliminazione dal citoplasma. In modo simile, sebbene per diverse funzioni,si comportano i canali del sodio dei neuroni.

6.3.6. I principali canali ionici delle membrane assoniche dei neuroni.La figura 22 illustra un canale del potassio controllato dal potenziale. Quando il campo elettrico di membrana aumentail canale si chiude, quando il campo diminuisce il canale si apre. Si ritiene che il meccanismo di attivazione dei canalidel calcio, del potassio e del sodio sia lo stesso. È stato dimostrato che la chiusura del canale è accompagnata da unospostamento di circa un’unità di carica elettrica positiva verso la parte esterna della membrana. Questo fenomeno èstato interpretato come effetto di uno scivolamento relativo di α-eliche dotate di cariche elettriche complementari(paragrafo 6.2.1). Un singolo canale di questo tipo presenta una conduttanza massima dell’ordine di 100 pS (picoSiemens), corrispondente a circa 1010 Ohm (107 ioni al secondo sotto la differenza di 100 mV).
125
Fig.22 Fig.23
La figura 23 illustra un canale delsodio che, dopo un processo diattivazione del tutto simile a quellodel potassio, viene disattivato da unmeccanismo a palla e catena.Questo modello di disattivazione èstata suggerito dal fatto che ladisattivazione viene abolita se nellospazio intracellulare è presente unenzima che agisce presumibilmentesu una parte del canale che sporgeall’interno della cellula. A confermadi questa ipotesi è stato notato checerti canali del sodio, che siottengono per mutazione geneticadel DNA che codifica per i canalidel sodio normali, mutazione checomporta la perdita di una parteterminale della proteina dimembrana, perdono la capacità didisattivarsi.

(1) ( )k k kI g E E= −
6.4. Proprietà elettriche delle membrane cellulari
6.4.1. Capacità elettrica e correnti ioniche.In prima approssimazione le proprietà elettriche di una membrana cellulare possono essere schematizzate come inFig.24. La capacità di membrana C è in parallelo con la resistenza (variabile) di membrana R e con un generatore diforza elettromotrice E. I valori di R ed E dipendono dagli equilibri elettrochimici della cellula. Ricordiamo che questi sistabiliscono in regime dinamico sotto l’azione delle pompe di membrana (equazione di Goldman, Capitolo IV, paragrafo4.5.5, Eq. 53). I valori tipici di C sono 1 µF /cm2, corrispondente a 10-200 pF per cellula, a seconda dell’area dellasuperficie cellulare. R varia da 10 Ω a 0.01 MΩ per cm2 di membrana. Valori tipici per una cellula variano tra 1 e 100MΩ. Corrispondentemente si ottengono costanti di tempo RC comprese tra centesimi di millisecondo e alcuni centesimidi secondo. Le correnti elettriche tipiche variano dalla frazione di nA a centinaia di nA. Questa rappresentazione delle proprietà elettriche cellulari, in cui tutte le forme di conduzione elettrica della cellulasono riassunte in un unico termine di resistenza R, è di scarsa utilità. Infatti la dinamica cellulare è dominata dai regimidi apertura e chiusura di canali diversamente permeabili alle diverse specie ioniche, la cui conduttività può dipendere davari fattori come lo stesso potenziale di membrana E, le concentrazioni di calcio o di altri ioni e agenti molecolari.
dove Ek è il potenziale di Nernst e gk è la conduttanza specifica dellamembrana per la specie k. Come è stato spiegato nel Capitolo IVparagrafo 4.4.4., Ek dipende dalla concentrazione interna Cki e da quellaesterna Cke della specie ionica k secondo la formula
Fig.24
126
E’ pertanto necessario disaggregare queste componenti considerandocome la corrente elettrica Ik trasportata da ciascuna specie ionica kdipenda dagli altri fattori. L’equazione standard è
(2) ln .kik
k ke
CkTE
ez C= −
dove zk è la valenza dello ione. Le caratteristiche di conduzione di uncanale ionico potranno pertanto essere raffigurate in digrammi cherappresentano Ik in funzione di E.

Fig.25
Nella figura 25, le specie di canali ionici 1, 2, …,N di una membrana cellulare sono rappresentate come un sistema diconduttanze variabili g1, g2, …, gN poste in serie con un pari numero di generatori di forza elettromotrice E1, E2,…, EN
che rappresentano i potenziali di Nernst. Il potenziale di membrana è rappresentato come un semplice generatore diforza elettromotrice E.
127
Aggiungendo alle correnti elettriche trasportate dai canali ionici quella accumulata orilasciata dalla capacità di membrana in corrispondenza a possibili variazioni di E, lacondizione di mantenimento dell’elettroneutralità intracellulare è espressa dallacondizione di corrente elettrica totale nulla
( )( ) ( )( ) ( )( )1 1 2 2, , , , ... , , 0,N N
dEC g E t E E g E t E E g E t E E
dtλ λ λ+ − + − + + − =
nella quale le conduttanze sono state rappresentate come funzioni dello stessopotenziale di membrana E, del tempo t e di altri parametri λ che influenzano laconduttanza ionica. E’ chiaro che per calcolare come evolve nel corso del tempo lostato elettrochimico del sistema bisognerà prendere in considerazione anche leequazioni che descrivono come le conduttanze dipendono dai loro argomenti.
6.4.2 Alcuni tipi di curve potenziale-corrente.Una prima caratterizzazione della dipendenza delle correnti ioniche dal potenziale dimembrana si può ottenere tracciando i profili delle correnti ioniche Ik, trasportate daicanali di membrana di una determinata specie k, in funzione del potenziale dimembrana E per fissati valori dei parametri λ e in corrispondenza del massimovalore che la conduttanza assume nel tempo. Cioè tracciando i diagrammi dellefunzioni
( )max
(3) , ( ).k k kI g E E Eλ= − Il riferimento a determinate condizioni di conduttanza, in questo caso la conduttanzamassima, è necessario perché in generale la conduttanza aumenta durante la fase diattivazione e diminuisce durante quella di disattivazione.
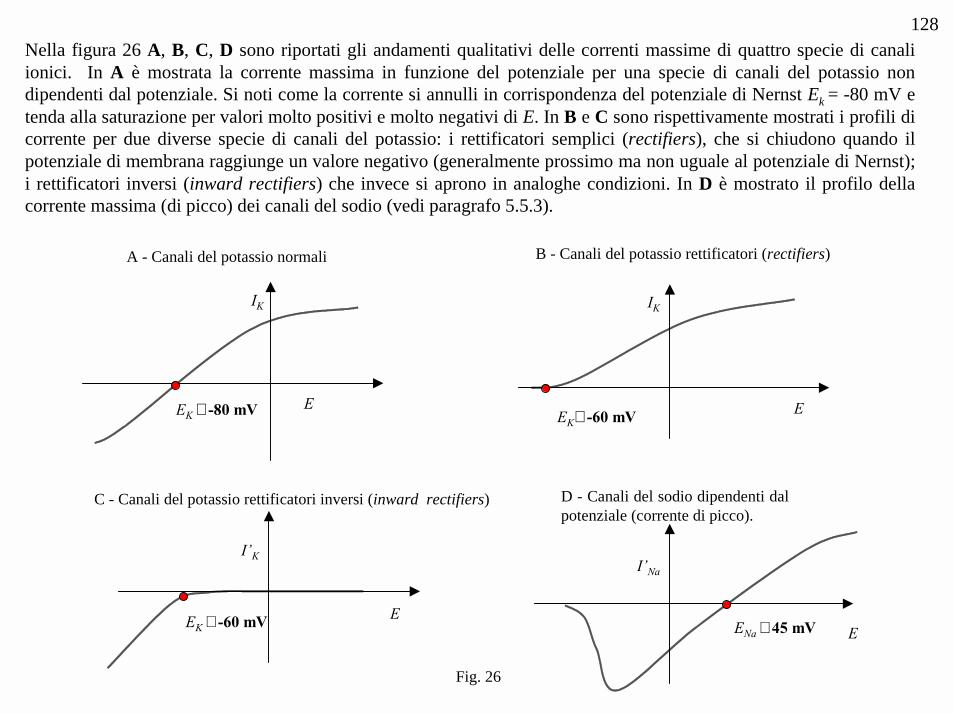
A - Canali del potassio normali
IK
EK ≅ -80 mV E
I’K
EK ≅ -60 mV E
C - Canali del potassio rettificatori inversi (inward rectifiers)
I’Na
ENa ≅ 45 mV E
D - Canali del sodio dipendenti dalpotenziale (corrente di picco).
IK
EK≅ -60 mV E
B - Canali del potassio rettificatori (rectifiers)
Fig. 26
128Nella figura 26 A, B, C, D sono riportati gli andamenti qualitativi delle correnti massime di quattro specie di canaliionici. In A è mostrata la corrente massima in funzione del potenziale per una specie di canali del potassio nondipendenti dal potenziale. Si noti come la corrente si annulli in corrispondenza del potenziale di Nernst Ek = -80 mV etenda alla saturazione per valori molto positivi e molto negativi di E. In B e C sono rispettivamente mostrati i profili dicorrente per due diverse specie di canali del potassio: i rettificatori semplici (rectifiers), che si chiudono quando ilpotenziale di membrana raggiunge un valore negativo (generalmente prossimo ma non uguale al potenziale di Nernst);i rettificatori inversi (inward rectifiers) che invece si aprono in analoghe condizioni. In D è mostrato il profilo dellacorrente massima (di picco) dei canali del sodio (vedi paragrafo 5.5.3).

(4) ( ) ( ).c aN N t N t= +
.k
c akN N
+
−
, .c aa c c a
dN dNk N k N k N k N
dt dt− + + −= − = −
( ) .aa a
dNk N N k N
dt + −= − −
6.5. Equazioni cinetiche dei canali ionici.
Consideriamo una popolazione di canali a due stati in una membrana cellulare. I canali transitano tra uno stato chiuso, chenon conduce corrente, a uno aperto che conduce. In questo modo essi modulano il flusso di corrente stazionaria attraversola membrana. La figura 27 mostra il tipico andamento della corrente elettrica di un singolo canale ionico in funzione deltempo. La corrente è interrotta dall’apertura e dalla chiusura dei canali e la statistica delle fluttuazioni di corrente riflettela statistica delle fluttuazioni dei canali. Il processo di apertura e chiusura si considera indipendente dal flusso di corrente.Ciò significa che il flusso di corrente non causa la transizione dei canali tra i due stati ma è semplicemente modulata daquel processo.
6.5.1. Le equazioni cinetiche a regime di transizione costante.Sia N il numero totale dei canali e siano Nc(t) e Na(t) rispettivamente i numeri dei canali chiusi e aperti al tempo t. Pertantoad ogni istante si ha
Nessun canale entra nello stato aperto senza aver lasciato lo stato chiuso, cioè N è costante. Sia k+ la costante cinetica della transizione tra uno stato chiuso e uno aperto e k- la costante cinetica tra uno aperto e uno chiuso. Allora il comportamento medio della popolazione di canali aperti e chiusi può essere descritta dallo schemacinetico
E’ chiaro che Nc sta per il numero probabile di canali chiusi al tempo t ed Na per il numero probabile si canali aperti altempo t; non il loro numero effettivo ma quello probabile. Poiché i canali che si aprono o si chiudono ad un certo istanteguadagnano solo gli uni dagli altri, o perdono gli uni dagli altri, la loro frequenza di transizione è governata dalleequazioni (Capitolo III).
Poiché i valori istantanei e quelli probabili di Na e Nc sono vincolati dalla(4), ciascuna di queste equazioni può essere scritta soltanto per Na o per Nc .Per esempio la seconda diventa
129
Fig.27
5 pA

( )(5) .aa
dNk k N k N
dt + − ++ + =
[ ] [ ] 0 0 0
1(7) ( ) ( ) exp ( ) / 1 exp ( ) / , .a aN t N t t t k N t t
k kτ τ τ τ+
+ −
= − − + − − − =+
[ ] [ ]0 0(8) ( ) ( ) exp ( ) / .a aN t N N t N t t τ∞ ∞− = − − −
( )(9) ( ) 1 exp / .aN t N t τ∞= − −
Seguiremo il comportamento dei canali aperti perché in definitiva siamo interessati alla statistica della corrente ionicache passa attraverso essi. La precedente equazione può essere risolta rispetto a Na. Riarrangiando i termini si trova
La soluzione generale di questa equazione è
E’ evidente che per t → ∞, Na(t) → τ k+N. Questo limite, che indicheremo con N∞, rappresenta il valore medio deicanali aperti per tempi lunghi. La (7) può anche essere scritta nella forma più economica
Supponiamo che all’istante iniziale t0=0 tutti i canali siano chiusi, cioè Na(0) = 0, allora il numero medio dei canali apertievolve secondo la legge illustrata nella figura 28
La figura 29 mostra come evolvono le popolazioni dei canali aperti nei casi in cui il numero di canali aperti inizialisiano maggiori (linea blu) o minori (linea rossa) del valore limite N∞.
Fig. 28 Fig. 29
130
N∞
N(t)
tτ
N∞
N(t)
t
N-
N+

6.5.2. Le equazioni cinetiche a regime di transizione variabile.Nel paragrafo precedente i coefficienti cinetici k+ e k- sono stati assunti rigorosamente costanti. In realtà essi possonodipendere da vari fattori come il potenziale di membrana E o la concentrazione di certe specie ioniche o molecolari.Poiché in generale E dipende a sua volta dalla cinetica di apertura e chiusura dei canali, le equazioni cinetiche nei casiconcreti devono essere incorporate in un più ampio sistema di equazioni che governano le variazioni temporali di tuttele grandezze in gioco. In questo paragrafo studieremo come k+ e k- possono dipendere da E nel caso che i canaliionici siano provvisti di una carica “portinaia” (gating charge) che influisce sulla loro probabilità di apertura e dichiusura. Come è stato già spiegato nel materiale integrativo Il modello di Iwasa, abbinato al Capitolo III, in seguitoad una variazione del campo elettrico di membrana, la carica portinaia di una proteina modifica le altezze delle barrieredi potenziale che separano le diverse configurazioni della proteina. Nel caso dei canali ionici, queste configurazionisono gli stati di apertura e chiusura del canale. Nei casi più semplici la teoria di Eyring (Capitolo III) prevede che icoefficienti cinetici a temperatura costante dipendono dal potenziale di membrana E secondo equazioni del tipo
( ) ( )0 0exp ; exp ;k k E k k Eα α+ + + − − −= − = −
dove k0+ , k0- e α+, α- sono indipendenti da E. La (5), riscritta per la frazione dei canali aperti w = Na/N, assume laforma 1
(10) ; , .dw w w k
wdt k k k k
ττ
∞ +∞
+ − + −
−= = =+ +
dove w∞ è il valore di w per t → ∞. Ponendo
( ) ( ) ( )( ) 0
0 0 0
1 tanh ( ) / 21 1(11) .
21 / exp 1 exp ( )
E Ew
k k E E E
α αα α α α
+ −∞
− + + − + −
+ − − = = =+ − + − −
( ) ( )( )
( ) ( )
( )
max0
0 0 0 0
0 max 0 0
0 0
exp / 21(12) ,
exp exp cosh ( ) / 2 cosh ( ) / 2
1 dove ; exp / 2 .
2
E
k E k E E E E E
Ek k
α α ττ τα α α α α α
τ τ τ α α
+ −
+ + − − + − + −
+ −+ −
+ = = ≅− + − − − − −
= = +
( )0 00
ln /k kE
α α+ −
+ −
=−
possiamo scrivere
131

In figura 30 A e B sono rappresentati i profili di w e τ/τ0 in funzione di E nei due casi: α+> α- e α+< α- in cui i canalirispettivamente si aprono o si chiudono quando la membrana cellulare viene depolarizzata. Il comportamentodescritto in A è caratteristico, ad esempio, dei canali del potassio rettificatori; quello descritto in B è caratteristico deicanali del potassio che rettificano in senso inverso (inward rectifiers). L’aspetto importante da tenere presente è chel’attivazione e la disattivazione non sono istantanee ma richiedono tempi τ che raggiungono i loro valori massimi τ0
quando la frazione dei canali aperti è circa 1/2. I valori tipici di τ0 variano da una frazione di ms ad alcuni ms.
Fig. 306.5.3. I canali del sodio.I canali del sodio hanno caratteristiche molto simili in tutte le cellule degli animali e la loro cinetica è più complessadi quella descritta nel precedente paragrafo.
Fig. 31
E
INa
Da un punto di vista fenomenologico, la corrente INa deicanali del sodio che viene attivata in risposta ad un gradinodel potenziale E presenta la dinamica qualitativa illustrata inFig.31. La corrente sale rapidamente per circa 0.5 ms, ma poidecade verso valori piccoli in un tempo dell’ordine di 5 ms.Potremmo dire che la corrente si comportaapprossimativamente come la derivata del potenziale.L’analisi comparativa del comportamento di questi canali invarie condizioni è consistente con l’esistenza di un doppiomeccanismo di attivazione-disattivazione.
t (msec)5
132

Nella figura 32 il profilo di attivazione wA e quello di disattivazione wD
di un canale del sodio (curve rosse), insiemi ai rispettivi tempi relatividi apertura τA/τ0A e τD/τ0D (curve blu), sono rappresentati nello stessodiagramma. I due meccanismi operano in serie, giacché la riattivazionedi un canale precedentemente attivato-disattivato ha luogo solo dopoche la disattivazione ha compiuto il suo corso. Ciò significa che ladisattivazione rende temporaneamente refrattario il canale rispetto apossibili attivazioni successive. La figura mostra anche come le curve wA e wD, essendo parzialmentesovrapposte in un intorno di E ~ -50 mV (area tratteggiata), creanouna piccola finestra di conduttanza. Ciò significa che per valori di Eintorno a -50 mV la disattivazione fa decadere INa verso valori piccoli(circa 0.05 volte il valore del picco), invece che verso valori nullicome indicato dalla Fig. 31. Nella figura 33 sono rappresentati i profili tipici della corrente delsodio in corrispondenza a vari gradini di potenziale applicati attraversola membrana cellulare di una cellula cardiaca. Per maggiore chiarezzasi tenga presente che il profilo della corrente del sodio in funzione delpotenziale rappresentato nella figura 26 D riguarda la corrente dipicco non quella stazionaria. Il duplice processo di attivazione veloce e disattivazione lenta deicanali del sodio svolge un ruolo importante nella generazione deipotenziali d’azione dei neuroni: una piccola depolarizzazioneimprovvisa della membrana attiva l’apertura del canale facendo primadepolarizzare ulteriormente e poi ripolarizzare in senso inverso lamembrana. La disattivazione della corrente del sodio fa in modo che laquantità di sodio che entra durante la fase di attivazione sia piccola. Seciò non accadesse la membrana si porterebbe a valori positivi delpotenziale facendo rimanere i canali del sodio sempre aperti.
Fig. 33
133
Fig. 32

6.5.4. Aspetti particolari della cinetica dei canali ionici.La permeabilità di un canale alle diverse specie ioniche e la sua dipendenza dal potenziale di membrana possono esserericondotte allo studio dei sistemi mesoscopici (Capitolo III) e in particolare delle barriere di potenziale che condizionanoil transito degli ioni. Il coefficiente cinetico che descrive la probabilità di apertura del canale del sodio presenta unadipendenza dal potenziale di membrana più ripida di quella che potrebbe essere spiegata con la transizione dello ioneattraverso un solo complesso di attivazione. Inoltre è stato constatato che l’aumento della concentrazione degli ioniidrogeno ha l’effetto di bloccare il canale (Woodhull, 1973). Questo fatto può essere spiegato assumendo che ilpassaggio dello ione sodio sia condizionato dalla presenza della carica negativa di un radicale acido (-COO-) che puòessere neutralizzata da un protone (Fig. 34). Uno studio del comportamento del canale rispetto a ioni di specie diversa haportato Bertil Hille (1975) a proporre il modello rappresentato in figura 34. Per passare dal fluido extracellulare (out) aquello intracellulare (in), lo ione attraversa una sequenza di barriere di varia altezza (Fig.35).
Fig.34
134
Fig.35

K K K K Ca Ca Ca Ca L L L L(13) ( ), ( ), ( ),I g w E E I g w E E I g w E E= − = − = −
6.6. Membrane eccitabili.
Membrane cellulari dotate di canali ionici dipendenti dal potenziale e con potenziali di Nernst di segno oppostopossono dar luogo a fenomeni di eccitabilità elettrica, che si manifesta con la spontanea generazione di variazioniimpulsive o periodiche del potenziale di membrana. Di questo tipo sono le membrane degli assoni neuronali cheprovvedono alla trasmissione dell’informazione nervosa. I meccanismi che generano questi potenziali sono statichiariti da Hodgkin e Huxley in una memorabile serie di lavori pubblicati nel periodo 1939-1952. Questi autori hannodimostrato come i canali del sodio e del potassio distribuiti sulle membrane degli assoni neuronali cooperino perprodurre i potenziali di azione. Le equazioni che governano la dinamica di queste membrane sono state costruite sullabase di osservazioni sperimentali riguardanti il comportamento delle correnti ioniche attraverso la membranadell’assone gigante del calamaro. Membrane eccitabili di tipo non neuronale si trovano ad esempio nelle fibremuscolari dei crostacei. Esse coinvolgono i canali di potassio e calcio dipendenti dal potenziale. Uno studio estensivosull’eccitabilità delle membrane è dovuto a Hagiwara e Naka (1964). Poiché i meccanismi che determinanol’eccitabilità di queste membrane sono più semplici di quelli delle membrane assoniche, cominciamo col presentareun modello che descrive il funzionamento di questo tipo di membrana.
6.6.1. Il modello di Morris-Lecar.Le fibre muscolari muscolari di un mollusco come il cirripede sono ricche principalmente di canali di calcio epotassio. Illustriamo qui come un semplice modello proposto da Morris e Lecar nel 1981 spieghi abbastanza bene lagenerazione di potenziali d’azione in questo tipo di cellule. Si assume che le equazioni che governano la dipendenzadella corrente elettrica totale I dal potenziale E siano riferibili alle correnti specifiche di tre tipi di canale: la correntedei canali del potassio IK; la corrente dei canali del calcio ICa; la corrente di perdita (leakage) Il della membrana, cherappresenta l’effetto medio di eventuli altri canali di membrana. La dipendenza di queste correnti dal potenziale dimembrana può scriversi con tutta generalità nella forma
dove gK, gCa, e gL sono rispettivamente le conduttanze massime dei canali del potassio, del calcio e dei fattori diperdita, wK, wCa e wL le rispettive frazioni dei canali aperti, ed EK, ECa, EL i rispettivi potenziali di Nernst. Si assumeche i canali del calcio siano attivati più velocemente di quelli del potassio e che i fattori di perdita non dipendano dalpotenziale.
135

Supporremo anzi per semplicità che il tempo di attivazione dei canali del calcio sia trascurabile rispetto a quello deicanali del potassio. Ciò significa che, in conformità con le (11) e (12) le frazioni dei canali aperti possono essereapprossimativamente scritte nella forma
( )( )
( )
3 4 K0K K KK K
K 3 4
1 2Ca Ca L L
1 tanh /(14) , dove e ,
2 cosh /
1 tanh /(15) ; 1.
2
E E Edw w ww
dt E E E
E E Ew w w w
τττ
∞∞
∞ ∞
+ − − = = ≅−
+ − ≅ = ≅ ≅Possiamo applicare queste assunzioni al modello di cellula rappresentato nella figura 36. Il sistema è costituito da tregeneratori specifici di forza elettromotrice, che rappresentano i potenziali Nernst, di conduttanze rispettive gK wK, gCa
wCa , gL, in parallelo alla capacità di membrana C. Il potenziale di membrana in condizioni stazionarie è regolato daun generatore di corrente esterno che fa passare attraverso una micropipetta una corrente costante I0. La corrente totaleche attraversa la membrana è governata dalla seguente equazione
K K K Ca Ca Ca L L 0(16) + ( ) ( ) ( ) ,dE
C g w E E g w E E g E E Idt ∞− + − + − =
dove CdE/dt = IC è la corrente capacitiva. Questa equazione,unitamente alle (14) e (15) formano un sistema dinamicoche evolve nel tempo in vari modi a seconda dei valori deiparametri. In certe condizioni possono aver luogo bruschevariazioni dei potenziali di membrana che vanno sotto ilnome di potenziali d’azione. In questo caso si dice che lemembrane sono eccitabili. La generazione dei potenziali di azione corrispondeall’esistenza di soluzioni instabili del sistema di equazioni.L’instabilità nasce dal fatto che combinando le correnti dicalcio e potassio si produce un profilo di corrente che percerti valori di I0 e durante certi intervalli temporali presentauna pendenza negativa in una regione del potenziale dimembrana.Fig.36
136
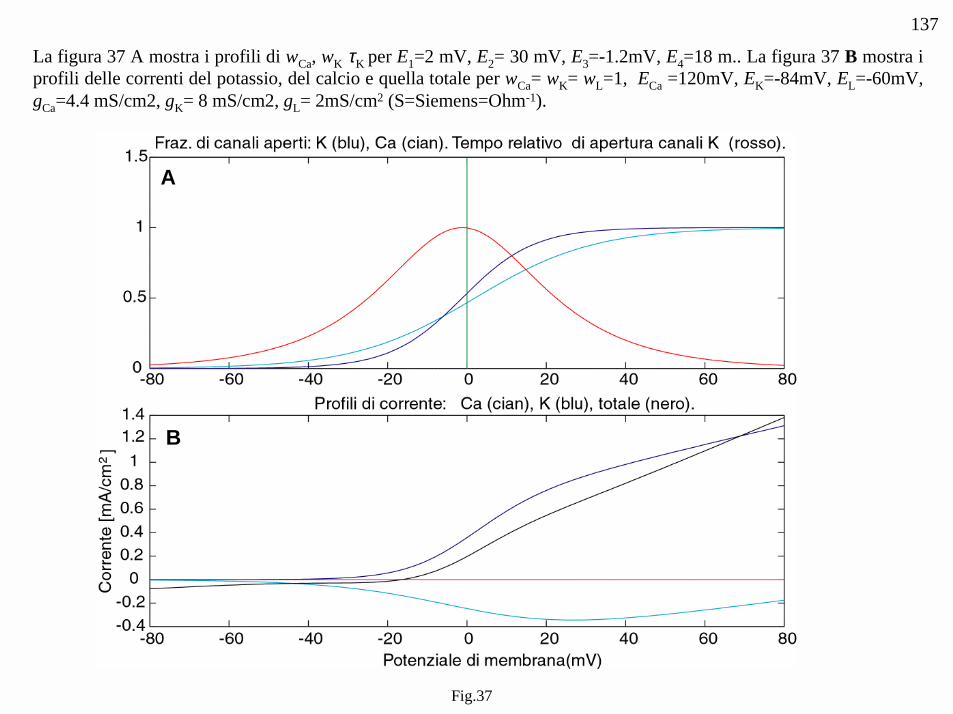
Fig.37
La figura 37 A mostra i profili di wCa, wK τK per E1=2 mV, E2= 30 mV, E3=-1.2mV, E4=18 m.. La figura 37 B mostra iprofili delle correnti del potassio, del calcio e quella totale per wCa= wK= wL=1, ECa =120mV, EK=-84mV, EL=-60mV,gCa=4.4 mS/cm2, gK= 8 mS/cm2, gL= 2mS/cm2 (S=Siemens=Ohm-1).
137
A
B

Nella figura 38 sono rappresentati i profili della corrente per τ0 = 25 msec e I0 = 35 microampere/cm2 ottenutimediante una simulazione con Matlab (routine MORLEC.M).
6.6.2. Il modello di Hodgkin-Huxley.Il modello di di HH (Hodgkin-Huxley) ha rappresentato una pietra miliare della fisiologia del sistema nervoso. Esso èstato elaborato da questi autori in seguito ad una indagine sistematica della membrana dell’assone gigante delcalamaro (diametro ≅ 0.5 mm, lunghezza = alcuni decimetri) in un periodo in cui non si sapeva nulla sui canaliionici, la cui esistenza viene ipotizzata proprio sulla base delle ricerche di HH. Attraverso una serie di esperimenti HHhanno dimostrato in primo luogo che la membrana conduce principalmente due tipi di corrente, quella del potassion equella del sodio. In seguito essi hanno cercato di stabilire come queste correnti variano quando al membrana èsottoposta a variazioni di potenziale. Il sistema di equazioni cinetiche proposto da questi autori caratterizza ladinamica elettrica della membrana dell’assone nervoso in condizioni varie e spiega la formazione dei potenzialid’azione interpretando con precisione le loro caratteristiche morfologiche. Si può dire che le equazioni di HH,determinate essenzialmente in modo empirico, hanno indicato quali sono le proprietà dei canali ionici.
Fig.38
138

Lo schema della membrana di HH è lo stesso di quello rappresentato in Fig.36 con “Na” al posto del simbolo“Ca”. Nel modello di HH i canali del potassio (K) possiedono la cinetica tipica di un canale a due stati attivato dapotenziali crescenti. Nel seguito la frazione dei canali K attivati al tempo t sarà indicata con n(t), quella stazionariacon n∞, il tempo di attivazione con τn. Naturalmente n∞ e τn dipendono dal potenziale (Fig.39 A). I canali del sodio(Na) possiedono una cinetica più complessa caratterizzata da un regime di attivazione veloce e uno di disattivazionelenta. Nel seguito la frazione dei canali Na attivati al tempo t sarà indicata con m(t), quella stazionaria con m∞, iltempo di attivazione con τm (Fig.39 B) ; la frazione dei canali Na disattivati al tempo t sarà indicata con h(t), quellastazionaria con h∞, il tempo di attivazione con τh (Fig.39 C) . L’analisi della dipendenza temporale dei processi di attivazione e disattivazione dei canali del potassio e del sodiohanno portato HH alla conclusione che la corrente K è proporzionale a n4(t) menre quella Na è proporzionale am3(t)h(t). Pertanto le equazioni HH hanno la forma
( ) ( ) ( )4 3K K Na Na L L 0 ,
0, 0, 0, n m h
dEC g n E E g m h E E g E E I
dtdn n n dm m m dh h h
dt dt dtτ τ τ∞ ∞ ∞
+ − + − + − =
− − −+ = + = + =
dove
[ ] [ ] [ ]1 tanh ( ) / 1 tanh ( ) / 1 tanh ( ) /; ; ;
2 2 2m m h h n nE E E E E E E E E
m h n∞ ∞ ∞
+ − ∆ + − ∆ + − ∆= = =
[ ] [ ] [ ]0 0 0
1 0.85 10.2 ( ) ; 0.15 ;
cosh ( ) / cosh ( ) / cosh ( ) /m m h h n nm m h h n n
m EE E E E E E E E E
τ τ τ τ τ τ = + = + = − ∆ − ∆ − ∆
Le espressioni dei tempi di attivazione e disattivazione sono state determinate empiricamente in modo dacorrispondere con buona approssimazione ad alcuni diagrammi riportati nel testo di Hille Ionic Channels ofExcitable Membranes (vedi routine Matlab HH.M).
139

140
Fig.39
A B C
D
E

141
Il pannello E della figura 39 mostra i potenziali d’azione evocati da una successione di piccoli impulsi di correnteesterna Io (linea rossa in basso). Il pannello D della stessa figura mostra come variano in corrispondenza le frazionidi canali aperti; la linea nera e quella blu rappresentano rispettivamente le frazioni dei canali K e Na attivati durantela formazione dei potenziali d’azione, mentre la linea rossa rappresenta la frazione di canali disattivati. Nel pannello E si può notare come un primo stimolo di corrente di ampiezza pari a 50 µA/cm2 e durata pari a0.25 ms non sia sufficiente ad innescare il potenziale d’azione (la corrente è rapportata a un cm2 di membranacellulare). I potenziali d’azione sono innescati invece dai successivi stimoli di ampiezze progressive pari a 75, 100,125 µA/cm2, tutti di durata pari a 0.25 ms, purché questi siano sufficientemente intervallati nel tempo. Lo stesso pannello mostra come uno stimolo che sia applicato troppo presto, prima che il processo didisattivazione sia completato, non riesca ad innescare il potenziale d’azione. Questo effetto è evidente incorrispondenza dello stimolo applicato subito dopo il terzo picco del potenziale d’azione. Si può notare comequesto stimolo inefficace capiti mentre il profilo della percentuale di disattivazione (pannello D, linea rossa) non haancora raggiunto il suo valore massimo. Questo spiega il fenomeno del periodo dei refrattarietà che caratterizza ilmeccanismo di sparo dei neuroni e che persiste per alcuni ms dopo la generazione del potenziale d’azione. Essospiega anche il fatto che la frequenza di sparo dei neuroni ha un valore massimo corrispondente al tempo direfrattarietà. Nella porzione di destra del pannello E (dopo la linea verticale rossa) sono simulati gli effetti di una sequenza distimoli di ampiezze decrescenti e di durata pari a 0.5 ms. Si può osservare come, in corrispondenza dell’ultimopicco del potenziale d’azione, uno stimolo di ampiezza pari a 50 µA/cm2 sia ora sufficiente ad innescare unpotenziale d’azione mentre uno di ampiezza pari a 25 µA/cm2 produca un effetto simile a quello di 50 µA/cm2 e0.25 ms applicato all’inizio della sequenza. Questo comportamento indica che la risposta della membrana èall’incirca proporzionale all’integrale temporale della corrente, in altri termini alla quantità di carica trasferita dagliimpulsi di corrente. Assieme alle routine in Matlab MORLEC.M e HH.M è disponibile la routine HH2.M che contiene una secondaversione del modello di HH. Questa seconda implementazione è stata parametrizzata con altri dati forniti dagli stessiHH e descritti in Keener e Sneyd (1998). La routine simula la generazione ripetitiva di potenziali d’azione diampiezza piuttosto elevata. Informazioni più dettagliate su questi modelli si trovano nei commenti delle routine menzionate.

Bibliografia• Aidley, D.J. e Stanfield, P.R. (1996) Ion channels - Molecules in Action. Cambridge University Press, UK.• De Felice, L. (1981) Introduction to Membrane Noise. Plenum Press, New York.• De Robertis, E.D.P, Nowinski, W.W. e Saez, F.A. (1972), Biologia della Cellula, Vol. I eII, Zanichelli, Bologna.• Fall, C.P., Marland E.S., Wagner, J.M. e Tyson J.J. Ed.s (2002) Computational Cell Biology, Springer-Verlag, NY.• Johnston, D. e Miao-Sin-Wu, S. (1995) Foundations of Cellular Neurophysiology. The MIT Press, USA.• Hille, B. (2001) Ionic Channels of Excitable Membranes. Terza edizione, Sinauer Associated Inc. Sunderland.• Keener, J. e Sneyd J. (1998) Mathematical physiology. Springer-Verlag, NY.• Tuszynski, J.A. e Kurzynski, M. F. (2003), Introduction to Molecular Biophysics. CRC Press LLC, USA.
142












![introduzione metabolismo [modalità compatibilità]tpall2015.altervista.org/alterpages/files/introduzionemetabolismo.pdf · glicoproteinee cerebrosidi 3. funzione disintossicante:](https://static.fdocumenti.com/doc/165x107/5c65f56409d3f2d0218b90f2/introduzione-metabolismo-modalita-compatibilita-glicoproteinee-cerebrosidi.jpg)




