
BIOTECNOLOGIE PER L’ENERGIA E LA...
45
Biotecnologie per l’energia e la chimica 1 BIOTECNOLOGIE PER L’ENERGIA E LA CHIMICA di: Roberto Avella Giovanni Giuliano Vito Pignatelli con il contributo di: Annamaria Bevivino Luigi Chiarini Nicola Colonna Claudia Dalmastri Rosella Franconi Giulio Izzo Silvia Tabacchioni 27/4/2006
Transcript of BIOTECNOLOGIE PER L’ENERGIA E LA...

Biotecnologie per l’energia e la chimica
1
BIOTECNOLOGIE PER L’ENERGIA
E LA CHIMICA
di:
Roberto Avella
Giovanni Giuliano
Vito Pignatelli
con il contributo di:
Annamaria Bevivino
Luigi Chiarini
Nicola Colonna
Claudia Dalmastri
Rosella Franconi
Giulio Izzo
Silvia Tabacchioni
27/4/2006

Biotecnologie per l’energia e la chimica
2
Indice dei contenuti
Riassunto p. 3 1. Lo scenario energetico p. 4 2. Il ciclo della CO2 p. 4 3. Caratteristiche strutturali dell’agricoltura italiana p. 5 4. Le piante come fonti di combustibili rinnovabili: biomasse e biocarburanti p. 6 5. Le piante per la “green chemistry” p. 9 6. Le biotecnologie e la transizione ad una società ad emissioni zero p. 9 7. Le microalghe come fonte energetica p. 10 8. La produzione biologica di idrogeno p. 12 9. La biodiversità microbica p. 14 10. Il posizionamento ENEA p. 15 11. Assi di ricerca strategici p. 15 11.1 Environmental systems biochemistry p. 15 11.2 Biomasse e biodiesel da piante p. 16 11.3 Sequestrazione biologica di CO2, biomasse e biodiesel da microalghe p. 17 11.4 Idrogeno biologico p. 18 12. Roadmap p. 20 13. Tabelle e Figure p. 22 14. Bibliografia p. 44

Biotecnologie per l’energia e la chimica
3
Riassunto
Il continuo aumento del prezzo dei combustibili fossili e l’impatto negativo sull’ambiente prodotto dalle emissioni antropiche di CO2 impongono approcci di ricerca nuovi, finalizzati a sostituire i combustibili e le materie prime per l’industria chimica di origine fossile con fonti rinnovabili, sequestrare la CO2 e consentire nel lungo periodo la transizione a vettori energetici nuovi, come l’idrogeno. Appare evidente che in Italia le biomasse e i biocombustibili di origine agricola e forestale possono contribuire in misura ridotta (<10%) al raggiungimento di questi obiettivi. Di conseguenza, occorre rivolgere la propria attenzione a nuove potenziali fonti energetiche rinnovabili, come quelle rappresentate dalle microalghe. Questo documento propone una “roadmap” di ricerca di 25 anni articolata su 4 assi di ricerca strategici: 1. Environmental systems biochemistry, finalizzato ad esplorare ed utilizzare la biodiversità genomica e chimica ambientale, biodiversità che rimane tuttora in massima parte inesplorata. 2. Biomasse e biodiesel da piante, finalizzato a sostituire, entro il 2015, il 3% dei combustibili fossili e il 6% dei carburanti per autotrazione con combustibili di origine agroforestale economicamente competitivi. 3. Biomasse e biodiesel da microalghe, finalizzato a sostituire, entro il 2030, il 20% dei combustibili fossili, il 30% della materia prima per l’industria chimica ed il 60% dei carburanti per autotrazione con fonti di origine microalgale, economicamente competitive, riducendo del 50% le emissioni di CO2 di origine antropica e depurando il 50% delle acque reflue urbane o agricole. 4. Idrogeno biologico, finalizzato a sostituire, entro il 2030, il 20% del carburante per autotrazione con idrogeno di origine biologica, economicamente competitivo.

Biotecnologie per l’energia e la chimica
4
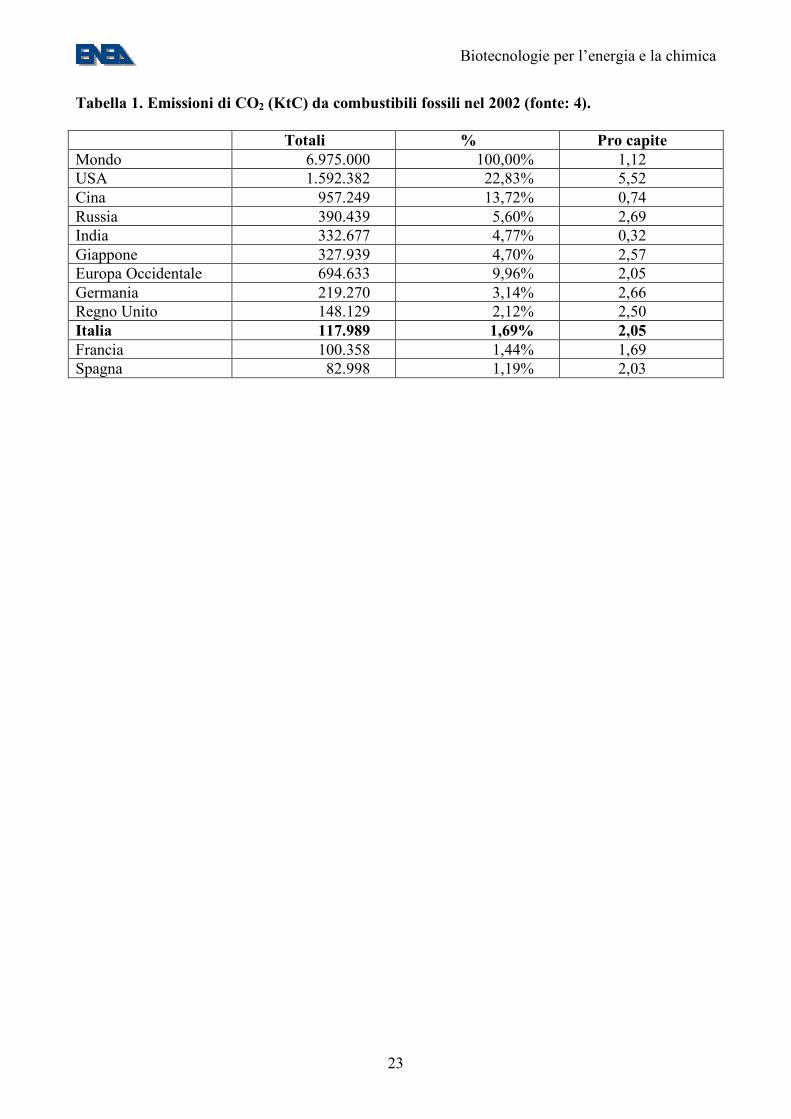
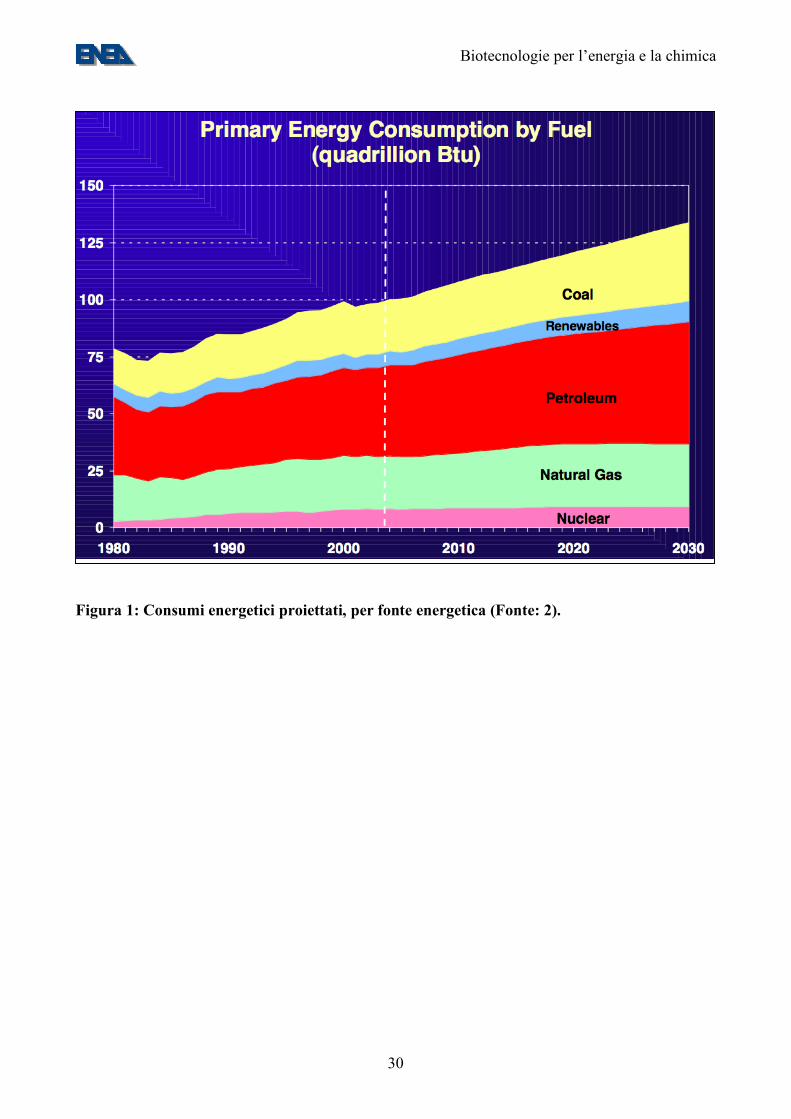
1. Lo scenario energetico La produzione di energia a livello mondiale si basa, in gran parte, sull’uso di combustibili fossili, fra cui un ruolo di spicco è giocato dal petrolio. Nella “Energy Outlook and Modeling Conference”, tenutasi nel Marzo 2006 a Washington DC ed organizzata dalla Energy Information Administration del DOE (1) è stato stimato che i consumi attuali sono di circa 100 “Quads”, di cui oltre l’80% da fonti (petrolio, carbone, gas) che producono CO2 (Figura 1) (2). 1 Quad corrisponde a 1015 Btu, ovvero circa 25 milioni di tonnellate equivalenti di petrolio (MTep) – cfr. Appendice 1. Le riserve attuali di petrolio, di 1.277 Mld di barili, sarebbero in grado di fornire 42 anni di copertura ai ritmi attuali di consumo e, nel medio periodo, è atteso che i consumi di petrolio cresceranno al ritmo di 1,6% annuo (3). Il consumo italiano di combustibili fossili nel 2001 è stato stimato in circa 160 MTep/anno, di cui circa 40 MTep/anno per i trasporti e 50-60 MTep/anno per la generazione di energia elettrica (4). 2. Il ciclo della CO2 Le emissioni mondiali di CO2 da attività umane nel 2002 sono state stimate in 6,9 GtC/anno, ovvero 1,12 tC/anno pro capite (Tabella 1) (5). Le emissioni italiane sono state stimate in 117,9 MtC/anno, ovvero 2,05 tC/anno pro capite. Questo valore è nella media Europea, anche se è decisamente superiore a quello di Paesi, come la Francia, che hanno investito nel nucleare (Tabella 1) (5). Secondo stime del DOE degli USA, che sono il maggiore produttore mondiale di CO2, le emissioni antropiche sono destinate a salire nel 2030 a 7.3 GtC/anno (scenario ottimistico) o 9.0 GtC/anno (scenario pessimistico) (2). Le emissioni antropiche costituiscono solo una piccola frazione della quantità totale di carbonio (210 GtC) che va in ciclo ogni anno sulla Terra, primariamente ad opera di fenomeni biologici come la fotosintesi clorofilliana e la respirazione. La captazione della CO2 ad opera della fotosintesi terrestre e marina ammonta, ripettivamente, a 90 e 120 GtC/anno (Figura 2). Di questi 210 GtC totali, 2-3 sono fissati in depositi a lungo termine e 207-208 sono riemessi, primariamente ad opera della respirazione e della decomposizione, ma anche della combustione e della deforestazione (Figura 2) (6,7). Il principale “sink” della CO2 antropogenica sono gli oceani, a causa della sedimentazione, in depositi a lungo termine, di una parte del carbonio fissato dalla fotosintesi marina. Si calcola che gli oceani fissino, da soli, da 1/3 (Figura 2) alla metà (8) della CO2 antropogenica. Si prevede che l'aumento della concentrazione atmosferica di anidride carbonica dovuta alle emissioni antropiche condurrà a cambiamenti significativi nel clima. Circa metà delle emissioni attuali di CO2 sono assorbite dai “sink” acquatici e terrestri, ma questo assorbimento è sensibile al clima e quindi alle stesse concentrazioni atmosferiche di CO2, creando un “feedback”. Esistono diversi modelli sull’evoluzione del clima, che tenere o non tengono o no conto degli effetti di questo “feedback”. Uno dei modelli che ne tiene conto predice che, intorno all’anno 2050, l’ecosistema terrestre si trasformerà da un debole “sink” ad una importante “source” di CO2 (principalmente a causa della progressiva scomparsa delle foreste pluviali tropicali e all’aumento della respirazione nel suolo in seguito del cambiamento del clima), risultando in un aumento della CO2 dalle 290 ppm dell’era pre-industriale a oltre 900 ppm nel 2100. Secondo il modello, tale aumento risulterebbe in aumenti catastrofici delle temperature medie, di +5°C a livello globale e di +8°C negli ecosistemi terrestri (9).

Biotecnologie per l’energia e la chimica
5
Qualunque sia il modello da prendere in considerazione, il livello di incertezza nella stima degli effetti dell’aumento della CO2 sul clima è ancora estremamente alto. Questa incertezza rende probabili scenari secondo cui, per stabilizzare il clima, si debba arrivare entro pochi decenni a ridurre quasi a zero la produzione antropogenica di CO2 (10). 3. Caratteristiche strutturali dell’agricoltura italiana L’Italia è un paese il cui territorio è più del 50% collocato in collina od in montagna. In tale condizione l’agricoltura assume una funzione che non è solo quella di produrre derrate alimentari – per le quali l’Europa risulta eccedentaria – ma, più che in altre nazioni Europee, di salvaguardia del territorio. L’Italia soffre di una crisi dell’economia rurale che genera dissesto territoriale dal punto di vista idrogeologico. Questo è imputabile anche all’abbandono del patrimonio boschivo, principalmente di quei 5 Mha dedicati al bosco ceduo. Il bosco ceduo, nato in epoca romana per sopperire a precise esigenze energetiche, ha la caratteristica di generare biomassa fissando CO2 se sottoposto a tagli periodici, ma di perdere questa caratteristica se alcuni turni di taglio vengono saltati, trasformandosi dapprima in un debole “sink” e, in casi estremi, in una “source” di CO2. Lo stato di abbandono del patrimonio boschivo è dovuto principalmente alla perdita di interesse della proprietà, costituita da un gran numero di piccoli proprietari detentori in media di pochi ettari. In Italia esistono circa 1.800.000 aziende agricole (2003, Universo statistico CEE che esclude le aziende piccolissime). Si stima che le aziende presenti sul mercato siano solo circa 600.000, prevalentemente localizzate in pianura e collina. La Superficie Agricola Utilizzata (SAU) complessiva italiana è diminuita fortemente negli ultimi decenni e ora si è assestata vicino ad un valore pari a 12,2 milioni di ettari. La superficie aziendale totale media (SAT) è pari a circa 8 ha mentre quella effettivamente utilizzata e sottoposta a coltura (SAU) è limitata e pari a circa 6,2 ha (dato inferiore alla media europea). Prevalgono aziende di dimensioni piccole o piccolissime. Le aziende con una superficie utilizzata superiore a 50 ha sono solo il 2% del totale (dato 2003). L’estrema eterogeneità delle condizioni pedoclimatiche sulle quali viene esercitata l’attività agricola rende difficile da una parte proporre “ricette” valide per tutto il paese dall’altra limita il numero di bacini o distretti agroenergetici sui quali avviare attività estese nel campo delle bioenergie. Ogni considerazione sull’agricoltura italiana che conosciamo deve però confrontarsi con il fatto che l’introduzione dell’ultima riforma della PAC sta modificando fortemente lo scenario della nostra agricoltura tanto che ci si attendono forti discontinuità nei dati statistici. In Italia, nel corso del 2005, è entrata in vigore la nuova PAC introducendo da subito il disaccoppiamento totale (ovvero disaccoppiando completamente il contributo ricevuto dall’azienda agricola e la produzione effettiva) mentre in Spagna e Francia l’introduzione è stata rinviata di un anno sfruttando i margini di flessibilità concessi dalla riforma della PAC. Nelle condizioni di incertezza in cui si sono trovati gli agricoltori, dato che ormai il contributo risulta svincolato da cosa effettivamente coltivano, si è assistito a dei profondi cambiamenti delle superfici investite. Ad esempio, nel giro di due anni la superficie investita a grano duro è diminuita secondo le stime ISMEA di un terzo, da 1.772.000 ha nel 2004 a 1.118.000 nel 2006.

Biotecnologie per l’energia e la chimica
6
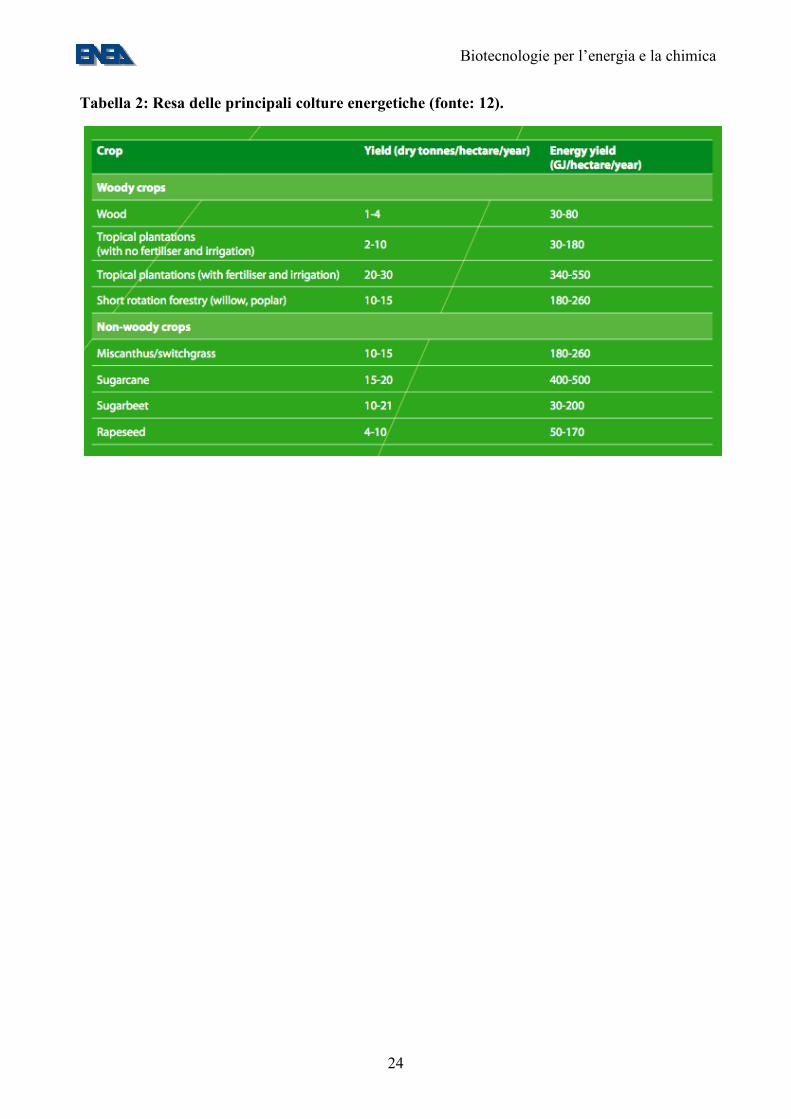
A ciò si associa la crisi del settore bieticolo saccarifero, con l’accordo europeo che ha portato nel corso di questo anno alla chiusura di 13 dei 19 zuccherifici italiani con un forte ridimensionamento delle superfici investite e quindi una disponibilità potenziale di terreni da dedicare a nuove colture. La crisi dell’industria alimentare della trasformazione del pomodoro, per la competizione internazionale, ha portato gli agricoltori a rivedere le proprie scelte e a modificare i propri ordinamenti produttivi. I fatti elencati, ai quali dobbiamo aggiungere anche le difficoltà che incontra il settore della maiscoltura da granella, stanno provocando un profondo e rapido riassetto del settore dei seminativi nel nostro paese con una disponibilità potenziale di elevate quantità di terreni da dedicare alle colture energetiche. In questo momento non esiste conflitto tra usi diversi dei terreni anzi gli agricoltori cercano conferme e rassicurazioni per poter intraprendere strade nuove (colture energetiche e filiera no-food) dato che dispongono di seminativi lasciati incolti. 4. Le piante come fonti di combustibili rinnovabili: biomasse e biocarburanti Come detto sopra, la fotosintesi clorofilliana fissa CO2 dall’atmosfera, restituendola sotto forma di biomassa. La produttività massima delle principali colture energetiche è di circa 20 T/ha/anno (peso secco) (11). Tale biomassa è composta principalmente da lignina, cellulosa ed emicellulose. In alcuni casi (mais, canna e barbabietola da zucchero) i principali prodotti di riserva sono zuccheri semplici (saccarosio) o complessi (amido) fermentabili per la produzione di bio-etanolo. Infine, nelle colture oleaginose, come il colza, la soia, e la palma da olio, i principali composti di riserva sono i trigliceridi, materia prima per la produzione del biodiesel. La produttività energetica stimata/ettaro delle principali colture energetiche è riportata nella Tabella 2. Assumendo una produttività media di 200 Gj/ettaro/anno, e una radiazione solare incidente di 4 kWh/m2/giorno, la efficienza energetica di tali colture è circa lo 0,38%. Questa percentuale è una sovrastima, in quanto non tiene conto degli input energetici per la coltivazione. Con questa produttività ed un fattore di conversione di 42,41 Gj/Tep, per coprire completamente il fabbisogno energetico italiano, stimato in 160 MTep (4), servirebbero 33 milioni di ettari, ovvero il 270% della superficie coltivabile e il 110% del territorio nazionale. Appare quindi evidente che, allo stato attuale delle tecnologie, le biomasse di origine agricola possono coprire solo una piccola parte del nostro fabbisogno energetico. Un discorso diverso riguarda invece, le biomasse residue dalle attività agricole e forestali: tali biomasse, composte per la maggior parte di residui legnosi, paglia ed altri residui cellulosici, e in piccola parte di residui animali, se non utilizzate vengono comunque attaccate dai microorganismi, rientrando nella atmosfera sotto forma di CO2 o altri gas serra, come il metano. Questi residui, con l’esclusione di quelli agro-industriali, che sono in gran parte autoconsumati per le esigenze energetiche della fabbrica, vengono attualmente bruciati in campo, non contribuendo in alcun modo al bilancio energetico del Paese. Si calcola che tali residui, se utilizzati come fonti di energia, possono fornire, nel territorio della UE, circa 6,75 Ej/anno (circa 160 MTep/anno, ovvero il fabbisogno energetico di un Paese come l’Italia) (12). Tuttavia, a causa della loro distribuzione sul territorio, il loro utilizzo presenta una serie di problemi logistici. L’obiettivo fissato dalla UE è di recuperare per scopi energetici 1,5 di questi 6,75 Ej potenziali entro l’anno 2010 (12).

Biotecnologie per l’energia e la chimica
7
A livello italiano, i residui agricoli, al netto del materiale riciclato, ammontano a circa 12 MT/anno (37). Questa cifra è probabilmente una sottostima. Assumendo un contenuto medio di cellulosa ed altri polisaccaridi pari al 40% ed una efficienza di conversione a zuccheri fermentabili dell’80% (efficienza che, per essere raggiunta, richiede una serie di investimenti in ricerca) questo dato equivale a circa 3,8 MT di equivalenti di glucosio/anno. Un uso già oggi tecnologicamente maturo di tali biomasse è la loro combustione in piccole centrali (potenza media 5 MW) con co-generazione di elettricità e calore. Mentre il rendimento, in termini di energia elettrica, di tali piccole centrali è chiaramente inferiore a quello delle grandi centrali termiche e assai inferiore a quello delle moderne centrali a ciclo combinato, esse presentano il vantaggio di poter essere delocalizzate sul territorio, minimizzando i costi di trasporto della biomassa dal luogo di produzione alla centrale e offrendo migliori possibilità di utilizzo del calore generato per teleriscaldamento di abitati o serre e di fissazione della CO2 prodotta (la quale comunque, essendo derivata dalla fotosintesi, non contribuisce al bilancio di emissioni antropiche di CO2). E’ comunque necessario condurre studi che elenchino da una parte i costi energetici ed economici per la produzione di biomassa e il suo trasporto alla centrale e dall’altra le rese energetiche ed economiche dell’uso dei prodotti della sua combustione (elettricità, calore e ceneri di combustione). Tali studi, noti come “life cycle inventories”, debbono stabilire le rese energetiche del processo e il costo economico per MW elettrico e termico generato, rispetto all’analogo MW generato con combustibili fossili. Ovviamente, il costo della generazione con combustibili fossili andrebbe gravato del costo per la sequestrazione della CO2 prodotta, che, a causa delle considerazioni fatte nella sezione 2, diventerà nel medio periodo una necessità. I due principali biocarburanti per cui la tecnologia di produzione è ormai matura sono il bioetanolo ed il biodiesel. Il bioetanolo è prodotto tramite fermentazione diretta da colture ad elevato tenore zuccherino (barbabietola, canna da zucchero) o tramite conversione dell’amido di colture amidacee (mais, grano) a zucchero prima di fermentarlo, seguita da distillazione e disidratazione. Questi ultimi processi richiedono una grande quantità di energia, che è la causa principale delle scarse rese energetiche del bioetanolo europeo e statunitense. In Brasile, il bioetanolo è principalmente prodotto da canna da zucchero, negli USA da mais, in Europa da barbabietola a grano. Il bioetanolo può essere ulteriormente trasformato in ETBE (etil-tertio-butil-etere), un additivo che aumenta il numero di ottano della benzina, o aggiunto al carburante come tale. Il biodiesel è prodotto tramite transesterificazione con metanolo dei trigliceridi contenuti negli oli vegetali, una reazione che produce esteri degli acidi grassi (biodiesel) e glicerina (coprodotto di lavorazione) (Figura 3). Negli Stati Uniti il biodiesel è prodotto principalmente da olio di soia, in Europa da olio di colza. La Direttiva n° 2003/30/CE prevede il raggiungimento per ogni stato membro di obiettivi indicativi di sostituzione dei carburanti derivanti dal petrolio con biocarburanti e/o altri carburanti da fonti rinnovabili per una quota pari al 2% (sulla base del contenuto energetico) nel 2005 fino al 5,75% nel 2010. L’Italia ha recepito questa Direttiva con il decreto legislativo n° 128 del 30 maggio 2005 stabilendo, però, obiettivi indicativi nazionali più bassi, pari rispettivamente all’1% nel 2005 e al 2,5% nel 2010. Assumendo una produzione di bioetanolo da barbabietola di 4 T/ha o di biodiesel da colza o girasole di 1 T/ha, e un consumo stabile di carburanti di origine fossile grazie all’introduzione di nuove tecnologie di trasporto, per raggiungere questi obiettivi con biocarburanti di produzione nazionale si dovrebbe destinare, rispettivamente, alla coltura di barbabietole o oleaginose rispettivamente 250.000 o 1 M ha,

Biotecnologie per l’energia e la chimica
8
ovvero ripettivamente il 2% e l’8% circa della superficie coltivabile italiana. Per recepire pienamente la direttiva CE, le superfici salirebbero al 5% e al 18%. Due problemi fondamentali nell’uso di biocarburanti sono la resa energetica e il costo. La resa energetica, calcolata come il rapporto fra tutti gli “input” energetici (dalla coltivazione della materia prima, al trasporto, alla trasformazione) e tutti gli “output” (il combustibile, ma anche tutti i coprodotti della lavorazione, come per esempio la glicerina dall’industria del biodiesel), è oggetto di complessi studi sulla filiera produttiva, noti come “life cycle inventories”. I risultati di questi studi sono altamente influenzati dai parametri presi in considerazione, dai confini del sistema e dalle assunzioni circa l’uso finale dei coprodotti. Uno dei primi studi è stato condotto sul biodiesel da olio di soia negli Stati Uniti (13). Il risultato di questo studio è che il bilancio energetico del biodiesel da soia è estremamente favorevole: per ogni unità di energia fossile immessa nel processo di produzione del biodiesel, se ne ottengono 3,2 di energia dal biodiesel prodotto. Dato che le emissioni di CO2 sono approssimativamente proporzionali alla quantità di energia fossile impiegata, se ne deduce che l’uso del biodiesel da soia in forma pura ridurrebbe a circa 1/3 le emissioni di CO2 di un veicolo. Studi analoghi condotti sul bioetanolo da mais hanno dato risultati meno incoraggianti: la maggioranza degli studi riferiscono bilanci leggermente maggiori di 1, mentre due studi riferiscono addirittura bilanci minori di 1 (ovvero che per produrre il bioetanolo da mais servirebbe più energia di quella che si produce). Un meta-studio pubblicato di recente (14), ha analizzato sei studi precedenti ed ha concluso che il bilancio energetico del bioetanolo da mais è da neutro a leggermente positivo (da 1 a 1,26) a seconda delle assuzioni fatte, mentre le emissioni dei gas serra sarebbero da neutre a leggermente negative (da 1,02 a 0,86). Bilanci estremamente interessanti (un bilancio di 10 per l’energia e di 0,1 per i gas serra) sono invece proiettati da questo studio per il bioetanolo da colture cellulosiche, che tuttavia non è una tecnologia matura. A livello Europeo, i bilanci energetici per i biocarburanti sono comparabili, nonostante la migliore efficienza energetica del sistema agricolo europeo rispetto a quello americano: un recente studio INRA cita bilanci di 1,19 per l’etanolo da grano, 1,28 per quello da barbabietola e 2,5 per il biodiesel da colza (15). Lo stesso studio conclude che i biocarburanti di produzione europea, al prezzo attuale del petrolio, non sono economicamente competitivi. Il biodiesel diventerebbe competitivo ad un prezzo del petrolio >80 $/barile, mentre il bioetanolo, a causa del suo bilancio energetico più sfavorevole, lo sarebbe a prezzi del petrolio ancora più alti. Di conseguenza, allo stato attuale i biocarburanti debbono essere sostenuti, rispetto ai carburanti tradizionali, da un regime di esenzioni fiscali. I biocarburanti di origine tropicale (bioetanolo da canna da zucchero, biodiesel da olio di palma) hanno sicuramente produttività per ettaro e bilanci energetici migliori di quelli prodotti alle nostre latitudini. Occorre però tenere presente che, in termini globali, le coltivazioni di canna da zucchero in Brasile o di palme da olio nel Sud-Est asiatico competono per le stesse risorse (acqua, fertilizzanti, terra coltivabile, manodopera agricola) con le principali colture alimentari; inoltre, rischiano di incoraggiare la deforestazione delle foreste pluviali tropicali che sono uno dei principali “sink” di CO2 del pianeta. Prima quindi di incoraggiare l’importazione di questi biocarburanti dal Sud del mondo in Europa, occorre valutare attentamente gli effetti ambientali e sociali di un aumento della loro diffusione.

Biotecnologie per l’energia e la chimica
9
5. Le piante per la “green chemistry” Le piante sono i migliori chimici organici del pianeta: mentre il batterio E. coli, con i suoi circa 4.000 geni, é capace di sintetizzare circa 800 metaboliti diversi, una pianta, con i suoi circa 30.000 geni, ne sintetizza ben oltre 100.000 (16). Le piante sono in grado di compiere complesse sintesi organiche, come quella del taxolo (un farmaco antitumorale che ha un costo estremamente elevato e la cui sintesi organica è stata eseguita in 3 soli laboratori al mondo) a spese dell’energia gratuita del sole. Oltre il 30% dei principi farmacologici attualmente in uso, nonché la grande maggioranza dei nutrienti che consumiamo, sono direttamente o indirettamente di origine vegetale. Il metabolismo vegetale è oggetto dello studio di tre discipline:
• la metabolomica (16) che, tramite l’uso di tecnologie di “profiling” ad elevato parallelismo punta, alla individuazione di tutti i metaboliti presenti in un tessuto vegetale;
• la genomica che permette di studiare i profili di espressione genica, associandoli ai profili metabolici di una pianta/tessuto;
• l’ingegneria metabolica che, basandosi sulla conoscenza delle pathway biochimiche e delle tecnologie di ingegneria genetica, modifica, tramite approcci “trial and error”, il contenuto in metaboliti di un determinato tessuto (17).
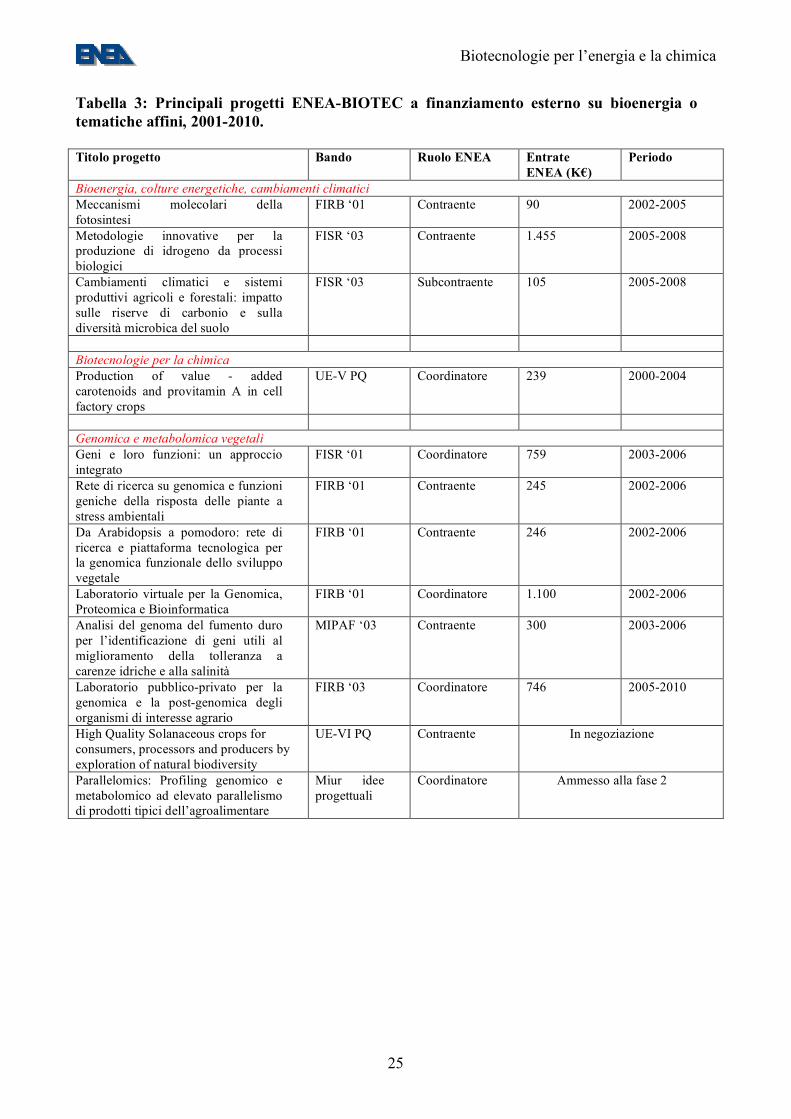
L’ENEA ha una posizione di eccellenza, a livello italiano, nella genomica e nella ingegneria metabolica vegetali (Tabelle 3 e 4). Occorre invece costruire delle competenze nel campo della metabolomica. Tali competenze andranno focalizzate e indirizzate verso attività di ingegneria metabolica su piante e microalghe, volte da una parte ad ottimizzare la produzione di biocombustibili (biodiesel, biodrogeno) e dall’altra ad ottimizzare l’uso delle biomasse coprodotte, sostituendo gradualmente le materie prime di origine fossile, usate attualmente nell’industria chimica, con quelle di origine biologica. 6. Le biotecnologie e la transizione ad una società ad emissioni zero Da quanto detto nella sezione 4 appare evidente che, con le tecnologie attuali, i combustibili di origine agricola possono contribuire solo in misura minore (<10%) al nostro bilancio energetico. I principali problemi sono la bassa efficienza energetica della fotosintesi (<1%, comparata con un >10% del fotovoltaico), la scarsità di terre destinabili a colture energetiche (il raggiungimento di un obiettivo modesto come quello UE di sostituire il 5,75% dei carburanti fossili con biocarburanti entro il 2010 richiederebbe un uso di terreni agricoli maggiore di quelli attualmente “set aside” (12, 15)), la competizione con le colture alimentari tradizionali. Una gran parte di questo contributo può essere realizzato con tecnologie già esistenti: l’uso più razionale delle biomasse di origine agricola e l’introduzione di colture oleaginose con una resa migliore. Un ulteriore contributo (2-3% sul bilancio energetico globale) potrebbe essere ottenuto con l’introduzione di colture cellulosiche e tecnologie per la efficiente fermentazione della cellulosa ad etanolo, obiettivi a cui punta una gran parte dello sforzo di ricerca sulle biomasse finanziato dal DOE. Tuttavia, in assenza di significative innovazioni rispetto a quanto esposto finora, la transizione ad una società ad emissioni zero sembrerebbe affidato a tecnologie non biologiche come il nucleare, il solare termico, il fotovoltaico.

Biotecnologie per l’energia e la chimica
10
Questo significa forse che la ricerca biotecnologica a scopo energetico debba essere abbandonata in favore di ricerche su tecnologie più promettenti? La risposta è no, per almeno tre ordini di motivi:
• Il bilancio di CO2 dell’atmosfera terrestre è determinato, in misura preponderante, da due processi biologici: fotosintesi e respirazione (Figura 2). Il mantenimento di questo bilancio entro limiti ben determinati è essenziale per la stabilità del clima. Le nostre conoscenze sugli effetti del cambiamento climatico sulla ecologia del nostro pianeta sono ancora estremamente rudimentali.
• Le proiezioni disponibili sulla produzione di CO2 antropica (2) rendono essenziale l’introduzione di nuove tecnologie per la sequestrazione attiva della CO2, in cui le tecnologie biologiche ricoprono un ruolo potenzialmente di spicco
• Gli ecosistemi acquatici, che coprono la maggior parte della superficie del pianeta, sono, in termini di bioenergia, una risorsa del tutto inesplorata.
Nel settore biotecnologico, appaiono quindi strategici approcci di ricerca che consentano:
a) una migliore capacità di previsione degli effetti dei cambiamenti climatici sugli ecosistemi terrestri ed acquatici; b) una aumentata efficienza della fotosintesi clorofilliana in sistemi chiusi (obiettivo: >8%) e una efficace fissazione biologica della CO2 direttamente ai punti di produzione (centrali termoelettriche); c) la possibilità di spostare, almeno parzialmente, la produzione di biocarburanti e la fissazione della CO2 in ecosistemi acquatici; d) a lungo termine, la transizione a vettori energetici diversi dal carbonio, come l’idrogeno.
7. Le microalghe come fonte energetica La ricerca sulle microalghe appare strategica per il raggiungimento degli obiettivi b)-d); esse infatti:
• hanno, in impianti dimostrativi, dimostrato efficienze fotosintetiche sostenute del 10%, in assenza di un qualsiasi processo di ottimizzazione genetica;
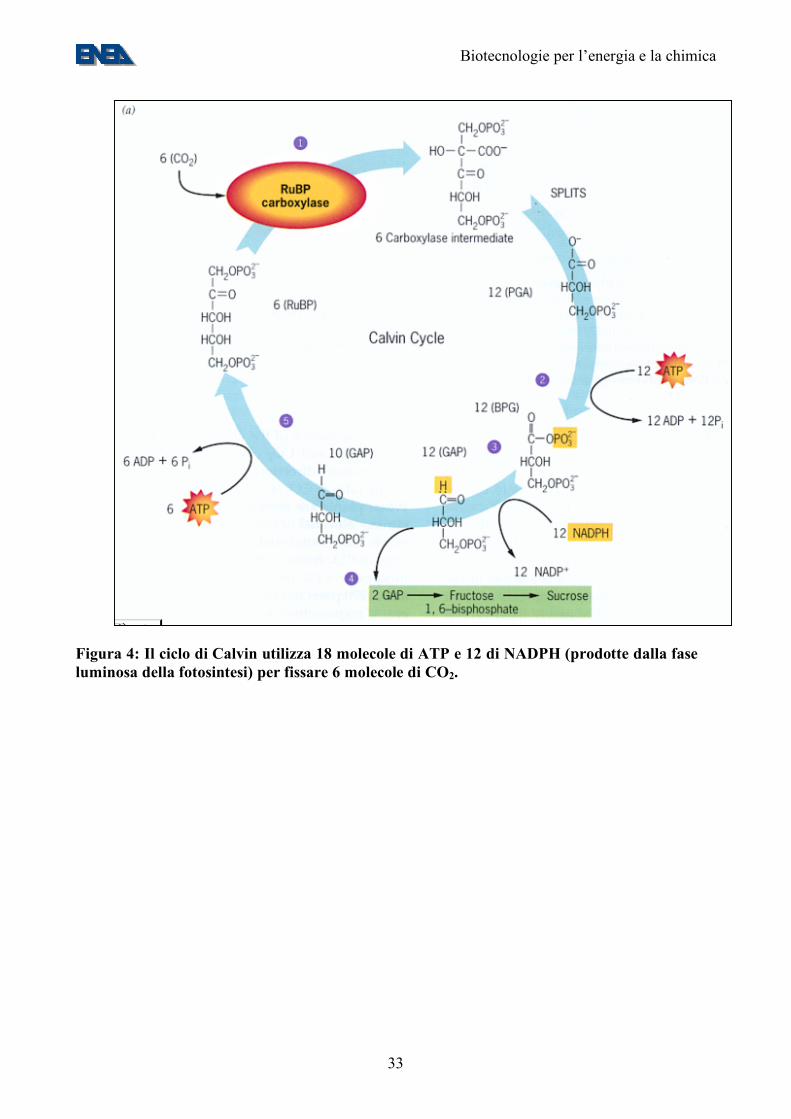
• al pari delle piante superiori, fissano CO2 tramite il ciclo di Calvin (Figura 4); al contrario delle piante superiori, in cui la fotosintesi raggiunge un punto di saturazione intorno alle 500-1000 ppm di CO2, sono capaci di crescere su CO2 altamente concentrata e di sopravvivere in un vasto “range” di temperature, salinità, pH e tensioni di ossigeno;
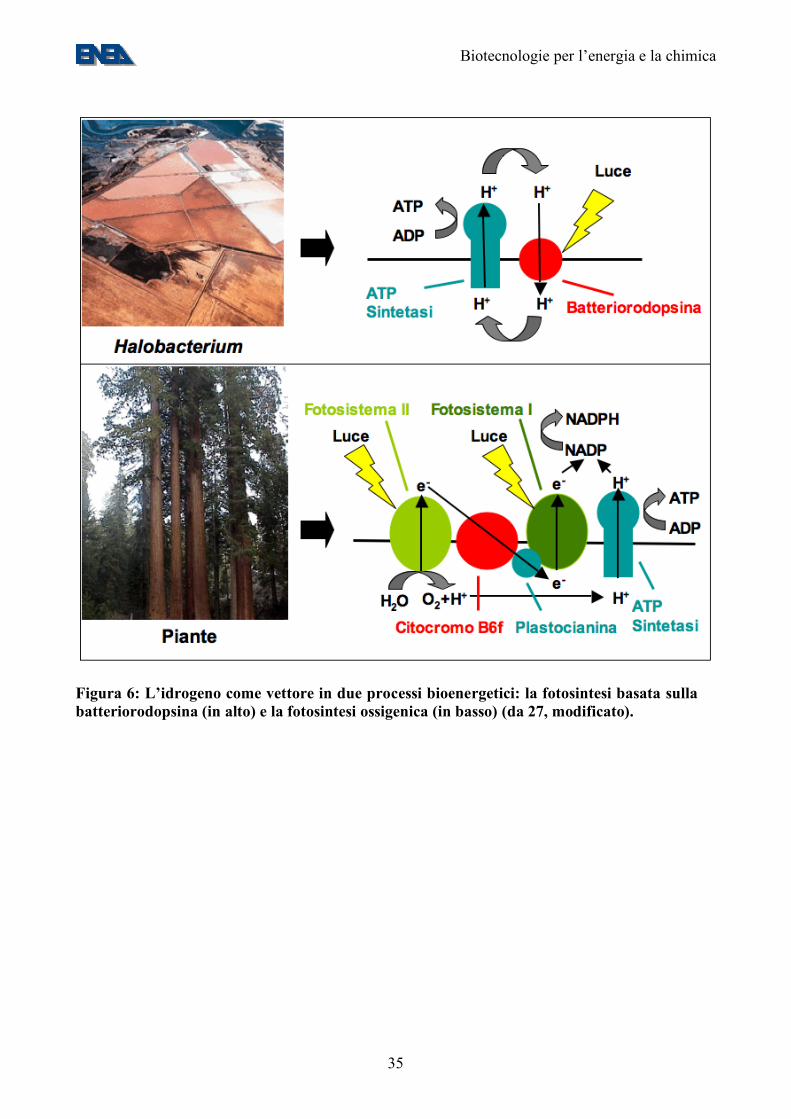
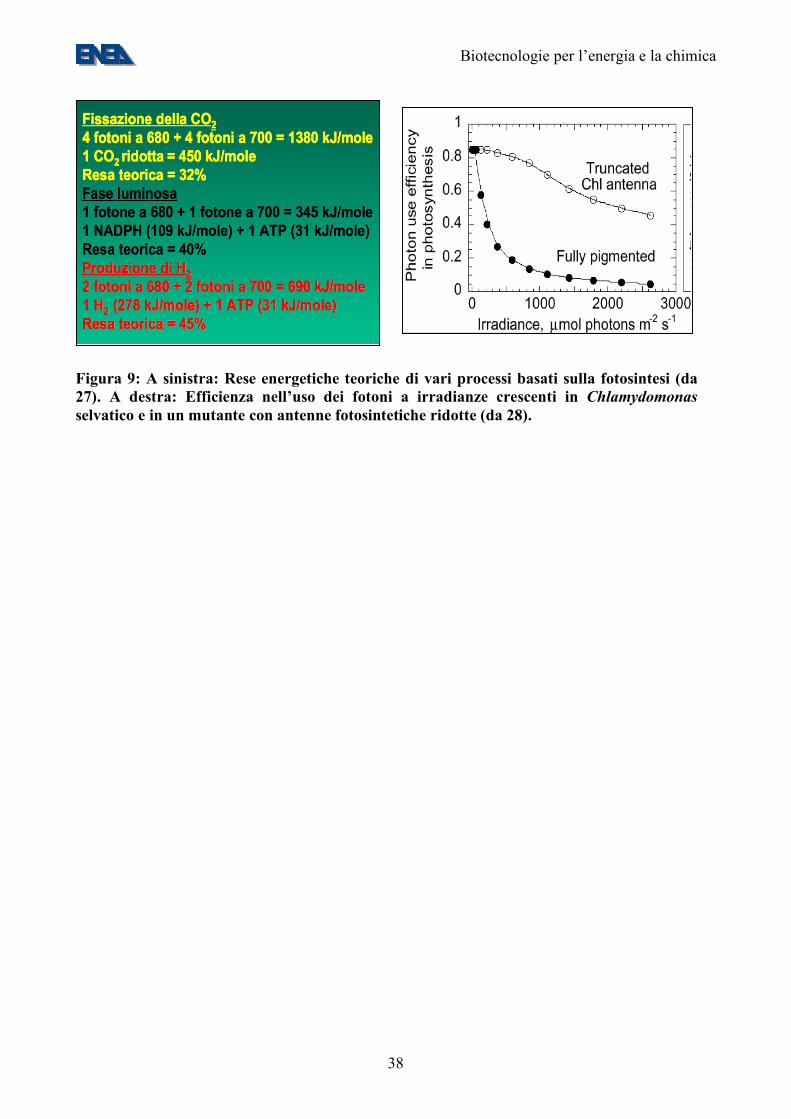
• al pari delle piante superiori, sono in grado di scindere l’acqua in protoni, elettroni ed ossigeno molecolare, con una efficienza energetica teorica di circa il 40% (Figure 6-9);
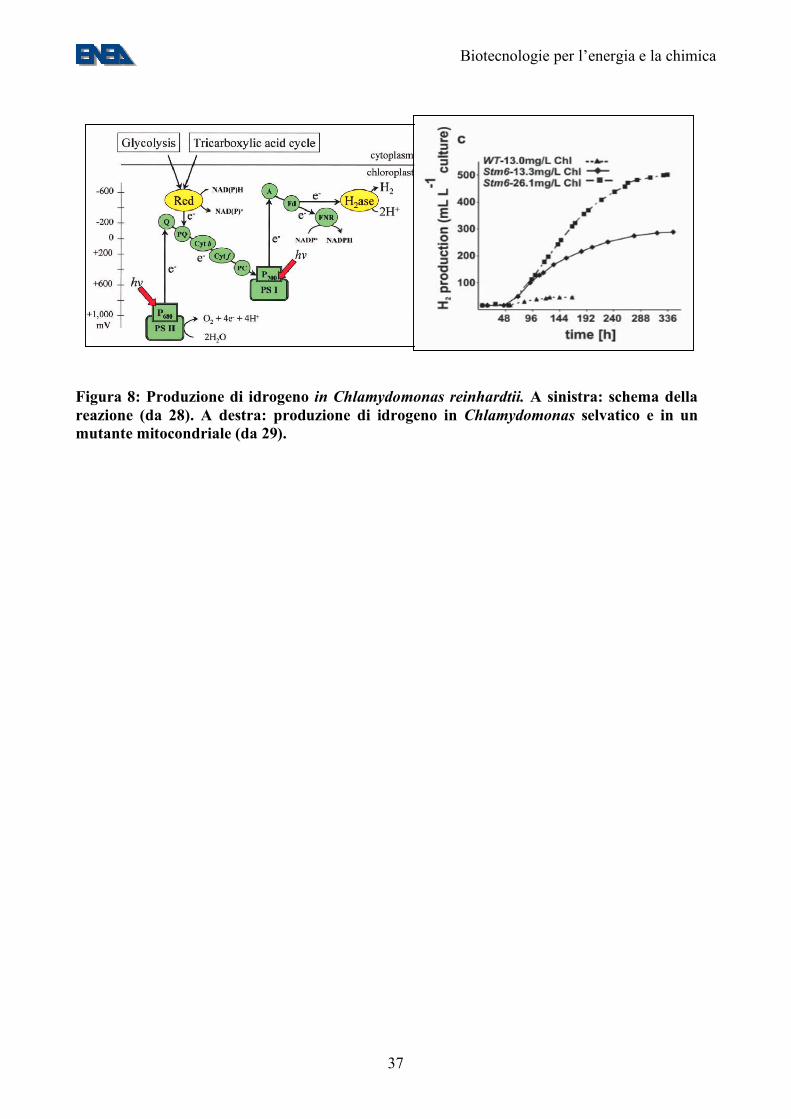
• alcune di esse, in condizioni anaerobiche, sono in grado di ricombinare i protoni e gli elettroni, con la produzione di idrogeno molecolare (Figura 8);
• alcune classi di microalghe accumulano trigliceridi (utilizzabili per la produzione di biodiesel) silicio (utilizzabile nell’industria farmaceutica, chimica e nella nanocomponentistica) carotenoidi (utilizzabili nell’industria cosmetica ed alimentare), bioplastiche (18-20); la biodiversità chimica delle microalghe, e dei microorganismi in genere, resta comunque largamente inesplorata.

Biotecnologie per l’energia e la chimica
11
Dal 1978 al 1996, l’Office of Fuels Development del DOE finanziò un programma, noto come l’Aquatic Species Program, per la produzione di biodiesel da microalghe, cresciute in piccoli specchi d’acqua e alimentate da CO2 prodotta da piccoli impianti a carbone (21). In questi 18 anni, il programma assorbì un finanziamento relativamente modesto (circa 25 M$). I principali risultati furono la produzione di una grande collezione di microalghe, tuttora disponibile presso l’Università delle Hawaii, la messa a punto di metodi di coltura, il clonaggio molecolare di geni che controllano la biosintesi di lipidi, la messa a punto di metodi di trasformazione genetica e test sulla efficienza produttiva in coltura massale. Test condotti in condizioni controllate, in grandi vasche a forma di “autodromo” (Figura 5) dimostrarono produzioni di punta di biomassa di 180 T/ha/anno (equivalente a un’efficienza fotosintetica di circa il 10%), un’utilizzazione della CO2 insufflata di oltre il 80%, ed una capacità di punta di fissare CO2 di 120 T/ha/anno (21). Assumendo una composizione di trigliceridi della biomassa algale del 30%, l’intero fabbisogno italiano di carburanti per autotrazione sarebbe coperto con 750.000 ettari di colture di microalghe (ovvero il 2,4% del territorio nazionale, senza contare i mari). Questa stessa superficie avrebbe la capacità di fissare il 75% delle emissioni di CO2 del Paese. Un fattore importante nei calcoli di fattibilità della produzione di biodiesel da alghe è il costo. L’Aquatic Species Program venne chiuso nel 1996, principalmente a causa del fatto che, sulla base dei dati raccolti fino a quel momento con ceppi non ottimizzati da un punto di vista genetico, il prezzo dell’olio da microalghe avrebbe oscillato fra 39 e 69 $ al barile, contro un prezzo del petrolio (1996) di 19 $ al barile (21). Il prezzo del petrolio, al momento in cui questo “report” è stato scritto, supera i 70 $ al barile. I costi di sequestrazione della CO2 tramite microalghe sono stati valutati fra 9 e 77 dollari/ tonnellata CO2 con un prezzo del petrolio di 35$/barile (21). Tale costo si abbassa al salire del prezzo del barile di petrolio e potrebbe diventare, ai prezzi attuali del petrolio, competitivo con quello del sequestro geologico, stimato in 3-10 $/tonnellata CO2 (22). Un modo per rendere il processo ulteriormente vantaggioso da un punto di vista economico è quello di combinare la fissazione di CO2 e per la produzione di biodiesel con la depurazione di rifiuti liquidi urbani o agricoli. Nei test condotti nell’Aquatic Species Program, tali rifiuti liquidi furono impiegati con successo come fonte di nutrienti (principalmente azoto e fosforo). Un ulteriore fattore di convenienza economica potrebbe essere l’utilizzo di tecniche di ingegneria metabolica per arricchire la biomassa algale residua di sostanze ad elevato valore aggiunto (ad es bioplastiche) o renderla maggiormente suscettibile a processi di fermentazione a metano o idrogeno. Una parte delle emissioni di CO2 italiane sono troppo disperse sul territorio per essere convogliate, in maniera economicamente sostenibile, in un sito di sequestrazione biologica basato su microalghe (questo discorso vale, a maggior ragione, per la sequestrazione geologica, i cui siti potenziali sono ancora meno diffusi sul territorio). Quindi, quello che può essere prospettato è un modello di tipo “aperto” in cui una frazione dei combustibili fossili attualmente consumati verrebbero bruciati in centrali accoppiate a siti di sequestrazione biologica, i quali a loro volta genererebbero biodiesel per gli usi più dispersi (centrali in cui la sequestrazione sia impraticabile, trasporti, riscaldamento per usi civili). Questo modello è attraente per vari motivi:
• esso abbatterebbe le emissioni di CO2 e migliorerebbe la bilancia commerciale in misura direttamente proporzionale all’abbattimento dell’uso dei combustibili fossili.
• non richiederebbe, a medio termine, l’introduzione di modifiche sostanziali nel sistema industriale e di autotrasporti.

Biotecnologie per l’energia e la chimica
12
• gli investimenti richiesti sarebbero quelli per la costruzione degli impianti per la coltura di microalghe (calcolati in 70-80.000 $/Ha per la costruzione su terraferma (21)) e per il potenzialmento della filiera di produzione del biodiesel che, essendo una filiera industriale ormai matura, ha dei costi preventivabili con precisione. Gli impianti oggi in funzione hanno una capacità produttiva stimata in circa 700.000-1.000.000 di t/anno, ben superiore alle produzioni attuali (320.000 t nel 2004). Restano invece totalmente da esplorare i costi per la coltura confinata di microalghe in mare aperto, che per un Paese come l’Italia costituirebbe una grande risorsa.
• farebbe maturare tecnologie di coltura massale di microalghe, successivamente utilizzabili per la produzione biologica di idrogeno.
La coltura massale di microalghe è già una realtà industriale, condotta in Paesi come gli USA, Israele e l’Australia, in “facilities” fino a 50 Ha di superficie, per almeno 4 specie di microalghe (Spirulina, Dunaliella, Haematococcus e Chlorella) (Figura 5). Per almeno 2 di queste (Dunaliella e Haematococcus) la coltura è finalizzata alla produzione di sostanze per la chimica fine (beta-carotene nel caso di Dunaliella, astaxantina nel caso di Haematococcus). Negli ultimi anni sono stati sviluppati metodi di trasformazione genetica per microalghe appartenenti alle principali famiglie: alghe blu-verdi, alghe verdi e diatomee (23-25), rendendo possibile l’ingegnerizzazione di questi organismi . 8. La produzione biologica di idrogeno L’idrogeno è uno dei vettori energetici più ampiamente utilizzati dagli organismi viventi (Figura 6). E’ quindi ipotizzabile la ingegneria metabolica della materia vivente per la produzione diretta di idrogeno. Tale produzione può avvenire attraverso due processi distinti: la fermentazione delle biomasse e la fotobiolisi. Fermentazione: Ultimamente, la produzione fermentativa di idrogeno ha acquisito nuovo impulso con la scoperta di idrogenasi parzialmente resistenti all’ossigeno in batteri estremofili come la Thermotoga neapolitana (30). La produzione fermentativa di idrogeno avviene a partire dai carboidrati, i quali sono un’importante componente delle biomasse ed una grande riserva di energia, che può essere restituita attraverso i processi di fermentazione batterica in forma di idrogeno. Una mole di glucosio può restituire in via teorica 12 moli di idrogeno. Per ottenere una resa vicina a quella teorica è necessario sviluppare un impianto che combini almeno due processi batterici principali: la fermentazione al buio in cui la mole di glucosio genera 4 moli di idrogeno + 2 di un sale di acido organico come ad es. l’acetato e la fermentazione alla luce con i batteri foto-organotrofi in cui le due moli di acetato generano 8 moli di idrogeno. La resa ottenibile è interessante: Calcoli condotti presso ENEA tramite uno scale-up virtuale di esperimenti di laboratorio fanno stimare che un reattore cilindrico di circa 4 metri di diametro e 4 di altezza alimentato da 42Kg/h di combustibile organico può produrre circa 15 kWh giornalieri di corrente elettrica attraverso una cella a combustibile con efficienza del 50%. Tali dimensioni sono interessanti per l’applicazione in allevamenti animali dove potrebbero soddisfare i bisogni energetici di grandi fattorie e di piccole comunità. Tale approccio consentirebbe di affrontare due temi strategici per il paese: la microgenerazione d’idrogeno da rifiuti organici attraverso un processo complesso di fermentazione e la contemporanea soluzione al grave problema dello smaltimento di liquami derivanti da allevamenti animali. Per quanto riguarda quest’ultimo punto, in Italia è in vigore

Biotecnologie per l’energia e la chimica
13
un regolamento nazionale non molto dissimile da quello comunitario: La Direttiva Europea sui Nitrati (91/676/EEC) e il Regolamento Comunitario sull’agricoltura biologica (EU Regulation n°1804/1999 on Organic Agriculture) che dettano i limiti di massimo quantitativo accettabile per il seppellimento delle deiezioni animali nei terreni agricoli. Il rispetto di queste leggi è impossibile negli allevamenti intensivi in quanto richiederebbe una superficie di circa un ettaro per capo. E’ importante notare che attualmente gli allevatori sono costretti, per rispettare le leggi nazionali e Comunitarie di dotarsi di impianti di costruzione austriaca o tedesca che convertono il rifiuto animale in metano e generano energia con elettrogeneratori a combustione interna del costo di circa un milione di euro ad impianto. E’ ben noto a tutti gli operatori del settore che tali impianti hanno una scarsissima efficienza e sono difficili da condurre: il processo batterico che porta alla formazione di metano è un processo molto instabile, inibito da numerose sostanze e dal variare delle condizioni chimico-fisiche ed inoltre ha un bassissimo OLR (Organic Loading Rate) ed un alto tempo di ritenzione; è in definitiva una scelta poco vantaggiosa, ma comunque l’unica scelta per la maggior parte degli allevatori. Proporre di produrre idrogeno con il letame è tutt’altro che utopico, in quanto dal punto di vista strettamente microbiologico è più conveniente e dal punto di vista impiantistico è molto attuale perché esistono sul mercato celle a combustibile di piccole dimensioni (10-20 kWh) che hanno costi intorno ai 100.000 euro. La maggiore difficoltà consiste nel combinare, ingegnerizzare e scalare a dimensione industriale due processi batterici ben studiati separatamente in laboratorio. Tuttavia, assumendo una resa realistica (10 moli idrogeno/mole di glucosio), i 3,8 MT di equivalenti di glucosio disponibili dagli scarti agricoli italiani (sezione 4) genererebbero 0,4 MT H2, ovvero circa 1,1 MTep (vd. Appendice 1). E’ quindi evidente che, per raggiungere quote significative del fabbisogno energetico del Paese, occorra basarsi su biomasse diverse dagli scarti agricoli, come quelle provenienti dalle colture energetiche o dalla coltura massale di microalghe. A lungo termine, la strategia più promettente appare quella della fotobiolisi diretta dell’acqua, che elimina due step poco efficienti dal punto di vista energetico come la fissazione della CO2 in zuccheri e la loro successiva fermentazione. Fotobiolisi: L’atmosfera terrestre primordiale era essenzialmente priva di ossigeno. Circa 3 Mld di anni fa comparvero i cianobatteri, dotati, come le microalghe e le piante verdi da essi derivate, del fotosistema II e quindi della capacità di ossidare l’acqua in protoni, elettroni, ed ossigeno molecolare (Figure 6 e 7). Tale capacità ha modificato la composizione dell’atmosfera terrestre, arricchendola di ossigeno e permettendo la comparsa di organismi che lo respirano (Figura 7) (26). Alcune alghe verdi, come Chlamydomonas reinhardtii, hanno mantenuto la capacità di crescere sia in condizioni aerobiche, nelle quali fissano CO2 producendo zuccheri, sia in condizioni anaerobiche, nelle quali gli zuccheri accumulati sono sottoposti a glicolisi. Nel secondo tipo di condizioni, Chlamydomonas attiva un ciclo futile di produzione di idrogeno che, probabilmente, serve ad utilizzare gli elettroni in eccesso prodotti dalla glicolisi (28) (Figura 8). Di conseguenza, la produzione di idrogeno di Chlamydomonas può essere vista come un processo di autofermentazione degli zuccheri accumulati durante la fase fotosintetica. La produttività in idrogeno di Chlamydomonas si attesta sui 5 ml/litro di coltura/giorno in ceppi selvatici e può essere decuplicata in ceppi geneticamente modificati (29), ovvero, facendo le stesse assunzioni impiantistiche fatte per la coltura di alghe da biodiesel, 5 litri/m2/giorno. Questo equivale ad una efficienza energetica, rispetto alla energia solare incidente, dello 0,35%. Restano quindi da compiere grandi passi avanti nell’aumento dell’efficienza del sistema. Uno dei passi più promettenti sembra quello dell’ingegnerizzazione dell’enzima produttore di idrogeno, l’idrogenasi, in modo che sia presente anche nella fase aerobica e sia resa resistente

Biotecnologie per l’energia e la chimica
14
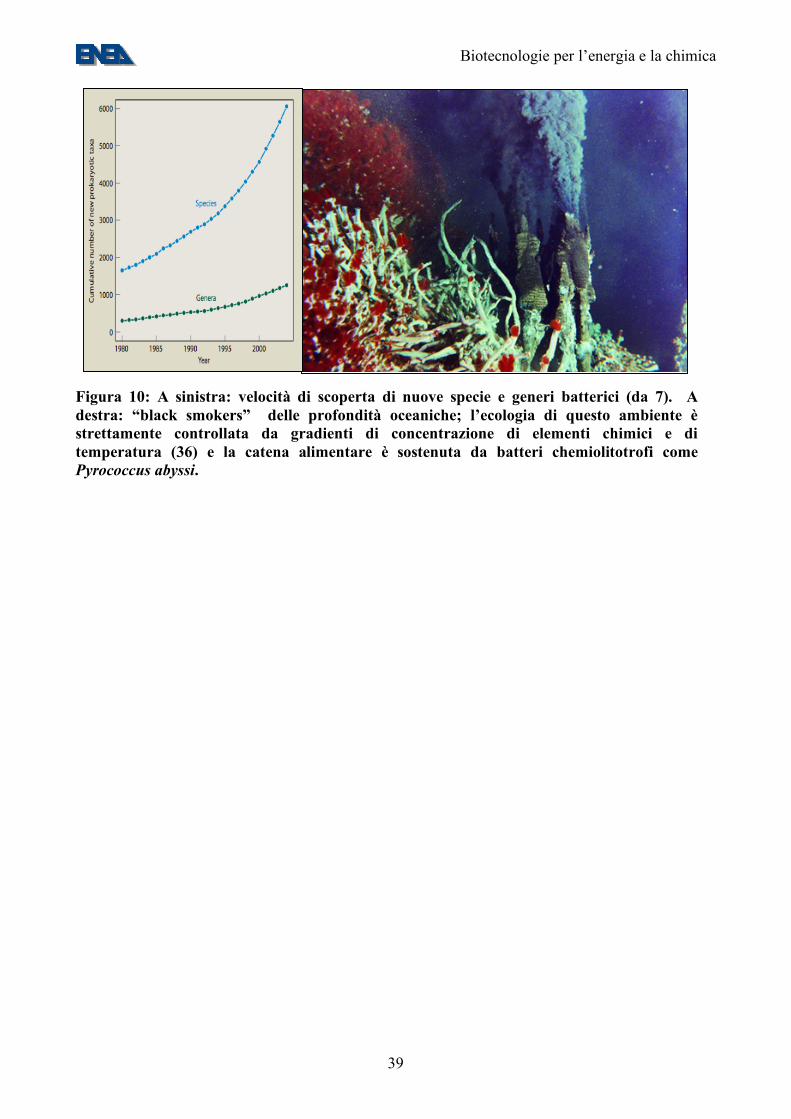
all’inibizione da parte dell’ossigeno (che inibisce l’attività di tutte le idrogenasi attualmente conosciute, con l’eccezione di quelle di alcuni batteri termofili (30). In tal modo, la cellula produrrebbe una mistura di ossigeno e idrogeno molecolari per fotobiolisi diretta (che ha una efficienza teorica del 40%, Figura 9). Tali gas potrebbero essere separati tramite sistemi di esclusione molecolare, basati sulla differente massa delle due molecole. Un secondo passo importante è quello della riduzione delle antenne fotosintetiche. Questo intervento, apparentemente contro-intuitivo, si basa sul fatto che molte microalghe hanno un “optimum” di luminosità estremamente basso: a luminosità simili a quelle del pieno sole, l’efficienza dell’uso dei fotoni cala drasticamente (Figura 9), in quanto la maggior parte della energia assorbita viene riemessa passivamente tramite un meccanismo noto come “non photochemical quenching” (38). Il risultato è che in una coltura algale densa i pochi cm superficiali della coltura assorbono (e disperdono) la maggior parte della luce incidente, utilizzandola in maniera sub-ottimale e lasciando al buio le alghe sottostanti. La riduzione della dimensione delle antenne fotosintetiche riduce la dispersione della luce da parte delle cellule superficiali ed aumenta l’utilizzo da parte di quelle più profonde, aumentando la resa fotosintetica della coltura (Figura 9). La riduzione delle antenne allevierebbe anche problemi impiantistici relativi alla costruzione delle vasche, che attualmente debbono avere una profondità di poche decine di cm. Un terzo passo importante appare la riduzione di vie di trasporto di elettroni che competono con l’idrogenasi per gli elettroni necessari alla produzione di idrogeno (29). 9. La biodiversità microbica I microbi popolano il mondo intorno a noi e contribuiscono, in maniera sostanziale, a renderlo abitabile. I microorganismi, benchè dotati di un metabolismo molto più semplice di quello delle piante superiori, sono in grado di adattarsi a condizioni fisico-chimiche estreme. Un esempio è quello di Pyrococcus abyssi, un batterio ipertermofilo abissale che prospera nei pressi dei “black smokers” oceanici (Figura 10). Malgrado la scoperta di nuove specie e generi microbici sia in piena espansione (Figura 10), è comunemente accettato che circa il 99,5% delle specie microbiche non sia ancora stato scoperto (7). Uno dei motivi per questo “gap” conoscitivo è che la maggior parte dei microorganismi presenti nell’ambiente (>99,5%) non sono coltivabili nelle normali condizioni di laboratorio. D’altronde, proprio la totale assenza di informazioni al riguardo non consente di approntare idonei terreni di coltura. Attualmente, l’approccio preferito per lo studio delle popolazioni microbiche presenti in ambiente naturale si basa sull’analisi del DNA, che viene estratto direttamente da campioni (acqua, suolo, etc.) senza passare per la fase di coltivazione. Questo tipo di approccio permette di prendere in considerazione tutti i componenti della microflora indigena nel sito esaminato. Il DNA viene analizzato sia per ottenere informazioni sulla composizione delle popolazioni naturali, sia per studiarne la funzionalità. Nel primo caso viene esaminata l’eterogeneità della sequenza del gene per il 16S rDNA, che fornisce anche informazioni sulla filogenesi, nel secondo viene studiata la diffusione e la variabilità di geni coinvolti nelle vie metaboliche oggetto di studio. Non solo moltissimi microorganismi, ma anche la stragrande maggioranza dei loro geni sono ancora sconosciuti. Pertanto un ulteriore approccio allo studio delle popolazioni batteriche naturali consiste nell’analisi di intere sequenze genomiche da campioni ambientali allo scopo di individuare i geni presenti e metterne in evidenza la variabilità. Una disciplina che studia geneticamente i geni dei microorganismi mediante un approccio indipendente dalla coltivazione in laboratorio è la Metagenomica (nota anche come “Genomica ambientale” o “Genomica delle Comunità”, 31, 32). La metagenomica consiste nell’analisi del metagenoma, definito come l’insieme dei genomi di una comunità microbica presente in un determinato ambiente. Il termine è stato usato per la prima volta da Jo Handelsman e colleghi (33) e

Biotecnologie per l’energia e la chimica
15
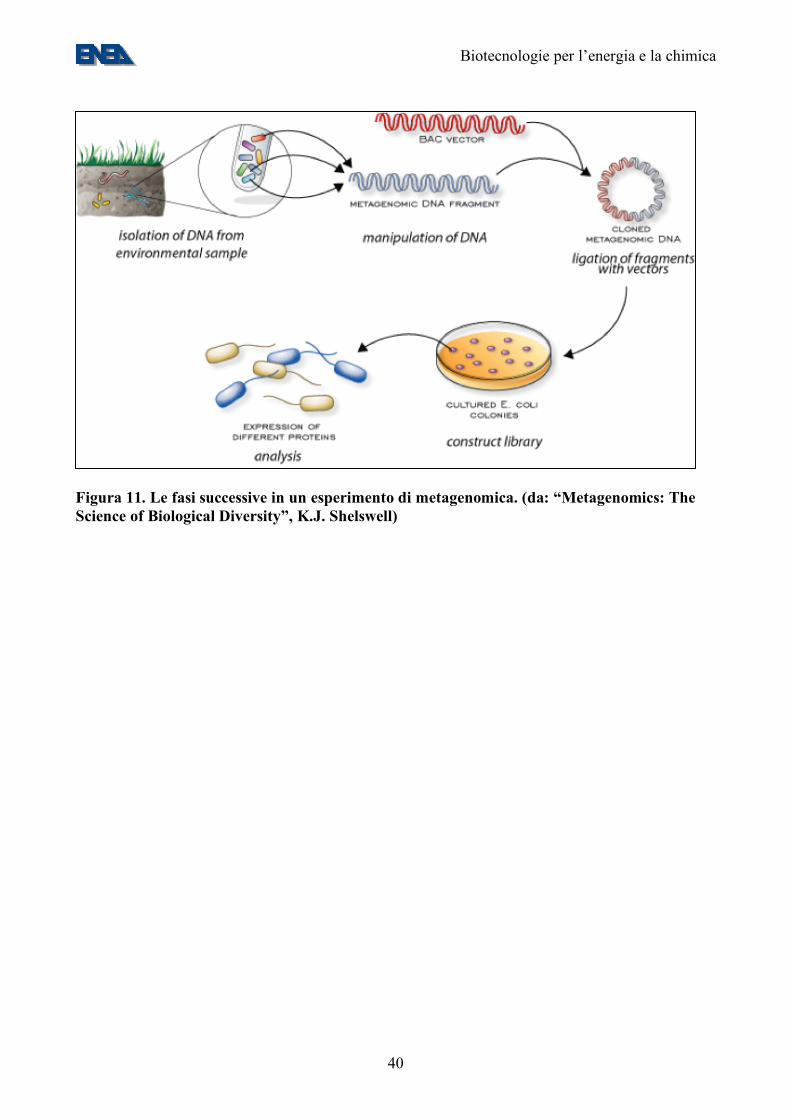
definisce un approccio che permette lo studio di genomi anche di organismi non facilmente coltivabili in laboratorio mediante il recupero del DNA totale e l’analisi dei genomi in esso contenuti (31,32) (Figura 11). La metagenomica permette, fra l’altro, di esplorare la presenza di nuovi geni nel DNA della comunità microbica totale. In tal modo, fornisce una visione completa della comunità microbica in quanto permette di valutare estesamente il grado di diversità filogenetica e di polimorfismo intraspecifico; inoltre, può essere utile per ricostruire sequenze pressoché complete di genomi di microrganismi refrattari alla coltivazione (34). In una recente campagna di metagenomica applicata all’ecologia del mar dei Sargassi, sono state sequenziate oltre 1 Mld di basi di DNA da campioni ambientali, derivati da oltre 1.800 specie e oltre 1,2 milioni di geni non descritti in precedenza (34). Con la stessa metodologia, sono stati ricostruiti genomi di batteri che prosperano in siti altamente contaminati sfruttando gli inquinanti ambientali come fonte di nutrimento (35). Nuovi geni e prodotti genici scoperti comprendono la prima batteriorodopsina di origine batterica, molecole ad attività antimicrobica, nuovi membri di famiglie di proteine note, come le Na+(Li+)/H+ antiporter, RecA, DNA polimerasi e geni codificanti per nuove resistenze ad antibiotici. I risultati ottenuti dall’analisi genomica possono essere anche utilizzati in sinergia con i dati derivanti dallo studio dei metaboliti prodotti negli ambienti esaminati (un approccio che, per analogia al termine “metagenomica” viene qui definito “metachimica”). All’interno di un ecosistema esistono complesse correlazioni tra le vie metaboliche dei diversi microrganismi, che vengono messe in evidenza attraverso lo studio comparativo dei geni e dei loro prodotti di espressione. La conoscenza genomica della comunità permette di modellare, con maggiore accuratezza, le vie di processamento dei metaboliti all’interno della comunità stessa (Figura 12). Questo approccio combinato, è utile per mettere in evidenza le vie metaboliche coinvolte nella produzione di determinati metaboliti e per comprendere quali meccanismi metabolici permettono l’adattamento di determinate specie batteriche ad ambienti estremi (35). In conclusione, le nuove tecnologie di metagenomiche e di “profiling” metabolico ad elevato parallelismo, se applicate a comunità ambientali di microorganismi, aprono vaste prospettive di caratterizzazione di nuovi prodotti e processi utilizzabili nell’industria chimica, nella bioenergia, nella “bioremediation” e, soprattutto, dei geni che ne controllano il funzionamento. La genomica e metabolomica ambientali si presentano quindi come la chiave di volta di uno sforzo di ricerca volto a capire la biosfera ed il suo potenziale contributo alla risoluzione dei problemi energetici ed ambientali. 10. Il posizionamento ENEA In campo vegetale, l’ENEA ha una posizione di eccellenza nei settori della genomica e dell’ingegneria metabolica vegetale, malgrado un investimento di personale finora inadeguato all’output in termini di prodotti CIVR e dei finanziamenti competitivi attirati (Tabelle 3 e 4). Un secondo punto di forza ENEA è la microbiologia degli ambienti naturali (Tabelle 3 e 4). Esistono competenze nel settore delle colture energetiche e della coltura di microalghe, le quali non hanno dato luogo ad una consistente produzione CIVR. Sono in corso di sviluppo competenze sulla produzione biologica di idrogeno, sia tramite fermentazione che tramite fotobiolisi, nell’ambito del progetto FISR Idrobio (Tabella 3). Le competenze nel settore della metabolomica sono invece allo stadio embrionale e vanno sviluppate. Inoltre, sono disponibili una serie di infrastrutture di ricerca importanti (Tabella 5).

Biotecnologie per l’energia e la chimica
16
Queste competenze, opportunamente rafforzate e coordinate ed in collaborazione con opportuni “partners” esterni, possono dar luogo agli assi di ricerca qui di seguito proposti. 11. Assi di ricerca strategici 11.1. Environmental systems biochemistry Il potenziamento di piattaforme tecnologiche avanzate nel campo delle scienze -omiche è trasversale a tutto l’intervento ENEA in campo biotecnologico. Tali piattaforme sono attualmente usate per le attività di biologia avanzata ENEA in campo agroalimentare (sequenziamento del genoma del pomodoro, caratterizzazione del proteoma della vite). L’applicazione di tali metodologie ai settori della bioenergia e della microbiologia ambientale sono essenziali per far compiere alla ricerca ENEA in questi settori un salto di qualità. Da una parte, andrà potenziata la posizione ENEA nel settore della genomica. Dall’altra, andranno sviluppate competenze nel settore della metabolomica e della analisi bioinformatica dei genomi e delle “pathway” metaboliche. La piattaforma comprenderà:
• il sequenziamento di geni e l’identificazione di metaboliti direttamente da campioni ambientali tramite tecnologie ad elevato parallelismo
• l’attribuzione bioinformatica di funzioni metaboliche ai geni sequenziati, e l’ordinamento dei metaboliti in potenziali catene metaboliche, sulla base di analisi di somiglianza con enzimi a funzione nota, di una compilazione delle reazioni chimiche conosciute e catalizzate da enzimi, della struttura dei metaboliti identificati
• la verifica della funzione delle proteine tramite l’espressione sistematica in E. coli, l’aggiunta delle proteine espresse a campioni ambientali, lo studio delle alterazioni metaboliche indotte in seguito a tale aggiunta.
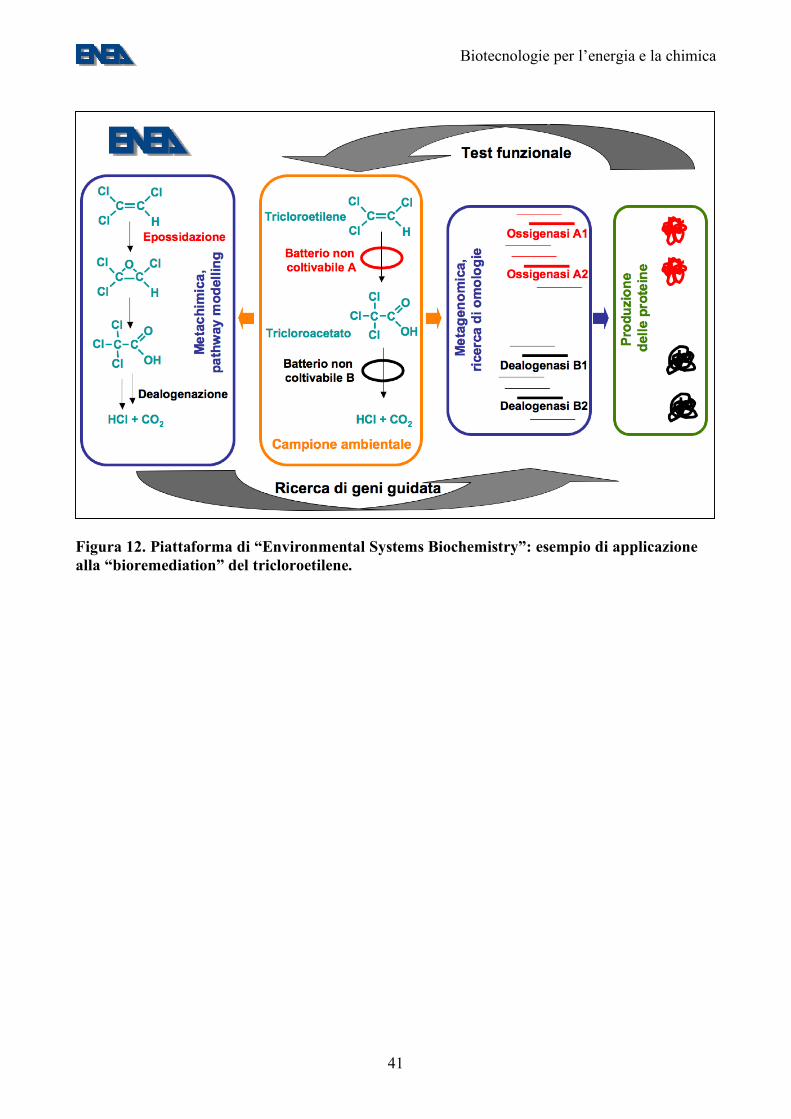
L’obiettivo di questa attività di ricerca sarà quello di mettere in piedi un approccio innovativo alla caratterizzazione di campioni ambientali: l’identificazione diretta di geni in campioni ambientali (metagenomica) unita alla identificazione diretta di nuovi gruppi di metaboliti (metachimica) e l’identificazione, tramite una combinazione di approcci bioinformatici e sperimentali, di nuove catene metaboliche in campioni ambientali. Lo schema di una piattaforma di “Environmental systems biochemistry” e della sua possibile applicazione alla “bioremediation” è riportato nella figura 12. Tale piattaforma ha un numero elevato di potenziali applicazioni, come l’individuazione di nuove pathway biochimiche vegetali e nell’isolamento dei relativi geni, da utilizzare in esperimenti di ingegneria metabolica. Interventi di ricerca e obiettivi: Potenziamento e completa robotizzazione della piattaforma di sequenziamento automatizzato esistente. Suo affiancamento con una piattaforma di sequenziamento ad elevato parallelismo, capace di sequenziare completamente un genoma batterico o un trascrittoma eucariotico in poche ore e ad un costo inferiore di 500 €/megabase sequenziata. Sequenziamento completo di almeno 2 genomi batterici di interesse energetico/ambientale. Installazione di piattaforme per il profiling metabolomico ad elevato parallelismo, basate sulla LC-MS e per l’espressione di proteine ad elevato parallelismo ed il “testing” funzionale delle stesse. Elaborazione di codici bioinformatici per attribuire, sulla base della omologia di sequenza e dei metaboliti presenti in un campione, putative funzioni metaboliche ad un gene. Tempi: Entro il 2010. 11.2. Biomasse e biodiesel da piante

Biotecnologie per l’energia e la chimica
17
Biomasse: Un problema delle biomasse, come combustibile da usare nella cogenerazione, è la loro bassa densità energetica, che richiede impianti voluminosi e quindi dispersivi. In tali impianti il rendimento elettrico è basso ed il costo non competitivo in assenza di incentivi e dell’utilizzo del calore generato. Inoltre, va definito lo stato di efficienza della filiera delle biomasse agricole. In assenza di colture dedicate e di legna proveniente dai boschi cedui, i residui delle lavorazioni agricole (paglie, tutoli, sansa) non sono presenti con la continuità e nella quantità necessaria ad alimentare una filiera energetica economicamente sostenibile. Anche a causa della piccola dimensione delle aziende agricole, l’utilizzo di tali residui agricoli è tutt’ora male organizzato, con il risultato che molte biomasse per alimentare le centrali termiche vengono tuttora importate dall’estero, prevalentemente da Paesi dell’America Latina. Queste constatazioni anno dato forma ad un ipotesi di progetto per la creazione di una filiera integrata di colture energetiche ed utilizzo di residui agroforestali, indirizzata alla cogenerazione di energia dispersa sul territorio. Tale progetto viene allegato al presente documento insieme ad un’ulteriore ipotesi progettuale legata alla produzione di idrogeno e contemporanea ricostituzione della sostanza organica dei terreni. Biodiesel: Da quanto discusso nella sezione 6, appare evidente la scarsa resa energetica, ai nostri climi, del bioetanolo, principalmente a causa dell’energia richiesta per i processi di distillazione e disidratazione. Questo suggerisce di concentrare gli sforzi di ricerca sul biocarburante più promettente e diffuso a livello Europeo, il biodiesel che, oltre ad una elevata resa energetica di partenza, presenta due ulteriori vantaggi: la sua utilizzabilità, con piccole modifiche, sul parco auto circolante e la migliore resa energetica dei motori diesel rispetto a quelli a benzina. La principale coltura Europea in questo settore, il colza, ha una resa bassa sul nostro territorio, per una serie di motivi (scarsa adattabilità alle alte temperature, deiscenza dei frutti). Altre colture oleaginose più adatte ai nostri climi, come la Brassica carinata o il girasole presentano difetti di altro tipo (non utilizzabilità del panello proteico, residuo della produzione di olio, per la mangimistica animale, a causa dell’elevato contenuto di glucosinolati nella Brassica carinata, composizione chimica dell’olio di girasole non pienamente adatta alla produzione di biodiesel). Interventi di ricerca ed obiettivi: Dopo una fase di “screening” agronomico iniziale di varie specie oleaginose (condotto in collaborazione con Istituti di ricerca, come il CRA, dotati delle infrastrutture necessarie) gli sforzi di ricerca si dovranno concentrare su una (max due) specie. Andranno sviluppate strategie, basate su metodi genetici tradizionali o di ingegneria genetica, per l’ottimizzazione della composizione dell’olio, la resistenza ad avversità biotiche e abiotiche (compresa l’eliminazione della deiscenza) l’ottimizazione dell’uso dei coprodotti (panello proteico per mangimistica, paglia per generazione distribuita di energia). Gli obiettivi sono di rendere la produzione di biodiesel da colture oleaginose economicamente conveniente, con una filiera interamente basata sul territorio nazionale, rispetto al diesel da petrolio a 60 $ al barile, stabilizzando la produttività media sopra le 1,3 T olio/ha e la resa energetica sopra 2,5, entro il 2015. 11.3. Sequestrazione biologica di CO2, biomasse e biodiesel da microalghe
Da quanto discusso nella sezione 7, le microalghe fotosintetiche hanno la potenzialità di produrre elevate quantità di trigliceridi, utilizzabili per la produzione di biodiesel, per unità di superficie. Per consentire una rapida crescita delle microalghe, nelle vasche di coltura viene fatta gorgogliare CO2 prodotta da piccole centrali di combustione. Questo fatto apre la possibilità di produrre biodiesel a partire dai fumi di combustione delle centrali termoelettriche. Calcoli condotti nell’ambito del programma DOE sul biodiesel hanno dimostrato, in impianti a cielo aperto, una produttività di biomassa di >180 T/ha/anno. Tale produttività è severamente limitata da fattori ambientali, principalmente le basse temperature

Biotecnologie per l’energia e la chimica
18
notturne dell’aria che, unitamente alla scarsa profondità delle vasche, causa grandi escursioni termiche all’interno delle colture. Questo inconveniente può essere ovviato con interventi di natura impiantistica, come l’uso di impianti coperti e l’uso parziale del calore di risulta delle centrali. Nell’ambito dello stesso programma, è stata dimostrata la fattibilità dell’utilizzo di microalghe per la depurazione di acque di scarico ricche in nitrati e fosfati. Interventi di ricerca ed obiettivi: Dopo una fase di “screening” iniziale di varie specie di microalghe gli sforzi di ricerca si dovranno concentrare su una (max due) specie, capaci di produrre. già in partenza, elevate quantità di trigliceridi per unità di superficie. Le ricerche proposte dovranno sviluppare (ove non disponibili) tecnologie di trasformazione genetica delle microalghe. Andranno sviluppate strategie genetiche per l’ottimizzazione della quantità e della composizione dell’olio, della sua estraibilità, dell’utililizzo della biomassa residua dopo l’estrazione, ad esempio tramite fermentazione batterica a metano o idrogeno oppure tramite la produzione di materiali per la “bulk chemistry” (bioplastiche). Andranno trovate soluzioni impiantistiche o genetiche al problema della discontinuità dell’attività di fissazione della CO2 (attiva solo durante i periodi di luce). Le soluzioni impiantistiche comprendono l’immagazzinamento fisico della CO2 durante la notte, mentre quelle genetiche comprendono l’uso di cicli biochimici, come il metabolismo degli acidi crassulacei, che permettono la fissazione della CO2 anche nei periodi di buio (Figura 13). Gli aspetti impiantistici riguarderanno anche la creazione di impianti a basso costo, basati su varianti del disegno ad “autodromo”, coperti, che tramite l’utilizzo del calore refluo e della CO2 generata in impianti di cogenerazione e del trattamento di reflui agricoli o urbani ricchi in azoto e fosforo, consentano una stabilità della temperatura ed una crescita vigorosa della biomassa minimizzando le emissioni nell’ambiente. Gli obiettivi sono di:
• sviluppare ceppi ottimizzati per la produzione di biodiesel, la fissazione di CO2 e il trattamento dei reflui
• stabilizzare la resa di olio su >30 T/ha/anno • fissare >100 T/ha/anno di CO2 ad un costo <20 €/T • rendere il costo del biodiesel competitivo con petrolio ≈ 50 $/barile • Ottimizzare l’uso della biomassa coprodotta tramite fermentazione a idrogeno o
metano o il suo utilizzo come materia prima sostitutiva del petrolio per l’industria chimica
Tempi: Entro il 2020. 11.4. Idrogeno biologico Le ricerche saranno finalizzate alla ottimizzazione della produzione di idrogeno per via fermentativa e per via fotosintetica (fotobiolisi). La via fermentativa comporterà da una parte l’esplorazione di consorzi microbici naturali produttori di idrogeno, utilizzando la piattaforma tecnologica messa a punto nell’asse 1, con l’obiettivo di ottenere consorzi microbici stabili capaci di convertire la biomassa in idrogeno in tempi rapidi e con elevate efficienze oppure, in alternativa, di trasferire le funzioni geniche presenti nel consorzio in un unico microorganismo adatto alla fermentazione. Dall’altra, punterà all’ottimizzazione genetica del processo in batteri che producono idrogeno in coltura pura (Thermotogales, Rhodobacteraceae). La via fotosintetica punterà su tre linee principali: la produzione di microalghe con una ottimizzata capacità di utilizzazione dell’energia luminosa (antenna troncata, migliorata risposta allo stress fotoossidativo). Questa linea è essenziale per aumentare la efficienza fotosintetica del sistema e sarà affrontata anche nell’asse 3 (biodiesel da microalghe).

Biotecnologie per l’energia e la chimica
19
la eliminazione di pathway intracellulari di utilizzazione di elettroni (pathway mitocondriale, trasporto di elettroni ciclico) in grado di inibire la produzione di idrogeno (29) la ingegnerizzazione di idrogenasi resistenti all’ossigeno ed in grado di essere indotte in risposta a segnali chimici o fisici semplici. Obiettivi: Ottenimento di consorzi microbici o di microorganismi ingegnerizzati in grado di trasformare biomasse vegetali o algali con elevata efficienza (>10 moli idrogeno/mole equivalente di glucosio) a costi contenuti e con efficienze elevate: l’idrogeno prodotto per via fermentativa dovrà avere costi ed efficienze energetiche uguali o migliori del metano prodotto a partire dagli stessi materiali (entro il 2010). Ottenimento di microalghe capaci di produrre idrogeno per via fotobiolitica con una elevata efficienza (>8% di conversione dell’energia solare) e a costi contenuti (un Mj di idrogeno dovrà avere un costo uguale o inferiore a 1 Mj di biodiesel) entro il 2020.

Biotecnologie per l’energia e la chimica
20
12. Roadmap
2008: •“Life cycle studies” su combustibile per autotrazione, cogenerazione da biomasse, riduzione delle emissioni di CO2 completati. •Modelli sugli effetti della evoluzione del clima sulla produzione primaria di biomassa in Italia. •Individuazione di aree marginali e di colture da destinare alla produzione di biomasse da combustione. •Individuazione di una/due oleaginose da biodiesel su cui puntare. Messa a punto delle tecnologie di base (trasformazione genetica, prove colturali, tecnologie analitiche dell’olio e dei coprodotti) per il miglioramento della filiera.
2010: •Piattaforma per la “Environmental Systems Biochemistry” attiva. Isolamento di geni microbici e vegetali per l’ingegnerizzazione della biomassa algale. Studi di genetica di popolazioni di microorganismi ambientali. •Sequenziamento di almeno due genomi batterici di interesse energetico/ambientale. •Impianti di cogenerazione (rendimento elettrico >25%) basati su una filiera nazionale di colture energetiche/sfruttamento forestale/residui agroindustriali, con sfruttamento del calore. •Individuazione di una/due microalghe da biodiesel su cui puntare. •Primi round di ottimizzazione della filiera oleaginose (incluso il processamento). •Primo round di ottimizzazione della produzione di idrogeno da processi fermentativi e fotobiolisi (resa fermentativa> 10 moli idrogeno/mole eq. di glucosio; resa fotosintetica >0,8%).
2015: •Sostituzione di 3% dei combustibili fossili con combustibili di origine agroforestale impiegati in sistemi di cogenerazione, con costi al kW competitivi. •Filiera biodiesel nazionale competitiva a partire da oleaginose migliorate (6% del fabbisogno di carburante, resa >1,3 T/ha, resa energetica >2,5, competitività con petrolio a 60 $/barile, uso efficiente dei coprodotti) •Dimostrazione su scala pilota della cogenerazione accoppiata con vasche per il trattamento dei reflui agroindustriali/urbani, fissazione della CO2, produzione di biodiesel •Nuovi geni/processi per la bioremediation e la ingegneria metabolica di microalghe •Idrogeno da fotobiolisi con resa >2%
2020: •Cogenerazione su scala industriale economicamente competitiva (10% del fabbisogno energetico nazionale)

Biotecnologie per l’energia e la chimica
21
•Fissazione della CO2 e trattamento di reflui urbani/agricoli con produzione di biomassa e biodiesel da microalghe (30% del fabbisogno di carburante, competitivo col petrolio a 50 $/barile, resa >30 T/ha, resa energetica>3) •Uso della biomassa algale come materiale di partenza per l’industria chimica o per la produzione di metano e idrogeno). •Idrogeno da fotobiolisi con resa>10%
2030: Coltivazione contenuta di microalghe in mare aperto. >60% del carburante per autotrazione = biodiesel da microalghe >30% del petrolio per l’industria chimica sostituito da prodotti di origine algale. >20% del carburante per autotrazione = idrogeno da biomassa algale e da fotobiolisi. Abbattimento delle emissioni di CO2 al 50% di quelle attuali.

Biotecnologie per l’energia e la chimica
22
13. Tabelle e Figure
Biotecnologie per l’energia e la chimica
23
Tabella 1. Emissioni di CO2 (KtC) da combustibili fossili nel 2002 (fonte: 4). Totali % Pro capite Mondo 6.975.000 100,00% 1,12 USA 1.592.382 22,83% 5,52 Cina 957.249 13,72% 0,74 Russia 390.439 5,60% 2,69 India 332.677 4,77% 0,32 Giappone 327.939 4,70% 2,57 Europa Occidentale 694.633 9,96% 2,05 Germania 219.270 3,14% 2,66 Regno Unito 148.129 2,12% 2,50 Italia 117.989 1,69% 2,05 Francia 100.358 1,44% 1,69 Spagna 82.998 1,19% 2,03
Biotecnologie per l’energia e la chimica
24
Tabella 2: Resa delle principali colture energetiche (fonte: 12).
Biotecnologie per l’energia e la chimica
25
Tabella 3: Principali progetti ENEA-BIOTEC a finanziamento esterno su bioenergia o tematiche affini, 2001-2010. Titolo progetto Bando Ruolo ENEA Entrate
ENEA (K€) Periodo
Bioenergia, colture energetiche, cambiamenti climatici Meccanismi molecolari della fotosintesi
FIRB ‘01 Contraente 90 2002-2005
Metodologie innovative per la produzione di idrogeno da processi biologici
FISR ‘03 Contraente 1.455 2005-2008
Cambiamenti climatici e sistemi produttivi agricoli e forestali: impatto sulle riserve di carbonio e sulla diversità microbica del suolo
FISR ‘03 Subcontraente 105 2005-2008
Biotecnologie per la chimica Production of value - added carotenoids and provitamin A in cell factory crops
UE-V PQ Coordinatore 239 2000-2004
Genomica e metabolomica vegetali Geni e loro funzioni: un approccio integrato
FISR ‘01 Coordinatore 759 2003-2006
Rete di ricerca su genomica e funzioni geniche della risposta delle piante a stress ambientali
FIRB ‘01 Contraente 245 2002-2006
Da Arabidopsis a pomodoro: rete di ricerca e piattaforma tecnologica per la genomica funzionale dello sviluppo vegetale
FIRB ‘01 Contraente 246 2002-2006
Laboratorio virtuale per la Genomica, Proteomica e Bioinformatica
FIRB ‘01 Coordinatore 1.100 2002-2006
Analisi del genoma del fumento duro per l’identificazione di geni utili al miglioramento della tolleranza a carenze idriche e alla salinità
MIPAF ‘03 Contraente 300 2003-2006
Laboratorio pubblico-privato per la genomica e la post-genomica degli organismi di interesse agrario
FIRB ‘03 Coordinatore 746 2005-2010
High Quality Solanaceous crops for consumers, processors and producers by exploration of natural biodiversity
UE-VI PQ Contraente In negoziazione
Parallelomics: Profiling genomico e metabolomico ad elevato parallelismo di prodotti tipici dell’agroalimentare
Miur idee progettuali
Coordinatore Ammesso alla fase 2

Biotecnologie per l’energia e la chimica
26
Tabella 4: Principali prodotti CIVR ENEA . Ingegneria metabolica titolo: To dye or not to dye: biochemistry of annatto unveiled codice: 040496 tipologia: Articolo su rivista categoria ISI: Plant Sciences autori: GIULIANO G; ROSATI C; BRAMLEY PM. anno: 2003 rivista: "TRENDS IN BIOTECHNOLOGY" (ISSN:0167-7799 volume:21 pagine:513-516 impact factor: 7.517)
titolo: Carotenoid oxygenases: cleave it or leave it codice: 040443 tipologia: Articolo su rivista categoria ISI: Plant Sciences autori: GIULIANO G; AL-BABILI S; VON LINTIG J. anno: 2003 rivista: "TRENDS IN PLANT SCIENCE" (ISSN:1360-1385 volume:8 pagine:145-149 impact factor: 13.405)
titolo: Centro di Innovazione Integrato "Agrobiopolis" codice: 040957 tipologia: Risultato di valorizzazione applicativa categoria ISI: Agriculture/Agronomy autori: BALDUCCHI R; BIANCIFIORI MA; DI GIORGIO A; DONADIO G; GAZZOLA G; PIGNATELLI V; VITALI F. anno: 2003
titolo: Metabolic engineering of xanthophyll content in tomato fruits codice: 040490 tipologia: Articolo su rivista categoria ISI: Plant Sciences autori: DHARMAPURI S; ROSATI C; PALLARA P; AQUILANI R; BOUVIER F; CAMARA B; GIULIANO G. anno: 2002 rivista: "FEBS LETTERS" (ISSN:0014-5793 volume:519 pagine:30-34 impact factor: 3.912)
titolo: Carotenoid isomerase: a tale of light and isomers codice: 040493 tipologia: Articolo su rivista categoria ISI: Plant Sciences autori: GIULIANO G; GILIBERTO L; ROSATI C. anno: 2002 rivista: "TRENDS IN PLANT SCIENCE" (ISSN:1360-1385 volume:7 pagine:427-429 impact factor: 12.414)
titolo: Impianto per la preparazione di materiali di riferimento per il settore agroalimentare codice: 040970 tipologia: Manufatto ed opera d'arte

Biotecnologie per l’energia e la chimica
27
categoria ISI: Chemistry & Analysis autori: ZAPPA G; CARCONI P; GATTI R; PALAZZO S; PORTARO N; TROISI L. anno: 2001
titolo: Recombinant plants and DNA constructs codice: 040933 tipologia: Brevetto categoria ISI: Biotechnology & Applied Microbiology autori: GIULIANO G; ROSATI C; DHARMAPURI S; PALLARA P; CAMARA B. anno: 2001
Ingegneria proteica
titolo: Engineering stable cytoplasmic intrabodies with designed specificity codice: 040528 tipologia: Articolo su rivista categoria ISI: Molecular Biology & Genetics autori: DONINI M; MOREA V; DESIDERIO A; PASHKOULOV D; VILLANI ME; TRAMONTANO A; BENVENUTO E. anno: 2003 rivista: "JOURNAL OF MOLECULAR BIOLOGY" (ISSN:0022-2836 volume:330 pagine:323-332 impact factor: 5.239)
titolo: A semi-synthetic repertoire of intrinsically stable antibody fragments derived from a single-framework scaffold codice: 040733 tipologia: Articolo su rivista categoria ISI: Molecular Biology & Genetics autori: DESIDERIO A; FRANCONI R; LOPEZ M; VILLANI ME; VITI F; CHIARALUCE R; CONSALVI V; NERI D; BENVENUTO E. anno: 2001 rivista: "JOURNAL OF MOLECULAR BIOLOGY" (ISSN:0022-2836 volume:310 pagine:603-615 impact factor: 5.826)
titolo: Evidence for cysteine clustering in thermophilic proteomes codice: 040513 tipologia: Articolo su rivista categoria ISI: Biochemistry & Biophysics autori: ROSATO V; PUCELLO N; GIULIANO G. anno: 2002 rivista: "TRENDS IN GENETICS" (ISSN:0168-9525 volume:18 pagine:278-281 impact factor: 13.216)
Microbiologia ambientale titolo: Burkholderia cepacia Complex Bacteria from Clinical and Environmental Sources in Italy: Genomovar Status and Distribution of Traits Related to Virulence and Transmissibility. codice: 040451 tipologia: Articolo su rivista categoria ISI: Microbiology autori: BEVIVINO A; DALMASTRI C; TABACCHIONI S; CHIARINI L; BELLI ML;

Biotecnologie per l’energia e la chimica
28
PIANA S; MATERAZZO A; VANDAMME P; MANNO G. anno: 2002 rivista: "JOURNAL OF CLINICAL MICROBIOLOGY" (ISSN:0095-1137 volume:40 pagine:846-851 impact factor: 3.565) titolo: A rhizospheric Burkholderia cepacia complex population: genotypic and phenotypic diversity of Burkholderia cenocepacia and Burkholderia ambifaria codice: 040449 tipologia: Articolo su rivista categoria ISI: Microbiology autori: DALMASTRI C; FIORE A; ALISI C; BEVIVINO A; TABACCHIONI S; GIULIANO G; SPROCATI AR; SEGRE L; MAHENTHIRALINGAM E; CHIARINI L; VANDAMME P. anno: 2003 rivista: "FEMS MICROBIOLOGY ECOLOGY" (ISSN:0168-6496 volume:46 pagine:179-187 impact factor: 2.947) titolo: Burkholderia cepacia complex: distribution of genomovars among isolates from the maize rhizosphere in Italy codice: 040462 tipologia: Articolo su rivista categoria ISI: Microbiology autori: FIORE A; LAEVENS S; BEVIVINO A; DALMASTRI C; TABACCHIONI S; VANDAMME P; CHIARINI L. anno: 2001 rivista: "ENVIRONMENTAL MICROBIOLOGY" (ISSN:1462-2912 volume:3 pagine:137-143 impact factor: 3.276) titolo: Effects of two different application methods of Burkholderia ambifaria MCI 7 on plant growth and rhizospheric bacterial diversity codice: 040456 tipologia: Articolo su rivista categoria ISI: Microbiology autori: CICCILLO F; FIORE A; BEVIVINO A; DALMASTRI C; TABACCHIONI S; CHIARINI L. anno: 2002 rivista: "ENVIRONMENTAL MICROBIOLOGY" (ISSN:1462-2912 volume:4 pagine:238-245 impact factor: 3.649) titolo: Stimulation of carotenoid metabolism in arbuscular mycorrhizal roots. codice: 040505 tipologia: Articolo su rivista categoria ISI: Plant Sciences autori: FESTER T; SCHMIDT D; LOHSE S; WALTER MH; GIULIANO G; BRAMLEY PM; FRASER PD; HAUSE B; STRACK D. anno: 2002 rivista: "PLANTA" (ISSN:0032-0935 volume:216 pagine:148-154 impact factor: 2.96)

Biotecnologie per l’energia e la chimica
29
Tabella 5. Principali infrastrutture disponibili presso ENEA CR Trisaia:
• Centro di innovazione integrato Agrobiopolis dotato di: • hall tecnologica con impianti pilota di fermentazione (capacità fino a 500 L),
cromatografia preparativa su colonna e rotante, e downstream processing (estrazione a fluidi supercritici, separazione a membrana, separazioni solido/liquido)
• laboratori dedicati di genomica (sequenziamento automatizzato, analisi microarray) proteomica (LC-MS), microbiologia, analisi NMR (600 MHz) anche su matrici solide e semisolide
• Camere di crescita in condizioni controllate, serre e campi sperimentali. • Impanto pilota di “steam explosion” in batch (10 L). • Impianto dimostrativo di “steam explosion” in continuo (300 Kg/h).
CR Casaccia:
• Laboratori tecnologici con impianti di separazione a membrana, estrazione con fluidi supercritici.
• “Facilities” per la coltura di microalghe su scala pilota. • Laboratori dedicati di microbiologia ed ecologia microbica (ca 200 mq). • Laboratori dedicati di genomica (sequenziamento automatizzato “high throughput” ad
elevata robotizzazione, analisi microarray).
Biotecnologie per l’energia e la chimica
30
Figura 1: Consumi energetici proiettati, per fonte energetica (Fonte: 2).

Biotecnologie per l’energia e la chimica
31
Figura 2: Ciclo del carbonio (Fonte: 6).

Biotecnologie per l’energia e la chimica
32
Figura 3: Produzione del biodiesel. A sinistra: ciclo industriale semplificato. A destra: dettaglio della reazione di transesterificazione.
Biotecnologie per l’energia e la chimica
33
Figura 4: Il ciclo di Calvin utilizza 18 molecole di ATP e 12 di NADPH (prodotte dalla fase luminosa della fotosintesi) per fissare 6 molecole di CO2.

Biotecnologie per l’energia e la chimica
34
Figura 5: “Facilities” per la crescita massale di microalghe. In alto: schema di vasca ad “autodromo” per la crescita di microalghe con insufflazione di CO2 (fonte: 21). In basso a sinistra: impianti di produzione di Spirulina (blu-verde), Chlorella (verde), Haematococcus (viola). In basso a destra: stagni per la produzione di Dunaliella (rossa). In sovraimpressione sono riportate le strutture dell’astaxantina (il principale prodotto di Haematococcus) e del beta-carotene (il principale prodotto di Dunaliella).
Biotecnologie per l’energia e la chimica
35
Figura 6: L’idrogeno come vettore in due processi bioenergetici: la fotosintesi basata sulla batteriorodopsina (in alto) e la fotosintesi ossigenica (in basso) (da 27, modificato).

Biotecnologie per l’energia e la chimica
36
Figura 7: A sinistra: Struttura del fotosistema II (PSII), il complesso fotosintetico dove avviene l’ossidazione dell’acqua nella fotosintesi ossigenica. A destra: aumento dell’ossigeno nell’atmosfera terrestre in seguito alla comparsa di organismi dotati di PSII (da 26).
Biotecnologie per l’energia e la chimica
37
Figura 8: Produzione di idrogeno in Chlamydomonas reinhardtii. A sinistra: schema della reazione (da 28). A destra: produzione di idrogeno in Chlamydomonas selvatico e in un mutante mitocondriale (da 29).
Biotecnologie per l’energia e la chimica
38
Figura 9: A sinistra: Rese energetiche teoriche di vari processi basati sulla fotosintesi (da 27). A destra: Efficienza nell’uso dei fotoni a irradianze crescenti in Chlamydomonas selvatico e in un mutante con antenne fotosintetiche ridotte (da 28).
Biotecnologie per l’energia e la chimica
39
Figura 10: A sinistra: velocità di scoperta di nuove specie e generi batterici (da 7). A destra: “black smokers” delle profondità oceaniche; l’ecologia di questo ambiente è strettamente controllata da gradienti di concentrazione di elementi chimici e di temperatura (36) e la catena alimentare è sostenuta da batteri chemiolitotrofi come Pyrococcus abyssi.
Biotecnologie per l’energia e la chimica
40
Figura 11. Le fasi successive in un esperimento di metagenomica. (da: “Metagenomics: The Science of Biological Diversity”, K.J. Shelswell)
Biotecnologie per l’energia e la chimica
41
Figura 12. Piattaforma di “Environmental Systems Biochemistry”: esempio di applicazione alla “bioremediation” del tricloroetilene.

Biotecnologie per l’energia e la chimica
42
Figura 13. Metabolismo degli acidi crassulacei. A sinistra: una pianta crassulacea. A destra: schema del metabolismo. Durante la notte, la CO2 viene temporaneamente fissata in acidi organici, accumulata nel vacuolo e liberata nuovamente durante il giorno per essere utilizzata dal ciclo di Calvin.

Biotecnologie per l’energia e la chimica
43
Appendice 1. Unità di misura: 1 Tep (tonnellata equivalente di petrolio = 7,18 bep (barili equivalenti di petrolio). 1 Tep = 42,41 Gj = 11,78 MWh = 40,19 MBtu = 10,13 Gcal 1 MWh = 3,6 Gj=0,085 Tep 1 Quad = 109 MBtu = 24,88 MTep = 1,05 Ej 1 ettaro (ha) = 10.000 mq 1 tC= tonnellata di carbonio. Unità di misura delle emissioni di CO2. Densità energetica dell’idrogeno = 33 kWh/Kg Densità energetica degli idrocarburi= 12 kWh/Kg Peso specifico della CO2 = 1,97 Kg/m3.

Biotecnologie per l’energia e la chimica
44
14. Bibliografia:
1. http://www.eia.doe.gov/oiaf/aeo/conf/ 2. http://www.eia.doe.gov/oiaf/aeo/conf/pdf/conti.pdf 3. http://www.eia.doe.gov/oiaf/aeo/conf/pdf/petak.pdf 4. ENEA: Rapporto Energia Ambiente. Roma, 2001. 5. Marland, G., T.A. Boden, and R.J. Andres. 2005. Global, Regional, and National CO2
Emissions. In Trends: A Compendium of Data on Global Change. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy, Oak Ridge, Tenn., U.S.A. http://cdiac.ornl.gov/
6. Climate Change 2001. Working Group I: The Scientific Basis. http://www.grida.no/climate/ipcc_tar/wg1/index.htm
7. DOE Genomics:GTL Roadmap: Systems Biology for Energy and the Environment. US Dept. of Energy, August 2005. http://doegenomestolife.org/roadmap/pdf/GenomicsGTL_Roadmap.pdf
8. Sabine CL, Feely RA, Gruber N, Key RM, Lee K, Bullister JL, Wanninkhof R, Wong CS, Wallace DW, Tilbrook B, Millero FJ, Peng TH, Kozyr A, Ono T, Rios AF (2004) The oceanic sink for anthropogenic CO2. Science 305: 367-371
9. Cox PM, Betts RA, Jones CD, Spall SA, Totterdell IJ (2000) Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature 408: 184-187.
10. Caldeira K, Jain AK, Hoffert MI (2003) Climate sensitivity uncertainty and the need for energy without CO2 emission. Science 299: 2052-2054.
11. Ragauskas AJ, Williams CK, Davison BH, Britovsek G, Cairney J, Eckert CA, Frederick WJ, Jr., Hallett JP, Leak DJ, Liotta CL, Mielenz JR, Murphy R, Templer R, Tschaplinski T (2006) The path forward for biofuels and biomaterials. Science 311: 484-489
12. BIOMASS – Green Energy for Europe. European Commission, Luxembourg, 2005. ISBN 92-894-8466-7. http://www.biomatnet.org/publications/1896rep.pdf
13. Life cycle inventory of biodiesel and petroleum diesel for use in an urban bus. USDA-USDOE, May 1998. http://www.nrel.gov/docs/legosti/fy98/24089.pdf
14. Farrell AE, Plevin RJ, Turner BT, Jones AD, O'Hare M, Kammen DM (2006) Ethanol can contribute to energy and environmental goals. Science 311: 506-508
15. L’ambivalence des filières biocarburants. INRA sciences sociales Decembre 2005. ISSN 0988-3266. http://www.inra.fr/esr/publications/iss/pdf/iss05-2.pdf
16. Oksman-Caldentey KM, Saito K (2005) Integrating genomics and metabolomics for engineering plant metabolic pathways. Curr Opin Biotechnol 16: 174-179
17. Giuliano G, Aquilani R, Dharmapuri S (2000) Metabolic engineering of plant carotenoids. Trends Plant Sci 5: 406-409.
18. Miyake M, Takase K, Narato M, Khatipov E, Schnackenberg J, Shirai M, Kurane R, Asada Y (2000) Polyhydroxybutyrate production from carbon dioxide by cyanobacteria. Appl Biochem Biotechnol 84-86: 991-1002
19. Lebeau T, Robert JM (2003) Diatom cultivation and biotechnologically relevant products. Part II: current and putative products. Appl Microbiol Biotechnol 60: 624-632
20. Rabbani S, Beyer P, Lintig J, Hugueney P, Kleinig H (1998) Induced beta-carotene synthesis driven by triacylglycerol deposition in the unicellular alga Dunaliella bardawil. Plant Physiol 116: 1239-1248
21. A look back at the US DOE’s aquatic species program: Biodiesel from Algae. National Renewable Energy Laboratory TP-580-24190. http://www1.eere.energy.gov/biomass/pdfs/biodiesel_from_algae.pdf

Biotecnologie per l’energia e la chimica
45
22. Hearings, US House Committee on Science. November 6, 2003. Hearing, Dr. Sally M. Benson. http://www.house.gov/science/hearings/energy03/nov06/benson.htm
23. Apt KE, Kroth-Pancic PG, Grossman AR (1996) Stable nuclear transformation of the diatom Phaeodactylum tricornutum. Mol Gen Genet 252: 572-579
24. Kawata Y, Yano S, Kojima H, Toyomizu M (2004) Transformation of Spirulina platensis strain C1 (Arthrospira sp. PCC9438) with Tn5 transposase-transposon DNA-cation liposome complex. Mar Biotechnol (NY) 6: 355-363
25. Tan C, Qin S, Zhang Q, Jiang P, Zhao F (2005) Establishment of a micro-particle bombardment transformation system for Dunaliella salina. J Microbiol 43: 361-365.
26. Xiong J, Bauer CE (2002) Complex Evolution of Photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 53: 503-521.
27. Giacometti G., Giuliano G. (2005) Chimica-fisica e biologia dell’idrogeno. In: Accademia Naz. dei Lincei (ed.) Giornate della Chimica, IV ed.
28. Melis A, Happe T (2001) Hydrogen production. Green algae as a source of energy. Plant Physiol 127: 740-748.
29. Kruse O, Rupprecht J, Bader KP, Thomas-Hall S, Schenk PM, Finazzi G, Hankamer B (2005) Improved photobiological H2 production in engineered green algal cells. J Biol Chem 280: 34170-34177.
30. Van Ooteghem SA, Jones A, Van Der Lelie D, Dong B, Mahajan D (2004) H(2) production and carbon utilization by Thermotoga neapolitana under anaerobic and microaerobic growth conditions. Biotechnol Lett 26: 1223-1232.
31. Handelsman J (2004) Metagenomics: application of genomics to uncultured microorganisms. Microbiol Mol Biol Rev 68: 669-685
32. Streit WR, Schmitz RA (2004) Metagenomics - the key to the uncultured microbes. Curr Opin Microbiol 7: 492-498.
33. Handelsman J, Rondon MR, Brady SF, Clardy J, Goodman RM (1998) Molecular biological access to the chemistry of unknown soil microbes: a new frontier for natural products. Chem Biol 5: R245-249
34. Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, Eisen JA, Wu D, Paulsen I, Nelson KE, Nelson W, Fouts DE, Levy S, Knap AH, Lomas MW, Nealson K, White O, Peterson J, Hoffman J, Parsons R, Baden-Tillson H, Pfannkoch C, Rogers YH, Smith HO (2004) Environmental genome shotgun sequencing of the Sargasso Sea. Science 304: 66-74.
35. Tyson GW, Chapman J, Hugenholtz P, Allen EE, Ram RJ, Richardson PM, Solovyev VV, Rubin EM, Rokhsar DS, Banfield JF (2004) Community structure and metabolism through reconstruction of microbial genomes from the environment. Nature 428: 37-43
36. Luther GW, 3rd, Rozan TF, Taillefert M, Nuzzio DB, Di Meo C, Shank TM, Lutz RA, Cary SC (2001) Chemical speciation drives hydrothermal vent ecology. Nature 410: 813-816.
37. APAT, rapporto rifiuti 2004. http://www.apat.gov.it/site/_contentfiles/00139200/139261_RapportoRifiuti2004Completo.pdf
38. Muller P, Li XP, Niyogi KK: Non-photochemical quenching. A response to excess light energy. Plant Physiol 2001, 125(4):1558-1566.


















