
Biotecnologie Genetiche Agrarie - DistaGenomics · di interesse tramite procedure di clonaggio...
15
Esercitazione di Biotecnologie Genetiche Agrarie “Mappe genetiche”
-
Upload
duongkhanh -
Category
Documents
-
view
216 -
download
0
Transcript of Biotecnologie Genetiche Agrarie - DistaGenomics · di interesse tramite procedure di clonaggio...

Esercitazione di
Biotecnologie Genetiche Agrarie
“Mappe genetiche”

Mappa con marcatori morfologici (sinistra) e mappa con marcatori
molecolari RFLP e SSR (destra) del cromosoma 1 di mais

Produzione di una mappa genetica
La produzione di un mappa genetica passa
attraverso tre fasi:
1. Calcolo della distanza tra tutte le coppie
di loci (geni o marcatori) e valutazione
della presenza di associazione.della presenza di associazione.
2. Suddivisione dei loci in gruppi di
associazione (anche detti di
concatenazione).
3. Ordinamento dei loci all’interno di
ciascun gruppo di associazione.

Calcolo delle distanze tra coppie di loci
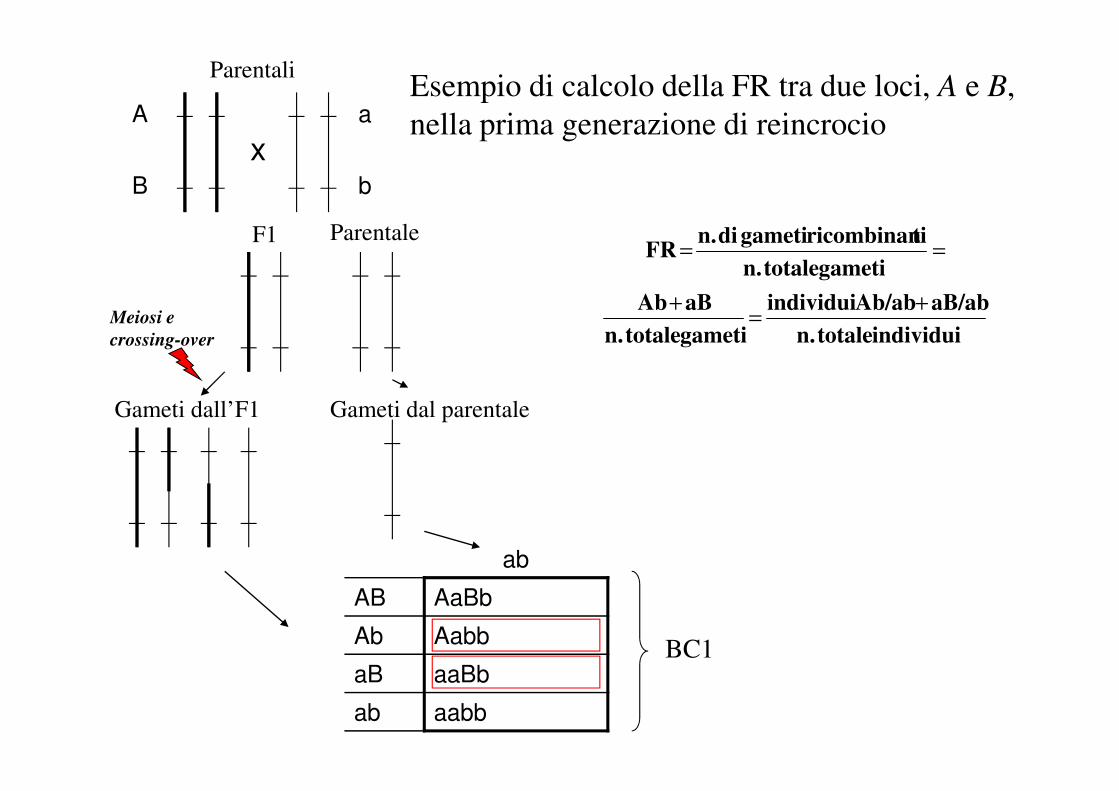
La determinazione della distanza di mappa tra due lociinizia con il calcolo della frequenza di ricombinazione(FR) tra i loci stessi.
FR = proporzione di gameti ricombinanti sul totale deigameti analizzati
0 ≤ FR ≤ 0,5
In semplici popolazioni sperimentali (prima generazione direincrocio o “BC1” ed aploidi raddoppiati) i gametiricombinanti sono riconoscibili dal genotipo degli individui.In altre situazioni (es. popolazioni F2) la proporzione deigameti ricombinanti deve essere stimata con metodi piùcomplessi.
associazione indipendenza
individui totale n.
aB/ab Ab/abindividui
gameti totale n.
aB Ab
gameti totale n.
tiricombinan gameti di n.FR
+=
+
==
Esempio di calcolo della FR tra due loci, A e B,
nella prima generazione di reincrocio
Parentali
ParentaleF1
x
Meiosi e
crossing-over
a
b
A
B
ab
AB AaBb
Ab Aabb
aB aaBb
ab aabb
Gameti dall’F1 Gameti dal parentale
BC1
2 x individui totalen.
ti?ricombinan
gameti totalen.
aB Ab
gameti totalen.
tiricombinan gameti di n.FR
=+
==
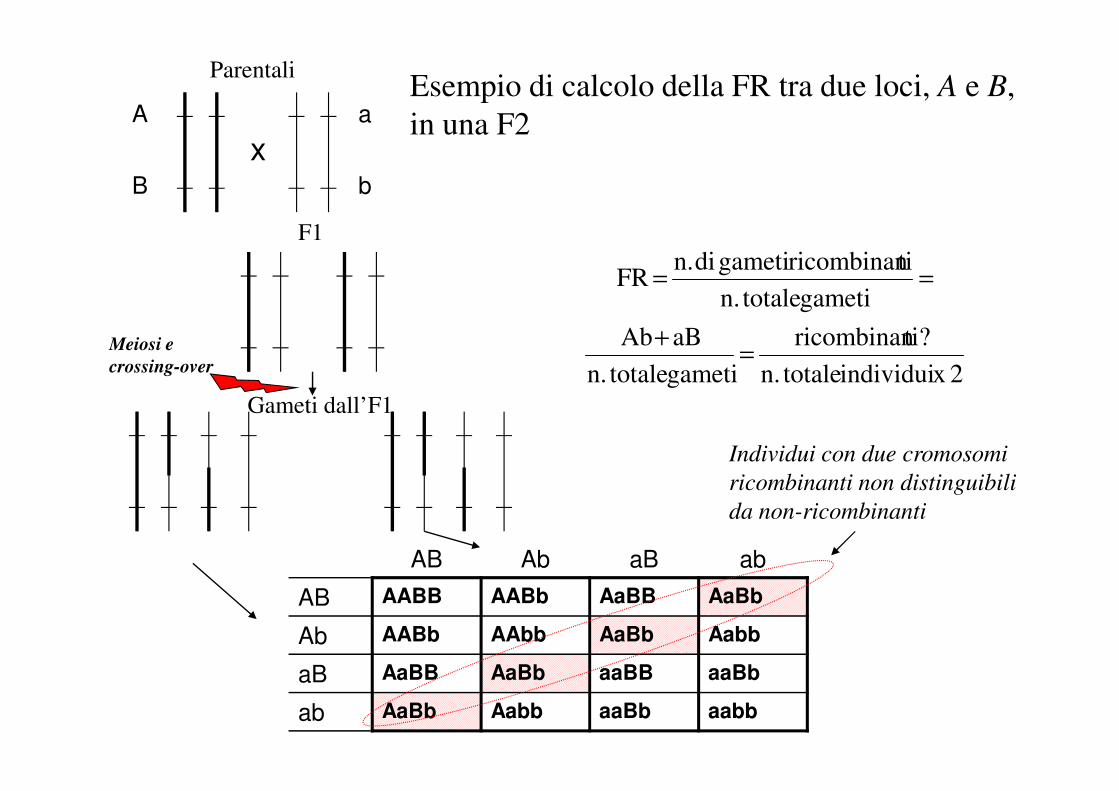
Esempio di calcolo della FR tra due loci, A e B,
in una F2
Parentali
F1
Gameti dall’F1
x
Meiosi e
crossing-over
a
b
A
B
AB Ab aB ab
AB AABB AABb AaBB AaBb
Ab AABb AAbb AaBb Aabb
aB AaBB AaBb aaBB aaBb
ab AaBb Aabb aaBb aabb
Individui con due cromosomi
ricombinanti non distinguibili
da non-ricombinanti
Gameti dall’F1

Valutazione della presenza di associazione:
1. Metodo del test del χ2
Es.: popolazione BC1
Classe fenotipica N. osservati N. attesi (E-O)2/E
AaBb 47 25 19,36AaBb 47 25 19,36
Aabb 4 25 17,64
aaBb 6 25 17,64
aabb 43 25 12,96
Totali 100 67,60
Il valore calcolato del χ2, 67,6 è molto maggiore del valore critico della distribuzione
per 3 gradi di libertà (7,81 per P = 0.05), quindi possiamo rigettare l’ipotesi di non
associazione: i loci A e B sono associati, ad una FR di 0,1 (10%).

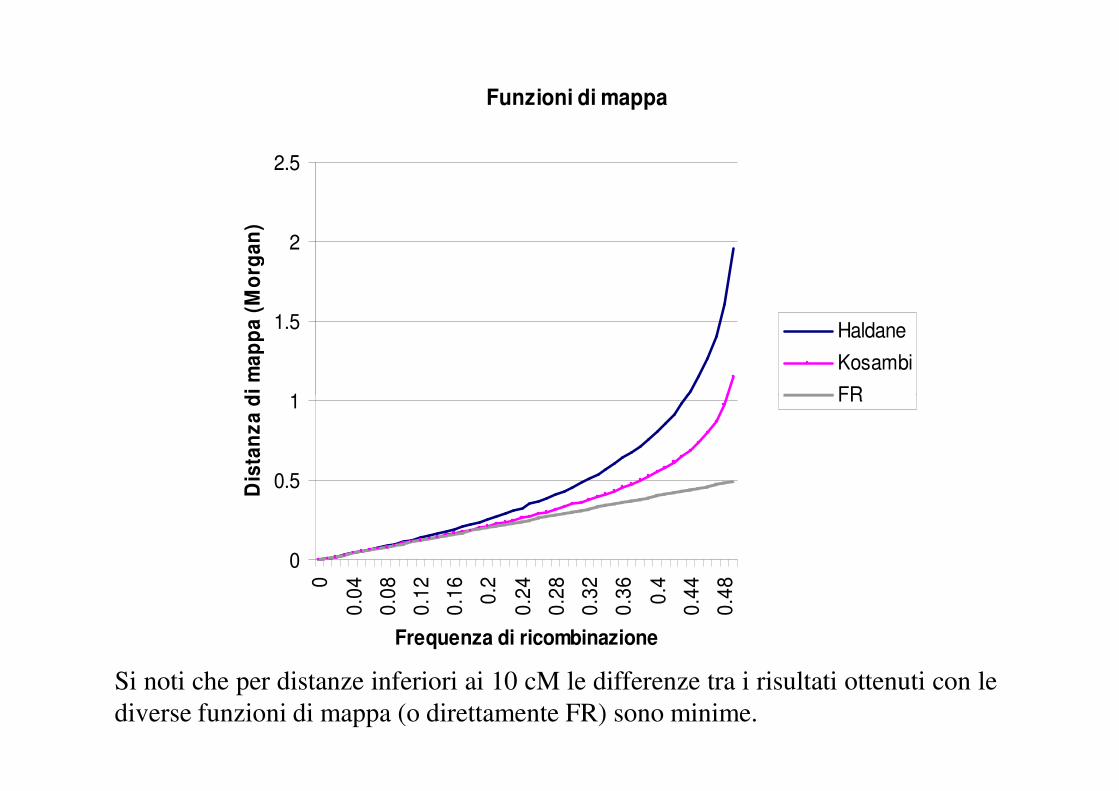
Funzione di mappa
Le distanze tra loci espresse sulla base della FR non sono addittive, in quanto non
sono in grado di tenere conto dei doppi crossover.
Una funzione di mappa è una funzione matematica che tenta di predire il numero
effettivo di crossover tra due loci a partire dal valore di FR.
Alcune possibili progenie di
popolazione BC1 (solo un
cromosoma)
Questi ricombinanti sfuggono al
conteggio quando si considera la
distanza A-B, ma vengono
considerati per A-D e B-D.
A
D
B
effettivo di crossover tra due loci a partire dal valore di FR.
Per una previsione del numero effettivo dei crossover tra due loci occorre fare delle
assunzioni sul livello di interferenza (= i = misura dell’indipendenza di un crossing-over
rispetto ad un altro). La funzione di Haldane assume totale indipendenza degli eventi di
crossover (i = 0), la funzione di Kosambi assume un livello di interferenza intermedio, (i =
1- 2FR), proporzionale ad FR.
Le distanze tra loci basate sul numero di crossover sono maggiormente addittive e
sono riportate sulla mappa come unità di mappa o centimorgan (cM).
2
2FR)-ln(1-(Haldane) cM =
+=
2FR-1
2FR 1ln
4
1(Kosambi) cM
Funzioni di mappa
1
1.5
2
2.5
Dis
tan
za
di
ma
pp
a (
Mo
rga
n)
Haldane
Kosambi
FR
0
0.5
1
0
0.0
4
0.0
8
0.1
2
0.1
6
0.2
0.2
4
0.2
8
0.3
2
0.3
6
0.4
0.4
4
0.4
8
Frequenza di ricombinazione
Dis
tan
za
di
ma
pp
a (
Mo
rga
n)
FR
Si noti che per distanze inferiori ai 10 cM le differenze tra i risultati ottenuti con le
diverse funzioni di mappa (o direttamente FR) sono minime.

Aggiungiamo un terzo
locus (D)….
dove mappa?
Parentali
ParentaleF1
xa
b
A
B
Ordinamento di 3 loci (incrocio a tre punti)
Gameti dall’F1 Gameti dal parentale
Meiosi e
crossing-over
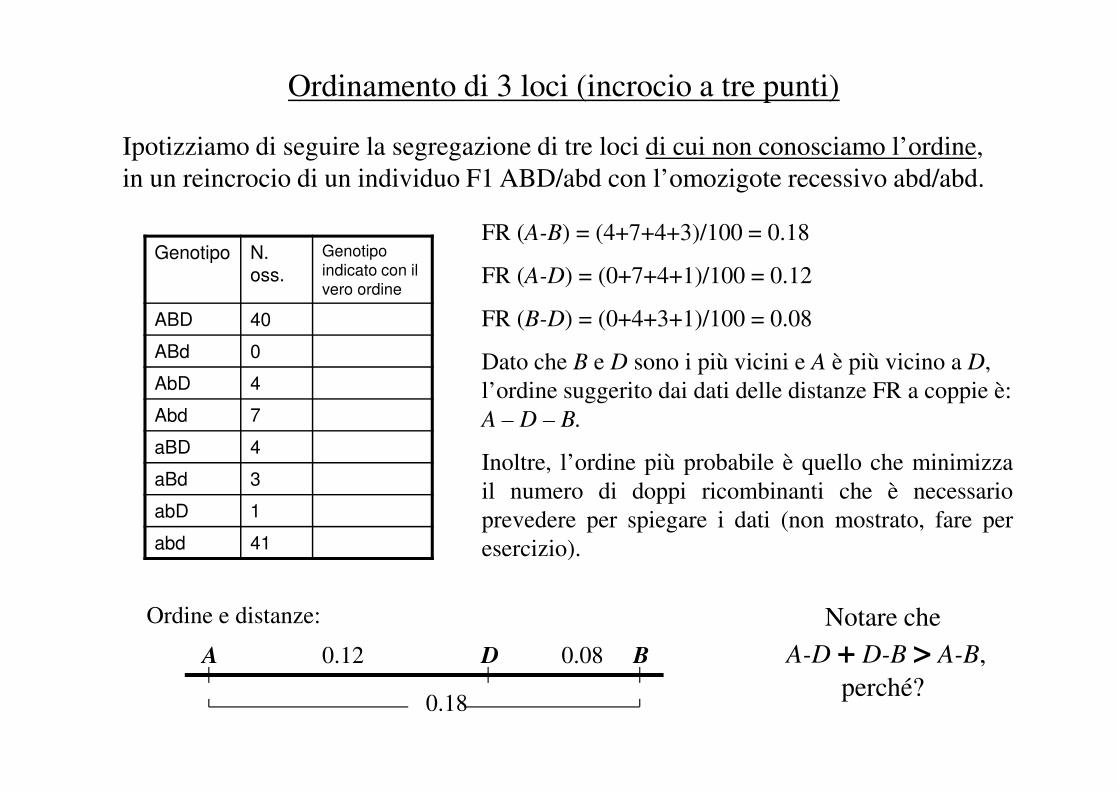
Ordinamento di 3 loci (incrocio a tre punti)
Ipotizziamo di seguire la segregazione di tre loci di cui non conosciamo l’ordine,
in un reincrocio di un individuo F1 ABD/abd con l’omozigote recessivo abd/abd.
Genotipo N. oss.
Genotipo
indicato con il
vero ordine
ABD 40
ABd 0
AbD 4
FR (A-B) = (4+7+4+3)/100 = 0.18
FR (A-D) = (0+7+4+1)/100 = 0.12
FR (B-D) = (0+4+3+1)/100 = 0.08
Dato che B e D sono i più vicini e A è più vicino a D,
l’ordine suggerito dai dati delle distanze FR a coppie è: Abd 7
aBD 4
aBd 3
abD 1
abd 41
l’ordine suggerito dai dati delle distanze FR a coppie è:
A – D – B.
Inoltre, l’ordine più probabile è quello che minimizza
il numero di doppi ricombinanti che è necessario
prevedere per spiegare i dati (non mostrato, fare per
esercizio).
0.12 0.08
0.18
A D B
Notare che
A-D + D-B > A-B,
perché?
Ordine e distanze:

Ordinamento dei loci all’interno di un gruppo di associazione
L’ordinamento di molti loci all’interno di un gruppo di
associazione richiede l’utilizzo di procedure ed algoritmi
avanzati. In generale il compito è quello di arrangiare
linearmente i loci in modo che le distanze risultanti sulla
mappa siano il più possibile in accordo con i dati, già a
disposizione, sulle distanze tra le singole coppie di loci.
Le principali strategie utilizzate sono due:Le principali strategie utilizzate sono due:
a) La minimizzazione del numero di crossover che si accettano
definendo uno specifico ordine di loci (strategia utilizzata dal
programma MapMaker)
b) La minimizzazione delle differenze tra le distanze genetiche
calcolate confrontando le singole distanze a coppie con le
distanze misurate, per le stesse coppie, sulla mappa finale
(strategia utilizzata dal programma JoinMap)

Perché costruire una mappa genetica
La disponibilità di una mappa genetica:
1. funge da punto di partenza per l’isolamento di geni
di interesse tramite procedure di clonaggio
posizionale;
2. consente di ottimizzare la scelta di marcatori per lo2. consente di ottimizzare la scelta di marcatori per lo
svolgimento di procedure di selezione assistita da
marcatori molecolari;
3. consente l’identificazione di regioni cromosomiche
sede di loci che controllano i caratteri quantitativi
(QTL: Quantitative Trait Loci);
4. facilita il trasferimento di informazione tra specie
sfruttando le relazioni di sintenia tra le mappe
genetiche.

Esempio di profilo AFLP su
popolazione di mappa in maisPrimer combination: PstGA/MseCGG
Popolazione: F2 (67 piante) dall’incrocio delle linee pure di mais Lo964 (P1) x Lo1016 (P2)
P1
P2

Esempio di file che raccoglie i dati genotipici, da
utilizzarsi per il calcolo di una mappa di linkage tramite
il software JoinMap.


















