Bio-anatomia delle vie ottiche -...
22
Introduzione Per comprendere compiutamente le sofisticate finalità strutturali della grande via sensitiva attor- no alla quale si è costruita nei millenni la civiltà dell’uomo, bisogna cercare di capirne la progetta- zione. È necessario allora risalire all’origine della luce e degli esseri elementari che da essa traggo- no letteralmente la vita. L’architettura delle vie ottiche segnano, passo per passo, l’evoluzione degli organismi viventi sul pianeta, ampliandosi, modificandosi, adattandosi sulla spinta di una pro- spettiva teleologica condizionata da circostanze peristatiche e dall’enorme implementazione delle informazioni sensoriali. Prospettiva sempre in bilico, parafrasando Jaques Monod, tra il caso e la necessità. Vedere è molto di più che percepire gli stimoli luminosi, le stupefacenti strutture della via ottica ne sono la esplicita attestazione materializzata. Si rendono perciò indispensabili i riferimenti retro- spettivi o paralleli alla filogenesi, ontogenesi, organogenesi, anatomia comparata; discipline tutte che troppo spesso sfiorano appena i giovani nei banchi dell’Università, lasciando un sottile “We are highly visual creatures” Charles Darwin “La morfologia è l’immagine plastica della funzione” Angelo Ruffini PARTE CLINICA Bio-anatomia delle vie ottiche Giuseppe Carella · Luigi Cavanna · Egidio Carella · Angelica Cerulli · Giovanni Peretto
Transcript of Bio-anatomia delle vie ottiche -...

Introduzione Per comprendere compiutamente le sofisticatefinalità strutturali della grande via sensitiva attor-no alla quale si è costruita nei millenni la civiltàdell’uomo, bisogna cercare di capirne la progetta-zione. È necessario allora risalire all’origine dellaluce e degli esseri elementari che da essa traggo-no letteralmente la vita. L’architettura delle vieottiche segnano, passo per passo, l’evoluzionedegli organismi viventi sul pianeta, ampliandosi,modificandosi, adattandosi sulla spinta di una pro-spettiva teleologica condizionata da circostanzeperistatiche e dall’enorme implementazione delleinformazioni sensoriali.Prospettiva sempre in bilico, parafrasando JaquesMonod, tra il caso e la necessità. Vedere è molto di più che percepire gli stimoliluminosi, le stupefacenti strutture della via otticane sono la esplicita attestazione materializzata. Sirendono perciò indispensabili i riferimenti retro-spettivi o paralleli alla filogenesi, ontogenesi,organogenesi, anatomia comparata; disciplinetutte che troppo spesso sfiorano appena i giovaninei banchi dell’Università, lasciando un sottile
“We are highly visual creatures”
Charles Darwin
“La morfologia
è l’immagine plastica della funzione”
Angelo Ruffini
PARTE CLINICA
Bio-anatomia delle vie otticheGiuseppe Carella · Luigi Cavanna · Egidio Carella · Angelica Cerulli · Giovanni Peretto

senso di nostalgia per le scienze biologiche purein chi, ha poi intrapreso un più pragmatico emeno soffice indirizzo clinico. Non ci sembrano perciò mal spesi tali riferimen-ti propedeutici, specie se appena abbozzati.Come non ci sembra uno sterile ed irrichiestoesercizio il seguire, per grandi cenni, l’evoluzio-ne del pensiero scientifico sul tema, percorsoperaltro illuminato dai nomi più prestigiosi dellastoria della conoscenza. Pensiamo, anzi, che i quadri di bio-anatomia cheoggi l’imaging consente e che consegneremo allavostra attenzione, possano guadagnare in vivez-za interpretativa ed acquistare un retrogustoculturale, che ci auguriamo gradevole. Solo con la comparsa della luce sulla terra, labiologia potrà scrivere la prima pagina della vita.Il “fiat lux” quindi, fu il fiat vita della Creazione.Tutto ciò incominciò ad accadere nell’eraArchezoica, quando ogni essere vivente iniziò adimparare ad interagire con la luce dalla quale eranato. Dapprima organizzando siti foto-sensoriali(figura 1) che servivano da recettori di impulsiper risposte elementari e stereotipate, e poistrutturando sistemi e percorsi sempre più com-plessi atti non solo a registrare, ma ad articola-re, elaborare ed integrare l’energia luminosa: levie ottiche. In alcuni protisti unicellulari, il corpo della cellu-la è differenziata in due poli opposti: uno dista-le, dove si organizzano specializzazioni cuticola-
ri adatte alla ricezione dello stimolo l’altro pros-simale che si prolunga in un filamento protopla-smatico che si può considerare la più sempliceespressione di fibra nervosa. Questa può colle-garsi con una seconda cellula, configurando cosìun sistema dove il primo corpo cellulare assumeil ruolo di ricettore ed il secondo la funzione dieffettore dello stimolo, a lui trasferito dalla fibradi interconnessione. Se poi un terzo elementocellulare si inserisce tra i precedenti, assumen-do il ruolo di trasformare lo stimolo ricevutodalla prima cellula, prima di trasmetterlo allaseconda, si può in esso riconoscere il primo rudi-mento di sistema nervoso che consente la realiz-zazione di un arco riflesso completo. Già neicelenterati più infimi, quali le idromeduse onelle amebe, si individua il disegno di vie otticherudimentali. Le cellule di senso sono situate insuperficie nell’epitelio;le ganglionari, allogate aldi sotto di questo, collegano le prime con gliorgani reagenti centrali. Salendo la scala zoolo-gica, si assiste vieppiù alla cefalizzazione del cer-vello ed alla periferizzazione degli organi visivi disenso ad esso connessi. Questo grazie alla seg-mentazione metamerica del corpo.Le reazioni che la luce determina in tutti gliorganismi viventi, dai più elementari ai più com-plessi, costituiscono una delle più entusiasman-ti osservazioni scientifiche di ogni tempo. La sua influenza sul movimento è un fenomenotra i più appariscenti e conosciuti: il fototropismo
26 PARTE CLINICA
Figura 1Luce su biostrutture primo arco riflesso
FOTONELa eccitabilità di una cellula determina in essa una serie di risposte, attraverso unmacchinario molecolare che le è proprio. Il risultato sarà la generazione di un impulsobio-elettrico cioè un segnale stereotipato, a velocità finita, estremamente propagabile

(φοτοζ = luce τρεποζ = direzione) degli organi-smi sessili, specie vegetali, che si voltano verso ilsole; la fototaxis (ταξιζ = movimento traslatorio)di certi organuli che corrono verso la luce; lafotokinesi = φοτοκινεσιζ = movimento accelera-torio) di altri, che sollecitano il loro abituale com-portamento motorio in presenza e direzione diuna fonte luminosa di alcuni insetti e molluschi,che si orientano verso la luce con un angolo diincidenza costante (angolo α), configurando spi-rali sorprendentemente logaritmiche. È di comu-ne osservazione che altri insetti si gettano irresi-stibilmente verso sorgenti luminose incande-scenti suicidandosi. Il prodigioso senso di orien-tamento degli uccelli (navigational sense) nelleloro trasmigrazioni, a volte intercontinentali, èdovuto al fatto che il loro sistema visivo usa lesorgenti di luce naturali (sole, stelle di primagrandezza) come un compasso geografico checonsegna loro infallibili coordinate di direzione. Jacques Loeb (1859-1924), il Linneo del regnoanimale, sostiene addirittura che tutti gli esseriviventi sono sostanzialmente macchine chimico-meccaniche azionate dalla energia della luce. Il biologo di Wurzburg, da buon tedesco, affermache il movimento di un animale è coordinato
dalla differenza quantitativa di stimolo luminosotra i due occhi, per cui si attiverebbero solo gliapparati locomotori di un emi-soma, determi-nando così lo spostamento automatico verso unadirezione obbligata. Un’altra grande influenzache la luce esercita su ogni cosa che abbia vita èil fotoperiodismo, cioè un ritmo biologico a cuiogni essere vivente, uomo incluso, deve sotto-stare, modulando il suo comportamento durantele stagioni e, sopratutto, nell’arco del giorno, conun eterno bilanciamento tra il dì e la notte. Alleorigini della vita i bioritmi venivano regolati nondalla luce, ma dalla temperatura del pianeta. Allafuoriuscita dallo stagnante mare caldo delPaleozoico, per guadagnare la terra, gli organi-smi viventi hanno subito per milioni di anni ildramma esistenziale delle radicali variazioni ter-miche, tra il giorno torrido (in cui tutte le attivi-tà metaboliche erano esaltate) e la gelida notte(durante la quale i processi energetici venivanodrasticamente ridotti). Solo alcune specie dianfibi, rettili ed uccelli sono riuscite a sviluppa-re la condizione di omeotermia, mentre gli ani-mali pecilotermi (ad esempio i grandi sauri) sisono estinti. Con il raffreddamento del pianeta ela conseguente variazione della sua atmosfera, il
27Bio-anatomia delle vie ottiche
Figura 2Foraminifero Copilia quadrata: via ottica primitiva
ritmo biologico venne orchestrato non più dallealternanze termiche, ma della luminosità. Le vie visive quindi diventarono il tramite fonda-mentale per la sopravvivenza e non stupisce quin-di che queste si siano evolute, adattate e semprepiù specializzate nei millenni (figura 2). È notoche nei mammiferi superiori, la crasi ematica(emoglobina, ematocrito, proteine plasmatiche)subisce variazioni significative nel corso del nicte-mero, come pure la temperatura corporea.Nell’uomo si rileva eosinopenia nelle prime ore delmattino, mentre negli animali notturni invertebra-ti si ha eosinofilia. Le mitosi cellulari sono partico-larmente esaltate nel periodo di riposo (curiosa-mente le cellule neoplastiche non obbediscono aquesta legge). L’oftalmologo d’altra parte sa chel’oftalmotono subisce variazioni nell’arco nicteme-rale. Gli intimi meccanismi che regolano questibioritmi scanditi dalle condizioni di luce non sono,a tutt’oggi, soddisfacentemente acclarati. Si sup-pone che, verosimilmente, l’asse diencefalo-ipofi-sario moduli le condizioni neuro-ormonali dell’ani-male, sull’informazione degli stimoli luminosi chepervengono ai suoi centri superiori attraverso levie ottiche. Per quanto riguarda gli animali inferio-ri, è noto sin da Aristotele (384-322 a.C.) che i lorocicli riproduttivi sono strettamente correlati allecondizioni di luce che ne stimolano le gonadi.Le grandi migrazioni stagionali degli uccelli, nonhanno sostanzialmente altro scopo che quello diportare le coppie in luoghi dove la luce è tale etanta da poter essere convenientemente veicola-ta attraverso le vie ottiche ai centri neuro¬endo-crini, la cui attività garantisce la conservazionedella specie. Da ultimo va segnalata l’influenzadelle radiazioni luminose sulla distribuzione deipigmenti ai vari livelli, da quello cutaneo a quel-lo retinico. La dispersione pigmentaria nella fasediurna e la concentrazione in quella notturna,obbedisce alle leggi di un bioritmo finalistico cheintende affievolire e proteggere le funzionidurante il periodo di riposo.
Profilo storico generaleLa storia delle vie ottiche corre ovviamente nel-l’alveo di conoscenza delle scienze mediche edin particolare della neuro-anatomia e della oftal-
28 PARTE CLINICA
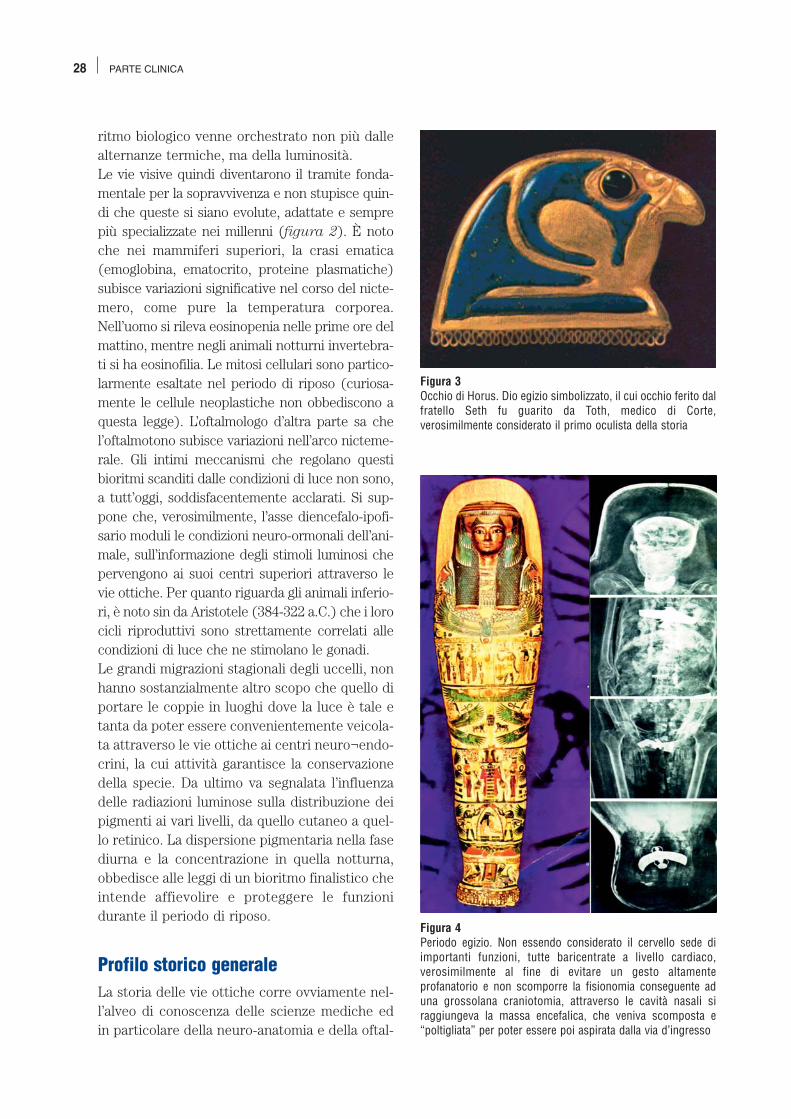
Figura 3Occhio di Horus. Dio egizio simbolizzato, il cui occhio ferito dalfratello Seth fu guarito da Toth, medico di Corte,verosimilmente considerato il primo oculista della storia
Figura 4Periodo egizio. Non essendo considerato il cervello sede diimportanti funzioni, tutte baricentrate a livello cardiaco,verosimilmente al fine di evitare un gesto altamenteprofanatorio e non scomporre la fisionomia conseguente aduna grossolana craniotomia, attraverso le cavità nasali siraggiungeva la massa encefalica, che veniva scomposta e“poltigliata” per poter essere poi aspirata dalla via d’ingresso

Figura 5Papiro di Ebers, 1550 a.C. Prende il nome dal suo acquirente,l’inglese John Ebers, che lo comprò a Tebe nel 1873. Lungo 20metri e largo 20 centimetri e suddiviso in 108 pagine, è databilealla XVII dinastia egizia nel regno di Amenhotep. Si rilevanoosservazioni prevalentemente bulbari con accenno ad un“peduncolo posteriore”. Conservato al Museo dell’Universitàdi Lipsia
mologia, ma con percorsi cronologici abbastanzadifferenziati. Le origini della scienza oftalmologica sono avvol-te nella leggenda e ci rimandano all’antico Egittodi 5000 anni fa.La tradizione indica il medico egizio Thot comeil primo oculista della storia: fu lui, infatti,come vuole la leggenda, a guarire l’occhio diHorus (figura 3), il dio sparviero ferito dal fra-tello Seth.L’Oftalmologia, al pari di altre branche della
Medicina, nasce e si sviluppa in Egitto, cometestimoniano dipinti, bassorilievi, oggetti scoper-ti nelle tombe dei faraoni (figura 4), ma soprat-tutto, i papiri più famosi. Quello di Ebers (1500a.C.) (figura 5) è il più ricco di notizie oftalmo-logiche: gli oculisti egiziani, come Pepi Ankh Ri,Medu Nefere, Khuy, erano noti in tutto il mondoallora conosciuto. Nella stessa epoca, l’oftalmologia si sviluppavanel medio oriente, in Mesopotamia, come ricor-dato in alcuni paragrafi della stele del codice diHammurabi (1792-1730 a.C.) (figura 6). Dall’Egitto e dalla Mesopotamia, attraverso laScuola alessandrina, le conoscenze di oftalmolo-gia vengono trasferite in Grecia e successiva-mente ai romani, alla Gallia e, grazie ai mediciarabi, giungeranno all’occidente in epocamedioevale. Le prime notizie fondate sulla concezione anato-mica delle vie ottiche risalgono alla civiltà elleni-ca ed in particolare ai pensatori ionici del VIsecolo avanti Cristo. Il primo riferimento storico si riscontra negliscritti di Alcmeone, medico di Crotone vicinoalla Scuola pitagorica, vissuto nel 500 a.C.. Egli,per primo, affermò l’esistenza di una connessio-ne tra nervo ottico e cervello, e formulò la teo-ria, allora rivoluzionaria, secondo cui era il cer-vello (e non il cuore) l’organo centrale delle
29Bio-anatomia delle vie ottiche
Figura 6Codice di Hammurabi. È il primo codice organico di 300 leggi che riguardano diversi ambiti della vita sociale. Tra queste vengonoillustrate le più conosciute malattie oculari ed i relativi provvedimenti terapeutici
Hammurabi (1792-1750 a.C.). Il nome del re babilonese deriva dall’amorreo Amnu
che significa “guarire”
Stele Testo
facoltà sensitive e motorie. Il nervo otticodescritto da Alcmeone consisteva in un tubocavo in cui lo “pneuma” (figura 7 a, b, c, d) cir-colante nell’occhio, considerato come il princi-pio della visione, fluiva dal cervello. Una descrizione grossolanamente più dettagliatadell’organo della visione ci è trasferita dallaScuola atomistica, che aveva in Leucippo diMileto (440 a.C.) ed in Democrito di Abdera(460-370 a.C.) i suoi maggiori esponenti. ADemocrito è attribuita la prima descrizione delbulbo oculare, da lui concepito come una strut-tura relativamente semplice, costituita da duestrati: uno esterno, la sclero-cornea, ed unointerno, la corio-retina; in essi fluiva un umoreomogeneo convogliato da un condotto, il nervoottico, in accordo con la teoria di Alcmeone. Nonvi è, a quell’epoca, una neppure approssimativaconcezione delle vie ottiche poste distalmente alnervo ottico. Nel 460 a.C, nasce a Kos Ippocrate, e con lui lavera scienza medica, in quanto veniva sostituitoalla superstizione ed alle leggende un metododescrittivo-empirico, volto alla ricerca dellecause naturali delle malattie. Se Ippocrate è universalmente considerato ilpadre della Medicina, Aristotele (384-322 a.C.) èconsiderato il fondatore della Storia Naturale edella Anatomia Comparata. Il grande filosofo diStagira, basandosi su studi anatomici (dissezioni
di occhi animali), fornì la più completa descrizio-ne, per l’epoca, della anatomia macroscopicaoculare. L’occhio era descritto come una sferacostituita da tre strati intimamente giustappostil’uno sull’altro, e riempita da un fluido omoge-neo, connesso con il cervello da tre tubi cavi,uno dei quali in collegamento con l’occhio con-trolaterale. In questa descrizione si possono pro-babilmente riconoscere nuove strutture anato-miche quali il nervo trigemino, i vasi oculari, ilchiasma ottico. In epoca romana, sia Rufo di Efeso che CornelioCelso (25 a.C.-50 d.C.), diedero nei loro scrittiuna descrizione più sistematica del sistema ner-voso centrale e delle connessioni oculo-cerebra-li attraverso il nervo ottico. Tali descrizioni nonaggiungono però nulla di nuovo rispetto alle con-cezioni greche, sembrando essere una merarielaborazione di testi già conosciuti. In epoca imperiale, Claudio Galeno di Pergamo(131-201 d.C.) (figura 8), considerato, conIppocrate, il più grande medico dell’antichità,descrisse il nervo ottico come una struttura cavaed anch’egli sostenne che proprio nel nervo otti-co scorrevano gli “umori” in eccesso, causa dellemalattie oculari. Se questa concezione anatomi-ca ricalca le teorie tradizionali, a Galeno si deveuna teoria della fisiologia della visione del tuttonuova e, sebbene molto lontana dalla realtà,destinata a rimanere pressoché inalterata per
30 PARTE CLINICA
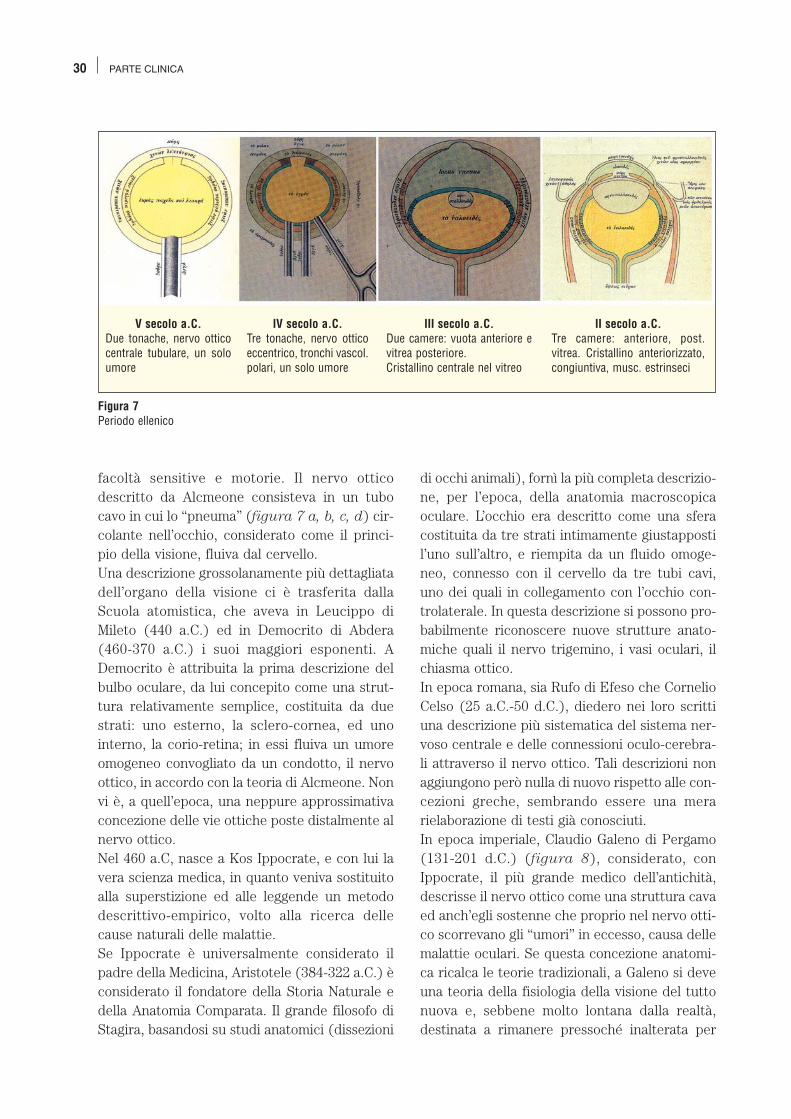
Figura 7Periodo ellenico
V secolo a.C.Due tonache, nervo otticocentrale tubulare, un soloumore
IV secolo a.C.Tre tonache, nervo otticoeccentrico, tronchi vascol.polari, un solo umore
III secolo a.C.Due camere: vuota anteriore evitrea posteriore. Cristallino centrale nel vitreo
II secolo a.C.Tre camere: anteriore, post.vitrea. Cristallino anteriorizzato,congiuntiva, musc. estrinseci

molti secoli. Egli riteneva che la retina fosseun’espansione del nervo ottico deputata a nutri-re il corpo vitreo, il quale a sua volta era il soste-gno trofico del cristallino; proprio il cristallinoera considerato come l’organo essenziale dellavisione (divinum oculi): in esso si formavano lesensazioni visive, e da esso venivano inviateverso gli oggetti da cui ritornavano al cristallino,per essere condotte lungo il nervo ottico al ven-tricolo laterale omolaterale, ritenuto la sede del-l’anima. I nervi ottici non subivano una decussa-zione a livello di un chiasma, ma erano semplice-mente accostati e collegati in modo che lo “spi-rito visivo” di emanazione cerebrale, potesse dis-tribuirsi ai due occhi così da ottenere una visio-ne binoculare singola; se un occhio era danneg-giato o perso, la capacità visiva dell’altro venivaaumentata. Solamente nell’undicesimo secolo, Ibn Rushd(Averroè) (1126-1198) (figura 9), dotto araboiberico, filosofo e medico, sostenne che fosse la
retina, dotata di fotorecettori, e non il cristalli-no, la sede di origine delle sensazioni visive, con-vogliate al cervello dal nervo ottico cavo. AncheAverroè che, per dirla con l’Alighieri, “il grancommento feo”, tuttavia, non postulò l’esistenzadi altre strutture anatomiche relative alle vieottiche. Durante il Rinascimento, Leonardo daVinci (1452-1519) (figura 10), avvalorando ingran parte le teorie di Galeno, descrisse la strut-tura cava del nervo ottico in collegamento noncon il ventricolo laterale, ma con il centrale deitre ventricoli, sede (secondo la cultura medioe-vale) delle tre principali facoltà mentali (“cellu-la phantastica” per la percezione, “cellula logisti-ca” per la ragione, “cellula memorialis” per lamemoria). Leonardo introdusse una concezionefisiologica del meccanismo della visione di gran-de progresso. Abbandonando il concetto del cri-stallino organo principale della visione, egli con-cepì e disegnò la luce provenire dall’esterno,passare attraverso la pupilla, compiere una dop-
31Bio-anatomia delle vie ottiche
Figura 8Claudio Galeno
Figura 10Leonardo, 1490. Nervo ottico tubulare, emergente al polo posteriore. Vescicole cerebrali: sensazione, percezione, elaborazione
Figura 9Periodo arabo. Rhazes (865-925 d.C.)

pia refrazione nella parte mediana del bulbo alivello del cristallino, e cadere direttamente sullaparte posteriore del globo oculare in corrispon-denza dell’emergenza del nervo ottico.Leonardo, dunque, spostò l’attenzione sulla reti-na ed suoi fotorecettori, ritenendo per primo,nella cultura occidentale, che questa strutturafosse la sorgente della visione. Nello stessoperiodo, Gregor Reisch (figura 11) consegnò lasua memorabile concezione delle 3 “facultates”memo-percettive. Fu il medico ed anatomistabelga Andrea Vesalio (1514-1565), grazie adaccurate dissezioni, a dubitare dell’esistenza diuna cavità all’interno dei nervi ottici; Vesaliodescrisse nervo ottico e chiasma; struttura ana-tomica, quest’ultima, forse dotata di una cavitàinterna e dove, comunque, le vie ottiche non siincrociavano, ma erano apparentemente collega-te. Bartolomeo Eustachi (1500-1574), contem-poraneo di Vesalio, non mise in discussione lacavità dei nervi ottici; tuttavia a suo merito vaascritta l’ipotesi che l’origine di tali nervi fosse laporzione posteriore del talamo e non le paretidei ventricoli cerebrali. Nel secolo successivo, Cartesio (1596-1650)(figura 12) finalmente smentì l’inveterata con-cezione del nervo ottico come struttura tubula-
re cava. Nel suo “Discorso sul Metodo” (1637),riprese la teoria dell’’origine delle vie ottiche daiventricoli laterali e descrisse le fibre del nervoottico decorrere parallele senza decussarsi alchiasma, per terminare in punti omologhi dellaretina. Le retine, stimolate dalla luce, liberavano“spiriti visivi” che seguivano le vie ottiche fino aiventricoli e da qui venivano trasmessi alla ghian-dola pineale, ritenuta la sede dell’anima. A livel-lo epifisario, le impressioni visive monoculari simiscelavano per formare una singola immagine
32 PARTE CLINICA
Figura 11Gregor Reisch (margarita philosophica, 1503)Le tre cavità cerebrali, senso percettive
Figura 12René Décartes, 1664. Les voies visuelles.

visiva binoculare, depositata poi nel cervellocome substrato mnemonico. Sempre nel ’500,Vesalio e Varolio descrivono la faccia inferioredell’encefalo dove si rilevano chiari decorsi dellevie ottiche.Thomas Willis (1621-1675), scopritore del circo-lo anastomotico arterioso cerebrale che porta ilsuo nome, ebbe il merito di fornire una più det-tagliata descrizione delle vie ottiche chiasmati-che e retro-chiasmatiche allora conosciute. Eglisottolineò l’origine anatomica dei nervi ottici(ormai considerati come solidi fasci di fibre) daltalamo ottico, dove si riteneva terminassero levie ottiche. William Briggs (1650-1704) (figura
13), professore a Cambridge ed al St. ThomasHospital di Londra, ripropose lo schema anato-mico disegnato da Willis e per primo descrisse laconvergenza retinica delle fibre ottiche sullapapilla.Gli anatomisti Zumbo, Pinson, Celenzuolo edancora lo stesso Briggs evidenziarono il fatto chele fibre ottiche, indecussate al chiasma, sembra-vano avere origine da due protuberanze del tala-mo ottico (i corpi genicolati). Questa regioneanatomica venne considerata come la sede dellasensazione visiva, originata dalle vibrazioni reti-niche indotte dalla luce, e trasmesse dalle fibreottiche che vibrerebbero in sintonia. La ricerca anatomo-funzionale sulle vie ottichesubì una fase di stasi che durò oltre un secolo.
Solo Raymond Vieussens (1641-1716) ipotizzòl’esistenza di una via ottica retro-talamica. Èmerito di Jean Pierre Marie Flourens (1794-1867) l’aver fornito la dimostrazione che l’abla-zione selettiva di alcune parti della cortecciacerebrale influiva sulla funzione visiva e che,pertanto, il talamo non era la sola sede della per-cezione visiva; le vie ottiche non potevano finirenel talamo, ma dovevano continuare fino allacorteccia cerebrale. Parallelamente presero lucei risultati delle ricerche di Gautier d’Agoty(1775) e Tarin (1750).Diversi studiosi antecedenti a Flourens, comeGennari (1782), Vicq d’Azyr (1786-89) e Reil(1809-12) avevano individuato numerose strut-ture anatomiche all’interno dell’encefalo, descri-vendo sempre più dettagliatamente i sistemi difibre cerebrali grossolanamente osservati dalWillis: per primo Reil individuò il sistema genico-lo-calcarino, senza però dame una interpretazio-ne funzionale. Fu Pierre Gratiolet (1833-1892)(figura 14), professore di anatomia allaSorbona, a delimitare ancor più dettagliatamen-te la connessione anatomica genicolo-corticale,descrivendo le radiazioni omonime sopratuttonella parte posteriore, mentre Theodor Meynert(1833-1892) (figura 15) precisò il loro decorsonella porzione anteriore. Ma il pioniere dellamoderna ricerca anatomica in questo campo puòessere considerato Bernhard Aloys Von Gudden
33Bio-anatomia delle vie ottiche
Figura 13William Briggs, 1650

(1824-1886) che, attraverso minuziose indaginicondotte su sottili sezioni di tessuto cerebraleottenute con microtomia fine, dimostrò inconfu-tabilmente la parziale decussazione delle fibreottiche nel chiasma, già ipotizzata da IsaacNewton nel ‘700 e William Hyde Wollaston (figu-
ra 16) nell’800. Egli inoltre descrisse la com-messura ottica superiore ed il tratto peduncola-re trasverso, strutture anatomiche oggi denomi-nate con i suoi eponimi. Infine, fu tra i primi afar luce sulla complessa organizzazione anato-mo-funzionale del talamo, osservando la degene-razione selettiva retrograda dei suoi nuclei dopodistruzione sperimentale di aree circoscritte dicorteccia cerebrale. Le scoperte di Von Gudden furono avvaloratedalle ricerche di Palli Emil Flechsig (1847-1929), che condusse un esauriente studio sullamielinogenesi dei sistemi di fibre corticali.Osservando che il processo della mielinizzazioneavviene in diversi momenti della vita fetale ecoinvolge cronologicamente sistemi ben precisidi fibre, Flechsig riuscì a suddividere la cortec-cia cerebrale in aree sensitivo-motorie ed inaree associative. Basandosi sul dato che il siste-ma delle fibre sensoriali visive al polo occipitaleera il primo a circondarsi di mielina, esaminò leradiazioni ottiche di Gratiolet per dimostrare
come tali fibre terminassero nella cortecciaoccipitale, in una regione circoscritta denomina-ta “area striata” da EIlit Smith (1904). Nellostesso periodo in cui si attuava la sistematizza-zione anatomo-topografica delle vie otticheposteriori ricerche parallele venivano dedicate
34 PARTE CLINICA
Figura 14Pierre Gratiolet (1815-1865). Professore di anatomia a Parigi, descrissenel 1856 la connessione genicolo-corticale,a cui diede il nome. Illustrazione dallacopertina di “memoires de la SociétéFrançaise d’Anthropologie”
Figura 15Theodor Meynert (1833-1892). 1858, prima descrizione della laminazionecorticale
Figura 16William Hyde Wollaston (1766-1828). Chimico di Cambridge, ipotizzò la parzialedecussazione chiasmatica delle fibre ottiche,studiando su se stesso una formaintermittente di emianopsia omonima, divutaad un tumore talamico di cui morì

alle vie ottiche anteriori, Otto Deiters rivendica-va assertivamente la individualità morfo-funzio-nale del neurone (1865) e Vicq d’Azyr distinsele fibre visive retro-chiasmatiche come “trattoottico”, suddividendo la parte extracerebraledelle vie ottiche nelle porzioni a cui ancora oggisi fa riferimento. Nell’ultima parte del XIX secolo, vari studiosistabilirono rigorosamente le connessioni tra leterminazioni dei nervi ottici e le tre principalistazioni sotto-corticali del sistema visivo (pulvi-nar talamico, corpo geni colato laterale ed colli-colo superiore). Salomon Henschen (1847-1930) e Mieczyslaw Minkowsky (1884-1934),dimostrarono la struttura lamellare del corpogenicolato laterale e la connessione ordinata diquesti strati con le terminazioni delle fibre deitratti ottici. Nel febbraio del 1904, FriedrichDimmer (figura 17), dimostrando la connessio-ne delle fibre ottiche ascendenti al corpo geni-colato laterale ed, in piccola parte, al collicolosuperiore, negò l’arresto di queste stesse fibre altalamo vero e proprio, come fino ad allora lascienza ufficiale supponeva. In questo modo latopografia generale delle vie ottiche veniva ana-tomicamente stabilita; gli ulteriori progressi sisarebbero limitati allo studio della cito-tettonicacorticale più fine e delle aree associative, grazie
al miglioramento delle tecniche di conservazio-ne dei tessuti e di osservazione dei preparatiistologici (connessione talamo-corticale di VonMonakov nel 1910).Alla fine del XIX secolo, ed ancor più negli annisuccessivi, l’attenzione dei ricercatori si trasferìdalla anatomia vera e propria del sistema visivoalla sua organizzazione funzionale: sono statedefinite la rappresentazione corticale delle fibreottiche, la funzione del giro angolare nei movi-menti di deviazione coniugata dello sguardo, l’e-sistenza di cecità psichica in seguito a lesioni diaree corticali non occipitali, la possibilità di dif-ferenti tipi di anopsie in rapporto alle diversesedi di lesione. All’inizio del XX secolo è stata precisata l’attua-le rappresentazione retinotopica nelle vie otti-che e la distribuzione assonomica delle fibreottiche nei vari segmenti. Dalla metà del ‘900, la micro-anatomia, l’elettro-fisiologia e la citochimica si sono fuse in un pro-digioso insieme finalistico di ricerche che haaperto prospettive radicalmente innovative. È in queste prospettive che si stanno scrivendole nuove pagine della storia delle vie ottiche, icui temi continueranno a mobilitare i protagoni-sti più illuminati delle scienze bio-mediche.
Filogenesi delle vie otticheQuando, nel 1865, Ernst Haeckel stabiliva.che laontogenesi (ον = individuo, γενεσιζ = sviluppo)è la breve ricapitolazione della filogenesi(φυλοζs= specie), voleva forse anche significareche l’impronta degli echi filogenetici lontanisulla successione delle fasi embriologiche, rap-presentano un prezioso filo conduttore per com-prendere la morfologia delle strutture anatomi-che definitive per l’oggi evoluzionale.L’assunto è particolarmente calzante per le con-figurazioni delle vie ottiche che solcano tuttol’encefalo, con il ruolo di grande baricentro sen-soriale a sua volta condizionante e correlante lealtre neuro-strutture. A questo proposito corre alla mente la teoriadella neuro-biotaxis, enunciata nel 1909 daCornelius Arien Kappers (figura 18), dalla cat-tedra di neuro-anatomia dell’Università di
35Bio-anatomia delle vie ottiche
Figura 17Friedrich Dimmer (1855-1926). Nel febbraio 1904, dimostrò la connessionedelle fibre ottiche ascendenti con il corpogenicolato laterale, negando l’arresto dellestesse al talamo, come era sino ad allorasupposto dalla scienza ufficiale

Amsterdam: nel divenire filogenetico, le vie ed icentri nervosi si sono raggruppati, affiancati,correlati, sistematizzati, in riferimento alle lorosuperiori funzioni comuni. Ad esempio, i nucleidei nervi oculomotori si sono gradatamenteposti in relazione con il fascicolo longitudinaleposteriore ed il sistema vestibolare, quest’ultimoin parallelo sviluppo con le vie ottiche. La pro-gressiva sempre maggiore decussazione dellefibre ottiche al chiasma con il salire la scala zoo-logica, va letta come la risposta alla necessitàfilogenetica di far decorrere fianco a fianco vieche provengono da regioni (le emiretine omoni-me) che lavorano in concomitanza. Negli inver-tebrati e nei vertebrati inferiori ogni nervo disenso o di moto fa capo ad un proprio centro dif-ferenziato, centro che si specializza in rapportoalle esigenze peristatiche proprie dell’animale edella sua attività biologica. Quando queste atti-vità diventano complesse, i centri sentono l’esi-genza di coordinarsi tra loro e di riferirsi vieppiùa centri di comando superiori. Le cellule senso-riali visive degli invertebrati si dispongono sullasuperficie ectodermica e solo secondariamentesi connettono con il sistema nervoso centrale(occhio tegumentale). All’opposto, nei vertebrati la retina origina dal-l’ectoderma neurale come sua estroflessone(occhio cerebrale).
Come già accennato, ci sembra interessantetratteggiare per schemi l’evoluzione delle vieottiche nel loro svolgersi filogenetico, delinean-do così il profilo di una opso-neuro-biotaxis cherappresenta le radici di un albero morfologicoche ci apprestiamo, molto succintamente adaccennare. Nei ciclostomi (pre-pesci sopravvissuti di anti-chissimi vertebrati del paleozoico), le fibre otti-che che provengono dalle cellule visuo-sensoriali,sono direttamente proiettate nel tectum opticum(soffitto del mesencefalo primitivo), dove entranoin relazione con le vie nervose spino- e bulbo-tet-tali, ed inoltre con quelle del V e VIII paio. Nei pesci compare un abbozzo rudimentale dicorpo genicolato laterale. Gli assoni visivi termi-nano ancora al tectum ed ai suoi prolungamentidorsali, che costituiscono i lobi ottici, particolar-mente evidenti. I nervi ottici sono molto svilup-pati, come pure il midollo allungato ed i nucleidel V e VII paio. Fa apparizione il lemnisco late-rale ed il fascio longitudinale posteriore, cherappresenta la prima importante via di intercon-nessione tra centri di senso e centri motori. Negli anfibi le fibre del II paio fanno stazione nelcorpo genicolato laterale, da cui si dipartonofibre genicolo-tettali che terminano ai lobi ottici. Nei rettili compare un primo accenno di cortec-cia (corteccia olfattoria). In connessione con le
36 PARTE CLINICA
Figura 18Cornelius Ubbo Ariens Kappers (1877-1946). Professore di neuro-anatomia - Università di Amsterdam

cellule del pallium, si osservano fasci di fibre ner-vose che afferiscono ai gangli della base e chevengono considerati come il primitivo abbozzodella corona raggiata. I lobi ottici sono piuttostosviluppati, discreta l’evidenziazione dei talami;accenno ai tubercoli quadrigemelli posteriori inalcuni di essi, ed alla grande via del lemnisco. Negli uccelli si ha un enorme sviluppo dellaparte superiore del cervello anteriore e del cer-vello medio, con lobi ottici cospicui. Gli emisfericerebrali coprono gran parte dei lobi ottici,lasciandone scoperta solo un’area laterale. Ilsistema talamico è ora prettamente fotostatico.La decussazione dei nervi ottici è completata. Sisistematizzano le vie di interconnessione, grazieal fascio longitudinale posteriore. Fasci di asso-ciazione collegano i lobi ottici con la cortecciaoccipitale, i talami ed il tetto mesencefalico conil cervelletto ed il midollo. Fibre intra-corticalicreano rapporti tra le varie aree della corteccia.In sostanza, vi è comunicazione ed integrazionetra i centri di moto cerebrali e spinali, nonchécon la corteccia l’inizio di quella organizzazionearticolata e complessa, che troverà la sua massi-ma espressione nei mammiferi superiori. Nei mammiferi il progressivo aumento dell’ence-falo, che abbiamo sin qui seguito nelle primeclassi dei vertebrati, continua si specializza viep-più dagli inferiori ai superiori. Tale sviluppo ècaratteristicamente disomogeneo, privilegiandoalcuni distretti e penalizzandone altri che siarrestano od addirittura regrediscono. La maggior parte delle fibre ottiche (80%) siporta al nucleo dorsale del corpo genicolato late-rale, da cui poi si proiettano verso la cortecciavisiva. II nucleo ventrale del CGL, filogenetica-mente più antico, perde progressivamente diruolo man mano che il baricentro della via visivasi sposta dal tetto alla corteccia. Dei corpi geni-colati, si accrescono principalmente i laterali adetrimento dei mediali. Le vie visive dei verte-brati inferiori dedicano scarsi contingenti al CGL,terminando la maggior parte ai corpi quadrigemi-ni. Nei superiori si ha una netta inversione di ten-denza, man mano che si perfeziona la visione cor-ticale, in rispetto alla legge che Ludwig Edingerformulò nel 1897. I quadrigemini sono estrema-mente ridotti d’importanza nei confronti degli
imponenti lobi ottici dei vertebrati più bassi. Inervi ottici sono meno sviluppati che nelle classiprecedenti, ma evidenziano chiaramente la lorocorrelazione con i tre nuclei della base: genicola-to laterale, corpo quadrigemello anteriore e tala-mo (pulvinar). A queste vie ottiche “basilari”, siaggiungono le superiori o “corticali” che costitui-scono il ventaglio di fibre ottiche corticipete, for-mato dalle radiazioni di Gratiolet. Nei mammife-ri si ha un imponente incremento volumetrico,soprattutto in superficie, degli emisferi cerebrali.La corteccia, liscia nei mammiferi inferiori (lis-sencefali), diventa anfrattuosa e convoluta neisuperiori (girencefali). A Bartolomeo Panizza va riconosciuta la paterni-tà della scoperta del centro corticale della visio-ne. Nel 1855, dopo aver accecato un occhio dialcuni piccoli pesci ed uccelli, notò ad un annodi distanza non solo l’atrofia del nervo ottico cor-rispondente, ma anche della metà di quello dellato opposto. Le sue ricerche seguirono quelle diLuigi Rolando il quale dimostrò, nel 1828, chealcuni selaci ed anfibi privati del cervello ante-riore non manifestavano significative alterazionidel senso visivo, senso gravemente compromes-so invece dalla ablazione dei lobi ottici. La circoscrizione delle funzioni delle aree corti-cali si ha soltanto nei primati. Nei mammiferimeno evoluti le varie zone di competenza sonosempre meno circoscritte e specializzate. Nei cani, il centro visivo arriva fin quasi in zonafrontale e deborda anche ai lati ed al di sottodella regione occipitale. Nelle scimmie è estesoanche al giro angolare. Nell’uomo è limitatoattorno alla scissura calcarina. Gli emisferi cerebrali sono costretti ad aumenta-re sempre più il loro volume per contenere fibree centri nervosi che lo sviluppo filogenetico for-nisce. La corteccia è obbligata a contorcersi suse stessa per rannicchiarsi nell’angusta cavitàcranica. A lei giungono e da lei partono tutte leinformazioni che regolano le innumerevoli fun-zioni dell’animale. Al senso dell’odorato (anima-li “osmatici”) ed al senso della vista (animali“opsici”) non viene dedicato come per gli altrisensi, un nervo, ma addirittura una porzione del-l’encefalo, tanta è l’importanza che queste infor-mazioni estesiche rivestono negli ipovertebrati. I
37Bio-anatomia delle vie ottiche

nervi olfattivi e ottici non si possono comunqueconsiderare nervi, ma fasci di fibre nervose, ana-logamente a quelle che formano le commessurefra le diverse parti del cervello. Negli animaliopsici più evoluti, il nervo ottico poi originadirettamente dal diencefalo e si ammanta diguaine meningee. Per cui si è tentati di affermare che “il nervoottico non è un nervo, ma un cervello lungo”. Alla fine di questo breve profilo filogeneticodelle vie ottiche, è importante sottolineare che,agli albori, il sistema visivo aveva come baricen-tro il tectum, crocevia elementare, ma fonda-mentale, degli impulsi statici, ottici, tattili,gustativi e propriocettivi. Con lo snodarsi dell’e-voluzione, il baricentro si sposta sempre più inalto, verso le regioni talamiche ed infine cortica-li, dove si realizzano le funzioni visive epicriti-che, denobilitando il precedente a mere funzio-ni riflessogene fotostatiche. La rimozione del cervello olfattivo di un pesce odi un rettile non influenza la funzione visiva. Larana decerebrata continua a cacciare agevol-mente le mosche, il piccione decorticato evitaperfettamente gli ostacoli, denunciando solo unaffievolimento delle facoltà di ricognizione piùalte. La rimozione della corteccia di un mammi-fero lo rende praticamente cieco. Quindi, il gran-de evento nel corso del tormentato arco dellafilogenesi è rappresentato dal passaggio di pre-dominanza dal cervello olfattivo a quello visivo. Lo svilupparsi della via ottica come protagonistadelle funzioni più evolute, ha quindi condiziona-to la morfologia e la fisiologia e il comportamen-to degli organismi viventi che, per mezzo suo,hanno avuto la possibilità di passare dal ruolo diesseri condizionati a quello di esseri creativi.
Ontogenesi delle vie ottichePer comprendere convenientemente questoargomento, si rende indispensabile un ricordopropedeutico di embriologia generale. Allo stadio di neurula (tre-quattro settimane), lavescicola ectodermica si appiattisce per costitui-re una specie di piastra (placca neurale) dappri-ma rotonda, poi progressivamente ellittica, dis-ponendosi lungo un asse che sarà la lunghezza
del corpo. Su questa placca dapprima compareuna striscia di cellule opache (stria primitiva),che termina nella sua parte anteriore con un pic-colo ispessimento bottonuto (nodo di Hensen).Nella parte anteriore della placca si individuanodue piccole depressioni (fossette ottiche). In seguito i bordi della stria si sollevano costi-tuendo il solco neurale, che poco a poco si chiu-de per formare il tubo neurale (primo abbozzodel sistema nervoso). Verso la fine della quartasettimana, il tubo subisce tre dilatazioni che rea-lizzano così le tre vescicole cerebrali primitive(prosencefalo, mesencefalo, rombencefalo). Ilsegmento cefalico si inflette secondo due curva-ture: la cefalica e la cervicale. Alla quinta setti-mana, la prima vescicola si sdoppia (telencefalo,diencefalo), la seconda rimane tale (mesencefa-lo) e la terza pure si divide (metencefalo e mie-lencefalo). Sempre nel corso della quinta setti-mana, gemmano dalla vescicola diencefalica dueespansioni (vescicole ottiche primitive) che,strangolandosi a livello dei loro peduncoli, costi-tuiranno i futuri nervi ottici. Le vescicole ottiche subiscono rapidamente uninfossamento al loro apice, trasformandosi incupole a concavità esterna. Come la costituzionedelle vescicole è indotta dalla presenza delmesoderma precordale, che è situato sotto laplacca neurale, così il loro infossamento a cupo-la è indotto dalla presenza dell’abbozzo del cri-stallino, che le antistà. La cupola è costituita dadue foglietti accollati: dal primo si differenzieràla retina sensoriale, dal secondo l’epitelio pig-mentato.L’invaginazione della vescicola ottica avvienesecondo una modalità particolare, finalizzata aporre in comunicazione occhio ed encefalo: lafessura embrionale. Questa infatti deve conside-rarsi come un “espediente embriologico” tempo-raneo (da 4-5 mm. a 15-20 mm. di lunghezza del-l’embrione) che rimane aperta per 4-5 settima-ne, al fine di connettere la retina al cervello tra-mite il peduncolo ottico. Attraverso di essa haaccesso l’arteria jaloidea, che entra alla estremi-tà prossimale della fessura stessa. Ida Mann (figura 19), attraverso lo studio dimodelli ipotetici, ha dimostrato che tale espe-diente era l’unico che poteva essere messo in
38 PARTE CLINICA

atto per conseguire le finalità morfogenetichedovute. Prima di passare alla descrizione del-l’embriogenesi dettagliata dei vari segmentidelle vie ottiche, merita una riflessione la parti-colare ed apparentemente irrazionale disposizio-ne della retina negli animali superiori. Lo spartiacque morfologico differenziale tra l’oc-chio epiteliale degli invertebrati e l’occhio cere-brale dei vertebrati, è costituito dalla localizza-zione dei fotorecettori nello spessore retinico.Nei primi, le cellule epiteliali sensoriali si molti-plicano e si infossano nella loro parte distale adiretto contatto con la luce, mentre le fibre ner-vose delle parti prossimali veicolano poi lo sti-molo luminoso verso le strutture cerebrali (reti-na vertita). Nei secondi invece, la retina gemmadal diencefalo e la modalità di infossamentodella vescicola ottica primitiva in cupola, porta ifotorecettori nella profondità dello spessore reti-nico, per cui la luce deve attraversare tutti gli
strati loro antistanti prima di attivarli (retinainversa). È suggestivo rilevare (come piccola ricapitola-zione filogenetica haeckeliana) che, nelle primefasi dell’embriogenesi umana, a livello delle fos-sette ottiche, le cellule ectodermiche (epiteliali)sono situate all’esterno (come negli invertebra-ti), mentre nelle fasi successive, con la costitu-zione del tubo neurale e delle vescicole ottiche,queste vengono invaginate e disposte all’interno. Si diceva poc’anzi che la retina inversa può sem-brare un atteggiamento irrazionale nella sofisti-cata organizzazione istogenetica dei vertebratisuperiori. Ma la retina inversa ha buone ragionifinalistiche per essere tale: il fotorecettore, con-siderate le sue raffinate esigenze funzionali,abbisogna di un imponente e massivo apportoossigenativo, che solo l’adiacenza della corioca-pillare può garantire. Inoltre il contatto direttocon l’epitelio pigmentato (mangia-dischi) hagrande importanza nel loro metabolismo. Ed èchiaro che, per ragioni ottiche di trasparenza, siala coriocapillare che l’epitelio pigmentato nonpossono essere che situati dietro le cellule visuo-sensoriali. Da ultimo, la disposizione invertitaconsente un aumento del potere risolutivo cen-trale, aprendo gli strati istologici davanti ai foto-recettori, per costituire la fovea (l’occhio dell’oc-chio).
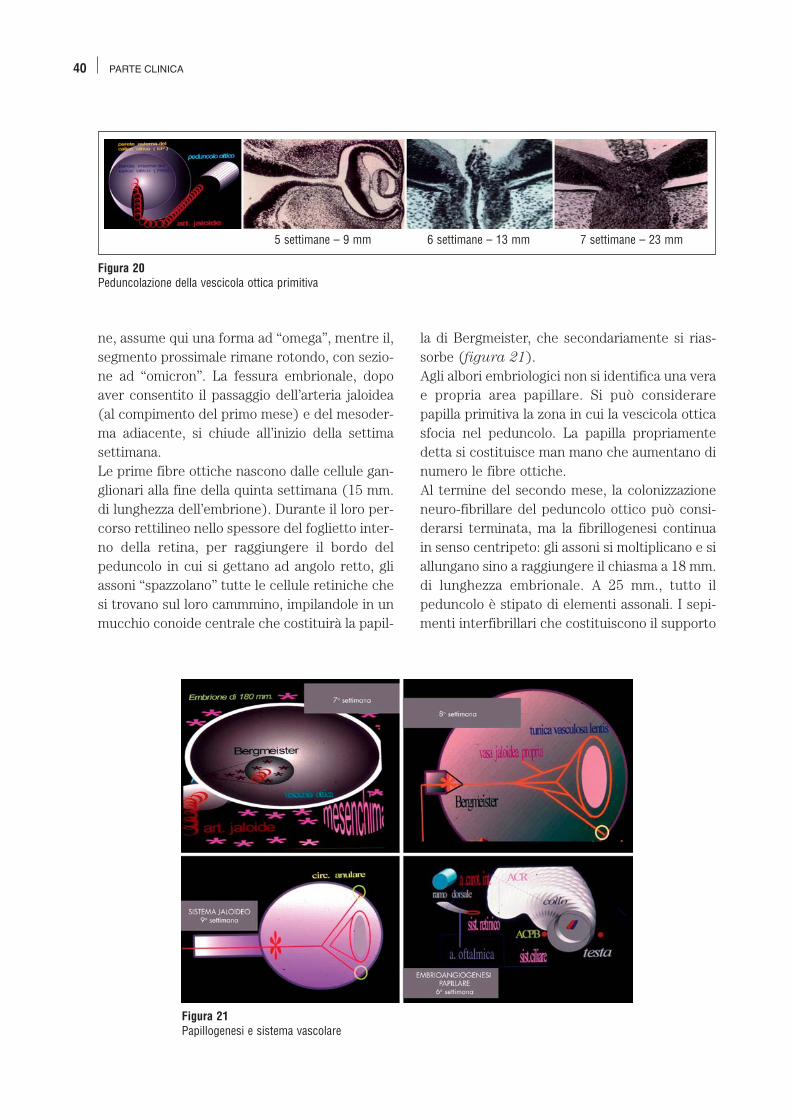
Embriogenesi del nervo ottico Abbiamo in precedenza visto che, nel corso dellaquinta settimana di vita embrionale, la vescicolaottica primitiva, gemmata dalla vescicola dience-falica, si peduncolizza progressivamente. Ilpeduncolo è un tubo sostanzialmente tappezza-to da un monostrato di cellule epiteliali cilindri-che che mette in comunicazione le cavità cere-brali e la futura cavità oculare (figura 20). Laparete peduncolare servirà da guida al costi-tuendo nervo ottico, alla cui formazione contri-buirà offrendo i sepimenti neurogliali interfibril-lari. La fessura embrionale interessa il pedunco-lo connettendolo con il foglietto invaginato, cheverrà a costituire il futuro strato interno dellaretina. L’invaginazione fissurale interessa solo laparte distale del peduncolo ottico che, in sezio-
39Bio-anatomia delle vie ottiche
Figura 19Ida Mann. Necessità ontogenetica della fessura embrionaria.Modello ipotetico. La fessura embrionale è un meraviglioso“espediente embriologico” che realizza e tutela l’evoluzioneontogenetica
ne, assume qui una forma ad “omega”, mentre il,segmento prossimale rimane rotondo, con sezio-ne ad “omicron”. La fessura embrionale, dopoaver consentito il passaggio dell’arteria jaloidea(al compimento del primo mese) e del mesoder-ma adiacente, si chiude all’inizio della settimasettimana. Le prime fibre ottiche nascono dalle cellule gan-glionari alla fine della quinta settimana (15 mm.di lunghezza dell’embrione). Durante il loro per-corso rettilineo nello spessore del foglietto inter-no della retina, per raggiungere il bordo delpeduncolo in cui si gettano ad angolo retto, gliassoni “spazzolano” tutte le cellule retiniche chesi trovano sul loro cammmino, impilandole in unmucchio conoide centrale che costituirà la papil-
la di Bergmeister, che secondariamente si rias-sorbe (figura 21). Agli albori embriologici non si identifica una verae propria area papillare. Si può considerarepapilla primitiva la zona in cui la vescicola otticasfocia nel peduncolo. La papilla propriamentedetta si costituisce man mano che aumentano dinumero le fibre ottiche. Al termine del secondo mese, la colonizzazioneneuro-fibrillare del peduncolo ottico può consi-derarsi terminata, ma la fibrillogenesi continuain senso centripeto: gli assoni si moltiplicano e siallungano sino a raggiungere il chiasma a 18 mm.di lunghezza embrionale. A 25 mm., tutto ilpeduncolo è stipato di elementi assonali. I sepi-menti interfibrillari che costituiscono il supporto
40 PARTE CLINICA
Figura 20Peduncolazione della vescicola ottica primitiva
5 settimane – 9 mm 6 settimane – 13 mm 7 settimane – 23 mm
Figura 21Papillogenesi e sistema vascolare

gliale e derivanti, come già detto, dalle celluledell’epitelio primitivo, nella parte distale delnervo si orientano perpendicolarmente allefibre, abbozzando al secondo mese la primalamina cribrosa (Haden, 1947). All’inizio costi-tuita essenzialmente da nevroglia, la lamina sidota, dal quarto al settimo mese, di elementiconnettivo-elastici. Nel corso del secondo mesedi vita embrionale, il mesoderma peri-pedunco-lare si condensa in due strati: l’uno esterno,spesso ed avascolare (dura madre, 65 mm.), l’al-tro interno, sottile e vascolarizzato (pia madre,5° mese). L’aracnoide si differenzia fra i due soloal 6°-7° mese, quale tessuto lacunare derivantedalla nevroglia periferica. Nel corso del 5° mesesi ha un notevole incremento in lunghezza delnervo che passa da 3 a 8 mm. (Hervouet, 1958).All’8° mese, l’architettura del II paio può consi-derarsi definitiva e completa. Alla nascita, la sualunghezza è di circa 24 mm. per 2 mm. di diame-tro. Alla pubertà, è lungo circa 40 mm con undiametro di 4 mm.
Embriogenesi del chiasma e della bandelletta ottica Prima del 2° mese di vita embrionale, la zonachiasmatica è semplicemente costituita daammassi cellulari della parete ventricolare deldiencefalo, posta tra i due peduncoli ottici. Talicellule serviranno in seguito come apparatoneuro-gliale di sostegno alle fibre ottiche, chearrivano in questa zona solo verso la 7° setti-mana (22 mm.). In seguito, le fibre visive cir-cumnavigano la parte laterale del diencefaloper raggiungere un raggruppamento cellulareche si è differenziato dalla parte dorso lateraledel talamo, al fine di costituire il nucleo dorsa-le del Corpo Genicolato Laterale (30 mm.).Queste fibre si moltiplicano e si ispessisconoper costituire il tratto ottico (bandelletta), cheè bene evidente a 48 mm. di lunghezza embrio-nale (figura 22). Un’altro esempio di ricapito-lazione filogenetica dell’ontogenesi, è fornitodai tempi di decussazione delle fibre al chia-sma. Sino alla 11a settimana (59 mm. di lun-ghezza embrionale) le fibre ottiche non siincrociano (come negli invertebrati). Solo alla
13a settimana (80 mm.), la decussazione hainizio (Sakamoto, 1952).
Embriogenesi delle vie ottiche centrali In estrema sintesi cercheremo qui di enuc1eare,dal complesso divenire embriogenetico del siste-ma nervoso centrale, lo sviluppo di quelle aree edistretti che sono toccati od ospitano le variestrutture della via ottica. Il mesencefalo (sede delle principali funzioni visi-ve nei vertebrati inferiori e relegato a sede diriflessi fotostatici e visuo-sensoriali nei superio-ri), prende origine dalle pareti della vescicolamesencefalica primitiva e si stipa attorno all’esi-le acquedotto di Silvio. La parte dorsale dell’ac-quedotto, che corrisponde al tetto del tubo neu-rale, si differenzia in tectum opticum (vedi anchecapitolo riguardante la filogenesi). A questo livel-lo compaiono due elevazioni longitudinali, a lorovolta poi divise da un solco in quattro tubercoli,chiamati appunto quadrigemelli per la loro simi-larità, e dei quali i superiori (collicoli visivi) fannoda perno ai riflessi fotomotori distinguendosifunziona1mente dagli inferiori (collicoli auditivi). Il prosencefalo, vescicola primitiva dell’estremocefalico dell’embrione, si divide precocemente(11 mm di lunghezza embrionale) in una vesci-cola prossimale (diencefalica) ed una distale(telencefalica). Il diencefalo, circondante quel tratto di canaleneurale che si dilata per formare il 3° ventricolo,contiene diverse strutture di pertinenza visiva. A
41Bio-anatomia delle vie ottiche
Figura 22Embriogenesi del chiasma e del tratto ottico

13 mm (5 settimana), si assiste ad una intensaproliferazione cellulare della parete laterale, chepoco dopo (16 mm) viene suddivisa da due sol-chi in tre zone: il talamo centralmente, sormon-tato dall’epitalamo e sovrastante l’ipotalamo. Inqueste tre zone hanno sede importanti ammassicellulari che costituiscono nuclei di grandeimportanza funzionale.Di nostro particolare interesse è il corpo genico-lato laterale (CGL), situato nella parte esternadel talamo dorsale e già distinguibile a 22 mm,epoca in cui questa zona viene raggiunta dallefibre ottiche del tratto, che probabilmente neinducono la strutturazione. Dapprima si differenzia il nucleo dorsale delCGL e solo secondariamente il ventrale (35mm). La tipica architettura laminare non si rea-lizza che al 6° mese di vita intrauterina, quandola formazione si inarca caratteristicamente ad“U” rovesciata con apertura latero-ventrale. Un breve cenno alla embriologia dell’ipofisi che,pur non appartenendo alla via ottica, frequente-mente ne condiziona la patologia. La neuroipofisitrae origine dalle cellule gliali del pavimento dien-cefalico, che alla quinta settimana (8 mm) evagi-na un piccolo diverticolo. L’adeno-ipofisi nasceinvece dalla borsa di Rathke, diverticolo ectoder-mico che gemma dal pavimento stomodeum. Leporzioni anteriore e posteriore dell’ipofisi vengo-no in contatto a 14 mm di lunghezza embrionale. A 9 mm, la parte più anteriore del prosencefalosi espande nelle due vescicole telencefaliche, lecui pareti costituiranno gli emisferi cerebrali e lecavità i ventricoli laterali. La faccia basale delle pareti si ispessisce per for-mare la zona striata, entro cui trovano posto inuclei del corpo striato. La parte laterale delcorpo striato prende il nome di “paleo-pallium”,primo abbozzo di corteccia a finalità puramenteolfattiva; come pure destinato a funzioni olfattivee reflessogene è “l’archi-pallium” che, a 12 mm,si espande centralmente attorno alle cavità ven-tricolari. Tutto il resto del telencefalo va a costi-tuire la grande massa del “neo-pallium”, nella cuizona marginale i neuroblasti si moltiplicano, for-mando la materia grigia corticale. La differenzia-zione corticale si realizza dapprima (30 mm) alivello dell’area parietale, per guadagnare preco-
cemente la superficie mediale e superiore del-l’encefalo. Solo più tardivamente (50 mm) ven-gono interessate le aree frontale e, soprattutto,occipitale (visiva). Quindi, ad ennesima confer-ma dell’assunto di Ernst Haeckel, dapprima siorganizzano i centri olfattivi, poi quelli della sen-sibilità somatica ed infine quelli della visione. Nel prematuro di 28 settimane, la corteccia occi-pitale è praticamente lissencefalica, riconoscendo-si a stento solo la fessura calcarina e parieto-occi-pitale. L’estensione dell’area striata si quadruplicada questa data sino al compimento del 9° meseregolare di gestazione (girencefalia) e si espandevieppiù sino al 4° mese di vita extrauterina.Alla nascita, i dendriti appaiono spinosi cioèprovvisti. di coni di crescenza, che conferisconoall’assone un aspetto villoso. Le spine aumenta-no progressivamente sino al 4° mese per scom-parire definitivamente al 9°. Il periodo neo- nata-le è caratterizzato da una intensa proliferazionesinaptica a livello di tutti gli strati della cortec-cia visiva. Questa “sinaptogenesi” diminuisceprogressivamente sino ad un anno di vita, perridursi al 60% all’età di l0 anni. Ciò perché allaelaborazione sinaptica anatomica dell’età neona-tale, si sostituisce una sinaptogenesi funzionale,poiché si organizzano i circuiti associativi chematerializzano le “ipercolonne di orientamento”così caratteristiche della corteccia visiva, per laformazione delle quali è peraltro fondamentalel’esperienza visiva (epigenesi). Le colonne corti-cali di orientamento visivo sono estremamentesensibili, quindi, alle penalizzazioni sensoriali. Sesi considera che la plasticità neuronale è parti-colarmente formativa nell’arco di tempo com-preso tra la nascita ed il 70 anno, è intuitivo ildanno irreversibile che si determina nel bambi-no in cui non è stata per tempo depistata unadisfunzione visiva.
MielinogenesiIl processo di mielinizzazione delle vie otticheinferiori avviene nel senso inverso a quello delloro sviluppo. Al 5° mese, la mielina compare alivello del corpo genicolato, al 6° della bandellet-ta, al 7° raggiunge il chiasma ed all’8° il nervoottico. Per le vie ottiche centrali, la mielinizza-
42 PARTE CLINICA

zione inizia generalmente all’epoca della nascitae procede centrifugamente dall’area occipitale,per completarsi verso la fine del 40 mese di vitaextrauterina. Per molto tempo si è voluto correlare la mieliniz-zazione di un segmento delle vie ottiche alla suafunzionalità. Oggi si propende a negare tale cor-relazione, in considerazione del fatto che vi sonoassoni ancora spogli di guaina mielinica e giàfunzionanti mentre, per converso, neurofibre giàmielinizzate non sono ancora in grado di veico-lare lo stimolo nervoso.
Cenni di anatomia comparata Lo studio della bio-anatomia delle vie otti-che, pensiamo non possa sottrarsi alla curio-sità di vedere come queste sono strutturateai vari livelli della scala zoologica (figura
23, figura 24). È sorprendente infatti scoprire come l’atto visi-vo si serva di strumenti ed artifizi tanto fantasio-si e diversificati nella loro unicità teleologica.Anche in questo campo è il caso di dire che ilbuon fine giustifica i mezzi.E di questi ne daremo qualche rapido cenno
salendo, come si conviene, la scala dal bassoverso l’alto. Ciclostomi = (κυκλοζ = rotonda, στομα = bocca)Il nervo ottico ha fibre non mielinizzate chedecorrono solo al centro del nervo, senza sepi-menti intersettali. Il chiasma è intracerebrale edi nervi ottici si incrociano in modo elementare,senza mostrare alcuna fascicolazione al lorointerno. Pesci = Il nervo è avascolare. Le fibre sonomielinizzate e variamente sepimentate.Assenza della lamina cribrosa. A livello delchiasma le fibre nervose assumono la confor-mazione a vimini intrecciato. Nei teleostei ilnervo ottico visto dall’esterno appare tondeg-giante o piatto. All’interno assume un partico-lare aspetto nastriforme; i sepimenti interfa-scicolari sono così sviluppati da suddividere ifasci in tante falde che lo fanno assomigliaread un nastro ripetutamente ripiegato su sestesso. Nel perioftalmo, ed ancor più nello sta-filoftalmus paradoxus, il nervo, al centro di unlunghissimo cono di sottili muscoli extraocula-ri, è contenuto in un altrettanto lungo pedun-colo alla sommità del quale è posto il bulbooculare. Totale decussazione al chiasma a sem-
43Bio-anatomia delle vie ottiche
Figura 23Via ottica comparata

plice sovrapposizione crociata, ad asola o adoppia asola. Anfibi = (ανφο = doppia; βιοζ = vita)Esseri di transizione tra la primitiva vita acquaticae la successiva terrestre, nell’era carbonifera ave-vano dimensioni gigantesche. Le loro fondamenta-li tappe evolutive sono consistite nella trasforma-zione delle pinne in arti, delle branchie in polmonie cuore tricamerale; inoltre adattamento dell’orec-chio alle vibrazioni dell’etere e dell’occhio allavisione extra-acquatica. Nervo ottico sottile e cilin-drico con sepimentazioni interfascicolari; chiasmacon decussazione totale di larghi fasci interdigita-ti. Le fibre sono solo 900 in derivazione di circa90.000 fotorecettori retinici (Palmer, 1918). Rettili = In questi animali la retina appare mero-angiotica (vascolarizzata solo in una parte) a dif-ferenza della olo-angiotica in cui la vascolarizza-zione è uniformemente distribuita. Il nervo otti-co, esile e sottile, possiede fibre che non mostra-no un andamento ordinato e parallelo, infram-mezzate da cellule di oligodendroglia, senzasepimenti interfascicolari apparenti. Al chiasma la decussazione è totale. In alcuniserpenti (Natrix) rare fibre raggiungono diretta-mente il corpo genicolato.
Uccelli = All’osservazione funduscopica balzaevidente una formazione caratteristica di questianimali: il “pecten” (pettine), formazione chedalla retina aggetta nel vitreo portandosi daldisco ottico verso l’equatore. È costituita da unfitto arabesco capillare intralicciato da tessuto disostegno. La sua discussa funzione sembra lega-ta alla nutrizione della retina attraverso la suavastissima superficie di scambio ossigenativo etermico, che tra l’altro consente di tenere co-stante la temperatura anche quando l’animalevola ad alta quota. Sembra inoltre che serva daspecchio riflettente sulla retina di immagini chesovrastano, come i predatori che in genereattaccano dall’alto (Thompson, 1928). La spazia-tura regolare dei denti del pettine, determine-rebbe la proiezione sulla retina di una immaginestroboscopica degli oggetti in movimento nellospazio, rendendoli così percepibili più rapida-mente e dettagliatamente (Menner, 1928). Daultimo consentirebbe una perfetta analisi deipunti geografici “di compasso” per l’orientamen-to di navigazione (Wilkinson, 1949). Il nervo ottico mostra sepimentazioni settali difattura e dimensioni variabili. Spesso un unicosepimento divide in due contingenti i fasci di
44 PARTE CLINICA
Figura 24Via ottica: decussazione comparata

fibre ottiche, cellule di oligodendroglia appaionoirregolarmente sparse negli spazi interfibrillari.La decussazione al chiasma è totale con interdi-gitazione complicata dei fasci. Mammiferi = Le loro caratteristiche biologichefondamentali possono essere così riassunte: dia-framma che divide il celoma in due cavità, cuoretetra-camerale, orecchio tri-camerulare, mandi-bola semplice, tegumenti ricoperti da peli, nutri-zione attraverso le ghiandole mammarie dellafemmina. La relativa semplicità delle strutturerecettive oculari è compensata dalla particolaredifferenziazione ed articolazione delle vie otti-che. Nei monotremi il nervo ottico ospita 32.000assoni (Bruesh e Arey, 1942). Nei placentati(che includono la maggior parte dei mammiferi)la struttura del nervo non si scosta da quella del-l’uomo, se non per le proporzioni. Gli assonisono raggruppati in piccoli fasci separati da settiinterfibrillari, la cui componente gliale cresce colprogredire nella scala biologica. Anche a questoproposito è interessante seguire, passo passo, ilcammino evolutivo. Dapprima (ciclostomi) ilsistema inter-fibrillare è costituito da una colon-na di cellule ependimali, che decorrono al cen-tro del nervo e da cui irradiano tralicci seconda-ri che si portano alla superficie. In seguito, neipesci e nei rettili, questo sistema centrale si rad-doppia e la sepimentazione diviene più spiccatae complicata. Nei vertebrati più evoluti, la fasci-colazione diventa sempre meno manifesta, sinoad arrivare nell’uomo a costituire una architettu-ra di sostegno tenue ed uniformemente distri-buita. Per quanto riguarda la testa del nervo otti-co, va ricordato che la lamina cribrosa esiste solonei mammiferi a prevalente visione diurna. Nonsi rinviene infatti negli animali che vivono inhabitat a bassa luminanza (certi roditori) in cuila retina trova varco per erniare negli spazimeningei perineurali.Relativamente al chiasma, si può in generaleaffermare che il numero delle fibre diretteaumenta con il grado di frontalizzazione degliocchi. Negli animali con occhi lateralizzati que-ste sono relativamente rare in rapporto tra fibredirette e decussate è di 1/6 nel cavallo, 1/4 nelcane e nel gatto, 1/3 nei primati più evoluti e di1/2 nell’uomo. La percentuale di fibre decussate
è in stretto rapporto con la organizzazione strut-turale del corpo genicolato a cui pervengono.Nei mammiferi inferiori le fibre ottiche si con-nettono con le cellule di un corpo genicolatoarchitettonicamemte elementare, che non haancora realizzato il processo di laminazione. Inmammiferi più evoluti si contano 4 strati delCGL, che raggiungono il numero di 6 nei prima-ti. Nei tetra-stratificati le fibre crociate perven-gono al l° e 3° strato e le dirette al 2° e 4°. Negliesa-stratificati. le prime approdano al 1°, 4° e 6°;le seconde al 2°, 3°, 5°. Nei primati ogni camporecettivo si proietta a livello del CGL “point topoint” (o “aire par aire”). Ne consegue che,mentre i settori sensoriali di ogni singola emire-tina riverberano lo stimolo su tre strati cellularigenicolati, a livello della corteccia vengonoproiettati gli stimoli di tutti i sei strati del corpogenicolato stesso. Vi sono animali dove manca un vero e proprionervo ottico, essendo gli occhi letteralmenteinseriti nella massa celebrale o appena salientisulla sua superficie (mesostomum). In alcuni ilnervo è sottilissimo e lungo (gasteropodi), inaltri grosso e tozzo (crostacei), in altri ancora inervi ottici possono essere multipli ed afferisco-no a gangli diversi che, essendo però molto vici-ni, costituiscono un complesso funzionale deno-minato lobo ottico cerebrale (aracnidi). Negliinvertebrati i centri nervosi appaiano sempreintimamente collegati con gli organi visuosenso-riali, sia che questi siano incastonati nei ganglisuperiori, sia che afferiscano ad un ganglio pro-prio che aderisce alla massa cerebrale o che neè separato da un peduncolo ottico. Nei vertebra-ti non esiste il ganglio ottico e gli assoni incana-lati nel nervo ottico si portano dalla retina aicentri visivi superiori. La forma, le dimensioni e l’architettura di questosegmento nervoso sono quanto mai variabili nelpanorama dell’anatomia comparata. La sua por-zione orbitaria, cortissima negli uccelli e lunganell’uomo circa 3 cm. è nell’elefante oltre 11 cm.Il suo diametro, di 3 mm. nell’uomo, è di 5, 5 nelcavallo, 6 nel bue, 2, 8 nel maiale, 1, 1 nel gattoe 50 nella balena. In alcuni rettili (tartarughe) èpercorso da una lunga incisione che gli conferi-sce una forma di doccia. Negli uccelli la pia
45Bio-anatomia delle vie ottiche

meninge si affonda nel suo contesto dal latonasale sino a raggiungere l’asse. Nel tritone hauna struttura particolarmente grossolana e pri-mitiva, è costituito da un canale tappezzato daelementi neurogliali, che sfocia nel 3° ventricolo.Nel proteus è tanto sottile e frammisto ad altritronchi nervosi da renderne difficile l’identifica-zione. J.Deyl, già nel 1895, stabilì che le singolefibre si raggruppano in fascetti assonali tanto piùidentificabili quanto più si sale la scala biologica. Nei ciclostomi l’incrocio chiasmatico avvienedentro la massa cerebrale. Nei pesci teleostei si realizza semplicemente colpassaggio di un nervo sopra all’altro. General-mente quello che proviene dall’occhio destroscavalca quello sinistro. Nella trota avviene inve-ce l’inverso. Nella figura 24 sono illustrati le piùcomuni modalità di decussazione. Nella maggiorparte dei rettili, uccelli ed anfibi i nervi otticisono costituiti da un diverso numero di fasci chesi interdigitano con quello dell’altro nervo percui, in una sezione trasversa, si configura unacaratteristica linea a zig-zag. I sepimenti interfa-scicolari sono in questi casi molto pronunciati edevidenti. Nei mammiferi i fasci sono molto più piccoli enumerosi ed i sepimenti sono molto più tenui. Lefibre ottiche nel chiasma non decorrono tuttesullo stesso piano, ma le superficiali si approfon-dano, le profonde si superficializzano e molte sisuddividono e si biforcano. Il parziale incrociodelle fibre nel chiasma del cavallo fu dimostratonel 1886 da G. Colin che osservò, oltre all’atrofiadel nervo ottico di un occhio cieco, la concomi-
tante atrofia della bandelletta del lato opposto.Nel ratto e nel coniglio l’incrocio avviene solo inparte a livello del tuber cinereum. Nel 1909,BossaIino stabilì che l’incrocio non avviene maiper singole fibre, ma per fasci di assoni. Nell’uomo il nervo ottico penetra nel bulbo nellasua parte posteriore, un po’ nasalmente. L’ec-centricità dell’impatto si fa invece temporalenella pecora, nel bue e nella foca. Nella balena è,per contro, superiore. La lamina cribrosa, robusta nei ruminanti, èrudimentale nel coniglio e nei carnivori.Nell’elefante manca totalmente e la coroide siinsinua negli spazi intervaginali della meninge. In alcuni uccelli, il nervo ottico entra nella scle-ra, decorre sotto il pettine e perfora la coroide,formando cosi una lamina cribrosa sui generis.In molte specie di pesci e nei cefalopodi, il nervosi suddivide in numerosi rami che penetranonella sclera attraverso molti fori, costituendopertanto papille ottiche multiple. La papilla rotondeggiante nell’uomo, si ovalizzatrasversalmente nei ruminanti; obliqua nel maia-le, di forma bizzarra nel cervo, a mezzaluna nellapecora, reniforme nella renna, a striscia orizzon-tale nella marmotta, a triangolo nel cane, imbu-tiforme nel coniglio dove si espande nelle carat-teristiche ali bianche. A caldaia nel coccodrillo,è sporgente in alcuni pesci (squatina). In altripesci la retina è separata nettamente dai bordidella papilla da un anello scuro. La papilla verti-cale della rana, secondo C. Nicolai, può variare isuoi diametri grazie ad un cercine muscolare cheè situato nella coroide circumpapillare.
46 PARTE CLINICA