
ANALISI DELLA CAPACITÀ DENITRIFICANTE DI AMBIENTI … · 5 le aree umide perifluviali, stagnanti e...
33
Convenzione “FILI D’ACQUA” tra il Dipartimento di Scienze Ambientali dell’Università degli Studi di Parma (DSA) e il Parco dell’Oglio Sud ANALISI DELLA CAPACITÀ DENITRIFICANTE DI AMBIENTI UMIDI PERIFLUVIALI Erica Racchetti, Elisa Soana, Daniele Longhi, Monica Pinardi, Rossano Bolpagni, Marco Bartoli, Pierluigi Viaroli Prof. Pierluigi Viaroli [email protected] tel. 0521 905683, fax 0521 905402 Novembre 2007
Transcript of ANALISI DELLA CAPACITÀ DENITRIFICANTE DI AMBIENTI … · 5 le aree umide perifluviali, stagnanti e...

Convenzione “FILI D’ACQUA” tra il Dipartimento di Scienze Ambientali
dell’Università degli Studi di Parma (DSA) e il Parco dell’Oglio Sud
ANALISI DELLA CAPACITÀ DENITRIFICANTEDI AMBIENTI UMIDI PERIFLUVIALI
Erica Racchetti, Elisa Soana, Daniele Longhi, Monica Pinardi, Rossano Bolpagni, Marco Bartoli, Pierluigi Viaroli
Prof. Pierluigi Viaroli [email protected]. 0521 905683, fax 0521 905402
Novembre 2007

2
INDICE 1. Introduzione.....................................................................................................3 2. Aree di studio ...................................................................................................7 3. Materiali e metodi .............................................................................................9
3.1 Determinazione delle caratteristiche chimico-fisiche dei sedimenti superficiali ..9 3.2 Quantificazione del metabolismo bentico e dei tassi di denitrificazione .......... 10 3.3 Calcoli utilizzati per la stima dei tassi di denitrificazione................................ 13 3.4 Determinazioni analitiche sulla colonna d’acqua........................................... 14
4. Risultati.......................................................................................................... 15 4.1 Caratterizzazione delle acque di fondo e dei sedimenti superficiali ................ 15 4.2. Tassi di respirazione dei sedimenti superficiali ............................................ 18 4.3. Flussi netti di azoto inorganico all’interfaccia acqua-sedimento .................... 22 4.4. Tassi di denitrificazione invernali ed estivi .................................................. 23
5. Discussione e conclusioni ................................................................................ 26 6. Riferimenti bibliografici.................................................................................... 29 APPENDICE ........................................................................................................ 31 localizzazione delle aree di studio ........................................................................ 31
3
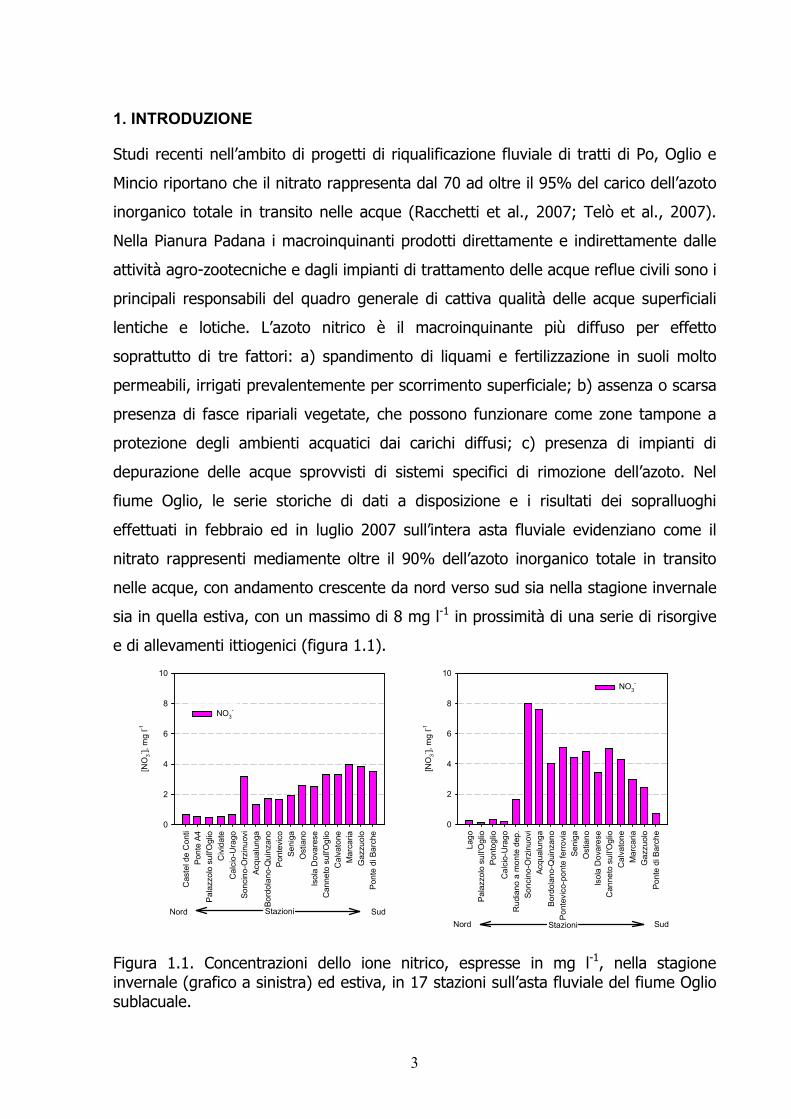
1. INTRODUZIONE Studi recenti nell’ambito di progetti di riqualificazione fluviale di tratti di Po, Oglio e Mincio riportano che il nitrato rappresenta dal 70 ad oltre il 95% del carico dell’azoto inorganico totale in transito nelle acque (Racchetti et al., 2007; Telò et al., 2007). Nella Pianura Padana i macroinquinanti prodotti direttamente e indirettamente dalle attività agro-zootecniche e dagli impianti di trattamento delle acque reflue civili sono i principali responsabili del quadro generale di cattiva qualità delle acque superficiali lentiche e lotiche. L’azoto nitrico è il macroinquinante più diffuso per effetto soprattutto di tre fattori: a) spandimento di liquami e fertilizzazione in suoli molto permeabili, irrigati prevalentemente per scorrimento superficiale; b) assenza o scarsa presenza di fasce ripariali vegetate, che possono funzionare come zone tampone a protezione degli ambienti acquatici dai carichi diffusi; c) presenza di impianti di depurazione delle acque sprovvisti di sistemi specifici di rimozione dell’azoto. Nel fiume Oglio, le serie storiche di dati a disposizione e i risultati dei sopralluoghi effettuati in febbraio ed in luglio 2007 sull’intera asta fluviale evidenziano come il nitrato rappresenti mediamente oltre il 90% dell’azoto inorganico totale in transito nelle acque, con andamento crescente da nord verso sud sia nella stagione invernale sia in quella estiva, con un massimo di 8 mg l-1 in prossimità di una serie di risorgive e di allevamenti ittiogenici (figura 1.1).
Figura 1.1. Concentrazioni dello ione nitrico, espresse in mg l-1, nella stagione invernale (grafico a sinistra) ed estiva, in 17 stazioni sull’asta fluviale del fiume Oglio sublacuale.
Stazioni
Caste
ldeC
onti
Ponte
A4Pa
lazzo
losu
ll'Ogli
oCi
vidate
Calci
o-Urag
oSo
ncino
-Orzi
nuov
iAc
qualu
nga
Bordo
lano-Q
uinza
noPo
ntevic
oSe
niga
Ostia
noIso
laDo
vares
eCa
nneto
sull'O
glio
Calva
tone
Marca
riaGa
zzuo
loPo
ntedi
Barch
e
[NO 3
- ],mgl
-1
0
2
4
6
8
10
NO3-
Nord SudStazioni
Lago
Palaz
zolo
sull'O
glio
Ponto
glio
Calci
o-Ur
ago
Rudia
noam
onte
dep.
Sonc
ino-O
rzinu
ovi
Acqu
alung
aBo
rdola
no-Q
uinza
noPo
ntevic
o-po
ntefer
rovia
Senig
aOs
tiano
Isola
Dova
rese
Cann
etosu
ll'Ogli
oCa
lvaton
eMa
rcaria
Gazz
uolo
Ponte
diBa
rche
[NO 3
- ],mgl
-1
0
2
4
6
8
10NO3
-
Nord Sud

4
L’azoto nitrico nei campioni di acqua prelevati in prossimità delle sorgive raggiunge concentrazioni oltre 12 mg l-1 e conferma la probabile origine diffusa di questo ione. I carichi di nitrato veicolati in Po variano dalle 3 alle 24 tonnellate al giorno, rispettivamente per il periodo estivo ed invernale 2007. L’analisi di oltre 50 immissari del fiume Oglio, sia in sponda destra che in sponda sinistra, rivela inoltre che le concentrazioni di azoto nitrico sono maggiori o uguali a quelle del corpo idrico recettore ad indicare la generale compromissione o vulnerabilità da nitrati dell’intero bacino. L’unico processo in natura in grado di rimuovere il nitrato dal comparto acquoso e sedimentario e trasferirlo in atmosfera come N2 gassoso è la denitrificazione, un processo mediato da batteri specializzati che avviene esclusivamente in condizioni di anaerobiosi (figura 1.2).
Figura 1.2. E’ illustrato il ciclo dell’azoto. Il processo di denitrificazione converte l’azoto nitroso e nitrico in azoto molecolare o protossido d’azoto, che vengono rilasciati in atmosfera. Negli ecosistemi acquatici, ambienti elettivi per le comunità di batteri denitrificanti sono le zone riparie a basso idrodinamismo e accumulo di sostanza organica labile e
Sedim
ento
Sedim
ento
anos
sico
anos
sico
Sedim
ento
ossic
oSe
dimen
toos
sico
Acqu
aAc
qua
ATM.
ATM. Ciclo dellCiclo dell’’azotoazoto
N organico DON NH4+
Argille/acidi umiciseppellimento
ammonificazioneNO2- NO3-
N2/N2ODENITRIFICAZIONE
N organico DON NH4+ NO2- NO3-
N organico DON NH4+ NO2- NO3-
NN22/N/N22OONN22

5
le aree umide perifluviali, stagnanti e generalmente caratterizzate da ipossia o anossia delle acque (Seitzinger, 1988; Leonardson, 1994). In sedimenti di aree umide perifluviali la disponibilità di substrati e le condizioni riducenti stimolano generalmente il metabolismo anaerobico che spiega una larga frazione della sostanza organica degradata (Seitzinger, 1988; Brusch et al., 1991). Ulteriormente, la presenza di macrofite in grado di trasferire ossigeno ai sedimenti mediante diffusione attraverso il parenchima aerifero, stimola processi aerobici quali la nitrificazione in micronicchie presenti nei sedimenti profondi che forniscono “dall’interno” una sorgente di nitrato ai batteri denitrificanti. In questo modo viene rimosso sia azoto nitrico dalla colonna d’acqua che azoto ammoniacale previa ossidazione (Reddy et al., 1989; Bodelier et al., 1996). I tassi di denitrificazione sono generalmente correlati positivamente al carico organico sedimentario, alle concentrazioni di azoto nitrico nella colonna d’acqua e alle temperature. Il carico organico e in particolare il carbonio organico disciolto stimola i processi respiratori aerobi ed anaerobi. La scarsa solubilità dell’ossigeno e la limitata disponibilità in acqua di questa molecola fa si che i tassi respiratori aerobi possano determinarne l’esaurimento a livello dei sedimenti. Minore è lo spessore dell’orizzonte ossico nel sedimento e minore è il percorso che il nitrato presente in acqua deve effettuare per raggiungere lo strato anossico dove avviene la denitrificazione. Concentrazioni elevate di azoto nitrico in acqua determinano inoltre in forte gradiente di concentrazione tra l’acqua ed i sedimenti (che sono generalmente privi di nitrato) ed una conseguente grande spinta diffusiva. La riduzione del nitrato è quindi direttamente proporzionale alla richiesta di accettori di elettroni e alla disponibilità di NO3-. Le piante acquatiche e le microalghe bentoniche possono competere con le comunità di batteri nitrificanti e denitrificanti in ambienti caratterizzati da basse concentrazioni di azoto inorganico e limitarne l’azione (Bartoli et al., 2003; Risgaard-Petersen, 2003). Lungo il corso dei fiumi Oglio e Po si alternano sequenze di ambienti umidi relitti parzialmente o completamente scollegati al corso d’acqua principale, testimonianza dell’evoluzione degli ambiti fluviali verso condizioni di pensilità. Si tratta di lanche, bodri, aree di esondazione e casse di espansione caratterizzate da età e quindi stadi di interramento e trofia differente. Alcuni di questi ambienti hanno le caratteristiche

6
degli ambienti distrofici e sono quindi perennemente anossici a causa dell’eccessivo carico organico, altri al contrario hanno substrati più minerali a seguito di pratiche gestionali in atto o di fasi successionali differenti. Le diverse fasi successionali e la connessione idraulica al fiume o alle falde superficiali selezionano inoltre nei diversi ambienti umidi comunità macrofitiche differenti che includono vegetazione sommersa, emergente o pleustofitica. Non esistono in bibliografia studi relativi alla misura dei tassi di denitrificazione di aree umide in Pianura Padana e tanto meno ricerche mirate a chiarire la regolazione del processo. In questa relazione vengono presentati i risultati di una serie di misure dei tassi di denitrificazione in 20 ambienti umidi relitti situati in ambiti pianeggianti compresi tra i bacini dell’Oglio e del Po, nelle province di Mantova e Cremona. In una relazione precedente, al termine del primo anno del progetto Fili d’Acqua, gli stessi ambienti umidi sono stati inquadrati dal punto di vista idrochimico e vegetazionale. Scopi di questo lavoro sono: a) valutare in modo comparativo i tassi di rimozione dell’azoto nitrico; b) caratterizzare i fattori che regolano il processo; c) valutare la denitrificazione potenziale dei vari ambienti mediante arricchimento del nitrato nella colonna d’acqua; d) discutere i risultati ottenuti in relazione a possibili interventi di connessione idraulica tra ambiente fluviale e zone umide perifluviali al fine di controllare i carichi diffusi di azoto nitrico.

7
2. AREE DI STUDIO La maggior parte degli ambienti indagati sono aree umide relitte caratterizzate da elevata eutrofia, avanzato stato di interramento e isolamento parziale o totale dal fiume. Un quadro sintetico degli ambienti studiati è riportato nella tabella 2.1, dove sono distinti per tipologia di ambiente umido (torbiere, bodri, canali o lanche), estensione, profondità, alimentazione (acque di falda, acque di scarico o piene eccezionali del fiume), isolamento o collegamento costante al fiume, caratteristiche dei sedimenti e popolamenti di produttori primari. La localizzazione approssimativa dei siti di indagine è riportata in appendice. Tabella 2.1. Caratteristiche delle zone umide perifluviali campionate. Codice
sito
Località Coordinate (Gauss-Boaga)
Tipologia
S(ha)
h(m)
Alimentazione Sedimento Vegetazione
OG1 Gabbioneta 1595982 5008085
lanca 8.07 0.5 falda, depuratore
limoso organico
Phragmites australis, alghe
filamentose OG2 Foce
Gambara 1602192 5004875
Rientranza foce
0.41 0.5 canale, fiume sabbioso argilloso
Microalghe bentoniche
OG3 Gerre Gavazzi
1605969 5002880
lanca 3.85 1.02 falda, fiume limoso organico
Microalghe
OG4 Runate ds 1606494 5002809
lanca 0.18 0.07 falda, fiume, collegamento con lanca sx
sabbioso limoso
Nuphar luteum
OG5 Runate sx 1606636 5002813
lanca 0.29 0.08 fiume, falda, apporti
puntiformi da abitazioni
limoso Nuphar luteum
0G6 Bicocca 1605383 5002214
lanca 0.29 0.03 falda limoso Phragmites australis, Nuphar luteum, Lemna minor, Spirodela
polyrrhiza

8
OG7 Castelfranco 2
1606066 5001731
lanca 0.37 0.02 falda limoso Phragmites australis, Nuphar
luteum OG8 Castelfranco
11606564 5001663
lanca 0.60 0.02 falda limoso Phragmites australis, Lemna minor, Salvinia
natans OG9 Lago Pesca 1610524
5003055 stagno 0.66 1.20 falda fangoso
organico Phragmites
australis, Nuphar luteum,
Ceratophyllum demersum
OG10 Bine 1612876 4999079
lanca 12.08
0.06 falda, fiume limoso organico
Phragmites australis
OG11 Mosio 1617484 4999276
torbiera-lanca
0.23 0.04 falda limoso organico
Phragmites australis
OG12 Marcaria 1621140 4997260
torbiera 1.09 0.03 falda limoso organico
Phragmites australis, Carex
riparia OG13 Belforte 1621023
4992786 torbiera 0.39 0.02 falda limoso
organico Microalghe bentoniche, Carex riparia
OG14 Bogina 1625592 4986548
canale 6.62 0.05 fiume limoso organico
Phragmites australis
PO1 Gerre de Caprioli
1582474 4992209
lanca 0.15 0.04 falda, fiume fangoso limoso
Assente, microfitobenthos
PO2 Lancone 1582437 4986916
lanca 6.19 0.05 falda, fiume limoso organico
Trapa natans
PO3 Lazzaretto 1584958 4991424
bodrio 1.25 0.1 falda, fiume, canale
limoso organico
Spirodela polyrrhiza,
Wolffia arrhiza, Lemna gibba
PO4 Gerre Ugolani
1584997 4989539
bodrio 0.39 0.1 falda, fiume limoso Myriophyllum sp. Ceratophyllum d. Potamogeton cr.

9
PO5 Cà de Gatti 1589282 4991353
bodrio 0.28 0.06 falda, fiume, canale
organico Lemna gibba, Spirodela polyrrhiza,
Wolffia arrhiza PO6 Cascina
Margherita 1590295 4991274
bodrio 0.2 0.1 falda, fiume limoso argilloso
Nuphar luteum, Ceratophyllum
demersum
3. MATERIALI E METODI Sono state effettuate due campagne di campionamento di acqua e sedimento, una nel periodo invernale tra febbraio e marzo 2007 e una nel periodo estivo tra giungo e agosto 2007; sono state campionate da 1 a 3 stazioni per uscita. Durante ogni sopralluogo sono stati raccolti campioni di acqua di fondo per misure di temperatura, ossigeno disciolto e azoto nitrico e campioni di sedimento. Sedimenti indisturbati sono stati prelevati manualmente mediante tubi in Plexiglas trasparente; in ogni stazione sono state raccolte 4 carote (i.d. 8 cm, lunghezza 30 cm) per la misura del metabolismo bentico e 3 carote (i.d. 4 cm, lunghezza 30 cm) per la caratterizzazione del substrato. Una volta campionate, le carote sono state trasferite in vasche contenenti acqua del sito alla stessa temperatura misurata nel punto di campionamento. Il rimescolamento della colonna d’acqua è stato assicurato attraverso una pompa da acquario posizionata all’interno delle vasche di mantenimento. 3.1 Determinazione delle caratteristiche chimico-fisiche dei sedimenti superficiali In laboratorio, il giorno successivo al prelievo, le carote sono state estruse ed il primo centimetro è stato sezionato ed omogeneizzato. Dallo strato selezionato sono state prelevate aliquote di sedimento per la misura della porosità, contenuto d’acqua, sostanza organica e clorofilla. La porosità del sedimento e la percentuale di sostanza secca sono state misurate come perdita in peso dopo essiccazione a 105°C per 24 ore di 1 ml di sedimento. La porosità viene espressa come ml di acqua per 1 ml di sedimento. La densità come g di sedimento umido per ml di sedimento.

10
La sostanza organica è stata misurata come perdita di peso di circa 0.2 – 0.3 g di sedimento secco per incenerimento a 350 °C per 2 ore. Il contenuto di clorofilla viene misurato tramite estrazione in acetone al 90% (24 ore al buio) e lettura spettrofotometrica a 4 lunghezze d’onda (Lorentzen, 1967). 3.2 Quantificazione del metabolismo bentico e dei tassi di denitrificazione Le incubazioni sono state effettuate il giorno successivo alla raccolta dei sedimenti mantenendo le carote al buio, secondo la metodica proposta da Dalsgaard et al (2000). Al momento dell’avvio delle incubazioni, il livello dell’acqua delle vasche è stato abbassato al di sotto del bordo superiore delle carote; l’apertura superiore delle stesse è stata quindi chiusa con un tappo galleggiante in plexiglas. Durante l’incubazione una barretta magnetica sospesa a circa 5 cm dall’interfaccia acqua-sedimento è stata fatta ruotare ad una velocità di 60 rpm da un sistema di magneti guidati da un motore esterno per evitare la stratificazione dell’acqua presente all’interno delle carote (figura 3.1). Come tempo iniziale (To) è stato considerato il momento della chiusura delle carote. Appena prima della chiusura sono stati prelevati 60 ml di acqua per le determinazioni analitiche. Al termine dell’incubazione è stato effettuato da ogni carota un secondo prelievo per la determinazione della concentrazione finale. La durata dell’incubazione è stata programmata in modo tale da ottenere una variazione della concentrazione dell’ossigeno significativa, ma non superiore al 20% della concentrazione iniziale (2-3 ore).
magnete Vasca di incubazione
Figura 3.1. Set up sperimentale per la misura di flussi bentonici e tassi di denitrificazione in carote intatte (per maggiori dettagli vedere il testo).

11
L’acqua prelevata è stata in parte fissata per le misure di ossigeno e del carbonio inorganico (TCO2 = H2CO3+HCO3-+CO32-) e in parte filtrata per le misure dei nutrienti azotati. Le analisi per le misure delle concentrazioni di ossigeno e TCO2 sono state eseguite subito dopo il prelievo, mentre i campioni per le misure di ammonio (NH4+), nitriti (NO2-) e nitrati (NO3-), sono stati congelati ed analizzati successivamente. I flussi sono stati misurati come variazione nel tempo della concentrazione del composto considerato e calcolati secondo la seguente formula:
( )AtVCiCfF *
*−=
Dove: F è il flusso del composto in esame (µmol m-2 h-1); Ci è la concentrazione (µmol/l) del composto in esame al tempo iniziale; Cf è la concentrazione (µmol/l) del composto in esame al tempo finale; V è il volume dell’acqua all’interno della carota (l); A è l’area della superficie del sedimento nella carota (m2). t è il tempo di incubazione (h) La quantificazione dei tassi di denitrificazione è stata effettuata utilizzando la tecnica dell’accoppiamento isotopico proposta da Nielsen (1992) (figura 3.2).
Figura 3.2. Misura dei tassi di denitrificazione mediante impiego di nitrato marcato. L’assunzione principale del metodo è la perfetta miscela dei due isotopi nella fase acquosa e la loro diffusione nell’orizzonte anossico dove vengono ridotti in base ai rispettivi rapporti stechiometrici (per maggiori dettagli vedere il testo).
SedimentoSedimentoAnossicoAnossico
2828NN222929NN223030NN22
1515NONO33--1414NONO33--
AcquaAcqua
SedimentoSedimentoOssicoOssico

12
Il metodo adottato consente di quantificare i tassi totali di denitrificazione e di distinguere tra quelli dovuti al nitrato che diffonde nei sedimenti dalla colonna d’acqua e quelli dovuti ai processi accoppiati di nitrificazione-denitrificazione. Il metodo si basa sull’aggiunta di nitrato marcato (15NO3-) all’interno di ogni singola carota, in modo da avere un arricchimento di nitrato marcato di almeno il 30% del pool di nitrato non marcato (14NO3-) presente nel sito in esame. Nelle 4 carote sono state aggiunte in realtà quantità crescenti di azoto marcato al fine di stimare l’attività potenziale di denitrificazione in relazione alla disponibilità di substrato. L’assunzione principale è che il nitrato che diffonde dalla colonna d’acqua (15NO3- + 14NO3-) equello che si origina nel sedimento per nitrificazione (14NO3-) si distribuiscano omogeneamente nella zona in cui avviene il processo. In queste condizioni appena al di sotto dell’interfaccia ossico/anossica, i batteri denitrificanti riducono la miscela isotopica di NO3- a 28N2 + 29N2 + 30N2.Le incubazioni sono state condotte successivamente a quelle per la misura dei flussi utilizzando le stesse carote e lo stesso set up sperimentale, tra i due esperimenti le carote sono state aperte e nuovamente sommerse in acqua del sito di campionamento per circa due ore. L’esperimento vero e proprio è iniziato con l’aggiunta di 15NO3- nella colonna d’acqua delle carote. Successivamente, dopo aver atteso un tempo di 10-15 minuti per permettere al 15NO3- di raggiungere la zona anossica e di equilibrarsi con il nitrato presente nell’acqua interstiziale, le carote sono state chiuse per iniziare l’incubazione. Il tempo di incubazione massimo è stato calibrato in modo che il consumo di ossigeno non fosse superiore al 20% del contenuto iniziale (tipicamente, 3-4 ore). Al termine dell’incubazione sono stati prelevati campioni per l’analisi del 28N2 + 29N2 + 30N2. Le carote sono state avvelenate con circa 2 ml di una soluzione di ZnCl2 7M e il sedimento è stato mescolato per ottenere una miscela omogenea. Un campione della miscela è stato quindi prelevato per le analisi dell’azoto molecolare prodotto per denitrificazione e immediatamente trasferito in provette Exetainer a tenuta (Labco) a cui sono stati aggiunti 100 µl di ZnCl2 7M. I campioni di N2 per l’analisi isotopica sono stati analizzati presso il National Environmental Research Institute di Silkeborg (DK) con un gascromatografo in linea con uno spettrofotometro di massa.

13
3.3 Calcoli utilizzati per la stima dei tassi di denitrificazione I tassi di produzione di 14N15N, 15N15N sono stati calcolati tramite l’equazione generale per il calcolo dei flussi
AtVVCCNP aqsedif
x1)()(15 ×
+×−=
φ
dove: P15Nx = tasso di produzione dell’isotopo di interesse (14N15N o 15N15N µmol m-2 h-1)A = area della superficie di sedimento della carota Ci = concentrazione iniziale di 15Nx (µM) Cf = concentrazione finale di 15Nx (µM) t = tempo di incubazione Vsed = volume di sedimento nella carota Vaq = volume di acqua nella carota Φ = porosità del sedimento La produzione di gas 28N2 (D14) rappresenta la denitrificazione in situ ed è stata calcolata con le seguenti formule in accordo con quanto descritto da Nielsen (1992):
D p N N p N N15 214 15 15 15= +( ) ( )
15)(2)(14 1515
1514DNNp
NNpD ×=
dove: D15 =tasso di denitrificazione di 15NO3- (µmol m-2 h-1)D14 = tasso di denitrificazione di 14NO3- (µmol m-2 h-1). Il tasso di denitrificazione del nitrato proveniente dalla colonna d’acqua (Dw), conoscendo la percentuale di 15NO3- aggiunto, è stato quantificato tramite la seguente formula:
1515%100
315 DDNONatomDw −
×= −
dove: 15Natom% NO3- = percentuale di 15NO3- nella colonna d’acqua Il tasso di denitrificazione accoppiata alla denitrificazione è stato di conseguenza stimato sottraendo Dw al tasso di denitrificazione in situ:

14
Dn=D14-Dw I tassi di denitrificazione misurati mediante isotopi sono stati confrontati con i tassi teorici ottenuti mediante l’equazione proposta da Christensen et al. (1990):
−
××+××=− 18.0
182.018.02
32
O
NOOW C
CFD
Dove FO2 è la domanda sedimentaria di ossigeno misurata al buio, CNO3- e CO2 sono le concentrazioni di nitrato e ossigeno nell’acqua di fondo adiacente ai sedimenti. 3.4 Determinazioni analitiche sulla colonna d’acqua Temperatura: misura diretta nell’ambiente mediante sonda multiparametrica (YSI 556 MPS); L’ossigeno disciolto è stato determinato per titolazione secondo il metodo Winkler (A.P.H.A., 1981). Il carbonio inorganico (H2CO3+HCO3-+CO32-) è stato determinato tramite titolazione con HCl 0.1N a sei punti finali di pH (Anderson et al., 1986). L’ammonio (NH4+) è stato determinato spettrofotometricamente con il metodo del blu indofenolo secondo Koroleff (1970). Il nitrito (NO2-) è stato determinato spettrofotometricamente secondo il metodo proposto da A.P.H.A., (1981). Il nitrato (NO3-) è stato determinato spettrofotometricamente dopo riduzione a nitrito con cadmio attivato con rame per diazotazione (A.P.H.A., 1981).

15
4. RISULTATI 4.1 Caratterizzazione delle acque di fondo e dei sedimenti superficiali I prelievi per la caratterizzazione delle acque di fondo sono avvenuti durante il giorno e le concentrazioni di ossigeno riscontrate sono sicuramente influenzate dall’attività delle alghe in colonna d’acqua e del microfitobentos. La tabella 4.1 riporta le concentrazioni di ossigeno disciolto e di nitrato per la stagione invernale ed estiva per i 20 siti d’indagine. In inverno l’ossigeno disciolto è risultato prossimo alla saturazione (300<x<400 µM) nella maggior parte dei siti mentre valori lievemente inferiori sono stati misurati nei siti OG3, OG8 e OG10 (~160, ~216, ~180 µM rispettivamente), con un sito decisamente sottosaturo OG11, ~20 µM. Nel campionamento estivo i valori di ossigeno diminuiscono in accordo alla minore solubilità alle maggiori temperature dell’acqua in 12 siti su 15, mentre aumentano in OG5, OG11, OG12 probabilmente a causa dell’attività fotosintetica. Nel periodo estivo circa la metà dei siti indagati ha tenori di ossigeno prossimi alla saturazione (~270 µM). Le concentrazioni dell’azoto nitrico sono risultate estremamente eterogenee tra siti. In OG1 e PO2 il tenore dei nitrati è risultato constante nelle due stagioni (~360 e ~2 µM, rispettivamente) probabilmente a causa dell’apporto costante di acque ricche di questo ione dal depuratore, per il primo, e dalla mancanza di apporti dal fiume Po, per il secondo. In OG3, OG4 e OG5 i valori invernali (820, 810 e 1150 µM rispettivamente) si sono ridotti sensibilmente durante il periodo estivo (380, 670 e 600 µM rispettivamente), così come per i siti in cui la concentrazione invernale è molto più bassa, tra 3 e 30 µM (OG10, OG13, PO1, PO3, PO4 e PO6), in estate si sono rilevati valori di nitrato minori di 1 µM. Si sono riscontrati anche aumenti delle concentrazioni di nitrato dall’inverno all’estate nei siti OG2, OG11 e in modo particolare nei siti OG12 e PO5 dove le concentrazioni invernali (10 e 3 µM, rispettivamente) sono aumentate di un fattore 10 e 7 durante l’estate. Per i siti OG6, OG7 e OG8, non è stato possibile effettuare il campionamento estivo per la mancanza di acqua nelle lanche. I siti OG9 e OG14 sono stati inseriti solo nel campionamento estivo. Gli ambienti studiati sono caratterizzati da sedimenti superficiali soffici, con densità nello strato tra 0 e 1 cm simili a quella dell’acqua, indici di ambienti ad elevato apporto particellato (tabella 4.2). Eccezioni sono rappresentate dal sito OG12, PO3 e PO5 e dal sito OG4. I primi,

16
estremamente riducenti, hanno densità lievemente inferiori a 1 g cm-1 per la presenza di bolle di gas (probabilmente metano) negli interstizi dei sedimenti. Il secondo, alimentato da acque di risorgiva, ha al contrario sedimenti compatti, sabbiosi, con densità superiori a 1.5 g cm-3. Il contenuto elevato di acqua dell’orizzonte superficiale è confermato dai valori di porosità, mediamente superiori a 0.8 con l’eccezione del sito OG4 per i motivi detti precedentemente. Tabella 4.1. Concentrazioni dell’azoto nitrico e dell’ossigeno disciolto nelle acque adiacenti ai sedimenti degli ambienti umidi studiati. 71.4 µM di nitrato corrispondono a 1 mg l-1; 31.2 µM di ossigeno corrispondono a 1 mg l-1.
sito NO3- (µM) O2 (µM) inverno estate inverno estate
OG1 354,6 362,7 327,3 118,7 OG2 364,5 478,8 408,1 146,8 OG3 1133,0 681,0 160,9 86,8 OG4 819,0 369,3 364,9 316,8 OG5 821,0 689,1 300,2 602,9 OG6 20,6 n.d. 743,7 n.d. OG7 <1 n.d. 569,2 n.d. OG8 8,3 n.d. 216,2 n.d. OG9 n.d. 960,3 n.d. 252,8 OG10 3,5 1,0 181,3 192,2 OG11 5,0 8,4 21,0 782,5 OG12 9,6 110,5 353,1 606,3 OG13 13,3 <1 313,1 298,0 OG14 n.d. 9,4 n.d. 545,1 PO1 30,8 <1 401,7 245,3 PO2 2,1 2,4 608,6 506,6 PO3 3,1 0,4 500,6 70,9 PO4 12,3 0,6 482,00 290,6 PO5 3,3 21,1 485,4 63,4 PO6 4, 0 0,2 352,6 328,8
17
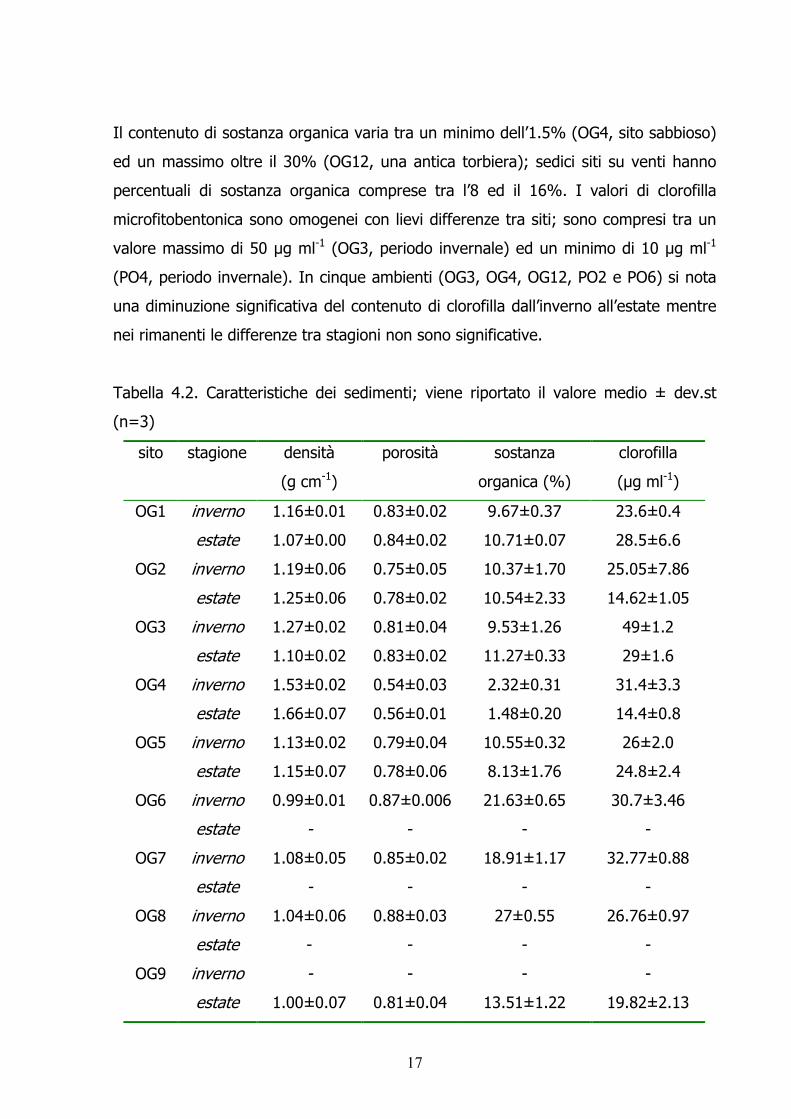
Il contenuto di sostanza organica varia tra un minimo dell’1.5% (OG4, sito sabbioso) ed un massimo oltre il 30% (OG12, una antica torbiera); sedici siti su venti hanno percentuali di sostanza organica comprese tra l’8 ed il 16%. I valori di clorofilla microfitobentonica sono omogenei con lievi differenze tra siti; sono compresi tra un valore massimo di 50 µg ml-1 (OG3, periodo invernale) ed un minimo di 10 µg ml-1
(PO4, periodo invernale). In cinque ambienti (OG3, OG4, OG12, PO2 e PO6) si nota una diminuzione significativa del contenuto di clorofilla dall’inverno all’estate mentre nei rimanenti le differenze tra stagioni non sono significative. Tabella 4.2. Caratteristiche dei sedimenti; viene riportato il valore medio ± dev.st (n=3)
sito stagione densità (g cm-1)
porosità sostanza organica (%)
clorofilla (µg ml-1)
inverno 1.16±0.01 0.83±0.02 9.67±0.37 23.6±0.4 OG1 estate 1.07±0.00 0.84±0.02 10.71±0.07 28.5±6.6 inverno 1.19±0.06 0.75±0.05 10.37±1.70 25.05±7.86 OG2 estate 1.25±0.06 0.78±0.02 10.54±2.33 14.62±1.05 inverno 1.27±0.02 0.81±0.04 9.53±1.26 49±1.2 OG3 estate 1.10±0.02 0.83±0.02 11.27±0.33 29±1.6 inverno 1.53±0.02 0.54±0.03 2.32±0.31 31.4±3.3 OG4 estate 1.66±0.07 0.56±0.01 1.48±0.20 14.4±0.8 inverno 1.13±0.02 0.79±0.04 10.55±0.32 26±2.0 OG5 estate 1.15±0.07 0.78±0.06 8.13±1.76 24.8±2.4 inverno 0.99±0.01 0.87±0.006 21.63±0.65 30.7±3.46 OG6 estate - - - -inverno 1.08±0.05 0.85±0.02 18.91±1.17 32.77±0.88 OG7 estate - - - -inverno 1.04±0.06 0.88±0.03 27±0.55 26.76±0.97 OG8 estate - - - -inverno - - - -OG9 estate 1.00±0.07 0.81±0.04 13.51±1.22 19.82±2.13

18
inverno 1.10±0.02 0.91±0.06 16.10±0.69 23.6±3.4 OG10estate 1.03±0.01 0.85±0.02 16.32±0.47 25.8±0.7 inverno 1.07±0.02 0.81±0.01 8.90±0.71 27.11±1.55 OG11estate 1.23±0.01 0.76±0.03 5.77±0.42 35.99±6.65 inverno 0.94±0.04 0.85±0.06 33.58±0.72 34.8±4 OG12estate 0.92±0.02 0.85±0.04 33.18±0.43 22±2 inverno 1.13±0.002 0.91±0.008 15.73±0.02 18.68±4.27 OG13estate 0.97±0.03 0.42±0.02 19.20±0.23 13.1±0.93 inverno - - - -OG14estate 0.99±0.002 0.84±0.006 22.72±1.83 23.98±1.98 inverno 1.16±0.02 0.81±0.01 7.31±0.81 24.14±1.88 PO1 estate 1.03±0.01 0.77±0.007 8.61±0.38 33.16±0.7 inverno 1.01±0.003 0.89±0.008 12.14±0.34 33.25±7.03 PO2 estate 0.96±0.02 0.83±0.01 12.13±0.26 18.66±0.48 inverno 0.93±0.003 0.89±0.01 34.35±1.26 19.50±3.62 PO3 estate 0.88±0.08 0.83±0.08 23.4±0.47 13.97±1.24 inverno 1.44±0.28 0.66±0.13 5.20±4.65 10.27±4.13 PO4 estate 0.99±0.03 1.52±0.03 8.81±0.22 18.27±1.51 inverno 0.96±0.05 0.86±0.05 27.26±1.26 17.05±3.38 PO5 estate 0.90±0.08 1.41±0.08 27.51±2.32 19.90±1.39 inverno 1±0.04 0.86±0.03 15.10±1.35 37.22±9.68 PO6 estate 1.17±0.26 0.85±0.22 13.57±3.90 24.21±4.17
4.2. Tassi di respirazione dei sedimenti superficiali Il consumo bentico di ossigeno misurato nei due periodi mediante incubazione di carote intatte è riportato in figura 4.1. La respirazione sedimentaria dipende dal carico organico, dalle temperature dell’acqua e dalla presenza di organismi bioturbatori. I risultati indicano in generale tassi di respirazione decisamente elevati (valore medio 3.3 mmol m-2h-1) in accordo con le caratteristiche distrofiche della maggior parte dei siti analizzati e tendenzialmente correlati al carico organico nei sedimenti e alle temperature dell’acqua. Nei siti in cui i tassi respiratori bentonici

19
sono risultati maggiori in inverno rispetto all’estate è ipotizzabile la scomparsa del macrobenthos. Osservazioni dei sedimenti hanno infatti permesso di notare dense comunità di chironomidi solo nei campioni prelevati a febbraio-marzo; l’anossia notturna e le condizioni riducenti del periodo estivo escludono probabilmente questi organismi dai sedimenti. Il forte rischio di anossia di questi ambienti si può desumere dal rapporto tra disponibilità di ossigeno in colonna d’acqua e respirazione bentonica. Nel periodo estivo, considerando un metro quadrato di superficie e una profondità di mezzo metro, la colonna d’acqua contiene circa 3 g di ossigeno disciolto mentre la respirazione dei sedimenti sottrae circa 0.2-0.3 g di ossigeno all’ora. In queste condizioni, in assenza di vento, in dieci ore di buio la riserva di ossigeno presente in acqua può essere completamente esaurita. E’ quindi probabile che all’alba la maggior parte degli ambienti studiati sia senza ossigeno. Questo non avviene nel periodo invernale quando la riserva in acqua è doppia, le temperature e le attività metaboliche sono minori e la frequenza di giornate ventose è maggiore. Il collegamento con ambienti fluviali e quindi la minore stagnazione eviterebbe l’instaurarsi dell’anossia a favore delle comunità animali e dei processi aerobici.
Figura 4.1. Consumo bentonico di ossigeno da parte dei sedimenti superficiali nel periodo invernale ed estivo del 2007.
OG1
OG2
OG3
OG4
OG5
OG6
OG7
OG8
OG9
OG10
OG11
OG12
OG13
OG14 PO
1PO
2PO
3PO
4PO
5PO
6
mmol
O 2m-2
h-1
-8
-6
-4
-2
0
2
4 inverno estate

20
I flussi netti di carbonio inorganico misurati nei due periodi sono riportati in figura 5; la produzione di anidride carbonica è, analogamente al consumo di ossigeno, una misura indiretta dei tassi respiratori degli organismi bentonici (batteri, protozoi, meio e macrofauna, produttori primari). Considerando che anche diversi processi microbici anaerobici sono in grado di ossidare completamente il carbonio organico ad anidride carbonica i flussi riportati in figura 4.2 sono la risultante dei processi respiratori aerobici ed anaerobici.
Figura 4.2. Flussi netti di anidride carbonica all’interfaccia acqua-sedimento misurati nel periodo invernale ed estivo del 2007 in ambienti umidi perifluviali di Oglio e Po. Analogamente a quanto discusso precedentemente i tassi di produzione di CO2 sono estremamente elevati, generalmente maggiori in valore assoluto a quelli dell’ossigeno ad indicare una quota importante dei processi anaerobici nel metabolismo bentonico complessivo dei sistemi analizzati. In 6 siti l’efflusso di CO2 è superiore a 10 mmol m-
2h-1 e in 4 siti addirittura superiore a 15 mmol m-2h-1 mentre il massimo consumo di ossigeno non supera 7 mmol m-2h-1. Tendenzialmente i flussi di anidride carbonica sono correlati con il carico organico e con le temperature dell’acqua. Il rapporto in valore assoluto tra i flussi di anidride carbonica ed i flussi di ossigeno, definito come quoziente respiratorio (RQ), è utilizzato come indice per valutare
OG1
OG2
OG3
OG4
OG5
OG6
OG7
OG8
OG9
OG10
OG11
OG12
OG13
OG14 PO
1PO
2PO
3PO
4PO
5PO
6
mmol
CO2
m-2h-1
-10
0
10
20
30
40inverno estate

21
l’importanza relativa dei processi aerobici rispetto a quelli anaerobici nel metabolismo del sistema bentonico. Generalmente un quoziente respiratorio unitario è tipico di sedimenti ossidati, senza accumulo di prodotti finali del metabolismo anaerobico; al contrario sedimenti con RQ>1 indicano condizioni riducenti con assenza di riossidazione e tendenza alla distrofia. Combinando i flussi riportati nelle figure 4.1 e 4.2 sono stati calcolati i quozienti respiratori invernali ed estivi per i siti di studio (figura 4.3).
Figura 4.3. Quoziente respiratorio (RQ = flusso dell’anidride carbonica/flusso dell’ossigeno) calcolato nei periodi invernale ed estivo nei siti di indagine. Come lecito aspettarsi nella grande maggioranza dei casi il quoziente respiratorio è risultato maggiore di 1, sia in inverno che in estate, ad indicare l’importanza della componente anaerobica nel metabolismo generale del sistema.
OG1
OG2
OG3
OG4
OG5
OG6
OG7
OG8
OG9
OG10
OG11
OG12
OG13
OG14 PO
1PO
2PO
3PO
4PO
5PO
6
RQ
0
5
10
15
20 inverno estate
22
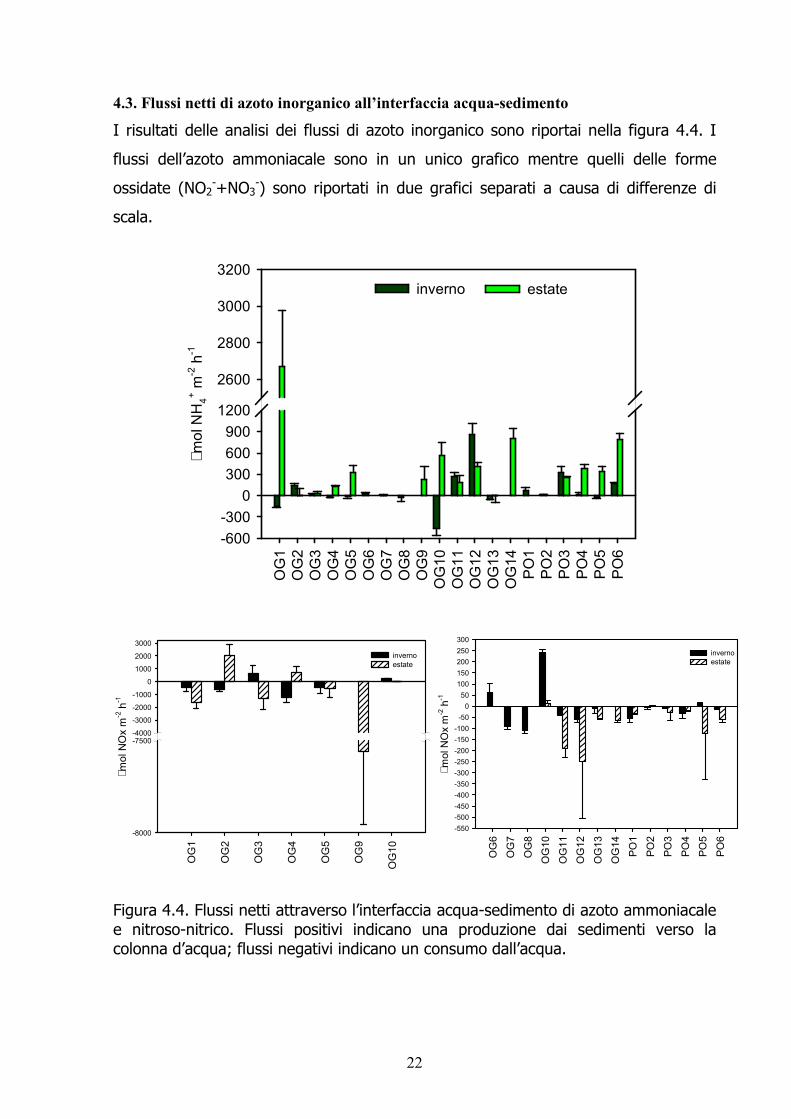
4.3. Flussi netti di azoto inorganico all’interfaccia acqua-sedimento I risultati delle analisi dei flussi di azoto inorganico sono riportai nella figura 4.4. I flussi dell’azoto ammoniacale sono in un unico grafico mentre quelli delle forme ossidate (NO2-+NO3-) sono riportati in due grafici separati a causa di differenze di scala.
Figura 4.4. Flussi netti attraverso l’interfaccia acqua-sedimento di azoto ammoniacale e nitroso-nitrico. Flussi positivi indicano una produzione dai sedimenti verso la colonna d’acqua; flussi negativi indicano un consumo dall’acqua.
OG1
OG2
OG3
OG4
OG5
OG9
OG10
µ mol
NOxm
-2h-1
-8000
-7500-4000-3000-2000-1000
0100020003000
invernoestate
OG6
OG7
OG8
OG10
OG11
OG12
OG13
OG14 PO1
PO2
PO3
PO4
PO5
PO6
µ mol
NOxm
-2h-1
-550-500-450-400-350-300-250-200-150-100
-500
50100150200250300
invernoestate
OG1
OG2
OG3
OG4
OG5
OG6
OG7
OG8
OG9
OG10
OG11
OG12
OG13
OG14 PO
1PO
2PO
3PO
4PO
5PO
6
µ mol
NH4+
m-2h-1
-600-300
0300600900
12002600
2800
3000
3200inverno estate

23
A conferma dei risultati relativi alle respirazioni, nella maggior parte dei siti si ha una rigenerazione netta di azoto ammoniacale, probabilmente a seguito del processo di ammonificazione dell’azoto organico labile presente nell’orizzonte più superficiale del sedimento. Solo in un numero estremamente basso di casi ed esclusivamente nel periodo invernale, sono stati misurati flussi negativi. In questi casi è probabile si tratti di assimilazione al buio da parte del microfitobenthos o di nitrificazione. I tassi di rigenerazione dell’ammonio sono estremamente rilevanti (in 10 siti gli efflussi sono maggiori di 300 µmol NH4+ m-2h-1) a conferma del carattere distrofico degli ambienti analizzati (=caratterizzati da un carico organico eccessivo e quindi da tassi elevati di sedimentazione di particellato labile). L’azoto ammoniacale rigenerato al buio è prontamente incorporato dai produttori primari nelle ore di luce e non si accumula nella colonna d’acqua. La grande disponibilità di questo ione spiega la presenza in numerosi ambienti di pleustofite, macrofite con foglie flottanti alla superficie dell’acqua. Questa tipologia di produttori primari si stabilisce in ambienti ricchi di nutrienti e tende a formare comunità monospecifiche che occupano l’intero specchio d’acqua escludendo essenze più pregiate e promuovendo ulteriormente condizioni di ipossia/anossia. La scarsa disponibilità di ossigeno nei sedimenti superficiali, nonostante la grande disponibilità di ammonio, si traduce in un limitato processo di nitrificazione. I flussi delle forme ossidate dell’azoto inorganico sono infatti prevalentemente negativi (figura 4.4) con tassi legati alle concentrazioni dell’azoto nitroso e nitrico in acqua. Il consumo del nitrito e del nitrato da parte dei sedimenti avviene probabilmente a seguito di processi dissimulativi quali la nitrato ammonificazione o la denitrificazione. 4.4. Tassi di denitrificazione invernali ed estivi Sette ambienti (OG1, OG2, OG3, OG4, OG5, OG9 e OG12) hanno concentrazioni di azoto nitrico in acqua decisamente superiori a quelle delle altre aree umide. Se per i siti OG2, OG3, OG4 e OG5 tali valori sono imputabili al fatto che sono idraulicamente collegati al fiume Oglio e in prossimità di risorgive, per OG1 il valore costante di circa 5 mg l-1 si pensa sia originato da un possibile scarico di acque reflue all’interno della lanca e dall’influenza dell’attività agrozootecnica. Anche il sito OG9, circondato da campi coltivati, viene influenzato dall’attività agrozootecnica e dalla presenza di
24
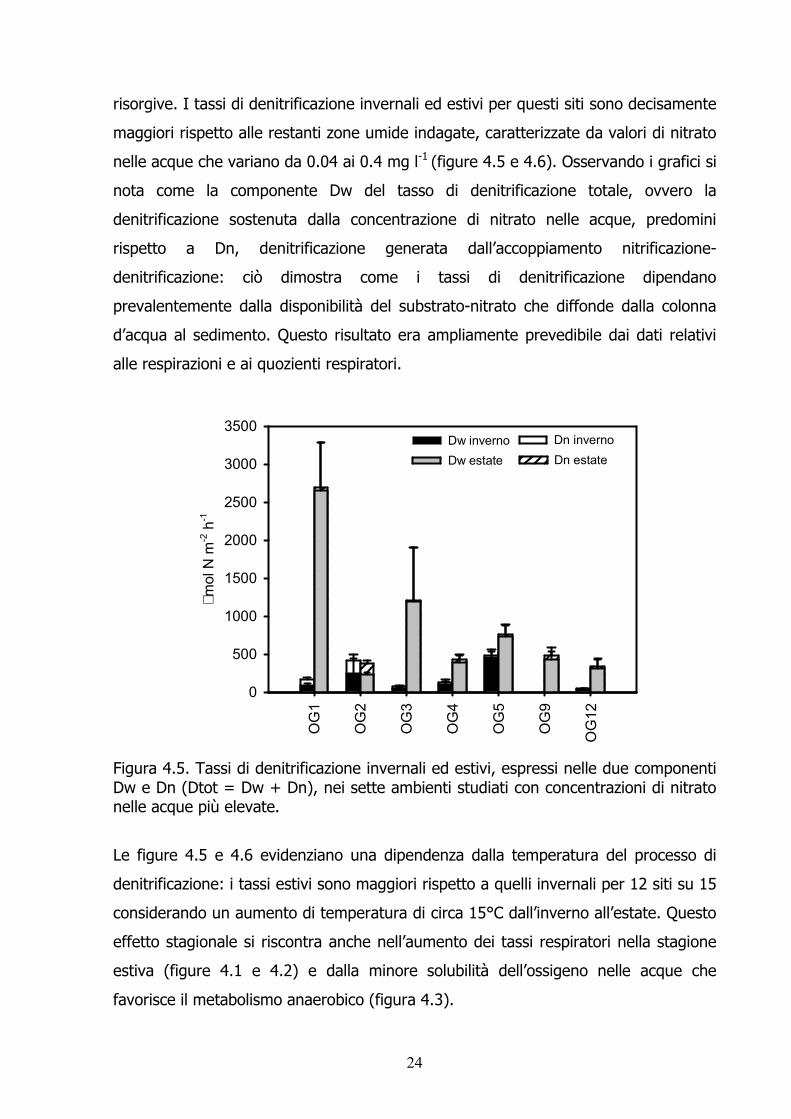
risorgive. I tassi di denitrificazione invernali ed estivi per questi siti sono decisamente maggiori rispetto alle restanti zone umide indagate, caratterizzate da valori di nitrato nelle acque che variano da 0.04 ai 0.4 mg l-1 (figure 4.5 e 4.6). Osservando i grafici si nota come la componente Dw del tasso di denitrificazione totale, ovvero la denitrificazione sostenuta dalla concentrazione di nitrato nelle acque, predomini rispetto a Dn, denitrificazione generata dall’accoppiamento nitrificazione-denitrificazione: ciò dimostra come i tassi di denitrificazione dipendano prevalentemente dalla disponibilità del substrato-nitrato che diffonde dalla colonna d’acqua al sedimento. Questo risultato era ampliamente prevedibile dai dati relativi alle respirazioni e ai quozienti respiratori.
Figura 4.5. Tassi di denitrificazione invernali ed estivi, espressi nelle due componenti Dw e Dn (Dtot = Dw + Dn), nei sette ambienti studiati con concentrazioni di nitrato nelle acque più elevate. Le figure 4.5 e 4.6 evidenziano una dipendenza dalla temperatura del processo di denitrificazione: i tassi estivi sono maggiori rispetto a quelli invernali per 12 siti su 15 considerando un aumento di temperatura di circa 15°C dall’inverno all’estate. Questo effetto stagionale si riscontra anche nell’aumento dei tassi respiratori nella stagione estiva (figure 4.1 e 4.2) e dalla minore solubilità dell’ossigeno nelle acque che favorisce il metabolismo anaerobico (figura 4.3).
OG1
OG2
OG3
OG4
OG5
OG9
OG12
µmol
Nm-2
h-1
0
500
1000
1500
2000
2500
3000
3500Dn invernoDn estate
Dw invernoDw estate

25
Figura 4.6. Tassi di denitrificazione invernali ed estivi, espressi nelle due componenti Dw e Dn (Dtot = Dw + Dn), nei 13 ambienti studiati con concentrazioni di nitrato nelle acque più basse. L’aggiunta di quantità crescenti di azoto nitrico ai sedimenti risulta in una stimolazione del processo con effetti differenti e sito-specifici. In altre parole, l’apporto puntiforme o diffuso di carichi azotati determina un aumento immediato dei tassi di denitrificazione ed una risposta positiva del sistema (meccanismo tampone). Gli ambienti che sono pensili rispetto al fiume o alle falde o scollegati ai corpi d’acqua principali hanno tenori di nitrati in acqua estremamente contenuti e tali da sostenere tassi di denitrificazione irrilevanti. Sono inoltre ambienti con sedimenti riducenti e nitrificazione scarsa: difficilmente il carico azotato interno al sistema sarà alleggerito dal processo accoppiato Dn di nitrificazione e denitrificazione. Più probabile un accoppiamento conservativo tra processi di ammonificazione, assimilazione, sedimentazione del materiale particellato e di nuovo ammonificazione. L’applicazione del modello di Christensen ai dati disponibili per gli ambienti umidi studiati conferma la predittibilità dei tassi di denitrificazione sulla base della domanda sedimentaria di ossigeno e del rapporto tra concentrazioni di nitrato e ossigeno nelle acque di fondo. I tassi teorici e quelli misurati mostrano un discreto accordo per gli ambienti con basse concentrazioni di nitrato in acqua (figura 4.7) mentre sono
OG6
OG7
OG8
OG10
OG11
OG13
OG14 PO
1PO
2PO
3PO
4PO
5PO
6
µ mol
Nm-2
h-1
0
50
100
150
200
250Dn invernoDn estate
Dw invernoDw estate

26
discrepanti i risultati ottenuti con i dati degli ambienti ad elevate concentrazioni di azoto nitrico. Questa discrepanza può essere dovuta ad un problema metodologico: negli ambienti dove il nitrati diffonde dal basso verso l’alto (falde superficiali ricche di NO3-) infatti le assunzioni del metodo isotopico non sono valide.
Figura 4.7. Confronto tra i tassi di denitrificazione misurati Dw ed i tassi di denitrificazione teorici previsti dal modello di Christensen sulla base della domanda sedimentaria di ossigeno e del rapporto tra nitrati e ossigeno in acqua.
5. DISCUSSIONE E CONCLUSIONI La grande maggioranza degli ambienti studiati è caratterizzata da condizioni di stato trofico estremamente elevato, con valori di sostanza organica fino a oltre il 30% e densità dei sedimenti paragonabili a quella dell’acqua. In questi ambienti i tassi di sedimentazione del particellato fine (microalghe, frammenti di macrofite, fiocchi di batteri) sono probabilmente maggiori dei tassi di decomposizione con accumulo di materiale indecomposto e grande domanda di accettori di elettroni per i processi di mineralizzazione (ossigeno ma anche nitrato). L’interramento rapido di queste zone umide, in particolare di quelle isolate come i bodri e le lanche pensili (OG3, OG6, OG7, OG8, OG10, OG13, PO1, PO2, PO3, PO4, PO5 e PO6) è conseguente agli apporti di nutrienti di origine alloctona e all’export limitato di sostanza organica.
[NO3-]/[O2]
0 1 2 3 4 5 6
Dw(m
molm
-2 h-1 )
0,0
0,5
1,0
1,5
2,0Winter period
0.51.01.5
Theoretical DwMeasured Dw
FlussodiO2
(mmolm-2h -1)
Dw teorico Dw misurato
[NO3-]/[O2]
0 1 2 3 4 5 6
Dw(m
molm
-2 h-1 )
0,0
0,5
1,0
1,5
2,0Winter period
0.51.01.5
Theoretical DwMeasured Dw
FlussodiO2
(mmolm-2h -1)
Dw teorico Dw misurato

27
I flussi di ossigeno e anidride carbonica misurati al buio sono tipici di ambienti a forte rischio anossico, dove quindi la riserva di ossigeno in acqua non è sufficiente a compensare la grande domanda sedimentaria. L’efflusso di anidride carbonica è stechiometricamente maggiore del consumo di ossigeno a conferma che gran parte dell’ossidazione del materiale organico presente nei sedimenti avviene per vie anaerobiche senza riossidazione dei prodotti finali e accumulo di potere riducente. Il metabolismo batterico rigenera nelle acque grandi quantità di azoto ammoniacale e consente il protrarsi di condizioni eutrofiche-distrofiche. In bibliografia sono riportati esempi di ambienti che permangono eutrofici per anni anche a seguito di interventi per il controllo degli inquinanti puntiformi e diffusi a causa dell’enorme carico interno. Il processo di denitrificazione, strettamente anaerobico, è stimolato in condizioni di grande domanda di accettori di elettroni (e quindi di elevato carico organico) ma deve essere presente anche nitrato in acqua poiché in condizioni riducenti il processo di nitrificazione non avviene. Se l’input di nitrato è occasionale (eventi piovosi intensi o esondazioni) in questi ambienti il processo di denitrificazione è immediatamente stimolato fino all’esaurimento del nitrato. Questa ipotesi è confermata dai nostri risultati che mostrano tassi di produzione di N2 crescenti al variare del substrato anche negli ambienti a minore concentrazione di NO3-. Quando però il nitrato è esaurito la funzione dell’area umida quale “reattore” naturale per la rimozione del nitrato rimane inespressa sino all’input successivo e la forma dominante dell’azoto inorganico è quella ammoniacale. Appare importante, nell’ottica di salvaguardare questi ambienti, intervenire attivamente nella gestione degli stessi controllando le sorgenti inquinanti, soprattutto gli apporti organici e particellati e alleggerendo il carico interno. Per considerazioni che saranno riprese in seguito inerenti i processi di denitrificazione, appare inoltre importante ove possibile ricollegare idraulicamente questi sistemi ai corpi d’acqua principali per favorire l’export dei nutrienti in eccesso e il rinnovo delle acque (e con esse l’apporto di ossigeno). I sistemi che non sono collegati al fiume stanno evolvendo rapidamente verso sistemi terrestri ed hanno una funzionalità come zone umide estremamente scarsa. Sono infatti grandi produttori di gas sera come anidride carbonica e metano (Bolpagni et al., 2007), sono probabilmente ipossici-anossici nelle ore notturne, selezionano

28
comunità poco pregiate di macrofite come comunità monospecifiche di pleustofite (OG6, PO3, PO5) e svolgono una limitata azione denitrificante poichè la forma dominante di azoto nelle acque è lo ione ammonio e non lo ione nitrato. Maggiormente funzionali appaiono invece gli ambienti costantemente o temporaneamente collegati al fiume (OG2, OG3, OG4, OG5); ricevono infatti apporti di molecole ossidate, esportano cataboliti e sostanza organica, permettono l’ingresso e l’uscita di organismi per motivi legati alla riproduzione, al rifugio o all’alimentazione e svolgono un’importante azione di rimozione del carico azotato. Il ripristino della funzionalità ed il mantenimento delle aree umide perifluviali è un target importante nell’ottica del controllo dei carichi azotati: i tassi di rimozione dell’azoto determinati in un ambiente umidi tipo (OG5) ammontano a oltre 900 kg ha-
1y-1 e sono quasi 4 volte superiori alla quantità di azoto fornito per concimare un ettaro di mais, la coltura più esigente in termini di azoto nei terreni (240 kg ha-1 y-1).

29
6. RIFERIMENTI BIBLIOGRAFICI A.P.H.A., A.W.W.A., W.P.C.F., 1981. Standard methods for the examination of water
and wastewater. Am. Publ. Health Ass., Washington. pp: 1-1134. Anderson, L.G., Hall, P.O.J., Iverfeldt, A., van der Loeff, M.M.R., Sundby, B.,
Westerlund, S.F.G., 1986. Benthic respiration measured by total carbonate production. Limnol. Oceanogr. 31: 319-329.
Bartoli, M., Nizzoli, D., Viaroli, P., 2003. Microphytobenthos activity and fluxes at the sediment-water interface: interactions and spatial variability. Aquatic Ecology. 37: 341-349.
Bodelier, P.L.E., Libochant, J.A., Blom, C.W.P.M., Laanbroek, H.J., 1996. Dynamics of nitrification and denitrification in root oxygenated sediments and adaptation of ammonia-oxidizing bacteria to low oxygen or anoxic habitats. Appl Environ Microbiol. 62: 4100–4107.
Brusch, W., Nilsson, B.,1991. Nitrate transformation and water movement in a wetland area. In Nitrogen and phosphorus in fresh and marine waters. Project abstracts of the Danish NPo research programme. Miljøstyrelsen, Copenhagen.
Christensen, PB, Nielsen, LP, Sorensen, J, Revsbech, NP, 1990. Denitrification in nitrate-rich streams: diurnal and seasonal variations related to benthic oxygen metabolism. Limnol Oceanogr. 35: 640-651.
Dalsgaard, T., Nielsen, L.P., Brotas, V., Viaroli, P., Underwood, G.J.C., Nedwell, D.B., Sundbäck, K., Rysgaard, S., Miles, A., Bartoli, M., Dong, L., Thornton, D.C.O., Ottosen, L.D.M., Castaldelli, G. & Risgaard-Petersen, N. 2000. Protocol handbook for NICE-Nitrogen cycling in estuaries: a project under the EU research programme. Marine Science and Technology (MAST III). National Environmental Research Institute, Silkeborg, Denmark, pp: 62.
Koroleff, F., 1970 - Direct determination of ammonia in natural waters as indophenol blue. Information on techniques and methods for seawater analysis. I.C.E.S. Interlaboratory Rep. No.3, pp:19-22.
Leonardson, L., 1994. Wetlands as nitrogen sinks: Swedish Environmental Protection Agency Report no. 4176. (In Swedish with English summary).
Lorenzen, C.J., 1967. Determination of chlorophyll and phaeo-pigments: spectrophotometric equations. Limnol. Oceanogr. 12:343-346.
Nielsen, L.P., 1992. Denitrification in sediment determined from nitrogen isotope pairing. FEMS (Fed. Eu. Microbiol. Soc.) Microbiol. Ecol. 86: 357-362.
Racchetti, E., Appiani, U., Soana, E., Longhi, D., Pinardi, M., Bolpagni, R., Baroli, M., Viaroli, P., Giugno 2007. “Relazione intermedia delle attività” – Convenzione tra il Dipartimento di Scienze Ambientali dell’Università degli Studi di Parma (DSA) e il Parco dell’Oglio Sud per la valutazione comparata dei carichi inquinanti diffusi e puntiformi generati nel bacino del fiume Oglio (tratto sublacuale) finalizzata ad individuare linee d’azione per la riduzione del carico inquinante.
Reddy, K.R., Patrick, W.H. Jr, Lindau, C.W., 1989. Nitrification-denitrification at the plant root–sediment interface in wetlands. Limnol Oceangr. 34: 1004–1013.
Seitzinger, S.P., 1988. Denitrification in freshwater and coastal marine ecosystems: Ecological and geochemical significance. Limnol. Oceanogr. 33(4, part 2): 702-724.

30
Regione Lombardia, Marzo 2006. “Programma di Tutela e Uso delle Acque” – Relazione di sintesi. pp: 1- 26.
Telò, R., Pinardi, M., Bodini, A., Bartoli, M., Racchetti, E., Viaroli, P., Cuizzi, D., Vannucci, M., Previdi, L., Marzo 2006 – Maggio 2007. “Caratterizzazione dello stato ambientale del fiume Mincio e analisi della strategia di riqualificazione integrata e partecipata”- Progetto del Mincio “da Agenda 21 ad Azione 21 per il Mincio”.

31
APPENDICE
LOCALIZZAZIONE DELLE AREE DI STUDIO

32

33








![[PPT]Ma il clima sta davvero cambiando? Conferenza a … · Web viewIl vero pericolo: Anossia ed Estinzioni di massa Conferenza a cura di Orleo Marinaro “...i dinosauri scomparvero](https://static.fdocumenti.com/doc/165x107/5c6b30f909d3f287198b7588/pptma-il-clima-sta-davvero-cambiando-conferenza-a-web-viewil-vero-pericolo.jpg)









