
Amplificazione non lineare e risonanza stocastica: come l...
23
Amplificazione non lineare e risonanza stocastica: come l'orecchio sente suoni deboli Seminari degli studendi di Fisica 30 maggio 2008 Ludovico Silvestri
Transcript of Amplificazione non lineare e risonanza stocastica: come l...

Amplificazione non linearee risonanza stocastica:come l'orecchio sente
suoni deboli
Seminari degli studendi di Fisica30 maggio 2008
Ludovico Silvestri

Qualche domanda...
Cosa vuol dire “suono piccolo”?
Come fa l'orecchio a trasdurre il segnale da acustico a elettrico?
Cosa c'entra la Fisica con tutto questo?

Cellule ciliate
Le cellule ciliate sono presenti in numerosi organi sensoriali, dalla coclea al labirinto ai
sensori cutanei.
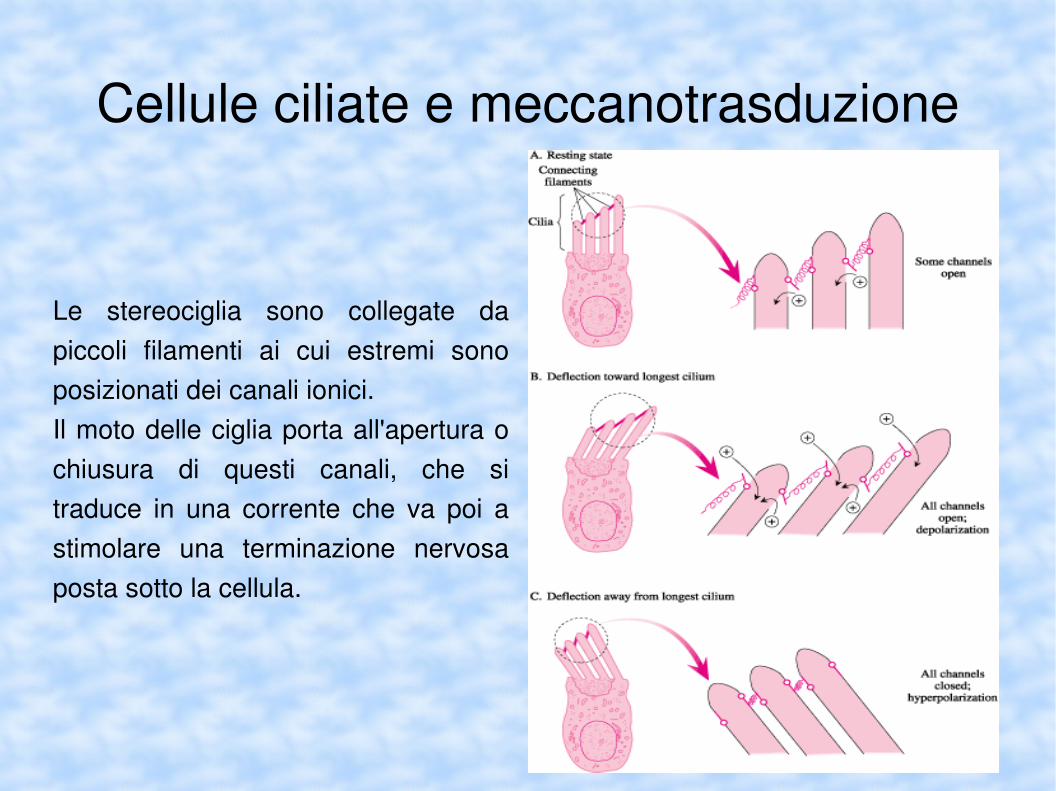
Cellule ciliate e meccanotrasduzione
Le stereociglia sono collegate da piccoli filamenti ai cui estremi sono posizionati dei canali ionici.Il moto delle ciglia porta all'apertura o chiusura di questi canali, che si traduce in una corrente che va poi a stimolare una terminazione nervosa posta sotto la cellula.

Qualche numeroMinima deflessione rilevabile = 0.3 nm (0.003°)
Deflessioni dovute al moto Browniano di equilibrio termico = 23 nm
Minima pressione di picco rilevabile come suono = 20 μPa
dalla legge di Stevino
si ha
ΔP = 20 μPa Δh = 2 μm
dPdh
=− g

Primo modello:l'oscillatore di Hopf forzato + rumore
Una biforcazione di Hopf è un cambiamento qualitativo della dinamica del sistema a seguito di un cambiamento quantitativo del valore di un certo parametro che trasforma un punto di equilibrio
stabile in un ciclo limite.

Un esempio di biforcazione di Hopf: il circuito di Van der Pol
d2x
dt2x2
−1dxdt
02xb=0

Oscillatore di Hopf forzato: sincronizzazione
Un sistema lineare forzato (una molla) presenta il noto fenomeno della risonanza.
Nel caso di un sistema non lineare questa scompare per lasciare il posto alla sincronizzazione.
Ci sono aspetti radicalmente diversi: ● la sincronizzazione è “repentina”, cioè esiste un'ampiezza critica della forzante;● la sincronizzazione “blocca” la fase relativa anche in presenza di rumore;● la sincronizzazione ha un rapporto ampiezza della forzante/ampiezza dell'oscillazione che (in generale) non è costante per ogni ampiezza.

Dinamica vicino ad una biforcazione di Hopf
Ogni sistema dinamico nei pressi di una biforcazione è equivalente (a meno di un diffeormorfismo) ad un sistema di equazioni caratteristico della biforcazione: la forma normale. Nel caso della biforcazione di Hopf l'equazione (una sola) è:
Vicini alla frequenza caratteristica si ha la sincronizzazione dell'oscillatore, quindi la risposta può essere sviluppata in serie di Fourier:
Scomponendo la forma normale del sistema nei vari modi si può dimostrare* che in caso di sincronizzazione 1:1 l'equazione per il modo dominante diventa:
f t = ∑n=−∞
∞
f n ei n t x t =∑n=−∞
∞
xn ei n t
f 1≃A ,C x1B ,C ∣x1∣2 x1
* J. Balakrishnan, J.Phys.A 38, 1627 (2005)S. Camalet, T. Duke, F. Julicher, J. Prost, PNAS 97, 3183 (2000)
z t =D 0 , C z−E 0 , C ∣z∣2 zO ∣z∣4 z f t

Dinamica vicino ad una biforcazione di HopfPer C = Cc , e ω vicina ad ω0 il termine lineare può essere sviluppato in serie:
Sostituendo nell'eq. Precedente si osservano due regimi.Per si ha:
Per si ha:
Quindi l'amplificazione è non lineare: per segnali piccoli va con una legge a potenza e per segnali piu' forti è lineare. Per ottenere questo effetto è necessario che il sistema rimanga nei pressi di Cc . Questo può essere garantito mediante un meccanismo di feedback. Una possibilità è data dalla seguente legge di evoluzione per Cc (in questo caso le oscillazioni si hanno al di sotto della soglia):
∣−0∣≪∣ f 1∣2/3∣B∣1/3/A1
∣−0∣≫∣ f 1∣2/3∣B∣1/3/A1
A ,C c≃A1−0
∣x1∣≃∣B∣−1/3
∣ f 1∣1/3
∣x1∣≃∣ f 1∣
∣−0A1∣
1C
dCdt
=1 x2
2−1

Oscillazioni di Hopf nelle cellule ciliateUn oscillatore di Hopf è caratterizzato dalla presenza di elementi attivi che sostengano le vibrazioni in presenza di viscosità. Nelle cellule ciliate questi elementi attivi sono i motori cellulari presenti nel chinociglio, cioè macromolecole che trasformano l'energia chimica dell'ATP in energia meccanica. Questi motori si muovono lungo i filamenti del citoscheletro della cellula, e il loro moto è contrastato da forze di tipo elastico; la presenza di queste due forze antagoniste può portare o ad un equilibrio statico o ad oscillazioni. Se si considera un modello a due stati (attaccato e staccato dal filamento) la forza totale che agisce sul chinociglio è:
dove W1 è il potenziale nello stato legato e W2 quello nell'altro stato. freaz é la reazione data dal sistema delle stereociglia. Studiando la stabilità del sistema intorno all'equilibrio (v = 0) si trova la possibilità di una biforcazione di Hopf regolata dal parametro di controllo e con frequenza caratteristica , dove α è la somma dei rate di attacco e distacco dal filamento e β il rate di distacco. Poiché i rate sono regolati dalla concentrazione del calcio, il parametro di controllo è proprio funzione della concentrazione di quest'ultimo.
f=−v−K z−N∫0
L
dP1dW1P2dW 2 f reaz
0=K / 1/2
C=22/

Selftuning nelle cellule ciliatePer quanto visto prima si ha amplificazione non lineare del segnale solo in prossimità della biforcazione di Hopf; è quindi necessario un sistema di feedback che mantenga il parametro di controllo (la concentrazione del calcio) vicino al valore critico. Quello che succede è questo: con i canali ionici chiusi il Ca viene lentamente espulso da alcune pompe biochimiche nella membrana, mentre l'apertura dei canali provoca un flusso entrante di ioni. L'equazione per la concentrazione può quindi essere scritta come segue:
dove J0 è il flusso di ioni a canale aperto e P0(x) è la probabilità che quest'ultimo si apra come conseguenza di una deflessione x. Assumendo un andamento periodico per la deflessione e scomponendo C in una parte lenta e in una di modulazione si ottiene:
dove è la probabilità di apertura dei canali in presenza di oscillazioni mediata su un periodo.
dCdt
=−CJ 0 P0 x
C≃C0 t C1 ei t
dC0
dt≃−
C0
J 0
P0 ∣x1∣2
P0

Comportamento in presenza di rumore
L'effetto del rumore su un oscillatore di Hopf forzato e selftuned è stato studiato sia analiticamente che numericamente. In assenza della forzante esterna, il sistema oscilla alla frequenza caratteristica in maniera incoerente. Se si assume (per semplicità) che il neurone sottostante emetta uno spike quando la corrente in ingresso supera una certa soglia, si vede che l'incoerenza della fase si manifesta nel firing, che avviene in maniera casuale.La forzante, seppur debole, sincronizza l'oscillatore: la fase viene gradualmente bloccata. Quindi la presenza del rumore è fondamentale per distinguere le oscillazioni dovute ad uno stimolo esterno da quelle spontanee.

Comportamento in presenza di rumore
S. Camalet, T. Duke, F. Julicher, J. Prost, PNAS 97, 3183 (2000)

Comportamento in presenza di rumore
S. Camalet, T. Duke, F. Julicher, J. Prost, PNAS 97, 3183 (2000)

Secondo modello: la risonanza stocastica
Nel 1981 Benzi, Parisi e Vulpiani, studiando la periodicità delle ere glaciali terrestri, scoprirono un fenomeno nuovo e a dir poco sconcertante. Il loro modello climatico
prevedeva due stati stabili: uno caldo (reso stabile dall'effetto serra) e uno freddo (reso stabile dall'aumento dell'albedo collegato alla comparsa dei ghiacci). Classicamente la
transizione tra questi due stati era stata spiegata con le variazioni di eccentricità dell'orbita terrestre, il cui periodo è proprio quello delle glaciazioni (circa 100.000 anni).
Ma i tre fisici romani si accorsero che quest'effetto è troppo piccolo per spiegare il fenomeno: in realtà delle simulazioni (e una teoria nata conseguentemente) hanno mostrato come il rumore dovuto alle fluttuazioni termiche e ad altre perturbazioni
nell'orbita terrestre è in grado di amplificare un “segnale” deterministico.Quest'effetto è stato chiamato risonanza stocastica.

Risonanza stocastica: la doppia bucaConsideriamo un sistema bistabile come in figura.
Se mettiamo una particella in questo potenziale, questa andrà a finire in una delle due buche. Se però aggiungiamo del rumore, la particella oscillerà intorno ai minimi fino a quando non arriva una fluttuazione di intensità sufficiente a farla saltare dall'altra parte. La frequenza media dei salti è data dal rate di Kramers:
r K=0b
2exp−V
D
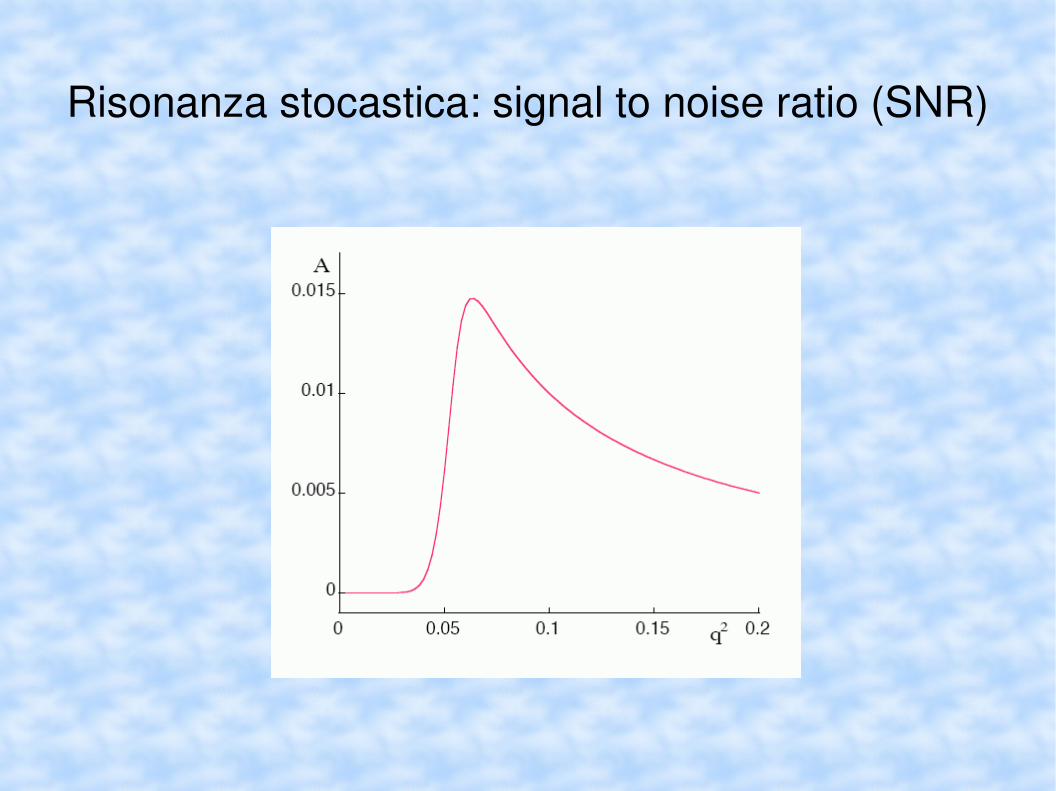
Risonanza stocastica: signal to noise ratio (SNR)

Bistabilità nelle cellule ciliate
Curva sperimentale forza – spostamentoda P. Nelson, Biological Physics, Freeman
Questa figura mostra come le stereociglia non si comportino in modo esattamente elastico. Anzi per piccoli spostamenti dalla posizione di equilibrio si ha una rigidità negativa. Questo può essere spiegato assumendo che lo stato del canale ionico influisca sulla rigidità del tip link (come pittorescamente mostrato dall'immagine del cancello) e che il canale stesso sia un sistema bistabile (aperto/chiuso).

Risonanza stocastica nelle cellule ciliateLe stereociglia sono quindi un sistema dinamico bistabile. In presenza di rumore (sia quello legato al moto delle ciglia che quello relativo al canale ionico) si può pensare di avere un fenomeno di risonanza stocastica.
Simulazioni fatte con parametri tipici per queste cellule* e dati sperimentali** sembrano confermare questa ipotesi.
* J. F. Lindner, M. Bennett, K. Wiesenfeld, Phys. Rev. E 72, 051911 (2005)** F. Jaramillo, K. Wiesenfeld, Chaos, Solitons and Fractals 11, 1869 (2000)

Modelli a confrontoIn questo seminario ho presentato due modelli correntemente utilizzati per cercare di interpretare la sorprendente capacità dell'orecchio di distinguere suoni debolissimi. Ecco a confronto gli aspetti salienti:
Oscillatore di Hopf forzato e selftuned
sincronizzazione deterministica(phaselocking)
amplificazione non lineare in ampiezza(i segnali piccoli sono più amplificati)
risonanza legata alla frequenza caratteristica del chinociglio
Risonanza stocastica
sincronizzazione statistica(no phaselocking)
amplificazione lineare in ampiezza
risonanza legata alla noise strength
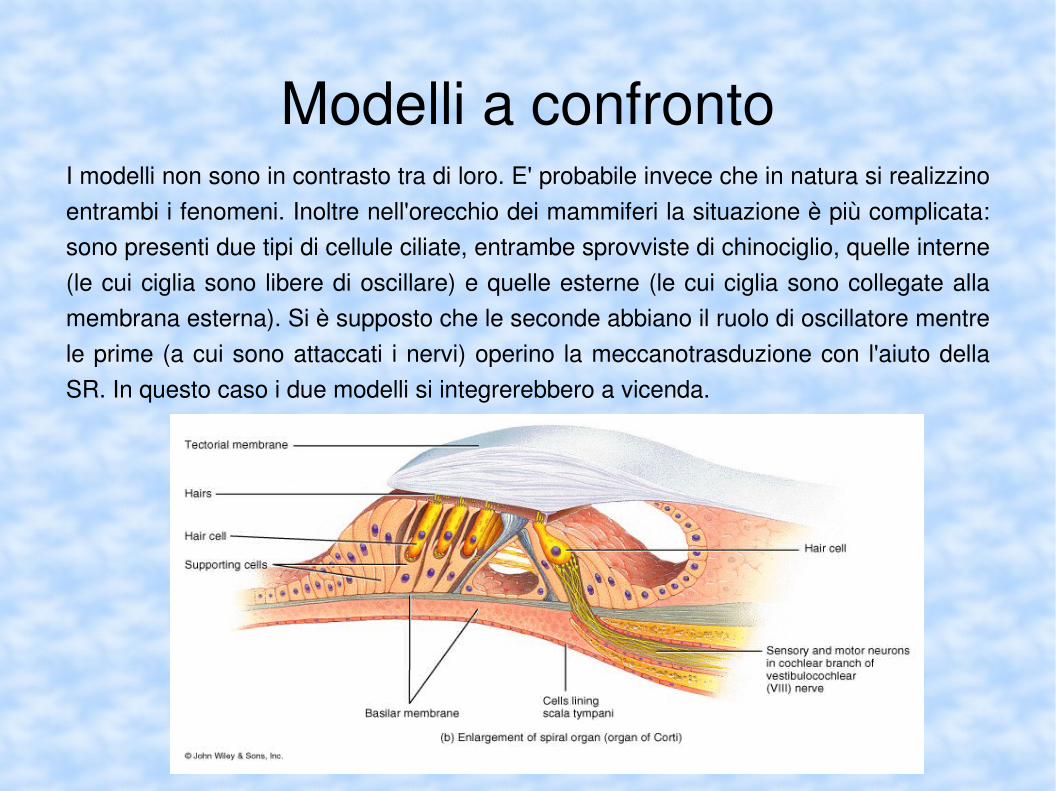
Modelli a confrontoI modelli non sono in contrasto tra di loro. E' probabile invece che in natura si realizzino entrambi i fenomeni. Inoltre nell'orecchio dei mammiferi la situazione è più complicata: sono presenti due tipi di cellule ciliate, entrambe sprovviste di chinociglio, quelle interne (le cui ciglia sono libere di oscillare) e quelle esterne (le cui ciglia sono collegate alla membrana esterna). Si è supposto che le seconde abbiano il ruolo di oscillatore mentre le prime (a cui sono attaccati i nervi) operino la meccanotrasduzione con l'aiuto della SR. In questo caso i due modelli si integrerebbero a vicenda.

BibliografiaM. Tabor, Chaos and integrability in nonlinear dynamics, WileyInterscience 1989;
Gammaitoni, P. Hanggi, P. Jung, F. Marchesoni, “Stochastic resonance”, Rev. Mod. Phys. 70, 223 (1998);
S. Camalet, T. Duke, F. Julicher, J. Prost, “Auditory sensitivity provided by selftuned critical oscillations of hair cells”, PNAS 97, 3183 (2000);
J. Balakrishnan, “Selftuning to the Hopf bifurcation in fluctuating systems”, J. Phys. A 38, 1627 (2005);
F. Jaramillo, K. Wiesenfeld, “Physiological noise level enhances mechanoelectrical transduction in hair cells”, Chaos, solitons and fractals 11, 1869 (2000);
J.F. Lindner, M. Bennett, K. Wiesenfeld, “Stochastic resonance in the mechanoelectrical transduction of hair cells”, Phys. Rev. E 72, 051911 (2005).


















