3. Fisiologia Cellulare Potenziale transmembranario ... Generale... · • Impulso nervoso: onda di...
25
3. Fisiologia Cellulare Potenziale transmembranario, potenziale d’azione Prof. Carlo Capelli Fisiologia Laurea in Scienze delle attività motorie e sportive Università di Verona
Transcript of 3. Fisiologia Cellulare Potenziale transmembranario ... Generale... · • Impulso nervoso: onda di...

3. Fisiologia Cellulare Potenziale
transmembranario, potenziale d’azione
Prof. Carlo Capelli Fisiologia
Laurea in Scienze delle attività motorie e sportive
Università di Verona

Obiettivi • Genesi e mantenimento del potenziale di membrana a
riposo • Potenziale di equilibrio ed equazione di Nernst • Equazione di Goldman e PMR del neurone • Potenziale di azione: descrizione delle fasi e
determinanti fisico-chimici • Andamento delle conduttanze del sodio e del potassio
nel corso del PA • Periodi refrattari • Propagazione del PA nelle fibre amieliniche e
mieliniche
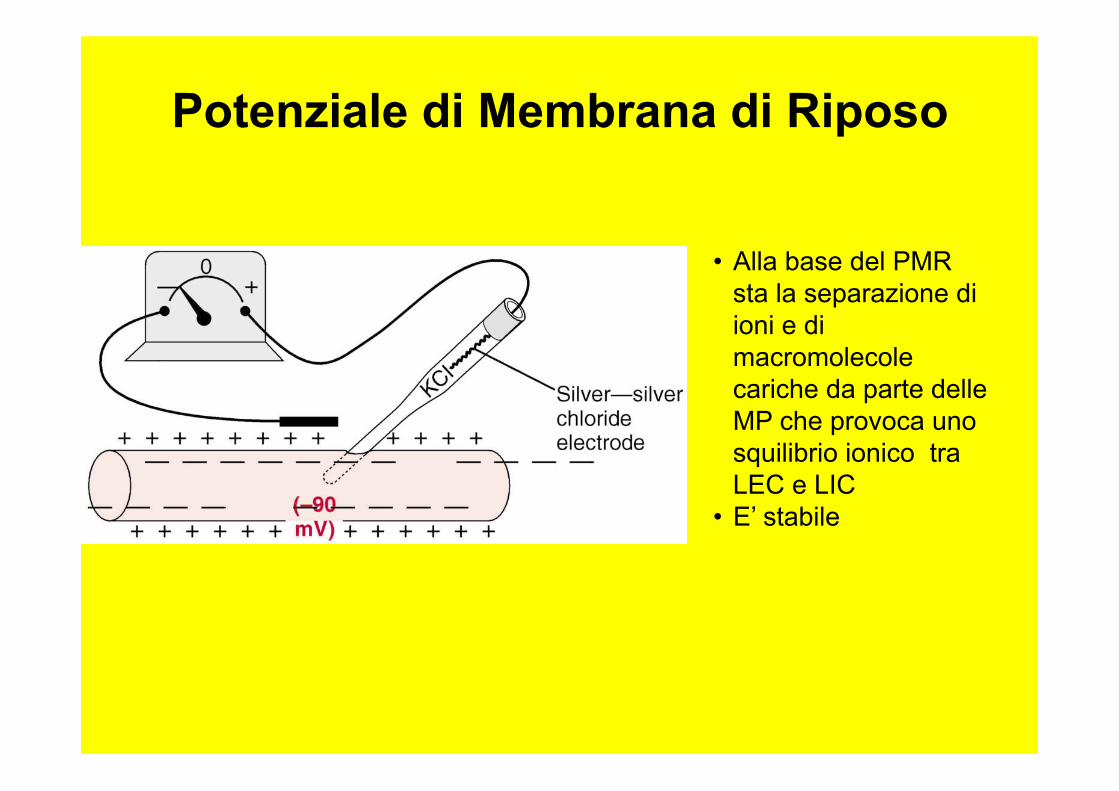
Potenziale di Membrana di Riposo
• Alla base del PMR sta la separazione di ioni e di macromolecole cariche da parte delle MP che provoca uno squilibrio ionico tra LEC e LIC
• E’ stabile

Potenziale di Membrana di Riposo
• Gli ioni organici (proteine) sono trattenuti all’interno delle cellule • A cavallo della membrana cellulare gli ioni NON sono presenti ad uguale
concentrazione (in uno strato sottilissimo), anche se in ciascun compartimento la somma delle cariche negative e di quelle negative è uguale a 0: elettroneutralità
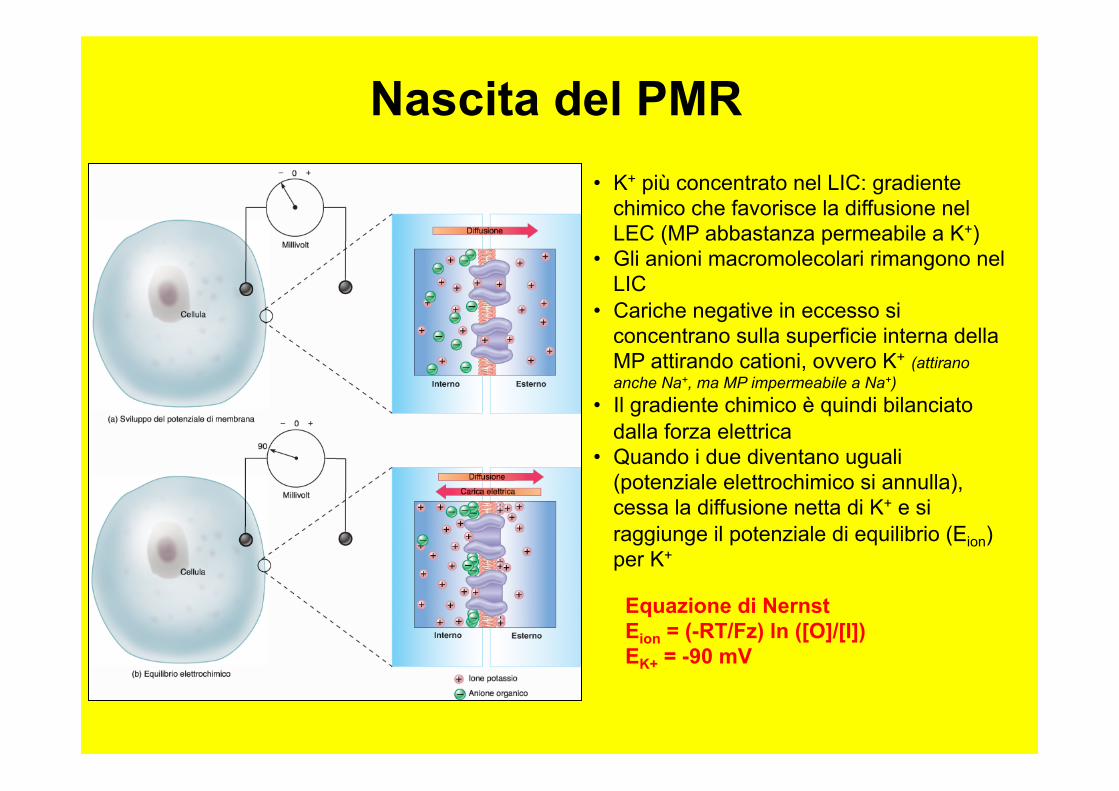
Nascita del PMR • K+ più concentrato nel LIC: gradiente
chimico che favorisce la diffusione nel LEC (MP abbastanza permeabile a K+)
• Gli anioni macromolecolari rimangono nel LIC
• Cariche negative in eccesso si concentrano sulla superficie interna della MP attirando cationi, ovvero K+ (attirano anche Na+, ma MP impermeabile a Na+)
• Il gradiente chimico è quindi bilanciato dalla forza elettrica
• Quando i due diventano uguali (potenziale elettrochimico si annulla), cessa la diffusione netta di K+ e si raggiunge il potenziale di equilibrio (Eion) per K+
Equazione di Nernst Eion = (-RT/Fz) ln ([O]/[I]) EK+ = -90 mV

µ = µo + R T l C + z F E
µ0. Potenziale elettrochimico della sostanza in condizioni di riferimento (1M , 0 °C, E = 0)
R: costante universale dei gas
T: temperatura assoluta in gradi K
lnC: logaritmo naturale della concentrazione C
Z: numero di cariche dello ione
F: numero di Faraday
E: potenziale elettrico
Il potenziale elettrochimico

µ è espresso in unità di energia per mole
Rappresenta l’energia potenziale posseduta da una mole di ioni ed è il risultato della concentrazione e del potenziale vigenti
Il flusso di ioni va dal sito in cui µ è più alto a quello in cui µ è più basso
Differenza di potenziale elettrochimico dello ione x a cavallo della membrana.
∆ µ = µ A(x) - µ B(x)
Il significato del potenziale elettrochimico

µA(x) = µoA(x) + RTln[x]A + zFEA
µB(x) = µoB(x) + RTln [x]B + zFEB
Δµ (x) = µA(x) - µB(x) = RTln [x]A/[x]B + zF (EA - EB)
Δµ (x) = 0 = µA(x) - µ B(x) = RTln [x]A/[x]B + zF (EA - EB)
Potenziale elettrochimico e condizioni di equilibrio

(EA - EB) = - RT/ zF ln [x]A/[x]B = RT/ zF ln [x]B/[x]A
EQUAZIONE di NERNST E’ valida solo per gli ioni all’equilibrio
• Quantifica la differenza di potenziale elettrico (EA - EB) richiesta per produrre una forza elettrostatica (zF (EA - EB)) uguale ed opposta a quella chimica (RTln ([x]A/[x]B)) che tende a far diffondere x da A a B
Risoluzione per condizione di equilibrio: l’Equazione di Nernst

Cl - extracellulare = 120 mM Cl - intracellulare = 3.5 mM
Pot equilibrio = -89 mV
• Cl - é in equilibrio EC: non tende a diffondere
• La membrana è permeabile a Cl-
Na + extracellulare = 120 mM Na + intracellulare = 9.2 mM
Pot equilibrio = +67 mV
• Na+ non é in equilibrio EC: tende a diffondere
• La membrana è poco permeabile a Na+
K + extracellulare = 2.5 mM K + intracellulare = 140 mM
Pot equilibrio = -105 mV
• K+ non é in equilibrio EC di poco: tende a diffondere meno del sodio
• La membrana é abbastanza permeabile a K+
• Poichè PMR è diverso da EK+, K+ tende a diffondere
Potenziale di riposo della membrana neuronale e potenziali di equililbrio
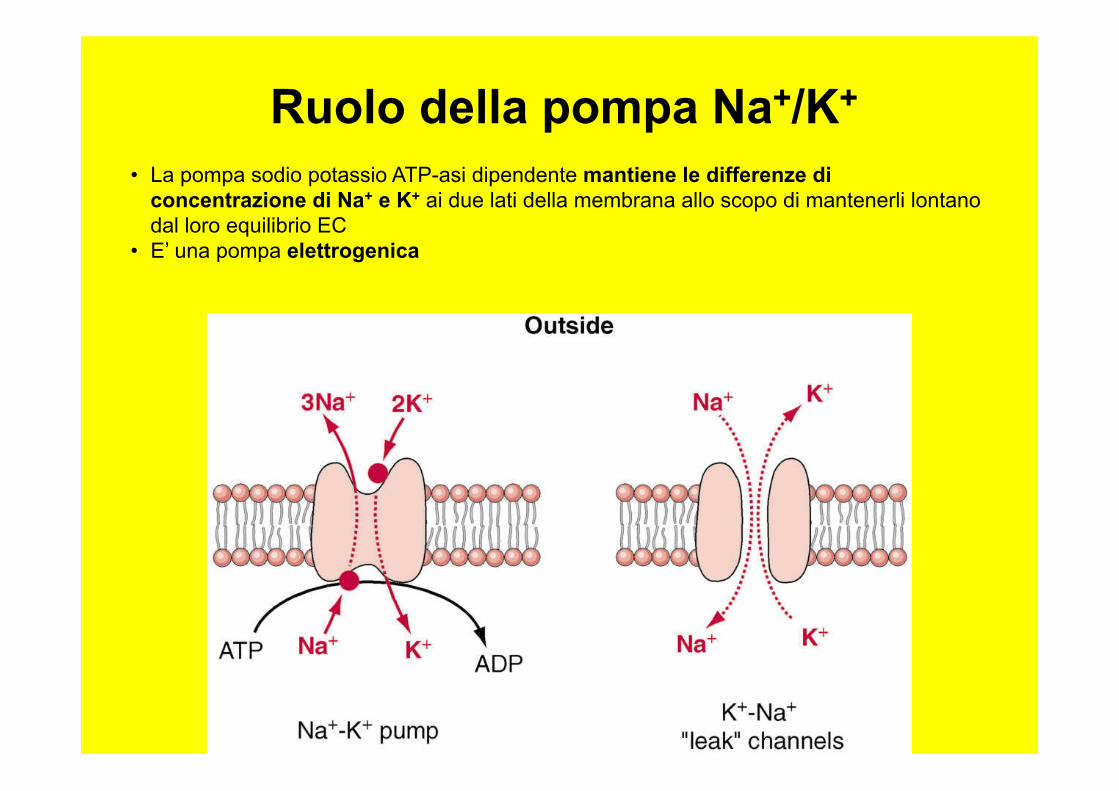
• La pompa sodio potassio ATP-asi dipendente mantiene le differenze di concentrazione di Na+ e K+ ai due lati della membrana allo scopo di mantenerli lontano dal loro equilibrio EC
• E’ una pompa elettrogenica
Ruolo della pompa Na+/K+

Equazione di Goldman
• Benchè il PRM sia molto vicino a EK+, e la pompa Na+/K+ mantenga il gradiente di concentrazione per la diffusione di K+, l’effettivo valore del potenziale è determinato dalle concentrazioni extra ed intracellulari di tutti gli ioni nelle immediate vicinanze della MP
• Per gli ioni potassio, sodio e cloro questa relazione è espressa dall’equazione di Goldman
• Di fatto è la somma delle equazioni di Nernst delle tre speci ioniche pesate per le rispettive conduttanze P
Er = RTF ln
PK K+⎡
⎣
⎢⎢⎢⎢
⎤
⎦
⎥⎥⎥⎥O
+PNa Na+⎡
⎣
⎢⎢⎢⎢
⎤
⎦
⎥⎥⎥⎥O
+PCl Cl−⎡
⎣
⎢⎢⎢⎢
⎤
⎦
⎥⎥⎥⎥I
PK K+⎡
⎣
⎢⎢⎢⎢
⎤
⎦
⎥⎥⎥⎥I
+PNa Na+⎡
⎣
⎢⎢⎢⎢
⎤
⎦
⎥⎥⎥⎥I
+PCl Cl−⎡
⎣
⎢⎢⎢⎢
⎤
⎦
⎥⎥⎥⎥O

Il potenziale d’azione • Depolarizzazione il PM diventa meno negativo (da -70 a -50) • Overshoot il PM supera lo zero elettrico • Iperpolarizzazione il PM diventa più negativo (da -70 a -90) • Ripolarizzazione il PM torna a PMR ( a -70 mV)

• I canali per il sodio hanno due cancelli uno di attivazione e uno di inattivazione. L’azione è simultanea e controllata dal voltaggio.
• Quelli di inattivazione sono però più lenti … il flusso di cariche positive si arresta prima che il sodio raggiunga il suo potenziale elettro chimico (+67 mV)
• Il processo induce un rapido, ma transitorio aumento della conduttanza per aumento della permeabilità di membrana per il sodio
Genesi del potenziale d’azione: canali voltaggio dipendenti per il sodio

Genesi del potenziale d’azione: canali voltaggio dipendenti per il sodio
• I canali per il sodio si aprono quando il potenziale di membrana diventa meno negativo. L’entrata di particelle positive riduce ulteriormente la negatività sul lato citoplasmatico e altri canali si aprono (effetto a cascata).
• Il PA è una risposta massimale tutto-o-nulla: una volta raggiunto il potenziale di soglia, il PA si autogenera ed evolve spontaneamente.
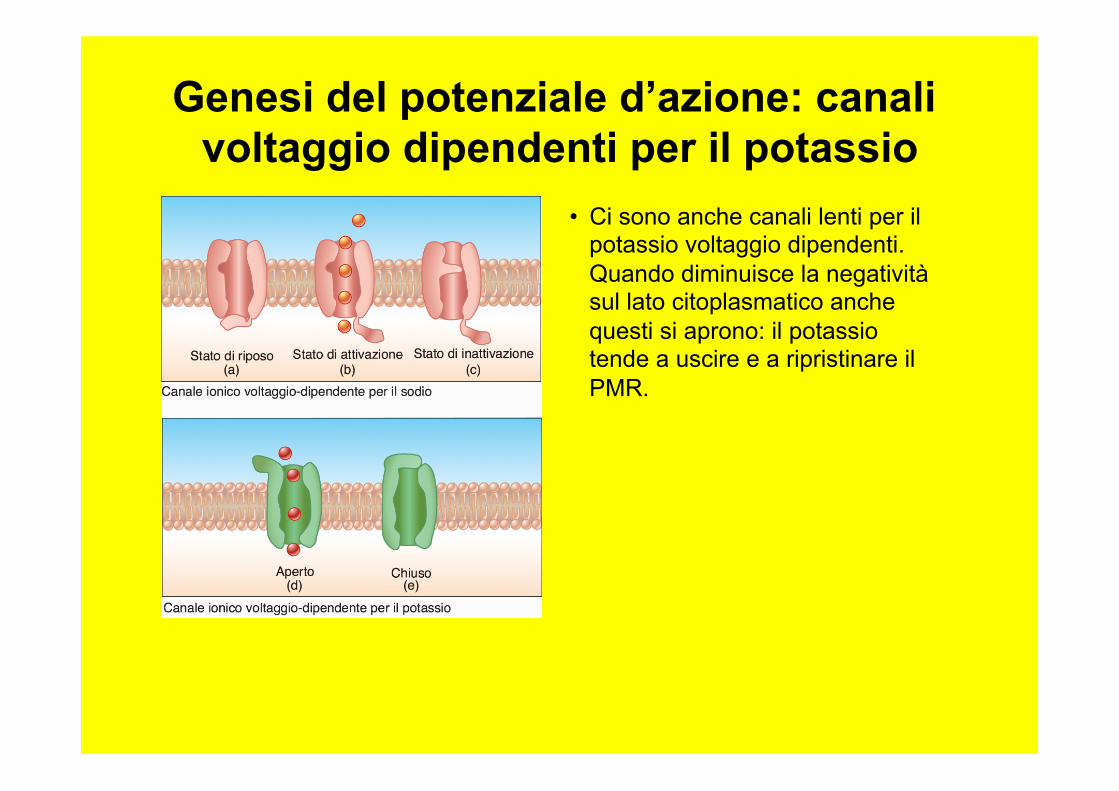
• Ci sono anche canali lenti per il potassio voltaggio dipendenti. Quando diminuisce la negatività sul lato citoplasmatico anche questi si aprono: il potassio tende a uscire e a ripristinare il PMR.
Genesi del potenziale d’azione: canali voltaggio dipendenti per il potassio
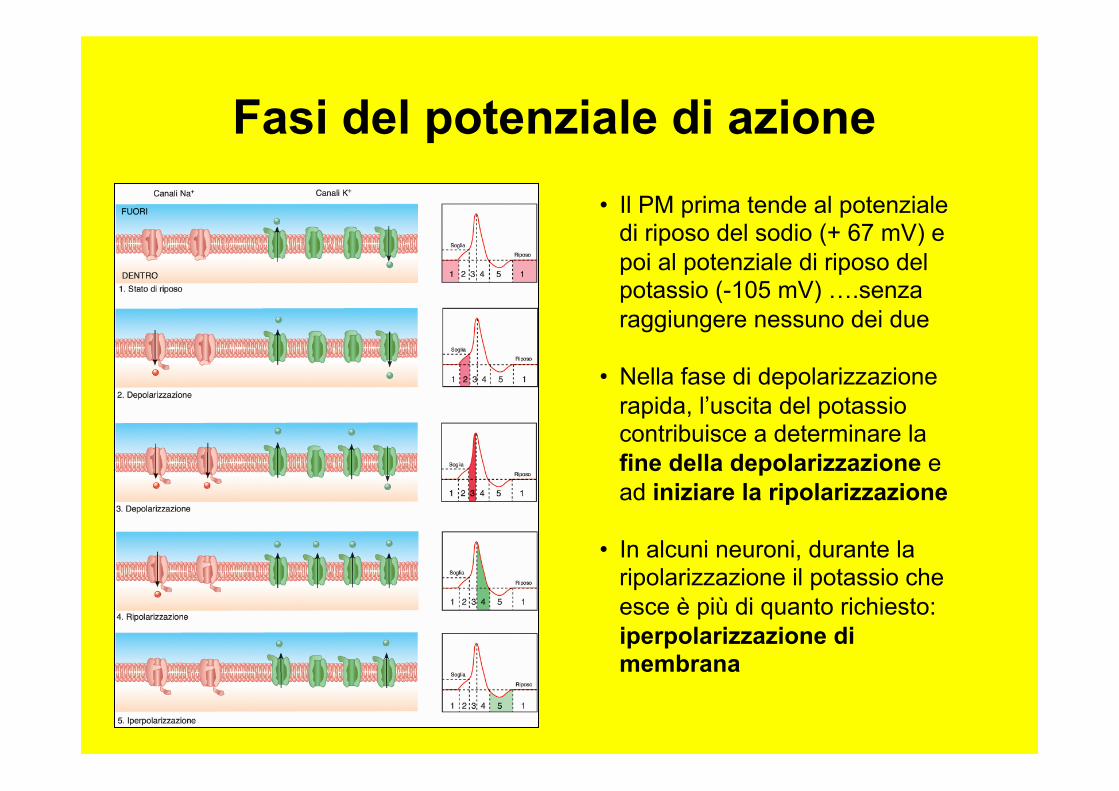
Fasi del potenziale di azione • Il PM prima tende al potenziale
di riposo del sodio (+ 67 mV) e poi al potenziale di riposo del potassio (-105 mV) ….senza raggiungere nessuno dei due
• Nella fase di depolarizzazione rapida, l’uscita del potassio contribuisce a determinare la fine della depolarizzazione e ad iniziare la ripolarizzazione
• In alcuni neuroni, durante la ripolarizzazione il potassio che esce è più di quanto richiesto: iperpolarizzazione di membrana
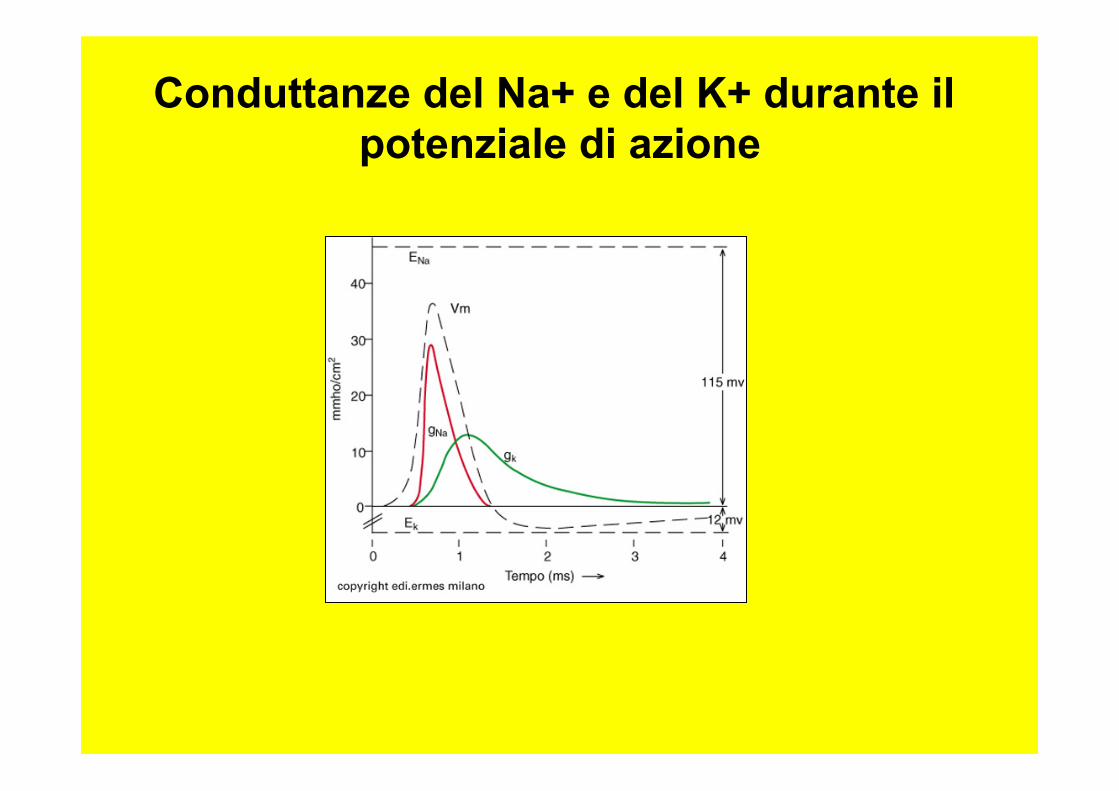
Conduttanze del Na+ e del K+ durante il potenziale di azione
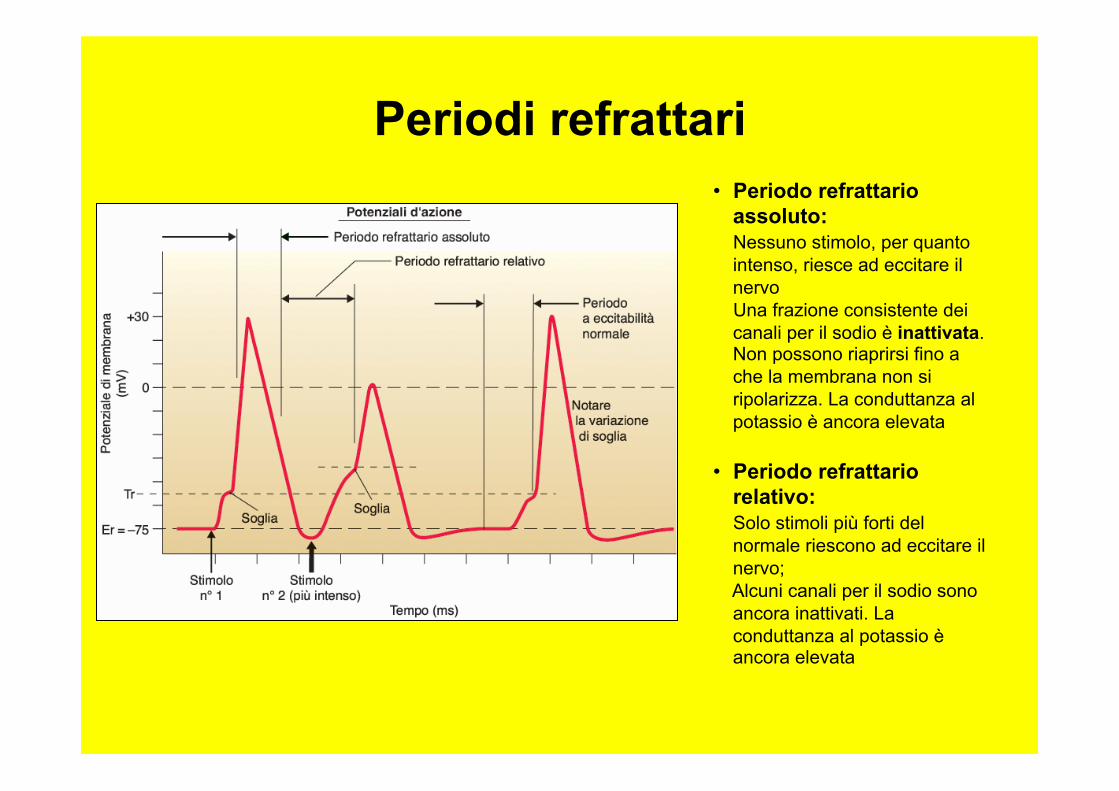
• Periodo refrattario assoluto: Nessuno stimolo, per quanto intenso, riesce ad eccitare il nervo Una frazione consistente dei canali per il sodio è inattivata. Non possono riaprirsi fino a che la membrana non si ripolarizza. La conduttanza al potassio è ancora elevata
• Periodo refrattario relativo: Solo stimoli più forti del normale riescono ad eccitare il nervo; Alcuni canali per il sodio sono ancora inattivati. La conduttanza al potassio è ancora elevata
Periodi refrattari
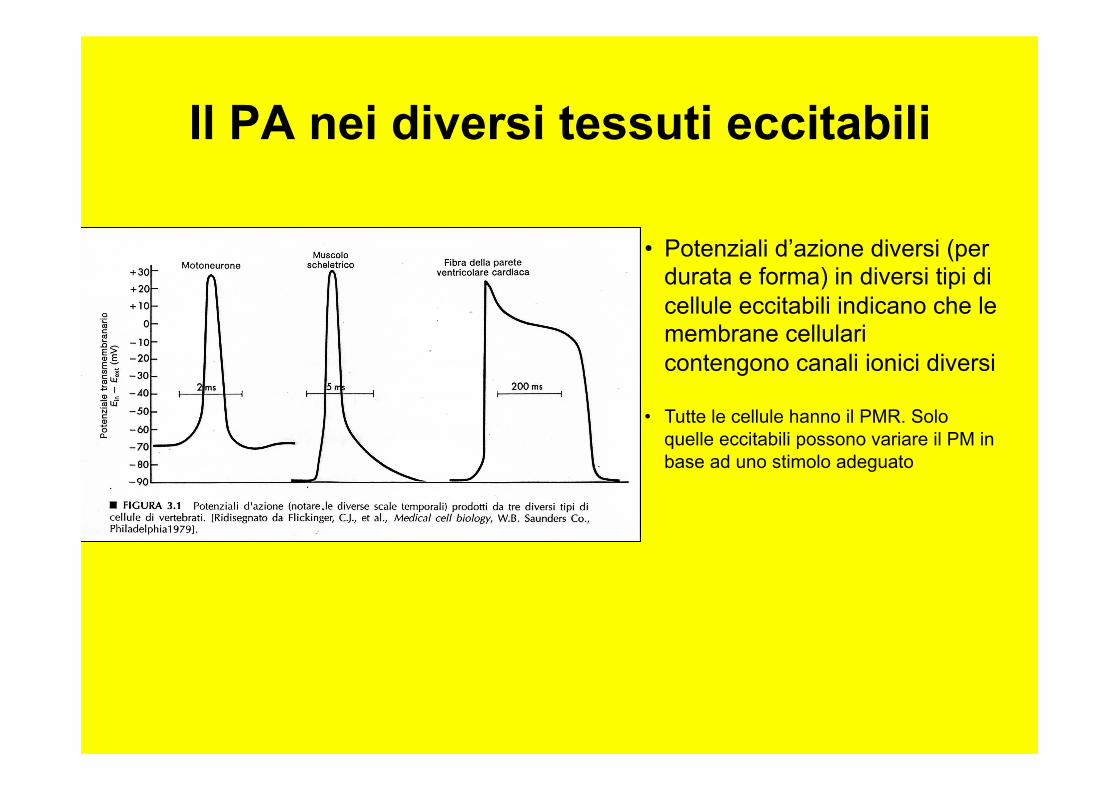
• Potenziali d’azione diversi (per durata e forma) in diversi tipi di cellule eccitabili indicano che le membrane cellulari contengono canali ionici diversi
• Tutte le cellule hanno il PMR. Solo quelle eccitabili possono variare il PM in base ad uno stimolo adeguato
Il PA nei diversi tessuti eccitabili

Propagazione di PA-fibre amieliniche
• Impulso nervoso: onda di depolarizzazione che corre lungo l’assone
• Zona del PA iniziale: punto di scarico in cui ioni Na+ fluiscono verso l’interno
• Ioni Na+ sono richiamati da zone adiacenti (sorgente) della membrana e la corrente assume un andamento circolare
• L’accumulo di cariche positive lungo l’assone dalla parte interna della membrana nelle zone sorgenti può raggiungere i potenziale di soglia ed innescare il PA
• Ripetendosi questo meccanismo, tratti successivi dell’assone possono depolarizzarsi e l’impulso si trasmette.
• La velocità di conduzione nelle fibre amieleniche è proporzionale al diametro delle fibre
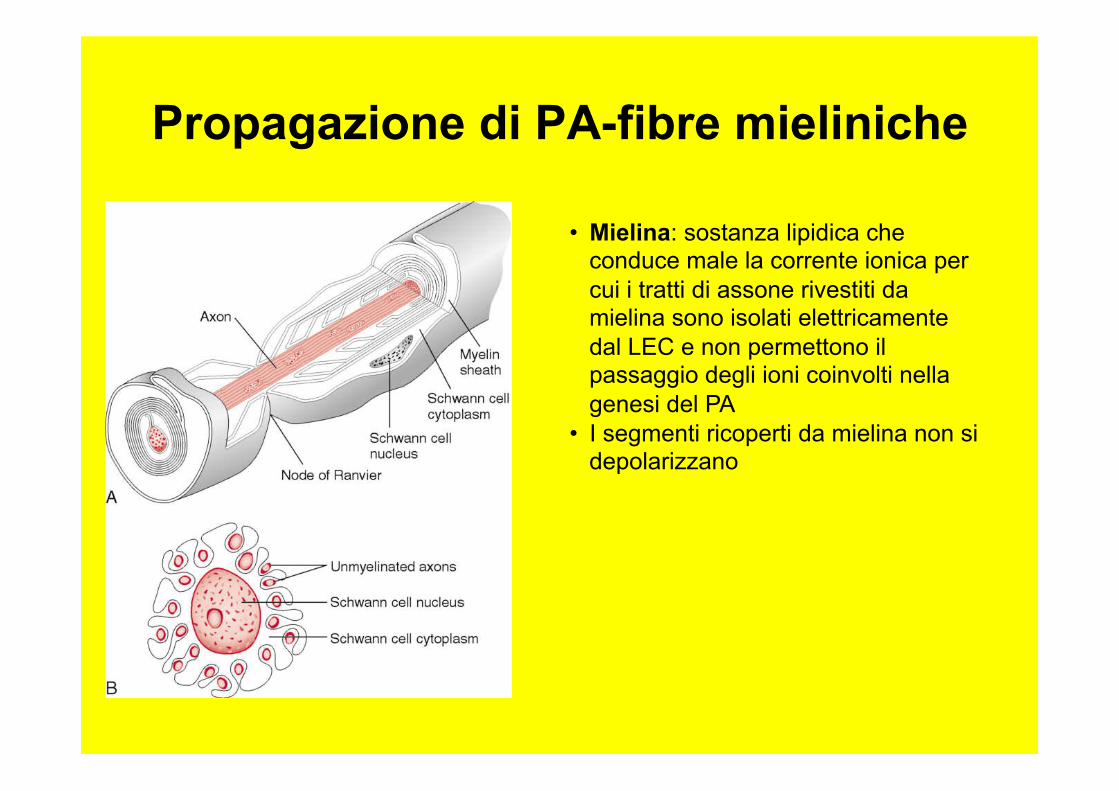
Propagazione di PA-fibre mieliniche
• Mielina: sostanza lipidica che conduce male la corrente ionica per cui i tratti di assone rivestiti da mielina sono isolati elettricamente dal LEC e non permettono il passaggio degli ioni coinvolti nella genesi del PA
• I segmenti ricoperti da mielina non si depolarizzano
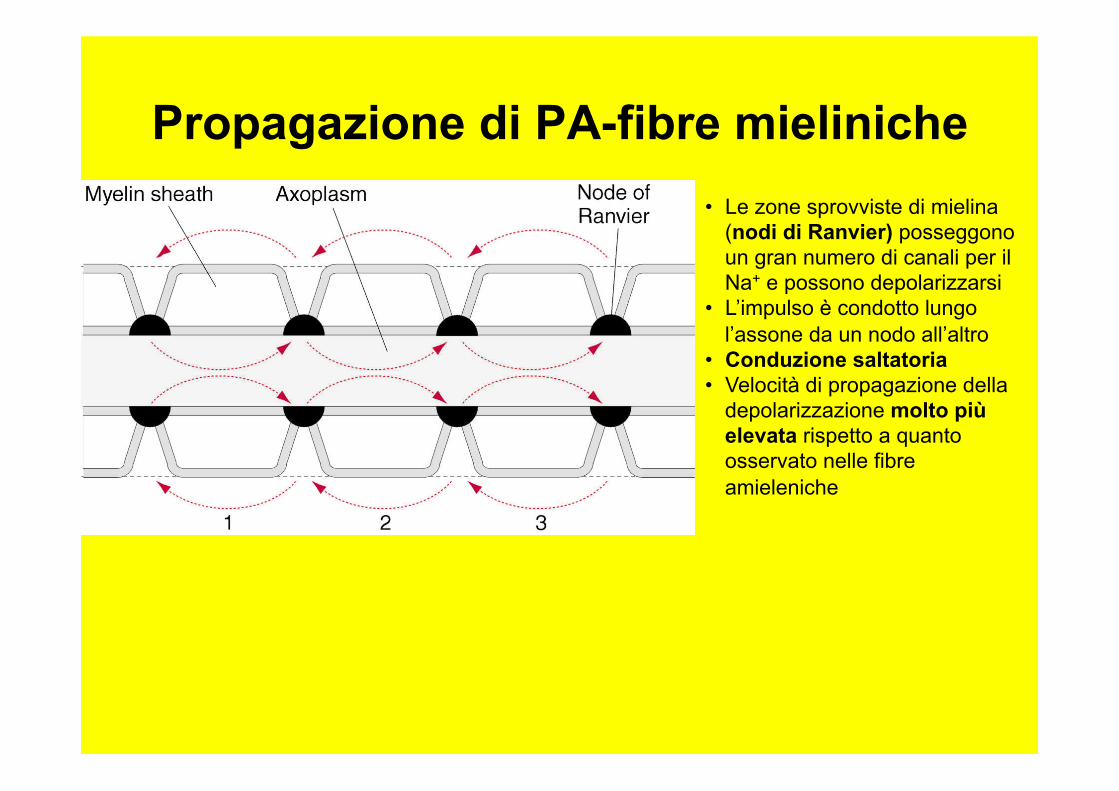
Propagazione di PA-fibre mieliniche • Le zone sprovviste di mielina
(nodi di Ranvier) posseggono un gran numero di canali per il Na+ e possono depolarizzarsi
• L’impulso è condotto lungo l’assone da un nodo all’altro
• Conduzione saltatoria • Velocità di propagazione della
depolarizzazione molto più elevata rispetto a quanto osservato nelle fibre amieleniche

Classificazione delle fibre nervose
• Fibre del gruppo A • Fibre mieliniche nervi periferici • 5- 120 m/s • Alfa: 80 – 120 m/s • Beta: 35-80 m/s • Delta: 5-30 m/s
• Fibre del gruppo B • Fibre mieliniche del sistema nervoso autonomo • < 50 m/s
• Fibre del gruppo C • Fibre amieliniche a lenta conduzione • < 2 m/s

Bibliografia
• Fisiologia dell’Uomo, autori vari, Edi.Ermes, Milano • Capitolo : Neurofisiologia generale (Capitoli 1.3, 1.4, 1.5)
• Fisiologia Generale ed Umana, Rhoades-Pflanzer • Capitolo 7: Organizzazione generale del sistema nervoso