UNIVERSITA’ DEGLI STUDI DI PAVIA - infn.it − Metodo Monte Carlo − Distribuzione in frequenza...
56
3 UNIVERSITA’ DEGLI STUDI DI PAVIA FACOLTA’ DI SCIENZE MATEMATICHE, FISICHE, NATURALI CORSO DI LAUREA IN FISICA PRINCIPI DI BASE DELLA MICRODOSIMETRIA ED APPLICAZIONI IN BNCT Relazione per la laurea di Elisa Bonasegla Relatore Prof. Saverio Altieri Dipartimento di Fisica Nucleare e Teorica Anno accademico 2007/2008
Transcript of UNIVERSITA’ DEGLI STUDI DI PAVIA - infn.it − Metodo Monte Carlo − Distribuzione in frequenza...

3
UNIVERSITA’ DEGLI STUDI DI PAVIA FACOLTA’ DI SCIENZE MATEMATICHE, FISICHE, NATURALI
CORSO DI LAUREA IN FISICA
PRINCIPI DI BASE DELLA MICRODOSIMETRIA ED APPLICAZIONI IN
BNCT Relazione per la laurea di Elisa Bonasegla
Relatore Prof. Saverio Altieri Dipartimento di Fisica Nucleare e Teorica
Anno accademico 2007/2008

4
INDICE
INDICE pag. 4
INTRODUZIONE 6
CAPITOLO 1. MICRODOSIMETRIA 9 1.1 CARATTERISTICHE ED APPLICAZIONI 9
1.2 GRANDEZZE FISICHE DI RIFERIMENTO CONVENZIONALI 10
− Fluenza
− Libero cammino medio e range
− Linear Energy Transfer (LET)
− Dose assorbita
1.3 GRANDEZZE STOCASTICHE E LORO DISTRIBUZIONE 15
− Energia specifica
− Energia lineale
CAPITOLO 2. IL PROCESSO COMPOSTO DI POISSON 20
2.1 CASUALITA’ DEGLI EVENTI 20
2.2 EQUAZIONI POISSONIANE 22
− Equazione base
− Relazioni per i momenti
2.3 IMPLICAZIONI BIOFISICHE DELLA MICRODOSIMETRIA 26
− Modello a più urti e modello a bersagli multipli
− Modello a soglia
CAPITOLO 3. APPLICAZIONE AI SISTEMI BIOLOGICI 30
3.1 MICRODOSIMETRIA NEI SISTEMI BIOLOGICI 30
3.2 LINEAR ENRGY TRANSFER (LET) 32
− Definizione e distribuzione

5
− Efficacia Biologica Relativa (RBE)
3.3 CONTATORI PROPORZIONALI 36
3.4 PROFILI DI TRACCIA ED ENTITA’ DI TRACCIA 38
3.5 SIMULAZIONE DELLA STRUTTURA DELLE TRACCE 39
− Metodo Monte Carlo
− Distribuzione in frequenza del deposito di energia
CAPITOLO 4. MICRODOSIMETRIA E BNCT 44
4.1 MICRODOSIMETRIA DELLA REAZIONE B10 44
4.2 MICRODOSIMETRIA SPERIMENTALE 47
4.3 STRUMENTAZIONI 48
− TEPC sferico
− TEPC piano
− TEPC cilindrico
− Mini TEPC
− Telescopio a pixel al silicio
CONCLUSIONI 56
BIBLIOGRAFIA 57

6
INTRODUZIONE
La microdosimetria studia le proprietà microscopiche del deposito di energia
nell’interazione delle radiazioni ionizzanti con la materia, ponendo l’accento sulla
disomogeneità e sulla natura stocastica delle interazioni; al contrario la dosimetria
convenzionale fa riferimento alla dose assorbita che è una quantità definita attraverso il
deposito di energia su volumi macroscopici. In molte situazioni la dose assorbita risulta
totalmente inadeguata a descrivere l’effetto della radiazione nei materiali biologici perché i
meccanismi e gli effetti sono dominati da depositi disomogenei in siti di dimensioni cellulari
o sub-cellulari.
Nonostante la microdosimetria sia nata sulla spinta degli studi radiobiologici essa risulta
utile in vari altri campi della fisica. Le tecniche ed i metodi adoperati, forniscono descrizioni
fisiche dei campi di radiazione e la loro interazione con la materia e questo è indipendente da
ogni particolare applicazione pratica o teorica a sistemi biologici o di qualsiasi altra natura.
Difatti, la microdosimetria offre un utile supporto a discipline quali la microelettronica e la
fisica dello stato solido, oltre a fornire contributi significativi nei campi ormai classici della
radiobiologia, della radioterapia e della radioprotezione.
Alcune delle applicazioni fondamentali della microdosimetria si hanno in
radioprotezione. In radioprotezione si differenzia di solito tra effetti stocastici, ad esempio
danni ereditari, ed effetti non-stocastici, ad esempio danni alla pelle, perdita delle fertilità,
malformazioni prenatali. L’obiettivo della radioprotezione è quello di evitare interamente gli
effetti non-stocastici, dipendenti dai danni provocati nella moltiplicazione cellulare in un
tessuto con una risultante dipendenza, curvilinea o a soglia, dalla dose. Si va quindi a
minimizzare l’esposizione alla radiazioni, riducendo il rischio di effetti stocastici ad un livello
simile od inferiore ad altri rischi associati ad attività umane. Gli effetti stocastici invece non
possono essere interamente eliminati. Essi risultano dal danno a cellule individuali ed hanno
una dipendenza lineare dalla dose quando questa è bassa.
Nell’ambito della radioterapia la microdosimetria trova importanti applicazioni nel
campo della terapia per cattura neutronica sul boro (BNCT). Questa tecnica, basata sulla
reazione B10 (n,α) Li7 , tratta i tessuti cancerosi con fasci di neutroni termici (En < 0.5 eV) od
epitermici (0.5 eV < En < 10 keV); l’irraggiamento neutronico viene effettuato dopo aver

7
prodotto nella massa tumorale (usando opportuni composti borotrasportatori arricchiti
dell’isotopo B10 ) una concentrazione di boro più alta che nei tessuti sani. La reazione di
cattura produce due ioni carichi che a causa del loro forte potere ionizzante e del loro breve
range nel tessuto (dell’ordine del diametro cellulare) cedono la maggior parte della loro
energia entro il volume della singola cellula in cui sono generate. Nel trattamento BNCT,
quindi, intervengono contemporaneamente diversi tipi di radiazioni (particelle alfa, ioni litio,
protoni, raggi gamma …) i cui depositi di energia hanno caratteristiche diverse. In questo
campo, perciò, la microdosimetria può fornire un utile supporto nella caratterizzazione dei
vari fasci usati per la terapia e per lo studio dell’efficacia biologica dei vari composti usati
per trasportare il boro.

8

9
1. MICRODOSIMETRIA
1.1 CARATTERISTICHE ED APPLICAZIONI
Lo studio degli effetti delle radiazioni sulla materia vivente è complicato dal fatto che
qualsiasi tentativo che cerca di spiegarli deve fare i conti con i fenomeni che insorgono in un
mezzo biologico in seguito ad un’irradiazione e deve tener presente la complessità insita nei
meccanismi biologici. Da una parte è necessaria una precisa descrizione del campo delle
radiazioni, e dall’altra servono informazioni sulle distribuzioni di energia in strutture
biologiche microscopiche, o sulle concentrazioni locali delle specie chimiche prodotte dalla
radiazione.
Per il primo punto si possono utilizzare quantità radiometriche, come la fluenza delle
particelle con la corrispondenti distribuzioni spaziali, temporali ed energetiche; la
distribuzione spaziale dovrebbe essere nota a livello cellulare o addirittura subcellulare.
D’altro canto è richiesta la conoscenza delle caratteristiche legate ai materiali irraggiati quali
ad esempio le sezioni d’urto e lo stopping power. In questo caso anche se oggi, grazie a
metodi di simulazione Monte Carlo, è possibile effettuare calcoli per ottenere una descrizione
dettagliata delle varie distribuzioni, la mole di dati da registrare è talmente grande che risulta
quasi impossibile estrarre delle informazioni utili alla comprensione dei fenomeni biologici.
Infatti a livello microscopico, i depositi energetici cambiano notevolmente in base ai diversi
tipi di radiazione e ai diversi livelli di risoluzione spaziale in modo tale che anche la
descrizione o simulazione più dettagliata contribuisce poco al riconoscimento di quei
parametri critici per gli effetti biologici.
Il ruolo della microdosimetria è quindi quello di individuare dei parametri essenziali che
possano aiutare ad interpretare gli effetti biologici partendo da diversi punti di vista. Una
semplificazione iniziale consiste, per esempio, nel trattare esclusivamente il campo di
radiazione delle particelle cariche direttamente ionizzanti. Si considerano solo le tracce di
queste particelle che hanno energia sufficiente per produrre la maggioranza delle eccitazioni e
ionizzazioni elettroniche responsabili degli effetti biologici.
Nello studio delle caratteristiche delle deposizioni di energia lungo le tracce delle
particelle cariche e la risultante efficacia biologica delle radiazioni, in microdosimetria

10
possono essere utilizzati due approcci differenti.
Il primo fissa determinate strutture nelle cellule, detti siti, e postula che l’effetto biologico
venga determinato dalla quantità di energia depositata in queste strutture. In prima
approssimazione si può trascurare la distribuzione spaziale del deposito di energia all’interno
del sito, nonostante questa, in generale possa, avere un’influenza nella determinazione
dell’efficacia biologica. Le distribuzioni di probabilità dell’energia depositata dipendono
infatti da forma e dimensione della struttura e dal tipo di radiazione.
Il secondo approccio, reso possibile dall’avanzamento di metodi computazionali e di
simulazione di tracce di particelle cariche, si occupa invece di determinare la struttura
microgeometrica dei depositi di energia. In questo secondo caso le caratteristiche dei depositi
di energia non sono più relazionati ad un sito, ovvero ad una geometria di riferimento, bensì
alla sola struttura spaziale delle tracce stesse.
1.2 GRANDEZZE FISICHE DI RIFERIMENTO CONVENZIONALI
Alcune caratteristiche basilari della microdistribuzione dei depositi di energia, da parte
delle radiazioni ionizzanti, possono essere descritte in termini di parametri convenzionali;
questo facilita l’introduzione dei concetti microdosimetrici e la loro interpretazione.
Fluenza
La quantità radiometrica più importante è la fluenza Φ di un tipo specifico di radiazione.
Essa è definita come numero medio o valore di aspettazione delle particelle entranti in una
sfera con cerchio massimo di area unitaria ovvero con diametro pari a π/2=d . Questa
definizione è equivalente a dire che, in media, Φ/2 particelle attraversano una superficie piana
di area unitaria con orientazione qualunque, o, in un campo unidirezionale, Φ particelle
attraversano un elemento con superficie unitaria ortogonale alla direzione del campo. Una
definizione alternativa è che la fluenza è uguale alla lunghezza totale media delle traiettorie
delle particelle per unità di volume.
In Fig. 1 è mostrata la fluenza per unità di dose assorbita nell’acqua per fotoni
monoenergetici, neutroni e particelle cariche. Si nota come la fluenza delle particelle neutre
sia molto maggiore rispetto a quella delle particelle cariche. A livello cellulare o subcellulare,
nei tessuti pur in presenza di molti attraversamenti di particelle neutre si possono verificare
poche interazioni da parte di particelle cariche che sono quelle che danno contributo alla dose.

11
Fig. 1. Fluenza per unità di dose assorbita nell’acqua di particelle cariche e non cariche con date energie iniziali [1].
Libero cammino medio e range
I grafici in Fig. 2 mostrano il range delle particelle cariche ed il libero cammino medio
delle particelle neutre nell’acqua. Il libero cammino medio di fotoni e neutroni è grande
(alcuni cm) rispetto alle dimensione delle cellule (alcuni micrometri) o dei siti cellulari come
quelli di cui si interessa la microdosimetria, pertanto, le correlazioni spaziali tra particelle
cariche messe in moto dalla stessa particella primaria non carica possono essere trascurate
nella maggior parte delle considerazioni microdosimetriche.
Per quanto riguarda le particelle cariche, l’interazione di queste con la materia differisce
molto da quella delle particelle prive di carica. Infatti tutte le particelle ad una data energia
vengono bloccate da un fissato spessore di un dato materiale e questo spessore viene appunto
detto range.

12
Fig. 2. (a) Range di elettroni e protoni e (b) cammino libero medio di fotoni e neutroni di data energia
nell’acqua [1].
LET
Il trasferimento lineare di energia ( in inglese Linear Energy Transfer da cui LET), o

13
stopping power di collisione, è definito come dxdEL = , dove dE è l’energia media persa da
una particella carica in collisioni elettroniche lungo un elemento dx della sua traiettoria. Il
valore di L dipende dall’energia della particella, come mostrato in Fig. 3 per protoni ed
elettroni.
Fig. 3. Linear energy transfer (LET) per
elettroni e protoni in acqua in funzione della loro energia [1].
Per caratterizzare la radiazione con un unico parametro, è utile utilizzare un valore medio
del LET. A questo proposito esistono due modi per definire la distribuzione di LET e quindi
esistono due diversi valori medi di LET associati.
La prima è la distribuzione in frequenza di LET, definita in termini della lunghezza totale
di traccia delle particelle cariche o, in modo equivalente, in termini della fluenza di particelle.
La funzione di distribuzione F(L) è la frazione di fluenza associata all’energia lineare
trasferita con un valore non superiore ad L:
φφLLF =)(
dove Φ è la fluenza totale e Lφ è la fluenza di particelle con LET non superiore L.
La densità di LET in lunghezza di traccia o fluenza è indicata da dL
LdFLf )()( = . La traccia
media, o frequenza media, è quindi il valore medio corrispondente alla distribuzione:
[ ]∫ ∫ −== dLLFdLLLfL F )(1)(
La seconda è la distribuzione in dose di LET o distribuzione pesata, definita in termini

14
della dose assorbita prodotta da particelle cariche con un LET specifico. Analogamente a
quanto detto sopra, la funzione di distribuzione D(L) è la frazione di dose assorbita dovuta a
particelle con energia lineare trasferita non superiore a L:
DD
LD L=)(
dove D è la dose assorbita totale, mentre LD è la dose assorbita associata a particelle con
LET non superiore a L.
Per cui la corrispondente densità di LET in dose è indicata con dL
LdDLd )()( = e la dose
media è
[ ]∫ ∫ −== dLLDdLLLdL D )(1)(
Dose assorbita
La dose assorbita può essere definita attraverso la correlata quantità stocastica energia
impartita. L’energia impartita dalla radiazione ionizzante ad un mezzo di massa m in un
volume finito V è definita come:
ε = − + − +∑( ) ( ) ( ) ( )R R R R Qin n out n in c out c
dove (Rin)n e (Rout)n rappresentano l’energia radiante associata rispettivamente alle particelle
neutre entranti e a quelle uscenti dal volume V; (Rin)c e (Rout)c l’energia radiante associata alle
particelle cariche entranti e a quelle uscenti dallo stesso volume, e ∑Q è la somma delle
trasformazioni di energia in massa e di massa in energia che avvengono in V. Si definisce

15
dose assorbita in ogni punto P appartenente a V la grandezza:
dmdD ε=
dove ε è il valore di aspettazione dell’energia impartita nel volume finito V, in un
determinato intervallo di tempo, e dε è la stessa quantità riferita ad un volume infinitesimo
dV, di massa dm, centrato intorno al punto P di interesse. Quindi la dose assorbita D è il
valore di aspettazione dell’energia impartita alla materia per unità di massa in un dato punto.
Le dimensioni e le unità di misura della dose sono uguali a quelle del kerma, e cioè,
rispettivamente, energia per unità di massa e gray (1Gy=1J/Kg).
E’ possibile scrivere una relazione che collega la dose alle grandezze radiometriche solo in
casi particolari, a differenza di quanto avviene per il kerma in cui abbiamo visto che esiste una
relazione fra il coefficiente di trasferimento e la fluenza di energia o di particelle. Questo
dipende dal fatto che, generalmente, non tutta l’energia ricevuta dalle particelle cariche in V
viene ceduta alla materia contenuta in V; una parte di essa può essere ceduta anche fuori da V;
inoltre in V può essere assorbita anche una certa quantità di energia che è stata ceduta alla
particelle cariche all’esterno di V.
1.3 GRANDEZZE STOCASTICHE E LORO DISTRIBUZIONE
Definiamo ora due grandezze normalmente utilizzate in microdosimetria: l’energia specifica
e l’energia lineale attraverso le quali si possono descrivere i depositi di energia in un sito
biologico con dimensioni e geometria prefissate.
Energia specifica
Una volta fissato un sito biologico, attraverso l’energia impartita viene definita una nuova
grandezza: l’energia specifica.
Le particelle ionizzanti, quando viaggiano attraverso un sito S, subiscono delle interazioni
e di conseguenza perdono energia. Se indichiamo con iε l’energia impartita nell’i-esimo urto,
l’energia totale impartita ε è la sommatoria delle iε :
iεε ∑= ( iε in S)
L’energia impartita assume un valore unicamente definito all’interno di una regione una volta

16
che l’esposizione alla radiazione ha avuto luogo. Ripetendo l’esposizione più volte alle stesse
condizioni, a causa della natura stocastica dei rilasci di energia, ogni volta si ottiene un valore
diverso. Le fluttuazioni dei valori assunti sono tanto più grandi quanto più piccolo è il sito,
quanto più piccola è la dose e quanto più densamente ionizzante è la radiazione in questione. I
valori variano secondo determinate distribuzioni di probabilità.
Una quantità legata all’energia impartita è l’energia specifica. Questa è definita come
l’energia impartita divisa per la massa m del sito in questione:
mz /ε=
Data una dose macroscopica D, fissato un sito biologico di massa m, si vede che in quel sito
biologico, z è distribuita secondo la funzione integrale di probabilità :
)();( DzzPDzF ≤=
che rappresenta la probabilità, per una data dose D, che l’energia specifica z non superi un
fissato valore di z.
Fig. 4. Distribuzioni somma F(z;D) di energia specifica in una sfera di tessuto di 6µm di diametro e densità
unitaria, esposto a dosi differenti di raggi γ 60Co e di neutroni a 15 MeV [1].
La densità di probabilità si ottiene differenziando );( DzF :
dzDzdFDzf );();( =

17
questa dà la probabilità di trovare z tra z e z+dz quando è stata assorbita una dose
macroscopica D.
Fig. 5. Densità delle energie specifiche corrispondenti alle distribuzioni somma di Fig. 4 [1].
Il valore medio o valore di aspettazione dell’energia specifica in un sito
∫= dzDzzfz );(
è uguale alla dose assorbita D quando il sito è uniforme ed è esposto ad un campo di
radiazione uniforme. Altrimenti z uguaglia la dose assorbita media nel sito: Dz = .
Attraverso l’energia specifica si può quindi arrivare a definire la dose assorbita:
FmzD
0lim
→=
Energia lineale
Un evento in un sito (traccia) è rappresentato dalla deposizione di energia dovuta a
particelle che sono statisticamente correlate fra loro. In genere i rilasci di energia possono
avvenire attraverso eventi singoli o eventi multipli (ossia in corrispondenza di ν =1 o ν >1,

18
traccia singola o tracce multiple).
La distribuzione di evento singolo, cioè quando una sola traccia è stata registra nel sito,
viene denotata con:
)1()(1 =<= νzzPzF e dz
zdFzf
)()( 11 =
Attraverso l’energia impartita nel caso di eventi singoli viene definita una nuova grandezza
microdosimetrica: l’energia lineale y.
Questa viene definita come il rapporto fra l’energia impartita nel singolo evento e la corda
media l del volume che occupa il sito:
ly /ε=
Anche per questa grandezza si possono definire le distribuzioni di probabilità. Data una dose
macroscopica D e fissato un sito biologico, si vede che y è distribuita secondo una funzione
integrale di probabilità:
)();( DyyPDyF ≤=
da cui si ricava la funzione differenziale:
dyDydF
Dyf);(
);( =
dove );( Dyf dà la probabilità di trovare y fra y e y +dy quando è stata impartita una dose
macroscopica D.
Per quanto riguarda eventi singoli, si può dimostrare che le distribuzioni dell’energia lineale e
dell’energia specifica, sono collegate tra loro:
)()( 1 zFyF = e )()( 1 zflVyf =
Analogamente alle distribuzioni in frequenza di z e y appena discusse, possono essere
definite le distribuzioni pesate in dose integrali D(y) e D(z) e le relative differenziali d(y) e
d(z). Se prima, fissata una dose macroscopica, guardavamo in un dato sito con quali valori di z
e y era stata impartita la dose, ora fissato il sito e la dose macroscopica, guardiamo quanta di
quella dose è stata impartita con un determinato valore di z o y.
Ad esempio, fissata una dose macroscopica D, yd(y) rappresenta la frazione di dose D
impartita in quel sito con un’energia lineale compresa tra y e y+dy; D(y) rappresenta invece la
frazione di dose impartita con energia lineale minore o uguale a y. Considerazioni simili si
possono fare per la variabile z.

19
Fig. 6. Distribuzioni di energia lineale in regioni sferiche di tessuto di 1 µm di diametro esposte a varie
radiazioni. Nel grafico inferiore sono rappresentate le distribuzioni come densità pesate in dose yd(y) rispetto ad una scala logaritmica dell’energia lineale y. Nel grafico superiore sono date le rispettive distribuzioni somma D(y) che specificano la frazione di eventi fino ad una energia lineale y [1].
Fra le distribuzioni in frequenza e quelle in dose esistono le seguenti relazioni:
Fyyyfyd )()( = e
Fzzzfyd )()( 1
1 =
I valori medi delle distribuzioni pesate sono:
dyyydyD )(∫=
dzzzdzD )(∫=

20
2. IL PROCESSO COMPOSTO DI POISSON
2.1 CASUALITA’ DEGLI EVENTI
Quando un sito microscopico è esposto al campo di radiazione gli eventi di deposizione
di energia avvengono in modo casuale e sono statisticamente indipendenti: ogni singolo
evento può essere dovuto all’apparizione di una o più particelle cariche in un sito con
conseguente deposizione di energia. Frequentemente si tratta il caso semplice in cui particelle
cariche indipendenti attraversano il sito in cammini rettilinei.
Durante l’intervallo di tempo che corrisponde al rilascio di una determinata dose
assorbita D, gli eventi si susseguono in modo casuale e, se l’energia specifica media per
evento è Fz , il numero di eventi che ci si aspetta è Fz
D=ν . Il numero di eventi ν è
soggetto a fluttuazioni statistiche e (essendo gli eventi indipendenti) segue la distribuzione di
Poisson. Per cui, conosciuta la frequenza di eventi per unità di dose assorbita Fz1 , con un
processo semplice di Poisson si calcola il numero di eventi.
La valutazione dell’energia impartita è tuttavia più complessa poiché dipende da un
processo composto di Poisson, cioè, un processo di eventi indipendenti ciascuno con intensità
variabile. Formalmente, questo può essere espresso dalla relazione
∑=
=ν
εε1i
i
Gli iε sono le energie impartite in eventi individuali, ν è il numero di eventi che segue la
distribuzione di Poisson.
In Fig. 7 a sinistra sono indicati gli eventi, cioè i passaggi delle particelle cariche, come
segmenti lineari. Nella parte destra uno di questi eventi è stato selezionato e rappresentato
come una successione di collisioni, cioè di perdite di energia da parte di una particella carica.

21
Fig. 7. Grafico schematico che spiega il doppio ruolo del processo composto di Poisson per depositi di
energia in un sito ad una data dose. Nel pannello di sinistra sono rappresentati ν eventi corrispondenti a traiettorie statisticamente indipendenti di particelle cariche. Nel pannello di destra è stato selezionato uno degli eventi e rappresentato come una sequenza di processi di µ collisioni statisticamente indipendenti lungo la traccia della particella [1].
Trattiamo ora il numero di collisioni µ lungo il segmento di traccia, anch’esso soggetto
alla distribuzione di Poisson. Il suo valore di aspettazione µ è proporzionale alla lunghezza
del segmento ed allo stopping power della particella (se il segmento della traccia nel sito è
molto più piccolo del range della particella, la variazione del LET della particella nel sito può
essere trascurata), mentre è inversamente proporzionale all’energia impartita media al sito in
una collisione. Le fluttuazioni di Poisson del numero µ di collisioni sono sempre presenti,
ma sono molto meno importanti rispetto alla variazione di energia persa dalla particella, o
energia impartita al sito, in collisioni individuali.
Infine, le variabili casuali iε (energia impartita in singoli eventi) sono esse stesse
risultato di un processo composto di Poisson
∑=
=i
jji
µ
εε1
per cui, l’energia impartita del sito sarà data dalla formula:
∑ ∑= =
=
ν µ
εε1 1i j
j
i
dove la sommatoria interna riguarda la il processo di Poisson sui segmenti delle tracce
individuali (energy-loss straggling), mentre la sommatoria esterna rappresenta il processo di
Poisson del numero di tracce cariche che attraversano il sito (sequenza casuale di eventi).

22
2.2 EQUAZIONI POISSONIANE
Equazione base
Ad una dose assorbita specifica, l’energia impartita al sito e la relativa variabile energia
specifica sono il risultato di un processo composto di Poisson. Il termine processo di Poisson
si riferisce all’indipendenza degli eventi; il termine composto si riferisce al fatto che il
numero degli eventi individuali è variabile. Lo spettro del processo composto di Poisson è la
distribuzione )(1 zf di singolo evento. Le soluzioni del processo composto di Poisson sono
le distribuzioni in dose );( Dzf .
La seguente trattazione matematica si occupa delle distribuzioni )(1 zf e );( Dzf , può
essere però estesa, in modo identico, al problema di energy-straggling, cioè la perdita casuale
di energia di una particella carica lungo un segmento di traccia.
L’energia specifica media prodotta da un evento singolo è il valore medio di )(1 zf .
Questo valore medio Fz è un parametro fondamentale perché determina la frequenza degli
eventi per unità di dose assorbita. Dato che gli eventi sono per definizione statisticamente
indipendenti, il loro numero ν in un determinato sito ad una determinata dose segue la
distribuzione di Poisson:
!)exp()(
νν
νnnp −= con Fz
Dn ==ν
Anche se il numero ν di eventi è fissato, l’energia specifica in un sito può variare
enormemente. La sua distribuzione è quindi la convoluzione ν -esima della distribuzione di
singolo evento. Questa convoluzione è indicata con )(zfν , e può essere definita dalla
formula ricorrente:
∫ −= −
z
odxxzfxfzf )()()( 11 νν (ν = 2, 3, …)
dzzf )(ν è la probabilità che l’energia specifica abbia un valore tra z e z +dz, se sono
avvenuti esattamente ν eventi nel sito.
Da qui si ricava la relazione per le distribuzioni in dose dell’energia specifica:
∑∞
=
−=0
)(!)exp();(
νν
ν
νzfnnDzf con
FzDn =
)(0 zf è uguale a )(zδ cioè alla funzione delta in z =0. );( Dzf contiene anche una

23
possibilità discreta ne− di evento non avvenuto per z =0.
Fig. 8. Grafici della distribuzione di energia specifica per una data dose assorbita in un sito sferico di 6 µm di diametro, esposti a neutroni di 15 MeV. (a) scala lineare, (b) scala logaritmica, (c) radice quadrata dell’energia specifica in funzione della radice quadrata della dose assorbita [1].

24
Fig. 9. Confronto fra le distribuzioni per siti piccoli (0.5 µm e siti con un diametro confrontabile con quello del nucleo cellulare (6 µm). I risultati sono dati per raggi γ del Co60 e neutroni di 15 MeV [1].
Esempi dei risultati ottenuti con calcoli Monte Carlo sono riportati nelle Fig. 8 e 9. Per
quanto riguarda la Fig. 8, il calcolo simula l’irraggiamento di un sito con un diametro di 6 µm;
il sito viene irraggiato numerose volte con i valori di dose riportati in ascissa, ed il programma
calcola l’energia specifica z che si ottiene in ogni irraggiamento nel sito. In Fig. 9 invece, è
riportato un confronto delle distribuzioni di z tra siti piccoli (0.5 µm) e siti con un diametro
simile a quello del nucleo cellulare (6 µm). Le figure mostrano i plot bidimensionali
dell’energia specifica in funzione della dose. Dato che gli eventi zero non sono rappresentati,
appaiono pochi punti nella parte sinistra dei grafici, dove la probabilità di evento è inferiore
all’unità. Il punto essenziale è visibile chiaramente nel grafico con scala logaritmica: a dose
assorbita sufficientemente bassa, diminuiscono le frequenze degli eventi ma non i valori
dell’energia specifica.

25
Relazioni per i momenti
Solitamente risultano molto importanti e ricchi di significato non tanto le distribuzioni
esplicite in dose dell’energia specifica quanto alcuni parametri di queste distribuzioni che
possono essere espressi per mezzo dei momenti della distribuzione di evento singolo. Il
secondo momento e la varianza di z giocano il ruolo più importante in molte applicazioni e
sono quindi qui derivate.
Il valore di aspettazione dell’energia specifica risulta uguale alla dose assorbita:
Dznz == 1
La varianza )(2 Dzσ , dell’energia specifica z per una dose assorbita D, si può facilmente
calcolare utilizzando il fatto che la varianza della somma di sue variabili casuali indipendenti
è uguale alla somma delle loro varianze. Segue che la varianza di z a dose 21 DD + è uguale
alla somma della varianze a dose 1D e 2D .
La varianza deve essere proporzionale alla dose assorbita:
cDz =2σ
La varianza di una variabile casuale è data da:
222222 )( Dzzzzzz −=−=−=σ
Il secondo momento può essere espresso in termini della distribuzione in dose dipendente
dell’energia specifica:
∑∫∑∞
=
−∞∞
=
− ==1
2
0
2
0
2
!)(
! νν
ν
νν
ν
ννznedzzfznez nn
dove 2νz è il secondo momento della distribuzione )(zfν di ν -eventi. In contrasto con
2z , i valori di 2νz non dipendono dalla dose.
Lo sviluppo dell’equazione precedente è:
( ) =+−+=
++⋅
−+−= ......
21...
211 22
122
21
22
221
22 nzzznznznnnz
( )...2
21
21
22
1
21 +
−+
= D
z
zzDzz
Per cui si ha:
( )...2
1
21
22
1
21
2
+
−+== D
z
zzzz
Dc zσ

26
Dato che c è una costante, si può ottenere il suo valore dal limite 0→D :
1
21
0lim
zzcc
D==
→
Quindi si hanno le relazioni essenziali:
Dzz
z
=
1
212σ e 2
1
212 DD
zzz +
=
2.3 IMPLICAZIONI BIOFISICHE DELLA MICRODOSIMETRIA
La rappresentazione grafica delle frazione di cellule sopravvissute in funzione della dose
di radiazioni ricevuta è detta curva di sopravvivenza (Fig 10). Per spiegare il loro andamento,
sono state formulate diverse teorie. Qui di seguito vengono presentati alcuni modelli
matematici, che si basano su concetti di microdosimetria impiegati per l’analisi dei dati delle
curve di sopravvivenza e per la descrizione di relazioni dose assorbita – effetti biologici: il
modello a più urti, il modello a bersagli multipli e il modello a soglia.
Fig. 10. Curve di sopravvivenza per cellule di mammifero irraggiate con radiazioni a basso e alto LET.
0D ad alto LET rappresenta il reciproco della pendenza della curva; a basso LET rappresenta il reciproco della pendenza nella parte finale della curva, 1D è il reciproco della pendenza della parte iniziale della curva [2].

27
Modello a più urti e modello a bersagli multipli
Il modello a più urti prevede che la cellula contenga un ipotetico bersaglio critico. Il
campione può cioè tollerare (n -1) hits o collisioni e, se avvengono n o più collisioni la cellula
si disattiva. Quando il numero di urti nel bersaglio è al di sotto di quelli necessari per ottenere
l’effetto letale il danno prodotto viene detto subletale. La probabilità di sopravvivenza della
cellula in funzione della dose può essere calcolata con la statistica di Poisson :
∑−
=
−=1
0 !)()(
nD DeDS
ν
να
να
Per n > 1 questa equazione produce curve dose - effetto con una spalla nella zona iniziale
dove S(D) decresce con la dose con un andamento poco meno che esponenziale. La pendenza
della curva cresce all’aumentare del numero n di collisioni. Per adattare l’equazione con
l’andamento sperimentale della parte iniziale della curva di sopravvivenza cellulare, bisogna
moltiplicare la probabilità di sopravvivenza per un fattore addizionale De γ− , legato alla
pendenza della curva di sopravvivenza, espressione grafica del tasso di spopolamento
cellulare. E’ interpretata come una disattivazione indotta a collisione singola da parte della
componente densamente ionizzante della radiazione:
∑−
=
+−=1
0
)(
!)()(
nD DeDS
ν
νγα
να
Il modello a bersagli multipli postula la presenza di m ipotetici bersagli uguali, ciascuno
dei quali può essere disattivato da una collisione singola. In questo caso, il danno subletale si
ha quando non tutti i bersagli sono colpiti. Viene assunta uguale probabilità di collisione e
l’indipendenza statistica delle collisioni su campioni individuali. L’equazione risultante per la
sopravvivenza delle cellule è
( )mDeDS α−−−= 11)(
Ancora viene introdotto un termine esponenziale per tener conto della pendenza iniziale
(spalla) presente nella maggior parte delle curve di sopravvivenza:
( )[ ]mDD eeDS αγ −− −−= 11)(
Anche in questo caso, all’aumentare dell’esponente m si ha un allargamento della spalla, il
che può significare due cose strettamente legate tra loro: o una maggiore difficoltà
nell’accumulo del danno subletale o una maggiore efficienza nei processi di recupero.
Purtroppo queste due teorie sono crude approssimazioni e quindi possono avere solo un

28
valore euristico. Un trattamento stocastico più generale non presenterebbe problemi nella
trattazione matematica, ma ogni analisi realistica richiede un’adeguata considerazione dei
modelli microscopici della deposizione di energia per le diverse radiazioni. Queste
caratteristiche microscopiche determinano le forti differenze degli effetti biologici delle
radiazioni ionizzanti ed è evidente che un’analisi biofisica senza dati microdosimetrici può
avere poco valore. E’ quindi interessante analizzare l’idea di base della teoria del bersaglio
(target theory) per mezzo della microdosimetria.
Modello a soglia
Una relazione dose-effetto è l’espressione di vari processi stocastici. La deposizione di
energia nelle cellule varia in modo casuale, ma anche la risposta della cellula stessa può
essere descritta in termini probabilistici. I modelli a più urti ed a bersagli multipli trascurano
appunto la risposta stocastica della cellula e la relazione dose-effetto è intesa solo in termini
della statistica dei depositi di energia. Questa è ovviamente un’interpretazione incompleta e
poco realistica; anche se questa stessa obiezione può essere mossa anche al modello a soglia
formulato in termini microdosimetrici, in quest’ultimo caso è possibile effettuare un
trattamento molto più realistico.
Si consideri un dosimetro microscopico in grado di registrare l’energia specifica in una
regione sferica di tessuto di diametro d e che risponda solo quando viene raggiunto un valore
critico Cz . La probabilità di non avere risposta è:
∫ == Cz
C DzFdzDzfDS0
);();()(
In Fig. 11 è rappresentata questa funzione nel caso di irraggiamento con raggi X per un sito
sferico di diametro 1 µm e per diversi valori critici Cz . Nonostante sia stata stato richiesto il
superamento di una soglia ben precisa, la dipendenza dalla dose risultante non è una funzione
a gradini. Se Cz è simile o più piccolo del valore Cz dell’evento medio associato alla
radiazione usata, la dipendenza è quasi esponenziale e si è in presenza di una reazione a
singola collisione. Per valori più grandi di Cz , la relazione dose-risposta, ha una spalla come
risultato di un accumulo di danni.

29
Fig. 11. Sono qui rappresentate le probabilità 1-F(z;D) di non superare un valore critico z dell’energia
specifica in funzione della dose assorbita in un sito sferico di tessuto con diametro di 1 µm esposto ai raggi X [1].
In generale, per ogni relazione dose-effetto S(D) osservata si possono determinare i valori
Cz e d tali che facciano aderire il più possibile la funzione F(z;D) a S(D). Queste due
funzioni possono essere viste come distribuzioni somma della dose richiesta per produrre
l’effetto.
Il diametro d qui trovato non è la vera dimensione dei bersagli critici né del volume
sensibile nella cellula, ma un valore più basso. Infatti il volume sensibile può essere uno solo
più grande, oppure una molteplicità di piccoli bersagli dispersi in un volume maggiore.
Inoltre, solo una parte della deviazione osservata rispetto alla funzione a gradino è dovuta alla
deposizione stocastica di energia nel sito, l’altra parte invece può riflettere la risposta
stocastica della cellula, anche ad energia specifica fissata.
Nel caso di studi di disattivazione cellulare, questo risultato è in accordo con la teoria che
il DNA, disperso nell’intero nucleo, sia il bersaglio dell’azione delle radiazioni.

30
3. APPLICAZIONE AI SISTEMI BIOLOGICI
3.1 MICRODOSIMETRIA NEI SISTEMI BIOLOGICI
Si sa che le radiazioni ionizzanti inducono una grande varietà di effetti biologici come
morte cellulare, mutazioni, aberrazioni cromosomiche e carcinogenesi. La maggioranza degli
effetti direttamente rilevanti nell’uomo sono dovuti a danni a singole cellule. E’ quindi chiaro
come i caratteri microscopici dell’interazione e della deposizione di energia da parte delle
radiazioni sia cruciale per comprendere interamente i meccanismi con cui vengono indotti
questi effetti. Le descrizioni fisiche di queste caratteristiche costituiscono il campo della
microdosimetria nel suo significato più ampio.
Ci sono molte applicazioni biologiche in cui le tecniche di dosimetria (macroscopica)
convenzionale sono in grado di dare un’adeguata descrizione fisica dell’interazione tra il
campo di radiazioni ed il materiale biologico, come ad esempio nella radioterapia con fotoni.
Tuttavia, anche nel campo della radioterapia, fluttuazioni a livello cellulare o sub-cellulare
possono essere di cruciale importanza, specialmente quando la deposizione di energia
localizzata è concentrata come nel caso di particelle densamente ionizzanti (ad alto LET).
Quando per il trattamento vengono usati fasci di radiazioni miste che includono fotoni,
neutroni e particelle pesanti la dosimetria convenzionale risulta inadeguata per calcolare la
probabilità di danno.
Ogni tentativo di confrontare diversi tipi di radiazione, inevitabilmente porta all’utilizzo
di descrizioni microdosimetriche dell’interazione radiazioni-materia per poter predire gli
effetti biologici. Campi di radiazione che possono sembrare uniformi a livello di tessuto o di
cellula (qualche decina di micrometri), risultano invece essere completamente disomogenei
scendendo a livello subcellulare (manometri).

31
Fig. 12. Rappresentazione schematica delle caratteristiche microscopiche delle tracce di radiazioni
corrispondenti a 1 cGy di dose assorbita di tre diverse radiazioni [3].
Conseguentemente il concetto di dose perde significato, in quanto, passando da un volume
elementare all’altro, ad esempio in siti con dimensioni lineari di 2 nm, si vede che la dose
varia di ben sei ordini di grandezza.
Una risposta a questi problemi può essere data dalla microdosimetria. In Fig. 13 sono
illustrate graficamente alcune delle descrizioni in termini microdosimetrici applicate ai
sistemi biologici e che verranno brevemente descritte nei paragrafi successivi.

32
Fig. 13. Descrizioni microscopiche alternative della qualità della radiazione. Il grafico rappresenta le
interazioni atomiche individuali lungo un segmento di 120 nm del cammino di una particella α di 8 MeV e le varie descrizioni microdosimetriche che vi si possono utilizzaree. Per confronto è presente un’elica di DNA (2 nm) nella stessa scala [3].
3.2 LINEAR ENERGY TRANSFER (LET)
Definizione e distribuzione
Il LET è stata la prima grandezza introdotta per spiegare la diversa efficacia biologica
delle radiazioni; tuttavia essa è una grandezza media che non tiene conto che il rilascio di
energia è discreto ed è variabile lungo la singola traccia divenendo più intenso verso la fine
della traccia. Ciononostante ci sono varie situazioni in cui si è rivelato utile per descrivere le
proprietà medie delle varie radiazioni.
A fianco al LET viene introdotto il LET ristretto o restricted linear collision stopping
power ( ∆L ); esso rappresenta in un mezzo il rapporto tra dE e dl, dove dl è la distanza
attraversata dalla particella e dE è la sua energia media persa in seguito a collisioni con
trasferimenti di energia inferiori ad un dato valore ∆:
∆∆
=
dldEL
Il LET infinito ( ∞L ) è quello che si ottiene quando vengono considerati tutti i possibili
trasferimenti di energia senza porre nessun valore di soglia; esso corrisponde allo stopping

33
power di collisione.
Quando una radiazione rilascia in un tessuto una dose macroscopica, si può vedere che
questa viene rilasciata con diversi valori di LET a causa della variazione di energia subita
dalle particelle cariche man mano che rallentano nel tessuto.
Si può definire una distribuzione della dose in LET e cercare di sfruttare l’andamento che
assume questa distribuzione per interpretare gli effetti biologici; una distribuzione
normalizzata della dose in funzione del LET è data da:
∫∞
⋅=0
)(
)()(dLLt
LtLLd
essa rappresenta la frazione di dose assorbita con determinati valori di LET; t(L) rappresenta
la frequenza con cui la dose viene rilasciata da una particella carica con LET tra L e L +dL (o
la frazione di lunghezza di traccia con LET tra L e L +dL). In Fig. 14 sono riportati degli
esempi di distribuzioni cumulative.

34
Fig. 14. Distribuzioni cumulative di dose assorbita in funzione del LET in acqua per diverse radiazioni.
(a)Distribuzioni di LET non ristretto per lo spettro di particelle cariche da raggi γ del 60Co, neutroni monoenergetici di 14.6 e 1.8 MeV e per brevi tratti di tracce di ioni pesanti monoenergetici. (b) Distribuzioni di LET ristretto, con un’energia di soglia per i raggi δ di ∆=100 eV, per lo spettro di rallentamento di particelle cariche da raggi γ del 60Co, per tracce di particelle α da 5.3 MeV, e per brevi tratti di tracce di alcuni ioni pesanti con energia 10 o 100 MeV/amu [3].
Si vede che la maggioranza delle radiazioni rilascia la dose attraverso particelle cariche con
diversi valori di LET molto diverso e la cui dispersione è tanto più grande quanto più le tracce
sono descritte a livello microscopico.
Il limite principale di cui soffre il LET è legato al fatto che esso viene calcolato con
l’ipotesi del rallentamento continuo, ossia senza tener conto che i rilasci lungo la traccia della

35
particella sono discreti. Il LET, quindi, finisce per essere una grandezza media che spesso non
è in grado di descrivere adeguatamente l’effetto biologico. L’adeguatezza o meno del LET per
la descrizione microscopica in una data applicazione dipende dalle dimensioni lineari in
gioco, per esempio la lunghezza della traccia rispetto alle dimensioni lineari della struttura
biologica esaminata.
Efficacia biologica relativa (RBE)
Un’altra grandezza introdotta per tener conto della diversa capacità di produrre effetti
biologici, a parità di dose, dai vari tipi di radiazione è la Relative Biological Effectiveness
(Efficacia Biologica Relativa RBE). L’RBE di una radiazione, per un dato tipo e severità di
effetto biologico, è il rapporto della dose assorbita di una radiazione di riferimento, che
produce quell’effetto, e la dose assorbita della radiazione test che produce un effetto identico.
Come radiazione di riferimento di solito si usano fotoni a basso LET (raggi X da 250 kV
oppure i raggi γ del 60Co). In genere la RBE varia sia in base al tipo e alla severità dell’effetto
biologico, sia in base al tipo della radiazione.
Il LET è stato spesso usato come parametro di qualità della radiazione per spiegare i
diversi valori di RBE; ma purtroppo si è visto che l’RBE non è una funzione univoca del
LET, anche per un dato tipo e severità di effetto biologico. Tuttavia, pur con i suoi limiti, il
LET viene usato ancora soprattutto in quei casi in cui non sono stati individuati altri parametri
in grado di descrivere adeguatamente i vari fenomeni.
Ci sono numerosi studi che riportano l’RBE in funzione del LET, ma solo un numero
limitato di questi contengono dati sistematici per particelle monoenergetiche di LET ben
definito. Queste ultime devono necessariamente essere ristrette a sistemi in vitro di cellule di
mammifero o altri organismi. Le irradiazioni esterne di sistemi in vivo sono invece
irradiazioni costituite sia da fotoni che da neutroni o ioni pesanti energia ma la dose in questi
casi, come detto prima, viene rilasciata con diversi valori di LET.
Un esempio di grafico RBE-LET è mostrato nella figura sottostante per cellule di
mammifero.

36
Fig. 15. Esempio di dipendenza dell’RBE dal LET non ristretto, in questo caso, per la sopravvivenza del
50% di una popolazione di cellule di criceto V79 irradiate. Altre caratteristiche sono: (i) aumento dell’RBE con la diminuzione della dose (frecce tratteggiate) e (ii) aumento (ad alto LET) o abbassamento (a LET intermedio) dell’RBE con la velocità degli ioni ad un dato LET [3].
E’ utile ricordare che l’RBE per un dato ∞L o ∆L assume diversi valori con variazioni
dipendenti da fattori come la velocità della particella carica, tipo e magnitudine dell’effetto
biologico, tipo di cellula e stato di crescita.
3.3 CONTATORI PROPORZIONALI
La scoperta di larghe fluttuazioni statistiche sui depositi di energia di radiazione su
distanze di micrometri ha portato ad un bisogno di migliorare le descrizioni fisiche a livello
microscopico. Le deposizioni di energia possono essere misurate direttamente in rivelatori che
simulano le dimensioni cellulari o sub cellulari fino a circa 1 µm. Questi studi culminarono
con la microdosimetria dei contatori proporzionali Rossi, sviluppata da Rossi e Kellerer.
Nella forma base, il contatore di Rossi consiste in una camera (tipicamente una sfera di 2
cm di diametro circa) tessuto equivalente (TE) contente due elettrodi, riempita con un gas a
pressione sufficientemente bassa tale che la lunghezza di traccia di una particella che lo
attraversa, sia la stessa che la particella avrebbe in una piccola sfera di tessuto. Quando la
radiazione interagisce con il gas per ionizzare le sue molecole o atomi, le cariche primarie

37
vengono accelerate dal campo elettrico presente tra gli elettrodi, causando ionizzazioni
secondarie delle molecole del gas, che risultano proporzionali alla quantità di ionizzazioni
primarie. Questo fenomeno è detto valanga elettronica o moltiplicazione del gas. Quindi, si
possono attuare misurazioni dirette dell’energia depositata all’interno del piccolo volume di
tessuto simulato.
Per ottenere una simulazione quantitativa di deposizione di energia in piccoli volumi di
tessuto, la perdita di energia di una particella carica passante attraverso il gas deve essere
identica a quella corrispondente ad una traiettoria nel volume di tessuto:
tttggg pSpS ρρ =
dove S è lo stopping power di massa, ρ la densità e p la lunghezza di percorso con gli indici
che denotano gas (g) o tessuto (t). Quindi
Kpp
SS
g
t
g
t
t
g 1=
⋅
=
ρρ
Se gli stopping power di massa sono identici allora la densità del gas richiesta per la
simulazione è più bassa di quella del tessuto secondo lo stesso fattore con cui scalano le
distanze. In queste condizioni si trova che la massa del gas nel contatore è molto K2 volte
quella del volume di tessuto simulato; quindi, per una data dose assorbita, il numero di
interazioni nel rivelatore sarà K2 volte più grande di quelle che si verificano nel sito; ma allo
stesso tempo la deposizione di energia delle singole interazioni sarà simulata correttamente
perché la perdita di energia lungo traiettorie corrispondenti è la stessa.
I contatori proporzionali a bassa pressione possono essere usati, come scritto sopra, per
misurare eventi di deposizioni di energia, in piccoli volumi di tessuto simulato, sull’assunto
che l’energia totale depositata da un evento nel contatore sia direttamente proporzionale al
numero di ionizzazioni prodotte nel gas del contatore, e che questo, a sua volta, sia
proporzionale al all’altezza dell’impulso misurato dopo la moltiplicazione del gas. In questo
modo, gli eventi di deposizione in energia nei contatori proporzionali, danno una descrizione
microscopica diretta di quello che avviene nel tessuto.
La maggior limitazione di questa descrizione è dovuta ad un limite inferiore di circa 0.3
µm sulle dimensioni che possono essere accuratamente simulate.
Un'altra limitazione è il cosiddetto “effetto pareti” creato dalle pareti solide del contatore
proporzionale, anche quando le pareti ed il gas hanno la stessa composizione elementare. La
densità più alta delle pareti può causare un aumento del deposito di energia nel gas. Per
esempio, un raggio δ , prodotto nella parete appena prima che una particella primaria entri nel

38
gas, può anch’esso entrare nel gas, nonostante questo non possa accadere nel volume
corrispondente del mezzo a densità uniforme lungo il percorso della particella primaria. Per
evitare questi inconvenienti, sono stati creati dei contatori wall-less, in cui per definire i
contorni del volume di gas si usano o una griglia fine o un campo elettrico creato da piccoli
elettrodi.
Accurate simulazioni di microscopici volumi di tessuto biologico, richiedono che il gas
del contatore proporzionale e le pareti siano sufficientemente tessuto-equivalenti. L’approccio
convenzionale è quello di scegliere materiali con una composizione elementare simile a quella
del tessuto rappresentativo quale grasso, osso o muscolo. Una soluzione approssimativa tipica
è per esempio quella di costruire le componenti solide con plastica A-150 (considerata
equivalente al tessuto muscolare) ed una miscela di gas come propano (o metano), ossigeno e
azoto.
Quando un contatore proporzionale di Rossi è esposto ad un campo di radiazione di
intensità bassa, ogni interazione della radiazione con il gas produce un impulso di altezza
proporzionale all’energia depositata nel contatore. Per descrivere le corrispondenti
deposizioni di energia in volumi di tessuti simulati a densità unitaria, sono state introdotte
appropriate quantità, ovvero l’energia lineale e l’energia specifica, già viste nel primo
capitolo.
3.4 PROFILI DI TRACCIA ED ENTITA’ DI TRACCIA
Una limitazione aggiuntiva del concetto di LET è che esso ignora la larghezza finita delle
tracce di radiazione, difatti il LET non ristretto ∞L la ignora totalmente mentre il LET
ristretto ∆L considera la componente principale delle deposizioni di energia localizzate
lungo il percorso delle particelle primarie, ma blocca i depositi di energia degli elettroni
secondari con energia >∆.
Esiste tuttavia una descrizione microscopica alternativa attraverso le distribuzioni radiali
dei depositi di energia attorno alle tracce delle particelle.
La larghezza massima radiale della traccia di una particella carica è determinata dai range
degli elettroni δ secondari ad energia più alta che essa produce. Questa larghezza massima
cresce all’aumentare della velocità della particella. Una semplice descrizione della traccia è
quella di considerarla come due volumi cilindrici coassiali centrati sul percorso della
particella. Il cilindro più interno, o core di traccia, è definito come la regione che include tutte

39
le energie trasferite dai raggi δ aventi energie al di sotto di una soglia arbitraria.
Il cilindro più esterno, o penombra, ha raggio uguale al range dei raggi δ a massima
energia. Nell’approssimazione più semplice le deposizioni di energia nella penombra sono
considerate uniformi.
Le entità di traccia forniscono una descrizione geometrica delle distribuzioni di energia.
Gli spurs (impulsi), da lungo tempo usati in chimica delle radiazioni di soluzioni diluite, sono
usati per descrivere piccole regioni sferiche in cui una singola perdita di energia da parte di
una particella carica produce poche ionizzazioni e/o eccitazioni.
In generale, sono state considerate le fluttuazioni nelle caratteristiche dei depositi di
energia, da parte delle tracce delle particelle, andando a suddividere ciascuna traccia in un
numero di entità discrete come segue:
- una perdita di energia <100 eV produce uno spur;
- una perdita di energia compresa tra 100 e 500 eV forma un blob (macchia). Questa è
una regione locale ad alta densità di ionizzazione;
- una perdita di energia, dovuta ad una collisione, tra i 500 eV ed i 5 keV forma una
short track (traccia breve). Il range di questo elettrone è sufficientemente corto in
modo che i suoi spurs costituenti si sovrappongano e creano una regione locale con
alta densità di ionizzazione.
Spesso i concetti di entità di traccia degli elettroni sono stati combinati con quelli di core
e penombra per descrivere tracce di particelle pesanti. L’evoluzione temporale del core è
diversa per particelle a LET basso, medio o alto, a causa dei diversi gradi di separazione o
sovrapposizione degli spurs costituenti.
3.5 SIMULAZIONE DELLA STRUTTURA DELLE TRACCE
Metodo Monte Carlo
I codici Monte Carlo per la simulazione delle tracce di radiazione, oggi forniscono la
descrizione più dettagliata per i caratteri microscopici delle interazioni di radiazione e della
deposizione di energia, fino a distanze di circa 1 nm, tanto che è difficile prevedere tecniche
future in grado di dare una descrizione ancora più particolareggiata. Nuovi sviluppi si
potranno avere da studi teorici e sperimentali per migliorare l’accuratezza e la rilevanza
biologica dei dati fisici da inserire nel codice Monte Carlo, e da studi biofisici per
determinare quali proprietà delle tracce sono le più rilevanti per gli effetti biologici.

40
Fig. 16. Segmento di traccia simulato
con Monte Carlo di una particella α confrontato con un cilindro di diametro 2 nm [3].
Fare una descrizione completa e precisa delle interazioni che avvengono durante una data
irradiazione non è possibile a causa della natura probabilistica di ogni interazione. Ogni
traccia, anche se di particelle identiche, è differente da tutte le altre.
Metodi Monte Carlo sono stati usati per produrre storie di tracce in mezzi omogenei
simulate a computer. I dati in input consistono nelle varie sezioni d’urto di interazione sia in
forma tabulare che analitica; un generatore di numeri casuali poi, seleziona tra queste sezioni
d’urto per determinare tipo, posizione ed effetti di ogni interazione in sequenza. In questo
modo, le particelle primarie e secondarie sono seguite nelle successive interazioni fino a
quando raggiungono un’energia trascurabilmente bassa. Piena informazione su ogni
interazione può essere immagazzinata per essere successivamente visualizzata ed analizzata.
La quantità di informazioni in gioco in questo tipo di calcolo risulta essere enorme, per cui,
per ragioni pratiche di tempo e di memoria del computer, lo schema delle interazioni e le
informazioni richieste in uscita sono spesso semplificati. Ad esempio, un segmento di traccia
lungo 1 µm di una particella α da 4 MeV in acqua, è costituito da circa 104 punti di interazioni
anelastiche per la particella primaria e per gli elettroni secondari quando sono seguiti fino ad
un’energia residua di 10 eV. Ogni punto è comunemente descritto con un minimo di otto
caratteristiche per poter specificare la sua posizione, identità della particella incidente, natura
dell’interazione, energia trasferita e direzione delle particelle secondarie.
Tracce simulate individualmente, di solito, non possono essere applicate direttamente a
problemi pratici. Essi contengono un tale assortimento di dati che si possono associare ad una
traccia reale solo statisticamente. Il numero di tracce simulate richieste come campione

41
statisticamente rappresentativo dipende dalla particolare applicazione. Ad esempio, per
ottenere una stima accurata di una proprietà media generale quale può essere il ∞L , è
richiesta una traccia con lunghezza di qualche micrometro anche per particelle con piccolo
straggling come nel caso di particelle α da 4 MeV. Solo questo implica ~104-105 punti di
interazione, ciascuno dei quali con otto parametri caratterizzanti. Altre proprietà, come la
concentrazione media di energia ad una certa distanza dal cammino primario, possono
richiedere un campione molto maggiore.
Un problema di primaria importanza nell’applicazione (a sistemi biologici) di tracce
simulate col Monte Carlo, è la scelta di una forma appropriata di riduzione dei dati; ogni
scelta particolare, infatti, limita molto la generale applicabilità dei risultati. E’ importante
perciò conoscere quali sono le proprietà di una traccia che più delle altre influiscono sugli
effetti biologici e la loro determinazione; di questo si occupano molti studi di radiobiologia.
Distribuzione in frequenza del deposito di energia
Studiare i depositi di energia su distanze molto piccole di circa < 10 nm è apparentemente
di grande importanza nel determinare gli effetti biologici delle radiazioni. Di conseguenza,
tramite Monte Carlo, sono state studiate le distribuzioni in frequenza di questi depositi di
energia per target con forma arbitraria. Inizialmente ci si è concentrati su campioni sferici,
per poi passare a volumi cilindrici, toroidali e addirittura con strutture simili al DNA o alla
cromatina.

42
Fig. 17. Distribuzioni di frequenza assoluta di eventi di deposizione di energia, da parte di diverse
radiazioni in acqua, in volumi cilindrici posizionati in modo casuale corrispondenti alle dimensioni della struttura del DNA. L’asse a destra mostra le frequenze rispettive a questi eventi nel DNA di cellule tipiche di mammifero [3].
Sono stati quindi fatti confronti diretti tra gli effetti biologici osservati di vari tipi di
radiazioni con le frequenze assolute di deposizione di energia calcolate. Si è potuto
concludere che:
- Modelli semplici dell’effetto della radiazione, basati sulla teoria di singola collisione,
non sono in grado di spiegare gli effetti sulle cellule di mammifero;

43
- i primi tentativi di calcoli di deposizione di energia in campioni piccoli effettuati nel
passato, basati su eventi di perdita di energia, sono troppo poco accurati per un’analisi
quantitativa dei dati radiobiologici;
- la frequenza di depositi di energia di circa 100 eV o superiore concentrati all’interno
di una sfera di circa 3 nm di diametro, è ben correlata con quella dei relativi effetti
biologici prodotti da fotoni; questo implica che ionizzazioni singole ed isolate
abbiano una rilevanza piccola o nulla su cellule di mammifero; inoltre è stato visto
che la suddetta correlazione viene persa se si cambiano questi parametri (100 eV, 3
nm);
- le suddette proprietà, però, non sono in grado di spiegare l’RBE delle particelle α
lente; sembra che siano necessari cambiamenti in entrambi i parametri. Questa
mancanza di accordo è dimostrata in vari esperimenti in cui una larga porzione del
danno immediato prodotto da radiazioni ad alto LET risulta qualitativamente
differente da quello prodotto da irradiazione con fotoni o elettroni. Gli effetti prodotti
da radiazioni ad alto LET sembrano essere correlati con rilasci di energia dell’ordine
di 340 eV in siti di 5-10 nm (nucleo somi);
- è stato stimato (usando particelle α o I125 ) che la mancanza di integrità in una
piccola parte di DNA (dell’ordine di ~1-4%) può risultare critica per la sopravvivenza
di una cellula di mammifero; in questo caso la regione bersaglio risulta avere
dimensioni dell’ordine di ~105 kb di DNA.

44
4. MICRODOSIMETRIA E BNCT
4.1 MICRODOSIMETRIA DELLA REAZIONE B10
Un’ideale terapia antitumorale è tale da distruggere selettivamente tutte le cellule
malate salvaguardando i tessuti sani.
Le cure oncologiche tradizionali sostanzialmente sono tre: l’esportazione chirurgica, la
chemioterapia e la radioterapia. Questi tipi di trattamenti di solito si integrano tra loro per
perseguire l’obiettivo finale di guarire il paziente e migliorarne le condizioni di vita. Una
continua ricerca sperimentale è tuttavia necessaria al fine di trovare nuove terapie ancora più
efficienti, che permettano di curare i tumori che mostrano una spiccata resistenza a quelle
tradizionali.
Una di queste terapie potrebbe essere la Boron Neutron Capture Therapy (BNCT), una
tecnica che sfrutta la combinazione di una sorgente di neutroni, di opportuna energia, e di un
composto biocompatibile arricchito in B10 , isotopo stabile del boro naturale. La BNCT fa
parte delle terapie mirate in cui l’agente viene inoculato nei tessuti cancerosi attraverso un
composto trasportatore e qui attivato ad esempio dalla luce o, come in questo caso, da
neutroni. Sebbene ancora in fase di sperimentazione, la boroterapia è una delle tecniche
oncologiche più promettenti per il trattamento di alcune forme di tumori radio resistenti..
Essa è attualmente orientata anche verso il trattamento di neoplasie collocate nel cervello, nel
capo-collo, nei polmoni, nel fegato o nella pelle.
La BNCT cerca di superare le limitazioni della radioterapia tradizionale riguardo alla
selettività (ovvero la capacità della terapia di colpire solo o prevalentemente i tessuti tumorali
e non quelli sani), somministrando al paziente un composto che concentra il B10 soprattutto
nei tessuti cancerosi. Se quest’ultimo viene bombardato con neutroni termici (En < 0.5 eV)
ha luogo una reazione nucleare che produce una particella alfa e uno ione litio, i quali
rilasciano la maggior parte della loro energia all’interno della cellula contente il B10 . Di
conseguenza se il rapporto della concentrazione di boro tra i tessuti malati e i tessuti sani è
dell’ordine di qualche unità una dose maggiore sarà rilasciata, durante l’irradiazione
neutronica, alle cellule tumorali rispetto al resto.

45
La Fig. 18 illustra il processo fisico alla base della boroterapia.
Fig. 18. Reazione nucleare caratterizzante la BNCT, tratta da [4].
Quando un neutrone termico viene catturato dal B10 si ha la formazione di un nucleo di
B11 che decade secondo due canali di reazione (Q=2.79 MeV):
B10 +1n→ B11 **→ Li7 +α + γ 94%
B10 +1n→ B11 **→ Li7 +α 6%
Nel 94 % dei casi in cui la reazione ha luogo il litio si trova in uno stato eccitato e la sua
successiva diseccitazione dà luogo ad un fotone da 0.48 MeV. Nel 6 % dei casi invece il litio
viene formato nel suo stato fondamentale.
Se il fascio di neutroni incidente sul bersaglio è epitermico (0.5 eV < En < 10 keV), i
neutroni, man mano che penetrano in profondità nel bersaglio, rallentano e quando
raggiungono l’energia propria dei neutroni termici danno luogo alle reazioni sopra descritte.
Le particelle cariche emesse nella reazione hanno traiettorie sostanzialmente rettilinee e
un range ( 9 µm per le α, 5 µm per il litio) confrontabile con il diametro medio cellulare (10
µm, circa): ne consegue che il rilascio energetico avviene quasi totalmente nella cellula
contenente il boro. Queste particelle inoltre hanno un LET medio di circa 200 keV/µm, perciò
la loro efficacia non è influenzata dalla presenza di ossigeno come invece accade per altri tipi
di radiazioni ionizzanti come i fotoni; risultano dunque più efficaci contro i tessuti tumorali,

46
solitamente meno irrorati rispetto a quelli sani e dunque più resistenti al trattamento con
fotoni o elettroni.
Nell’ambito della BNCT la dosimetria dipende dal tipo di sorgente di neutroni usata ed
è complessa per diversi motivi: si tratta di fasci misti che contengono cioè sia neutroni, che
fotoni, che particelle alfa e ioni litio (particelle ad alto e basso LET. Inoltre bisogna
considerare che penetrando nei tessuti biologici le diverse componenti del fascio
interagiscono in modo differente (la parte termica del fascio neutronico è assorbita, quella
veloce viene moderata…) e quindi il campo di radiazione e lo spettro energetico dei neutroni
variano con la profondità.
La qualità biologica delle particelle cariche secondarie conseguenti le interazioni
neutroniche con i tessuti è diversa, così come è diversa la distribuzione spaziale dei depositi
energetici. I principali contributi alla dose totale assorbita dai tessuti irraggiati sono dovuti ai
raggi gamma (dose gamma), alle interazioni neutroniche con gli elementi costituenti i tessuti
biologici (dose neutronica) e alle particelle cariche emesse nella reazione di cattura
B10 (n,α) Li7 (dose BNCT). Riepilogando abbiamo:
- Dose gamma: un campo neutronico è sempre accompagnato da raggi gamma a cui si
aggiungono i fotoni emessi dalle reazioni di cattura neutronica da parte dei materiali
costituenti la struttura d’irraggiamento (il reattore o l’acceleratore).
- Dose neutronica: i neutroni, attraversando la materia biologica, non producono
ionizzazione direttamente ma le loro interazioni hanno luogo attraverso meccanismi
di diverso tipo. La dose depositata da neutroni epitermici e veloci (En > 10 keV) è
principalmente dovuta ai nuclei di rinculo conseguenti le interazioni di scattering. I
neutroni termici si propagano nella materia finchè non sono catturati da un nucleo che
generalmente emette un fotone.
- Dose BNCT: è la dose dovuta agli eventi di deposito energetico delle particelle
conseguenti la reazione B10 (n,α) Li7 di cui si è parlato in precedenza.
L’analisi del campo di radiazione caratteristico della BNCT può essere fatta attraverso
codici di calcoli Monte Carlo, che simulano le storie individuali di centinaia di neutroni e
delle loro tracce all’interno del bersaglio. Questi calcoli devono però sempre essere
convalidati da misure di tipo dosimetrico fatte in aria e in fantocci tessuto-equivalenti, che
hanno cioè una composizione elementale la più simile possibile a quella del bersaglio da
irraggiare.
La microdosimetria, che cerca di descrivere le fluttuazioni dei depositi energetici in
volumi microscopici, può essere una soluzione adeguata alla caratterizzazione del campo di

47
radiazione della boroterapia. Per esempio, attraverso un contatore proporzionale a gas e
tessuto equivalente è possibile misurare la dose assorbita e lo spettro microdosimetrico dei
depositi energetici delle componenti del campo di radiazione. Pesando gli spettri con una
opportuna funzione è possibile determinare direttamente l’RBE delle singole componenti.
4.2 MICRODOSIMETRIA SPERIMENTALE
La possibilità di usare i concetti e il metodo sperimentale della microdosimetria nel
monitoraggio del campo di radiazione caratteristico della BNCT comporta dei vantaggi.
L’utilizzo di contatori proporzionali tessuto-equivalenti (TEPC) con due catodi di cui uno
arricchito con boro e l’altro non, le informazioni che si ricavano da uno spettro
microdosimetrico, assieme a misure di controllo sul fascio e alle informazioni riguardanti la
distribuzione del boro nelle cellule permettono di raggiungere una conoscenza accurata del
campo.
La prima applicazione di questa disciplina per la boroterapia risale al 1992 quando Wuu
et al. al BNL analizzarono gli spettri microdosimetrici ottenuti con un contatore
proporzionale tessuto equivalente (Rossi-type) con il catodo, costituito dalla plastica
SHONKA-A150, arricchito con 50 ppm di boro. Il gas di riempimento era propano-TE
mescolato con trifloruro di boro in una quantità equivalente a 50 ppm di B10 . Le misure
furono fatte sfruttando il fascio epitermico del BMRR (Brookhaven Medical Research
Reactor) a una potenza compresa tra 0.1 e 5 kW. In queste condizioni il rivelatore simulava
una cellula contenente del boro. La differenza tra lo spettro microdosimetrico ottenuto con il
rivelatore con il catodo arricchito in B10 e quello ottenuto con un contatore identico in tutto
ma senza boro è dovuta solamente all’effetto della cattura neutronica del boro.
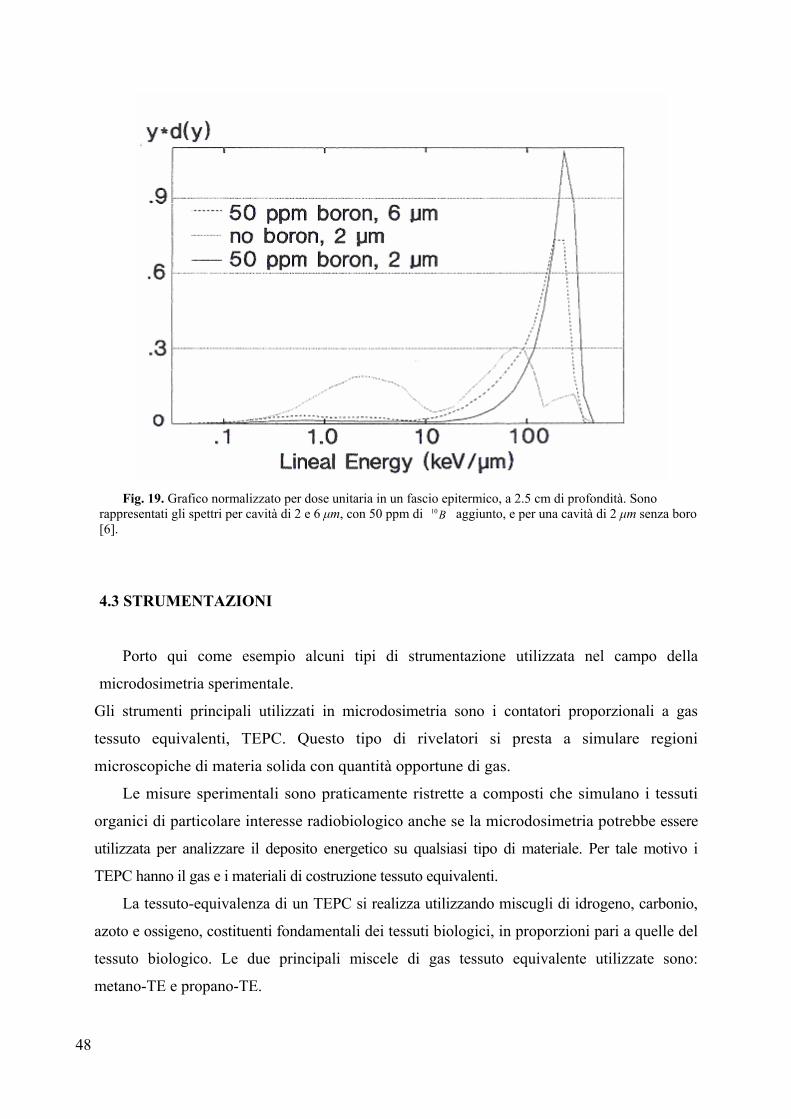
La Fig. 19, presa dall’esperimento di Wuu, mostra un grafico di energia lineale per
volumi di tessuto equivalente di 2 e 6 µm. E’ anche presente la curva per il volume di 2 µm in
assenza di boro.
Dalla figura emerge chiaramente come la presenza del boro aumenti notevolmente la
frazione della dose rilasciata con alti valori dell’energia lineale, ossia la componente a
maggiore efficacia biologica; quest’effetto è ancora più evidente nei siti di minore
dimensione.
48
Fig. 19. Grafico normalizzato per dose unitaria in un fascio epitermico, a 2.5 cm di profondità. Sono
rappresentati gli spettri per cavità di 2 e 6 µm, con 50 ppm di B10 aggiunto, e per una cavità di 2 µm senza boro [6].
4.3 STRUMENTAZIONI
Porto qui come esempio alcuni tipi di strumentazione utilizzata nel campo della
microdosimetria sperimentale.
Gli strumenti principali utilizzati in microdosimetria sono i contatori proporzionali a gas
tessuto equivalenti, TEPC. Questo tipo di rivelatori si presta a simulare regioni
microscopiche di materia solida con quantità opportune di gas.
Le misure sperimentali sono praticamente ristrette a composti che simulano i tessuti
organici di particolare interesse radiobiologico anche se la microdosimetria potrebbe essere
utilizzata per analizzare il deposito energetico su qualsiasi tipo di materiale. Per tale motivo i
TEPC hanno il gas e i materiali di costruzione tessuto equivalenti.
La tessuto-equivalenza di un TEPC si realizza utilizzando miscugli di idrogeno, carbonio,
azoto e ossigeno, costituenti fondamentali dei tessuti biologici, in proporzioni pari a quelle del
tessuto biologico. Le due principali miscele di gas tessuto equivalente utilizzate sono:
metano-TE e propano-TE.

49
TEPC Sferico
Storicamente i primi TEPC ad essere costruiti furono quelli sferici. I primi di questi
rivelatori avevano volumi sensibili che arrivavano anche a 8-10 cm di diametro. Di
conseguenza, oltre ad avere una pessima risoluzione spaziale, erano limitati a misure di campi
con intensità estremamente basse: poche particelle al secondo per cm2. I rivelatori sferici
sono tuttora quelli più commercializzati e disponibili anche se hanno dimensioni più
piccole (1-3cm) dei loro precursori (8-10cm).
Il volume sensibile è una sfera di plastica A-150 di 1.3 cm di diametro, riempita con
propano-TE (Fig. 20).
Fig. 20. Schema del rivelatore sferico, modello LET1/2", della Far West Technology [7]
TEPC Piano
Un microdosimetro piano è stato sviluppato da Cosgrove. Il rivelatore ha una cavità
cilindrica di 8 cm di diametro e 3 cm di profondità. Per evitare di non rilevare i protoni di più
bassa energia, il rivelatore ha una finestra di ingresso in Mylar alluminato da 50 µm. Dal lato
opposto all’entrata si ha una finestra di perspex in modo da poter misurare i protoni di più alta

50
energia che riescono a passare; in tal modo è possibile eseguire misure microdosimetriche in
coincidenza. Per simulare 2 µm è stato riempito con 70 mbar di metano-TE. L’anodo è un filo
centrale da 25 µm. Una schematizzazione di TEPC piano è data in Fig. 21:
Fig. 21. Schema del microdosimetro piano sviluppato da Cosgrove [7].
TEPC Cilindrico
Presso il GSI in Darmstadt (Germania) si trova una facility sperimentale per adroterapia
con fasci di ioni carbonio. Proprio per caratterizzare la qualità di questi fasci sono stati
sviluppati dei piccoli microdosimetri cilindrici.

51
Fig. 22. Microdosimetro cilindrico sviluppato da Gerlach per adroterapia con ioni carbonio [7]
Il volume sensibile di questo rivelatore è un cilindro di diametro 3 mm, lunghezza 30 mm
ed un filo centrale, anodo, da 20 µm. Le pareti del catodo sono realizzate con plastica
A-150 tessuto equivalente. Dentro al rivelatore viene fatto fluire di continuo metano tessuto
equivalente e la pressione è aggiustata in modo tale da simulare 1 µm. Le piccole dimensioni
del rivelatore consentono di farlo alloggiare in un piano di perspex e di posizionarlo a varie
profondità in un fantoccio sempre in perspex.
Le misure microdosimetriche ottenute con tale rivelatore sono state utilizzate per calcolare
l’RBE dei fasci di Carbonio.
Mini TEPC
In questi ultimi anni le applicazioni della microdosimetria si stanno rendendo sempre più
necessarie al fine di caratterizzare la qualità dei fasci impiegati sia in BNCT che in
adroterapia. Nel caso della terapia protonica si prevede l’utilizzo di flussi di circa
7108 ⋅sec2 ⋅cm
particelle . Con i TEPC a gas oggi disponibili tale monitoraggio non risulta
possibile senza attenuare il fascio.
Un altro limite riscontrato in letteratura e che si sta cercando di ovviare è la bassa

52
risoluzione spaziale. Accade infatti che a fronte di una precisione diagnostica del tumore del
paziente dell’ordine del millimetro, non si riesce a monitorare con altrettanta precisione, a
volte indispensabile, il campo di radiazione poichè i rivelatori hanno dimensioni superiori a tale
limite.
Un modo per evitare questi inconvenienti è quello di diminuire il volume sensibile del
TEPC in modo che il numero di particelle intercettate rimanga basso; sempre riducendo il
volume sensibile si risolve anche il secondo problema cioè si aumenta la precisione spaziale con
cui si misura il campo di radiazione.
Sfruttando le tecnologie usate alla Coulombia University furono costruiti nel 1998 presso
i LNL-INFN due mini TEPC con lo scopo di monitorare i fasci terapeutici. I due
rivelatori così costruiti sono stati denominati AMiCo1 e AMiCo2. I due rivelatori
risultano quasi identici. Esternamente essi hanno dimensioni di quasi 4 cm in lunghezza e 1.6
cm in spessore.
La differenza principale è: AMiCo1 possiede tubi di guardia (o di campo) mentre
AMiCo2 è stato costruito con la possibilità di mettere e togliere i tubi di guardia.
I tubi di guardia (o di campo) sono elettrodi che vengono mantenuti, relativamente
all’anodo, ad un potenziale uguale a quello che sarebbe esistito in corrispondenza della loro
superficie esterna se non fossero stati presenti e se il contatore fosse stato un cilindro
infinito. Uno studio ha evidenziato che in assenza di tubi di campo, quando un TEPC è
sottoposto ad un fascio di radiazione intenso, il segnale non si stabilizza. La necessità di
miniaturizzare i TEPC ha portato a studiare la possibilità di costruire dei rivelatori senza tubi
di guardia che funzionino in maniera stabile. AMiCo1 e AMiCo2 hanno permesso tale
studio.

53
Fig. 23. Disegno tecnico di AMiCo1 / 2, con il particolare del volume sensibile [7].
I rivelatori costruiti sono caratterizzati da un volume sensibile di 0.8 mm3 costituito da un
cilindro retto di 1 mm di diametro e 1 mm di altezza. L’anodo è un filo di tungsteno dorato da 10
µm. Nella Fig. 23 è rappresentato un disegno tecnico del rivelatore e in particolare i tubi di
campo da 250 µm di diametro. Il catodo è costituito da una plastica denominata Shonka A-150.
Il catodo è contenuto in un cilindro di Rexolite di 0.45 mm di spessore. Questo, oltre a dare
solidità al sistema, permette di isolare il catodo dal cilindro di alluminio dello spessore di
0.2 mm, che contiene il tutto. Alla base del rivelatore si trovano le connessioni elettriche per
anodo, catodo e tubi di campo, e i fori da 1 mm per il flussagio del gas.
In entrambi i rivelatori è stata inserita una sorgente alfa, Cm244, che può essere usata per fare
un’accurata calibrazione energetica. Le particelle alfa passano attraverso una finestra di
mylar da 1.4 µm di spessore e di 300 µm di diametro.

54
Telescopio a pixel al silicio
Fig. 24. (a) Schema del telescopio a pixel di silicio. (b) Schema del singolo elemento ∆E [12].
In Fig. 24 è rappresentato un telescopio in silicio composto da pixel per la
microdosimetria allo stato solido usato in un fascio di neutroni veloci. Esso consiste di una
matrice di elementi cilindrici ∆E di spessore pari a 2 µm impiantati in un supporto singolo E
con spessore di circa 500 µm. Più di 7000 pixel sono stati connessi tra loro in parallelo per
creare un’area sensibile efficace di circa 0.5 mm2. L’energia minima distinguibile è limitata a
20 keV dal rumore elettrico, per cui l’applicabilità di questo microdosimetro in silicio è utile
per particelle ad alto LET.
Il rivelatore ∆E- E è stato accoppiato con un convertitore in polietilene spesso 1 mm ed è
stato irradiato con neutroni a 2.7 MeV. La risposta al campo monodirezionale di neutroni
quasi monoenergetici è stato misurato all’acceleratore Van De Graaff dei Laboratori
Nazionali di Legnano (LNL) dell’INFN. Per l’energia del neutrone lo spettro dell’energia
impartita nel supporto ∆E dalle particelle secondarie generate nel convertitore, è stato
confrontato con quello misurato con un TEPC cilindrico tessuto equivalente. A questo
proposito la distribuzione di energia impartita negli elementi ∆E deve essere corretta per
equivalenza con tessuto in modo da ottenere uno spettro confrontabile con quello del TEPC.
Nella Fig. 25 è riportato il confronto tra le due distribuzioni in dose dell’energia impartita per
evento.

55
Fig. 25. Distribuzione in dose dell’energia impartita per evento acquisita con un TEPC (linea grigia) e con
un singolo ∆E del telescopio (linea nera). I rivelatori sono stati irradiati con neutroni mono energetici da 2.7 MeV [12].
Si possono notare delle discrepanze nella forma dello spettro del singolo supporto ∆E,
specialmente nelle alte energie. Le differenze sono dovute ad effetti geometrici legati alla
vasta area sensibile dei ∆E.

56
CONCLUSIONI
In questa tesi sono state esaminate rapidamente le problematiche connesse
all’interpretazione degli effetti biologici delle radiazioni. Abbiamo visto come spesso la dose
assorbita definita su domini macroscopici non sia in grado di fornire adeguate risposte. Si è
passati quindi ad esaminare i concetti di base della microdosimetria che affronta tali
problematiche da un punto di vista completamente diverso. In microdosimetria si fissa un
sito biologico con determinate dimensioni e si va a studiare direttamente la distribuzione dei
rilasci di energia in esso. Dalle distribuzioni di grandezze quali l’energia specifica o l’energia
lineale si possono trarre spunti utili a comprendere l’interazione radiazione – materia vivente.
La microdosimetria continuerà a supportare modelli biofisici nati per la formulazione di
spiegazioni del danno a DNA e cromosomi o per spiegare vari effetti biologici (ad esempio
morte cellulare in campi misti).
La microdosimetria sperimentale alle dimensioni dei micrometri continua a dimostrare il
suo valore nei campi della dosimetria, della radioprotezione e della radioterapia ovunque
nasca l’esigenza di separare le varie componenti di un campo di radiazioni rivelando le
particelle cariche responsabili del deposito di energia.
E’ portato avanti il continuo sviluppo dei metodi di misurazione microdosimetrici nei
campi della dosimetria spaziale e delle terapie ad alto LET presso impianti di adroterapia o
presso reattori nucleari usati in BNCT.
Sono stati quindi descritte le principali tecniche e alcuni tipi di strumenti utilizzati in
questo campo. I contatori proporzionali a bassa pressione continuano a rappresentare il
metodo dominante utilizzato anche se stanno emergendo rivelatori più sofisticati di nuova
generazione in grado di effettuare misure per siti a livello sub-micrometrico.
In un prossimo futuro si prevede di effettuare delle misure con un microdosimetro
miniaturizzato costruito presso il Laboratori Nazionali di Legnaro dell’INFN per
caratterizzare il campo neutronico della colonna termica del reattore Triga Mark II utilizzato
per la BNCT a Pavia.

57
BIBLIOGRAFIA
[1]. A. M. Kellerer: Fundamentals of Microdosimetry. In: K. R. Kase, B. E. Bjarngard, F. H.
Attix: The dosimetry of ionizing radiation, volume I. Academic Press, INC. Orlando, Florida 1987.
[2]. J. E. Coggle: Effetti biologici delle radiazioni. Edizioni Minerva Medica, Torino 1998.
[3]. D. T. Goodhead: Relationship of microdosimetric techniques to applications in biological systems. In: K. R. Kase, B. E. Bjarngard, F. H. Attix: The dosimetry of ionizing radiation, volume II. Academic Press, INC. Orlando, Florida 1987.
[4]. E. Seravalli: Monitoraggio microdosimetrico del campo di radiazione nella terapia per cattura neutronica del B-10. Tesi di laurea, Università degli studi di Padova 2003.
[5]. C. S. Wuu, H. I. Amols, P. Kliauga, L. E. Reinstein, S. Saraf: Microdosimetry for Boron Neutron Capture Therapy. Radiation Research, vol. 130, No. 3 (Jun 1992), pp. 355-359.
[6]. H. I. Amols, L. E. Reinstein: Stochastic considerations in BNCT: microdosimetry and cell survival. In: R. G. Zamenhof, G. R. Solares, O. K. Harling: Topics in dosimetry & treatment planning for Neutron Capture Therapy. Advanced Medical publishing, Madison, Wisconsin 1994.
[7]. D. Moro: Determinazione del fattore di qualità mediante TEPC miniaturizzati nella
radioterapia adronica. Tesi di laurea, Università degli studi di Padova 2003.
[8]. H. H. Rossi: Specification of radiation quality. Radiation Research 10,(1959), pp. 522-531.
[9]. V.P.Cosgrove, S.Delacroix et al.: Microdosimetric Studies on the Orsay proton synchrocyclotron at 73 and 200 MeV. Radiat. Prot. Dos., vol.70 (1997), Nos. 1-4, pp. 493-496.
[10]. R.Gerlach, H.Roos, A.M.Kellerer: Heavy ion RBE and Microdosimetric spectra. Radiat.
Prot. Dos., vol.99 (2002), No. 1-4, pp. 413-418.

58
[11]. P. Kliauga: Measurement of Single Event Energy Deposition Spectra at 5nm to 250 nm
Simulated site sizes. Radiat Prot. Dos., vol.31 (1990), No.1/4, pp 119-123.
[12]. S. Agosteo, P. G. Fallica, A. Fazzi, M. V. Introini, A. Pola, G. Valvo: A pixelated silicon telescope for solid state microdosimetry. Radiation Measurements 43 (2008) pp. 585-589.
[13]. A. J. Waker: Techniques for radiation measurements: microdosimetry and dosimetry. Radiation Protection Dosimetry, vol. 122, No. 1-4 (2006), pp. 369-373.