
Ultradian corticosterone pulses balance glutamatergic … · 2014-09-26 · when we quantified the...
6
Ultradian corticosterone pulses balance glutamatergic transmission and synaptic plasticity Ratna Angela Sarabdjitsingh a,1 , Julie Jezequel b,c , Natasha Pasricha a,b,c , Lenka Mikasova b,c , Amber Kerkhofs a,b,c , Henk Karst a , Laurent Groc b,c,1,2 , and Marian Joëls a,1,2 a Department of Translational Neuroscience, Brain Center Rudolf Magnus, University Medical Center Utrecht, 3584 CG, Utrecht, The Netherlands; b Interdisciplinary Institute for Neuroscience, Unité Mixte de Recherche 5297, Université de Bordeaux, F-33000 Bordeaux, France; and c Interdisciplinary Institute for Neuroscience, Unité Mixte de Recherche 5297, Centre National de la Recherche Scientifique, F-33000 Bordeaux, France Edited* by Donald W. Pfaff, The Rockefeller University, New York, NY, and approved August 18, 2014 (received for review June 16, 2014) The rodent adrenal hormone corticosterone (CORT) reaches the brain in hourly ultradian pulses, with a steep rise in amplitude before awakening. The impact of a single CORT pulse on gluta- matergic transmission is well documented, but it remains poorly understood how consecutive pulses impact on glutamate receptor trafficking and synaptic plasticity. By using high-resolution imag- ing and electrophysiological approaches, we report that a single pulse of CORT to hippocampal networks causes synaptic enrich- ment of glutamate receptors and increased responses to sponta- neously released glutamatergic vesicles, collectively abrogating the ability to subsequently induce synaptic long-term potentia- tion. Strikingly, a second pulse of CORT one hour after the first— mimicking ultradian pulses—completely normalizes all aspects of glutamate transmission investigated, restoring the plastic range of the synapse. The effect of the second pulse is precisely timed and depends on a nongenomic glucocorticoid receptor-dependent path- way. This normalizing effect through a sequence of CORT pulses— as seen around awakening—may ensure that hippocampal glutama- tergic synapses remain fully responsive and able to encode new stress-related information when daily activities start. hippocampus | AMPA receptor trafficking T he rodent stress hormone corticosterone (CORT) is synthe- sized in the adrenal glands. Through the circulation, it reaches the brain, where it binds to intracellular receptors, i.e., the high- affinity mineralocorticoid receptor (MR) and lower affinity glu- cocorticoid receptor (GR) (1). These receptors are enriched in the hippocampus and act as transcriptional regulators (2). Intracellular MRs are already considerably occupied with low (i.e., nonstress) levels of CORT, whereas GRs become substantially bound only when corticosteroid levels increase (1). After stress, hormone levels slowly increase and normalize after 2 h as a result of negative feedback actions in the pituitary and hypothalamus (3). Corticosteroid levels also show an endogenous circadian variation, which in fact is carried by brief hourly (ultra- dian) pulses (4). Pulse amplitudes steeply rise some hours before awakening and then slowly diminish again (5). The functional relevance of ultradian pulses is only starting to be understood; recent evidence suggests that pulses are necessary to maintain optimal transcriptional activity of GRs (4–8), thus coordinating essential bodily functions in preparation of the active phase. It is well known that, through DNA binding and transcriptional regulation, GRs slowly and persistently alter glutamatergic trans- mission in the hippocampus (9–11). A brief CORT pulse increases the surface expression and synaptic localization of AMPA receptor (AMPAR) subunits (10) and increases the amplitude of miniature excitatory postsynaptic currents (mEPSCs) (11)—the postsynaptic response to a spontaneously released glutamate-containing vesicle. CORT-dependent signaling is thought to (partly) overlap with pathways giving rise to long-term potentiation (LTP) (12, 13). In agreement, administration of CORT occludes the subsequent in- duction of chemically (10) and electrically evoked LTP (9, 14). In line with GRs acting as transcriptional regulators, these effects become apparent ∼1 h after CORT administration and last for many hours. We here examined how glutamatergic transmission in the hippocampus is affected when cells are exposed to a second pulse of CORT 1 h after the first, mimicking the ultradian pattern of endogenous CORT release (6). Results Consecutive CORT Pulses Normalize AMPAR Surface Dynamics and Synaptic Content. Previously, we showed in hippocampal neurons that a single pulse of CORT drastically increases the surface dif- fusion and synaptic content of GluA2- and GluA1-containing AMPARs several hours later (10). This CORT-induced synaptic potentiation of AMPARs was prevented by selectively and artificially blocking AMPAR surface diffusion. Furthermore, the increased surface trafficking and synaptic accumulation of AMPARs after CORT exposure likely contributed to the occlusion of classical synaptic LTP (10, 13). To investigate the impact of CORT pulsatility, we first examined how two 10-min pulses of CORT (100 nM) with an interval of 1 h affect the surface dynamics of AMPARs. For this, we took advantage of the single particle tracking approach to monitor individual particle/receptor com- plexes in hippocampal cultured neurons (Fig. 1A). Hippocampal networks were exposed to one or two pulses, and single nano- particle imaging was performed 120 min after onset of the first exposure (Fig. 1B). Consistent with previous results (10), a single CORT pulse significantly increased the surface diffusion of Significance A pulse of the adrenal hormone corticosterone (CORT) changes hippocampal glutamate transmission for many hours. CORT is normally released in hourly pulses, with a steeply rising am- plitude just before awakening. How organisms can be pre- pared for imminent danger if the first high-amplitude pulse of CORT would lastingly change glutamate transmission—thus potentially deadlocking the system—has remained an enigma. We show that exposure of hippocampal cells to a second high- amplitude CORT pulse completely normalizes all aspects of glutamate transmission (including synaptic plasticity), thus lift- ing the potential deadlock caused by a first pulse. This ensures that the system remains fully responsive to any stressful event that requires encoding of information, an important principle that promotes survival of individuals. Author contributions: R.A.S., J.J., N.P., L.M., H.K., L.G., and M.J. designed research; R.A.S., J.J., N.P., L.M., A.K., and H.K. performed research; L.G. contributed new reagents/analytic tools; R.A.S., J.J., N.P., L.M., A.K., H.K., and L.G. analyzed data; and R.A.S., J.J., N.P., L.M., A.K., H.K., L.G., and M.J. wrote the paper. The authors declare no conflict of interest. *This Direct Submission article had a prearranged editor. 1 To whom correspondence may be addressed. Email: [email protected], [email protected], or [email protected]. 2 L.G. and M.J. contributed equally to this work. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1411216111/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1411216111 PNAS | September 30, 2014 | vol. 111 | no. 39 | 14265–14270 NEUROSCIENCE Downloaded by guest on April 14, 2020
Transcript of Ultradian corticosterone pulses balance glutamatergic … · 2014-09-26 · when we quantified the...

Ultradian corticosterone pulses balance glutamatergictransmission and synaptic plasticityRatna Angela Sarabdjitsingha,1, Julie Jezequelb,c, Natasha Pasrichaa,b,c, Lenka Mikasovab,c, Amber Kerkhofsa,b,c,Henk Karsta, Laurent Grocb,c,1,2, and Marian Joëlsa,1,2
aDepartment of Translational Neuroscience, Brain Center Rudolf Magnus, University Medical Center Utrecht, 3584 CG, Utrecht, The Netherlands;bInterdisciplinary Institute for Neuroscience, Unité Mixte de Recherche 5297, Université de Bordeaux, F-33000 Bordeaux, France; and cInterdisciplinaryInstitute for Neuroscience, Unité Mixte de Recherche 5297, Centre National de la Recherche Scientifique, F-33000 Bordeaux, France
Edited* by Donald W. Pfaff, The Rockefeller University, New York, NY, and approved August 18, 2014 (received for review June 16, 2014)
The rodent adrenal hormone corticosterone (CORT) reaches thebrain in hourly ultradian pulses, with a steep rise in amplitudebefore awakening. The impact of a single CORT pulse on gluta-matergic transmission is well documented, but it remains poorlyunderstood how consecutive pulses impact on glutamate receptortrafficking and synaptic plasticity. By using high-resolution imag-ing and electrophysiological approaches, we report that a singlepulse of CORT to hippocampal networks causes synaptic enrich-ment of glutamate receptors and increased responses to sponta-neously released glutamatergic vesicles, collectively abrogatingthe ability to subsequently induce synaptic long-term potentia-tion. Strikingly, a second pulse of CORT one hour after the first—mimicking ultradian pulses—completely normalizes all aspects ofglutamate transmission investigated, restoring the plastic range ofthe synapse. The effect of the second pulse is precisely timed anddepends on a nongenomic glucocorticoid receptor-dependent path-way. This normalizing effect through a sequence of CORT pulses—as seen around awakening—may ensure that hippocampal glutama-tergic synapses remain fully responsive and able to encode newstress-related information when daily activities start.
hippocampus | AMPA receptor trafficking
The rodent stress hormone corticosterone (CORT) is synthe-sized in the adrenal glands. Through the circulation, it reaches
the brain, where it binds to intracellular receptors, i.e., the high-affinity mineralocorticoid receptor (MR) and lower affinity glu-cocorticoid receptor (GR) (1). These receptors are enriched in thehippocampus and act as transcriptional regulators (2). IntracellularMRs are already considerably occupied with low (i.e., nonstress)levels of CORT, whereas GRs become substantially bound onlywhen corticosteroid levels increase (1).After stress, hormone levels slowly increase and normalize after
2 h as a result of negative feedback actions in the pituitary andhypothalamus (3). Corticosteroid levels also show an endogenouscircadian variation, which in fact is carried by brief hourly (ultra-dian) pulses (4). Pulse amplitudes steeply rise some hours beforeawakening and then slowly diminish again (5). The functionalrelevance of ultradian pulses is only starting to be understood;recent evidence suggests that pulses are necessary to maintainoptimal transcriptional activity of GRs (4–8), thus coordinatingessential bodily functions in preparation of the active phase.It is well known that, through DNA binding and transcriptional
regulation, GRs slowly and persistently alter glutamatergic trans-mission in the hippocampus (9–11). A brief CORT pulse increasesthe surface expression and synaptic localization of AMPA receptor(AMPAR) subunits (10) and increases the amplitude of miniatureexcitatory postsynaptic currents (mEPSCs) (11)—the postsynapticresponse to a spontaneously released glutamate-containing vesicle.CORT-dependent signaling is thought to (partly) overlap withpathways giving rise to long-term potentiation (LTP) (12, 13). Inagreement, administration of CORT occludes the subsequent in-duction of chemically (10) and electrically evoked LTP (9, 14). Inline with GRs acting as transcriptional regulators, these effects
become apparent ∼1 h after CORT administration and last formany hours. We here examined how glutamatergic transmission inthe hippocampus is affected when cells are exposed to a secondpulse of CORT 1 h after the first, mimicking the ultradian patternof endogenous CORT release (6).
ResultsConsecutive CORT Pulses Normalize AMPAR Surface Dynamics andSynaptic Content. Previously, we showed in hippocampal neuronsthat a single pulse of CORT drastically increases the surface dif-fusion and synaptic content of GluA2- and GluA1-containingAMPARs several hours later (10). This CORT-induced synapticpotentiation of AMPARs was prevented by selectively andartificially blocking AMPAR surface diffusion. Furthermore, theincreased surface trafficking and synaptic accumulation ofAMPARs after CORT exposure likely contributed to the occlusionof classical synaptic LTP (10, 13). To investigate the impact ofCORT pulsatility, we first examined how two 10-min pulses ofCORT (100 nM) with an interval of 1 h affect the surface dynamicsof AMPARs. For this, we took advantage of the single particletracking approach to monitor individual particle/receptor com-plexes in hippocampal cultured neurons (Fig. 1A). Hippocampalnetworks were exposed to one or two pulses, and single nano-particle imaging was performed 120 min after onset of the firstexposure (Fig. 1B). Consistent with previous results (10), a singleCORT pulse significantly increased the surface diffusion of
Significance
A pulse of the adrenal hormone corticosterone (CORT) changeshippocampal glutamate transmission for many hours. CORT isnormally released in hourly pulses, with a steeply rising am-plitude just before awakening. How organisms can be pre-pared for imminent danger if the first high-amplitude pulse ofCORT would lastingly change glutamate transmission—thuspotentially deadlocking the system—has remained an enigma.We show that exposure of hippocampal cells to a second high-amplitude CORT pulse completely normalizes all aspects ofglutamate transmission (including synaptic plasticity), thus lift-ing the potential deadlock caused by a first pulse. This ensuresthat the system remains fully responsive to any stressfulevent that requires encoding of information, an importantprinciple that promotes survival of individuals.
Author contributions: R.A.S., J.J., N.P., L.M., H.K., L.G., and M.J. designed research; R.A.S.,J.J., N.P., L.M., A.K., and H.K. performed research; L.G. contributed new reagents/analytictools; R.A.S., J.J., N.P., L.M., A.K., H.K., and L.G. analyzed data; and R.A.S., J.J., N.P., L.M.,A.K., H.K., L.G., and M.J. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.1To whom correspondence may be addressed. Email: [email protected],[email protected], or [email protected].
2L.G. and M.J. contributed equally to this work.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1411216111/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1411216111 PNAS | September 30, 2014 | vol. 111 | no. 39 | 14265–14270
NEU
ROSC
IENCE
Dow
nloa
ded
by g
uest
on
Apr
il 14
, 202
0

GluA2–AMPAR (Fig. 1 C and D). Strikingly, the introduction ofa second pulse of CORT—i.e., 60 min after the first—abolishedthis effect, yielding a surface diffusion comparable to the vehiclecontrol situation (Fig. 1 C and D). Of note, the second pulse didnot simply delay the CORT-induced AMPAR surface diffusionincrease, as the dynamics measured 120 min after the second pulse(at t 180 min) were undistinguishable from the one measured60 min after two pulses at t 120 min (value at t 180 min GluA2–AMPAR surface diffusion: median, 0.092 μm2/s; interquartilerange, 4.10−3 to 0.14 μm2/s; n = 670 trajectories; P > 0.05 vs.CORT–CORT diffusion at 120 min; Fig. 1). The diffusion changesobserved after one or two CORT pulses at t 120 min appear tobe mostly caused by a change in the mobile fraction (diffusion >0.005 μm2/s), as control (54%) and two-pulse (47%) valueswere lower than the fraction seen after one pulse (76%). To-gether, to our knowledge, these data provide the first evidencethat consecutive pulses of CORT restore the surface traffickingof AMPARs toward a basal range.
We next investigated the impact of consecutive CORT pulseson synaptic AMPAR signaling by using high-resolution imagingand electrophysiological means. First, the single particle trackingapproach allows to point, with subwavelength accuracy (∼20 nmresolution), the location of tagged-receptors in specific mem-brane compartments, such as the synapse. Based on that, weestimated that, on average, fewer than 10% of the labeledAMPARs are present in the synapse under control conditions.This value significantly increased after exposure to a singleCORT pulse whereas it was comparable to baseline after twopulses (Fig. 1E), indicating that the second pulse reverses ornormalizes the CORT-induced GluA2-AMPAR synaptic accu-mulation caused by the first pulse. A similar effect was observedwhen we quantified the area of the postsynaptic density (PSD) ofsynapses exposed to a single or two CORT pulses (Fig. 1F). Ofnote, the increase in GluA2–AMPAR synaptic fraction observedafter one pulse (+304%) was larger than the increase of the PSDarea (122%), suggesting that the altered AMPAR content is anactive process that cannot solely be explained by a change in PSDarea. Together, these data demonstrate that a second pulse ofCORT (60 min after the first pulse) reverses or prevents thesynaptic accumulation of glutamate synapses observed 120 minafter a single pulse.We predicted that functional indices of glutamatergic trans-
mission would follow a similar pattern, i.e., that signals viaAMPAR subunits would be enhanced 2 h after a single pulse butnormalized when tested after two consecutive pulses. In CA1pyramidal neurons, recorded in slices from 2-mo-old mice, wefirst replicated that, ∼120 min after a single brief (10 min) pulseof CORT, the mEPSC amplitude was significantly increased(vehicle, 14.3 ± 0.6, n = 9; CORT, 18.4 ± 1.3, n = 7; P < 0.01;Fig. 2). As predicted, such increase was not seen in a secondseries of experiments in which the 2-h delay after the first pulsewas interrupted by another CORT pulse (60-min interval be-tween pulses; vehicle, 16.4 ± 1.0, n = 7; CORT, 16.9 ± 1.0, n = 8;P = 0.73; Fig. 2). In none of the conditions did CORT treatmentaffect mEPSC frequency (all P > 0.05; Table S1). Altogether,these data provide direct evidence that two consecutive pulses,60 min apart, restore the AMPAR surface dynamics and synapticcontent to values close to those seen under basal conditions.
CORT Pulses Restore the Plastic Range of Hippocampal GlutamateSynapses. Earlier studies showed that exposure of hippocampalslices to a single pulse of CORT impairs the induction of LTP insubsequent hours (9, 14). This was interpreted as a CORT-induced
Fig. 1. Consecutive CORT pulses normalize AMPAR surface distribution andsynaptic content. (A) Surface GluA2 was labeled with a single quantum dot(QD)-antibody complex (Left), allowing single particle tracking. (Right)Representative trajectories (yellow lines; 40-s duration imaging; 20-Hz ac-quisition rate) in cultured hippocampal neurons (15 d in vitro). (Scale bar:5 μm.) (B) Schematic representation of the experimental protocol. Neuronswere exposed to one pulse of 100 nM CORT (or vehicle) for 10 min or twoconsecutive 100 nM CORT pulses (or vehicle) with an interval of 60 min. In allexperiments, surface AMPAR trafficking was assessed at 120 min relative tothe onset of the first CORT application. (C) Representative single GluA2–AMPAR trajectories in the different experimental paradigms described in B.Note the increased lateral diffusion 120 min after a single pulse, an up-regulation not observed in presence of two pulses (100–100 nM CORT).(Scale bar: 500 nm.) (D) Distribution (median ± 25–75% interquartile range)of the instantaneous diffusion coefficients of surface GluA2–AMPAR, mea-sured 120 min after one pulse of 100 nM CORT or two consecutive 100 nMCORT pulses (or vehicle). The diffusion coefficient was significantly increasedafter a single CORT pulse (0′; n = 2,790 trajectories; 100 nM CORT, n = 916;100–100 nM CORT, n = 449; vehicle–vehicle, n = 452; Kruskal–Wallis test,P < 0.05; post hoc tests, **P < 0.01, 0′ control vs. 100 nM CORT; P > 0.05,100 nM CORT vs. other conditions). (E ) Single QDs (black dots) can belocated with a high pointing accuracy in a given membrane compart-ment, such as the synapse (syn., gray area). (Left) Examples of 500 framestacks obtained while tracking down single GluA2–AMPAR/QD com-plexes. Those 500 locations are then projected on a single image, pro-viding the successive positions of this receptor/particle complex in thevarious experimental paradigms. (Right) Relative fraction of synapticGluA2–AMPAR/QD particles (control, n = 28 neuronal fields; 100 nMCORT, n = 19; 100–100 nM CORT, n = 27; Kruskal–Wallis test, P < 0.05;post hoc tests, **P < 0.01, control vs. 100 nM CORT or 100 nM CORT vs.100–100 nM CORT; P > 0.05, control vs. 100–100 nM CORT). (F ) The PSDarea was quantified by using immunocytochemical staining of Homer 1cor Shank proteins (control, n = 315 PSDs; 100 nM CORT, n = 259; 100–100nM CORT, n = 212; one-way ANOVA test, P < 0.05; *P < 0.05, control vs.100–100 nM CORT or 100 nM CORT vs. 100–100 nM CORT).
Fig. 2. A single CORT pulse increases mEPSC amplitude, whereas two pulsesdo not. (A) Typical traces showing mEPSCs of a mouse CA1 hippocampal cell∼120 min after exposure to a brief (10 min) pulse of vehicle (Left) or 100 nMCORT (Right). (B) The mEPSC amplitude was significantly enhanced in neu-rons recorded ∼120 min after a single pulse of CORT as opposed to vehicle(Left). By contrast, two consecutive CORT pulses, with a 1-h interval, causedno change in mEPSC amplitude compared with two vehicle pulses whenneurons were recorded ∼120 min after onset of the first pulse. Data repre-sent the mean ± SEM of the mEPSC amplitudes recorded 5–10 min afterestablishing the whole cell configuration. To allow easy comparison, wenormalized the effects of CORT exposure to those of vehicle in bothexperiments (see main text for absolute values). *P < 0.01, unpaired t test.
14266 | www.pnas.org/cgi/doi/10.1073/pnas.1411216111 Sarabdjitsingh et al.
Dow
nloa
ded
by g
uest
on
Apr
il 14
, 202
0

occlusion of pathways necessary for the development of LTP (12,13). We therefore examined how synaptic plasticity is affected byconsecutive pulses. First, we tested the impact of consecutive pulseson LTP expression in hippocampal cultured neurons by usinga previously described chemical LTP (cLTP) induction protocol(10) and measuring the synaptic content (synapses were identifiedby Homer 1c detection) of AMPARs through live immunocyto-chemical staining (Fig. 3A). We focused our attention on GluA1–AMPAR because of their primary and well-characterized recruitmentto synapses following LTP (15). Application of a single pulse ofCORT prevented the cLTP-induced GluA1–AMPAR synapticrecruitment, as previously described (10) (Fig. 3 B and C).However, when neurons were exposed to two consecutive pulses,full-blown synaptic recruitment was observed 120 min after the firstpulse (Fig. 3B), indicating that the second pulse restores the po-tentiation range of the glutamate synapses. Consistent with theaforementioned electrophysiological data, two consecutive pulsesof CORT maintained the synaptic content of GluA1–AMPARclose to basal values (Fig. 3C). Together, these data indicate thata second pulse of CORT normalizes the potentiation range ofglutamate synapses, which was occluded by the first pulse.Hippocampal cultured cells are not connected to their normal
afferent and efferent fibers as in the intact brain. We thereforeswitched to acutely prepared hippocampal slices from youngadult mice and used a high-frequency paradigm that (in vehicle-treated slices) causes mild LTP in the CA1 area (Fig. 4 A1 andB). Induction of LTP was fully prevented by a single 10-min pulseof CORT applied to the slices 120 min before high-frequencystimulation [F(2, 21) = 15.77; P < 0.001; vehicle vs. one CORTpulse; P < 0.001; Fig. 4 A1, B, and C]. However, if we applied twoCORT pulses (with a 1-h interpulse interval), synaptic potentiationcaused by high-frequency stimulation 120 min after the first pulse
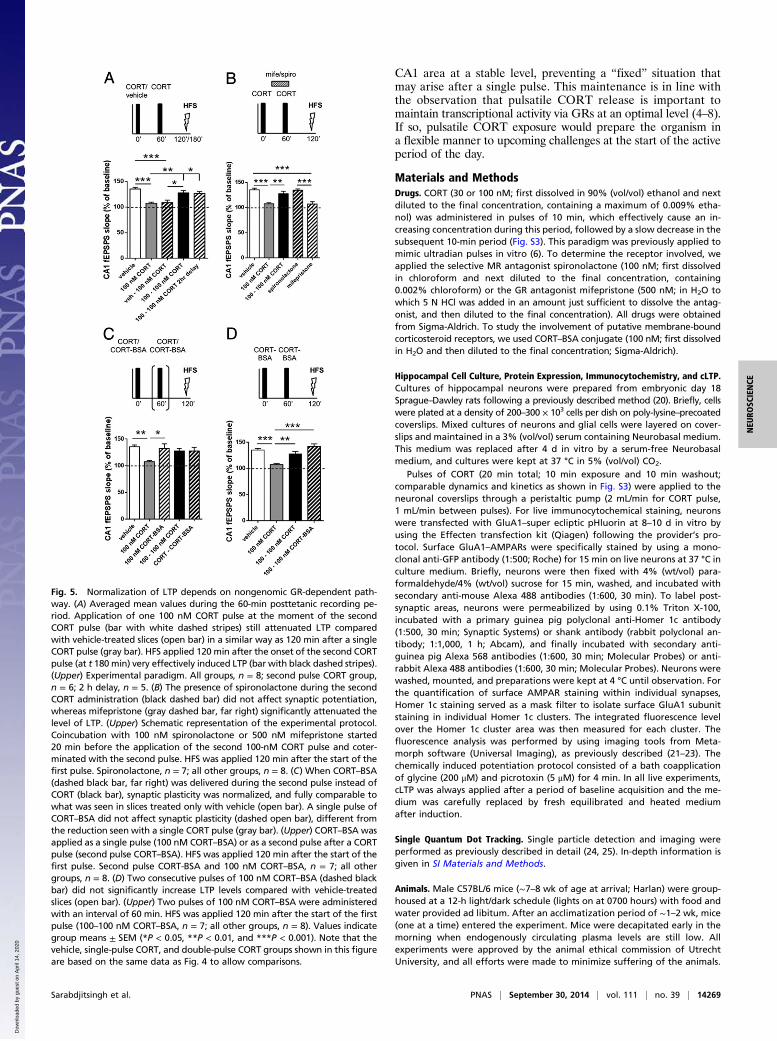
was comparable to that seen in the vehicle group (P > 0.1 vs.vehicle; Fig. 4 A2, B, and C). As 100 nM CORT is most likelya concentration not reached under physiological conditions, wealso tested two pulses of 30 nM CORT. This resulted in compa-rable results [F(2, 21) = 9.87; P < 0.01], i.e., suppression of LTP 120min after a single CORT pulse (P < 0.01 vs. vehicle), but efficientLTP after two pulses (P > 0.1 vs. vehicle; Fig. 4D and Fig. S1A). Wealso wondered to what extent the naturally occurring 1-h interval isrelevant for the observed phenomenon. As is evident from Fig. 4Eand Fig. S1B, the interval is indeed important [F(4, 29) = 12.52; P <0.001]: no normalization of LTP was seen when the second pulsewas given 10 min after the first pulse (P < 0.001 vs. vehicle), andonly partial normalization was found with an interpulse interval of30 min (P < 0.05 vs. vehicle). With intervals of 60 or 90 min, LTPinduced 120 min after onset of the first pulse was comparable tothat in the vehicle control group (P > 0.1 vs. vehicle). Together,these results suggest that the CORT-induced restoration of LTP isphysiologically relevant and depends on the timing of the secondCORT pulse.
CORT Pulses’ Effects on the Plastic Range Are Nongenomic andGR-Dependent. To better understand the mechanism contributingto normalization of LTP as seen after two pulses of CORT, wefirst questioned whether the second pulse reversed or preventedthe effect of the first pulse. Therefore, we tested if suppression ofLTP after a single pulse was already seen after 60 min, i.e., at thetime point at which the second CORT pulse normally would bedelivered. As shown in Fig. 5A and Fig. S2A, this was indeed thecase [F(4, 30) = 10.75; P < 0.001; P < 0.001 vs. vehicle). Addi-tionally, to exclude time-dependency of CORT effects, we testedfor LTP 120 min after the second pulse (at t 180 min). At thistime point, LTP was efficiently induced, suggesting that thesecond pulse lastingly reverses or compensates for the con-sequences of the first CORT application (P < 0.05 vs. 100 nMCORT; Fig. 5A and Fig. 2A). The results from the experiment inwhich two pulses were applied with a 90-min interval (leavingonly 20 min between the end of the second pulse and LTP in-duction; Fig. 4E) suggest that this reversal or compensationdevelops rapidly, in a time frame that seems incompatible withthe classic genomic pathway. Such nongenomic signaling byCORT has indeed been described in recent years (16–18). In thehippocampus, nongenomic signaling so far was found to involveMRs (16), but, in other areas of the brain (e.g., the hypothalamusor amygdala), rapid effects through GRs have been described(17, 18). To determine the receptor mediating the reversal orcompensation, we applied the second pulse of CORT in thepresence of an MR or GR antagonist. As is clear from Fig. 5Band Fig. S2B, we observed significant differences [F(4, 34) = 13.68;P < 0.001] between the treatment groups. The MR antagonistspironolactone was entirely ineffective in blocking the effect ofCORT (P > 0.1 vs. vehicle), whereas the GR antagonist mife-pristone fully prevented the normalizing effect of the secondCORT pulse (P < 0.001 vs. vehicle), supporting involvement ofGRs. Nongenomic, as opposed to genomic, actions of CORT canbe mimicked by the membrane impermeable conjugate of CORTto BSA (CORT–BSA). In agreement with a nongenomic pathway,normalization of LTP was replicated when slices were exposedto CORT–BSA, delivered 60 min after a first pulse of CORT[F(4, 33) = 4.16; P < 0.01; P > 0.1 vs. vehicle or 100–100 nM CORT;Fig. 5C and Fig. S2C]; in agreement with the genomic pathwayunderlying the response to the first pulse, a single pulse of CORT–BSA was unable to suppress LTP induced 120 min later (P > 0.1 vs.vehicle). Finally, we reasoned that, if the second pulse of CORTwould compensate for (i.e., act in the opposite direction) ratherthan reverse the response to the first pulse, a sequence of twopulses of CORT–BSA might result in enhanced LTP comparedwith the effect seen with two pulses of CORT. Although LTPlevels were, on average, slightly increased after two pulses of
Fig. 3. Consecutive CORT pulses restore cLTP-induced AMPAR potentiation.(A) (Left) Representative dendritic fragment of hippocampal neurons express-ing GluA1–SEP AMPAR. (Scale bar: 1 μm.) (Right) Enlargement of a synapticGluA1–SEP cluster, with the postsynaptic area labeled by the detection ofHomer-1c (dashed black line). (B) (Left, Center) GluA1–SEP dendritic frag-ments from neurons under basal condition or after the application of a cLTPprotocol with or without the presence of CORT pulses [protocols (Left):control cLTP, 100 nM CORT cLTP, 100–100 nM CORT cLTP, or 100–100 nMCORT]. The pseudocolor GluA1–SEP subunit representation shows the dif-ferent intensity levels of the GluA1–SEP staining. (Scale bar: 1 μm.) (Right)Enlargement of a GluA1–SEP cluster. (Scale bar: 500 nm.) Note that cLTPfails to potentiate GluA1–SEP fluorescence after a single CORT pulse.(C) Normalized measures of GluA1–SEP clusters intensity compared withbaseline (n = 3,495 clusters). The cLTP protocol induced an increasedGluA1–SEP expression in control condition (one-way ANOVA test, P < 0.05;control cLTP, n = 2,619 clusters; **P < 0.01) and after consecutive CORTpulses (100–100 nM CORT cLTP, n = 1,458 clusters; **P < 0.01), whereasGluA1–SEP fluorescence remained unchanged after a single pulse (100 nMCORT cLTP, n = 4,372 clusters).
Sarabdjitsingh et al. PNAS | September 30, 2014 | vol. 111 | no. 39 | 14267
NEU
ROSC
IENCE
Dow
nloa
ded
by g
uest
on
Apr
il 14
, 202
0

CORT–BSA, especially directly after high frequency stimulation, wedid not observe a significant effect [F(3, 27) = 14.67; P < 0.001; P >0.1 for CORT–BSA/CORT–BSA vs. vehicle or 100 nM CORT–CORT; Fig. 5D and Fig. S2C]. Together, these experiments indicatethat the normalizing effect of consecutive CORT pulses is mediatedby a noncanonical membrane GR-dependent signaling pathway.
DiscussionA single pulse of CORT facilitates glutamatergic transmission inhippocampal cells (9–11, 13), a condition that is associated with
an impaired ability to subsequently induce synaptic potentiation(12, 13). This has been interpreted earlier as a means to promotethe encoding of stress-related information, by protecting theencoding process from interfering input reaching the same syn-apses at a later point in time (12). Although this may be bene-ficial when CORT is released as part of a stress response, it maydeadlock the system when CORT is increased as a result of en-dogenous circadian variations: this would fully prevent encoding ofinformation for many hours after the first high-amplitude ultra-dian CORT pulse before awakening.We show that a second pulse of CORT, hitting the cells at a time
point when the effects of the first pulse have already developed, canfully normalize all investigated aspects of glutamatergic transmissionin the hippocampus and restore responsiveness to high-frequencydependent encoding. The system does so by triggering an entirelydifferent signaling pathway after the second pulse than the oneinvolved in the first pulse. Although, in the absence of CORT,surface expression of corticosteroid receptors is apparently low(because CORT–BSA was ineffective in mimicking the effect ofCORT in the single-pulse paradigm), the first pulse of CORT maytrigger surface localization of GRs, thus priming the system toa nongenomic effect that effectively reverses the genomic suppres-sion of LTP.The observation that limbic cells respond differently to a sec-
ond pulse of CORT than to the first is not unprecedented. Re-cently, we reported that a single pulse of CORT quickly butpersistently increases mEPSC frequency in amygdala cells (18);through this persistent effect, amygdala cell characteristics areslowly changed such that they respond in an opposite manner toa second pulse arriving >1 h after the first. Here we show that, inthe hippocampus, metaplastic responses can be observed notonly at the level of mEPSC amplitude, but also with respect toAMPAR trafficking and synaptic plasticity.The selected CORT application paradigm was based on ear-
lier studies (e.g., ref. 6) and mimics as closely as possible thenatural pulsatile pattern with gradually rising/falling flanks andvery low CORT levels during the interpulse interval (4) (Fig. S3).It therefore seems unlikely that our observations are a mereproduct of an artificial “on/off” paradigm. Although our reducedpreparations—cultured cells and brain slices—allow a straight-forward interpretation of the results, we are fully aware that theylack the complex connectivity of the brain and connections withperipheral organs. It would thus be interesting and important tosee if similar phenomena occur in the intact organism. Un-fortunately, this is currently not feasible at the single-cell levelbecause of technical issues.Most of the experiments described here were performed with
100 nM CORT because earlier studies have shown very clearactions with this concentration. Most likely, this is a supra-physiological concentration (19). The fact that we also sawnormalization of LTP with two pulses of 30 nM CORT supportsphysiological relevance, but future experiments would need toexplore the concentration dependency in greater detail. Simi-larly, it would be of interest to follow metaplastic changes formore than the current 2-h range. Finally, several studies haveshown that nuclear GR signaling—e.g., translocation, binding,and transcription of target genes—normalizes to baseline levelsbetween pulses (4, 6–8). It is therefore of great interest to resolvein detail if and particularly how a first pulse of CORT could leadto enhanced surface expression of GRs and which signalingpathways underlie the reversal exerted by the second pulse onprocesses activated by the first pulse. In that respect, additionalexperiments that interfere with transcriptional and/or trans-lational regulation by CORT would strengthen the notion ofnongenomic signaling regarding the latter, which is presentlybased only on our results with CORT–BSA.In summary, we show that pulsatile exposure to CORT has the
potential to maintain glutamatergic transmission in the hippocampal
Fig. 4. Synaptic plasticity in hippocampal slices is restored by consecutiveCORT pulses. (A1 and A2) Schematic representation of the experimentalprotocol. (A1) Mouse brain slices were exposed to one pulse of 100 nM CORT(or vehicle) for 10 min. (A2) Brain slices were exposed to two consecutive100 nM CORT pulses (or vehicle) with an interval of 60min. In both experiments,high-frequency stimulation (HFS) was applied at 120 min relative to theonset of the first CORT application. (B) HFS (10 Hz, 90 s) resulted in signifi-cant potentiation of synaptic responses at CA1 synapses in vehicle-treatedbrain slices (open circles). LTP was attenuated by 100 nM CORT given 120 minbefore HFS (gray circles). A second application of 100 nM CORT 60 min afterthe first pulse reversed this effect (black circles). (Upper) Representative in-dividual fEPSP traces for the vehicle group taken during baseline or ∼20 minafter HFS. (Scale bars: horizontal, 5 ms; vertical, 0.5 mV.) (C) Averaged meanvalues during the 60-min posttetanic recording period indicate significantlylower LTP in slices treated 120 min earlier with one 100 nM CORT pulse (graybar) compared with vehicle (open bar). A second 100-nM CORT pulse applied60 min after the first yielded LTP comparable to that seen in the vehiclegroup (black bar). (Upper) Experimental paradigm (see also A1 and A2).Thesecond CORT pulse in between brackets was applied for only the group re-ceiving two CORT pulses (100–100 nM CORT). All groups, n = 8. (D) Similarresults were found with lower, physiologically more relevant CORT con-centrations (30 nM). (Upper) Schematic representation of the experimentalprotocol in which slices were exposed to one or two 30 nM CORT pulses withan interval of 60 min. The second CORT pulse between brackets was appliedfor only the group receiving two CORT pulses (30–30 nM CORT). HFS wasapplied 120 min after the start of the first pulse (vehicle and 30 nM CORT,n = 8; 30–30 nM CORT, n = 6). (E) Variations in the duration of the intervalbetween the two 100-nM CORT pulses. Normalization of LTP was seen with60- and 90-min intervals but not with shorter (10 or 30 min) intervals be-tween the two CORT pulses. (Upper) Schematic representation of the ex-perimental protocol in which the delay between application of the two100-nM CORT pulses was varied between 10 and 90 min. In all cases, HFS wasapplied 120 after the start of the first pulse. Vehicle and 60 min delay group(100–100 nM CORT), n = 8; all other groups, n = 6. Values indicate groupmeans ± SEM (***P < 0.001, **P < 0.01, and *P < 0.05).
14268 | www.pnas.org/cgi/doi/10.1073/pnas.1411216111 Sarabdjitsingh et al.
Dow
nloa
ded
by g
uest
on
Apr
il 14
, 202
0
CA1 area at a stable level, preventing a “fixed” situation thatmay arise after a single pulse. This maintenance is in line withthe observation that pulsatile CORT release is important tomaintain transcriptional activity via GRs at an optimal level (4–8).If so, pulsatile CORT exposure would prepare the organism ina flexible manner to upcoming challenges at the start of the activeperiod of the day.
Materials and MethodsDrugs. CORT (30 or 100 nM; first dissolved in 90% (vol/vol) ethanol and nextdiluted to the final concentration, containing a maximum of 0.009% etha-nol) was administered in pulses of 10 min, which effectively cause an in-creasing concentration during this period, followed by a slow decrease in thesubsequent 10-min period (Fig. S3). This paradigm was previously applied tomimic ultradian pulses in vitro (6). To determine the receptor involved, weapplied the selective MR antagonist spironolactone (100 nM; first dissolvedin chloroform and next diluted to the final concentration, containing0.002% chloroform) or the GR antagonist mifepristone (500 nM; in H2O towhich 5 N HCl was added in an amount just sufficient to dissolve the antag-onist, and then diluted to the final concentration). All drugs were obtainedfrom Sigma-Aldrich. To study the involvement of putative membrane-boundcorticosteroid receptors, we used CORT–BSA conjugate (100 nM; first dissolvedin H2O and then diluted to the final concentration; Sigma-Aldrich).
Hippocampal Cell Culture, Protein Expression, Immunocytochemistry, and cLTP.Cultures of hippocampal neurons were prepared from embryonic day 18Sprague–Dawley rats following a previously described method (20). Briefly, cellswere plated at a density of 200–300 × 103 cells per dish on poly-lysine–precoatedcoverslips. Mixed cultures of neurons and glial cells were layered on cover-slips and maintained in a 3% (vol/vol) serum containing Neurobasal medium.This medium was replaced after 4 d in vitro by a serum-free Neurobasalmedium, and cultures were kept at 37 °C in 5% (vol/vol) CO2.
Pulses of CORT (20 min total; 10 min exposure and 10 min washout;comparable dynamics and kinetics as shown in Fig. S3) were applied to theneuronal coverslips through a peristaltic pump (2 mL/min for CORT pulse,1 mL/min between pulses). For live immunocytochemical staining, neuronswere transfected with GluA1–super ecliptic pHluorin at 8–10 d in vitro byusing the Effecten transfection kit (Qiagen) following the provider’s pro-tocol. Surface GluA1–AMPARs were specifically stained by using a mono-clonal anti-GFP antibody (1:500; Roche) for 15 min on live neurons at 37 °C inculture medium. Briefly, neurons were then fixed with 4% (wt/vol) para-formaldehyde/4% (wt/vol) sucrose for 15 min, washed, and incubated withsecondary anti-mouse Alexa 488 antibodies (1:600, 30 min). To label post-synaptic areas, neurons were permeabilized by using 0.1% Triton X-100,incubated with a primary guinea pig polyclonal anti-Homer 1c antibody(1:500, 30 min; Synaptic Systems) or shank antibody (rabbit polyclonal an-tibody; 1:1,000, 1 h; Abcam), and finally incubated with secondary anti-guinea pig Alexa 568 antibodies (1:600, 30 min; Molecular Probes) or anti-rabbit Alexa 488 antibodies (1:600, 30 min; Molecular Probes). Neurons werewashed, mounted, and preparations were kept at 4 °C until observation. Forthe quantification of surface AMPAR staining within individual synapses,Homer 1c staining served as a mask filter to isolate surface GluA1 subunitstaining in individual Homer 1c clusters. The integrated fluorescence levelover the Homer 1c cluster area was then measured for each cluster. Thefluorescence analysis was performed by using imaging tools from Meta-morph software (Universal Imaging), as previously described (21–23). Thechemically induced potentiation protocol consisted of a bath coapplicationof glycine (200 μM) and picrotoxin (5 μM) for 4 min. In all live experiments,cLTP was always applied after a period of baseline acquisition and the me-dium was carefully replaced by fresh equilibrated and heated mediumafter induction.
Single Quantum Dot Tracking. Single particle detection and imaging wereperformed as previously described in detail (24, 25). In-depth information isgiven in SI Materials and Methods.
Animals. Male C57BL/6 mice (∼7–8 wk of age at arrival; Harlan) were group-housed at a 12-h light/dark schedule (lights on at 0700 hours) with food andwater provided ad libitum. After an acclimatization period of ∼1–2 wk, mice(one at a time) entered the experiment. Mice were decapitated early in themorning when endogenously circulating plasma levels are still low. Allexperiments were approved by the animal ethical commission of UtrechtUniversity, and all efforts were made to minimize suffering of the animals.
Fig. 5. Normalization of LTP depends on nongenomic GR-dependent path-way. (A) Averaged mean values during the 60-min posttetanic recording pe-riod. Application of one 100 nM CORT pulse at the moment of the secondCORT pulse (bar with white dashed stripes) still attenuated LTP comparedwith vehicle-treated slices (open bar) in a similar way as 120 min after a singleCORT pulse (gray bar). HFS applied 120 min after the onset of the second CORTpulse (at t 180min) very effectively induced LTP (bar with black dashed stripes).(Upper) Experimental paradigm. All groups, n = 8; second pulse CORT group,n = 6; 2 h delay, n = 5. (B) The presence of spironolactone during the secondCORT administration (black dashed bar) did not affect synaptic potentiation,whereas mifepristone (gray dashed bar, far right) significantly attenuated thelevel of LTP. (Upper) Schematic representation of the experimental protocol.Coincubation with 100 nM spironolactone or 500 nM mifepristone started20 min before the application of the second 100-nM CORT pulse and coter-minated with the second pulse. HFS was applied 120 min after the start of thefirst pulse. Spironolactone, n = 7; all other groups, n = 8. (C) When CORT–BSA(dashed black bar, far right) was delivered during the second pulse instead ofCORT (black bar), synaptic plasticity was normalized, and fully comparable towhat was seen in slices treated only with vehicle (open bar). A single pulse ofCORT–BSA did not affect synaptic plasticity (dashed open bar), different fromthe reduction seen with a single CORT pulse (gray bar). (Upper) CORT–BSA wasapplied as a single pulse (100 nM CORT–BSA) or as a second pulse after a CORTpulse (second pulse CORT–BSA). HFS was applied 120 min after the start of thefirst pulse. Second pulse CORT-BSA and 100 nM CORT–BSA, n = 7; all othergroups, n = 8. (D) Two consecutive pulses of 100 nM CORT–BSA (dashed blackbar) did not significantly increase LTP levels compared with vehicle-treatedslices (open bar). (Upper) Two pulses of 100 nM CORT–BSA were administeredwith an interval of 60 min. HFS was applied 120 min after the start of the firstpulse (100–100 nM CORT–BSA, n = 7; all other groups, n = 8). Values indicategroup means ± SEM (*P < 0.05, **P < 0.01, and ***P < 0.001). Note that thevehicle, single-pulse CORT, and double-pulse CORT groups shown in this figureare based on the same data as Fig. 4 to allow comparisons.
Sarabdjitsingh et al. PNAS | September 30, 2014 | vol. 111 | no. 39 | 14269
NEU
ROSC
IENCE
Dow
nloa
ded
by g
uest
on
Apr
il 14
, 202
0

Electrophysiology. Early in the morning, the mouse was decapitated and thebrain was rapidly dissected and placed in ice-cold artificial cerebrospinal fluid(aCSF) containing (in mM) 120 NaCl, 3.5 KCl, 1.3 MgSO4, 1.25 NaH2PO4,2.5 CaCl2, 10 glucose, and 25 NaHCO3, and continuously gassed [mixture of 95%(vol/vol) O2/5% (vol/vol) CO2]. Next, dorsal hippocampal slices (350 μm) weremade by using a Vibratome (VT 1000S; Leica) and stored in aCSF at roomtemperature for >1 h before recording commenced at a bath temperature of30–32 °C (16).
For patch-clamp electrophysiology, one slice at a time was transferred tothe recording chamber mounted on an upright microscope (Axioskop 2 FSplus; Zeiss) with differential interference contrast and a water immersionobjective (×40) to identify the cells. Slices were continuously perfused (flowrate, 1.5 mL/min) with warm aCSF (temperature, 30 °C; pH 7.4) containingTTX (0.5 μM; Latoxan) to block sodium channels and bicuculline (50 μM;Enzo) to block GABAa receptors. Slices were exposed to a single 10-min CORT(100 nM; or vehicle as control) pulse or two consecutive pulses (of CORT orvehicle) with a 60-min interval. Unless stated otherwise, all recordings wereperformed ∼2 h after onset of the first pulse.
Patch pipettes (borosilicate glass pipettes; inner diameter, 0.86 mm; outerdiameter, 1.5 mm; Hilgenberg) were pulled on a Sutter micropipettepuller (resistance; 3–6 MΩ).The intracellular solution contained (in mM):120 Cs methane sulfonate, 17.5 CsCl, 10 Hepes, 2 MgATP, 0.1 NaGTP, 51,2-bis(o-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid (BAPTA), pH 7.4,adjusted with CsOH. BAPTA was obtained from Molecular Probes, and allother chemicals were obtained from Sigma-Aldrich. An Axopatch 200Bamplifier (Axon Instruments) was used for whole-cell recordings, operatingin the voltage-clamp mode. The patch-clamp amplifier was interfaced toa computer via a Digidata (type 1200; Axon Instruments) analog-to-digitalconverter. Data acquisition and storage was done by using PClamp(version 9.2).
Holding potential was −70 mV. The liquid junction potential caused a shiftof 8 mV at most, for which we did not compensate. Recordings with anuncompensated series resistance of <2.5 times the pipette resistance wereaccepted for analysis. In view of the small current amplitudes, the recordingswere not corrected for series resistance. Currents were identified as mEPSCswhen the rise time was faster than the decay time; mEPSC frequency andpeak amplitude were determined.
Field excitatory postsynaptic potentials (fEPSPs) were recorded in theSchaffer collateral-CA1 pathway of coronal mouse brain slices as describedpreviously (26). Briefly, a bipolar stimulation electrode (60 μm stainless-steelwires insulated except for the tip) was placed on the Schaffer collaterals, and
glass recording pipettes (filled with aCSF; 2–5-MΩ impedance) were posi-tioned in the CA1 stratum radiatum. At the start of each experiment, aninput–output curve was established to record the slope of the fEPSP, fromwhich maximal and half-maximal slope as well as the corresponding maxi-mal and half-maximal stimulus intensity were determined. The half-maximalstimulus intensity that was calculated was used throughout the remainderof the recording session. For each experimental group, baseline synaptictransmission was recorded with a frequency of 0.033 Hz (0.15 ms) for 10 min.Thereafter, repetitive tetanic stimulations (10 Hz; 90 s) were applied, afterwhich recordings proceeded for another 60 min at a frequency of 0.033 Hz;this stimulation paradigm is very sensitive to the effects of CORT (26). Twoconsecutive traces were averaged to represent the mean per minute. Datawere acquired, stored, and analyzed by using Signal 2.16 (Cambridge 159Electronic Design).
Data Analysis. Because data of diffusion coefficients are not normally dis-tributed, comparisons between groups for instantaneous diffusion coef-ficients were performed by using a Mann–Whitney test (pair comparison) orKruskal–Wallis test followed by Dunn multiple comparison test (groupcomparison). All other group comparisons for live cell imaging or single-cell electrophysiology were performed by using parametric statistical tests,Student t test (unpaired or paired comparison as appropriate), ANOVAfollowed by Newman–Keuls multiple comparison test (group comparison),or Kolmogorov–Smirnov test (distribution comparison). Significance levelswere defined at P < 0.05, P < 0.01, or P < 0.001.
For mEPSC properties, data for both series (a single CORT/vehicle pulse ortwo CORT/vehicle pulses) were compared with an unpaired Student t test. Forthe fEPSP recordings, all statistical analyses were done with SPSS 21.0. Groupvalues are expressed as mean ± SEM. A one-way ANOVA was used to assessgroup differences in the level of potentiation. Where applicable, Tukey posthoc tests were used. Student paired t tests was used to compare within-group baseline values with the level of LTP. P values <0.05 were consideredto indicate a significant difference.
ACKNOWLEDGMENTS. This work was supported by Netherlands Organiza-tion for Scientific Research Grants 863.13.021 (to R.A.S.), 817.02.017 (to H.K.),and 024.001.003 (to M.J.); by travel grants from Federation of EuropeanNeuroscience Societies/Network of European Neuroscience Schools (to N.P.and A.K.); and by Centre National de la Recherche Scientifique, ConseilRégional d’Aquitaine, Agence National de la Recherche (ANR-JC08-329238)and France BioImaging (L.G.).
1. de Kloet ER, Joëls M, Holsboer F (2005) Stress and the brain: From adaptation todisease. Nat Rev Neurosci 6(6):463–475.
2. Oakley RH, Cidlowski JA (2011) Cellular processing of the glucocorticoid receptorgene and protein: New mechanisms for generating tissue-specific actions of gluco-corticoids. J Biol Chem 286(5):3177–3184.
3. Ulrich-Lai YM, Herman JP (2009) Neural regulation of endocrine and autonomic stressresponses. Nat Rev Neurosci 10(6):397–409.
4. Lightman SL, Conway-Campbell BL (2010) The crucial role of pulsatile activity of theHPA axis for continuous dynamic equilibration. Nat Rev Neurosci 11(10):710–718.
5. Qian X, Droste SK, Lightman SL, Reul JM, Linthorst AC (2012) Circadian and ultradianrhythms of free glucocorticoid hormone are highly synchronized between the blood,the subcutaneous tissue, and the brain. Endocrinology 153(9):4346–4353.
6. Stavreva DA, et al. (2009) Ultradian hormone stimulation induces glucocorticoidreceptor-mediated pulses of gene transcription. Nat Cell Biol 11(9):1093–1102.
7. Sarabdjitsingh RA, et al. (2010) Disrupted corticosterone pulsatile patterns attenuateresponsiveness to glucocorticoid signaling in rat brain. Endocrinology 151(3):1177–1186.
8. Conway-Campbell BL, Pooley JR, Hager GL, Lightman SL (2012) Molecular dynamics ofultradian glucocorticoid receptor action. Mol Cell Endocrinol 348(2):383–393.
9. Joëls M, Sarabdjitsingh RA, Karst H (2012) Unraveling the time domains of cortico-steroid hormone influences on brain activity: Rapid, slow, and chronic modes. Phar-macol Rev 64(4):901–938.
10. Groc L, Choquet D, Chaouloff F (2008) The stress hormone corticosterone conditionsAMPAR surface trafficking and synaptic potentiation. Nat Neurosci 11(8):868–870.
11. Karst H, Joëls M (2005) Corticosterone slowly enhances miniature excitatory post-synaptic current amplitude in mice CA1 hippocampal cells. J Neurophysiol 94(5):3479–3486.
12. Diamond DM, Park CR, Woodson JC (2004) Stress generates emotional memories andretrograde amnesia by inducing an endogenous form of hippocampal LTP. Hippo-campus 14(3):281–291.
13. Krugers HJ, Hoogenraad CC, Groc L (2010) Stress hormones and AMPA receptortrafficking in synaptic plasticity and memory. Nat Rev Neurosci 11(10):675–681.
14. Kim JJ, Diamond DM (2002) The stressed hippocampus, synaptic plasticity and lost
memories. Nat Rev Neurosci 3(6):453–462.15. Huganir RL, Nicoll RA (2013) AMPARs and synaptic plasticity: The last 25 years. Neuron
80(3):704–717.16. Karst H, et al. (2005) Mineralocorticoid receptors are indispensable for nongenomic
modulation of hippocampal glutamate transmission by corticosterone. Proc Natl Acad
Sci USA 102(52):19204–19207.17. Tasker JG, Di S, Malcher-Lopes R (2006) Minireview: Rapid glucocorticoid signaling via
membrane-associated receptors. Endocrinology 147(12):5549–5556.18. Karst H, Berger S, Erdmann G, Schütz G, Joëls M (2010) Metaplasticity of amygdalar
responses to the stress hormone corticosterone. Proc Natl Acad Sci USA 107(32):
14449–14454.19. Droste SK, et al. (2008) Corticosterone levels in the brain show a distinct ultradian
rhythm but a delayed response to forced swim stress. Endocrinology 149(7):
3244–3253.20. Mikasova L, et al. (2012) Disrupted surface cross-talk between NMDA and Ephrin-B2
receptors in anti-NMDA encephalitis. Brain 135(pt 5):1606–1621.21. Park M, Penick EC, Edwards JG, Kauer JA, Ehlers MD (2004) Recycling endosomes
supply AMPA receptors for LTP. Science 305(5692):1972–1975.22. Petrini EM, et al. (2009) Endocytic trafficking and recycling maintain a pool of mobile
surface AMPA receptors required for synaptic potentiation. Neuron 63(1):92–105.23. Wang Z, et al. (2008) Myosin Vb mobilizes recycling endosomes and AMPA receptors
for postsynaptic plasticity. Cell 135(3):535–548.24. Groc L, Choquet D (2008) Measurement and characteristics of neurotransmitter re-
ceptor surface trafficking (review). Mol Membr Biol 25(4):344–352.25. Groc L, et al. (2007) Surface trafficking of neurotransmitter receptor: Comparison
between single-molecule/quantum dot strategies. J Neurosci 27(46):12433–12437.26. Wiegert O, Joëls M, Krugers H (2006) Timing is essential for rapid effects of cortico-
sterone on synaptic potentiation in the mouse hippocampus. Learn Mem 13(2):
110–113.
14270 | www.pnas.org/cgi/doi/10.1073/pnas.1411216111 Sarabdjitsingh et al.
Dow
nloa
ded
by g
uest
on
Apr
il 14
, 202
0


















