T1 - Glicólise e Oxidação do Piruvato
6
Click here to load reader
Transcript of T1 - Glicólise e Oxidação do Piruvato

8/19/2019 T1 - Glicólise e Oxidação do Piruvato
http://slidepdf.com/reader/full/t1-glicolise-e-oxidacao-do-piruvato 1/6
Ana Clara Oliveira e Andreia Preda 2013/2014
Glicólise e Oxidação do Piruvato
A glicose, quando entra dentro das células, entra através de um transportador e vocês
já viram, no semestre passado, como é que a glicose entrava dentro das células do epitélio
intestinal. Quer para entrar quer para sair, a glicose atravessa a membrana através de um
transportador. Para ter a certeza que a glicose não volta a sair das células, ela tem que ser
fosforilada, porque quando a glicose está fosforilada, não atravessa a membrana, não passa
pelos transportadores. Senão o que é que acontecia? A glicose entrava dentro da célula, se
acumulasse muita glicose aqui, ela tenderia a sair porque estes transportadores são a favor do
gradiente de concentração. Depois, a glicose fosforilada pode entrar em várias em vias. Vamos
considerar só a glicólise. A glicose é transformada em piruvato e, conforme foram as condições
de estado redox ou a maior ou menor quantidade de oxigénio ou o facto de a cadeia
transportadora de eletrões funcionar mais ou menos, o piruvato pode ter dois destinos: pode
ser convertido em lactato ou entrar dentro da mitocôndria, sendo convertido em acetil-CoA,
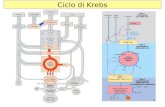
que pode entrar no ciclo de Krebs, havendo depois a produção de NADH ou , os
eletrões passam para a cadeia de eletrões com a produção de ATP, o que significa que a glicose
é totalmente oxidada a dióxido de carbono e água.
Hoje vamos falar detalhadamente sobre a transição de glicose a piruvato e de piruvato
a lactato ou a acetil-CoA. Adicionalmente, o acetil-CoA pode ser utilizado para a síntese de
ácidos gordos.Agora vocês têem aqui as várias vias ou os vários destinos da glicose. A glicose entra
para dentro das células, é fosforilada e, depois, a glicose-6-fosfato, se for convertida em
glicose ou fosfato, pode ser armazenada na forma de glicogénio, pode ser transformada em
frutose-6-fosfato, ou a glicose-6-fosfato pode ser transformada em 6-fosfogliconato.
Na passagem de glicose a glicose-6-fosfato, as enzimas que fazem esta fosforilação não
são específicas da glicólise porque se se inibir a passagem de glicose a glicose-6-fosfato, não
se estará só a inibir a glicólise. Inibe-se também, por exemplo, a síntese de glicogénio ou a
produção da ribose-5-fosfato.
Para termos mais ou menos glicose-6- fostato dentro das células, ela poderá vir da
alimentação, da degradação do glicogénio ou então da gliconeogénese, que é a síntese de
glicose a partir de substratos que não são glícidos.
Numa situação de anaerobiose, o piruvato pode ser convertido em lactato, em alanina
ou a vários outros aminoácidos, em oxalo-acetato e daqui começa uma das possibilidades da
gliconeogénese, em acetil-CoA, que pode ser utilizado na síntese de ácidos gordos ou, se
entrar no ciclo de Krebs, leva à oxidação completa da glicose.

8/19/2019 T1 - Glicólise e Oxidação do Piruvato
http://slidepdf.com/reader/full/t1-glicolise-e-oxidacao-do-piruvato 2/6
Ana Clara Oliveira e Andreia Preda 2013/2014
Dentro das vias associadas ao metabolismo dos glícidos, temos a glicólise. Todas as
reações da glicólise ocorrem no citoplasma e são, ao todo, dez. Primeiro, a glicose é convertida
em glicose-6-fosfato. A principal função desta reação é o impedimento da saída da glicose das
células, bem como a ativação da glicose de modo que esta possa entrar numa via metabólica.
Em todas as vias metabólicas, as moléculas que nelas participam são, de alguma forma,
ativadas. Esta ativação pode ocorrer, por exemplo, por fosforilação, por adição da coenzima A,
por ligação a um nucletídeo. As moléculas são sempre, de algum modo, ativadas e, neste caso,
a glicose é ativada por fosforilação. Depois, a glicose-6-fosfato é transformada em frutose-6-
fostato por uma isomerase, ou seja, isto é uma isomerização. Neste caso, é a isomerização de
uma aldose (glicose) em cetose (frutose). De seguida, a frutose-6-fosfato vai sofrer nova
fosforilação, passando assim a frutose-1,6-difosfato, que vai posteriormente ser quebrada ao
meio, formando duas trioses, cada uma monofosfatada. Uma das trioses é o gliceraldeído-3-
fosfato e a outra é a dihidroxiacetona-fosfato. A dihidroxiacetona é convertida em
gliceraldeído-3-fosfato, pela fosfotriose isomerase, portanto, daqui para a frente é sempre a
dobrar. O gliceraldeído-3-fosfato é convertido em 1,3-difosfoglicerato, depois em 3-
fosfoglicerato, 2-fosfoglicerato, fosfoenolpiruvato e, finalmente, (enol)piruvato. Além da
glicose, para que a glicólise possa ocorrer, é ainda necessário ADP, ATP e NAD +. Para além
disso, a glicose-6-fosfato, a frutose-6-fosfato, a frutose-1,6-difosfato e o gliceraldeído-3-fosfato
são também intermediários da Via das Pentoses-fosfato.
As reações onde vai ocorrer gasto de ATP correspondem àquelas em que vai haver um
investimento na ativação das moléculas para que as reações a seguir se possam dar. Temos,
depois, uma molécula em que, ao ser quebrada em duas, cada uma das partes já se vai
encontrar fosforilada, logo já está ativada.
O NAD é um transportador de equivalentes redutores e, quando está na forma de
NAD+, encontra-se oxidado e, na forma de NADH, encontra-se reduzido.
Então, na glicólise, vai-se produzir NADH, a glicose é oxidada e vai-se gastar NAD +, e é
nesta mesma fase, em que se reduz o NADH, que se produz o ATP. Então, existe uma parte em
que se investe ATP, outra em que a molécula é quebrada em duas e há depois uma
interconversão, e, por último, uma em que é produzida energia. Esta secção da glicólise em
que há produção de energia é que é a secção em que há produção de ATP e de NADH.
Voltando agora à primeira reação, esta é uma reação considerada, em termos
fisiológicos, irreversível. Por isso, a reação ocorre sempre no sentido da glicose para a glicose-
6-fosfato. Em quase todas estas reações da glicólise, ou as enzimas em si precisam de
magnésio ou o ATP e o ADP, para reagirem, precisam de estar complexados com o magnésio.O que esta enzima, a hexocínase, tem de característico é o facto de ser inibida pelo seu

8/19/2019 T1 - Glicólise e Oxidação do Piruvato
http://slidepdf.com/reader/full/t1-glicolise-e-oxidacao-do-piruvato 3/6
Ana Clara Oliveira e Andreia Preda 2013/2014
produto, o que significa que quando a célula tem no seu interior grande quantidade de glicose-
6-fosfato, vai inibir a fosforilação da glicose em glicose-6-fosfato. Se este mecanismo não
existisse, quando houvesse um excedente de glicose na célula, ela voltaria a sair.
No entanto, este mecanismo de feedback negativo não é compatível com o
funcionamento do fígado. Por exemplo, se se comer muitos chocolates sucessivamente, o
fígado tem de ter a capacidade de retirar a glicose em excesso da corrente sanguínea. Por
outro lado, em jejum, o fígado tem de ser capaz de repôr e manter os níveis basais de glicose
no sangue. Ou seja, o facto de haver uma enzima que é inibida pelo seu produto não é
co 1mpatível com o funcionamento do fígado.
No fígado e nas células pancreáticas, entre outras, a entrada da glicose é independente
da insulina. No tecido adiposo, a entrada da glicose depende da insulina. Isto significa que a
expressão dos transportadores na membrana aumenta quando aumenta a insulina na corrente
sanguínea. No fígado e nas células pancreáticas isto não acontece. O músculo e o tecido
adiposo só expressam a hexocínase, enquanto que o fígado e as células pancreáticas
expressam a hexocínase e a glicocínase. 1 O que acontece é que para as concentrações normais
de glicose, a hexocínase já está a trabalhar perto da sua velocidade máxima e a glicocínase
não. Das duas isoformas, a que tem maior afinidade para a glicose é a que tem um K m menor,
logo é a hexocínase. Deste modo, a glicocínase no fígado vai continuar a permitir a retirada de
glicose da corrente sanguínea, uma vez que esta enzima, ao contrário da hexocínase, não é
inibida pela glicose-6-fosfato. A regulação da glicocínase faz-se pela quantidade de glicose na
corrente sanguínea: quando esta é elevada, a enzima é ativada; quando a glicemia é baixa, a
enzima é inibida.
Prosseguindo agora na glicólise, a glicose-6-fosfato sofre uma isomerização em
frutose-6-fosfato e, de seguida, a segunda fosforilação em frutose-1,6-difosfato, sendo gastas,
até esta etapa, duas moléculas de ATP. Depois, havia a quebra de frutose-1,6-difosfato em
gliceraldeído-3-fosfato e dihidroxiacetona-fosfato e a conversão desta no primeiro produto,
ficando-se assim com duas moléculas de gliceraldeído-3-fosfato. Cada um dos gliceraldeídos
vai ser fosforilado a 1,3-fosfoglicerato e, depois, a passagem deste a 3-fosfoglicerato com a
produção de duas moléculas de ATP (uma por cada molécula de gliceraldeído-3-fosfato).
Então, até esta etapa, já houve compensação das moléculas de ATP que foram gastas para
ativar esta via. Até este momento, houve também a produção de duas moléculas de NADH.
1 Alguns autores consideram apenas hexocínases, enumerando-as para as diferenciar. Nestes casos, a glicocínase
corresponde à hexocínase-4.

8/19/2019 T1 - Glicólise e Oxidação do Piruvato
http://slidepdf.com/reader/full/t1-glicolise-e-oxidacao-do-piruvato 4/6
Ana Clara Oliveira e Andreia Preda 2013/2014
Depois, há a passagem de 3-fosfoglicerato a 2-fosfoglicerato, de seguida, a fosfoenolpiruvato
e, finalmente, a piruvato (ou ácido pirúvico), com a produção de mais duas moléculas de ATP.
Em termos de rendimento energético, há a produção de duas moléculas de ATP, e em
termos de equivalentes redutores, são produzidas duas moléculas de NADH.
O ATP é produzido na fosforilação oxidativa, em função do funcionamento da cadeia
transportadora de eletrões, na glicólise ou em consequência de alguma reações do ciclo de
Krebs. Ou seja, o ATP pode ser formado quer por fosforilação nível do substrato, quer dizer
que as reações não estão associadas à cadeia transportadora de eletrões nem à síntase de
ATP, quer por fosforilação oxidativa, onde vai ser consumido oxigénio que depois é convertido
em água.
Ao longo da glicólise ocorrem duas reações que podem ser chamadas fosforilações ao
nível do substrato. No caso da glicólise a produção de ATP pode ocorrer em situações de
anaerobiose.
As reações que ocorrem nos dois sentidos representam reações reversíveis e são as
mesmas da gliconeogénese. Quando as setas na reação aparecem apenas num sentido
significa que a reação é fisiologicamente irreversível e quando se pretende produzir glicose há
enzimas que fazem exactamente a ação contrária a estas enzimas.
Se todas as enzimas tivessem ação reversível não era possível inibir a glicólise sem
inibir a gliconeogénese. Assim, as enzimas de ação irreversível são os pontos de regulação das
vias.
O primeiro ponto especifico de regulação da glicólise é a cínase 1 da frutose-6-fosfato
que catalisa a reação frutose-6-fosfato + ATP frutose-1,6-bisfosfato + ADP uma vez que a
hexocinase não é especifica da glicólise.
A enzima desidrogenase do gliceraldeido 3-fosfato tem vários resíduos de cisteína e a
cisteína na extremidade da cadeira tem grupos tiol e um destes grupos está no centro ativo
tendo a vantagem que o substrato se liga covalentemente. Os equivalentes redutores que se
perdem são captados pelo NAD que passa de NAD+ a NADH, ocorrendo uma oxidação do
substrato e uma oxidação do NAD. O NAD na sua forma oxidada está associado à enzima mais
fortemente.
A fosforólise (muito importante no metabolismo do glicogénio) é semelhante a uma
hidrólise mas há quebra de uma ligação por entrada de um fosfato. O grupo carbonilo que
antes está ligado à enzima passa a estar ligado ao ácido fosfórico; assim o fosfato serve para
quebrar a ligação do substrato que entretanto foi oxidado à enzima.
3 enzimas são responsáveis pela regulação da glicólise mas apenas 2 são específicaspata a glicólise.

8/19/2019 T1 - Glicólise e Oxidação do Piruvato
http://slidepdf.com/reader/full/t1-glicolise-e-oxidacao-do-piruvato 5/6
Ana Clara Oliveira e Andreia Preda 2013/2014
A passagem do 1,3-bifosfoglicerato para o 3-fosfoglicerato dá-se pela ação de uma
cínase mas nos eritrócitos pode haver um mecanismo alternativo para esta passagem. Pode
ser uma mútase que passa o 1,3-bifosfoglicerato para 2,3-bifosfoglicerato e depois temos uma
fosfatase que tira o fosfato da posição 2 e passa a ser 3-fosfoglicerato.
Em termos de contas para a glicólise, no processo de glicólise normal há nesta fase
produção de 2 ATP que dá para compensar os ATP gastos, enquanto que nesta via alternativa o
rendimento da glicólise nos eritrócitos poderá ser 0 (não quer dizer que seja sempre). A
vantagem para os eritrócitos desta via alternativa é que o 2,3-bifosfoglicerato diminui a
afinidade da hemoglobina para o oxigénio ajudando a libertar o oxigénio da hemoglobina. Esta
via não é a predominante nos eritrócitos porque, uma vez que os eritrócitos não têm
mitocôndrias não podem obter ATP pela cadeia transportadora de eletrões e então a produção
de ATP pela glicólise “normal” é fundamental.
Se os eritrócitos não têm mitocôndrias eles não conseguem transformar o NADH a
NAD+, ou seja a passagem de NAD reduzido a NAD oxidado não ocorre, mas tem de ocorrer
porque senão os eritrócitos deixam de produzir ATP. Este problema resolve-se com a
passagem de piruvato a lactato, em que o NADH é oxidado. Isto tem sempre de ocorrer nos
eritrócitos e também ocorre noutras células onde não haja mitocôndrias ou haja algum
problema no ciclo de Krebs ou no aceitador final (O 2).
Recapitulando: da glicose até ao piruvato existem reações reversíveis e irreversíveis (
na aula de gliconeogénese, vamos ter a passagem de piruvato entre outros substratos a glicose
e as reações reversíveis vão ser as mesmas e as reação irreversíveis não vão ser as mesmas, o
que significa que se se pretender ter a glicólise muito ativa ou a gliconeogénese mito inibida
ou ao contrário, teremos de simultaneamente estar a ativar as enzimas irreversíveis da
glicólise e a inibir as enzimas irreversíveis da gliconeogénese).
Agora vamos ver como é que o piruvato pode ser oxidado a acetilCOA.
A passagem de piruvato a lactato acontece em tecidos que não há mitocôndrias ou
quando a disponibilidade de oxigénio não acompanha a produção de piruvato. Mas o piruvato
pode ser oxidado a aceitlCOA para depois entrar no ciclo de krebs.
O piruvato é produzido no citoplasma e depois entra por um transportador na
mitocôndria onde é oxidado. A enzima é um complexo enzimático e inicialmente o piruvato é
descarboxilado e o que “sobra” da descarboxilação vai f icar ligado à tiamina difosfato. Este
complexo enzimático (estas três atividades enzimáticas distintas) ao conjunto todo chama-se
desidrogenase do piruvato que é também o nome da primeira enzima que catalisa a primeira
reação de descarboxilação.

8/19/2019 T1 - Glicólise e Oxidação do Piruvato
http://slidepdf.com/reader/full/t1-glicolise-e-oxidacao-do-piruvato 6/6
Ana Clara Oliveira e Andreia Preda 2013/2014
O piruvato é descarboxilado, fica ligado à tiamina difosfato e depois a tiamina difosfato
vai transferir o resto do piruvato para a alipoamida oxidada.
As moléculas podem ser ativadas por fosforilação, por adição da coenzima A, por
ligação a um nucletídeo, como foi referido anteriormente, então o que sobra da
descarboxilação do piruvato liga-se agora à coenzima A para depois poder entrar no ciclo de
krebs. A ligação à coenzima A dá-se no segundo componente que é diidrolipoil transacetilase.
Os átomos de enxofre passam a estar reduzidos, ou seja, ligados a hidrogénios em vez de
estarem ligados um outro e os eletrões e protões passam para o NADH. A vantagem de termos
um complexo enzimático e dos intermediários estarem sempre ligados às enzimas é que não
se perdem pelo caminho tornando a reação muito mais eficiente.
Então, o piruvato é descarboxilado e é uma descarboxilação oxidativa porque para
além de haver uma descarboxilação também vai haver uma redução do NADH e oxidação do
substrato.
Produz-se NADH e acetilCOA o que significa que se houver muita acetilCOA e muito
NaDH este complexo enzimático vai ser inibido. O NADH e a acetilCOA inibem a conversão de
piruvato a acetilCOA.
As enzimas podem ser reguladas por alosteria. Este complexo enzimático pode ser
regulado por fosforilação e desfosforilação. Ela quando está desfosforilada está na sua forma
ativa e quando está fosforilada está na sua forma inativa.
O NADH e acetilCOA além de inibirem diretamente a enzima por alosteria também a
inibem indiretamente porque vão ativar a cínase que vai fosforilar a enzima (quando está
fosforilada está inativa).