9.2 La piruvato deidrogenasi è strettamente...
3
La piruvato deidrogenasi è strettamente controllata La decarbossilazione ossidativa degli - chetoacidi, di cui i tre metabolicamente più importanti sono il piruvato, l’ossalacetato e l’-chetoglutarato, è una reazione irre- versibile di fondamentale importanza bio- chimica che avviene entro i mitocondri. La decarbossilazione del piruvato, prodotto dal metabolismo dei carboidrati e, in mi- sura minore, di molti amminoacidi, produ- ce acetil CoA utilizzato a scopi energetici nel ciclo dell’acido citrico oppure per la sin- tesi di acidi grassi; pertanto questa reazio- ne rappresenta una fondamentale cernie- ra tra il metabolismo di carboidrati e am- minoacidi e quello dei lipidi. L’irreversibi- lità della reazione giustifica il fatto ben noto che la maggior parte degli organismi non possono produrre carboidrati da lipidi. In- vece, la decarbossilazione dell’-chetoglu- tarato trasforma quest’ultimo in succinil CoA in una delle reazioni del ciclo dell’aci- do citrico. Come esempio di decarbossilazione os- sidativa degli -chetoacidi prenderemo in esame la decarbossilazione del piruvato ca- talizzata dalla piruvato deidrogenasi. La reazione in realtà comprende sia una de- carbossilazione sia un’ossidazione a carico del substrato; essa pertanto richiede l’in- tervento di un complesso multienzimatico costituito da tre differenti tipi di subunità catalitiche e da cinque diversi cofattori (ta- bella; figura 1) che catalizzano la seguente reazione complessiva: Piruvato NAD CoA-SH 8n acetil CoA AMP PPi I cinque cofattori sono NAD , FAD, tiam- minapirofosfato (TPP, la forma attivata del- la vitamina B1, o tiamina), CoA e acido li- poico; quest’ultimo è un acido grasso a otto atomi di carbonio contenente due gruppi SCHEDA 9.2 Stefani-Taddei Percorsi di biochimica © Zanichelli 2011 1 50 nm Figura 1 Fotografia al microscopio elettronico mostrante la composizione in subunità della piruvato deidrogenasi di Escherichia coli. Tabella Funzione dei coenzimi e dei gruppi prostetici della reazione della piruvato deidrogenasi Coenzimi e gruppi prostetici Localizzazione Funzione Tiammina pirofosfato Legata alla piruvato Reagisce con il substrato, deidrogenasi piruvato Acido lipoico Covalentemente legato Accetta il gruppo acetile a un residuo di lisina dalla tiammina pirofosfato sulla diidrolipoil transacetilasi Coenzima A Libero in soluzione Accetta il gruppo acetile dalla lipoammide sulla transacetilasi Flavina adenin Fortemente legata alla Accetta equivalenti dinucleotide (FAD) diidrolipoil deidrogenasi dalla lipoammide ridotta Nicotinammide Libera in soluzione Accettore terminale degli adenin dinucleotide equivalenti riducenti dalla flavoproteina ridotta
Transcript of 9.2 La piruvato deidrogenasi è strettamente...

La piruvato deidrogenasi è strettamente controllataLa decarbossilazione ossidativa degli �-chetoacidi, di cui i tre metabolicamente piùimportanti sono il piruvato, l’ossalacetatoe l’�-chetoglutarato, è una reazione irre-versibile di fondamentale importanza bio-chimica che avviene entro i mitocondri. Ladecarbossilazione del piruvato, prodottodal metabolismo dei carboidrati e, in mi-sura minore, di molti amminoacidi, produ-ce acetil CoA utilizzato a scopi energeticinel ciclo dell’acido citrico oppure per la sin-tesi di acidi grassi; pertanto questa reazio-ne rappresenta una fondamentale cernie-ra tra il metabolismo di carboidrati e am-minoacidi e quello dei lipidi. L’irreversibi-lità della reazione giustifica il fatto ben notoche la maggior parte degli organismi nonpossono produrre carboidrati da lipidi. In-vece, la decarbossilazione dell’�-chetoglu-tarato trasforma quest’ultimo in succinilCoA in una delle reazioni del ciclo dell’aci-do citrico.
Come esempio di decarbossilazione os-sidativa degli �-chetoacidi prenderemo inesame la decarbossilazione del piruvato ca-talizzata dalla piruvato deidrogenasi. Lareazione in realtà comprende sia una de-carbossilazione sia un’ossidazione a caricodel substrato; essa pertanto richiede l’in-tervento di un complesso multienzimaticocostituito da tre differenti tipi di subunitàcatalitiche e da cinque diversi cofattori (ta-
bella; figura 1) che catalizzano la seguentereazione complessiva:
Piruvato � NAD� � CoA-SH 8nacetil CoA � AMP � PPi
I cinque cofattori sono NAD�, FAD, tiam-minapirofosfato (TPP, la forma attivata del-la vitamina B1, o tiamina), CoA e acido li-poico; quest’ultimo è un acido grasso a ottoatomi di carbonio contenente due gruppi
SCHEDA 9.2
Stefani-Taddei Percorsi di biochimica © Zanichelli 2011 1
50 nm
Figura 1 Fotografia al microscopio elettronicomostrante la composizione in subunità dellapiruvato deidrogenasi di Escherichia coli.
Tabella Funzione dei coenzimi e dei gruppi prostetici della reazione della piruvatodeidrogenasi
Coenzimi egruppi prostetici Localizzazione Funzione
Tiammina pirofosfato Legata alla piruvato Reagisce con il substrato, deidrogenasi piruvato
Acido lipoico Covalentemente legato Accetta il gruppo acetilea un residuo di lisina dalla tiammina pirofosfatosulla diidrolipoiltransacetilasi
Coenzima A Libero in soluzione Accetta il gruppo acetiledalla lipoammide sullatransacetilasi
Flavina adenin Fortemente legata alla Accetta equivalentidinucleotide (FAD) diidrolipoil deidrogenasi dalla lipoammide ridotta
Nicotinammide Libera in soluzione Accettore terminale degliadenin dinucleotide equivalenti riducenti dalla
flavoproteina ridotta

OSH (forma ridotta) oppure un gruppoSOS (forma ossidata).
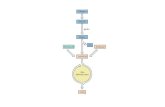
La reazione catalizzata dalla prima atti-vità enzimatica del complesso multienzi-matico consiste nella decarbossilazionedell’acido piruvico, il cui frammento bi-carbonioso residuo resta legato al TPP. Talecomplesso viene quindi trasferito dalla se-conda attività enzimatica su una moleco-la di acido lipoico ossidato (contenente unlegame SOS) che, a sua volta, lo trasferiscesu una molecola di CoA formando acetilCoA e la forma ridotta dell’acido lipoico. Laterza attività enzimatica rigenera la formaossidata dell’acido lipoico trasferendo i dueatomi di idrogeno della forma ridotta diquesto su una molecola di FAD formandoFADH2, a sua volta utilizzato per ridurreuna molecola di NAD� (questa reazione èanomala perché solitamente è il NADH �H� a ridurre il FAD). Durante tutte questereazioni i vari intermedi e cofattori resta-no sempre legati al complesso enzimaticoe al termine di un ciclo di reazione i cofat-tori acido lipoico, TPP e FAD vengono ri-generati in modo da poter partecipare a unnuovo ciclo (figura 2).
Data la sua importanza centrale nel me-tabolismo energetico e il suo carattere dicrocevia tra metabolismo di carboidrati,amminoacidi e lipidi, la reazione catalizza-ta dalla piruvato deidrogenasi è stretta-mente controllata. L’enzima è soggetto a
due tipi principali di regolazione, uno dainibizione da prodotto e uno legato a fo-sforilazione/defosforilazione del comples-so stesso. Il primo è il risultato dell’azioneinibitrice sul complesso di due dei suoi pro-dotti, l’acetil CoA e il NADH � H�, che agi-scono da inibitori competitivi.
Il secondo tipo di regolazione è legato alfatto che il complesso della piruvato dei-drogenasi è soggetto a reazioni di fosforila-zione su specifici residui di serina cataliz-zate da una proteina cinasi ATP-dipenden-te legata al complesso, che ne modificanol’attività. In particolare, la forma fosforilataè inattiva mentre quella attiva è la forma de-fosforilata; quest’ultima si origina per azio-ne di una fosfoproteina fosfatasi Mg2�- eCa2�-dipendente. L’attività della piruvatodeidrogenasi risulta complessivamente mo-dulata dalla differente regolazione delle at-tività cinasica e fosfatasica. I modulatori piùimportanti sono, oltre al piruvato stesso, an-cora acetil CoA e NADH � H�, che stimola-no l’attività cinasica rafforzando così la loroazione inibitrice complessiva; invece l’atti-vità cinasica è inibita da CoA e NAD�, la cuipresenza indica la necessità che la reazio-ne catalizzata dal complesso proceda. Quin-di, l’aumento del rapporto mitocondrialeNADH � H�/NAD� e del rapporto acetilCoA/CoA, quale si verifica per esempio du-rante la rapida �-ossidazione degli acidigrassi, determina l’inattivazione del com-
2 Stefani-Taddei Percorsi di biochimica © Zanichelli 2011
Figura 2 Decarbossilazione ossidativa del piruvato ad acetil CoA da parte del complesso della piruvatodeidrogenasi.
Acil lipoil-lisina
Lipoil-lisina ossidata
Lipoil-lisina ridotta
CoA-SH
TPP
CO2
Piruvato
Idrossietil-TPP
Piruvato deidrogenasi,
E1
Diidrolipoil transacetilasi,
E2
Diidrolipoil deidrogenasi,
E3
TPP Lys
Acetil CoA
O
CH3 CoAS-CO
CH3 C
O
O–C
CHOH
CH3
FAD
FADH2
SH
SH
NADH + H+
NAD+
3
45
21
S
S
C
CH 3
SSH
O

Stefani-Taddei Percorsi di biochimica © Zanichelli 2011 3
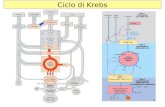
plesso ad opera della cinasi; al contrario, inpresenza di elevati livelli di piruvato la ci-nasi risulta inattivata e l’enzima presente informa defosforilata è pienamente attivo ecapace di smaltire il carico di piruvato (fi-gura 3).
Infine, il complesso della piruvato dei-drogenasi subisce anche una regolazioneormonale da insulina (nel tessuto adiposo)e da catecolammine (nel miocardio). Taleregolazione non dipende dall’intervento diproteina cinasi cAMP-dipendenti come nelcaso di altri enzimi, ma è piuttosto legataa modificazioni dei livelli di calcio intra-cellulare che modulano l’attività della fo-sfatasi.
Data l’importanza metabolica della pi-ruvato deidrogenasi, qualsiasi alterazionedella sua funzionalità comporta gravi dan-ni soprattutto a carico del cervello, in cuiquesta reazione è essenziale a causa dellastretta dipendenza dell’organo dal meta-
bolismo ossidativo del glucosio. Un esem-pio di questi danni è fornito dall’avvelena-mento da arsenito, una sostanza inorgani-ca che complessa irreversibilmente i grup-pi tiolici, per esempio dell’acido lipoico, im-pedendone la partecipazione alla reazionecatalizzata, che quindi viene a essere bloc-cata (vedi la scheda 7.1, «L’avvelenamentoda arsenico»). Un altro esempio è dato dal-la carenza alimentare di tiamina, che cau-sa una grave sindrome nota come beri-bericaratterizzata, tra l’altro, da gravi compli-canze neurologiche dovute alla mancatafunzionalità neuronale. Tale carenza puòessere causata da una dieta basata preva-lentemente sul consumo di riso brillato,particolarmente povero della vitamina, op-pure dall’alcolismo, in cui una frazione ri-levante del fabbisogno calorico è introdot-ta sotto forma di alcol, con la riduzione del-la frazione di calorie fornite da alimenti con-tenenti la vitamina.
Figura 3 Il complesso meccanismo di regolazione dell’attività della piruvato deidrogenasi.
Piruvatodeidrogenasi
inattiva
Piruvatodeidrogenasi
attiva
Cinasi
1
1
2
2
2
2
2
2
Fosfatasi
Pi
Pi
Ca2+Mg2+
Mg2+
Acetil CoA
CoASH
Piruvato
CO2
NAD+NADH
ATP
ADP