Regolazione dell'espressione genica
39
Le principali strategie di Le principali strategie di regolazione dell’espressione regolazione dell’espressione genica nei procarioti genica nei procarioti
-
Upload
alessandro-bellapianta -
Category
Science
-
view
708 -
download
0
Transcript of Regolazione dell'espressione genica

Le principali strategie di Le principali strategie di regolazione dell’espressione regolazione dell’espressione
genica nei procariotigenica nei procarioti

Regolazione metabolicaRegolazione metabolica
Nel Nel genomagenoma di un microorganismo sono presenti migliaia di geni di un microorganismo sono presenti migliaia di geni (3000(3000--6000). 6000).
Alcuni geni vengono espressi continuamente Alcuni geni vengono espressi continuamente (geni costitutivi);(geni costitutivi);altri solo quando è necessario altri solo quando è necessario (geni regolati).(geni regolati).
ENZIMI COSTITUTIVI ENZIMI COSTITUTIVI ((metabolismo del glucosiometabolismo del glucosio) ) ENZIMI INDUCIBILI ENZIMI INDUCIBILI ((prodotti in presenza del substrato
es. β–galattosidasi nel metabolismo del lattosio)

Regolazione metabolicaRegolazione metabolicaGli organismi sono in grado di regolare l’espressione dei propri geni Gli organismi sono in grado di regolare l’espressione dei propri geni
in modo che le proteine, e le altre molecole, vengano sintetizzate in modo che le proteine, e le altre molecole, vengano sintetizzate nella quantità esatta e nel momento esatto del ciclo cellulare.nella quantità esatta e nel momento esatto del ciclo cellulare.
I MECCANISMI DI REGOLAZIONEConsentono alla cellula di produrre proteine
(enzimi) solo quando servono
ECONOMIA CELLULARE

Meccanismi utilizzati nella regolazioneMeccanismi utilizzati nella regolazione
Nella cellula ci sono due strategie principali di regolazione:Controllo dell’attività di un enzima, dopo che la proteina è stata sintetizzata (regolazione post-traduzionale)Controllo della quantità sintetizzata di enzima, variando la quantità di mRNA oppure di proteina prodotti (regolazione della trascrizione e regolazione della traduzione)

Controllo dell’attività di un enzima
Inibizione dovuta a cambiamenti nell’enzima di tipo: Inibizione dovuta a cambiamenti nell’enzima di tipo:
covalente covalente (adenilazione, processamento delle proteine)(adenilazione, processamento delle proteine) non covalentenon covalente (inibizione da feedback o retroazione ; (inibizione da feedback o retroazione ;
isoenzimi)isoenzimi)

Inibizione da feedback dell’attività enzimaticaInibizione da feedback dell’attività enzimaticaMeccanismo tipico della regolazione di intere
vie biosintetiche che coinvolgono diversi passaggi enzimatici.
Il prodotto finale (es. amminoacido, nucleotide, etc.) è in grado di retroagire sul primo passaggio nella via biosintetica e di regolare la propria biosintesi (autoregolazione).
Il prodotto finale inibendo il primo enzima della via inibisce tutta la via biosintetica in quanto non vengono più generati gli intermedi necessari come substrato per gli altri enzimi coinvolti nel processo.
L’inibizione da feedback E’ REVERSIBILE:la sintesi riprende quando il prodotto finale si esaurisce

AllosteriaAllosteriaL’inibizione da feedback è resa possibile
da una proprietà dell’enzima definita ALLOSTERIA.
Un enzima allosterico possiede un sito attivo che lega il substrato e un sito allosterico dove si lega in modo reversibile l’inibitore.
Quando un inibitore si lega al sito allosterico, la conformazione dell’enzima cambia ed il substrato non riesce più a legarsi efficientemente al sito attivo.
Quando la concentrazione di inibitore diminuisce, si dissocia dal sito allosterico ed il sito attivo ritorna nella sua forma catalitica.

Inibizione da feedback Inibizione da feedback in una via biosintetica ramificatain una via biosintetica ramificata
Gli enzimi allosterici sono molto importanti nelle vie biosintetiche ramificate.Prolina e arginina sono sintetizzate a partire dall’acido glutamico.
Questi due aminoacidi possono controllare il primo enzima specifico (evidenziati in rosso) per la propria sintesi senza interferire con gli altri, in modo che, ad esempio l’eccesso di prolina
non provochi nell’organismo una carenza di arginina.
N-acetil glutamatosintetasi
g-glutamil chinasi

Isoenzimi ed inibizione da feedbackIsoenzimi ed inibizione da feedback
ISOENZIMI:Proteine diverse
che catalizzano la stessa reazione ma soggette a
sistemi di regolazione indipendenti
SINTESI AMINOACIDI AROMATICI IN E. COLIIn E. coli 3 diversi isoenzimi con attività di DAHP sintetasi catalizzano la prima reazione della
via biosintetica. Ognuno di questi enzimi è specificamente retroinibito da uno dei tre aminoacidi aromatici. In questo caso è richiesto un eccesso di tutti e tre i prodotti di reazione per spegnere
completamente la sintesi di DAHP
3-deossi-D-arabino-eptulosonato 7-fosfato

Modificazioni covalenti degli enzimiModificazioni covalenti degli enzimiI più comuni meccanismi di modificazioni enzimatica covalenti:I più comuni meccanismi di modificazioni enzimatica covalenti:
ADENILAZIONE:ADENILAZIONE: legame di un nucleotide adenosinmonofosfato (legame di un nucleotide adenosinmonofosfato (AMPAMP) o ) o adenosindifosfato (adenosindifosfato (ADPADP))
FOSFORILAZIONE:FOSFORILAZIONE: legame di un gruppo fosfato (legame di un gruppo fosfato (POPO4422--))
METILAZIONE:METILAZIONE: legame di un gruppo metilico (legame di un gruppo metilico (CHCH33))
Sono processi rapidamente reversibiliSono processi rapidamente reversibili
Il legame del gruppo modificante provoca un cambiamento conformazionale Il legame del gruppo modificante provoca un cambiamento conformazionale della proteina e può alterarne profondamente l’attività catalitica. della proteina e può alterarne profondamente l’attività catalitica.
La rimozione del gruppo modificante ripristina l’enzima nel suo stato attivo.La rimozione del gruppo modificante ripristina l’enzima nel suo stato attivo.

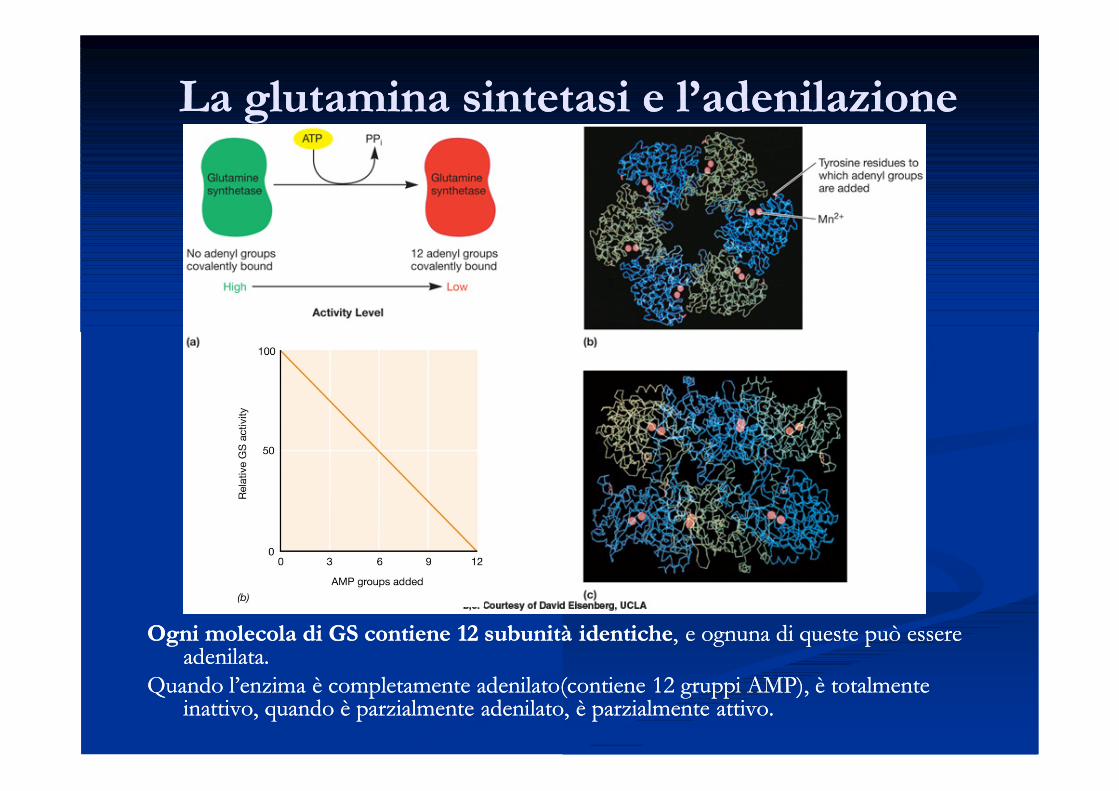
Regolazione della glutamina sintetasiRegolazione della glutamina sintetasi
La La glutamina sintetasi (GS)glutamina sintetasi (GS) è un enzima che svolge un ruolo chiave nel è un enzima che svolge un ruolo chiave nel metabolismo dell’azoto nella cellula.metabolismo dell’azoto nella cellula.
L’attività della GS può essere controllata mediante:L’attività della GS può essere controllata mediante:
Inibizione da feedback concertata:Inibizione da feedback concertata: ad opera di 9 composti ad opera di 9 composti diversi (aa, prodotti del metabolismo nucleotidico). L’attività diversi (aa, prodotti del metabolismo nucleotidico). L’attività enzimatica si riduce proporzionalmente al numero di inibitori enzimatica si riduce proporzionalmente al numero di inibitori presenti: la presenza di tutti e nove gli inibitori determina presenti: la presenza di tutti e nove gli inibitori determina l’inibizione completal’inibizione completa
Adenilazione:Adenilazione: modificazione covalentemodificazione covalente

La glutamina sintetasi e l’adenilazioneLa glutamina sintetasi e l’adenilazione
Il controllo della GS mediante modificazione covalente Il controllo della GS mediante modificazione covalente è determinato dalla è determinato dalla concentrazione di concentrazione di glutaminaglutamina e di un precursore della e di un precursore della glutaminaglutamina, l’, l’aa--chetoglutaratochetoglutarato, un , un intermedio del ciclo dell’acido citrico.intermedio del ciclo dell’acido citrico.
Quando il livello di Quando il livello di glutaminaglutamina nella cellula nella cellula diminuiscediminuisce, l’enzima che aggiunge e rimuove i , l’enzima che aggiunge e rimuove i
gruppi AMP (enzima Pgruppi AMP (enzima PIIII)) viene modificato viene modificato covalentementecovalentemente e e promuove la promuove la deadenilazionedeadenilazione della GS, e di conseguenza, un aumento della sua attivitàdella GS, e di conseguenza, un aumento della sua attività. .
Quando la concentrazione intracellulare di Quando la concentrazione intracellulare di glutaminaglutamina aumenta, la GS viene aumenta, la GS viene fortemente fortemente adenilataadenilata e la sua attività diminuisce.e la sua attività diminuisce.
GS GS-AMP(1-12)
ATP PPi
PiADP
GlutaminaAMP
La La glutaminaglutamina sintetasisintetasi e l’e l’adenilazioneadenilazione
Ogni molecola di GS contiene 12 Ogni molecola di GS contiene 12 subunitàsubunità identicheidentiche, e ognuna di queste può essere , e ognuna di queste può essere adenilataadenilata. .
Quando l’enzima è completamente Quando l’enzima è completamente adenilatoadenilato(contiene 12 gruppi AMP), è totalmente (contiene 12 gruppi AMP), è totalmente inattivo, quando è parzialmente inattivo, quando è parzialmente adenilatoadenilato, è parzialmente attivo. , è parzialmente attivo.

La La glutaminaglutamina sintetasisintetasi e l’e l’adenilazioneadenilazione
I livelli di attività della GS possono essere considerati per la I livelli di attività della GS possono essere considerati per la cellula una specie di barometro ad azotocellula una specie di barometro ad azoto::
N GS
Livelli di azoto Attività glutamina sintetasi
E VICEVERSA

Splicing delle proteineSplicing delle proteine
Nel processamento della subunità A della DNA girasi di M. leprae,codificata dal gene gyrA, le sequenze fiancheggianti del polipeptide GyrA, chiamate esteine,
vengono riunite per formare la forma attiva della proteina, mentre le inteine vengono scartate.Le inteine sono proteasi specifiche self-splicing (autoprocessanti).
RUOLO INTEINE?
Anche nei procarioti esistono alcuni rari geni con introni. Recentemente sono stati individuati numerosi casi in cui l’informazione “extra” viene
rimossa a livello della proteina e non dell’RNA. Il processo è stato definito “splicing delle proteine”, e la parte proteica interna che
viene rimossa è stata chiamata inteina.

La regolazione della trascrizioneLa regolazione della trascrizione
Controllo positivo:Controllo positivo: (operone maltosio)(operone maltosio)L’elemento di controllo, una proteina regolatrice, promuove il L’elemento di controllo, una proteina regolatrice, promuove il
legame dell’RNA polimerasi determinando un aumento legame dell’RNA polimerasi determinando un aumento della sintesi di mRNA della sintesi di mRNA
Controllo negativo:Controllo negativo: repressionerepressione (operone arginina) (operone arginina) ed ed induzione (induzione (operone lattosio)operone lattosio)
L’elemento di controllo, il repressore, inibisce la sintesi L’elemento di controllo, il repressore, inibisce la sintesi dell’mRNAdell’mRNA

Le proteine che legano il DNALe proteine che legano il DNA
Le interazioni tra proteine ed acidi nuclei sono Le interazioni tra proteine ed acidi nuclei sono essenziali per la replicazione, la essenziali per la replicazione, la trascrizione, la traduzione, nonché per la trascrizione, la traduzione, nonché per la regolazione di tali processi.regolazione di tali processi.
1.1. INTERAZIONI ASPECIFICHEINTERAZIONI ASPECIFICHE2.2. INTERAZIONI SEQUENZAINTERAZIONI SEQUENZA--
SPECIFICASPECIFICA
Le sequenze ripetute invertite costituiscono spesso siti specifici di legame per le proteine. Le proteine che interagiscono con il DNA sono spesso omodimeri, composte da due catene polipetidiche identiche. Su ogni catena polipeptidica c’è un dominio di legame al DNA che si inserisce nel solco maggiore e lungo l’impalcatura di fosfato e un dominio di interazione proteina-proteina che tiene insieme le due catene del dimero.
Sequenza nucleotitica della regione operatore dell’operone lattosio legata dal repressore lac

Struttura delle proteine che legano il DNAStruttura delle proteine che legano il DNA
Classi di domini proteici di fondamentale importanza per Classi di domini proteici di fondamentale importanza per la formazione del corretto legame con il DNA:la formazione del corretto legame con il DNA:
HelixHelix--turnturn--helix (elicahelix (elica--girogiro--elica)elica) Zinc finger (dito di zinco)Zinc finger (dito di zinco) Leucine zipper (cerniera di leucine)Leucine zipper (cerniera di leucine)

Struttura helixStruttura helix--turnturn--helixhelixIl Il motivo strutturale motivo strutturale helixhelix--turnturn--helixhelix
possiede:possiede: Un’ elica di riconoscimentoUn’ elica di riconoscimento ((aa--
elica)elica) che interagisce con il DNAche interagisce con il DNA Una breve sequenza di tre Una breve sequenza di tre aaaa,, il il
primo dei quali è generalmente una primo dei quali è generalmente una glicinaglicina con la funzione di “girare la con la funzione di “girare la proteina”, ovvero di curvarla a gomitoproteina”, ovvero di curvarla a gomito
Un’elica stabilizzanteUn’elica stabilizzante,, che stabilizza che stabilizza la prima interagendo con essa la prima interagendo con essa idrofobicamente e partecipando alla idrofobicamente e partecipando alla formazione del dimero. formazione del dimero.
Il riconoscimento di sequenze specifiche nel DNA avviene tramite interazioni non covalentiche comportano la formazione di legami idrogeno ed interazioni di van der Waals.
Nei Batteri ci sono diverse proteine con questo motivo:tra cui molti repressori (sistemi lac e trp di E. coli; repressore del batteriofago l)
Oltre 250 proteine con questo motivo si legano al DNA per regolare la trascrizione in E. coli.

Modello delle substrutture proteiche che si trovano Modello delle substrutture proteiche che si trovano nelle proteine eucariotiche che legano il DNAnelle proteine eucariotiche che legano il DNA
Nel motivo zinc fingerNel motivo zinc finger tra gli aa che tra gli aa che legano lo ione Znlegano lo ione Zn2+2+ vi sono vi sono sempre almeno due residui di sempre almeno due residui di cisteina (C), mentre gli altri residui cisteina (C), mentre gli altri residui sono generalmente istidine (H). sono generalmente istidine (H).
Nel motivo leucine zipperNel motivo leucine zipper i residui i residui di leucina sono regolarmente di leucina sono regolarmente ripetuti ogni sette aa formando una ripetuti ogni sette aa formando una struttura simile ad una cerniera struttura simile ad una cerniera lampo. La cerniera di leucine serve lampo. La cerniera di leucine serve a tenere unite due eliche di a tenere unite due eliche di riconoscimento che interagiscono riconoscimento che interagiscono con il DNA con il DNA

Il controllo negativo della trascrizione
Nei batteri sono noti diversi meccanismi per il controllo della sintesi di un enzima e tutti sono ampiamente influenzati dall’ambiente nel quale
l’organismo cresce ed in particolare dalla presenza o assenza di piccole molecole specifiche (come quelle che
legano il DNA) per controllare la trascrizione o più raramente la traduzione.
La repressione e l’induzione rappresentano le forme più semplici di regolazione che controllano
l’espressione genica a livello della trascrizione.

Repressione dell’attività enzimatica
Gli enzimi coinvolti nella sintesi dell’ arginina sono sintetizzati soltanto se l’arginina è assente nel mezzo di crescita; l’aggiunta di arginina nel mezzo reprime la loro sintesi.
E’da notare, come questo sia un effetto specifico, in quanto la sintesi di tutti gli altri enzimi continua regolarmente.
La repressione enzimatica è un fenomeno ampiamente diffuso nei batteri e viene utilizzato nel controllo della sintesi di
vari enzimi coinvolti nella biosintesi di aminoacidi, purine e pirimidine.
In quasi tutti i casi è il prodotto finale di una particolare via biosintetica a reprimere gli
enzimi della stessa.
Si può ovviamente intuire quanto sia importante questo processo per il
microrganismo, dato che gli permette di non sprecare energia sintetizzando
enzimi non necessari.

Induzione dell’attività enzimaticaInduzione dell’attività enzimatica
Gli enzimi coinvolti nel catabolismo del carbonio Gli enzimi coinvolti nel catabolismo del carbonio o di altre fonti di energia sono spesso o di altre fonti di energia sono spesso
inducibili. inducibili. Tipico esempio è dato dall’enzima Tipico esempio è dato dall’enzima
bb--galattosidasigalattosidasi,, coinvolto nella utilizzazione coinvolto nella utilizzazione del lattosio, che non viene sintetizzato se del lattosio, che non viene sintetizzato se lo zucchero è assente nel mezzo, mentre lo zucchero è assente nel mezzo, mentre
la sintesi inizia non appena al terreno la sintesi inizia non appena al terreno viene aggiunto lattosio.viene aggiunto lattosio.
Anche in questo caso si può facilmente Anche in questo caso si può facilmente comprendere il valore di tale comprendere il valore di tale
meccanismo, dal momento che esso meccanismo, dal momento che esso permette all’organismo di non permette all’organismo di non
sintetizzare un enzima fino a quando non sintetizzare un enzima fino a quando non gli sia necessario.gli sia necessario.
Un fenomeno complementare alla repressione è Un fenomeno complementare alla repressione è l’induzione di un enzimal’induzione di un enzima, ovvero la , ovvero la sintesi di un particolare enzima solo quando è presente il suo substrato.sintesi di un particolare enzima solo quando è presente il suo substrato.

L’induttore ed il corepressoreL’induttore ed il corepressoreEFFETTORIEFFETTORI
Induttore:Induttore: sostanza che dà inizio all’induzione di un enzima sostanza che dà inizio all’induzione di un enzima CorepressoreCorepressore:: sostanza che reprime la sintesi di un enzimasostanza che reprime la sintesi di un enzima
Gli induttori e i Gli induttori e i corepressoricorepressori in genere in genere agiscono in modo indirettoagiscono in modo indiretto, , combinandosi con specifiche proteine di regolazione che a loro volta combinandosi con specifiche proteine di regolazione che a loro volta influenzano la sintesi di influenzano la sintesi di mRNAmRNA. .
Non tutti gli effettori sono substrato o prodotti finali degli specifici enzimi Non tutti gli effettori sono substrato o prodotti finali degli specifici enzimi regolati.regolati.
Alcuni analoghi di queste sostanze sono in grado di provocare l’induzione o la Alcuni analoghi di queste sostanze sono in grado di provocare l’induzione o la repressione di un enzima senza esserne il substrato. repressione di un enzima senza esserne il substrato.
L’L’isopropiltiogalattosideisopropiltiogalattoside (IPTG),(IPTG), per esempio, è un induttore della per esempio, è un induttore della bb--galattosidasigalattosidasi, pur non essendo idrolizzato dall’enzima stesso. , pur non essendo idrolizzato dall’enzima stesso.

Regolazione dell’operone argininaRegolazione dell’operone arginina
Nel caso di un enzima reprimibile, il corepressore (per esempio l’arginina) si lega alla proteina repressore specifica (il repressore dell’arginina). Il repressore è una proteina allosterica la cui
conformazione può venire modificata in seguito al legame con il corepressore.Una volta modificato, il repressore può legarsi a una regione specifica di DNA vicina al promotore del gene stesso, la regione operatore che deve il suo nome all’operone, gruppo di geni la cui espressione è sotto il controllo di un singolo promotore. Se il repressore si lega all’operatore, la sintesi di mRNA è bloccata perché l’RNA polimerasi non si può legare o non può procedere nella trascrizione e tutte le proteine codificate da quell’mRNA policistronico saranno represse.

Regolazione dell’operone lattosioRegolazione dell’operone lattosio
Anche l’induzione di un enzima può Anche l’induzione di un enzima può essere controllata da un repressore; essere controllata da un repressore;
in questo caso il repressore è in questo caso il repressore è attivo in assenza dell’induttore e attivo in assenza dell’induttore e
capace quindi di bloccare la capace quindi di bloccare la sintesi di sintesi di mRNAmRNA..
Quando invece si aggiunge l’induttore, Quando invece si aggiunge l’induttore, questo si lega al repressore questo si lega al repressore
inattivandolo, rimuovendo il inattivandolo, rimuovendo il blocco blocco trascrizionaletrascrizionale e e
permettendo la sintesi dell’permettendo la sintesi dell’mRNAmRNA..
Dal momento che il ruolo dei Dal momento che il ruolo dei repressori è inibitorio repressori è inibitorio (reprimono la sintesi (reprimono la sintesi
dell’dell’mRNAmRNA), il tipo di ), il tipo di regolazione che li coinvolge regolazione che li coinvolge
viene spesso definito viene spesso definito controllo negativocontrollo negativo..
Nel caso dell’operone lac, il repressore è costituito dal repressore lac e l’induttore
dall’allolattosio

Regolazione dell’operone maltosioRegolazione dell’operone maltosio Il catabolismo del maltosio in E. Il catabolismo del maltosio in E.
coli è un tipico esempio di coli è un tipico esempio di sistema sottoposto a sistema sottoposto a regolazione regolazione positivapositiva. .
Gli enzimi necessari Gli enzimi necessari all’utilizzazione del maltosio in E. all’utilizzazione del maltosio in E. coli sono sintetizzati solo dopo coli sono sintetizzati solo dopo l’aggiunta di maltosio al mezzo.l’aggiunta di maltosio al mezzo.
La sintesi di questi enzimi è La sintesi di questi enzimi è controllata a livello controllata a livello trascrizionaletrascrizionaleda una da una proteina proteina attivatriceattivatrice che che non può legarsi al DNA se prima non può legarsi al DNA se prima non si è legata al non si è legata al maltosio,maltosio, il suo il suo induttore.induttore.
Il legame dell’attivatore al Il legame dell’attivatore al DNA induce un cambiamento DNA induce un cambiamento della sua struttura e permette della sua struttura e permette alla RNA polimerasi di iniziare alla RNA polimerasi di iniziare la trascrizionela trascrizione..
La sequenza specifica che serve La sequenza specifica che serve da sito di legame per l’attivatore si da sito di legame per l’attivatore si chiama chiama sito di legame sito di legame dell’attivatore.dell’attivatore.
I geni necessari per l’utilizzazione del maltosio sono dispersi I geni necessari per l’utilizzazione del maltosio sono dispersi in diversi operoni, ognuno dei quali possiede un sito di legame in diversi operoni, ognuno dei quali possiede un sito di legame
dell’attivatore.dell’attivatore.Quando diversi operoni si trovano sotto il controllo primario Quando diversi operoni si trovano sotto il controllo primario
della stessa proteina costituiscono un regulone.della stessa proteina costituiscono un regulone.
In assenza di induttore (maltosio)
In presenza di induttore (maltosio)
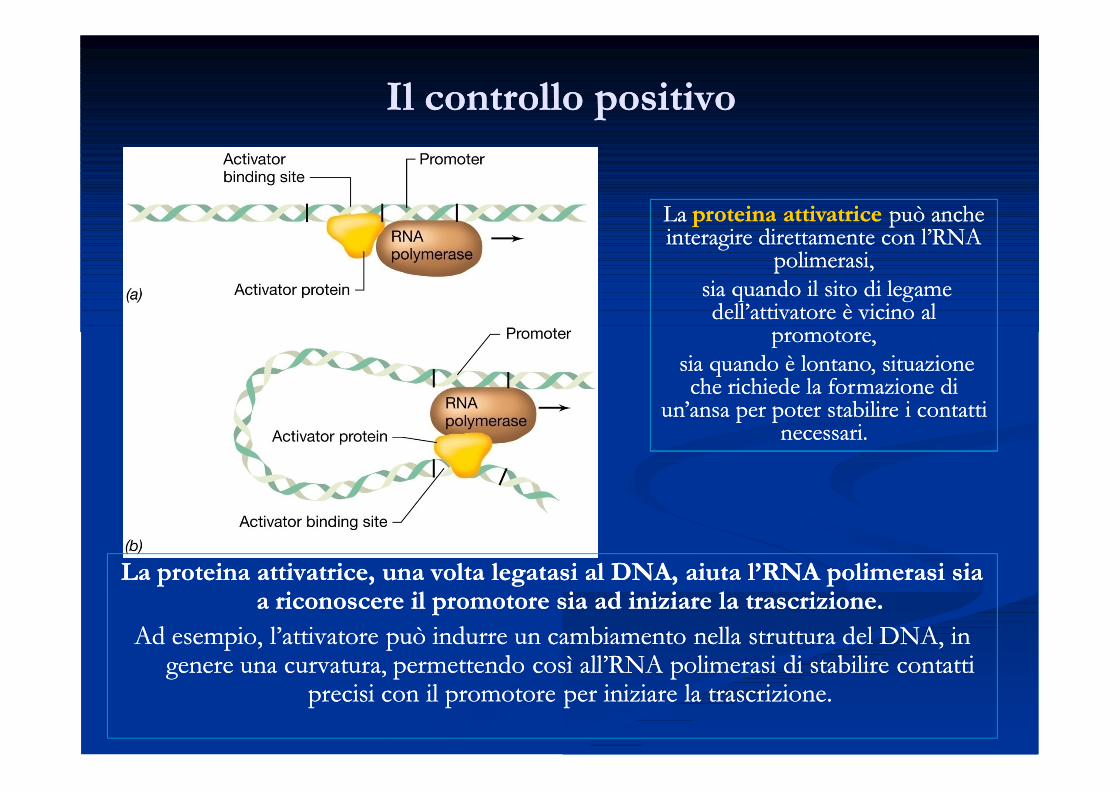
Il controllo positivoIl controllo positivo
La proteina attivatrice, una volta legatasi al DNA, aiuta l’RNA polimerasi sia La proteina attivatrice, una volta legatasi al DNA, aiuta l’RNA polimerasi sia a riconoscere il promotore sia ad iniziare la trascrizione.a riconoscere il promotore sia ad iniziare la trascrizione.
Ad esempio, l’attivatore può indurre un cambiamento nella struttura del DNA, in Ad esempio, l’attivatore può indurre un cambiamento nella struttura del DNA, in genere una curvatura, permettendo così all’RNA polimerasi di stabilire contatti genere una curvatura, permettendo così all’RNA polimerasi di stabilire contatti
precisi con il promotore per iniziare la trascrizione.precisi con il promotore per iniziare la trascrizione.
La La proteina attivatriceproteina attivatrice può anche può anche interagire direttamente con l’RNA interagire direttamente con l’RNA
polimerasi,polimerasi,sia quando il sito di legame sia quando il sito di legame dell’attivatore è vicino al dell’attivatore è vicino al
promotore,promotore,sia quando è lontano, situazione sia quando è lontano, situazione che richiede la formazione di che richiede la formazione di
un’ansa per poter stabilire i contatti un’ansa per poter stabilire i contatti necessari.necessari.

La regolazione globaleLa regolazione globale Spesso, in risposta ad una variazione ambientale, un Spesso, in risposta ad una variazione ambientale, un
organismo ha bisogno di regolare simultaneamente geni organismo ha bisogno di regolare simultaneamente geni diversi.diversi.
I meccanismi di regolazione che rispondono a I meccanismi di regolazione che rispondono a segnali ambientali sono definiti segnali ambientali sono definiti sistemi di controllo globalesistemi di controllo globale..
Nell’ambiente in cui crescono le cellule batteriche, infatti, Nell’ambiente in cui crescono le cellule batteriche, infatti, possono essere presenti diverse fonti di carbonio utilizzabili e possono essere presenti diverse fonti di carbonio utilizzabili e sarebbe uno spreco indurre, gli enzimi per il catabolismo del sarebbe uno spreco indurre, gli enzimi per il catabolismo del lattosio o del maltosio quando le cellule stanno utilizzando una lattosio o del maltosio quando le cellule stanno utilizzando una fonte di carbonio più facilmente metabolizzabile.fonte di carbonio più facilmente metabolizzabile.

La repressione da catabolitaLa repressione da catabolitaIl problema viene risolto da uno dei sistemi di Il problema viene risolto da uno dei sistemi di
regolazione globale definitoregolazione globale definitorepressione da catabolitarepressione da catabolita..
Nella repressione da catabolita la sintesi di numerosi enzimi Nella repressione da catabolita la sintesi di numerosi enzimi viene inibita quando le cellule crescono in un terreno che viene inibita quando le cellule crescono in un terreno che contiene una fonte di energia ottimale come il glucosio. contiene una fonte di energia ottimale come il glucosio.
La repressione da catabolita è stata anche definita La repressione da catabolita è stata anche definita effetto effetto glucosioglucosio,, perché questa è stata la prima sostanza per la quale si è perché questa è stata la prima sostanza per la quale si è dimostrato il fenomeno; anche se è noto che diverse fonti di dimostrato il fenomeno; anche se è noto che diverse fonti di carbonio determinano questo tipo di repressione. carbonio determinano questo tipo di repressione.

Crescita diauxicaCrescita diauxicain presenza di una miscela di glucosio e lattosioin presenza di una miscela di glucosio e lattosio
Nella Nella cresita diauxicacresita diauxica, l’organismo cresce , l’organismo cresce prima utilizzando una fonte di energia (ad prima utilizzando una fonte di energia (ad esempio il glucosio), e segue poi un lungo esempio il glucosio), e segue poi un lungo intervallo di tempo prima che la crescita intervallo di tempo prima che la crescita ricominci utilizzando la seconda fonte (ad ricominci utilizzando la seconda fonte (ad esempio lattosio). esempio lattosio).
Quando un microrganismo cresce in presenza Quando un microrganismo cresce in presenza di una miscela di glucosio e lattosio, finchè di una miscela di glucosio e lattosio, finchè il glucosio è presente nel mezzo, la il glucosio è presente nel mezzo, la bb--galattosidasigalattosidasi, codificata dal gene , codificata dal gene lacZ lacZ dell’operone lattosio e responsabile della dell’operone lattosio e responsabile della utilizzazione di questo zucchero, utilizzazione di questo zucchero, non non viene sintetizzataviene sintetizzata ed il microrganismo ed il microrganismo utilizza solo il glucosio lasciando intatto il utilizza solo il glucosio lasciando intatto il lattosio. lattosio.
Quando il glucosio è esaurito, Quando il glucosio è esaurito, la repressione la repressione da catabolita finisceda catabolita finisce e dopo un certo e dopo un certo intervallo si osserva sintesi di intervallo si osserva sintesi di bb--galattosidasi e inizia la crescita basata sul galattosidasi e inizia la crescita basata sul lattosio come fonte di carbonio.lattosio come fonte di carbonio.
Una conseguenza della repressione da catabolita è la crescita diauxica che si osserva quando Una conseguenza della repressione da catabolita è la crescita diauxica che si osserva quando due fonti di energia sono presenti contemporaneamente nel mezzo.due fonti di energia sono presenti contemporaneamente nel mezzo.
La repressione da catabolita permette alle cellule di utilizzare La repressione da catabolita permette alle cellule di utilizzare subito la migliore fonte di carbonio, infatti, i batteri crescono subito la migliore fonte di carbonio, infatti, i batteri crescono
più rapidamente in presenza di glucosio.più rapidamente in presenza di glucosio.

La repressione da catabolitaLa repressione da catabolita
La repressione da catabolita si basa sul controllo della trascrizione mediato da un La repressione da catabolita si basa sul controllo della trascrizione mediato da un attivatore ed è quindi attivatore ed è quindi un tipo di controllo positivo.un tipo di controllo positivo.
Nel caso di enzimi reprimibili da parte del catabolita, il legame della RNA polimerasi al Nel caso di enzimi reprimibili da parte del catabolita, il legame della RNA polimerasi al DNA avviene solo quando si è legata un’altra proteina, chiamata DNA avviene solo quando si è legata un’altra proteina, chiamata attivatore proteico attivatore proteico del catabolita (CAP),del catabolita (CAP), che a sua volta può legarsi al DNA solo se prima si è legato che a sua volta può legarsi al DNA solo se prima si è legato l’AMP ciclicol’AMP ciclico, un elemento chiave in numerosi sistemi controllo. , un elemento chiave in numerosi sistemi controllo.
L’entrata del glucosio nella cellula inibisce la sintesi dell’AMP ciclico e stimola il suo L’entrata del glucosio nella cellula inibisce la sintesi dell’AMP ciclico e stimola il suo trasporto al di fuori della cellula con riduzione della sua concentrazione intracellulare ed trasporto al di fuori della cellula con riduzione della sua concentrazione intracellulare ed inibizione del legame della RNA polimerasi ai promotori sensibili a questo tipo di inibizione del legame della RNA polimerasi ai promotori sensibili a questo tipo di controllo. controllo.
La repressione da catabolita, quindi, è in realtà dovuta alla mancanza di AMP La repressione da catabolita, quindi, è in realtà dovuta alla mancanza di AMP ciclico nella cellula e può essere superata aggiungendo tale composto al terrenociclico nella cellula e può essere superata aggiungendo tale composto al terreno..

La repressione da catabolitaLa repressione da catabolita Nonostante questo tipo di regolazione possa sembrare un semplice sistema di Nonostante questo tipo di regolazione possa sembrare un semplice sistema di
controllo positivo, ognuno degli operoni controllati da CAP è anche soggetto a controllo positivo, ognuno degli operoni controllati da CAP è anche soggetto a regolazione da parte dei suoi regolatori specifici. regolazione da parte dei suoi regolatori specifici.
Poiché la repressione da catabolita modula l’espressione di sistemi di Poiché la repressione da catabolita modula l’espressione di sistemi di regolazione non correlati essa rappresenta uno dei principali esempi di regolazione non correlati essa rappresenta uno dei principali esempi di
regolazione globale.regolazione globale. Finchè è presente glucosio viene impedita l’espressione di tutti gli altri Finchè è presente glucosio viene impedita l’espressione di tutti gli altri
operoni catabolici soggetti a questo elemento di controllo globale.operoni catabolici soggetti a questo elemento di controllo globale.
Affinché la trascrizione possa avvenire, devono essere rispettate due condizioni: Affinché la trascrizione possa avvenire, devono essere rispettate due condizioni: 1.1. il ivello di AMP ciclico deve essere abbastanza elevato, in modo da il ivello di AMP ciclico deve essere abbastanza elevato, in modo da
permettere alla proteina CAP di legarsi al sito di legame specificopermettere alla proteina CAP di legarsi al sito di legame specifico(controllo positivo);(controllo positivo);
2.2. deve essere presente un induttore, come il lattosio, in modo che il deve essere presente un induttore, come il lattosio, in modo che il repressore del lattosio non blocchi la trascrizione legandosi all’operatorerepressore del lattosio non blocchi la trascrizione legandosi all’operatore(controllo negativo).(controllo negativo).
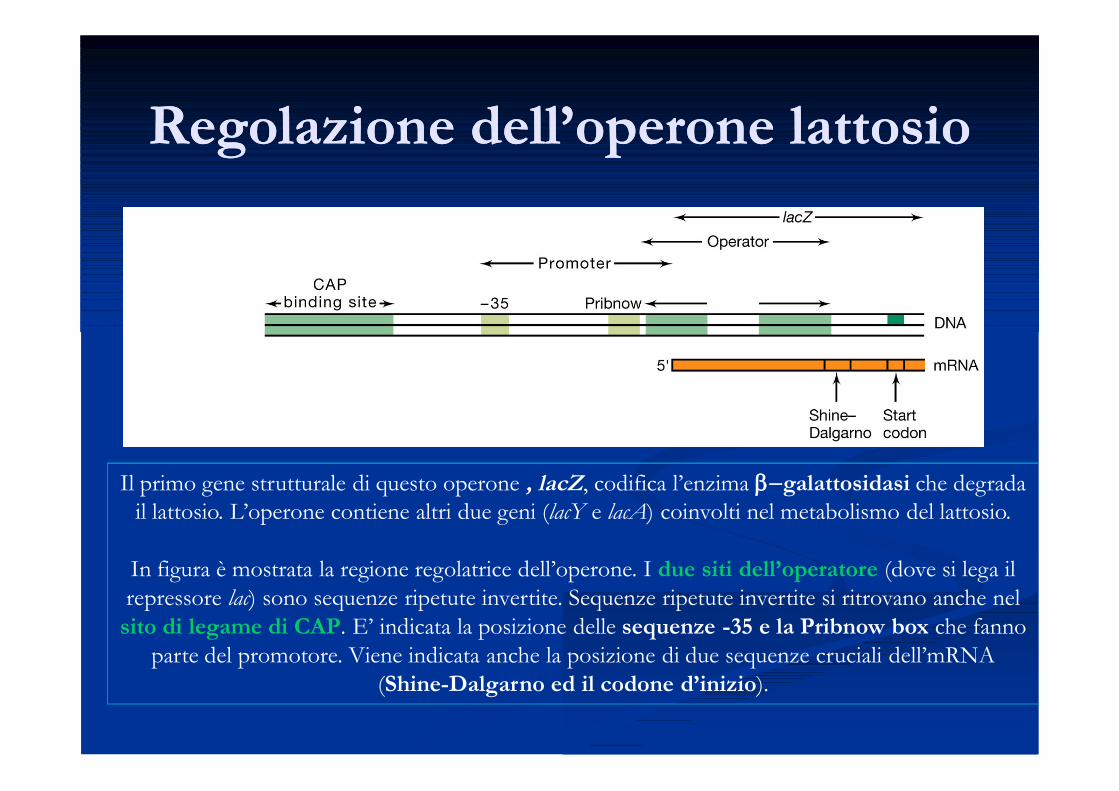
Regolazione dell’operone lattosioRegolazione dell’operone lattosio
Il primo gene strutturale di questo operone , lacZ, codifica l’enzima b-galattosidasi che degrada il lattosio. L’operone contiene altri due geni (lacY e lacA) coinvolti nel metabolismo del lattosio.
In figura è mostrata la regione regolatrice dell’operone. I due siti dell’operatore (dove si lega il repressore lac) sono sequenze ripetute invertite. Sequenze ripetute invertite si ritrovano anche nel
sito di legame di CAP. E’ indicata la posizione delle sequenze -35 e la Pribnow box che fanno parte del promotore. Viene indicata anche la posizione di due sequenze cruciali dell’mRNA
(Shine-Dalgarno ed il codone d’inizio).

L’attenuazione dell’operone triptofano in L’attenuazione dell’operone triptofano in E. coliE. coli
Oltre al promotore Oltre al promotore PP e all’operatore e all’operatore O O c’è c’è una sequenza leader una sequenza leader LL che codifica un che codifica un polipeptide contenente residui di polipeptide contenente residui di triptofano ripetuti in tandem vicino triptofano ripetuti in tandem vicino alla sua estremità terminale e operante alla sua estremità terminale e operante come attenuatore. come attenuatore.
Se nella cellula è il triptofano è Se nella cellula è il triptofano è abbondante, il peptide leader verrà abbondante, il peptide leader verrà sintetizzato, determinando la sintetizzato, determinando la terminazione della trascrizione del terminazione della trascrizione del restante operone trp, che restante operone trp, che comprende i geni strutturali per gli comprende i geni strutturali per gli enzimi biosintetici; enzimi biosintetici;
se invece il triptofano è scarso, il se invece il triptofano è scarso, il peptide leader, che è ricco in peptide leader, che è ricco in triptofano, non sarà prodotto, e la triptofano, non sarà prodotto, e la trascrizione della restante parte trascrizione della restante parte dell’operone potrà aver luogo. dell’operone potrà aver luogo.
L’operone triptofano contiene i geni strutturali L’operone triptofano contiene i geni strutturali per cinque proteine della via biosintetica del per cinque proteine della via biosintetica del triptofano, oltre al promotore e alle sequenze triptofano, oltre al promotore e alle sequenze regolatrici localizzate all’inizio dell’operone.regolatrici localizzate all’inizio dell’operone.
Questo operone è soggetto in realtà a diversi tipi di regolazione: il primo enzima della via, Questo operone è soggetto in realtà a diversi tipi di regolazione: il primo enzima della via, l’antranilato sintetasi è infatti, sottoposto l’antranilato sintetasi è infatti, sottoposto all’inibizione da feedback da parte del triptofanoall’inibizione da feedback da parte del triptofano, ed , ed
inoltre la trascrizione dell’operone è sotto il inoltre la trascrizione dell’operone è sotto il controllo del repressore del triptofanocontrollo del repressore del triptofano..

L’attenuazione dell’operone triptofanoL’attenuazione dell’operone triptofano
Se il triptofano è presente in abbondanza, il ribosoma potrà tradurre l’intero peptide leader Se il triptofano è presente in abbondanza, il ribosoma potrà tradurre l’intero peptide leader (codificato dalle regioni 1 e 2), e così la regione 2 non potrà appaiarsi con la regione 3. Le regioni (codificato dalle regioni 1 e 2), e così la regione 2 non potrà appaiarsi con la regione 3. Le regioni
3 e 4 si appaieranno a formare una struttura ansa3 e 4 si appaieranno a formare una struttura ansa--stelo, che costituirà un sito di pausa della stelo, che costituirà un sito di pausa della trascrizione. Poi trovandosi a valle di questa sequenza un tratto ricco in uracile, si avrà la trascrizione. Poi trovandosi a valle di questa sequenza un tratto ricco in uracile, si avrà la
terminazione.terminazione.
Per spiegare questo fenomeno bisogna tener conto che Per spiegare questo fenomeno bisogna tener conto che nelle cellule procariotiche i processi di traduzione e nelle cellule procariotiche i processi di traduzione e trascrizione avvengono simultaneamente:trascrizione avvengono simultaneamente: mentre la trascrizione di regioni a valle è ancora in corso, è già iniziata mentre la trascrizione di regioni a valle è ancora in corso, è già iniziata
la traduzione delle regioni trascritte.la traduzione delle regioni trascritte.L’attenuazione ha luogo perché una parte dell’mRNA neosintetizzato si ripiega a formare L’attenuazione ha luogo perché una parte dell’mRNA neosintetizzato si ripiega a formare un’ansa a doppia elica, grazie alla presenza di sequenze complementari, seguita da una un’ansa a doppia elica, grazie alla presenza di sequenze complementari, seguita da una sequenza di uracili che segnalano alla RNA polimerasi di cessare l’attività trascrizionale.sequenza di uracili che segnalano alla RNA polimerasi di cessare l’attività trascrizionale.

L’attenuazione dell’operone triptofanoL’attenuazione dell’operone triptofano
Se invece il triptofano è scarso, il Se invece il triptofano è scarso, il ribosoma rallenterà in ribosoma rallenterà in corrispondenza del codone corrispondenza del codone triptofano; la sosta del ribosoma in triptofano; la sosta del ribosoma in questa posizione permetterà la questa posizione permetterà la formazione di una struttura ansaformazione di una struttura ansa--stelo alternativa (regione 2stelo alternativa (regione 2--regione regione 3) che non rappresenta un segnale di 3) che non rappresenta un segnale di terminazione e previene terminazione e previene efficacemente la formazione del vero efficacemente la formazione del vero terminatore. terminatore.
L’RNA polimerasi oltrepasserà il sito L’RNA polimerasi oltrepasserà il sito di terminazione che non si è di terminazione che non si è correttamente ripiegato e inizierà la correttamente ripiegato e inizierà la trascrizione dei geni strutturali del trascrizione dei geni strutturali del triptofano triptofano

I sistemi di regolazione a due componentiI sistemi di regolazione a due componenti
Molti dei sistemi di regolazione tramite i quali i batteri sono in grado di rilevare i segnali
ambientali e fornire una risposta sono costituiti da due componeneti. Un sistema classico a due componenti è costituito da:1. Una proteina sensore (chinasi
sensore), locaizzata nelle membrana cellulare e in grado di autofosforiularsi in
risposta a un segnale ambienetale2. Una proteina regolatrice della risposta, generalmente una proteina di
legame al DNA che regola la trascrizione in modo + o – a seconda del sistema

La regolazione della chemiotassiLa regolazione della chemiotassi
Il grado di metilazione delle proteine MCP controlla la loro capacità di rispondere ad una sostanza chimica e determina il processo dell’ADATTAMENTO.