
POLIMORFISMI delle GST - Homepage |...
33
POLIMORFISMI delle GST
Transcript of POLIMORFISMI delle GST - Homepage |...

POLIMORFISMI delle GST

POLIMORFISMO: insieme di due o più fenotipi alternativi comuni nella popolazione.
Una misura della variabilità genetica è la quantità di eterozigosi in una popolazione, che è data dalla frequenza totale di eterozigoti in un locus. Se un
allele è presente in una frequenza altissima e tutti gli altri sono prossimi allo 0, ci sarà pochissima eterozigosi. Quest’ ultima sarà quindi più elevata quando ci sono
molti alleli per un locus, tutti con una stessa frequenza.Il polimorfismo, che è un’espressione della variabilità genica può riflettersi in uno o più livelli fenotipici all’interno di una stessa popolazione o tra popolazioni differenti.
POLIMORFISMI CROMOSOMICI: variazioni dei siti di restrizioneripetizioni in tandem
POLIMORFISMI IMMUNOLOGICI: tipi sanguigni del sistema AB0
POLIMORFOSMI PROTEICI: Glutatione trasferasi

Le GST sono divise in tre famiglie principali:
•Mitocondriali•Citosoliche•Microsomali
Alle sette classi individuate nei mammiferi, Alpha (a), Mu (µ), Pi (p ), Sigma (s ), Theta (?), Omega (? ) e Zeta (?), (Mannervik et al., 1985; Meyer et al., 1991; Meyer & Thomas 1995; Board et al., 1997, 2000), se ne aggiungono
altre, denominate Beta (ß), Delta (d), Epsilon (e), Lambda (?), Phi (f ) e Tau (t), identificate in batteri, insetti e piante (Sheehan et al., 2001; Edwards & Dixon,
2004).Sebbene i membri di famiglie diverse presentino una omologia di sequenza inferiore al 30%, tra quelli della stessa famiglia il valore sale tra il 60 e l’80%
(Hayes & Pulford, 1995; Armstrong, 1997).
In particolare la regione N-terminale è quella che sembra essere maggiormente conservata.
In questa regione è infatti presente un aminoacido importantissimo per l’attivitàcatalitica di questi enzimi ( Tyr per le classi Alpha, Mu, Pi e Sigma; Ser per le
classi Theta e Zeta e da un Cys per le classi Omega e Beta) in quanto capace di stabilizzare la forma anionica del glutatione (GS-) mediante la formazione di un legame idrogeno con il gruppo tiolico (SH) del glutatione stesso (Sheehan et al.,
2001).
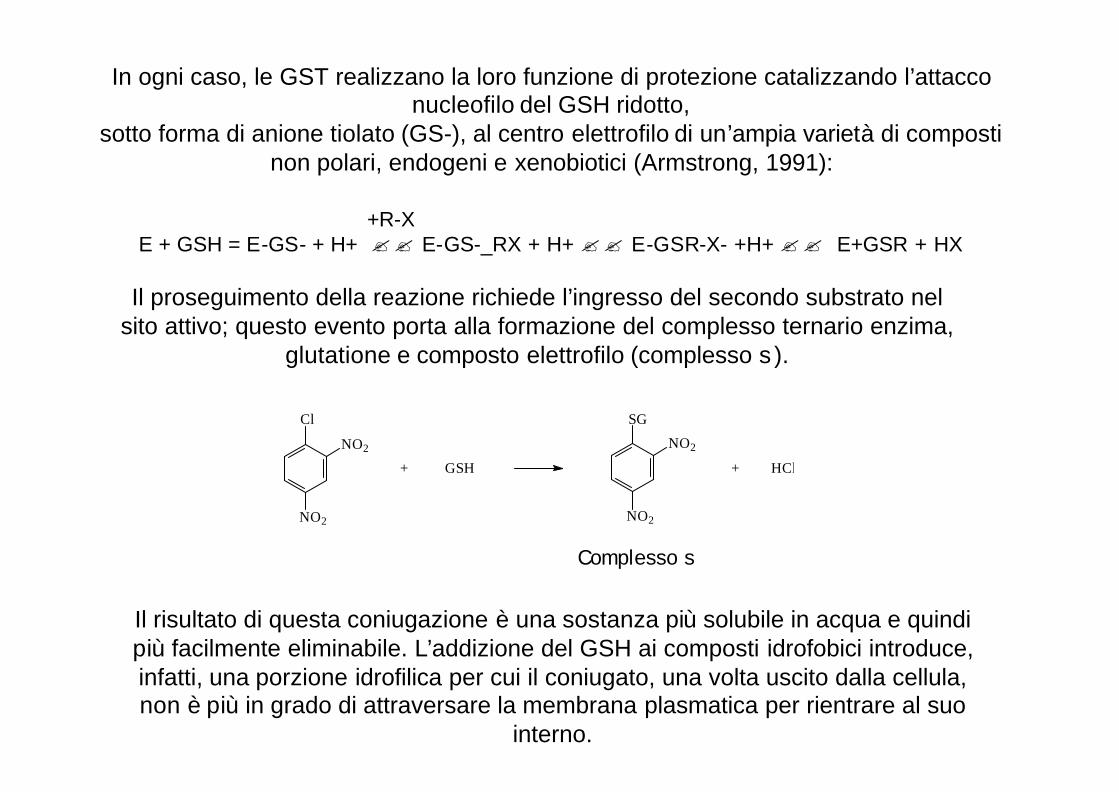
In ogni caso, le GST realizzano la loro funzione di protezione catalizzando l’attacco nucleofilo del GSH ridotto,
sotto forma di anione tiolato (GS-), al centro elettrofilo di un’ampia varietà di composti non polari, endogeni e xenobiotici (Armstrong, 1991):
+R-XE + GSH = E-GS- + H+ ?? E-GS-_RX + H+ ?? E-GSR-X- +H+ ?? E+GSR + HX
Il proseguimento della reazione richiede l’ingresso del secondo substrato nel sito attivo; questo evento porta alla formazione del complesso ternario enzima,
glutatione e composto elettrofilo (complesso s ).
Cl
NO2
NO2
+ GSH
NO2
NO2
SG
+ HCl
Complesso s
Il risultato di questa coniugazione è una sostanza più solubile in acqua e quindi più facilmente eliminabile. L’addizione del GSH ai composti idrofobici introduce, infatti, una porzione idrofilica per cui il coniugato, una volta uscito dalla cellula, non è più in grado di attraversare la membrana plasmatica per rientrare al suo
interno.

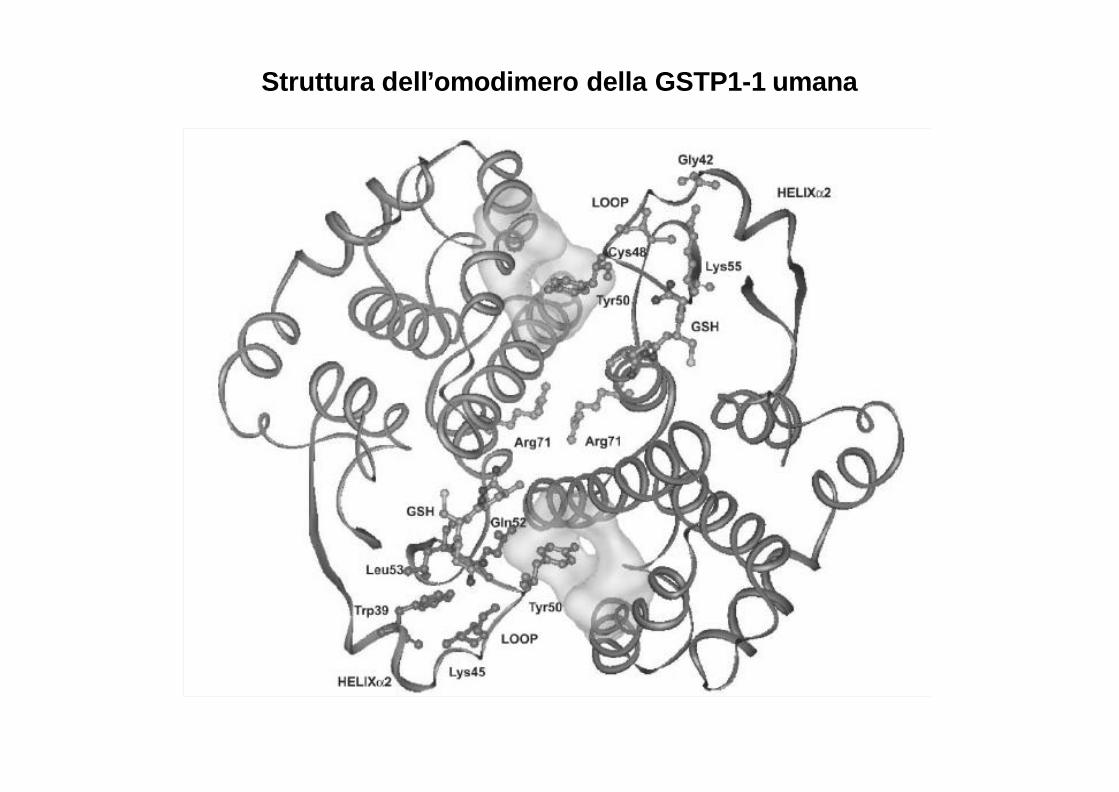
Struttura dell’omodimero della GSTP1-1 umana
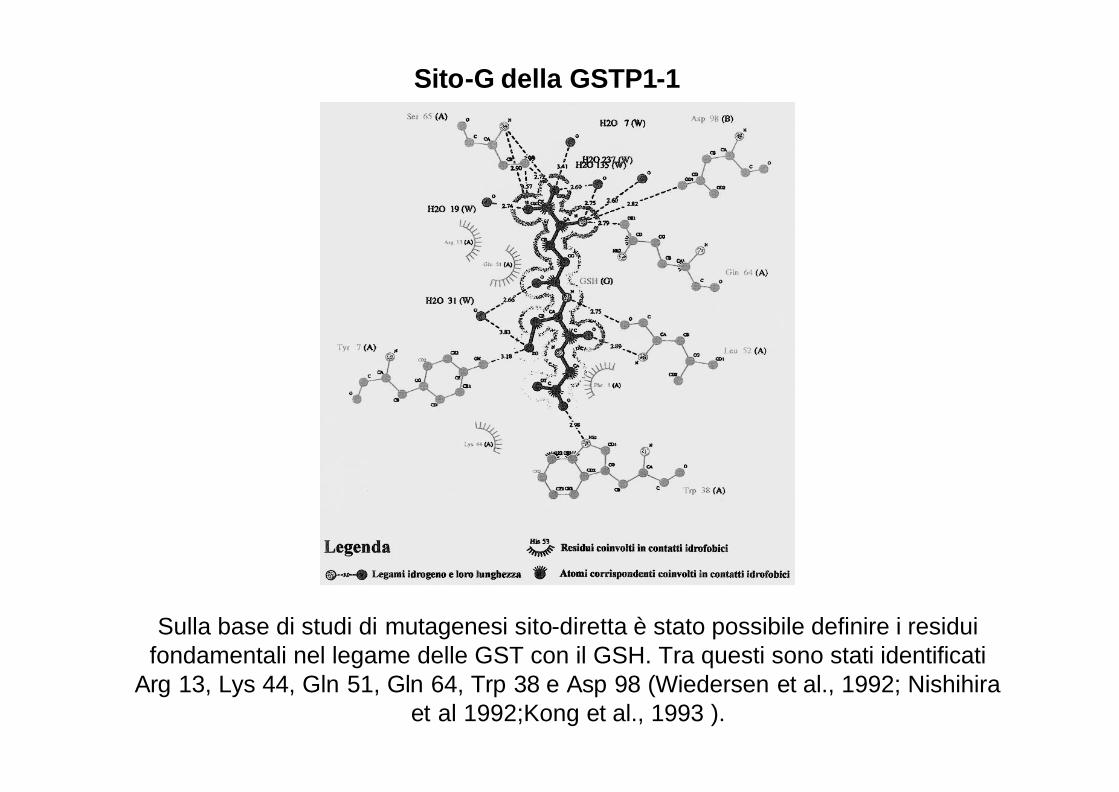
Sulla base di studi di mutagenesi sito-diretta è stato possibile definire i residui fondamentali nel legame delle GST con il GSH. Tra questi sono stati identificati
Arg 13, Lys 44, Gln 51, Gln 64, Trp 38 e Asp 98 (Wiedersen et al., 1992; Nishihiraet al 1992;Kong et al., 1993 ).
Sito-G della GSTP1-1
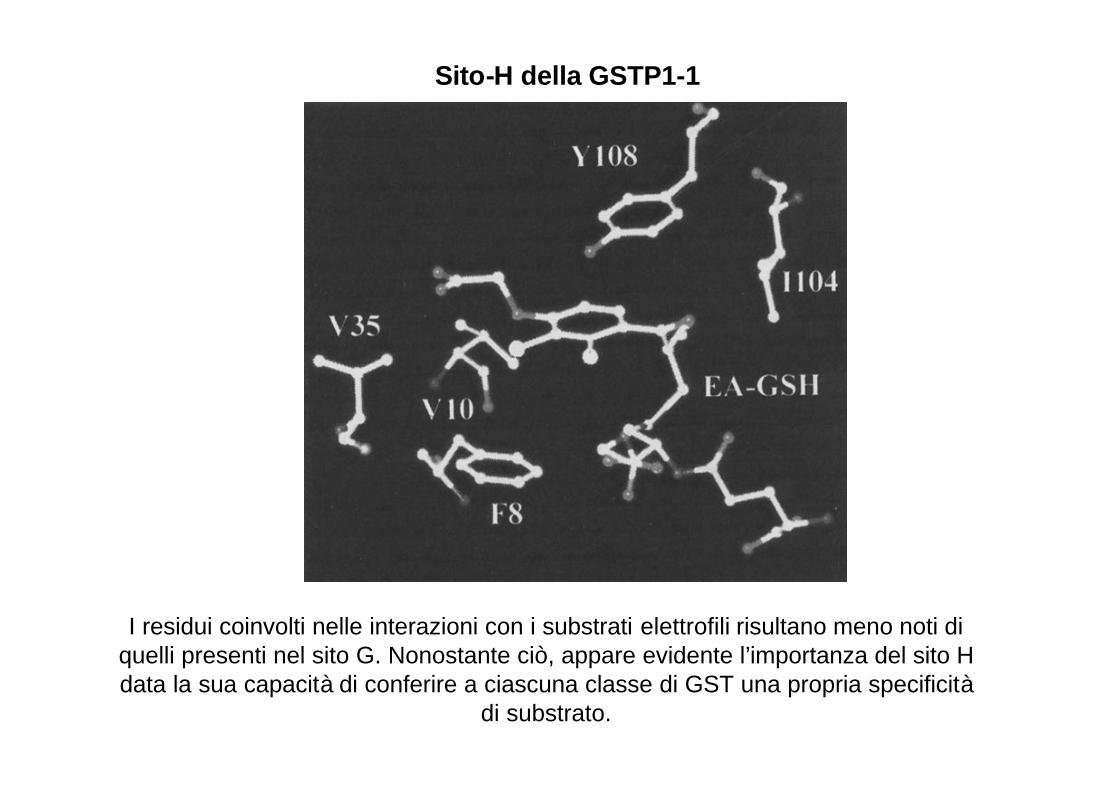
I residui coinvolti nelle interazioni con i substrati elettrofili risultano meno noti di quelli presenti nel sito G. Nonostante ciò, appare evidente l’importanza del sito H data la sua capacità di conferire a ciascuna classe di GST una propria specificità
di substrato.
Sito-H della GSTP1-1

Numerosi studi epidemiologici hanno indagato il ruolo dei polimorfismi delle glutatione trasferasi come determinanti genetici della suscettibilità verso
tumori indotti da carcinogeni endogeni ed esogeni. Probabilmente, il rischio di cancro dovuto alla variazione polimorfica dipende dalla differente abilità di
coniugare e detossificare composti elettrofili e i loro metaboliti. I polimorfismi di GSTM1 e GSTP1 sono stati particolarmente studiati per il
loro coinvolgimento nelle reazioni di detossificazione dei PAH poichémodulano il livello degli addotti DNA-PAH in cellule, come i globuli bianchi,
esposte a composti carcinogenici; individui aventi diverse combinazioni polimorfiche GSTM1 e GSTP1 mostreranno, quindi, una diversa suscettibilità
verso i carcinogeni (Butkiewicz et al., 2000). In particolare sono stati studiati i polimorfismi GSTM1, GSTT1 e GSTP1 e il
loro ruolo come fattori di rischio verso diverse forme di cancro.

A partire dagli anni ‘80 sono stati identificati numerosi polimorfismi a carico delle GST (Hayes et al. 2000). Tali polimorfismi contribuiscono a definire le
differenze interindividuali in risposta a numerosi composti xenobiotici, compresi farmaci e chemioterapici.
I primi studi in tale campo miravano a valutare se vi fosse un maggiore rischio di sviluppare alcuni tipi di tumore, quali quelli del seno, della prostata, del
polmone o il tumore colon-rettale, in individui privi dei geni per la GSTM1-1 e/o per la GSTT1-1 (definiti, rispettivamente, omozigoti per gli alleli GSTM1¯0 e/o GSTT1¯0). Successivamente, la scoperta delle varianti alleliche della GST P1-1 ha portato alla necessità di vagliare l’ipotesi che una maggiore suscettibilitàad alcune patologie potesse dipendere da una combinazione dei polimorfismi
di classe Mu, Pi e Theta delle GST.Ad oggi sono noti polimorfismi per la maggior parte delle GST citosoliche.
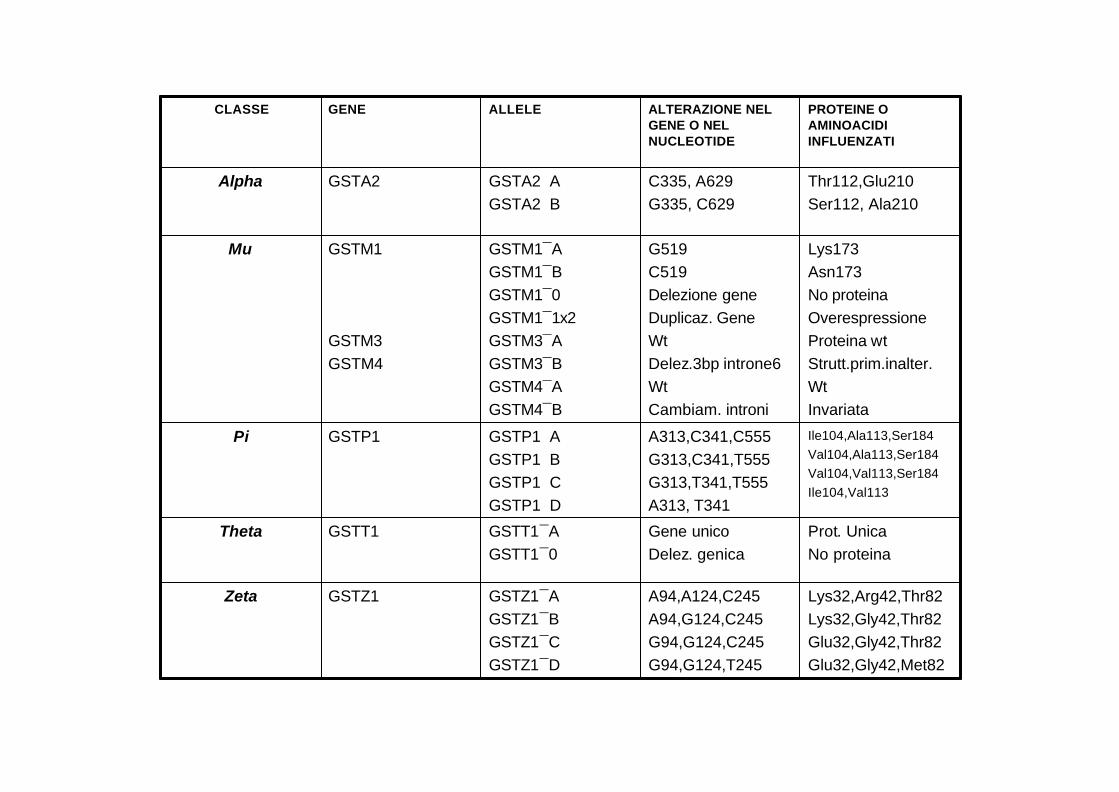
Lys32,Arg42,Thr82Lys32,Gly42,Thr82Glu32,Gly42,Thr82Glu32,Gly42,Met82
A94,A124,C245A94,G124,C245G94,G124,C245G94,G124,T245
GSTZ1¯AGSTZ1¯BGSTZ1¯CGSTZ1¯D
GSTZ1 Zeta
Prot. UnicaNo proteina
Gene unicoDelez. genica
GSTT1¯AGSTT1¯0
GSTT1 Theta
Ile104,Ala113,Ser184Val104,Ala113,Ser184Val104,Val113,Ser184Ile104,Val113
A313,C341,C555G313,C341,T555G313,T341,T555A313, T341
GSTP1� AGSTP1� BGSTP1� CGSTP1� D
GSTP1 Pi
Lys173Asn173No proteinaOverespressioneProteina wtStrutt.prim.inalter.WtInvariata
G519C519Delezione geneDuplicaz. GeneWtDelez.3bp introne6WtCambiam. introni
GSTM1¯AGSTM1¯BGSTM1¯0GSTM1¯1x2GSTM3¯AGSTM3¯BGSTM4¯AGSTM4¯B
GSTM1
GSTM3GSTM4
Mu
Thr112,Glu210Ser112, Ala210
C335, A629G335, C629
GSTA2� AGSTA2� B
GSTA2 Alpha
PROTEINE O AMINOACIDI INFLUENZATI
ALTERAZIONE NEL GENE O NEL NUCLEOTIDE
ALLELEGENECLASSE

Classe Alpha
Questa isoforma è principalmente espressa nel fegato ed è codificata da un cluster genico localizzato sul cromosoma 6p12; tale cluster contiene 5
geni che codificano per altrettante proteine (GSTA1-5).Un polimorfismo del gene GSTA1 è caratterizzato da due polimorfismi
(*A, *B) che presentano tre sostituzioni nucleotidiche a livello dei residui 567, 69 e 52, presenti sul promotore.
GSTA3-3 è selettivamente espressa tessuti steroidogenici e gioca un ruolo fondamentale nella biosintesi degli ormoni steroidei. Polimorfismi
nel gene GSTA3 possono influenzare la steroidogenesi e alterare i livelli proteici o la funzione proteica; inoltre, è stato ipotizzato che alterazioni in geni implicati nella sintesi degli ormoni steroidei e nel metabolismo degli ormoni sessuali possano potenziare la suscettibilità al tumore ovarico.
Thr112,Glu210Ser112, Ala210
C335, A629G335, C629
GSTA2� AGSTA2� B GSTA2 Alpha
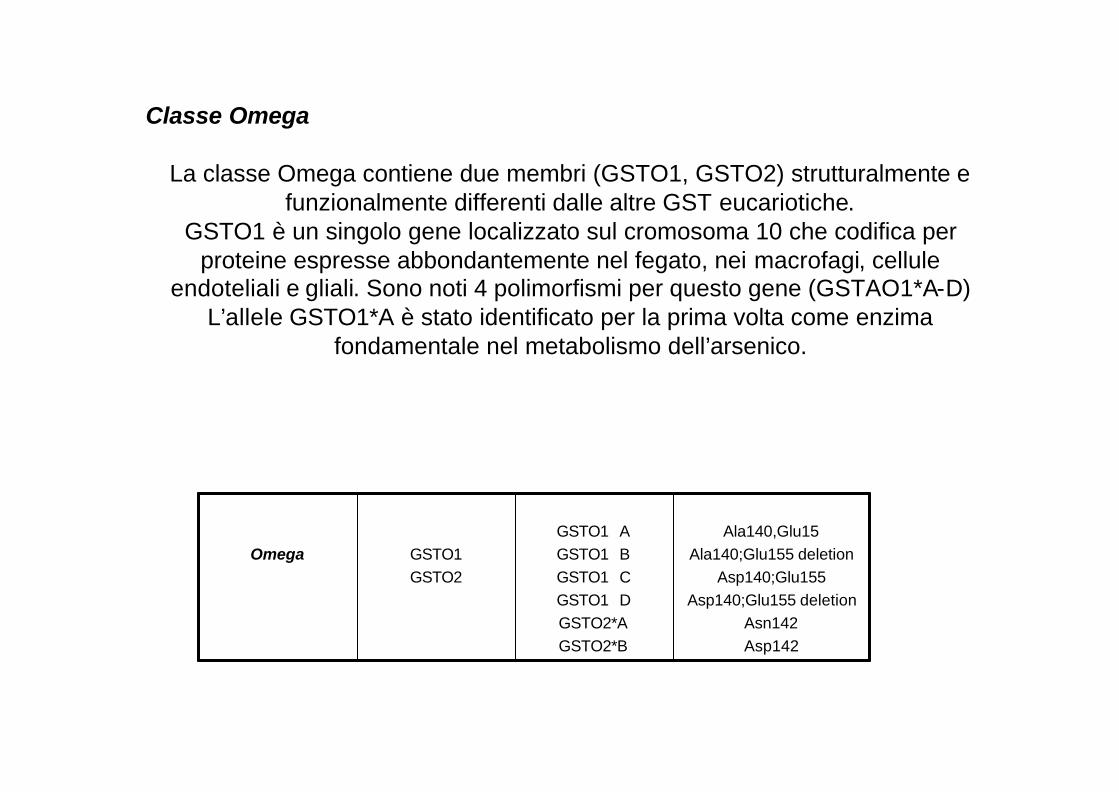
Classe Omega
La classe Omega contiene due membri (GSTO1, GSTO2) strutturalmente e funzionalmente differenti dalle altre GST eucariotiche.
GSTO1 è un singolo gene localizzato sul cromosoma 10 che codifica per proteine espresse abbondantemente nel fegato, nei macrofagi, cellule
endoteliali e gliali. Sono noti 4 polimorfismi per questo gene (GSTAO1*A-D)L’allele GSTO1*A è stato identificato per la prima volta come enzima
fondamentale nel metabolismo dell’arsenico.
Omega GSTO1GSTO2
GSTO1� AGSTO1� BGSTO1� CGSTO1� D GSTO2*AGSTO2*B
Ala140,Glu15Ala140;Glu155 deletion
Asp140;Glu155Asp140;Glu155 deletion
Asn142Asp142
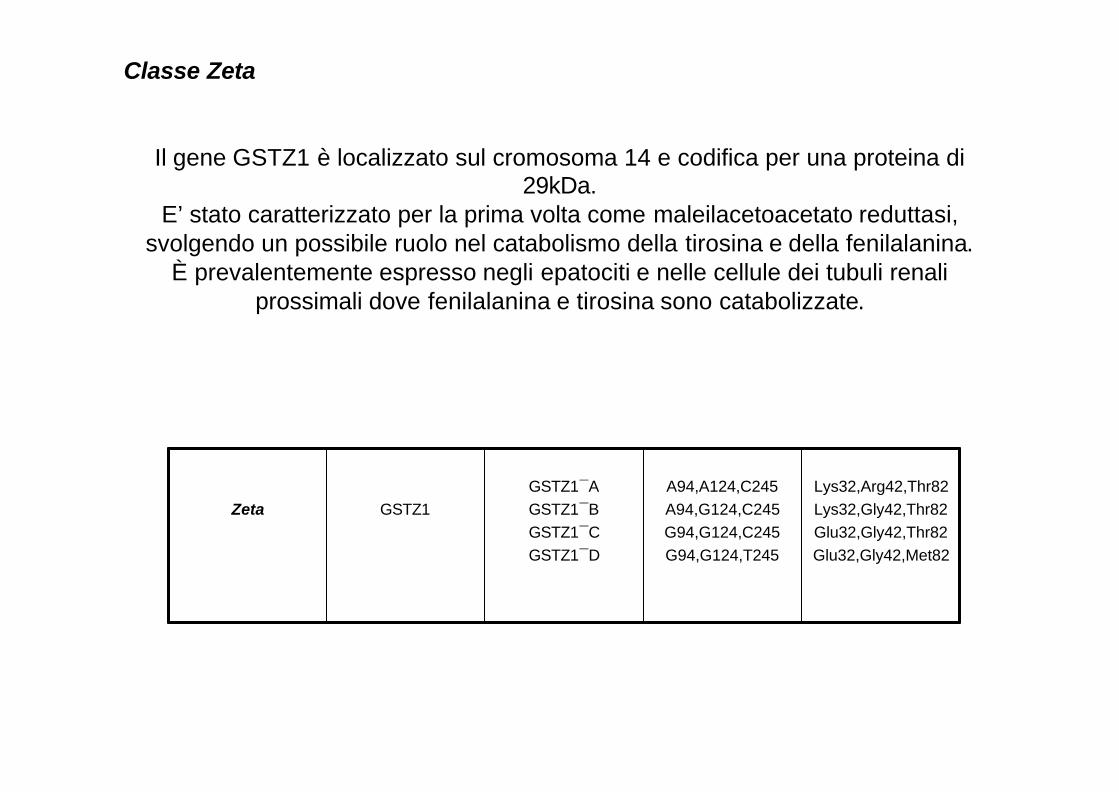
Classe Zeta
Il gene GSTZ1 è localizzato sul cromosoma 14 e codifica per una proteina di 29kDa.
E’ stato caratterizzato per la prima volta come maleilacetoacetato reduttasi, svolgendo un possibile ruolo nel catabolismo della tirosina e della fenilalanina.
È prevalentemente espresso negli epatociti e nelle cellule dei tubuli renali prossimali dove fenilalanina e tirosina sono catabolizzate.
Lys32,Arg42,Thr82Lys32,Gly42,Thr82Glu32,Gly42,Thr82Glu32,Gly42,Met82
A94,A124,C245A94,G124,C245G94,G124,C245G94,G124,T245
GSTZ1¯AGSTZ1¯BGSTZ1¯CGSTZ1¯D
GSTZ1Zeta

Classe MuIl gene GSTM1 contiene 4 alleli. Le varianti GSTM1*A e *B sono identiche dal punto di
vista funzionale e differiscono tra loro per una sola sostituzione aminoacidica in posizione 173 (K/N). La presenza dell’allele *A è stata associata con un decrementato
rischio di cancro della vescica.Al contrario, invece, il polimorfismo Mu null (Mu1*0) è associato con un maggiore
rischio di tumore a polmoni, colon e vescica; tuttavia pazienti con questo polimorfismo affetti da Leucemia Mieloide Acuta (AML) hanno mostrato una migliore risposta al
trattamento chemioterapico con Adriamicina e Ciclofosfamide.L’allele GSTM2*B sembra catalizzare la coniugazione del GSH all’aminocromo, un prodotto del ciclo redox delle dopamine (catecolamine); poiché i prodotti di questi
processi sono associati a malattie neurodegenerative (Parkinson), la GSTM*2 potrebbe svolgere un ruolo citoprotettivo nei confronti di tale patologia.
Lys173Asn173
No proteinaOverespressione
Proteina wtStrutt.prim.inalter.
WtInvariata
G519C519
Delezione geneDuplicaz. Gene
WtDelez.3bp introne6
WtCambiam. introni
GSTM1¯AGSTM1¯BGSTM1¯0
GSTM1¯1x2GSTM3¯AGSTM3¯BGSTM4¯AGSTM4¯B
GSTM1
GSTM3GSTM4
Mu

La mancanza dell’enzima GSTM1 è considerata un fattore di rischio per l’insorgenza del cancro ai polmoni (Hirvonen et al., 1993), alla
pelle (Heagerty et al., 1994), alla vescica (Bell et al., 1993) e al colon (Zhong et al., 1993) a causa della ridotta efficienza nel legame dei
substrati genotossici (Mc Williams et al., 1995).Poiché la GSTM1-1 catalizza il metabolismo di un gran numero di
composti potenzialmente genotossici, molti studi sugli effetti di questo polimorfismo genetico hanno riguardato la formazione di addotti di DNA e l’insorgenza di danni citogenetici. Alti livelli di addotti PAH-DNA sono stati trovati in campioni di polmone di donatori (fumatori) aventi genotipo GSTM1-1 null, rispetto ad
individui con genotipo GSTM1-1 positivo (Butkiewicz et al., 2000). Recentemente è stato scoperto che il numero di addotti di DNA
presenti nella placenta umana di individui con genotipo GSTM1-1 null è elevato se paragonato a quello di individui con genotipo
GSTM1-1 positivo (Sorsa et al., 1996). Da uno studio su un campione di popolazione Statunitense costituito da soggetti di origine europea e da afro-americani (Chun-Lin Chen
et al., 1996) è emerso che circa il 53.5% dei bianchi possiede genotipo GSTM1 null, contro il 27.6% dei neri, in accordo con i dati
riportati precedentemente da Bell et al. nel 1992.

Classe Theta
La classe Theta consiste di due sottofamiglie, GSTT1 e GSTT2, i cui geni sono localizzati sul cromosoma 22. Esistono polimorfismi per entrambi questi membri
compreso il tipo GSTT1*0, che presenta una ridotta attività catalitica ed èassociato con un aumentato rischio di tumore alla testa, al collo e alla cavità
orale.Una singola sostituzione nucleotidica localizzata sull’esone 3 porta allo sviluppo
di due polimorfismi: GSTT1*A (Treonina 104) e GSTT1*B (Prolina 104)
Prot. UnicaNo proteina
Gene unicoDelez. genica
GSTT1¯AGSTT1¯0 GSTT1 Theta
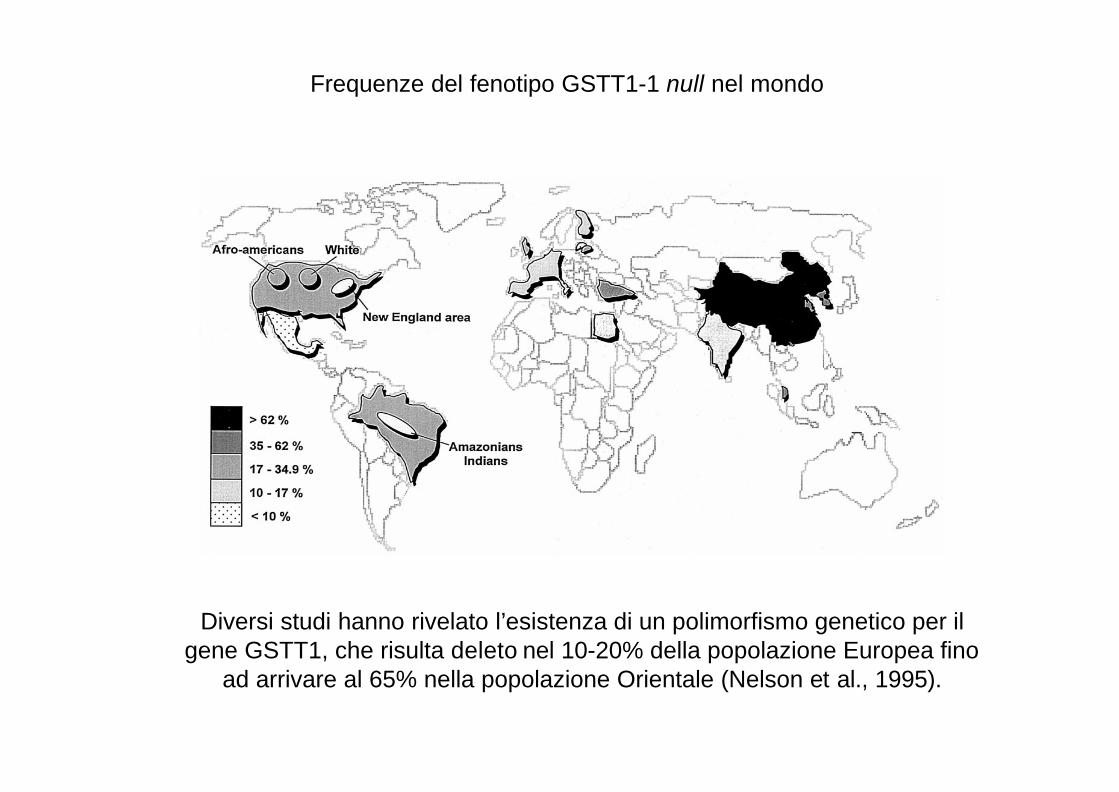
Frequenze del fenotipo GSTT1-1 null nel mondo
Diversi studi hanno rivelato l’esistenza di un polimorfismo genetico per il gene GSTT1, che risulta deleto nel 10-20% della popolazione Europea fino
ad arrivare al 65% nella popolazione Orientale (Nelson et al., 1995).

Gli effetti del polimorfismo GSTT1 sono stati oggetto di studi epidemiologici volti a determinare se nell’uomo la presenza o meno del
fenotipo GSTT1-1 null è associata alla suscettibilità al cancro, in relazione all’esposizione verso sostanze tossiche. In generale, nella popolazione Europea gli individui fumatori con genotipo GSTT1-nullsono considerati ad alto rischio per lo sviluppo di tumori all’apparato
respiratorio; diversi studi, inoltre, hanno dimostrato che la presenza del genotipo GSTT1-GSTM1 doppio null aumenta di 3 volte, rispetto al
genotipo GSTT1-GSTM1 doppio positivo, il rischio di tumori (Jourenkova-Mironova et al., 1999).
La GSTT1-1 sembra giocare un ruolo importante nello sviluppo del cancro alla cavità orale e alla laringe in Cina e in India, dove il
genotipo GSTT1-null ha la frequenza più alta nel mondo e dove sono diverse le abitudini alimentari ed il modo di fumare. Al contrario, il gene GSTM1 è un importante fattore di rischio di cancro nei paesi
industrializzati, dove il consumo di alcol e carne è alto, la frequenza del genotipo GSTT1-null è bassa e il fumo di sigaretta è la più
comune fonte di esposizione (Nair et al., 1999).

Classe Pi
La GSTPi è codificata da un singolo gene localizzato sul cromosoma 11 e presenta 4 polimorfismi attivi, ma funzionalmente differenti (GSTP1 *A-*D). Il
genotipo GSTP1 è associato con differenti risposte al trattamento chemioterapicoed è overespresso in tumori quali quello del seno, colon, avaie, sistema nervoso
centrale, pancreas e linfoma.La sostituzione I104V porta al polimorfismo *B che risulta in una sostanziale riduzione dell’attività catalitica e una diminuita capacità di detossificazione.L’allele *C (I104V/A113V) è predominante nelle cellule di glioma maligno.
L’allele *D presenta una singola mutazione in posizione 113 (A?V).
Ile104,Ala113,Ser184Val104,Ala113,Ser184Val104,Val113,Ser184Ile104,Val113
A313,C341,C555G313,C341,T555G313,T341,T555A313, T341
GSTP1� AGSTP1� BGSTP1� CGSTP1� D
GSTP1 Pi

• Tra le principali conseguenze biologiche a carico dei polimorfismi della GST P1-1, di particolare importanza risultano essere quelle riguardanti la
suscettibilità ai tumori e lo sviluppo di chemioresistenza.
Pazienti omozigoti per l’allele GSTP1¯B presentano una ridotta capacità di detossificare gli agenti chemioterapici a base di platino (Stoelmacher et al., 2002) e non solo. La sostituzione Ile104Val nell’allele GSTP1¯B comporta
una sostanziale riduzione dell’attività catalitica dell’enzima e una ridotta capacità di detossificazione negli individui che la portano (Watson et al., 1998). GSTP1¯C, la variante allelica dominante nelle cellule di glioma
maligno, sembra conferire agli individui che la portano una minore incidenza del cancro del seno (Maugard et al., 2001).
Da alcuni anni a questa parte si è diffuso l’uso di programmi terapeutici che prevedono l’associazione dei chemioterapici con la radioterapia e che
garantiscono una maggiore possibilità di sopravvivenza ai pazienti su cui vengono attuati. Tuttavia, nel lungo termine, pazienti con varianti alleliche
*B,*C e *D della GSTP1-1 hanno un aumentato rischio di sviluppare tumori secondari alla terapia stessa tra cui la leucemia mieloide acuta (AML) è il più comune (Kaldor et al., 1990; Travis et al., 1996; Travis et al., 1999;
Travis et al., 2000; Woldon et al., 1999).
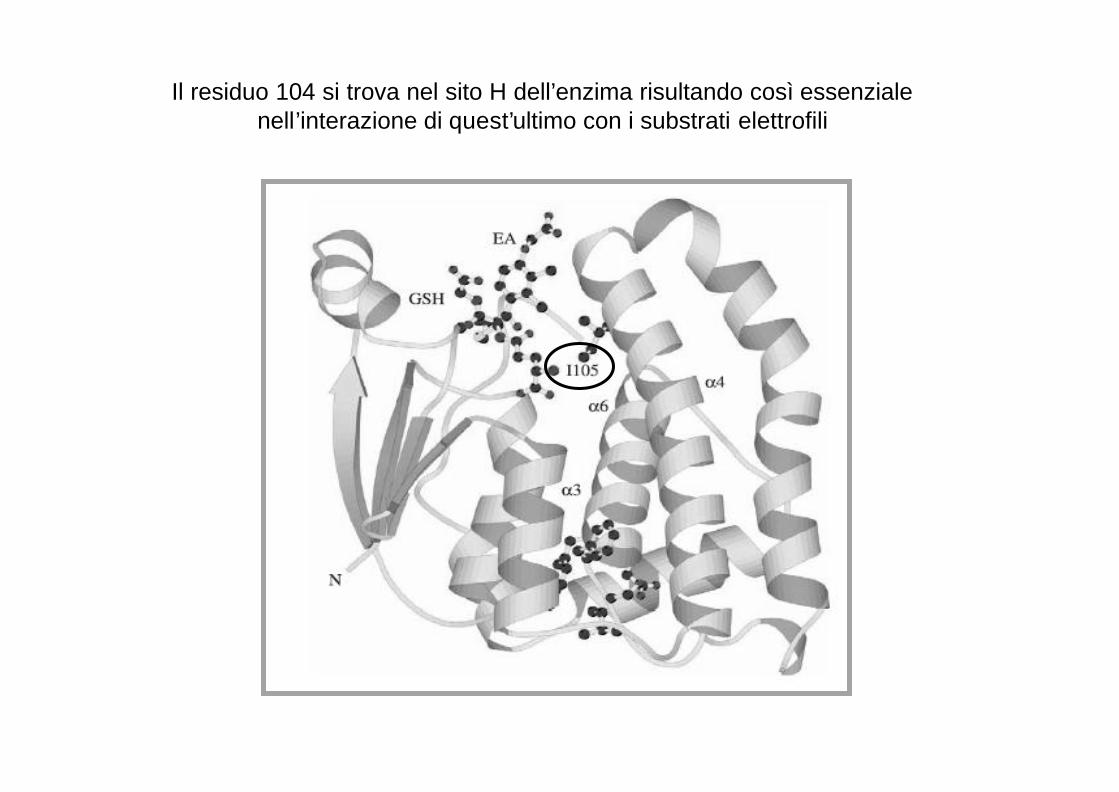
Il residuo 104 si trova nel sito H dell’enzima risultando così essenziale nell’interazione di quest’ultimo con i substrati elettrofili
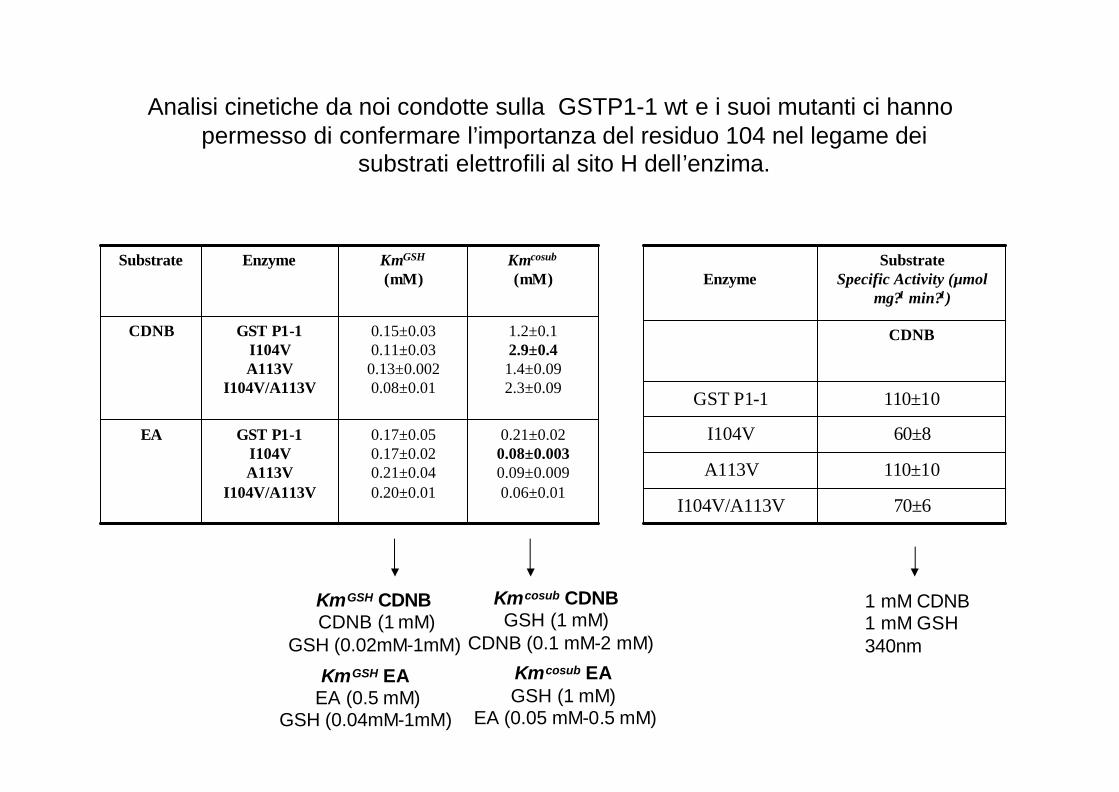
Analisi cinetiche da noi condotte sulla GSTP1-1 wt e i suoi mutanti ci hanno permesso di confermare l’importanza del residuo 104 nel legame dei
substrati elettrofili al sito H dell’enzima.
0.21±0.020.08±0.0030.09±0.0090.06±0.01
0.17±0.050.17±0.020.21±0.040.20±0.01
GST P1-1I104VA113V
I104V/A113V
EA
1.2±0.12.9±0.41.4±0.092.3±0.09
0.15±0.030.11±0.030.13±0.0020.08±0.01
GST P1-1I104VA113V
I104V/A113V
CDNB
Kmcosub
(mM)KmGSH
(mM)EnzymeSubstrate
70±6I104V/A113V
110±10A113V
60±8I104V
110±10GST P1-1
CDNB
Substrate Specific Activity (µmol
mg?1 min?1)Enzyme
1 mM CDNB1 mM GSH340nm
KmGSH CDNBCDNB (1 mM)
GSH (0.02mM-1mM)
Kmcosub CDNBGSH (1 mM)
CDNB (0.1 mM-2 mM)
KmGSH EAEA (0.5 mM)
GSH (0.04mM-1mM)
Kmcosub EAGSH (1 mM)
EA (0.05 mM-0.5 mM)
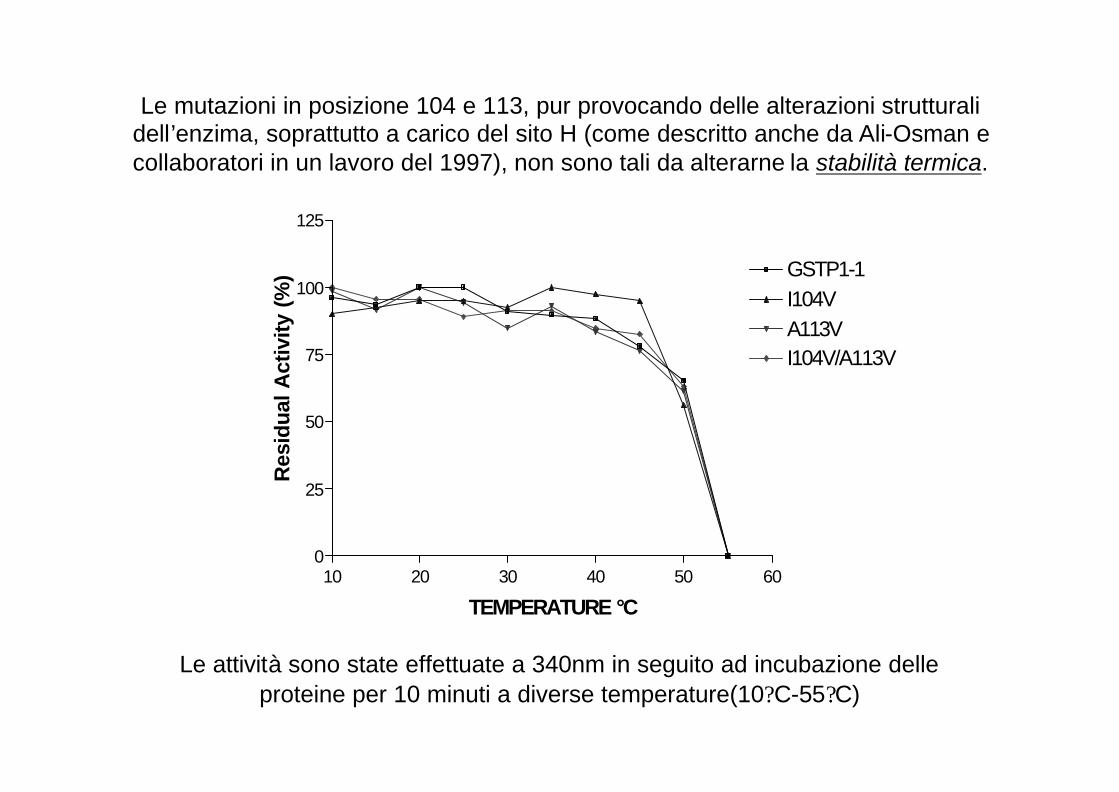
Le mutazioni in posizione 104 e 113, pur provocando delle alterazioni strutturali dell’enzima, soprattutto a carico del sito H (come descritto anche da Ali-Osman e collaboratori in un lavoro del 1997), non sono tali da alterarne la stabilità termica.
10 20 30 40 50 600
25
50
75
100
125
GSTP1-1I104VA113VI104V/A113V
TEMPERATURE °C
Res
idua
l Act
ivity
(%
)
Le attività sono state effettuate a 340nm in seguito ad incubazione delle proteine per 10 minuti a diverse temperature(10?C-55?C)
N
Cl
Cl
O
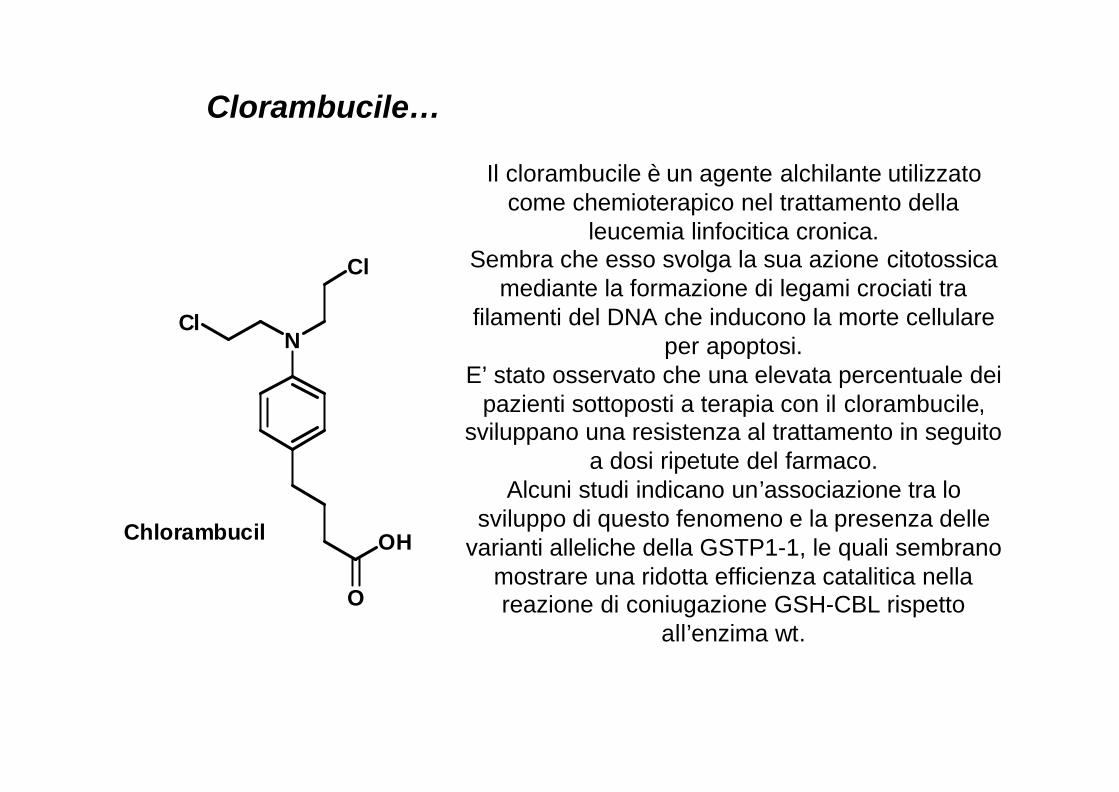
OHChlorambucil
Il clorambucile è un agente alchilante utilizzatocome chemioterapico nel trattamento della
leucemia linfocitica cronica.Sembra che esso svolga la sua azione citotossica
mediante la formazione di legami crociati tra filamenti del DNA che inducono la morte cellulare
per apoptosi.E’ stato osservato che una elevata percentuale dei
pazienti sottoposti a terapia con il clorambucile, sviluppano una resistenza al trattamento in seguito
a dosi ripetute del farmaco.Alcuni studi indicano un’associazione tra lo
sviluppo di questo fenomeno e la presenza delle varianti alleliche della GSTP1-1, le quali sembrano
mostrare una ridotta efficienza catalitica nella reazione di coniugazione GSH-CBL rispetto
all’enzima wt.
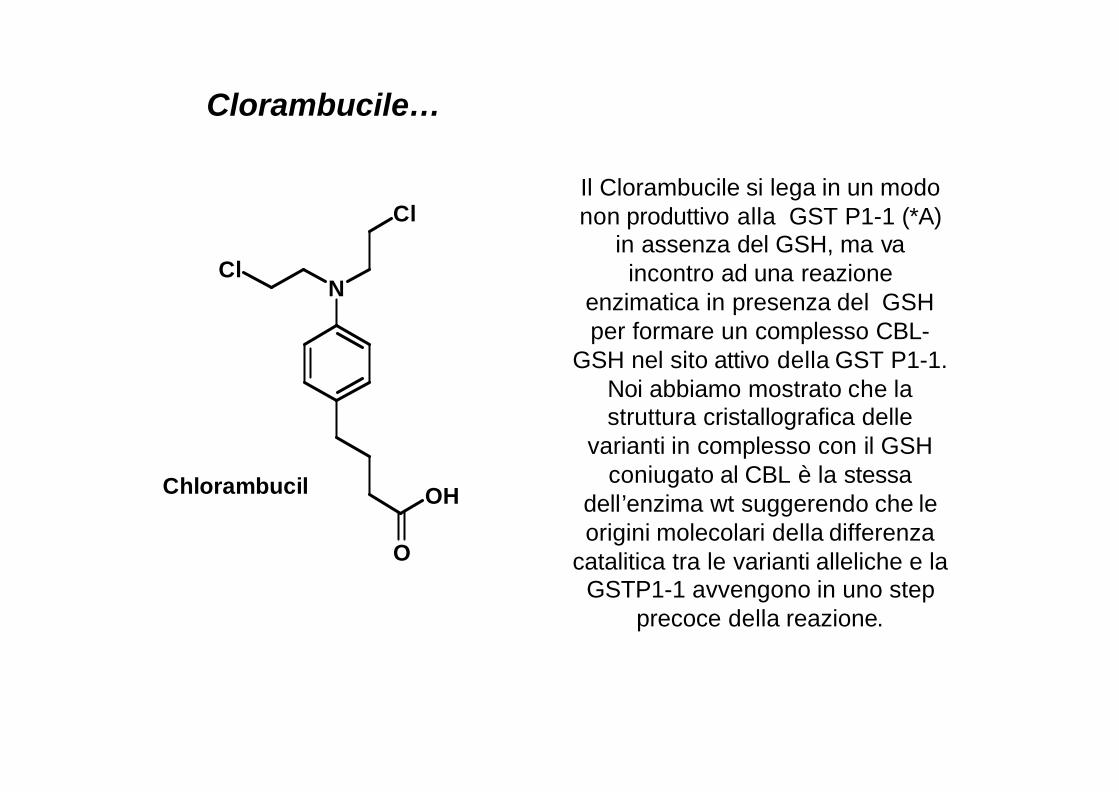
Clorambucile…
N
Cl
Cl
O
OHChlorambucil
Clorambucile…
Il Clorambucile si lega in un modo non produttivo alla GST P1-1 (*A)
in assenza del GSH, ma vaincontro ad una reazione
enzimatica in presenza del GSH per formare un complesso CBL-
GSH nel sito attivo della GST P1-1. Noi abbiamo mostrato che la struttura cristallografica delle
varianti in complesso con il GSH coniugato al CBL è la stessa
dell’enzima wt suggerendo che le origini molecolari della differenza
catalitica tra le varianti alleliche e la GSTP1-1 avvengono in uno step
precoce della reazione.
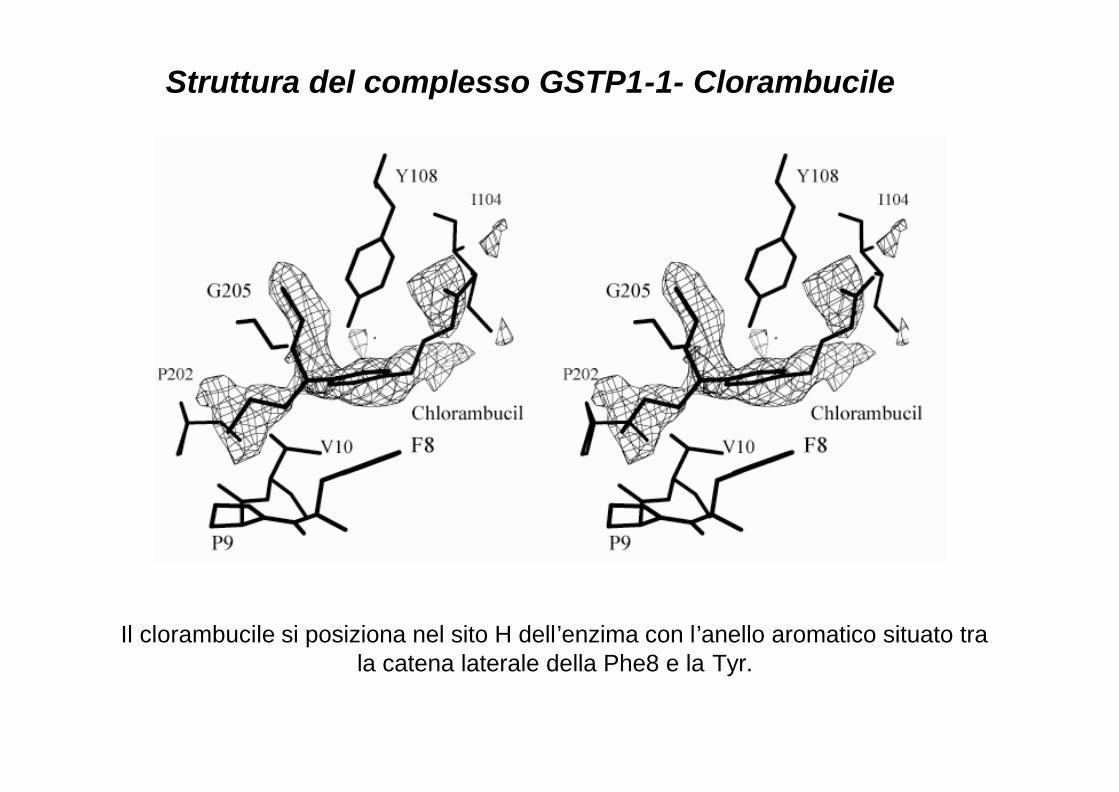
Il clorambucile si posiziona nel sito H dell’enzima con l’anello aromatico situato tra la catena laterale della Phe8 e la Tyr.
Struttura del complesso GSTP1-1- Clorambucile
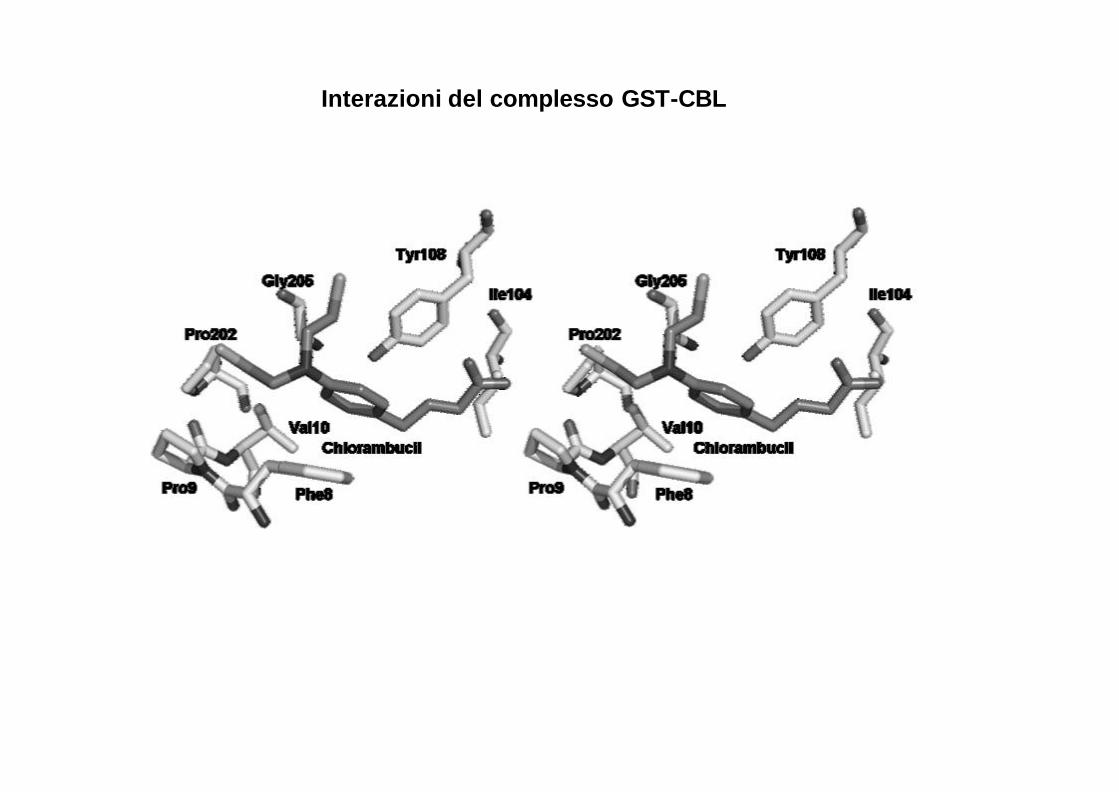
Interazioni del complesso GST-CBL
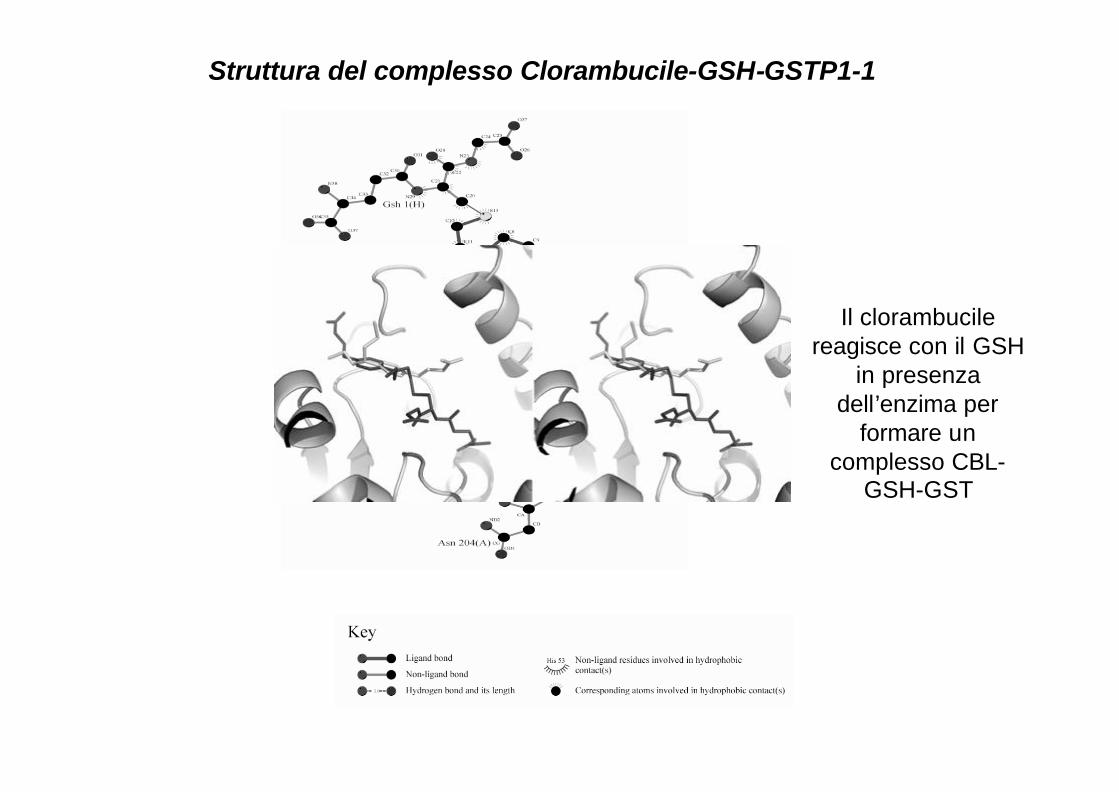
Struttura del complesso Clorambucile-GSH-GSTP1-1
Il clorambucilereagisce con il GSH
in presenzadell’enzima per
formare un complesso CBL-
GSH-GST
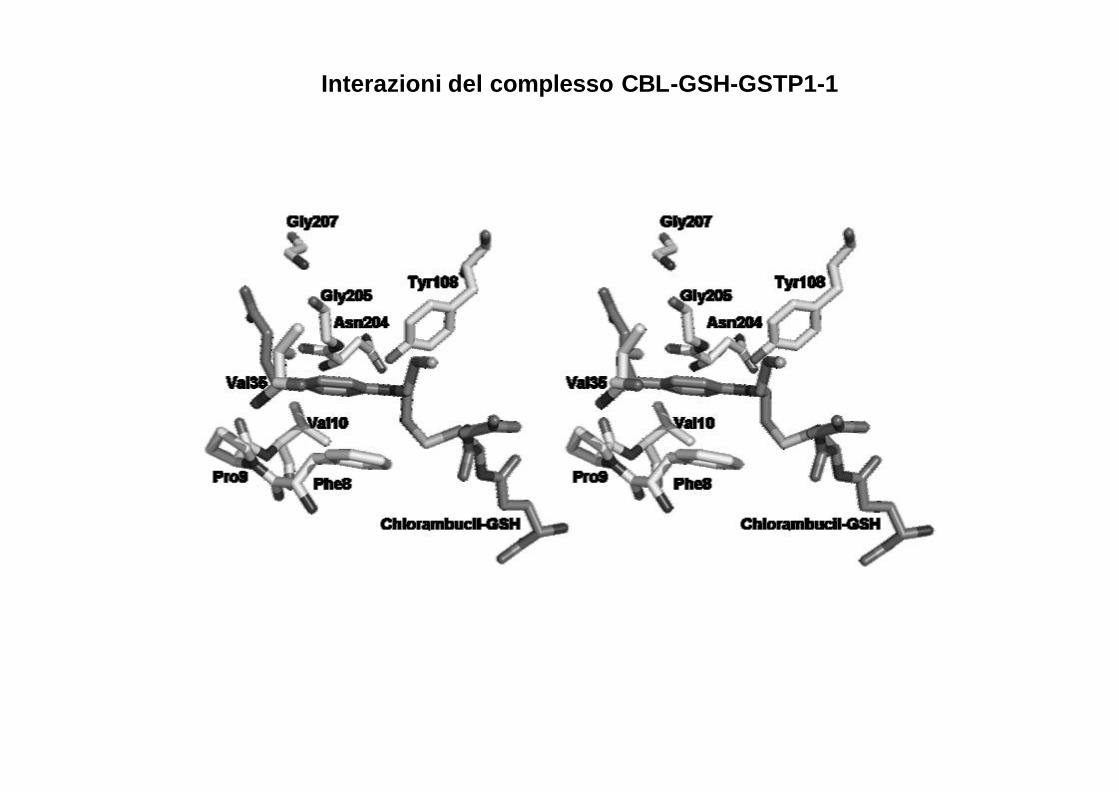
Interazioni del complesso CBL-GSH-GSTP1-1
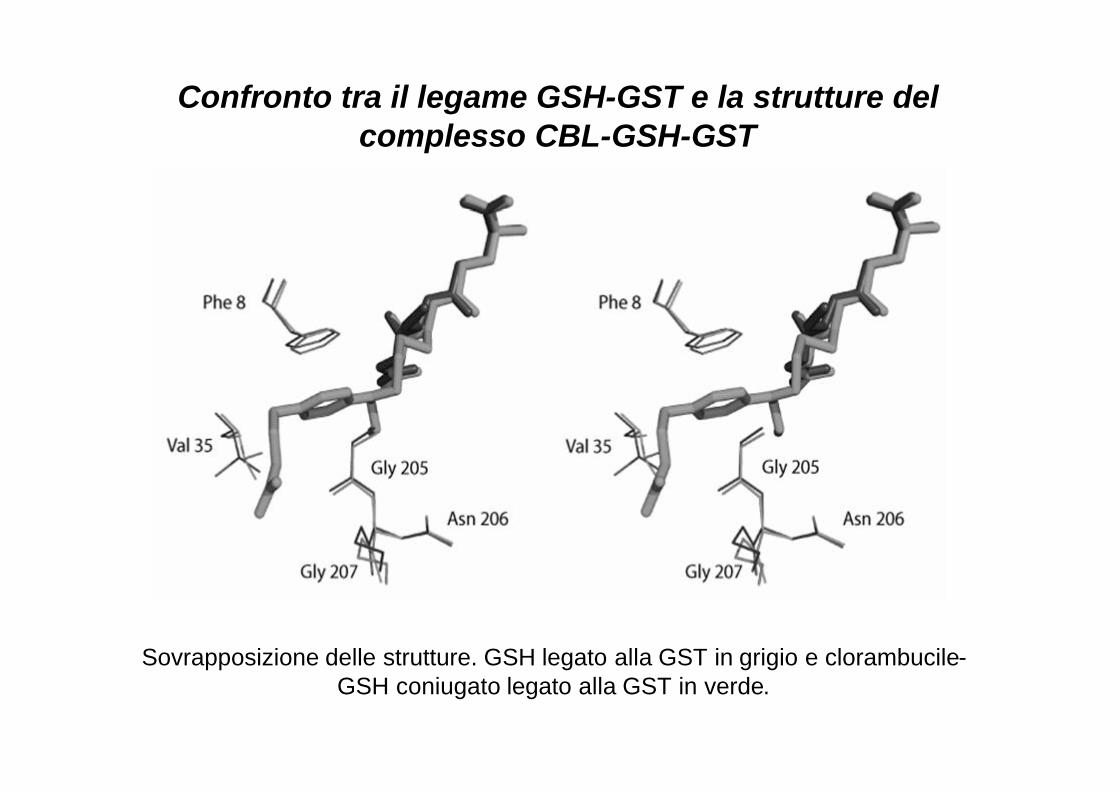
Sovrapposizione delle strutture. GSH legato alla GST in grigio e clorambucile-GSH coniugato legato alla GST in verde.
Confronto tra il legame GSH-GST e la strutture del complesso CBL-GSH-GST
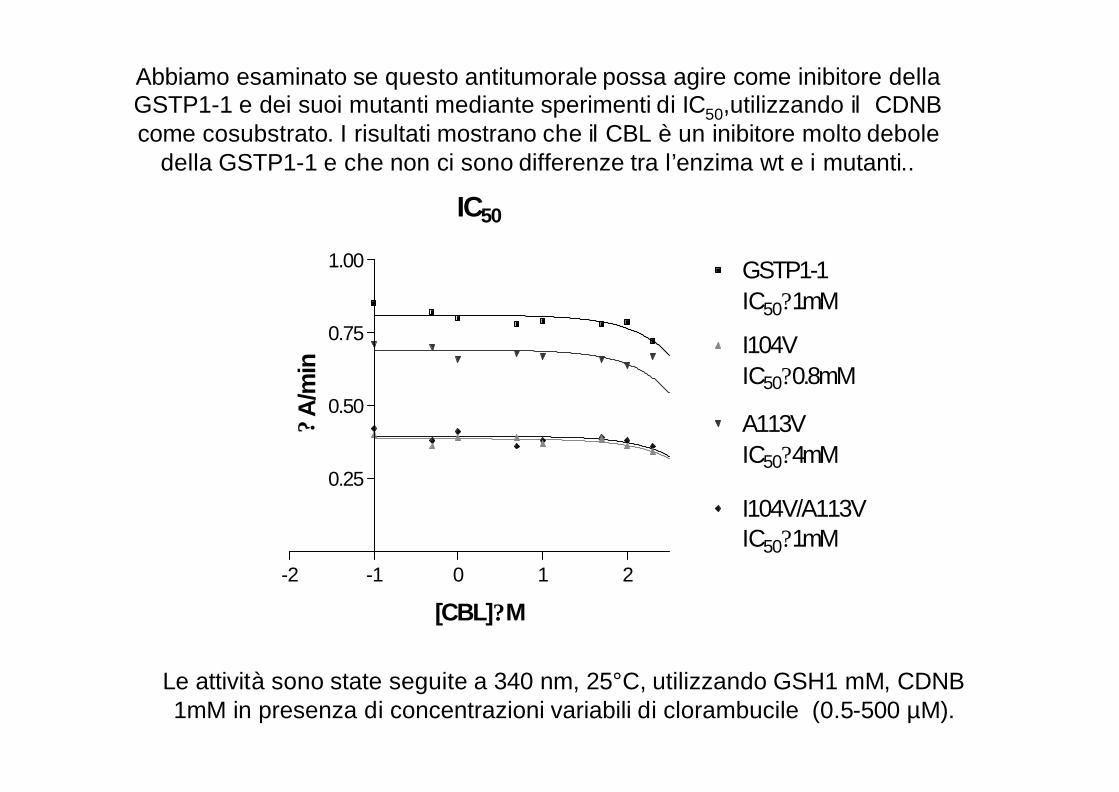
IC50
-2 -1 0 1 2
0.25
0.50
0.75
1.00 GSTP1-1IC50?1mM
I104VIC50?0.8mM
A113VIC50?4mM
I104V/A113VIC50?1mM
[CBL]?M
?A
/min
Abbiamo esaminato se questo antitumorale possa agire come inibitore dellaGSTP1-1 e dei suoi mutanti mediante sperimenti di IC50,utilizzando il CDNB come cosubstrato. I risultati mostrano che il CBL è un inibitore molto debole
della GSTP1-1 e che non ci sono differenze tra l’enzima wt e i mutanti..
Le attività sono state seguite a 340 nm, 25°C, utilizzando GSH1 mM, CDNB 1mM in presenza di concentrazioni variabili di clorambucile (0.5-500 µM).

Conclusioni…
Il CBL si lega al sito H della GSTP1-1 wt e dei suoi mutanti (I104V, A113V e I104V/A113V)
Questi enzimi sono capaci di catalizzare la reazione di coniugazione tra il GSH e il CBL in modo tale che il prodotto sia osservato nel sito attivo con il
clorambucile posizionato nel sito-H e il GSH nel sito G.
Le varianti alleliche della GST P1-1 vanno incontro alla stessa reazione con il clorambucile nel cristallo nonostante presentino una ridotta efficienza catalitica.
Il clorambucile è un inibitore debole della GSTP1-1 e non sono rilevate differenze tra l’enzima wt e i suoi mutanti.







![Farmaci corticosteroidei [modalità compatibilità] · FARMACI GLUCOCORTICOSTEROIDEI Cl ifi iClassificazione Anti-infiammotori steroidei Immunosoppressori. Ormoni steroidei Attivi](https://static.fdocumenti.com/doc/165x107/5c674ee509d3f22d638baad4/farmaci-corticosteroidei-modalita-compatibilita-farmaci-glucocorticosteroidei.jpg)










