

TESTI OPACHI-progetto finale del Corso Cognizione e Linguaggio

SOMMARIO Lezione del 5/11/2015:
Quadro generale
• Cenni di morfologia delle aree cerebrali
• Le aree motorie (nella scimmia e nell’uomo) – Neuroni canonici e affordances – Neuroni specchio
• Neuroni specchio, leLura delle intenzioni ed empaMa
• “Embodied cogni2on” e teoria della mente

SOMMARIO Lezione del 6/11/2015: Perché la scoperta dei neuroni specchio dovrebbe interessare al linguista? In che modo influenza la teoria del linguaggio?
• Filogenesi del linguaggio
• Ontogenesi del linguaggio
– Azione/Gesto/Linguaggio (studi di V. Volterra e ISTC-‐CNR)
– Case study acquisizione aMpica: l’auMsmo (V. Gallese; L. Brandi e S. Lucchesini)
• Neuroni specchio, percezione e comprensione linguisMca
– Liv. Fonologico (“Motor theory of speech percep2on”) – Liv. Lessicale-‐semanMco

Movimento, percezione, cognizione (Rizzola] e Sinigaglia, 2006)
“Per decenni ha dominato l’idea che le aree motorie della corteccia cerebrale sarebbero desMnate a compiM meramente esecuMvi, privi di alcuna valenza perce]va e, meno che mai, cogniMva” Schema tradizionale: Percezione è cognizione è movimento “Uno schema del genera poteva risultare convincente finché del sistema motorio si aveva un’immagine estremamente semplificata. Ma oggi non è più così. Sappiamo che tale sistema è formato da un mosaico di aree frontali e parietali streLamente connesse con le aree visive, udiMve, ta]li, e dotate di proprietà funzionali molto più complesse di quanto si potesse aspeLare” “Lo stesso rigido confine tra processi perce]vi, cogniMvi e motori finisce per rivelarsi in gran parte arMficioso: non solo la percezione appare immersa nella dinamica dell’azione […], ma il cervello che agisce è anche innanzituLo un cervello che comprende. Si traLa, come vedremo, di una comprensione pragmaMca, preconceLuale e prelinguisMca, e tuLavia non meno importante, poiché su di essa poggiano molte delle nostre tanto celebrate capacità cogniMve.”
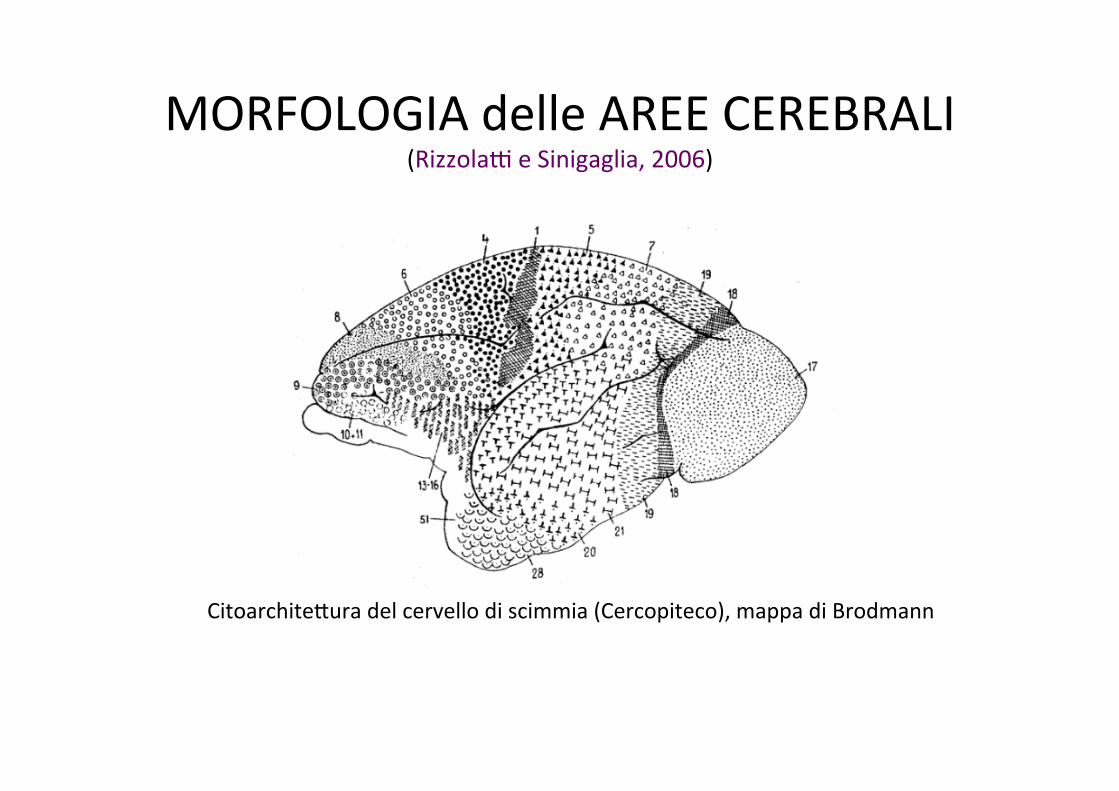
MORFOLOGIA delle AREE CEREBRALI (Rizzola] e Sinigaglia, 2006)
CitoarchiteLura del cervello di scimmia (Cercopiteco), mappa di Brodmann

MORFOLOGIA delle AREE CEREBRALI
CitoarchiteLura del cervello umano, mappa di Brodmann

AREE MOTORIE (nella scimmia) Aree 4 e 6 di Brodmann, “corteccia agranulare” • Area 4, corteccia motoria
primaria, F1 • Area 6, disMnta in 3 regioni
principali (ciascuna divisa a sua volta in una parte rostrale ed una caudale): – Regione Mesiale: F3, F6 – Regione Dorsale: F2, F7 – Regione Ventrale: F4, F5

AREE MOTORIE (nell’uomo) Aree 4 e 6 di Brodmann • Area 4, corteccia motoria primaria, M1
• Area 6: – Area PREMOTORIA (APM) – Area Motoria Supplementare (AMS)

AREE MOTORIE Area 4
• Controllo del movimento volontario • Organizzazione somatotopica: a gruppi di neuroni adiacenM corrispondono
aree del corpo conMgue. La quanMtà di neuroni assegnata a ciascuna area varia in funzione della finezza dei movimenM controllaM Input: corteccia somatosensoriale, APM, AMS Output: tronco dell’encefalo, midollo spinale
Homunculus motorio di W. Penfield Semiunculus di C. Woolsey

Controllo del Movimento
La mano ha una struLura anatomica molto complessa: • 39 muscoli (estrinseci ed
intrinseci) che agiscono su 18 giunM
• 23 gradi di libertà dal punto di vista cinemaMco
COME VIENE CONTROLLATO IL MOVIMENTO?

AREE MOTORIE Area 6
• Controllo del movimento volontario • Gioca un ruolo estremamente importante nella pianificazione delle
sequenze motorie complesse • Organizzazione somatotopica
• Input – corteccia parietale posteriore (aree 5-‐7): percezione e interpretazione delle relazioni
spaziali (localizzazione delle parM del corpo da muovere, e degli ogge] esterni con cui esse devono interagire)
– corteccia prefrontale (dorsolaterale): area associaMva polimodale, implicata con gli aspe] congniMvi del movimento (pianificazione ed esecuzione di movimenM complessi)
• Output – ProieLano su M1

AREE MOTORIE: LESIONI
Lesione M1 (oppure della via corMco-‐spinale): paresi (diminuzione parziale della moMlità) o paralisi (pardita completa non reversibile dell’a]vità motoria) Lesione APM / AMS: aprassia, incapacità di eseguire in modo correLo e coordinato il movimento, lentezza, goffaggine.

Area F5 PROPRIETA’ GENERALE DELL’AREA F5 (Rizzola] et al. 1988) Neuroni dell’area F5 hanno proprietà più complesse di quelle della corteccia motoria primaria. Codificano qualcosa di più astraLo del singolo movimento: scaricano in concomitanza di a] motori specificamente goal-‐related. “The funcMonal properMes of neurons located in the rostral part of inferior area 6 were studied in awake, parMally restrained macaque monkeys. The most interesMng property of these neurons was that their firing correlated with specific goal-‐related motor acts rather than with single movements made by the animal.” “All together these neurons form a vocabulary where proximal and distal movement necessary for reaching, grasping, holding and bringing the food to the mouth are represented.”

Una dimostrazione… Umiltà et al. 2008
“Primates are able to interact with objects not only by using their natural effectors, but also by using tools. Common tools, such as sMcks, stones, and rakes, act basically as funcMonal extensions of natural effectors. With pracMce, they become parts of the agent’s body schema.”
LO STUDIO: • Two adult macaque monkeys
(Macaca nemestrina) were used. • Single-‐unit acMvity was recorded
from areas F5 and F1 • Before recording, the monkeys
were trained to grasp food placed in front of them by using two types of tools.
• To grasp the object with normal pliers, the monkey has to close its hand (A), whereas with the reverse pliers, the monkey has to open its hand (B).
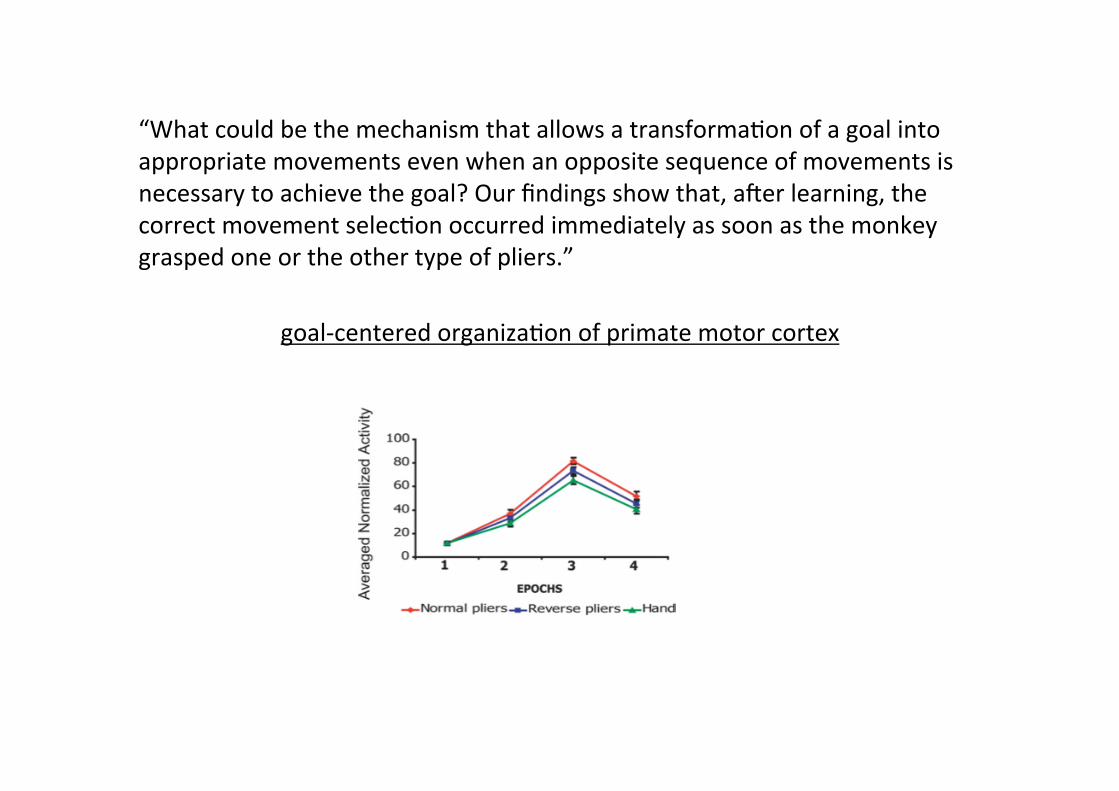
“What could be the mechanism that allows a transformaMon of a goal into appropriate movements even when an opposite sequence of movements is necessary to achieve the goal? Our findings show that, auer learning, the correct movement selecMon occurred immediately as soon as the monkey grasped one or the other type of pliers.”
goal-‐centered organizaMon of primate motor cortex

Neuroni canonici Murata et al. 1997 • Visual and motor properMes of single neurons of monkey ventral premotor cortex
(area F5) were studied • The employed objects were six different three-‐dimensional (3-‐D) geometric solids.
FINDINGS
• Two main types of neurons were disMnguished: motor neurons and visuomotor neurons. – Motor neurons discharged in associaMon with grasping movements. Most of them discharged
selecMvely during a parMcular type of grip. Different objects, if grasped in similar way, determined similar neuronal motor responses.
– Visuomotor neurons also discharged during acMve movements, but, in addiMon, they fired also in response to the presentaMon of 3-‐D objects.
• every Mme an object is presented, its visual features are automaMcally (regardless
of any intenMon to move) ‘‘translated’’ into a potenMal motor acMon. This potenMal acMon describes the pragmaMc physical properMes of the objects.

Neuroni canonici CaraLerisMche: • Neuroni visuomotori che scaricano sia durante gli a] motori, sia
alla presentazione di ogge] 3D.
• Ogni volta che un oggeLo viene percepito, le sue caraLerisMche fisiche vengono automaMcamente tradoLe in un potenziale motorio di azione.
• La visione di un oggeLo a]va immediatamente la selezione delle proprietà fisiche che permeLono di interagire con esso è AFFORDANCES

Affordance ConceLo introdoLo da Gibson “The affordances of the environment are what it offers the animal, what it provides or furnishes, either for good or ill. The verb to afford is found in the dicMonary, the noun affordance is not. I have made it up. I mean by it something that refers to both the environment and the animal in a way that no exisMng term does. It implies the complementarity of the animal and the environment.” (Gibson 1979, 127) DeFelice (2013) “L'autore definisce le affordances come “possibilità d'azione” offerte dagli ogge]: il conceLo non va ridoLo però alle semplici proprietà fisiche e visivamente percepibili degli ogge] in sé, ma è intrinsecamente relazionale, in quanto implica che sussista una complementarietà tra gli agenM e l'ambiente (e gli ogge] che ne fanno parte)” DeFelice (2014), Definizione di Affordance: “invariant set of objects’ physical properMes that offer possibiliMes for acMons”.

Affordances e neuroni canonici
• Where are Affordances grounded? CANONICAL NEURONS CIRCUITS
• La percezione di proprietà di ogge] manipolabili a]va una sorta di simulazione dell’azione nei circuiM cerebrali
• Stre]ssima relazione tra azione e percezione: l’abilità di agire dipende dalla abilità di percepire.

Neuroni Canonici nell’uomo Grezes et al. 2003a • S2muli: color video recordings (3.5 s each) of objects, grasping
pantomimes, and objects being grasped.
• The subjects viewed the videos and were required either to passively observe or to execute the appropriate grip on a manipulandum. – In the observaMon context, subjects observed an object (OO), observed a
grasp (OG), or observed an object being grasped (OGO). – In the execuMon condiMons, the subjects executed the grasp appropriate for
the object that they viewed (EO), imitated the pantomime they viewed (EG), or imitated the hand grasping an object (EGO).
• Two different objects were used; one had large opposiMon axes and would normally be grasped with a power grip. The other object was small and would normally be grasped by a precision grip.

Power grip (thumb + surface of the palm + all other fingers)
Precision grip (thumb + index finger)

Comparing those data with electrophysiological findings in monkeys, the concomitant acMvaMon of parietal and pre-‐motor areas during passive observaMon of an object and during execuMon of a grasp toward that object seems to correspond to the circuit shown in the macaque brain, that is, the parietal area AIP and the premotor area F5, where canonical neurons have been recorded.
• fMRI scanning
• The manipulandum employed for responses had two components. The first component consisted of a wooden cylinder […], that was used for the power grip response. ALached to the top part of the cylinder, the second component was made of a 2-‐cm-‐square and 1.5-‐cm-‐thick piece of wood and was used for the precision grip response. Both components contained a force transducer to record grip responses.

Buccino et al. 2009 • TMS: Transcranial MagneMc SMmulaMon1 • STIMULI: The sMmuli set was made of 6 × 2 × 2 sMmuli, each object being
represented in two horizontal orientaMons (one compaMble with a right-‐hand grasp, the other with a leu-‐hand grasp) and according to two different types of handle (one normal and compaMble with a hand grasp, the other broken).
1 Applicazione di un impulso magneMco alla corteccia. Se l’impulso è applicato alla corteccia motoria è possibile registrare i potenziali evocaM motori (MEP) nei muscoli controlaterali mediante eleLromiografia.
• Motor programs for grasping objects are specifically related to some specific pragmaMc features of objects like, for example, handle orientaMon (MEPs recorded from the right-‐hand were significantly larger when objects were presented with a normal handle on the right side versus when the handle was presented on the opposite side)
• The recruited motor programs are fine tuned to privileged components so that as soon as these are violated, the related motor programs are consequently interrupted, (This is tesMfied by MEPs recorded when objects were presented with the broken handle on the right side)

Neuroni Specchio ScoperM negli anni ‘90, a Parma [Rizzola], Gallese, Fadiga, Fogassi] Primi arMcoli: Rizzola] et al. 1996; Gallese et al. 1996 Dove? Area F5 della scimmia Cosa sono? sistema di neuroni parMcolari, a]vi sia quando il soggeLo compie l’azione, che quando l’azione è compiuta da un altro agente e semplicemente osservata. cfr. Gallese et al. 1996: Registrazione di 532 neuroni dell’area F5. 92 hanno proprietà “mirror”. Di quesM: • 51 si a]vano durante l’osservazione di una singola, specifica azione
compita con la mano. • 38 si a]vano per 2 o 3 azioni. • 3 si a]vano quando la scimmia osserva il ricercatore prendere cibo
con la bocca o con la mano 25: mirror-‐like, rispondono all’osservazione di azioni della mano ma non hanno proprietà motorie.

Neuroni specchio nella scimmia: funzione e caraLerisMche
HP: il sistema specchio gioca un ruolo nella comprensione delle azioni • Il traLo visivo che a]va i neuroni specchio è l’interazione tra agente e
oggeLo: la vista del solo agente o del solo oggeLo non produce scarica. • I neuroni specchio della scimmia non scaricano se l’azione viene mimata
dall’agente.
• I neuroni specchio si a]vano quando l’animale può prevedere la finalità dell’azione studi molto noM: – Umiltà et al. 2001 – Kohler et al. 2002; Keysers et al. 2003

Umiltà et al. 2001 • Two basic experimental
condiMons: – “full vision” condiMon [the monkey was shown a fully visible acMon directed toward an object] – “hidden” condiMon [the same acMon was presented but with its final criMcal part (hand-‐ object interacMon) hidden].
• The results showed that the majority of mirror neurons responded also in the hidden condiMon.

• “[…] a subset of mirror neurons becomes acMve during acMon presentaMon and also when the final part of the acMon, crucial in triggering the response in full vision, is hidden and can therefore only be inferred.”
• “This implies that the motor representaMon of an acMon
performed by others can be internally generated in the observer’s premotor cortex, even when a visual descripMon of the acMon is lacking. The present findings support the hypothesis that mirror neuron acMvaMon could be at the basis of acMon recogniMon.”
• “Full visual informaMon about an acMon is not necessary to recognize its goal. AcMon understanding could be based on a mechanism that can trigger the internal motor representaMon of the acMon.”

“Audiovisual” mirror neurons Kohler et al. 2002/Keysers et al. 2003 • STIMULI= “noisy acMons”
– Peanut breaking – Paper ripping – PlasMc crumpling – Metal hi]ng metal – Paper shaking – SMck dropping
• three ‘sensory’ condiMons: – vision-‐and-‐sound (‘V+S’) – vision-‐only (‘V’) – sound-‐only (‘S’)

• “[…] some neurons in the ventral premotor cortex (area F5) of the monkey responding during the execuMon of acMons also respond to the vision and/or the sound of these acMons.”
• “[…] for half of the tested audiovisual mirror neurons, the amplitude of the response does not differ significantly whether the preferred acMon is heard, seen or both heard and seen.”
• “This finding is important, as it suggests that the neurons code the acMon in an abstract way, which does not depend on the source of informaMon (auditory or visual) from which the evidence about the presence of the acMon is taken.”
• cornerstone in the evoluMon of language?

Non solo F5: Mirror-‐neuron circuit (Rizzola] e Sinigaglia, 2006)
Il sistema dei neuroni specchio è bilaterale e include ampie porzioni della corteccia parietale e premotoria.
• F5
• Area PF of Von Economo (rostral part of the inferior parietal lobule, IPL) “It receives input from STS and sends an important output to the ventral premotor cortex including area F5. PF neurons are funcMonally heterogeneous. Most of them (about 90%) respond to sensory sMmuli, but about 50% of them also have motor properMes discharging when the monkey performs specific movements or acMons”).
• STS (superior temporal sulcus) STS neurons respond to the observaMon of acMons done by but do not appear to be endowed with motor properMes.)

“The corMcal mirror neuron circuit is formed by two main regions: the rostral part of the inferior parietal lobule and the ventral premotor cortex. STS is strictly related to it but, lacking motor properMes, cannot be considered part of it.” (Rizzola] & Craighero, 2004)

Neuroni specchio: funzione
Generazione di una rappresentazione interna del movimento (“simulazione”), che può avere varie funzioni: • comprensione del significato dell’azione
osservata • leLura delle intenzioni dell’individuo che
agisce (Fogassi et al. 2005)
“The mirror system provides a way to match observa2on and execu2on od events.” [Gallese et al. 1996]

Fogassi et al. 2005 • IPL (inferior parietal lobe) • neurons were studied when monkeys performed motor acts embedded in different
acMons and when they observed similar acts done by an experimenter. • Most motor IPL neurons coding a specific act (e.g., grasping) showed markedly different
acMvaMons when this act was part of different acMons (e.g., for eaMng or for placing). • Many motor IPL neurons also discharged during the observaMon of acts done by others.
Most responded differenMally when the same observed act was embedded in a specific acMon.
e.g.: -‐ intense firing when the monkey grasped a fruit to bring it to
its mouth. -‐ weaker response when the monkey grasped the food to place
it in a container. -‐ intense response when the monkey watched an
experimenter perform the grasp-‐to-‐eat gesture. -‐ weaker response to the grasp-‐to-‐place acMon.
These neurons not only code the observed motor act but also allow the observer to understand the agent’s intenMons.

Neuroni specchio nell’uomo COME? Evidenze indireLe • EsperimenM di Mpo neurofisiologico
– EEG -‐ eleLroencefalografia: Registrazione del mu-‐rhythm (cfr. studi di Gastaut) – TMS -‐ Transcranial magneMc sMmulaMon
• brain-‐imaging – PET -‐ Tomografia a emissione di positroni – fMRI-‐ Risonanza magneMca funzionale
DOVE? Osservazione di azioni compiute da altri comporta anche nell’uomo l’a]vazione di un network complesso di aree: • Aree visive (occipitali, temporali e parietali) • regioni corMcali eminentemente motorie
– parte rostrale del lobulo parietale inferiore IPL – parte inferiore del giro precentrale – parte posteriore del giro frontale inferiore >> AREA DI BROCA

Area di Broca • Non è un’area esclusivamente linguisMca!
• Corrisponde alle aree di Brodmann 44 e 45. -‐ Area 44, Pars opercularis del giro frontale inferiore: in aggiunta alla rappresentazione del
linguaggio conMene, così come F5 nella scimmia, le rappresentazioni motorie dei movimenM della mano.
-‐ Area 45, pars triangularis del giro frontale inferiore: a]vata da elemenM linguisMci, sia verbali che segnaM.
• Connessa all’area di Wernicke aLraverso il fascicolo arcuato

scimmia
uomo

Buccino et al. 2001 • fRMI • AIM:
– to localize brain areas that were acMve during the observaMon of acMons made by another individual
– to assess whether the observaMon of acMons made with different effectors would acMvate specific parts of the premotor cortex in accord with the somatotopic motor organizaMon of the region
• STIMULI: Object-‐ and non-‐object-‐related acMons made with different effectors (mouth, hand and foot) were presented. – mouth acMons:
• biMng an apple • chewing
– hand acMons: • reaching and grasping a ball or a liLle cup with the hand • mimicking these acMons without the object
– foot acMons • kicking a ball • pushing a brake • mimicking these acMons without the object

FINDINGS: • “ObservaMon of both object-‐ and non-‐object-‐related acMons determined a
somatotopically organized acMvaMon of premotor cortex. The somatotopic paLern was similar to that of the classical motor cortex homunculus.”
• “During the observaMon of object-‐related acMons, an acMvaMon, also somatotopically organized, was addiMonally found in the posterior parietal lobe.”
• The mirror System “is not restricted to the ventral premotor cortex, but involves several somatotopically organized motor circuits.”
L’ESPERIMENTO DIMOSTRA INOLTRE CHE: • Il sistema specchio nell’uomo non è circoscriLo alla mano.
• Nell’uomo anche l’osservazione di pantomine determina l’a]vazione nei neuroni specchio. (cfr. Umiltà et al. 2001)

Buccino et al. 2004 • fRMI
• AIM: “to assess the corMcal areas acMve during the observaMon of mouth acMons performed by humans and by individuals belonging to other species (monkey and dog).”
• STIMULI: Two types of acMons were presented: biMng and oral communicaMve acMons (speech reading, lip-‐smacking, barking).

FINDINGS: • “The results showed that when the observed acMon is common to animals
and humans, there is a clear overlap between the acMvated areas, in spite of the enormous differences in the visual aspects of the observed sMmuli. In contrast, during the observaMon of acMons that, like oral communicaMve acMons, have a common goal, but are expressed differently in the three species, there is a clear difference in the distribuMon and extent of acMvaMons.”
• “Taken together, the results of the present experiment suggest that acMons made by other individuals may be recognized in different ways. AcMons belonging to the motor repertoire of the observer are mapped on his/her motor system. AcMons that do not belong to this repertoire appear to be recognized essenMally based on their visual properMes.”
• “AcMons that are not part of the motor repertoire of the observer and that therefore cannot be reproduced appear to be recognized in nonmotor terms. They are most likely understood based on visual descripMon of the observed events and inferences of their consequences and/or goals.”


Neuroni specchio ed empaMa Simulazione incarnata e rispecchiamento di emozioni e sensazioni. Comprensione esperienziale delle emozioni di base, ad es. Disgusto è BASE BIOLOGICA DELL’EMPATIA Wicker et al. 2003 • fRMI • STIMULI:
– parMcipants inhaled odorants producing a strong feeling of disgust.
– the same parMcipants observed video clips showing the emoMonal facial expression of disgust.
• Observing such faces and feeling disgust acMvated the same sites in the anterior insula and to a lesser extent in the anterior cingulate cortex.

“Simulazione Incarnata” • Il meccanismo funzionale che sta alla base del doppio paLern di a]vazione dei
neuroni specchio è stato definito “simulazione incarnata” (“embodied simula2on”)
• Nuova concezione delle a]vità mentali e della cognizione: embodied (V. Gallese) / grounded (Barsalou)
da meccanismo di manipolazione di simboli astra] e rappresentazioni, basato su regole formali e processi è a ciò che emerge dall’azione del corpo nel suo contesto reale
• Capacità perce]ve e motorie non sono separabili. Da esse si sono sviluppate
probabilmente anche le facoltà “superiori”. • Il sistema sensomotorio di cui fano parte i neuroni specchio ha caraLere
mulMmodale, e non modulare!

Riassumendo • Peculiarità dell’area F5: neuroni a]vi somatotopicamente in rapporto alla
funzione, ovvero “sparano” in concomitanza di a] motori, non di singoli movimenM.
• 2 popolazioni di neuroni nella scimmia nell’uomo, con distribuzione corMcale omologa: Neuroni Canonici (Neuroni visuomotori che scaricano sia durante gli a] motori, sia alla presentazione di ogge] 3D)
Neuroni Specchio (Neuroni visuomotori che scaricano sia durante gli a] motori, sia durante l’osservazione dello stesso aLo motorio compiuto da un altro agente)
• SoLoclasse dei neuroni specchio audiovisivi • Funzione dei Neuroni specchio : comprensione dell’azione + leLura dell’intenzione • Peculiarità del sistema specchio nell’uomo:
– Non è circoscriLo alla mano – Si a]va anche in caso di pantomina – Simulazione incarnata: le azioni che sono fuori dal repertorio della specie non hanno risonanza
motoria nell’osservatore. • Neuroni specchio ed empaMa • Simulazione incarnata: quali implicazioni per la teoria della mente?

Perché la scoperta dei neuroni specchio dovrebbe interessare al linguista?
• Filogenesi del linguaggio
• Ontogenesi del linguaggio
• Neuroni specchio, percezione e comprensione linguisMca
– Liv. Fonologico – Liv. Lessicale-‐semanMco

MA PRIMA… Neuroni specchio nella scimmia: altri effeLori Cfr. Buccino et al. 2001 il sistema specchio nell’uomo non è circoscriLo alla mano Ferrari et al. 2003: “NEURONI SPECCHIO DELLA BOCCA” Purpose of the experiments: “to study the responses of F5 neurons motorically coding mouth acMons to the observaMon of mouth acMons made by another individual.” Results: • “35% of mouth motor neurons discharged when the monkey observed
mouth acMons (mouth mirror neurons).” • “The majority of mouth mirror neurons (85%) discharged during the
observaMon of ingesMve mouth acMons, others (15%) during the observaMon of communicaMve acMons.”

INGESTIVE MOUTH-‐MIRROR NEURONS • “The funcMonal properMes of ingesMve mouth-‐mirror neurons are very
similar to those of hand mirror neurons.” – “As hand mirror neurons, mouth mirror neurons require an interacMon
between an effector and an object in order to be triggered. The presentaMon of an object alone or of a not goal-‐directed movement is insufficient to acMvate the neurons.”
– “They show congruence between the effecMve observed and the effecMve executed acMon.”
• “these findings extend the previous concept of the mirror system as a system matching acMon observaMon and acMon execuMon from hand to mouth acMons.”
COMMUNICATIVE MIRROR NEURONS • “communicaMve mirror neurons, unlike other mirror neurons, discharge in
response to acMons such as lip-‐smacking or tongue protrusion, acMons that are not object-‐directed.”
• “the effecMve motor acMon is not idenMcal to the effecMve observed one. All studied communicaMve mirror neurons discharged during acMve ingesMve acMons made by the monkey.”
Why should ingesMon and communicaMon share a common neural substrate?

Filogenesi del linguaggio Come è possibile che da tale sistema, così streLamente legato all’oggeLo, si sia sviluppato un sistema aperto capace di descrivere azioni e ogge] senza necessariamente riferirvisi? Rizzola] & Arbib 1998 The “observaMon/execuMon matching system provides a necessary bridge from doing to communica2ng” “The link between actor and observer becomes a link between the sender and the receiver of each message.”

Filogenesi del linguaggio • “The development of the human lateral speech circuit is a consequence of
the fact that the precursor of Broca’s area was endowed, before speech appearance, with a mechanism for recognizing acMons made by others. This mechanism was the neural prerequisite for the development of interindividual communicaMon and finally of speech.”
• HP: “the mimeMc capacity inherent to F5 and Broca’s area had the
potenMal to produce various types of closed systems related to the different types of motor fields present in that area (hand, mouth and larynx)”
• “the first open system to evolve en route to human speech was a manual
gestural system that exploited the observaMon and execuMon matching system described earlier”
• gestural communicaMon è vocalizaMons è speech

Filogenesi del linguaggio Neuroni specchio della bocca [Ferrari et al. 2003]
HP: il sistema ingesMvo, controllato dalle aree motorie ventrali, si è evoluto nel sistema comunicaMvo • “IngesMon and communicaMon share a common neural substrate” • “ethological and evoluMonary data that strongly suggest that these two
funcMons are strictly linked.” • “monkey communicaMve gestures (e.g. lip-‐smacking, lips protruded face)
are ritualizaMons of ingesMve acMons that are used for affiliaMve purposes. […] In other words, the knowledge common to the communicator and the recipient of communicaMon about food and ingesMve acMon became the common ground for social communicaMon. IngesMve acMons are the basis on which communicaMon is built.”
cfr. MacNeilage 1998: “human vocal communicaMon derived from the cyclic open-‐close mandibular alternaMon originally evolved for food ingesMon.”

Ontogenesi del linguaggio (Capirci et al. 2005; Stefanini et al. 2009; Congestrì et al. 2010; PeLenaM et al. 2010)
Ruolo centrale del gesto nell’ontogenesi del linguaggio, sia nelle primissime fasi dello sviluppo comunicaMvo, sia nelle fasi successive. ConMnuità tra le forme linguisMche e prelinguisMche (ovvero tra azione-‐gesto-‐parola) “Progression from acMon to word through gesture” “There is a conMnuity between the producMon of the first acMon schemes, the first gestures and the first words produced by children. The relaMonship between gesture and words changes over Mme.”

1 anno: comunicazione intenzionale inizia mediante vocalizzazione e gesM dei]ci (“poinMng”) • Gesto:
– ProdoLo da solo – ProdoLo in combinazione con vocalizzazioni – ProdoLo in combinazione con 1word-‐uLerance
• Cross modal combinaMon gesture/word combinaMon
• Two-‐word stage

Gestualità negli adulM Contributo delle neuroscienze: approccio globale, integrato e mulMmodale all’analisi dei processi comunicaMvi.
• I gesM sono parte integrante del processo comunicaMvo • Il linguaggio è un sistema integrato gesto-‐parlato
(cfr. Kendon 2004; McNeill 2000, 2005). “As a close examinaMon of the coordinaMon of gesture with speech suggests, these two forms of expression are integrated, produced together under the guidance of a single aim.” (Kendon, 2004: 2-‐3)
• Nel linguaggio adulto i gesM sono coespressivi e sincronici rispeLo al parlato: gesM e parole, pur usando modalità differenM, sono inseparabili (“Unbreakable bond”, McNeill 2005) ed esprimono un’unica idea soLostante, ovvero sono espressioni diverse dello stesso sistema.
• Ruolo del gesto nell’acquisizione di L1

Gestualità negli adulM: case study (Gagliardi, 2014)
• Analisi della gestualità di sogge] adulM ed anziani in un test di visual confronta2on naming di azioni (daM della taratura della baLeria SMAAV, Seman2c Memory Assessment on Ac2on verb)
• Analisi linguisMca + analisi delle produzioni gestuali degli informanM durante la somministrazione della baLeria.
• Focus sui “GesM rappresentaMvi” (“representa2onal”) ovvero gesM che forniscono una rappresentazione piLografica del significato associato ad un oggeLo o ad un evento. Denotano un preciso referente. Il contenuto semanMco di base rimane relaMvamente stabile nelle diverse situazioni.
– “Ac2on gesture” (o “enactment”): una parte del corpo compie un paLern di azioni che hanno caraLerisMche in comune con il paLern di azioni che funge da referente ;
– “size-‐shape gesture” (o “modeling / depic2on”): il gesto richiama dimensione, forma o caraLerisMche perce]ve dell’evento o dell’oggeLo. (Ad esempio, le mani assumono una configurazione che riproduce la forma dell’oggeLo a cui il gesto si riferisce o eseguono un movimento per creare l’oggeLo nell’aria.)

FINDINGS: • In accordo con quanto è riportato in leLeratura, i significaM convogliaM da
gesto e risposta vocale sono simili (match semanMco)
• Lo stroke del gesto (ovvero la parte centrale del gesto, obbligatoria, dotata di significato, in cui è concentrato lo sforzo energeMco) è prodoLo sincronicamente al parlato, con cui partecipa alla costruzione del senso degli enunciaM.
• La coespressivita di gesto e risposta vocale puo declinarsi in varie maniere: talvolta il gesto sembra configurarsi come espressione parallela, ridondante rispeLo al significato espresso verbalmente, mentre in altre occasioni precisa e restringe l’applicazione dell’eMcheLa lessicale.
• Sebbene in alcuni casi il gesto sembri subentrare nel momento in cui vi siano difficoltà nel recupero lessicale il rapporto tra esso e il parlato non è mai di pura compensazione.
• Probabile a]vazione somatotopica.

Acquisizione AMpica: Sindrome dello SpeLro AuMsMco
CARATTERISTICHE del DISTURBO: – incapacità di relazione interpersonale – mancanza di aLenzione – tendenza all’isolamento – evitamento dello sguardo – considerazione dell’altro come un oggeLo – …
Gallese 2006 • “embodied mechanisms involving the acMvaMon of the sensory-‐motor system
seem to play a major role in social cogniMon.” • “all the deficits can be explained as instanMaMons of intenMonal aLunement
deficits produced by defecMve embodied simulaMon, likely underpinned by a malfuncMoning of the mirror neuron systems.”
Evidenze che vi sia un malfunzionamento nel sistema specchio:
– EleLroencefalografia: gli individui auMsMci non mostrano soppressione del ritmo mu durante l’osservazione delle azioni

DapreLo et al. 2006 • fRMI
• 10 High-‐funcMoning children with auMsm and 10 typically developing children • AIM: “to invesMgate neural acMvity during the imitaMon and observaMon of facial
emoMonal expressions” • STIMULI: 80 faces expressing five different emoMons:
– anger – fear – happiness – neutrality – sadness
• FINDINGS: “The evidence […] suggests that although both groups performed the imitaMon task as requested, the neural strategies adopted by typically developing children and those with ASD are quite different.”

Reliable acMvity during imitaMon of emoMonal expressions. (a,b) AcMvity in bilateral pars opercularis of the inferior frontal gyrus is seen in the typically developing group (a) but not in the ASD group (b). A between-‐group comparison (c)
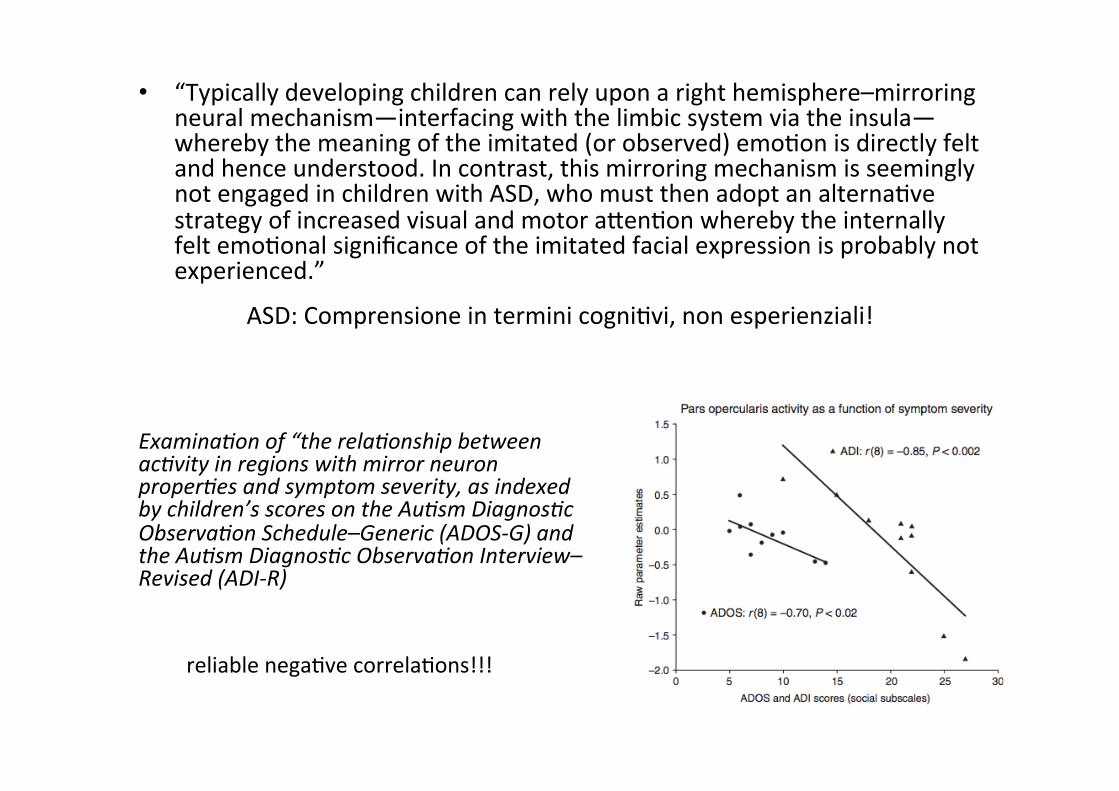
• “Typically developing children can rely upon a right hemisphere–mirroring neural mechanism—interfacing with the limbic system via the insula— whereby the meaning of the imitated (or observed) emoMon is directly felt and hence understood. In contrast, this mirroring mechanism is seemingly not engaged in children with ASD, who must then adopt an alternaMve strategy of increased visual and motor aLenMon whereby the internally felt emoMonal significance of the imitated facial expression is probably not experienced.”
ASD: Comprensione in termini cogniMvi, non esperienziali!
Examina2on of “the rela2onship between ac2vity in regions with mirror neuron proper2es and symptom severity, as indexed by children’s scores on the Au2sm Diagnos2c Observa2on Schedule–Generic (ADOS-‐G) and the Au2sm Diagnos2c Observa2on Interview–Revised (ADI-‐R)
reliable negaMve correlaMons!!!

Linguaggio nel bambino auMsMco Brandi & Bigagli 2004 “le caraLerisMche del disturbo portano a sospeLare la crucialità, anche per lo sviluppo del linguaggio, di ciò che cogniMvamente manca al bambino auMsMco: la capacità di relazione intersogge]va, la capacità di rappresentare lo stato mentale altrui, la capacità di imitare.” uno degli indicatori fondamentali della presenza del disturbo auMsMco è l’incapacità nell’esecuzione di giochi simbolici (tra 3 e 5 anni) LINGUAGGIO (Brandi 2005) “È ormai noto che l’auMsmo, oltre ad essere propriamente un deficit di Mpo socio-‐comportamentale, porM con sé anche disturbi di caraLere streLamente linguisMco: a seconda della gravità della sindrome è possibile assistere ad un vero e proprio muMsmo, oppure ad un notevole ritardo nello sviluppo del linguaggio (talvolta le prime sillabe appaiono intorno agli oLo anni), o ancora, nel caso di uno sviluppo linguisMco tardivo, esso può manifestarsi aLraverso la produzione di stereoMpie”

Disturbo complesso e di conseguenza eterogeneo per quanto riguarda la manifestazione dei sintomi. Varie tipologie di comportamento linguisMco. (vedi Brandi 2005) STEREOTIPIE: dissociazione tra forma e funzione nell’acquisizione del linguaggio è dissociazione tra abilità sinta]co-‐semanMca e abilità pragmaMca marcata asincronia tra lo sviluppo della funzione comunicaMva e l’acquisizione della grammaMca Sono gli aspe] pragmaMci del linguaggio a presentarsi sistemaMcamente disturbaM nei sogge] auMsMci

Neuroni specchio e linguaggio: livello fonologico
Fadiga et al. 2002 • TMS
• Subjects were instructed to listen carefully to a sequence of acousMcally presented verbal and nonverbal sMmuli. e.g. WORDS: birra, carro, ferro, porro, terra PSEUDO-‐WORDS: berro, furra, parro, vurro
• Motor-‐ evoked potenMals (MEPs) were recorded from opponens pollicis and
tongue muscles FINDINGS: • during speech listening, there is an increase of motor-‐evoked potenMals recorded
from the listeners' tongue muscles when the presented words strongly involve, when pronounced, tongue movements.
• when an individual listens to verbal sMmuli there is an acMvaMon of the speech related motor centres. Most interesMngly, they show also that this acMvaMon is highly specific.

Watkins et al. 2003 • TMS (Transcranial magneMc sMmulaMon) was applied to the face area of
primary motor cortex to elicit motor-‐evoked potenMals in the lip muscles.
• AIM: to examine whether auditory percepMon of speech modulated the excitability of the motor system underlying speech producMon.
• EXPERIMENTAL CONDITIONS – (1) Speech condiMon: listening to speech (conMnuous prose) while viewing
visual noise. – (2) Non-‐verbal condiMon: listening to non-‐verbal sounds (e.g. glass breaking,
bells ringing, guns firing) while viewing visual noise. – (3) Lips condiMon: viewing speech-‐related lip movements while listening to
white noise. – (4) Eyes condiMon: viewing eye and brow movements while listening to white
noise.
• FINDINGS: “The results demonstrate that speech percepMon, either by listening to speech or by visual observaMon of speech-‐related lip movements, enhanced excitability of the motor units underlying speech producMon.”

SinteMzzando: • Il sistema motorio si a]va durante percezione del linguaggio (effeLori: lingua, labbra).
• HP: Coinvolgimento del circuito dei neuroni specchio nel processing linguisMco: l’ascoltatore percepisce e decodifica i suoni linguisMci mappando l’input sul repertorio di comandi motori necessari per produrre qui suoni, che anche lui possiede.
• Simulazione incarnata della fonazione

CoarMcolazione e il “problema dell’invarianza”
COARTICOLAZIONE “A fundamental problem in speech percepMon is that the realizaMon of speech sounds is highly context dependent. Successive speech sounds are produced by vocal tract gestures that overlap temporally. The consequence of coarMculaMon is that there is no one-‐to-‐one correspondence between an acousMc event and the repertoire of phonemes or phoneMc features in the language.” “PROBLEMA DELL’INVARIANZA”: Invarianza del sistema che soggiace alla competenza linguisMca vs. variabilità del parlato. E’ comunque necessario un livello di rappresentazione fonologica? Soluzione classica: “A view that is shared by many in the field of speech percepMon is that speech is perceived by exploiMng the same auditory mechanisms that analyse other classes of environmental sounds. On the way to acMvaMng word forms in the mental lexicon (lexical access), the acousMc informaMon gives way to an intermediate input representaMon.” • Phonemic level of representaMon • AlternaMve “unit of percepMon” (e.g. syllabe, stress unit…)

“Motor theory of speech percepMon” ALTERNATIVA RADICALE! Haskins Laboratories Liberman et al. 1967; Liberman e Ma]ngly 1985, Galantucci et al. 2006 • PrimiMvi linguisMci: i gesM arMcolatori.
• Il linguaggio parlato viene percepito confrontando tali gesM arMcolatori con il repertorio motorio dell’ascoltatore.
• Percezione e produzione del linguaggio uMlizzano il medesimo repertorio di primiMvi motori.
• Carenza di evidenze sperimentali al momento della presentazione della teoria. La scoperta dei neuroni specchio ha portato nuovi argomenM.
• HP: A]vazione delle rappresentazioni motorie rilevanM fonologicamente. Risonanza motoria durante la percezione.

Toni et al. 2008 • “the listener does not solve the invariance problem in the auditory
domain, but instead in the motor domain. The acousMc paLerns might be different, but the arMculatory gestures that are needed to produce them are the same.”
• “A listener will understand speech by virtue of being a speaker.”
• “The neuromotor commands provide the required invariance” Fadiga et al. 2002 • “A cogniMve translaMon into phonology is not necessary because the
arMculatory gestures are phonologic in nature. Furthermore, speech percepMon and speech producMon processes use a common repertoire of motor primiMves that, during speech producMon, are at the basis of arMculatory gesture generaMon, while during speech percepMon, are acMvated in the listener as the result of an acousMcally evoked motor `resonance'.”
SI PUO’ FARE A MENO DI UN LIVELLO DI RAPPRESENTAZIONE FONOLOGICO?

Neuroni specchio e linguaggio: livello semanMco
Buccino et al. 2005 • TMS
• AIM: “to assess whether listening to acMon-‐related sentences modulates the acMvity of the motor system.”
• “By means of single-‐pulse TMS, either the hand or the foot/leg motor area in the leu hemisphere was sMmulated in disMnct experimental sessions, while parMcipants were listening to sentences expressing hand and foot acMons. Listening to abstract content sentences served as a control. Motor evoked potenMals (MEPs) were recorded from hand and foot muscles.”
• STIMULI: Fiueen hand-‐acMon-‐related and 15 foot-‐acMon-‐related sentences were presented. As control sMmuli, 15 sentences with verbs expressing an abstract content were delivered. All sentences were in Italian. All acMon-‐related sentences expressed a concrete acMon on an appropriate object (e.g., cuciva la gonna […]). All abstract content sentences expressed an abstract acMon on an appropriate object (e.g., amava la patria […]). All verbs were formed by three syllables and were conjugated in the third person of the past tense.

Hand-‐acMon-‐related sentences: Girava la chiave, Prendeva la tazza, scriveva il tema… Foot-‐acMon-‐related sentences: Marciava sul posto, saltava il muro, pestava le foglie… Abstract content sentences: Soffriva il freddo, scordava la data, temeva la pena…
• Results showed that MEPs recorded from hand muscles were specifically
modulated by listening to hand-‐acMon-‐related sentences, as were MEPs recorded from foot muscles by listening to foot-‐acMon-‐related sentences.

Hauk et al. 2004 • fMRI
• AIM: “The predicMon under invesMgaMon in the present study concerns possible differences between the corMcal acMvaMon paLerns elicited by acMon words of different semanMc subcategories and, more specifically, their relaMon to motor areas”
• STIMULI: “50 words from each of the three semanMc subcategories (i.e. acMon words referring to face, arm, leg acMons) were selected and presented in a passive reading task to 14 right-‐handed volunteers, while hemodynamic acMvity was monitored using event-‐related fMRI.”
• “To idenMfy the motor cortex in each volunteer individually, localizer scans were also performed, during which subjects had to move their leu or right foot, leu or right index finger, or tongue.”

FINDINGS: • “Our present results indicate that such acMon-‐related acMvaMon can
involve primary motor cortex and does not require a linguisMc task (e.g., naming) but is elicited by sMmulus words per se, even in a passive reading task.”
• “[…] the paLern of corMcal acMvaMon elicited by an acMon word reflects the corMcal representaMon of the acMon to which the word refers.”
Esperimento replicato molteplici volte! Ad esempio Te4aman5 et al. 2005

In sintesi… Pulvermuller 2005 “Hearing a word seems to be associated with acMvaMon of its arMculatory motor program, and understanding an acMon word seems to lead to the immediate and automaMc thought of the acMon to which it refers.” “If acMon words are semanMcally related to the movements of the face or arMculators, arm or hand, or leg or foot, the distributed neuronal ensembles would include semanMc neurons in perisylvian (face-‐related words), lateral (arm-‐related words) or dorsal (leg-‐related words) motor and premotor cortex. Therefore, this semanMc somatotopy model of acMon words implies that there are differently distributed networks for the English words ‘lick’, ‘pick’ and ‘kick’”

Memoria SemanMca: definizione • La memoria semanMca, o conoscenza conceLuale, è l’aspeLo della
memoria umana che corrisponde alla conoscenza generale del significato delle parole e di ogge], fa] e persone, senza connessione ad un parMcolare tempo o luogo. “Seman2c memory is the memory necessary for the use of language. It is a mental thesaurus, organized knowledge a person possesses about words and other verbal symbols, their meanings and referents, about rela2ons among them, and about rules, formulas, and algorithms for the manipula2on of these symbols, concepts and rela2ons.” (Tulving 1972)
• Vs. Memoria Episodica
• Substrato neurale?

Memoria SemanMca: correlaM neurali
• Le posizioni teoriche correnM sulla memoria semanMca condividono l’idea che gran parte del contenuto della conoscenza conceLuale sia collegata alla percezione ed all’azione, e sia dunque un network neurale distribuito, rappresentato somatotopicamente in regioni cerebrali sovrapponibili o coincidenM con le aree senso-‐motorie
• Due visioni contrapposte della rappresentazione neuroanatomica
della memoria semanMca (PaLerson et al. 2007): – distributed-‐only view: queste regioni cerebrali distribuite, e le
connessioni neurali che le collegano, esauriscono per intero la base neurale della memoria semanMca
– distributed-‐plus-‐hub view: le rappresentazioni provenienM dalle aree senso-‐motorie e linguisMche sarebbero connesse e coordinate da un “hub” amodale (o eteromodale) situato nel lobo temporale.

Distribuzione neuroanatomica dei network semanMci (PaLerson et al. 2007)

Classi di parole nel cervello Gli aLribuM semanMci connessi all’azione e al movimento, generalmente lessicalizzaM nei verbi, risiedono in struLure corMcali frontali, ovvero nelle aree corMcali primariamente deputate alla programmazione ed esecuzione motoria. Il processing dei nomi avviene nell’area temporale anteriore e mesiale. Però:
– La generalizzazione non sembra estendersi al lessico nominale e verbale astraLo (es. libertà, e2ca).
– La generalizzazione non sembra estendersi al linguaggio metaforico (cfr. ad esempio la review di Aziz-‐Zadeh & Damasio 2008)
– conce] logici (es. IF, NOT)? • Lesioni focali e “doppie dissociazioni” La scoperta di una doppia dissociazione tra nomi e verbi come forme lessicali è stata considerata a parMre dagli anni novanta un’evidenza del faLo che la classe grammaMcale sia un principio organizzaMvo della conoscenza lessicale nel cervello (Hillis & Caramazza, 1995), rappresentata in network neurali disMnM e separaM (Damasio & Tranel, 1993).

• Ipotesi alternaMve: – la differenza di classe lessicale emerge a livello morfologico: non sono nomi e
verbi in sé ad essere elaboraM in network separaM, ma i processi morfo-‐sinta]ci che si applicano al lessico nominale e verbale sarebbero computaM in reM neurali disMnte (Shapiro & Caramazza, 2003; Shapiro et al., 2006)
– La dissociazione non è tanto tra nomi e verbi come classi lessicali, quanto tra parole riferite ad ogge] e parole riferite ad azioni (Vigliocco et al. 2006, 2011)

Fino a che punto è possibile generalizzare?
Toni et al. 2008 • “The discovery of mirror neurons in macaques and of a similar
system in humans has provided a new and ferMle neurobiological ground for rooMng a variety of cogniMve faculMes.”
• Three claims that deal with the relaMonship between language and the motor system: – Does language comprehension require the motor system? – Was there an evoluMonary switch from manual gestures to speech as
the primary mode of language? – Is human communicaMon explained by automaMc sensorimotor
resonances?
CRITICAL REVIEW

• “We are not arguing against important links and interacMons between language and motor systems in the brain. For the semanMcs of certain word classes the acMon system might be invoked. Clearly at the level of phoneMc realizaMons the motor system plays a crucial role. This could even be in language comprehension when under condiMon of high predictability the predicted word forms might actually be produced in the form of internal speech. However, all this does not imply that the highly complicated communicaMon system of natural language can be fully reduced to sensorimotor properMes and the contribuMon of sensorimotor areas.”
• “Language goes beyond acMon. Without denying the enormous importance of the discovery of mirror neurons, their explanatory power for understanding human communicaMon is limited.”







![Cognizione - Enciclopedia Einaudi [1982]](https://static.fdocumenti.com/doc/165x107/55c3ecd6bb61ebf4088b4674/cognizione-enciclopedia-einaudi-1982.jpg)










