
PROVE DI PERMEABILITÀ BOUTWELL Dott. Geol. Raffaele Isolani.

Modulo 9: Membrane

Membrane formano barriere fisiche. Separano il citosol dall’ambiente esterno preservano
l’individualità della cellula mantenendola separata dall’ambiente circostante.
Le funzioni delle membrane biologiche
Permettono la
comunicazione tra ambiente
esterno ed interno attraverso
recettori proteici.
Attuano la conversione dell’
energia chimica attraverso la
formazione di gradienti
elettrochimici
Delimitano i compartimenti intracellulari (nucleo, mitocondri, ER, lisosomi ecc.)
Hanno funzioni di trasporto di metaboliti, di ioni.

Le membrane biologiche: proprietà
Le membrane biologiche sono strutture laminari simili a fogli con
spessori compresi tra 5 e 8 nm.
Costituite principalmente da glicerofosfolipidi, sfingolipidi, colesterolo.
Strutture senza legami covalenti tra le unità di
base, le unità che le compongono sono
tenute unite da interazioni non covalenti.
Le membrane sono asimmetriche e di norma
elettricamente polarizzate
Permeabilità molto bassa per la maggior parte delle
molecole. Contengono proteine specifiche e
particolari responsabili di funzioni specializzate delle
membrane.
Distribuzione dei lipidi negli strati interno ed esterno
della membrana plasmatica (eritrociti)

I principali componenti lipidici delle
membrane biologiche
Ogni membrana (plamatica o
interna) ha una sua
composizione lipidica ben
precisa.

Il modello a mosaico fluido
Le membrane sono strutture fluide e dinamiche. Fluidità di membrana: data dalle
interazioni non covalenti, lasciano libere le molecole di spostarsi lateralmente nel piano
della membrana.
I lipidi inseriti nelle membrane sono soggetti ad una
rapida diffusione laterale: 1m2 S-1 Mentre la diffusione
trasversale è molto limitata.
“Le membrane biologiche sono soluzioni
bidimensionali di lipidi orientati e proteine
globulari” (Singer & Nicholson1972)
Membrane paragonate a un
liquido bidimensionale. Le
proteine “galleggiano” nel
liquido bidimensionale.

La fluidità di membrana è controllata dalla
composizione lipidica
Le catene aciliche
sono in uno stato
ordinato
Le catene aciliche sono
in stato disordinato
La temperatura (Tm) di transizione fluido (liquido cristallino)-solido (gel) dipende
dalla lunghezza delle catene degli acidi grassi, dal loro grado di insaturazione e
dalla presenza di colesterolo.
La presenza di doppi legami rende le catene più irregolari diminuendo il numero
di legami possibili tra le catene, rendendo le catene più fluide
Gli animali a sangue freddo modificano la composizione in acidi grassi dei lipidi di
membrana a seconda della T.

Il colesterolo è il principale modulatore della
fluidità delle membrane degli animali
Il colesterolo si inserisce
nel doppio strato lipidico
e si impacca tra i
fosfolipidi/sfingolipidi
Ha un duplice effetto:
1. con la sua struttura rigida spezza la regolarità dei legami tra catene aciliche
sature rendendo la struttura meno ordinata (più fluida)
2. Aumenta il grado di impaccamento delle catene aciliche insature rendendo
meno fluida la membrana.
L’effetto finale è uno smorzamento dei cambiamenti di fluidità della membrana

Le proteine di membrana Ogni tipo di cellula possiede un set specifico di proteine di membrana che consente di
compiere funzioni specifiche
Le funzioni principali : trasporto, comunicazione, produzione di energia, locomozione

Proteine periferiche e integrali di membrana
Le proteine integrali attraversano la membrana attraverso lo strato lipidico formando
interazioni idrofobiche con i lipidi.
1 monopasso; 2 multipasso; 3. barile
ß; 4 Inserita su un solo lato
interazioni con
proteine integrali
(7,8) oppure
sono ancorate
alla membrana
da legami
covalenti con
lipidi (5,6).
Le proteine periferiche sono associate con le
membrane attraverso

Proteine di membrana ancorate ai lipidi
Vari modi con cui le proteine sono ancorate alla
membrana
GPI: glicosilfosfatidilinositolo (GPI) un fosfolipide
complesso legato all’estremità carbossiterminale
della proteina.
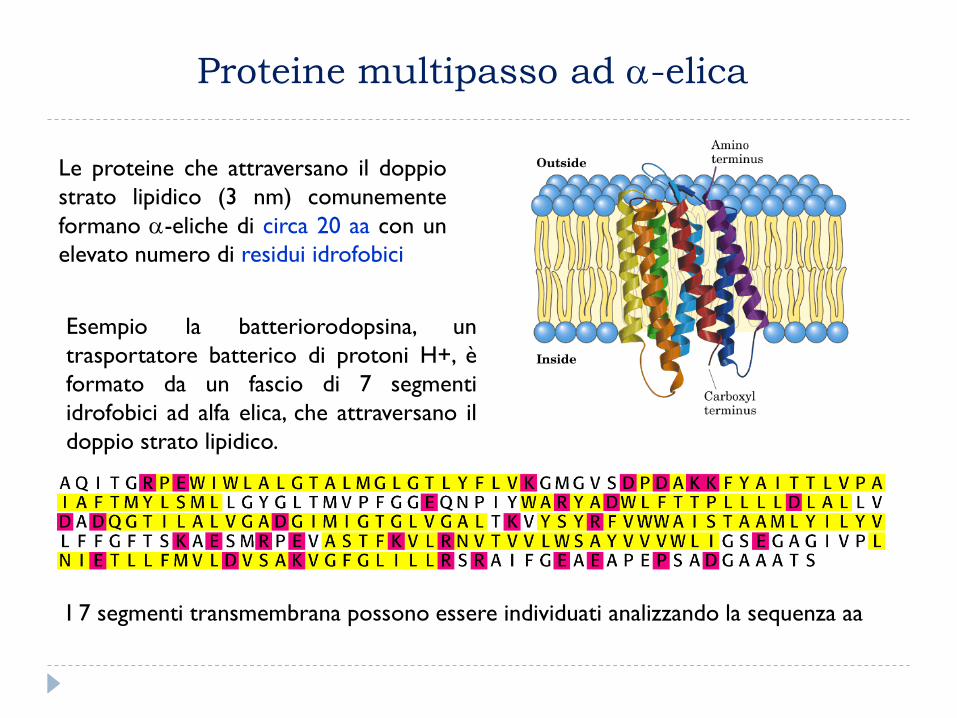
Proteine multipasso ad -elica
Esempio la batteriorodopsina, un
trasportatore batterico di protoni H+, è
formato da un fascio di 7 segmenti
idrofobici ad alfa elica, che attraversano il
doppio strato lipidico.
I 7 segmenti transmembrana possono essere individuati analizzando la sequenza aa
Le proteine che attraversano il doppio
strato lipidico (3 nm) comunemente
formano -eliche di circa 20 aa con un
elevato numero di residui idrofobici

I canali proteici possono essere formati da
catene β
Proteine di membrana con poro
centrale posso anche avere una
struttura formata esclusivamente da
foglietti beta di 16-18 catene
antiparallele. Il canale interno è
idrofilico. Le pareti esterne sono
idrofobiche.
Sequenza amino acidica
I residui in giallo (idrofobici) sono
rivolti all’esterno quelli bianchi
verso l’interno del canale della
porina
I trattini indicano i legami
idrogeno che stabilizzano la
struttura secondaria
La porina è una proteina di membrana esterna dei
batteri Gram-negativi.

Le membrane biologiche sono
selettivamente permeabili
I gas, l’acqua, e alcune sostanze lipofiliche (ormoni steroidei) hanno sufficiente
permeabilità e si muovono per diffusione semplice, dalle regioni di maggiore
concentrazione a quelle a minor concentrazione. Le molecole polari, idrofiliche e
ioniche (gli ioni hanno P di 109 volte inferiori a quello dell’acqua) hanno bisogno di
proteine di membrana per il loro movimento attraverso le membrane
La permeabilità delle piccole molecole è correlata alla loro solubilità in un solvente non
polare relativamente a quello dell’acqua. Coefficienti di permeabilità, P (cm S-1) di alcuni soluti
attraverso le membrane
Video su membrane transport :
https://www.youtube.com/watch?v=mnE_hT3eN6g

Meccanismi di trasporto trans-membrana Trasporto passivo
Il passaggio secondo concentrazione di
composti polari ed ioni avviene per diffusione
facilitata (trasporto facilitato, trasporto passivo)
ad opera di trasportatori proteici selettivi.
Il trasporto di ioni è mediato da trasportatori
(ionofori) o canali ionici.
Trasporto attivo
Alcune sostanze vengono concentrate: il
passaggio avviene con consumo di energia
(ATP) ad opera di specifici trasportatori
(pompe).
In base al numero di soluti i sistemi di trasporto sono classificabili in:
uniporto, simporto e antiporto.

Il trasporto passivo è facilitato da proteine di
membrana
molecola elettricamente neutra
Il passaggio transmembrana di molecole polari
richiede energia (energia di attivazione ΔGǂ ) .
Proteine che forniscono una via alternativa
(Trasportatori o permeasi) (in blu) facilitano il
trasporto. (trasportatori paragonabili a catalizzatori proteici).
Il movimento di una molecola elettricamente neutra avviene in direzione della
minor concentrazione (secondo gradiente di concentrazione = distribuzione
disuguale di concentrazione).

Il trasporto passivo avviene
per mezzo proteine di trasporto
Trasportatori (proteine trasportatrici): legano le molecole da un lato della membrana e
in seguito ad un cambiamento conformazionale della proteina le depositano dall’altro.
Il trasporto è saturabile.
Canali: formano un poro idrofilico all’interno della membrana che permette la
diffusione di ioni specifici attraverso la membrana.
I trasportatori sono proteine transmembrana multipasso.
Ogni trasportatore è altamente selettivo per uno o pochi tipi di composti, si lega al
ligando tramite uno specifico sito di legame.
Due tipi:

Il trasportatore del glucosio
Trasportatore (GLUT-1) proteina 55 kDa con 12 eliche
transmembrana. Il trasporto è 50000 volte superiore
alla diffusione semplice.
Il processo di trasporto è descritto come una reazione
enzimatica con Sesterno e Sinterno
Ci sono 2 conformazioni alternative, T1 sporge all’esterno, T2
sporge all’interno. Ognuna è in grado di legare il glucosio con
siti di legame esposti sulle due superfici.
* Il glucosio non è in scala rispetto alla membrana
*
Il trasporto avviene in
entrambe le direzioni dal
comparto in cui il glucosio è
più concentrato a quello in
cui lo è meno.

Variazione di energia (libera) durante il
trasporto un soluto non carico
La distribuzione disuguale di concentrazione
(gradiente), determina una condizione ricca di
energia. Per formare un gradiente è necessario
spendere energia (trasporto attivo).
Per una specie molecolare non carica il lavoro per
portare una molecola di concentrazione c1 da un
lato della membrana all’altro lato in cui si trova alla
concentrazione c2 è dato da:
ΔG= RTln(c2/c1) = 2,3 RT log10(c2/c1)
R= costante dei gas (8,3 x10-3 kJ mol-1 K-1)
T= Temperatura in gradi Kelvin
Un soluto elettricamente neutro si muove verso la
regione dove la sua concentrazione è minore fino
all’equilibrio.
molecola elett. neutra

Lo spostamento di una carica (senza spostare una
di segno opposto) genera un potenziale elettrico. Il
costo energetico del movimento di uno ione
dipende da:
ΔG= 2,3 RTlog10 (c2/c1) + ZF ΔV
Z = carica della specie trasportata
F = costante di Faraday = 96,5 kJ V-1 mol-1
V = potenziale elettrico (potenziale di
membrana)
Variazione di energia (libera) per il trasporto
di una molecola carica
potenziale elettrochimico = gradiente di concentrazione (chimico) + potenziale
elettrico
+ - molecola carica
Per potenziale di membrana si intende la potenziale elettrico , misurabile in una cellula. Tutte
le tipologie cellulari presentano un potenziale di membrana che si aggira tra -60 mV e -70 mV.
Il segno meno indica proprio che l'interno della cellula è carico negativamente.

Il trasporto attivo trasferisce un soluto contro
gradiente di concentrazione o elettrochimico
Trasporto primario: Accumulo della
molecola S1 è accoppiato ad una
reazione esoergonica :
Conversione di ATP in ADP
Trasporto secondario: Accumulo della
molecola S2 è accoppiato ad un
secondo trasporto esoergonico (in
direzione di gradiente) della molecola
S1:
ΔG= 2,3 RTlog10 (c2/c1) + ZF ΔV >0
Se per portare una molecola di concentrazione c1 da un lato della membrana
all’altro lato in cui si trova alla concentrazione c2 abbiamo che:
Ci troviamo in una condizione di trasporto contro gradiente elettrochimico e per
trasportare la molecola è necessario spendere energia (trasporto attivo). Trasportare
attivamente significa concentrare.

Composizione salina e trasporto di
ioni
25:1 4 mM 100 mM Cl-
25000:1 0.1 M 2.5 mM Ca++
1:35 140 mM 4 mM K+
10:1 14 mM 140 mM Na+
Rapporto
out/In
liquido
intracellulare
liquido
extracellulare
Ione I gradienti ionici negli
organismi sono mantenuti da
un sistema di trasporto:
pompa Na+-K+ (ATPasi)
ATPasi di tipo P comprendono trasportatori
di cationi. L’energia è fornita dall’idrolisi
dell’ATP che catalizza la formazione di un
composto fosforilato P.
Struttura generale delle ATPasi di tipo P Esempi:
uniporto ATPasi Ca++ (SERCA)
antiporto ATPasi Na+ K+ delle cellule animali
ATPasi H+ delle cellule delle piante
Dominio A (Fosfatasi)
Dominio T canale per lo ione (6 eliche)

Trasporto primario: il meccanismo della
pompa per il calcio (SERCA)
•Variazione conformazionale E1/E2
•Fosforilazione ciclica in Asp
•Trasformazione di ATP in ADP
•Uscita di 2 Ca++
SERCA: pompa ATPasi per il
calcio del reticolo endoplasmatico
e sarcoplasmatico

La struttura della pompa del calcio
Le ATPasi di tipo P sono simili tra loro. La Ca2+ ATPasi del reticolo sarcoplasmatico dei
miociti è stata studiata nel dettaglio. Rimuove rapidamente il Ca2+ dal citoplasma e lo
trasferisce al reticolo sarcoplasmatico promuovendo il rilassamento muscolare.
Ca ATPasi formata da un dominio transmembrana (giallo) con sito legame
Ca, e parte citoplasmatica con domini A (adattatore), P (legame P) e N
(legame Nucleotide ATP). Il trasportatore si può trovare nello stato E1
(non fosforilata) o E2

Trasportatori guidati da idrolisi dell’ATP
Le ATPasi sono caratterizzate a seconda della funzione, struttura e del tipo di ioni
che trasportano.
ATPasi di tipo V: (vacuolari). Responsabili
dell’acidificazione sei lisosomi, endosomi,
ATPasi di tipo F: Trasportano protoni .
Fattori di accoppiamento energetico:
utilizzano gradienti di H per formare ATP
(ATP sintasi).
Trasportatori ABC. Pompano amminoacidi, peptidi, proteine, ioni, vari lipidi, sostanze
idrofobiche.

Trasporto secondario: accumulo del glucosio
nelle cellule intestinali.
Il co-trasporto di nutrienti attraverso le membrane delle cellule intestinali è un
processo attivo che muove il glucosio contro gradiente di concentrazione. La fase
che richiede energia per questo co-trasporto è:
– La Na+K+ATPasi che pompa il Na+
dalle cellule epiteliali al sangue
mantenendo bassi i livelli di Na+ nella
cellula
– Il simporto Na+-glucosio sulla
superficie apicale che introduce
glucosio nelle cellule favorito dal flusso
di ioni Na+ secondo gradiente
–Il trasportatore del glucosio sulla
superficie basale che versa il glucosio
nel sangue secondo concentrazione.


















