
Metabolismo e attivazione cerebrale -...
22
Federico Giove Metabolismo e attivazione cerebrale Le tecniche fMRI basate sul contrasto BOLD hanno acquisito negli ultimi anni crescente importanza per le loro buone caratteristiche di definizione spaziale e temporale e per l’assenza di rischi per il paziente. Tuttavia la corretta interpretazione delle mappe risente dell’ambiguità sull’origine fisiologica del segnale BOLD, che presenta una complicata dipendenza da più parametri di tipo vascolare e metabolico. In particolare i meccanismi metabolici che sostengono l’attività neuronale sono stato oggetto di un esteso dibattito a partire dalla metà degli anni ’80, quando si è osservato che in fase di attivazione il flusso sanguigno e il consumo di glucosio aumentano molto di più del consumo di ossigeno 1,2 . Ciò ha suggerito l’ipotesi, inizialmente confermata da altre risultanze sperimentali, che l’energia necessaria al sostentamento dell’attivazione neuronale sia ottenuta da un metabolismo anaerobico, contrariamente a quanto atteso sulla base delle conoscenze sul metabolismo cerebrale a riposo. Coerentemente con questa interpretazione si sono osservati consistenti aumenti della concentrazione cerebrale di lattato (fino a +150%) durante una stimolazione prolungata 3 . Una nuova chiave interpretativa è stata fornita da Luc Pellerin e Pierre Magistretti, che hanno elaborato un modello, cosiddetto della “navetta del lattato” (Astrocyte–neuron lactate shuttle, ANLS), che ipotizza l’intervento obbligato degli astrociti nello svolgimento del metabolismo energetico neuronale 4 . Più in dettaglio secondo questa ipotesi i neurotrasmettitori (specificamente glutammato) rilasciati dai neuroni a seguito dell’attivazione verrebbero assorbiti dagli astrociti, dove stimolerebbero l’uptake di glucosio, l’avvio della glicolisi ed il rilascio di lattato, che si diffonderebbe nei neuroni e verrebbe sfruttato come substrato per il metabolismo aerobico. Per testare questa ipotesi abbiamo realizzato un esperimento di spettroscopia NMR sul protone con il quale abbiamo misurato la concentrazione di lattato nella corteccia visiva nei primi secondi successivi ad una stimolazione impulsiva, in modo da evitare tutte le difficoltà interpretative proprie delle stimolazioni prolungate. Abbiamo osservato una riduzione di circa il 50% della concentrazione di lattato entro 5 secondi dall’attivazione. Questo dato conferma che il lattato svolge un ruolo nell’energetica neuronale, ma non implica che esso sia l’esclusivo substrato neuronale, e d’altra parte non fornisce indicazioni su un eventuale incremento della glicolisi astrocitaria. Non si osserva infatti l’attesa sovrapproduzione iniziale di lattato, tendente a creare il gradiente di concentrazione indispensabile ad attivare il trasporto passivo del lattato fra astrociti e neuroni 5 . Per interpretare questo andamento sperimentale noi ipotizziamo che gli astrociti si attivino in presenza di glutammato al solo scopo di eliminare dallo spazio intersinaptico tale neurotrasmettitore, e che il lattato venga utilizzato dai neuroni nell’ottica dell’ottimizzazione dei substrati, restando il glucosio la principale fonte di energia neuronale, interamente metabolizzata all’interno dei neuroni stessi. Ciò conduce ad una revisione del modello ANLS, che tuttavia richiede il supporto di altri dati sperimentali, soprattutto necessari a separare gli eventi metabolici dei due comparti, astrocitico e neuronale.
Transcript of Metabolismo e attivazione cerebrale -...

Federico Giove
Metabolismo e attivazione cerebrale Le tecniche fMRI basate sul contrasto BOLD hanno acquisito negli ultimi anni
crescente importanza per le loro buone caratteristiche di definizione spaziale e temporale e per l’assenza di rischi per il paziente. Tuttavia la corretta interpretazione delle mappe risente dell’ambiguità sull’origine fisiologica del segnale BOLD, che presenta una complicata dipendenza da più parametri di tipo vascolare e metabolico. In particolare i meccanismi metabolici che sostengono l’attività neuronale sono stato oggetto di un esteso dibattito a partire dalla metà degli anni ’80, quando si è osservato che in fase di attivazione il flusso sanguigno e il consumo di glucosio aumentano molto di più del consumo di ossigeno1,2. Ciò ha suggerito l’ipotesi, inizialmente confermata da altre risultanze sperimentali, che l’energia necessaria al sostentamento dell’attivazione neuronale sia ottenuta da un metabolismo anaerobico, contrariamente a quanto atteso sulla base delle conoscenze sul metabolismo cerebrale a riposo. Coerentemente con questa interpretazione si sono osservati consistenti aumenti della concentrazione cerebrale di lattato (fino a +150%) durante una stimolazione prolungata3.
Una nuova chiave interpretativa è stata fornita da Luc Pellerin e Pierre Magistretti, che hanno elaborato un modello, cosiddetto della “navetta del lattato” (Astrocyte–neuron lactate shuttle, ANLS), che ipotizza l’intervento obbligato degli astrociti nello svolgimento del metabolismo energetico neuronale4. Più in dettaglio secondo questa ipotesi i neurotrasmettitori (specificamente glutammato) rilasciati dai neuroni a seguito dell’attivazione verrebbero assorbiti dagli astrociti, dove stimolerebbero l’uptake di glucosio, l’avvio della glicolisi ed il rilascio di lattato, che si diffonderebbe nei neuroni e verrebbe sfruttato come substrato per il metabolismo aerobico.
Per testare questa ipotesi abbiamo realizzato un esperimento di spettroscopia NMR sul protone con il quale abbiamo misurato la concentrazione di lattato nella corteccia visiva nei primi secondi successivi ad una stimolazione impulsiva, in modo da evitare tutte le difficoltà interpretative proprie delle stimolazioni prolungate. Abbiamo osservato una riduzione di circa il 50% della concentrazione di lattato entro 5 secondi dall’attivazione.
Questo dato conferma che il lattato svolge un ruolo nell’energetica neuronale, ma non implica che esso sia l’esclusivo substrato neuronale, e d’altra parte non fornisce indicazioni su un eventuale incremento della glicolisi astrocitaria. Non si osserva infatti l’attesa sovrapproduzione iniziale di lattato, tendente a creare il gradiente di concentrazione indispensabile ad attivare il trasporto passivo del lattato fra astrociti e neuroni5.
Per interpretare questo andamento sperimentale noi ipotizziamo che gli astrociti si attivino in presenza di glutammato al solo scopo di eliminare dallo spazio intersinaptico tale neurotrasmettitore, e che il lattato venga utilizzato dai neuroni nell’ottica dell’ottimizzazione dei substrati, restando il glucosio la principale fonte di energia neuronale, interamente metabolizzata all’interno dei neuroni stessi. Ciò conduce ad una revisione del modello ANLS, che tuttavia richiede il supporto di altri dati sperimentali, soprattutto necessari a separare gli eventi metabolici dei due comparti, astrocitico e neuronale.

BIBLIOGRAFIA 1 P. T. FOX, M. E. RAICHLE. Focal physiological uncoupling of cerebral blood flow and oxidative metabolism
during somatosensory stimulation in human subjects. Proc. Natl. Acad. Sci. USA, 83:1140–1144, 1986. 2 P. T. FOX et al. Nonoxydative glucose consumption during focal physiologic neural activity. Science, 241:462–
464, 1988. 3 J. PRICHARD et al. Lactate rise detected by 1H NMR in human visual cortex during physiologic stimulation. Proc.
Natl. Acad. Sci. USA, 88:5829–5831, 1991. 4 L. PELLERIN, P. J. MAGISTRETTI. Glutamate uptake into astrocytes stimulates aerobic glycolysis: a mechanism
coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA, 91:10625–10629, 1994. 5 P. J. MAGISTRETTI. Cellular basis of functional brain imaging: insights from neuron–glia metabolic coupling.
Brain Res., 886:108–112, 2000.

Federico GioveUniversità degli Studi di Roma “La Sapienza”
INFM – UdR Roma 1Museo Storico della Fisica e Centro di Studi e Ricerche “Enrico Fermi”
CISBCENTRO INTERDIPARTIMENTALE DI RICERCA PER L'ANALISI DEI MODELLI
E DELL'INFORMAZIONE NEI SISTEMI BIOMEDICI
WORKSHOPSviluppo di metodi per lo studio della funzione e dell'architettura cerebrale
Metabolismo e attivazione cerebraleMetabolismo e attivazione cerebrale

Federico GioveMetabolismo e attivazione cerebrale
Problema: l’attivazione cerebrale
Le tecniche di neuroimaging come PET e fMRI hanno un approccio metabolico-vascolare; in particolare l’fMRI si basa su un tipo di contrasto, il BOLD, che dipende contemporaneamente da:
CBFCBVCMRO2
La comprensione e l’affidabilità delle mappe di attivazione usate in ricerca e in clinica è dunque fortemente influenzata dalla corretta conoscenza del metabolismo che sostiene l’attività neuronale.
Federico GioveMetabolismo e attivazione cerebrale
Il metabolismo neuronale: i protagonisti…
La sintesi dell’ATP avviene nel cervello solo a seguito della demolizione del glucosio.
ATPglucosio
L’ATP è donatore immediato e quasi universale di energia libera. Ogni reazione che richiede energia può avvenire grazie all’accoppiamento con l’idrolisi dell’ATP.
0*2 7.3 /iATP H O ADP P H G kcal mol++ → + + ∆ = −
idrolisi dell’ATP

Federico GioveMetabolismo e attivazione cerebrale
… e la trama
La prima tappa della demolizione del glucosio è la glicolisi, che avviene nel cytosol:
22 2 2 2 2 2 2 2iGlc P ADP NAD Pyr ATP NADH H H O+ ++ + + + + + +„
Va notato il destino simmetrico degli atomi di carbonio dello scheletro del glucosio.
La glicolisi avviene:senza consumo di ossigenovelocemente con rendimento energetico relativamente scarso
Federico GioveMetabolismo e attivazione cerebrale
… e la trama
La glicolisi comporta necessariamente almeno un altro passo, destinato a ristabilire la concentrazione di NAD+:
fermentazione alcolicafermentazione lattica
Piruvato Acetil CoA
+ CO + NADH2+NAD+ + CoA SHformazione di acetil CoA e passaggio al ciclo di Krebs
Piruvato Acetil CoA
+ CO + NADH2+NAD+ + CoA SH

Federico GioveMetabolismo e attivazione cerebrale
… e la trama
La tappa centrale del metabolismo aerobico è il ciclo di Krebs, che avviene nei mitocondri ed in sé non consuma ossigeno
3 2
2 2
3
2 3 2iCH CO SCoA NAD FAD GDP P H O
CO NADH FADH GTP H CoA
+
+
+ + + + +
+ + + + +
: „
„
L’energia viene immagazzinata in elettroni ad alta energia di NADH e FADH2
Gli intermedi del ciclo sono precursori di varie macromolecole biologiche, tra cui aminoacidi e neurotrasmettitori
NADH FADH2
Federico GioveMetabolismo e attivazione cerebrale
… e la trama
La maggior parte della produzione di ATP e l’intero consumo di O2 avvengono durante la catena respiratoria.In questa fase gli elettroni ad alto potenziale generati durante il ciclo di Krebssono ceduti all’O2attraverso più tappe che determinano il mantenimento di un gradiente diconcentrazione protonica attraverso le membrane mitocondriali, a sua volta sfruttato da un enzima che sintetizza ATP.

Federico GioveMetabolismo e attivazione cerebrale
La questione cruciale
Il cervello a riposo consuma glucosio in modo esclusivamente aerobico, ma…
… ciò continua ad essere vero anche in condizioni di attivazione?
Federico GioveMetabolismo e attivazione cerebrale
Un po’ di numeri
scorrelato
scorrelato
scorrelato
scorrelato
scorrelato
trascurabile
ignoto
+30-80%
…… ed in attivazioneed in attivazione
2%Sintesi dei lipidi
1%Sintesi dei nucleotidi
5%Sintesi del glicogeno
6%Sintesi proteica
6%Trasporto assonale veloce
qualche %Sintesi neurotrasmettitori
<1%Messaggeri secondari
65%Pompe ioniche
a riposo…a riposo…Il consumo neuronale di ATPIl consumo neuronale di ATP

Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i dati sperimentali
Nel 1986 P. Fox e M. Raichle misurano con la PET* un disaccoppiamento in condizioni di attivazione fra le variazioni di CMRO2 da un lato e CMRglc e CBF dall’altro.
Ciò li portò ad ipotizzare che l’attivazione neuronale è sostenuta energeticamente da un metabolismo anaerobico.
Quest’idea è stata inizialmente confortata da altre indicazioni provenienti da:
*PNAS 83:1140 (1986)
fMRIRiflettanza otticaMRS
Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i dati sperimentali
Nel 1986 P. Fox e M. Raichle misurano con la PET* un disaccoppiamento in condizioni di attivazione fra le variazioni di CMRO2 da un lato e CMRglc e CBF dall’altro.
Ciò li portò ad ipotizzare che l’attivazione neuronale è sostenuta energeticamente da un metabolismo anaerobico.
Quest’idea è stata inizialmente confortata da altre indicazioni provenienti da:
*PNAS 83:1140 (1986)
fMRIRiflettanza otticaMRS
Federico GioveMetabolismo e attivazione cerebrale
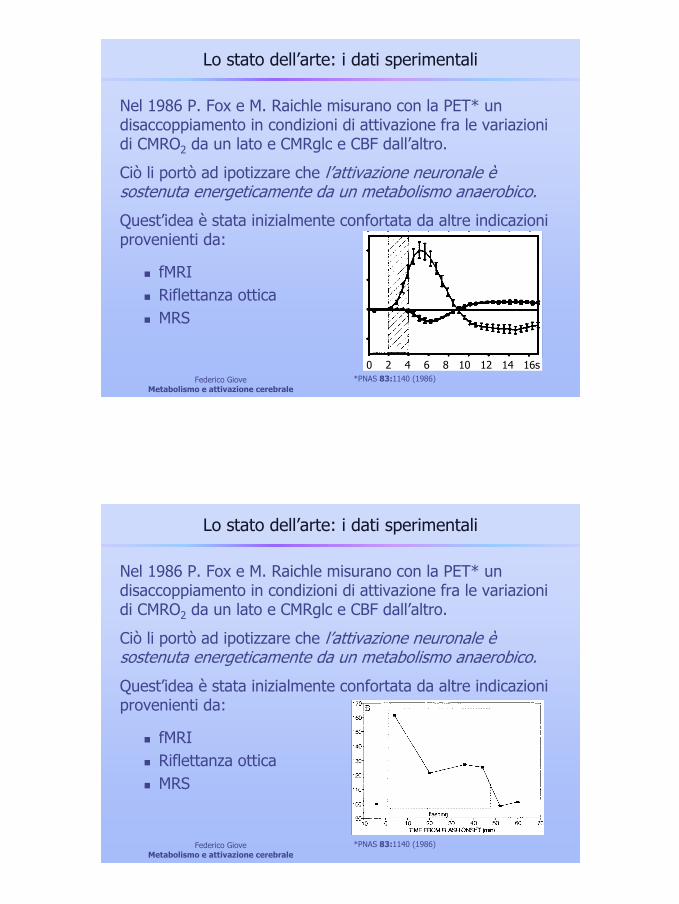
Lo stato dell’arte: i dati sperimentali
Nel 1986 P. Fox e M. Raichle misurano con la PET* un disaccoppiamento in condizioni di attivazione fra le variazioni di CMRO2 da un lato e CMRglc e CBF dall’altro.
Ciò li portò ad ipotizzare che l’attivazione neuronale è sostenuta energeticamente da un metabolismo anaerobico.
Quest’idea è stata inizialmente confortata da altre indicazioni provenienti da:
*PNAS 83:1140 (1986)
fMRIRiflettanza otticaMRS
0 2 4 6 8 10 12 14 16s
Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i dati sperimentali
Nel 1986 P. Fox e M. Raichle misurano con la PET* un disaccoppiamento in condizioni di attivazione fra le variazioni di CMRO2 da un lato e CMRglc e CBF dall’altro.
Ciò li portò ad ipotizzare che l’attivazione neuronale è sostenuta energeticamente da un metabolismo anaerobico.
Quest’idea è stata inizialmente confortata da altre indicazioni provenienti da:
*PNAS 83:1140 (1986)
fMRIRiflettanza otticaMRS

Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i dati sperimentali
25%48%BOLD-PERFUSION2%61%BOLD-PERFUSION
15.6%43.6%BOLD-PERFUSION4%74%BOLD-PERFUSION16%45%BOLD 5.7%43%BOLD
21%BOLD-PERFUSION-MRS0%15%12%Kety-Schmidt
totale:23,4%ossidativo:11,7%PET
0%43%27%PET15-25%15-25%PET
22%MRS50%MRS
28%40%PET5%50%51%PET13%7%PET5%29%PET
∆∆CMROCMRO22∆∆CBFCBF∆∆CMRglucCMRglucTecnicaTecnica
Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i dati sperimentali
GROUP PROTOCOL OF STIMULATION VOXEL D[LAC]% max
J. PRICHARD et al. 1991
VISUAL STIM.: 5×6 grids of red light emitting diodes, flashing at 16Hz for 48min; temp. res.:6-8min, 128 scan
13cm3 +57%
D. SAPPEY-MARINIER et al. 1992
VISUAL STIM.: Black/white checkerboard, flashing at 2Hz for 40min; temp res. 6.4min, 256 scan
31.5cm3 (3.5×3×3cm3) +150%
K. D. MERBOLDT et al. 1992
VISUAL STIM. : red LED matrix flashing at 4÷16 Hz for 6.5min 8÷64cm3 ±0%
M. SINGH et al. 1992
AUDITORY STIM.: 1KHz tone burst at a repetition rate of 1Hz and 40Hz, lasting30-60min; temp. res: 3.2 min, 64scan
27cm3
(3×3×3cm3) +15%
B. G. JENKINS et al. 1993
VISUAL STIM.: red LED matrix flashing at 7.8Hz for 3 or 6min; temp res: 3 or 6min, 90 or 180 scan, ~25 averages/spectrum
15.6cm3
(2×3×2.6cm3)+92% (3’ stim) +76% (6’ stim)
T. KUWABARA et al. 1995
SENSORYMOTOR STIM.: Finger movements at 1Hz; temp. ris.: 6.5min, 256scan
56cm3 (3.5×4×4cm3) +20%
R.S.MENON et al. 1997
VISUAL STIM.: radial checkerboard flashing at 8Hz for 40s, then 40s rest, for 8×2min; temp res.: 2s, 8 averages/spectrum
11.34cm3
(4.2×2.7× ×1cm3)
+24%
FRAHM J. et al. 1996-
1997
VISUAL STIM: pattern flickering at 10Hz for 6min; temp res: 30s, 25 averages/spectrum
11.2cm3 +70%

Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i dati sperimentali
Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i dati sperimentali

Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i modelli proposti
DUNQUEDUNQUE“Il substrato metabolico dei neuroni è lattato e non glucosio”“Il substrato metabolico dei neuroni è lattato e non glucosio”
(ANLS model)*(ANLS model)**PNAS 91:10625 (1994)
Gli astrociti assorbono glutammatoIl glutammato stimola uptake di glucosio, glicolisi e rilascio di lattato da parte degli astrocitiIl glutammato assorbito viene rilasciato come glutammina, ed il ciclo glu-gln è in rapporto 1:1 col CMRglc(O2)I neuroni esprimono solo LDH1, gli astrociti LDH5Nel cervello sono presenti MCT1 e MCT2Gli astrociti costituiscono una barriera tra vasi e neuroni…
Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i modelli proposti

Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i modelli proposti
*MRM 35:143 (1996)
Federico GioveMetabolismo e attivazione cerebrale
Lo stato dell’arte: i modelli proposti
Il modello di Magistretti ha rapidamente assunto dignità di “paradigma” nonostante i numerosi punti deboli
L’ipotesi di Fox è stata di fatto scartata, pur in assenza di evidenze sperimentali a diretta smentita, ed oggi si dà per scontato che il metabolismo cerebrale sia complessivamente aerobico.

Federico GioveMetabolismo e attivazione cerebrale
Un approccio originale: la spettroscopia MR event related
Vogliamo investigare la dinamica del lattato nei primi secondi dopo l’attivazione
TR>7 sTE 270 msdurata dello stimolo 1 sRitardo d’acquisizione (RA) 0, 3, 5, 8 sacquisizione singola ripetuta, tipicamente 128 voltestimolazione “interblob”MRS single voxel (PRESS)
Federico GioveMetabolismo e attivazione cerebrale
Un approccio originale: la spettroscopia MR event related
Stimolazione interblob di V16/°, eccentricità<5%, contrasto 100%
Acquisizione
RA RAAcquisizione
Stimolazione (1s) Riposo (≥6.5 s) Stimolazione (1s) Riposo (≥6.5 s)
TR (≥7.5 s) TR (≥7.5 s)

Federico GioveMetabolismo e attivazione cerebrale
Un approccio originale: la spettroscopia MR event related
Federico GioveMetabolismo e attivazione cerebrale
Un approccio originale: la spettroscopia MR event related

Federico GioveMetabolismo e attivazione cerebrale
Si osserva dinamica rapida del lattato, ciò dimostra senza ambiguità che nei primi secondi dopo l’attivazione
Il consumo è più rapido dell’eventuale produzioneIl lattato decresce sensibilmente, fino al 50% della sua concentrazione a riposo
Il lattato ha un ruolo metabolico nell’attivazione neuronaleMA…
Ciò non implica che il lattato sia l’unico substrato metabolico (anzi appare improbabile)Non si ha alcuna evidenza di un incremento del metabolismo glicolitico astrocitario tendente a rifornire i neuroni
Dilemmi interpretativi
Federico GioveMetabolismo e attivazione cerebrale
L’ANSL dovrebbe implicare un’iniziale sovrapproduzione di lattato e comunque uno stretto accoppiamento tra ∆CMRgluc e ∆CMRO2
Il picco di lattato che si osserva in stimolazioni prolungate hauna dinamica del tutto incompatibile con le necessità energetiche dei neuroni: potrebbe essere legato ad un altro fenomeno e comunque indicare lentezza di trasportoRisulta inspiegabile l’assenza di specifici meccanismi di trasportoI neuroni sono ben attrezzati a assorbire glucosio e non si vedeil vantaggio metabolico di usare lattatoLe indicazioni sull’adeguatezza del lattato a sostenere l’energetica neuronale sono ambigue
Dilemmi interpretativi

Federico GioveMetabolismo e attivazione cerebrale
Una nuova chiave di lettura
I nostri dati evidenziano il ruolo del lattato nell’energetica neuronale attivativa in vivo sull’uomo. Tuttavia riteniamo che siano meglio interpretabili in un contesto diverso dall’ANSL, inparticolare riguardo a…
Ruolo del lattatoProvenienza del lattato
Di conseguenza il ruolo degli astrociti viene ridimensionato: danutrici dei neuroni a spazzini intercellulari
Federico GioveMetabolismo e attivazione cerebrale
Una nuova chiave di lettura
Glucose of capillary (uptake stimulated by synaptically released glutamate)
Pyruvate
LactateLactate
Pyruvate
34ATP
1ATP 1ATP
NEURONNEURON ASTROCYTEASTROCYTE
An aero bic glyco lisis
Na+-K+
pumpGlu-glncycle
TCA cycle
glycogen
Anaero bic glyco lisis
ASTROCYTE FOOTASTROCYTE FOOT

Federico GioveMetabolismo e attivazione cerebrale
Una nuova chiave di lettura
Quest’ipotesi è consistente con le principali evidenze sperimentali:
L’aumento di [Lac] durante stimolazioni prolungateLa presenza di GLUT3 nei neuroniL’assenza di un meccanismo di efficiente trasporto del lattatoIl disaccoppiamento fra ∆CMRglc and ∆CMRO2
L’interpretazione dei dati nell’ambito ANLS è possibile, ma apparirebbe forzata
Federico GioveMetabolismo e attivazione cerebrale
Conclusioni
Il principale substrato metabolico dei neuroni resta il glucosioL’uso del lattato può essere rilevante solo nelle prime fasi dell’attivazioneIl metabolismo neuronale è strettamente aerobicoIl ruolo nutrizionale degli astrociti appare non accettabile

Federico GioveMetabolismo e attivazione cerebrale
Che fare dell’ANSL?
I nostri dati sono molto interessanti, ma sconvolgono le attualivisioni complessive sul metabolismo neuronale.
In letteratuta cominciano a trovarsi alcuni esperimenti che vanno nella direzione da noi suggerita; in ogni caso…
SERVONO NUOVI DATI!
Federico GioveMetabolismo e attivazione cerebrale
Prospettive future
Investigare ulteriori RAUtilizzare una stimolazione più lungaVerificare la dinamica del flusso con un esperimento di perfusione event-related realizzato nelle medesime condizioni sperimentaliCondurre uno studio event-related di 31P MRSEffettuare simulazioni numeriche sulla cinetica dei fenomeni osservatiPianificare studi in vitro per determinare il substrato preferenziale dei neuroni

Federico GioveMetabolismo e attivazione cerebrale
Ci proponiamo di trovare “gradi di compatibilità”La soluzione analitica della cinetica chimica coinvolta è difficileUna simulazione numerica appare fattibile in tempi ragionevoli.Tale simulazione dovrà tenere conto:
del termine “forzante” (i parametri ematici sistemici)del ritardo nel trasporto sanguignodelle strutture presentidei coefficienti di diffusione e di permeabilitàdel consumo di ossigeno regionale
Approssimazioni:compartimenti ben miscelati?reazioni limitanti
Simulazioni numeriche
Federico GioveMetabolismo e attivazione cerebrale
Simulazioni numeriche
Buffer
Circolo cerebrale
Neuroni Astrociti
O2GlcLac
Spazio intercellulare
CB(t)=cost
CC(t)=CB(t)θ(t-τ)+a

Federico GioveMetabolismo e attivazione cerebrale
La spettroscopia in vitro su colture cellulari
Colture primarie13C MRS, 31P come controlloU- 13C Glc e U- 13C LacStimolazione chimica (glutammato) o “elettrica” (K+)Volume del campione ~1 ml, concentrazione cellulare 107 /mlCondizioni fisiologiche, ipoglicemiche, postipossicheVari rapporti neuroni-astrocitiTempi di acquisizione da mezz’ora in su
*Brain Res. Prot. 3:183 (1994)
*
Federico GioveMetabolismo e attivazione cerebrale
La spettroscopia in vitro su colture cellulari

Federico GioveMetabolismo e attivazione cerebrale
La spettroscopia in vitro su colture cellulari
*Brain Res. Prot. 3:183 (1994)
Principali metaboliti osservabili:2-13C 3-13C 4-13C Glu2-13C 3-13C 4-13C Gln2-13C 3-13C 4-13C GABA2-13C 3-13C Ala3-13C Lac2-13C 3-13C Asp
Da intermedi del ciclo di Krebs derivano fra gli altri Glu, Gln, GABA e AspDa intermedi della glicolisi derivano fra gli altri AlaDunque abbiamo uno strumento eccezionalmente sensibile al metabolismo energetico mentre esso avviene
Bruno Maraviglia
Francesco Di Salle
Girolamo Garreffa
Marta Bianciardi
Renato Morrone
Silvia Mangia
e
Alfredo Colosimo
E infine, in ordine semialfabetico …


















