METABOLISMO DEL DNA
21
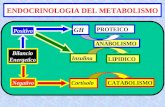
1 METABOLISMO DEL DNA Replicación de DNA Facultad de Química, UNAM 1630 Genética y Biología Molecular Durante el Ciclo celular se controla la replicación del DNA y la división de la célula P-M-A-T Energía Química derivada de la hidrólisis de ATP Enlaces fosfoanhídrido Enlaces fosfoéster ∆G 0 ’ = -14 kJ/mol ∆G 0 ’ = -30 kJ/mol ATP + H 2 O ↔ ADP + P i La hidrólisis de ATP favorece termodinámicamente las reacciones acopladas Energía Química derivada de la hidrólisis de ATP Muchas enzimas involucradas en el metabolismo del DNA requieren ATP. • Tienen sitios de unión a ATP • Tienen actividad de ATPasa • Transducen la energía química derivada de la hidrólisis de ATP a energía mecánica. • Se refleja como cambios conformacionales en la estructura de la proteína.
-
Upload
daniela-suarez-echalar -
Category
Documents
-
view
43 -
download
0
Transcript of METABOLISMO DEL DNA

1
METABOLISMO DEL DNA
Replicación de DNA
Facultad de Química, UNAM
1630 Genética y Biología Molecular
Durante el Ciclo celular se controla la replicación del DNA y la división de la célula
P-M-A-T
Energía Química derivada de la hidrólisis de ATP
Enlaces fosfoanhídrido
Enlaces fosfoéster∆G0’ = -14 kJ/mol
∆G0’ = -30 kJ/mol
ATP + H2O ↔ ADP + Pi
La hidrólisis de ATP favorece termodinámicamente las reacciones acopladas
Energía Química derivada de la hidrólisis de ATP
Muchas enzimas involucradas en el metabolismo del DNA requieren ATP.
• Tienen sitios de unión a ATP
• Tienen actividad de ATPasa
• Transducen la energía química derivada de la hidrólisis de ATP a energía mecánica.
• Se refleja como cambios conformacionales en la estructura de la proteína.

2
Generalidades de la Replicación de DNA
La replicación de DNA es:
• Semiconservativa
• Semidiscontinua
• Bidireccional
• Dependiente de un cebador de RNA
Mecanismo semiconservativo de la replicación del DNA
Estructura del DNA
Doble Hélice
Cadenas complementarias
¿Pruebas experimentales del mecanismo de replicación semiconservativo?
Hebras parentales Replicación Hebras hijas
Semiconservativo
Conservativo
Dispersivo al azar
Experimento de Meselson y Stahl
Separación de DNA por centrifugación en gradiente de CsCl
Alimentar cultivo de E. coli con fuente de 15N y purificar el DNA
Se deja otro ciclo de replicación
Experimento de Meselson y StahlComprobación del mecanismo semiconservativo de Replicación
Consistente con un mecanismo semiconservativo
DNA (15N)DNA híbrido
(15N/14N)

3
La replicación del DNA es semidiscontinua
Síntesis continua requiere que las DNA polimerasas sintetizen DNA de 5’->3’ y de 3’ a 5’
Síntesis semidiscontinuapredice que una cadena se sintetiza en forma continua y la otra en forma discontinua.
3’3’
3’
5’
5’
5’
Como la dirección de síntesis de la cadena discontinua es opuesta a la dirección de apertura de la horquilla, se
comienza una nueva cadena.
En cambio, la cadena continua simplemente se sigue elongando
Reiji Okazaki
*
3’3’
3’
5’
5’
5’
5’
5’
5’
3’
3’
3’
Como la dirección de síntesis de la cadena discontinua es opuesta a la dirección de apertura de la horquilla, se
comienza una nueva cadena.
En cambio, la cadena continua simplemente se sigue elongando
Reiji Okazaki
*
*
Durante la replicación del DNA se sintetizan fragmentos que varían entre 1000 a 2000 nucleótidos
Replicación semidiscontinua:
Una hebra se sintetiza en forma continua y la otra discontinua
Cadena adelantada (leading)/ Cadena retrasada (lagging)
Estos fragmentos corrsponden a la cadena discontinua.
Se llaman fragmentos de Okazaki.

4
¿Es la replicación del DNA unidireccional o bidireccional?
En bacterias, que tienen genoma circular cerrado
Replicación del DNA de E. coli por 1.5 generaciones en presencia de nucleótidos marcados
Autoradiografía
Demostración experimental de que la replicación del DNA es bidireccional
Experimento de Gyurasits y Lake
DNA de Bacillus subtilis (ventajas?)
Pulso de baja intensidad en el inicio
Pulso de alta intensidad
Resultados del Experimento de Gyurasits y Lake
La replicación de DNA requiere de la síntesis de RNA
• El antibiótico rifampicina inhibe la replicación del DNA de fago M13 cuando se usa un extracto de E. colicomo fuente de enzima.
• La DNasa no hidroliza completamente los fragmentos de Okazaki. Deja cadenas de 10 a 12 oligonucleótidos de largo.
• La naturaleza de los oligonucleótidos es RNA.
• La síntesis de DNA requiere ser comenzada por cebadores de RNA que luego son elongados por la DNA polimerasa.

5
Topología del DNA y Topoisomerasas
La naturaleza de doble hebra del DNA tiene varias implicaciones para su función biológica:
La Replicación se parece facilitar ya que la información genética está codificada por duplicado en la estructura del DNA.
También, las limitaciones impuestas por la naturaleza de doble hélice en el DNA implica que muchos eventos que involucran desenrollamiento (relajación) causan tensión en la molécula.
El DNA puede pasar de una forma superenrollada a una forma más relajada
Las distintas formas de enrollamiento de un DNA se pueden resolver por electroforesis en geles de
agarosa
El DNA superenrollado migra más rapidamente que una molécula relajada del mismo tamaño.
El DNA separado solamente difiere en su topología, por lo que a las distintas formas se les llama topoisómeros.
Aunque los cromosomas eucariontes son lineales, forman asas de DNA que están unidas a la matriz nuclear.
Estas asas representan dominios topológicos que sonequivalentes a los dominios de DNA circular.
Muchos eventos metabólicos del DNA requieren la apertura de la doble hélice.
• La apertura de la doble hélice está restringida puesto que las hebras que se separan no tienen rotación libre. El desenrollamiento de las dos hebras se convierte en un evento topológicamente imposible.
• A un segmento de DNA que está constriñido en la rotación de los extremos, se le llama dominio topológico.
• Un ejemplo canónico de un dominio topológico es un DNA circular, que es típico del genoma de bacterias, mitocondria, cloroplastos y los plásmidos.

6
Mecanismo de la Topoisomerasa
Una cadena de la doble hélice es cortada y la enzima permanece unida covalentemente al fosfato en el extremo 5’.
La hebra intacta del DNA pasa a través del corte y la enzima vuelve a formar el enlace fosfodiéster.
Relación entre las propiedades de la replicación de DNA y las enzimas que lo
realizan
¿Cómo se unen los fragmentos de Okazaki?
¿Cómo es reconocido el origen de replicación?
¿Cómo se separan las dos hebras de DNA?
¿Cómo se mantienen las dos hebras separadas?
¿Qué enzima sintetiza el cebador de RNA?
La replicación de DNA comienza en una zona definida del genoma llamada origen de replicación
En Escherichia coli el locus oriC es de 245 pb
La secuencia del origen de replicación en procariontes está altamente conservada
El origen de replicación en procariontes es muy conservado
GATC: Sitios de reconocimiento Metiltransferasa Dam
4 cajas dnaA (nonámeros)
3 repeticiones de 13 nucleótidos/ Región donde comienza a abrirse el DNA (65 – 70% A + T) Región P1

7
1. La proteína HU causa torsiones en el DNA y facilita su unión a la proteína DnaA
Reconocimiento del oriC
2. La unión del complejo HU-ATP-DnaA desestabiliza la doble hélice en la región P1
Proteína Dna A
• Tiene una muy alta afinidad por ATP y lo hidroliza
lentamente a ADP en una forma dependiente de
DNA.
• Actúa como regulador de la iniciación.
• Se une con alta afinidad y de forma cooperativa a
las cajas dnaA de oriC. Aprox. 30 subunidades de
DnaA por origen de replicación.
• Se une a DNA de cadena sencilla.
• Facilita la unión de la helicasa al origen de
replicación.
3. Unión de DnaB a la región abierta del DNA.

8
Proteína DnaB. Helicasa
Forma un homohexámero.
Tiene dominios que se requieren para:
•Unión a DNA de cadena doble
• Activación de la primasa
• Hidrólisis de ATP
La helicasa rodea una de las hebras del DNA duplex y se desplaza logrando la apertura de la doble hélica por exclusión estérica.
Una hebra es excluida del canal interno, mientras que la otra hebra es retenida en el interior del anillo.
Unión de la helicasa (dnaB) al origen de replicación y apertura de la doble hélice de DNA
Proteína SSB “Single-strand binding”
La proteína SSB se une al DNA de cadena sencilla con alta afinidad y así previene que se vuelva a formar el híbrido DNA-DNA. Actúa de manera cooperativa con la actividad de helicasa.
DnaG Primasa
La primasa sintetiza oligo-ribonucleótidos de 10 a 12 unidades de longitud, usando rNTPs. Es dependiente de molde y la secuencia corresponde al origen de replicación.
El extremo 3‘ del último ribonucleótido es extendido como DNA por la acción de la DNA polimerasa. Se forma una unión covalente entre RNA y DNA

9
INICIACION
Una vez que el cebador está en su lugar comienza la síntesis de DNA: ELONGACION
La actividad de DNA polimerasa cataliza la adición de un dNTP al extremo 3’-OH de un polidesoxinucleótido por un mecanismo de desplazamiento nucleofílico .
Éste es complementario al DNA molde.
ELONGACION
DNA POLIMERASAS
DNA polimerasa I (A. Kornberg)
Fue la primera DNA polimerasa descrita y caracterizada.
Representa la función de otras DNA polimerasas.
Enzima dependiente de molde (DNA)
Síntesis de una cadena de DNA en dirección de 5’ 3’
Es un monómero de 102 kDa
Tiene 3 actividades:
1. Polimerización DNA
2. Actividad de exonucleasa 3’-> 5’
3. Actividad de exonucleasa 5’ -> 3’
Fragmento Klenow de la DNA Pol I
324 520 928
Exo 5’-3’ Exo 3’-5’ Pol
N C

10
La FIDELIDAD se refiere al seguimiento exacto de la secuencia que sirve como molde. En promedio, las DNA pols, 1 error por cada 108 nts
La actividad de exonucleasa 3’ 5’ de la DNA polimerasa contribuye a la fidelidad pues tiene actividad correctora (proofreading).
Hay dos propiedades importantes de las DNA POLIMERASAS:
FIDELIDAD Y PROCESIVIDAD
La PROCESIVIDAD se refiere a la capacidad de una DNA polimerasa de elongar una cadena de DNA por muchos nucleótidos antes de disociarse del complejo que forma con el sustrato.
La DNA polimerasa I tiene una procesividad baja.
La DNA polimerasa III tiene una procesividad alta.
Competencia cinética entre la actividad de Polimerasa y de Exonucleasa
Descubrimiento de la DNA polimerasa III
Mutantes de E. coli en el gen de Pol I son viables, por lo que la DNA Pol I no es la principal enzima en replicación.
Sin embargo, en E. coli w.t.; por cromatografía de intercambio iónico, copurifica y enmascara otra actividad de DNA polimerasa.
Subunidad α: 129 kDa, tiene actividad de DNA polimerasa
Subunidad ε: 27.5 kDa, tiene actividad de exonucleasa 3’ 5’
La DNA polimerasa III está compuesta por muchas subunidades distintas. Holoenzima Pol III
La DNA polimerasa III requiere de la subunidad βpara incrementar su procesividad
Cuando se une la subunidad β se incrementa la procesividad de la DNA pol III de 10 nts. a mas de 50,000 nts. Así, puede sintetizar 1000 nts/seg.
La subunidad β es el factor de procesividad y actúa como una abrazadera o pinza móvil que se une al DNA.
Forma un homodímero funcional compuesto por dos subunidades de 40.6 kDa que permanecen unidos a DNA circular cerrado.
El ensamblado del complejo γ requiere la unión de ATP que es hidrolizado durante el cargado de la
subunidad β al DNA

11
En la DNA polimerasa III hay dos núcleos del elemento catalítico (core).
Para que la DNA polimerasa dimérica pueda sintetizar DNA sobre ambas cadenas, el molde de DNA de la cadena retrasada se debe doblar unos 180o para aparecer que va en la misma dirección que la cadena adelantada.
Función de la DNA pol III dimérica
TERMINACIÓN DE LA REPLICACIÓN EN PROCARIONTES
Las dos horquillas de replicación se aproximan a la misma región que contiene las cajas Ter. Son secuencias de 22 pb, tambien llamados sitios de terminación. Están presentes en tandem (seis) en forma invertida.
A estas secuencias se unen las proteínas TUS (TBP).
La presencia de estas proteínas de unión a DNA causa que se detenga el avance de las horquillas.
Tus: termination utilization substance TBP: Termination binding protein. Proteína de 36 kDa que afecta la actividad de la DNA helicasa (DnaB).
TERMINACIÓN. Desenrrollamiento y Síntesis reparativa.
Las dos hebras duplex, productos de la replicación están enrrolladas
La topoisomerasa IV contribuye a la desnaturalización y descatenación de las hebras. Mutantes en el gene de TopoIV exhiben cromosomas que no se han separado totalmente.
Ocurre síntesis reparativa para llenar los huecos

12
Función de la DNA polimerasa I en la replicación
Función de la DNA ligasa en la replicación
1. Formación del intermediario Enzima ATP
2. Transferencia del adenilo al 5’-P
3. Formación del enlace fosfodiéster
La DNA ligasa de E. coli es una enzima de 75 kDa. Es muy lábil
REPLICACIÓN DE DNA EN EUCARIONTES
En eucariontes, la replicación comienza en muchos sitios a lo largo de los cromosomas.
Los orígenes de replicación de metazuarios no están definidos por una secuencia específica, como el oriC.
Sino que consisten en:
• Sitios de inicio de alta frecuencia
• Sitios de inicio de baja frecuencia
Los orígenes de replicación se establecen durante la fase G1 del ciclo celular y dependen de mucho parámetros:
• Estructura nuclear
• Contenido de G+C ([G+C], mayor que el resto del genoma)
• Modificaciones en el DNA. Baja metilación en G y C.
•Estructura de la cromatina. Histonas H3 metiladas y H4 acetiladas.
Permite modificar el número y localización de los orígenes de rep.

13
ELONGACIÓN EN EUCARIONTES
DNA POLIMERASAS
Se han identificado mas de 20 DNA polimerasas en eucariontes.
La mayoría participa en eventos de reparación de DNA.
Las DNA polimerasas replicativas son:
DNA polimerasa α. Procesividad baja. Tiene actividad de primasa.
DNA polimerasa δ. Procesividad alta en presencia de PCNA.
DNA polimerasa ε. Procesividad alta. No requiere PCNA.
Proliferating-Cell Nuclear Antigen (PCNA)
• PCNA es una proteína de 29 kDa.
• Forma un trímero alrededor del DNA.
• Incrementa la procesividad de la DNA pol delta hasta 40 veces.
•Se ha demostrado su interacción in vitro con mas de 50 proteínas. Entre ellas:
• Ciclina D1, cdk2 y el inhibidor de cdks
TERMINACIÓN EN EUCARIONTES
El dilema de los cromosomas lineales
Durante la terminación en procariontes, hay hidrólisis del cebador pero el extremo 3’ de la cadena funciona para cebar la síntesis que así completa la cadena.
Sin embargo, en los cromosomas lineales de eucariontes, después de eliminar al cebador no hay forma de completar la síntesis.
Esto implica que los cromosomas se irían acortando después de cada ronda de duplicación
TERMINACIÓN EN EUCARIONTES
El dilema de los cromosomas lineales. La Solución
Telomerasa. Enzima que adiciona secuencias cortas que se repiten en los extremos de los cromosomas. Telómeros.
Las secuencias repetidas varían entre especies:
Tetrahymena TTGGGG
Humano TTAGGG
Paramecium TTGGGG
Trypanosoma TTAGGG
Arabidopsis TTTAGG

14
La telomerasa es una DNA polimerasa que utiliza RNA como molde
La telomerasa está compuesta por dos subunidades:
• Subunidad catalítica (proteína)
• Subunidad de RNA asociada
Funciona como molde para elongación de una de las cadenas.
Mecanismo de acción de la telomerasa
Mecanismo de acción de la telomerasa
El resultado:
La telomerasa está presente en células embrionarias, pero en células somáticas su actividad es muy baja.

15
METABOLISMO DEL DNA
Reparación de DNA
Facultad de Química, UNAM
1630 Genética y Biología Molecular
El DNA puede ser dañado de muchas maneras, pero este daño solamente conduce a una mutación cuando no es reparado.
• Daño: Se refiere a cambios químicos en el DNA.• Mutación: Se refiere a cambios en la secuencia de bases del DNA.
Agentes físicos naturales que dañan al DNA. Radiación UV solar.
Formación de dímeros de timina.
Entrecruzamiento covalente de pirimidinas adyacentes en la misma cadena de DNA. Generalmente son timinas.
Agentes químicos naturales que dañan al DNA. AFLATOXINAS
Formación de aductos covalentes con el DNA

16
Agentes químicos sintéticos que dañan al DNA.
EMS => Acetilación / Nitrosamina => Metilación
Metilación o acetilación de las bases en los átomos (O/N) que participan en la formación de puentes de H, desestabiliza la doble hélice de DNA y hace esa zona susceptible a mutación.
La presencia de estas regiones dañadas puede causar detención de la replicación, o si ésta continúa, está sujeta a errores.
Agentes físicos que causan daño al DNA.
Rayos X y rayos gamma.
Ionizan moléculas que rodean al DNA generando radicales libres, algunos de estos contienen oxígeno que tienen un electrón desapareado. Son especies altamente reactivas y pueden atacar la molécula del DNA causando rupturas en una o en las dos cadenas.
Los radicales libres que se forman también pueden modificar quimicamente una base, como la guanina.
Generación de 8-oxo-guanina. Causa transversiones: GC -> TA
Tipos de mutaciones de pares de bases.
CATTCACCTGTACCAGTAAGTGGACATGGT
CATGCACCTGTACCAGTACGTGGACATGGT
CATCCACCTGTACCAGTAGGTGGACATGGT
Transición (T-A to C-G) Transversión (T-A to G-C)
CATCACCTGTACCAGTAGTGGACATGGT
Deleción o eliminación
CATGTCACCTGTACCAGTACAGTGGACATGGT
Inserción
Sustituciones de pares de bases• transición: pirimidina a pirimidina• transversión: pirimidina a purina
Secuencia silvestre

17
Cuando los daños no son reparados, se pueden generar cambios en las secuencia de las bases durante la replicación, por lo lo que se genera una mutación.
REPARACIÓN.
•Corrección directa del DNA dañado.
• Eliminación de la región dañada del DNA y rellenarlo con DNA recien sintetizado.
Reparación Directa de Daños al DNA. Fotoreactivación
Sistema de reparación que se activa en presencia de luz.
Fotoliasa. Detecta al DNA dañado y se une a éste. La enzima absorbe luz azul y se activa. Rompe los enlaces covalentes entre los dímeros de timina.
La enzima se disocia y se separa del DNA.
Reacción dependiente de FADH
Reparación de DNA por EsciciónImplica la remoción del DNA dañado y de secuencias adyacentes también.
BER: “Base excision repair”
NER: “Nucleotide excision repair”
Reparación de DNA por Escición de Nucleótidos (NER)
Escinucleasa uvrABC realiza este tipo de reparación en dímeros de timina, otros fotoproductos y bases dañadas.
Escinucleasa (246 kDa) está compuesta por tres subunidades (A, B y C)
UvrA se une al DNA en la región dañada.
UvrB/UvrC tienen actividad de endonucleasa y corta en los lados adyacentes de la cadena liberando un oligonucleótido de unos 12-13 nts de largo.
La región “vacía” es rellenada por una DNA polimerasa I y sellada por una DNA ligasa.

18
• Cuando se expone E. colia altos niveles de radiación o a un mutágeno, ocurre un daño extensivo en el DNA. La célula responde induciendo la vía SOS de reparación.• Se activan los genes umuC y umuD cuyos productos componen las subunidades de la DNA polimerasa V. • DNA PolV replica el DNA, aun en regiones donde hay daño y es muy propensa a cometer errores. • Se observa una alta tasa de mutación
• Cepas de E. coli nulas en umuC son inmutables.
Reparación sujeta a errores
Reparación por escición de nucleótidos en eucariontes
1. Reparación global de genoma. El heterodímero XPC-hHR23B localiza lesiones que distorsionan a la doble hélice de DNA.
2. Reparación acoplada a la transcripción. La RNA polimerasa identifica el daño en el DNA. Se detiene la transcripción.
Reparación por escición de nucleótidos en eucariontes
Una vez que se ha reconocido el daño, participana las mismas proteínas en la reparación.
• TFIIH (XPB + XPD) que tiene actividad de helicasa es reclutado.
• XPA y RPA contribuyen a mantener abierta la doble hélice de DNA.
• XPA guía a las endonucleasas para realizar el corte del DNA.
• Endonucleasa XPG corta el DNA en el lado 3’.
• El complejo ERCC1-XPF corta del lado 5’ de la cadena dañada.
• El oligonucleótido (24-32nts) con la región dañada es liberado.
• El hueco es rellenado por la DNA polimerasa δ o ε.
Enfermedades asociadas con defectos en la replicación o reparación del DNA
Asociadas con una alta frecuencia de mutaciones en cromosomas y genes. Algunas se asocian a una alta predisposición a tumores.
• Xeroderma pigmentosum•Mutaciones en genes involucrados en reparación por escición de nucleótidos. Melanoma
• Ataxia telangiectasia• Mutaciones en genes que detectan daño al DNA.• Riesgo incrementado a rayos X.
•Anemia Fanconi.• Mutación en gene involucrado en reparación al DNA.
•Síndrome de Bloom.• Mutación en una helicasa de DNA. • Hipersusceptibilidad a rayos X y a la luz solar.
•Síndrome de Cockayne (CSA/CSB)• Defecto en reparación acoplada a transcripción. • Susceptibilidad a luz solar.
• Síndrome de Werner.• Mutación en un gen de DNA helicasa. • Envejecimiento prematuro.

19
Recombinación de DNA
1630 GENÉTICA Y BIOLOGÍA MOLECULAR
• RECOMBINACIÓN.Se refiere al rearreglo de la información en
moléculas de DNA e involucra la interacción de dos moléculas de DNA.
• RECOMBINACIÓN HOMÓLOGA. Involucra el intercambio genético entre dos moléculas de DNA (o dos segmentos de la misma molécula de DNA) que comparten una región de secuencias homólogas.
• RECOMBINACIÓN SITIO ESPECÍFICO. Involucra el intercambio genético en secuencias definidas de DNA (Ej. Integración del DNA de fago λ al genoma de E.coli)
• TRANSPOSICIÓN DE DNA. La inserción de una molécula de DNA (transposón) en el cromosoma.
Mecanismos de la recombinación.
Para que ocurra la recombinación deben darse los siguientes eventos:
1. Ruptura de las dos cadenas de DNA homólogas
2. Apareamiento de las dos cadenas de DNA homólogas
3. Regeneración de los enlaces fosfodiester para unir las cadenas homólogas
4. Ruptura de las otras dos cadenas y volver a unirlas.
Modelos de recombinación. Modelo de Holliday
Aunque el modelo de Holliday describe con precisión los eventos que ocurren, no explica algunos aspectos del mecanismo.
¿Cómo explicar que ocurran dos cortes en las posiciones exactas correspondientes en cadenas homólogas de DNA?
“Holliday junction”

20
Modelo de la vía RecBCD
• Proteína RecBCD:
• Corte en el extremo 3’ de la secuencia “Chi” χ
• RecBCD también tiene actividad de helicasa.
• Unión de RecA y SSB al DNA
• Invasión y desplazamiento del DNA duplex.
• Formación del asa D
• Apareamiento en la región homóloga.
Modelo de la vía RecBCD
• Apareamiento en la región homóloga.
• Corte en la cadena desplazada
• Apareamiento.
•Sellado por la DNA ligasa
• RuvA + RuvB = Helicasa dependiente de ATP. Desplazamiento de la rama.
• RuvC: Resolvasa, hace los cortes en las dos cadenas para la generación de las dos moléculas de DNA duplex recombinadas.
RecA y RecBCD
Se identificaron en mutantes de E. coli que aceptaban el plásmido F, pero eran incapaces de recombinarlo.
RecA es una proteína de 38 kDa. Promueve intercambio de cadenas in vitro. Se distinguen tres etapas.
Pre-sinápsis. RecA se une al DNAss
Sinápsis. Alineamiento de las secuencias complementarias que participan en el intercambio (50 –150 pb).
Post-Sinápsis. La cadena sencilla desplaza y reemplaza a la cadena (+) del DNA de doble cadena.
- + +
Esta enzima tiene cinco actividades:
exonucleasa V; helicasa,endonucleasa; ATPasa y exonucleasa sobre DNAss
La actividad de helicasa genera asas grandes antes de que el DNA se vuelva a renaturalizar.
Cuando RecBCD encuentra una secuencai Chi, se disocia la subunidad D y la la enzima RecBCactúa como helicasa para abrir el DNA en un evento dependiente de ATP. Se realiza el corte en el DNA y se genera la cadena sencilla de DNA que es estabilizada por RecA y SSB para iniciar el proceso de recombinación.
RecBCD

21
RuvA + RuvB. Desplazamiento de la rama.
Estas proteínas se unen en cruce de Holliday y son las responsables del desplazamiento de la rama.
RuvA es un homotetrámero con simetría cuadrada plana que reconoce y se une al cruce de Holliday. Esto facilita la unión del hexámero RuvB. Helicasa dependiente de ATP, abre la cadena y la desplaza lo necesario para la migración de la rama.
www.sdsc.edu/journals/mbb/ruva.html
Actividad de RuvA
RuvC Resolvasa.
Una vez que se han recombinado las cadenas y se han desplazado, debe ocurrir un corte para liberar el par de DNA duplex.
Hay secuencias consenso de reconocimiento:
5’- (A/T)TT (G/C)-3’